 Adrian Y. S. Lee
Adrian Y. S. Lee Dorothea Reimer4‡
Dorothea Reimer4‡ Ming Lu
Ming Lu Heinrich Körner
Heinrich Körner- 1Menzies Institute for Medical Research, University of Tasmania, Hobart, TAS, Australia
- 2Department of Internal Medicine, Western Hospital, Footscray, VIC, Australia
- 3Department of Medicine and Radiology, The University of Melbourne, Melbourne, VIC, Australia
- 4Division of Molecular Immunology, Internal Medicine III, University Hospital Erlangen, Nikolaus-Fiebiger-Center, University of Erlangen-Nuremberg, Erlangen, Germany
- 5Institute of Clinical Pharmacology, Anhui Medical University, Key Laboratory of Anti-Inflammatory and Immunopharmacology, Ministry of Education, Engineering Technology Research Center of Anti-inflammatory and Immunodrugs in Anhui Province, Hefei, China
The CC chemokine receptor 6 (CCR6) and its sole chemokine ligand CC chemokine ligand 20 (CCL20) display an emerging role in the coordination of humoral immune responses. Recent studies demonstrate a role of this chemokine axis in the migration of B cells to key immunological sites during an immune response, and facilitating the generation of high-quality antibodies. Very little, however, is known about CCL20 and its role in these functions. We undertook a preliminary investigation into the expression and function of CCL20 and demonstrate its well-noted upregulation in the spleen during immunization. Furthermore, we show that most follicular T helper (Tfh) cells can be CCR6+ and can produce CCL20. Surprisingly, CCL20 cannot only be found in the cytoplasm but also on the surface of these cells and their precursors. Analysis of T–B-cell conjugates revealed that mature Tfh cells, but not their precursors, are highly enriched in the conjugates. Further functional studies are needed to unravel the precise role of CCL20 in coordinating T and B cell interactions during the humoral immune response.
Introduction
CC chemokine ligand 20 (CCL20) is emerging as an important mediator in the molecular orchestration of immunological homeostasis as well as the inflammatory and the humoral immune response (1, 2). CCL20 is expressed in endothelial cells (3), macrophages (4), and Th17 cells (5) and is upregulated during inflammation (6). The only known receptor for CCL20 is the CC chemokine receptor 6 (CCR6) which is found predominantly on dendritic cells (DC) (7), and on various lympocyte populations such as B cells, regulatory T cells (Tregs), Th17, and memory T cells (2). Mice genetically deficient for CCR6 demonstrate structural and functional deficits in mucosal immunity (8) highlighting the importance of this chemokine axis in homeostasis. Under inflammatory conditions, the regulation of CCR6 on DC, CD4+ T cell subpopulations, and B cells controls the migration of immune cells for correct positioning facilitating an efficient response (9, 10).
The cellular expression of CCL20 and its putative function in the humoral immune response are still not understood. In comparison, its partner CCR6 on B cells has been characterized and its dynamic expression has been found to orchestrate B cells and the germinal center (GC) response. B cells that leave the bone marrow to mature upregulate CCR6, and become highly responsive to CCL20—the precise reason of which has not been fully elucidated (9, 11). In CCR6-deficient mice, the GC response was quantitatively enhanced but at the expense of efficient and specific humoral immune responses (11, 12) and memory B cell responses (10). This suggests an important role for the CCR6–CCL20 chemokine axis in humoral immunity. To further investigate these findings, we decided to investigate the potential role for CCL20 in coordinating T and B cell interactions.
Materials and Methods
Mice and Immunizations
C57BL/6 WT and CCR6−/− mice on a C57BL/6 background (B6.CCR6−/−) were sourced and generated as described (13). Animal experiments were approved by the University of Tasmania Animal Ethics Committee. Immunization was carried out by injecting 1.0 × 108 sheep red blood cells (SRBC) suspended in sterile PBS intraperitoneally.
Polymerase Chain Reaction (PCR)
Spleens from sacrificed mice were removed and homogenized in liquid nitrogen. Spleens were then added to 1 mL of TriReagent (Sigma-Aldrich, NSW, Australia) and RNA extraction carried out as per manufacturer’s instructions. cDNA was synthesized from total RNA using Tetro cDNA synthesis kit (Bioline, Alexandria, NSW, Australia). Template cDNA was stored at −20°C until further use. Quantitative real-time PCR was performed on cDNA templates using the SensiFAST SYBR No-Rox kit (Bioline). Results were normalized to β-actin (sense 5′- AGA GGG AAA TCG TGC GTG AC -3′ and anti-sense 5′- CAA TAG TGA TGA CCT GGC CGT -3′). The following primers were used: Ccr6 (sense 5′- TGT CCT CAC CCT ACC GTT CTG -3′ and anti-sense 5′- TAC AGG CCA GGA GCA GCA T -3′), and Bcl6 (sense 5′- CTG CAG ATG GAG CAT -3′ and anti-sense 5′- CGG CTG TTC AGG AAC -3′).
Antibodies
The following rat anti-mouse antibodies and conjugations were obtained from BioLegend (Australian Biosearch, WA, Australia), BD Biosciences (Sydney, NSW, Australia), or eBioscience (Sydney, NSW, Australia) and used for flow cytometry: B220-Biotin (clone RA3-6B2), CD19-APC Fire 750 (6D5), CCR6-PE (29-2L17), CCR6-AF647 (140706), CD11b-PerCP-Cy5.5 (M1/70), CD11b-BV510 (M1/70), CD4-APC (RM4-5), CD4-PerCP Cy5.5 (RM4-5), CD8α-PB (53-6.7), CXCR5-Biotin (2G8), CXCR5-PerCP-Cy5.5 (2G8), PD-1-PE (J43), PD-1-PE-Cy7 (J43), TCR-αβ-PB (HM3628, Thermo Fisher Scientific Australia, Soresby, VIC, Australia), hamster IgG1-λ isotype-PE (G235–2356), and rat IgG1-κ isotype-FITC (eBRG1). Cy5-conjugated streptavidin (Jackson Immuno Research, Pennsylvania, PA, USA) was used as secondary reagent. Unlabeled CCL20 (114906) was obtained from R&D Systems (Sydney, NSW, Australia) and labeled with DyLight 488 Microscale Antibody Labeling Kit (Thermo Fisher Scientific, Australia) according to the manufacturer’s instructions.
Flow Cytometry
Murine spleens were dissected and pushed through a 40 µm nylon cell strainer to obtain a single cell suspension. After washing, the cells were resuspended in 10 mL of red blood cell lysis buffer and left to incubate at room temperature for 10 min. For cells undergoing intracellular cytokine staining, 0.5 µL of 200 µg/mL PMA (Sigma-Aldrich) and 0.5 µL of 10 mM ionomycin (Thermo Fisher Scientific, Australia) were added to a 5 mL resuspension of the cells in RPMI medium (Thermo Fisher Scientific, Australia) and were incubated at 37°C for 1 h, 5% CO2. Following this, 1 µL of Golgi stop (BD Biosciences) (equivalent to 3.75 mM monensin) was added and the suspension incubated for further 3 h at 37°C.
Multicolor flow cytometry was performed on splenocytes using CyAn ADP and Gallios flow cytometers (Beckman Coulter, Inc., NSW, Australia). Post-acquisition analysis was performed using FlowJo v7.6 software (TreeStar, Inc., Ashland, OR, USA) and Kaluza 1.5a software (Beckman Coulter, Inc.). Fluorescent-activated cell sorting (FACS) was performed using the MoFlo Astrios cell sorter (Beckman Coulter, Inc.) with a sorting accuracy >95%.
For surface-bound chemokine studies, cells were blocked with Fc-receptor block (BD Biosciences) for 30 min on ice and antibody staining performed under sodium azide-free conditions. Fluorescent-minus-one and isotype controls were used to minimize non-specific binding and autofluorescence. To dissociate CCL20 from the surface cells were washed with acid as described (14). Briefly, cells were pelleted and resuspended in 200 µL ice-cold acid buffer for 2 min. Samples were neutralized with 5× volumes of R10 medium and cells were washed twice with FACS buffer (2% FCS, 0,01% NaN3 in PBS). In the following experiments, Fc-block and surface staining were performed as indicated.
Statistics
Statistical analyses using the non-parametric two-tailed Mann–Whitney U test were performed and graphs constructed using Prism software v5 (GraphPad Software). Graph columns represent median with error bars representing range. The alpha value was set at 0.05. In our figures these have been summarized as: *p < 0.05, **p < 0.01, ***p < 0.001.
Results and Discussion
CCL20 Expression in Different Splenic Immune Cell Types
CC chemokine ligand 20 is an inflammatory chemokine that is upregulated by the pro-inflammatory cytokine TNF (3) or induced by TGF-β and IL-6, or IL-21 during TH17 cell differentiation (5). To examine whether CCL20 is involved in humoral immune responses, we immunized mice with SRBC, known to trigger strong GC responses. Splenocytes from unchallenged mice expressed virtually no CCL20 (Figure S1A in Supplementary Material). After immunization with SRBC splenocytes showed an upregulation of CCL20 mRNA in a time-dependent manner reaching approximately ninefold expression by day 7 (Figure S1A in Supplementary Material). Interestingly, Ccl20 has a very low-constitutive expression in the spleen, relative to other organs such as Peyer’s patches (15) (Adrian Y. S. Lee and Heinrich Körner, unpublished data) suggesting a subordinate role in splenic immune homeostasis. In fact, the rapid and marked elevation of Ccl20 post-immunization suggests it has a very specific role in the inflammatory response. Therefore, we examined the expression of CCL20 in this organ for subsequent experiments.
To identify possible sources for splenic CCL20, we sorted day 5 SRBC-immunized splenocytes and performed quantitative real-time PCR on major cellular subpopulations found in the spleen: B cells (B220+TCR-β¯), CD4+ T cells (B220¯TCR-β+CD4+), CD8+ T cells (B220¯TCR-β+CD8+), and miscellaneous CD11b+ cell populations (B220¯TCR-β¯CD11b+) which predominantly consists of NK cells, splenic macrophages, and granulocytes (4, 16). An earlier study (10) demonstrated very low-constitutive Ccl20 expression in the CD4+ T cell compartment of the spleen and no expression in the CD8+ T cell or monocytic compartments. Our study, however, demonstrated Ccl20 mRNA highly expressed in CD11b+ cells after activation (Figure S1B in Supplementary Material). In comparison, total splenic Ccl20 expression was low suggesting a marked dilution by non-CCL20-producing cells, most likely the B cells. This is in line with early expression studies that found overall very low amounts of constitutive splenic Ccl20 compared with other organs (15). However, we show a substantially upregulated expression induced upon immunization (Figure S1A in Supplementary Material) or challenge using bacterial lipopolysaccaride (data not shown).
Intracellular CCL20 Expression in Different Subsets of CD4+ T Cells
CC chemokine receptor 6, the CCL20 receptor, modulates GC reactions and antibody affinity maturation, which appears to be regulated by follicular T helper (Tfh) cells (17). Thus, the CCR6/CCL20 axis might be involved in the GC response via Tfh cells (12) and we decided to focus our attention on CD4+ T cells, and specifically, Tfh cells. To initiate a strong humoral immune response, we used SRBC as a T cell-dependent antigen. We identified the Tfh population in spleen (Figure 1A) and sorted Tfh cells (defined as CD4+CXCR5hiPD-1hi) from SRBC-immunized mice (day 5 p.i.), an intermediate population Tfhint (CD4+CXCR5loPD-1lo) and the remaining CD4+ T cells (non-Tfh cells; CD4+CXCR5−PD-1−) (Figure 1A). These non-Tfh cells comprise other T helper subsets (e.g., Th1), Tregs, and memory T cells including memory Tfh cells (18). The classic effector Tfh cells were significantly increased following immunization, confirming their activation (Figure S2 in Supplementary Material). To validate the quality of the isolated cell populations, we subjected sorted Tfh, Tfhint, and non-Tfh cells to qRT-PCR for Bcl6, demonstrating a non-significant trend toward highest mRNA expression in the Tfh cells, moderate expression in the intermediary population, and lowest expression in the non-Tfh cells (Figure 1B). The fact that we detected more Bcl6 in sorted CD4+TCRb+CXCR5+PD-1+ cells corroborates their identity as Tfh as shown previously (19). Similarly, we also show that mRNA expression of Ccr6 tends to be highest in Tfh cells (Figure 1C), which corresponds to protein expression as determined by flow cytometry (Figure 1D).
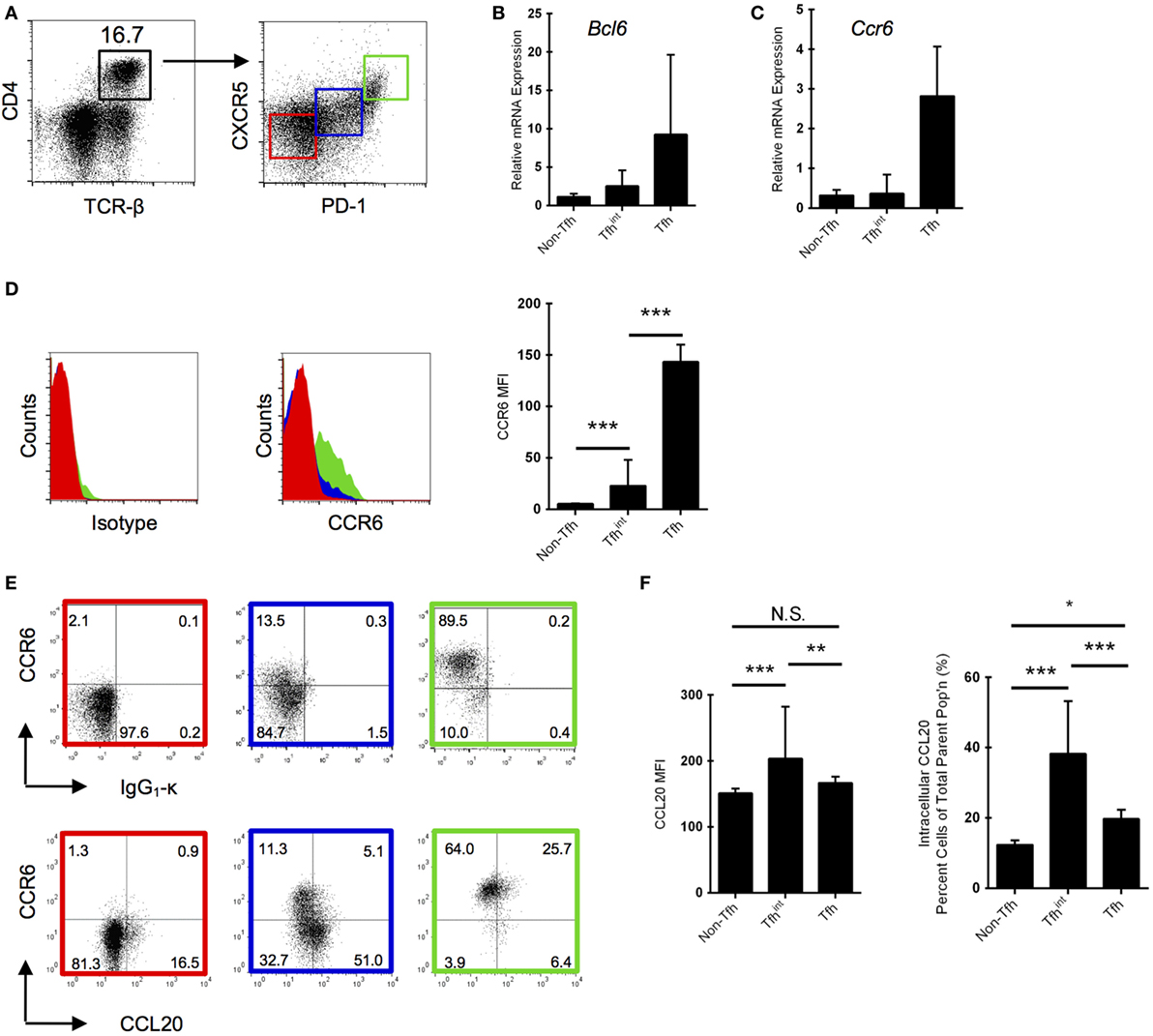
Figure 1. Follicular T helper (Tfh) cells express CC chemokine receptor 6 (CCR6) and are positive for CC chemokine ligand 20 (CCL20) protein. (A) Gating strategy for Tfh cells isolated from splenocytes by fluorescent-activated cell sorting. Non-Tfh cells (PD-1−CXCR5−) are marked in red, intermediary Tfh (Tfhint) (PD-1loCXCR5lo) cells are marked in blue, and Tfh cells (PD-1hiCXCR5hi) marked in green. mRNA expression of Bcl6 (B) and Ccr6 (C) normalized to Actb in the three isolated Tfh populations. One representative experiment is shown. Results are representative of two experiments with five to six mice each (Bcl6 and Ccr6) with 5-day sheep red blood cells (SRBC)-immunized mice. Means between each column in (B,C) are not significant (p > 0.05). (D) CCR6 protein expression in the three T cell populations via flow cytometry using an isotype control (left panel). Mean fluorescence intensity (MFI) of CCR6 is also provided (right panel). (E) Differential CCR6 and intracellular CCL20/isotype protein expression in the three splenic T cell populations via flow cytometry in day 5 SRBC-immunized mice. CCL20 protein expression was quantified as MFIs and percentages (F). CCL20 gating is based on an isotype control. Representative panels from five independent experiments with two to five mice each. *p < 0.05, **p < 0.01, ***p < 0.001.
The canonical CD4+CCR6+ T cell population has been attributed to Th17 cells (20, 21); however, we show approximately 90% of Tfh cells are positive for CCR6 (Figure 1E), consistent with our mRNA data for strong Ccr6 expression in this population (Figure 1C). We performed intracellular cytokine staining and show that of the Tfhint population, a marked proportion of CCR6− cells are CCL20+, whereas very little CCR6+Tfhint cells are positive (Figure 1E). In contrast, the CCL20 expression is less marked in Tfh cells, being present in approximately 30% of cells (Figure 1E). The dominant intracellular CCL20 expression in the intermediary population (Figure 1F) is congruent to microarray data that demonstrated that Ccl20 is upregulated in intermediate population and down-regulated in Tfh cells (19). Interestingly, in a second study, CCL20 expression has also been described to be higher in Tfr cells than in regular Tfh cells (22).
Surface Expression of CCL20 on Tfh Cells
Since Tfh cells are known to form conjugates with CCR6+ B cells, we examined surface expression of CCL20 on these cells. After eliminating autofluorescence (Figure S3 in Supplementary Material) and non-specific binding through isotype and fluorescence-minus-one controls, we show that compared with non-Tfh cells, approximately 4% of Tfhint and Tfh cells show CCL20 on their surface (Figures 2A,B). To eliminate the possibility of the flow cytometric assay detecting bound CCL20 to CCR6, we used CCR6−/− mice and repeated the experiments. CCR6−/− mice similarly demonstrated surface CCL20 expression indicating that CCL20 is bound independent of CCR6 (Figure 2B). The percentage of T cells with surface-bound CCL20 did not differ significantly between parent populations of WT and CCR6−/− mice (Figures 2B,C).
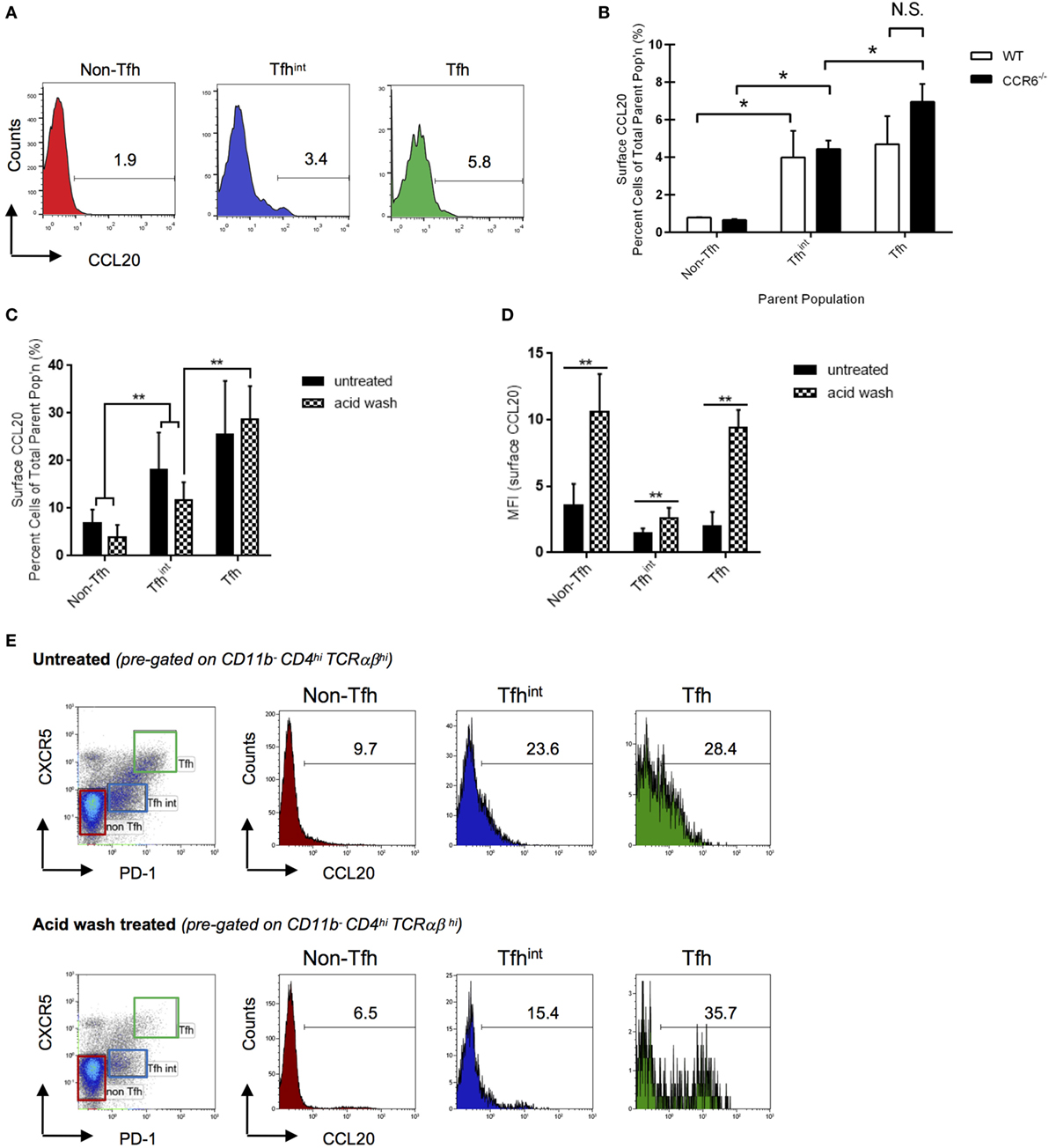
Figure 2. CC chemokine ligand 20 (CCL20) is found on the surface of splenic T cells. Surface-bound CCL20 was determined on T cells from sheep red blood cells-immunized WT and CCR6−/− mice at day 5 after immunization. Non-follicular T helper (Tfh) cells (CD11b−CD4hiTCRαβhiPD-1−CXCR5−), Tfhint (CD11b−CD4hiTCRαβhiPD-1loCXCR5lo), and Tfh cells (CD11b−CD4hiTCRαβhiPD-1hiCXCR5hi) were analyzed by flow cytometry. (A) Representative flow cytometry histograms of percent T cells with surface CCL20 in WT mice. The gate was made based on an isotype control. (B) Statistical analysis of the percentage of CCL20+ T cells in the total subpopulation comparing WT and CCR6−/− mice. Representative bar graph from two experiments with 10 mice of each genotype in total. (C) Percent surface CCL20 was analyzed in an independent experiment as well as CCL20 mean fluorescence intensities (D). Cells were left untreated or stripped with acid buffer to remove non-specifically bound CCL20 from the surface. Flow cytometry plots representative for two experiments with 9 mice in total are displayed (E). *p < 0.05, **p < 0.01.
To validate our differential flow cytometric detection of intracellular versus surface CCL20, we analyzed CCL20 expression on in vitro-generated Th17 cells (23, 24). As expected, after activation, a major percentage of Th17 cells (circa 90% of Th17 cells) display a strong expression of intracellular CCL20 as published previously (5). However, additionally we were able to detect surface CCL20 on about 25% of Th17 cells (Figure S4 in Supplementary Material).
Chemokines are known to bind sulfated polysaccharides [glycosaminoglycans (GAGs)] found on cell surfaces (25). The presence of GAGs cause polymerization of chemokines, thereby increasing local concentration and facilitating chemokine-receptor binding (26). In this case, the surface-bound CCL20 likely functions to increase local concentration of the chemokine in order to create a concentration gradient for CCR6+ cells [haptotaxis (27)]—a likely candidate being partnering CCR6+ B cells to T cells as T–B cell conjugates.
The likely source of surface-bound CCL20 appears to be Tfhint and Tfh cells as indicated by protein (Figures 1E,F) and mRNA expression studies (19, 22). However, it is equally possible that CCL20 originates from other sources and becomes tethered on their surfaces. The closely related CCL19 and CCL21, for example, could be detected on the luminal side of high-endothelial venules despite the fact that these cells do not seem to express mRNA for these chemokines. Instead, they rely on transcytosis of the ligands from the surrounding T cell zone, presenting them on their surface to attract CCR7+ T cells to secondary lymphoid organs (28, 29). Therefore, it is possible that the source of CCL20 is the GC as both CCR6 and CCL20 are found in GC in lymphoid tissue (30) (Adrian Y. S. Lee and Heinrich Körner, unpublished data). To test whether CCL20 on Tfhint and Tfh cells is covalently expressed on the cell surface or bound via GAGs or others, we treated cells with an acid wash before staining. This experiment confirmed the increase in CCL20 expression proportional to Tfh differentiation, with acid wash leaving the frequency of CCL20 expressing cells unchanged (Figure 2C). Unexpectedly, acid wash treatment enhanced the intensity of CCL20 staining (Figure 2D). Careful examination of the histograms showed that acid treatment removed the low-/intermediate surface CCL20 surface expression but not the high-CCL20 expression in the CCL20 positive cell fraction (Figure 2E). Hence, high-CCL20 surface expression represents covalently bound CCL20 and intermediate/low-expression represents CCL20 bound to GAG or others, but likely not in an autocrine or paracrine fashion to CCR6 (see Figure 2B). These data infer that a cell population with a low frequency of CCL20+ cells, for instance at a certain differentiation stage such as non-Tfh cells, can eventually provide as many CCL20 ligand binding sites as a population with a higher frequency of CCL20 positive cells. Cells displaying acid-sensitive CCL20 might serve as a mobile source of CCL20, thereby, providing spatio-temporal information during immune responses. Hence, even non-Tfh cells could have the capability of interacting with CCR6-expressing B cells; for instance at the entry point of the GC (12).
Tfh Cells in T–B Conjugates
To assess whether this surface-bound CCL20–CCR6 interaction is essential for the formation of T–B cell conjugates, we compared the formation of T–B cell conjugates in SRBC-immunized spleens from WT and CCR6−/− mice by flow cytometry. By gating on doublets and CD4+CD19+ cells, we determined a surrogate measure of T–B cell conjugates (Figure 3A)—a method performed previously (31). We found a significantly higher proportion of CD4+CD19+ conjugates in CCR6−/− mice compared with WT mice, indicating that the CCR6–CCL20 chemokine axis is not essential for the formation of these T–B cell conjugates (Figure 3B). This would be in accordance with the observation that mice deficient in CCR6 appeared to form more GCs in an accelerated fashion in the spleen than their WT counterparts (11). To determine which subpopulation of CCL20 expressing T cells (see Figures 2C,D) contributes to T–B-conjugate formation, we analyzed Tfh and Tfhint populations cells in CD11b-negative singlets or aggregates (Figure 3A, gating strategy). Whereas Tfh cells represent only 6–10% of CD4hiTCRhi cells within singlets or T cell duplets (Figures 3A,C), they are highly enriched over non-Tfh or Tfhint cells in T–B-conjugates (Figures 3D,E). The stability of the conjugates was addressed in control experiments using acid washing and treatment with EDTA. The proportion of T and B cells in conjugates was not changed by these treatments (data not shown).
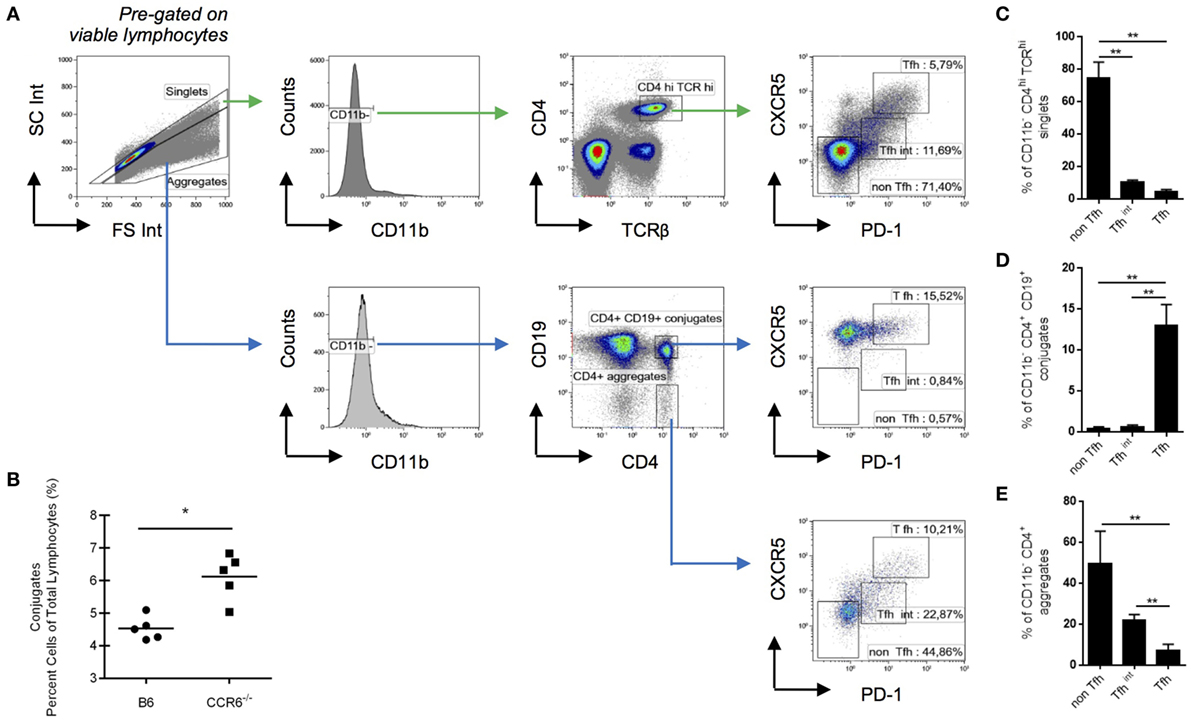
Figure 3. Follicular T helper (Tfh) cells represent the main CD4 subpopulation in T–B cell conjugates. Splenocytes from WT mice were analyzed at day 5 post-sheep red blood cells immunization for the composition of T–B cell conjugates. (A) Gating strategy for different CD4 subsets in single cells and in cell aggregates. (B) Conjugates are expressed as a percentage of total gated lymphocytes in WT and CCR6−/− mice. (C) The frequency of non-Tfh cells (PD-1−CXCR5−), Tfhint (PD-1loCXCR5lo), and Tfh cells (PD-1hiCXCR5hi) within CD11b− CD4hi TCRαβhi are shown for singlets. (D) Using the same gates for non-Tfh cells, Tfhin and Tfh cells, the frequency of these three subpopulations is shown in CD11b−CD4+CD19+ conjugates and, as a control, in CD11b−CD4+CD19− conjugates (E), both pre-gated on cell aggregates in the forward-side-scatter. Shown are experiments with five mice {both genotypes; (B) or six mice [only wt; (A,C,D,E)]}. *p < 0.05, **p < 0.01.
Thus, although the presence of CCR6 regulates T–B-conjugate formation negatively (Figure 3B), CCL20+ Tfh cells are most enriched in T–B-conjugates. Whether the B cells interacting with CCL20hi Tfh here represent light zone B cells that express CCR6 (17) remains to be tested. In this case, the CCR6–CCL20 axis could be responsible for maintenance of the pairing between Tfh and cognate light zone B cell, thereby, contributing to the selection of high-affinity B cells since CCR6−/− mice display poorer antibody quality production (11, 12).
On the other hand, our data show that Tfhint cells have a relatively high frequency of low-/intermediate CCL20 expressing cells (Figure 2E). It is known that T cells near the T–B border already express Bcl6 and are PD-1lo, preceding a Bcl6+PD-1hi phenotype (32), and therefore may correspond to our Tfhint. This Tfhint population appears to precede cognate B cell interaction (33) and appears to be a necessary step in the selection of B cells and hence, formation of a GC (34). This notion together with our data implicates a fine-tuned CCR6–CCL20 axis in this critical conjugation, which might be modulated by the presentation of non-covalently CCL20 on the surface. Thus, the CCL20–CCR6 axis likely functions at several GC checkpoints governing the efficacy of the GC reaction via B cells. It is tempting to speculate that two different mechanisms may act at the GC entry and at the GC selection point, depending on quantity and quality of surface-presented CCL20.
To conclude, we have shown that CCL20 is a chemokine that is produced by monocytes and T cells in the spleen. As subsets of the latter, Tfh and Tfhint cells are producers of CCL20 which is also tethered on their surfaces, an observation we have validated in vitro using Th17 cells. A likely function of this CCL20 is in the role of coordinating cognate CCR6+ B cells to receive T cell help for the formation of antibodies. As optimal humoral immune responses rely on this critical T–B cell interaction, further research into the role of the CCR6–CCL20 chemokine axis in this conjugation will be helpful in understanding suboptimal immune responses and the pathogenesis of autoimmune disease.
Ethics Statement
Animal experiments were approved by the University of Tasmania Animal Ethics Committee (A13934—Immunization in C57BL/6 and substrains).
Author Contributions
AL: designed experiments, carried out experiments, acquired data, analyzed data, wrote the manuscript, and edited the manuscript. DR: designed experiments, carried out experiments, acquired data, analyzed data, and edited the manuscript. AZ and ML: carried out experiments, acquired data, analyzed data, and edited the manuscript. DM: designed experiments, carried out experiments, acquired data, analyzed data, edited the manuscript, and revised the final version. HK: conception of the project, designed experiments, analyzed data, wrote the manuscript, edited the manuscript, and revised the final version.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Jocelyn Darby and Kathy Buttigieg for their technical assistance and Paul Scowen and his team for expert animal husbandry. AZ was recipient of a Tasmania Graduate Research Scholarship. ML is a recipient of a AMU/UTAS postgraduate scholarship. The study was funded by the NHMRC (APP1004592 to HK), the Menzies Institute for Medical Research Tasmania, the Lupus Association of Tasmania, and the Deutsche Forschungsgemeinschaft (DFG;TRR130/2, Project 3, to DM).
Supplementary Material
The Supplementary Material for this article can be found online at http://www.frontiersin.org/articles/10.3389/fimmu.2017.01871/full#supplementary-material.
Figure S1. CC chemokine ligand 20 (CCL20) mRNA becomes upregulated in splenocytes upon immunization and is highly expressed in splenic monocytes and T cells. (A) Time course of total spleen Ccl20 mRNA expression after SRBC intraperitoneal immunization. Graph is representative of six to eight mice at each timepoint. (B) Ccl20 mRNA expression in splenic cellular subpopulations. Day 5 SRBC-immunized spleens were sorted for the four subpopulations. Intact splenocytes were used for “total spleen” and expression was measured relative to Actb. Data from three mice and one experiment are shown. NTC, no-template control; ND, not detected. **p < 0.01, ***p < 0.001.
Figure S2. Upregulation of Tfh cells after mice immunization. B6 mice were immunized with intraperitoneal sheep red blood cells and splenocytes subjected to flow cytometry. A clear upregulation of Tfh cells can be detected after 5 days of immunization. Shown here are representative flow plots from five independent experiments.
Figure S3. Method of gating out autofluorescence in flow cytometry. To eliminate the possibility of non-specific fluorescence contributing to apparent cell surface chemokine expression, only cells negative for unutilized fluorescent channels were gated in for analyses (sample plot shown).
Figure S4. CC chemokine ligand 20 (CCL20) and Th17 cells. Assessment of CCL20 expression of Th17 cells in relation to cell proliferation was performed using flow cytometry. Isolated CD4+ lymph node T cells were labeled with cell trace violet (CTV) and were activated in vitro with CD3/CD28 in the presence of a cocktail of TGF-β, IL-6, IL-23 in combination with anti-IL-4 and anti-IFN-γ for 72 h following standard protocols. The expression of CCL20 on the surface (A) and intracellularly (B) was detected using a directly labeled anti-CCL20 mAb. The expression of IL-17 was independently verified using intracellular flow cytometry and is not shown. A representative result is shown.
References
1. Comerford I, Bunting M, Fenix K, Haylock-Jacobs S, Litchfield W, Harata-Lee Y, et al. An immune paradox: how can the same chemokine axis regulate both immune tolerance and activation? CCR6/CCL20: a chemokine axis balancing immunological tolerance and inflammation in autoimmune disease. Bioessays (2010) 32(12):1067–76. doi:10.1002/bies.201000063
2. Lee AY, Phan TK, Hulett MD, Korner H. The relationship between CCR6 and its binding partners: does the CCR6-CCL20 axis have to be extended? Cytokine (2015) 72(1):97–101. doi:10.1016/j.cyto.2014.11.029
3. Meissner A, Zilles O, Varona R, Jozefowski K, Ritter U, Marquez G, et al. CC chemokine ligand 20 partially controls adhesion of naive B cells to activated endothelial cells under shear stress. Blood (2003) 102(8):2724–7. doi:10.1182/blood-2003-01-0007
4. Schroder K, Irvine KM, Taylor MS, Bokil NJ, Le Cao KA, Masterman KA, et al. Conservation and divergence in toll-like receptor 4-regulated gene expression in primary human versus mouse macrophages. Proc Natl Acad Sci U S A (2012) 109(16):E944–53. doi:10.1073/pnas.1110156109
5. Yamazaki T, Yang XO, Chung Y, Fukunaga A, Nurieva R, Pappu B, et al. CCR6 regulates the migration of inflammatory and regulatory T cells. J Immunol (2008) 181(12):8391–401. doi:10.4049/jimmunol.181.12.8391
6. Lee AY, Eri R, Lyons AB, Grimm MC, Korner H. CC chemokine ligand 20 and its cognate receptor CCR6 in mucosal T cell immunology and inflammatory bowel disease: odd couple or axis of evil? Front Immunol (2013) 4:194. doi:10.3389/fimmu.2013.00194
7. Dieu MC, Vanbervliet B, Vicari A, Bridon JM, Oldham E, Ait-Yahia S, et al. Selective recruitment of immature and mature dendritic cells by distinct chemokines expressed in different anatomic sites. J Exp Med (1998) 188(2):373–86. doi:10.1084/jem.188.2.373
8. Cook DN, Prosser DM, Forster R, Zhang J, Kuklin NA, Abbondanzo SJ, et al. CCR6 mediates dendritic cell localization, lymphocyte homeostasis, and immune responses in mucosal tissue. Immunity (2000) 12(5):495–503. doi:10.1016/S1074-7613(00)80201-0
9. Bowman EP, Campbell JJ, Soler D, Dong Z, Manlongat N, Picarella D, et al. Developmental switches in chemokine response profiles during B cell differentiation and maturation. J Exp Med (2000) 191:1303–18. doi:10.1084/jem.191.8.1303
10. Elgueta R, Marks E, Nowak E, Menezes S, Benson M, Raman VS, et al. CCR6-dependent positioning of memory B cells is essential for their ability to mount a recall response to antigen. J Immunol (2015) 194(2):505–13. doi:10.4049/jimmunol.1401553
11. Wiede F, Fromm PD, Comerford I, Kara E, Bannan J, Schuh W, et al. CCR6 is transiently upregulated on B cells after activation and modulates the germinal center reaction in the mouse. Immunol Cell Biol (2013) 91(5):335–9. doi:10.1038/icb.2013.14
12. Reimer D, Lee AY, Bannan J, Fromm P, Kara EE, Comerford I, et al. Early CCR6 expression on B cells modulates germinal centre kinetics and efficient antibody responses. Immunol Cell Biol (2017) 95(1):33–41. doi:10.1038/icb.2016.68
13. Varona R, Villares R, Carramolino L, Goya I, Zaballos A, Gutierrez J, et al. CCR6-deficient mice have impaired leukocyte homeostasis and altered contact hypersensitivity and delayed-type hypersensitivity responses. J Clin Invest (2001) 107(6):37–45. doi:10.1172/JCI11297
14. Ishizaka T, Ishizaka K. Mechanisms of passive sensitization. IV. Dissociation of IgE molecules from basophil receptors at acid pH. J Immunol (1974) 112(3):1078–84.
15. Hieshima K, Imai T, Opdenakker G, Van Damme J, Kusuda J, Tei H, et al. Molecular cloning of a novel human CC chemokine liver and activation-regulated chemokine (LARC) expressed in liver. Chemotactic activity for lymphocytes and gene localization on chromosome 2. J Biol Chem (1997) 272(9):5846–53. doi:10.1074/jbc.272.9.5846
16. Hey YY, Tan JK, O’Neill HC. Redefining myeloid cell subsets in murine spleen. Front Immunol (2015) 6:652. doi:10.3389/fimmu.2015.00652
17. Victora GD, Schwickert TA, Fooksman DR, Kamphorst AO, Meyer-Hermann M, Dustin ML, et al. Germinal center dynamics revealed by multiphoton microscopy with a photoactivatable fluorescent reporter. Cell (2010) 143(4):592–605. doi:10.1016/j.cell.2010.10.032
18. Weber JP, Fuhrmann F, Hutloff A. T-follicular helper cells survive as long-term memory cells. Eur J Immunol (2012) 42(8):1981–8. doi:10.1002/eji.201242540
19. Liu X, Yan X, Zhong B, Nurieva RI, Wang A, Wang X, et al. Bcl6 expression specifies the T follicular helper cell program in vivo. J Exp Med (2012) 209(10):S1–24. doi:10.1084/jem.20120219
20. Acosta-Rodriguez EV, Rivino L, Geginat J, Jarrossay D, Gattorno M, Lanzavecchia A, et al. Surface phenotype and antigenic specificity of human interleukin 17-producing T helper memory cells. Nat Immunol (2007) 8(6):639–46. doi:10.1038/ni1467
21. Hirota K, Yoshitomi H, Hashimoto M, Maeda S, Teradaira S, Sugimoto N, et al. Preferential recruitment of CCR6-expressing Th17 cells to inflamed joints via CCL20 in rheumatoid arthritis and its animal model. J Exp Med (2007) 204(12):2803–12. doi:10.1084/jem.20071397
22. Chowdhury A, Del Rio Estrada PM, Tharp GK, Trible RP, Amara RR, Chahroudi A, et al. Decreased T follicular regulatory cell/T follicular helper cell (TFH) in simian immunodeficiency virus-infected rhesus macaques may contribute to accumulation of TFH in chronic infection. J Immunol (2015) 195(7):3237–47. doi:10.4049/jimmunol.1502269
23. Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature (2006) 441(7090):235–8. doi:10.1038/nature04753
24. Mangan PR, Harrington LE, O’Quinn DB, Helms WS, Bullard DC, Elson CO, et al. Transforming growth factor-beta induces development of the T(H)17 lineage. Nature (2006) 441(7090):231–4. doi:10.1038/nature04754
25. Kuschert GS, Coulin F, Power CA, Proudfoot AE, Hubbard RE, Hoogewerf AJ, et al. Glycosaminoglycans interact selectively with chemokines and modulate receptor binding and cellular responses. Biochemistry (1999) 38(39):12959–68. doi:10.1021/bi990711d
26. Hoogewerf AJ, Kuschert GS, Proudfoot AE, Borlat F, Clark-Lewis I, Power CA, et al. Glycosaminoglycans mediate cell surface oligomerization of chemokines. Biochemistry (1997) 36(44):13570–8. doi:10.1021/bi971125s
27. Weber M, Hauschild R, Schwarz J, Moussion C, de Vries I, Legler DF, et al. Interstitial dendritic cell guidance by haptotactic chemokine gradients. Science (2013) 339(6117):328–32. doi:10.1126/science.1228456
28. Baekkevold ES, Yamanaka T, Palframan RT, Carlsen HS, Reinholt FP, von Andrian UH, et al. The CCR7 ligand elc (CCL19) is transcytosed in high endothelial venules and mediates T cell recruitment. J Exp Med (2001) 193(9):1105–12. doi:10.1084/jem.193.9.1105
29. Weninger W, Carlsen HS, Goodarzi M, Moazed F, Crowley MA, Baekkevold ES, et al. Naive T cell recruitment to nonlymphoid tissues: a role for endothelium-expressed CC chemokine ligand 21 in autoimmune disease and lymphoid neogenesis. J Immunol (2003) 170(9):4638–48. doi:10.4049/jimmunol.170.9.4638
30. Buri C, Gutersohn A, Hauser C, Kappeler A, Mueller C. The chemokines CCL11, CCL20, CCL21, and CCL24 are preferentially expressed in polarized human secondary lymphoid follicles. J Pathol (2004) 204(2):208–16. doi:10.1002/path.1622
31. Reinhardt RL, Liang HE, Locksley RM. Cytokine-secreting follicular T cells shape the antibody repertoire. Nat Immunol (2009) 10(4):385–93. doi:10.1038/ni.1715
32. Lee SK, Rigby RJ, Zotos D, Tsai LM, Kawamoto S, Marshall JL, et al. B cell priming for extrafollicular antibody responses requires Bcl-6 expression by T cells. J Exp Med (2011) 208(7):1377–88. doi:10.1084/jem.20102065
33. Vinuesa CG, Cyster JG. How T cells earn the follicular rite of passage. Immunity (2011) 35(5):671–80. doi:10.1016/j.immuni.2011.11.001
Keywords: T–B cell communication, CC chemokine ligand 20, CC chemokine receptor 6, humoral immune response, T cells
Citation: Lee AYS, Reimer D, Zehrer A, Lu M, Mielenz D and Körner H (2017) Expression of Membrane-Bound CC Chemokine Ligand 20 on Follicular T Helper Cells in T–B-Cell Conjugates. Front. Immunol. 8:1871. doi: 10.3389/fimmu.2017.01871
Received: 01 May 2017; Accepted: 08 December 2017;
Published: 21 December 2017
Edited by:
Francisco Sanchez-Madrid, Universidad Autonoma de Madrid, SpainReviewed by:
Cosima T. Baldari, University of Siena, ItalyVasileios Bekiaris, Technical University of Denmark, Denmark
Copyright: © 2017 Lee, Reimer, Zehrer, Lu, Mielenz and Körner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Heinrich Körner, aGVpbnJpY2gua29ybmVyQHV0YXMuZWR1LmF1
†Present address: Annette Zehrer, LMU Biomedizinsches Centrum, Institut für Kardiovaskuläre Physiologie und Pathophysiologie, Walter-Brendel-Zentrum für Experimentelle Medizin, AG Walzog, Planegg-Martinsried, Germany
‡These authors share joint primary authorship.
§These authors share joint senior authorship.