 Sabine Mayer1
Sabine Mayer1 Rebecca Moeller1†
Rebecca Moeller1† João T. Monteiro1Kerstin Ellrott2,3
João T. Monteiro1Kerstin Ellrott2,3 Christine Josenhans2,3,4,5*
Christine Josenhans2,3,4,5* Bernd Lepenies1*
Bernd Lepenies1*
- 1Immunology Unit and Research Center for Emerging Infections and Zoonoses, University of Veterinary Medicine, Hannover, Germany
- 2Medical School Hannover, Institute for Medical Microbiology, Hannover, Germany
- 3German Center for Infection Research (DZIF), Partner Site Hannover–Braunschweig, Germany
- 4Max von Pettenkofer Institute, Ludwig Maximilian University Munich, Munich, Germany
- 5German Center for Infection Research (DZIF), Partner Site Munich, Germany
C-type lectin receptors (CLRs) are carbohydrate-binding receptors that recognize their ligands often in a Ca2+-dependent manner. Upon ligand binding, myeloid CLRs in innate immunity trigger or inhibit a variety of signaling pathways, thus initiating or modulating effector functions such as cytokine production, phagocytosis, and antigen presentation. CLRs bind to various pathogens, including viruses, fungi, parasites, and bacteria. The bacterium Campylobacter jejuni (C. jejuni) is a very frequent Gram-negative zoonotic pathogen of humans, causing severe intestinal symptoms. Interestingly, C. jejuni expresses several glycosylated surface structures, for example, the capsular polysaccharide (CPS), lipooligosaccharide (LOS), and envelope proteins. This “Methods” paper describes applications of CLR–Fc fusion proteins to screen for yet unknown CLR/bacteria interactions using C. jejuni as an example. ELISA-based detection of CLR/bacteria interactions allows a first prescreening that is further confirmed by flow cytometry-based binding analysis and visualized using confocal microscopy. By applying these methods, we identified Dectin-1 as a novel CLR recognizing two selected C. jejuni isolates with different LOS and CPS genotypes. In conclusion, the here-described applications of CLR–Fc fusion proteins represent useful methods to screen for and identify novel CLR/bacteria interactions.
Introduction
C-type lectin receptors (CLRs) are pattern recognition receptors and are known to sense pathogen-associated molecular patterns as well as danger-associated molecules. Upon ligand recognition, CLRs trigger a variety of functions, including the production of inflammatory mediators, the phagocytosis of pathogens, or intracellular signaling (1, 2). The carbohydrate recognition domain (CRD) mediates the binding of CLRs to their specific ligands. One well-described example for a CLR–ligand pair is the CLR Mincle and its ligand trehalose-6,6'-dimycolate (TDM), a unique glycolipid present in the cell wall of mycobacteria (3, 4). Crystal structure analyses of the bovine (5) and human (6) Mincle CRD revealed that the two glucose moieties and one acyl chain of TDM and its synthetic analog trehalose-6,6'-dibehenate interact with Mincle. However, for the majority of CLRs, their glycan ligands and binding mode to their respective ligands are still incompletely understood.
The main function of CLRs is the recognition of highly conserved glycans and glycoproteins located on the surface of pathogens including viruses, parasites, fungi, and bacteria (3, 7–9). However, CLRs may also sense endogenous danger signals released by damaged and necrotic cells (10–13). Dectin-1 is a CLR that is predominantly expressed by monocytes, dendritic cells (DCs), and macrophages (14). It has been described to bind to β-1,3-glucans present in the cell wall of several fungal pathogens. Dectin-1 was shown to recognize various fungal pathogens such as Pneumocystis carinii, Candida albicans, Aspergillus fumigatus, and Cryptococcus neoformans (15).
Fc fusion proteins are established tools to identify novel receptor–ligand interactions. To date, CLR–Fc fusion proteins have been successfully used to screen for novel CLR/pathogen interactions, as demonstrated for fungal interactions, such as the recognition of P. carinii by Mincle (16). In addition, several previously unknown CLR/bacteria interactions were identified using CLR–Fc fusion proteins, including the Mincle/Streptococcus pneumoniae (S. pneumoniae) interaction (17) or the SIGNR3/Lactobacillus acidophilus interaction (18). Moreover, CLEC5A (MDL-1) was found to be an important receptor for Listeria monocytogenes that impacts macrophage and neutrophil functions in Listeria-induced innate immunity (19). Recently, CLR–Fc fusion proteins were used to identify novel CLRs that interact with mycobacteria. Here, CLEC9A was identified as a receptor that was crucial for the regulation of signal transduction and cytokine production during Mycobacterium tuberculosis infection (20). Besides the identification of pathogen-derived CLR ligands, CLR–Fc fusion proteins also allow to screen carbohydrate libraries for novel CLR ligands. Identified ligands can then be further evaluated for their utility to target CLRs on antigen-presenting cells (21–24). Indeed, the ligation of cell surface CLRs can induce various responses such as phagocytosis, cell adhesion, cytokine and chemokine release, as well as antigen presentation (25), rendering CLRs promising targets for immune modulation.
The enteropathogen Campylobacter jejuni (C. jejuni) expresses various virulence factors that allow for motility, adhesion, and invasion of host tissue, leading not only to acute self-limiting gastrointestinal illness but also to autoimmune disorders like Guillain–Barré syndrome (26, 27). Diarrheal Campylobacter species can colonize the intestines of many different host species, ranging from farm animals such as cattle and chicken to humans. Interestingly, they persistently colonize most nonhuman species without overt symptoms, verging on commensalism, while they cause acute intestinal disease in humans (28). C. jejuni is genetically quite variable and exists as generalist variants able to colonize various host species equally well (29, 30) and as specialist variants, which usually have only one preferred host species (31). In addition, each individual C. jejuni strain has the intrinsic property to vary its phenotype, for instance, by phase variation and contingency genes (32, 33). Campylobacter sp. are the only bacteria to date which express functional N- and O-glycosylation modules (34). Surface-exposed glycolipids such as the capsular polysaccharide (CPS) (35) and the lipooligosaccharide (LOS) (36) of the outer membrane play a pivotal role in host interaction and evasion by C. jejuni. In addition, C. jejuni expresses several cell-envelope-located O- and N-linked glycoproteins (37, 38). Recent studies identified CLRs that are involved in C. jejuni recognition (39, 40). In one study, hMGL–Fc was shown to interact with C. jejuni-derived glycoproteins (39). Another study used a murine CLR–hFc fusion protein library and showed LMIR5 to bind to C. jejuni (40). These studies point to a role of CLRs in host interplay and modulating the host immune response against C. jejuni.
This “Methods” paper presents a combination of innovative techniques to screen for and study CLR/bacteria interactions, using C. jejuni as a representative example. All applied methods are based on CLR–hFc fusion proteins in which the extracellular part of the respective murine (m) or human (h)CLR containing the CRD has been fused to the Fc fragment of human IgG1 molecules, thus leading to dimer formation. ELISA-based methods allow for a high-throughput prescreening for potential CLR interactions with bacteria, followed by flow-cytometric analyses of identified candidates as a confirmatory method. To visualize and confirm binding of CLRs to bacteria in situ, confocal microscopy can be applied and was used in this study to visualize the binding of Dectin-1 to C. jejuni.
Materials and Methods
C. jejuni Strains, Culture Conditions, and Preparation of Bacteria for Interaction Studies
Campylobacter jejuni strains used were from two strain collections (29, 41) assembled in Germany between 2011 and 2016. We selected two different, but related, generalist strains that are from two frequent C. jejuni lineages that can colonize well with various animal species including humans and that cause frequent diarrheal diseases in the latter. Strain MHH-24 is equivalent to isolate FBI-Zoo 06025 (ST22) from Ref. (29) and was isolated from raw milk (cattle), and strain MHH-19, a human enteritis isolate, is equivalent to isolate FBI-Zoo 07079 (ST19) from the more recent case–control study (41). MHH-19 has well-recognized genetic determinants for capsule and LOS types (own unpublished data); however, strain MHH-24 has not been typeable so far by molecular methods for LOS or CPS (own unpublished data). Both strains’ LOS and CPS glycans or other surface glycans have not been characterized biochemically so far. For the plate assay and FACS-based assay, heat-fixed bacteria were used, which allow for better staining of the bacteria with the fluorescent Syto61 dye (Thermo Scientific), while for immunofluorescent individual bacterial labeling, PFA-fixed bacteria were prepared, which permit a superior surface preservation of the cells and better storage capacity. Bacteria were grown on blood agar plates (Columbia agar, supplemented with 5% sheep blood, Oxoid, Germany) and diluted in sterile 1x PBS at an OD600 of 1. Heat fixation was performed at 65°C in a heating block for 5 h. Heat-inactivated bacteria were stored at 4°C for a maximum of 3 weeks. Fresh 2% PFA as an alternative fixing agent for immunofluorescent labeling was prepared in 100 mM sterile-filtered potassium phosphate buffer, pH = 7.0, and bacteria were fixed twice for 1 h at room temperature (RT), with centrifugation (6.000 × g, 10 min, RT) and one change of fixing agent in between. Afterward, the bacteria were centrifuged again and resuspended in a sterile solution of 0.1% glycine in PBS to quench the fixing agent, which prevents nonspecific attachment of proteins or cell clumping. Ultimately, the bacteria were resuspended in pure, sterile 1× PBS (pH = 7.4) after a final centrifugation step and stored at 4°C until further use, with a high storage capacity of several months.
Generation of CLR–hFc Fusion Proteins
The production of the CLR–hFc fusion proteins was performed as previously described (23). Briefly, RNA was isolated from murine spleen and reverse-transcribed into cDNA using a reverse transcriptase (New England Biolabs, Ipswich, MA, USA). Polymerase chain reaction was applied to amplify the cDNA encoding the extracellular part of each CLR using specific primers (Table 1). The respective cDNA fragments were ligated into a pFuse-hIgG1-Fc expression vector (InvivoGen, San Diego, CA, USA). Next, CHO-S cells were transiently transfected with the vector construct using MAX reagent (InvivoGen). CLR–hFc fusion proteins were purified after 4 days of transfection from the cell supernatant using HiTrap protein G HP columns (GE Healthcare, Piscataway, NJ, USA). To confirm the purity of each CLR–hFc fusion protein, the protein was analyzed by dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and subsequent Coomassie staining as well as Western blot using an anti-human IgG- horseradish peroxidase (HRP) antibody (Dianova, Hamburg, Germany).
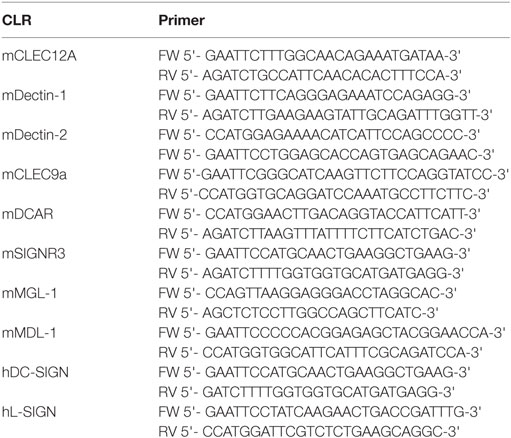
Table 1. DNA sequences of primers used for amplification of the extracellular domain of the respective CLRs.
Western Blot
After protein separation using SDS-PAGE, the proteins were transferred to a nitrocellulose membrane for 1 h at 5 V. The membrane was blocked for 1 h with 5% milk powder in TBS and 0.1% Tween-20 (TBS-T) followed by a 1-h incubation with an anti-human IgG antibody conjugated to HRP (Dianova). The membrane was washed three times with TBS-T, for 5 min each. Detection of the CLR–hFc fusion proteins was performed using the Amersham ECL Western blotting detection reagent (GE Healthcare).
ELISA-Based Binding Studies
A half-area microplate (Greiner Bio-One GmbH, Frickenhausen, Germany) was coated with 3 × 108 CFU/ml heat-inactivated C. jejuni for 3.5 h at RT. Non-adherent bacteria were washed away, and the plate was blocked with buffer containing 1% BSA (Thermo Fisher Scientific/Invitrogen, Darmstadt, Germany) in 1x PBS for 2 h at RT. After washing the wells, 200 ng of each respective CLR–hFc fusion protein in lectin-binding buffer (50 mM HEPES, 5 mM MgCl2, and 5 mM CaCl2) was added to the bacteria and incubated for 1 h at RT. Then, a 1:5.000-diluted HRP-conjugated goat anti-human IgG antibody (Dianova) was added for 1 h at RT. Finally, the substrate solution [o-phenylenediamine dihydrochloride substrate tablet (Thermo Fisher Scientific), 24 mM citrate buffer, 0.04% H2O2, 50 mM phosphate buffer in H2O] was added to the samples, and the reaction was stopped with 2.0 M sulfuric acid. Data was collected using a Multiskan Go microplate spectrophotometer (Thermo Fisher Scientific) at a wavelength of 495 nm. Four independent experiments were performed with technical triplicates each.
Flow Cytometry-Based Binding Studies
To detect the bacteria and exclude them from debris, 3–6 × 107 CFU/ml heat-inactivated C. jejuni were stained with 1 µM of the DNA-staining dye Syto61 (Thermo Fisher Scientific) and incubated for 30 min at RT. Subsequently, samples were incubated for 1 h with 200 ng of the respective CLR–hFc fusion protein in lectin-binding buffer. After washing once with lectin-binding buffer, the bacterial pellet was stained with a PE-conjugated goat anti-human Fc (Dianova) antibody solution and incubated for 25 min at 4°C. Finally, flow-cytometric analysis was performed using an Attune NxT Flow Cytometer (Thermo Fisher Scientific). Data analysis was performed using the FlowJo Software (FlowJo, Ashland, OR, USA). As a control, hFc protein was used to exclude the nonspecific binding of C. jejuni to the Fc part of the CLR–Fc fusion proteins. Besides the use of the hFc protein, the secondary antibody alone served as an additional negative control. Three independent experiments were done with technical duplicates each.
Confocal Fluorescence Microscopy-Based Binding Studies
Cover slides (Thermo Fisher Scientific) were cleaned with 70% ethanol and coated with poly-L-lysine solution (Sigma-Aldrich, St. Louis, MO, USA) for 30 min at 60°C. 6 × 107 CFU/ml C. jejuni isolate MHH-19 fixed with 2% PFA was washed with 1x PBS and incubated overnight (o.n.) with 500 ng Dectin-1–hFc and hFc in lectin-binding buffer at 4°C. After washing two times with lectin-binding buffer, samples were incubated for 2 h with 1:200-diluted goat anti-human Fc Alexa Fluor (AF) 488-conjugated antibody (Dianova) at 4°C. Next, samples were washed with 1x PBS, applied onto poly-L-lysine-coated cover slides, and incubated for 45 min at 37°C. In addition, a sample with C. jejuni and the secondary antibody only was used as a negative control. Finally, the cover slides were mounted on microscopic slides (Roth, Karlsruhe, Germany) with proLong™ gold antifade mountant containing DAPI (Thermo Fisher Scientific), sealed and visualized using a TCS SP5 confocal inverted-base fluorescence microscope (Leica, Nussloch, Germany) equipped with a HCX PL APO 63 × 1.4 oil immersion objective. To avoid the detection of artifacts, PFA-fixed bacterial samples were inspected visually by a high-magnification microscopy (100× lens magnification) for clumps before performing hFc fusion protein co-incubation. Only bacterial preparations without any visible clumps were further used. Three independent experiments were performed, each with three randomly selected pictures.
Statistical Analysis
All data are presented as mean ± SD. Unpaired, one-tailed Student’s t-test was applied to determine the significance between CLR candidates and the hFc control. Data were analyzed using the GraphPad Prism software (version 7.02).
Results
Generation and Detection of CLR–hFc Fusion Proteins Used in This Study
The generation of CLR–hFc fusion proteins required several steps (Figure 1A, 1–4). The first step was the cloning of the cDNA fragment encoding for the extracellular part of each CLR (containing the CRD) and its fusion to the Fc fragment of human IgG1 in the pFuse-hIgG1-Fc expression vector (1). Next, mammalian CHO-S cells were transfected with this vector construct (2). The use of a mammalian cell line such as CHO-S cells ensured that soluble CLR–hFc fusion proteins were secreted into the supernatant that carried mammalian-type glycosylation. Finally, the supernatant was harvested, followed by purification of the respective fusion proteins using protein G columns (3). To confirm the presence and purity of the respective CLR–hFc fusion proteins after purification, SDS-PAGE and subsequent Coomassie staining and a Western blot were performed (Figure 1B). Bands at the expected size of the respective CLR–hFc fusion protein showed the presence of each CLR–hFc fusion protein.
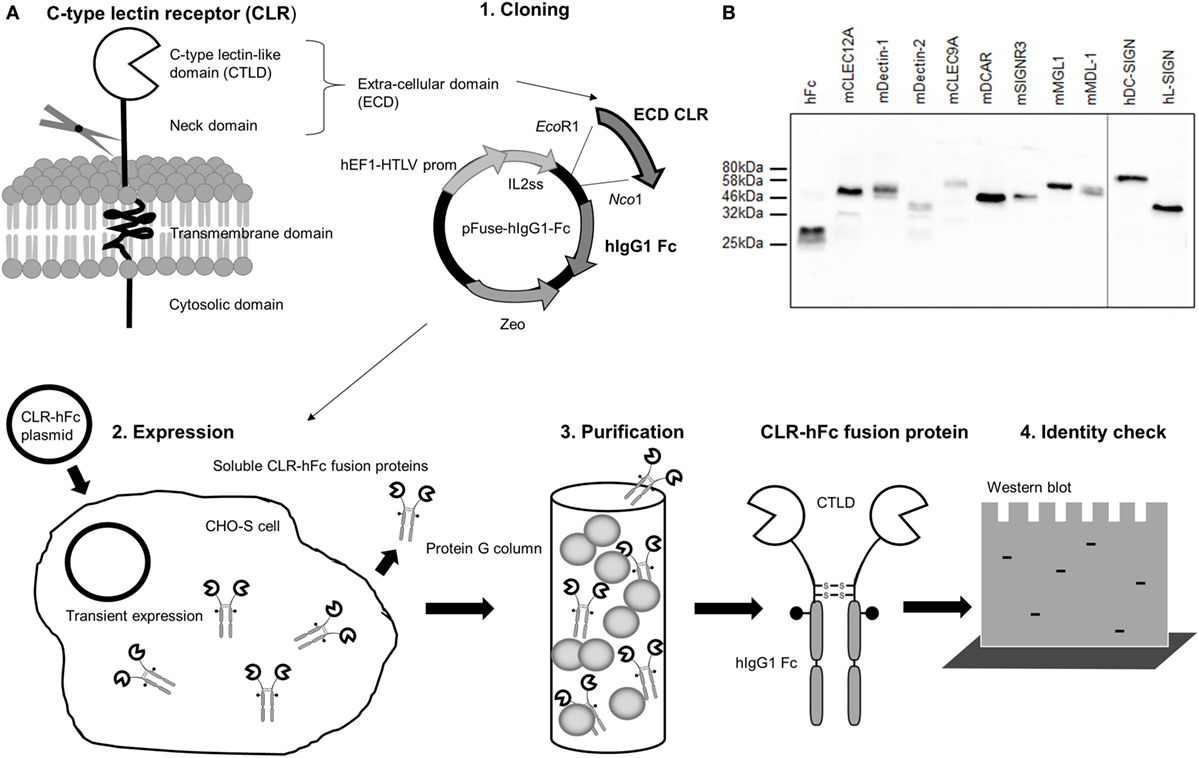
Figure 1. (A) Overview of the generation and production of C-type lectin receptor (CLR)–hFc fusion proteins. (1) First, the cDNA encoding for the carbohydrate recognition domain of the respective CLR is cloned into the expression vector and thereby fused to the cDNA fragment encoding for human IgG1-Fc. (2) CHO-S cells are transiently transfected with the expression vector construct, and the respective soluble CLR–hFc fusion proteins are secreted. (3) The cell supernatant is purified, and (4) the presence of the CLR–Fc fusion proteins is detected. (B) Western blot of selected CLR–hFc fusion proteins. To confirm the successful CLR–hFc fusion protein production, supernatants of transiently transfected cells were purified using protein G columns, and detection by Western blot was performed using a horseradish peroxidase-conjugated anti-hFc antibody.
Prescreening Using an ELISA-Based Assay
In a first screening, the binding of the CLR–hFc fusion proteins to two different C. jejuni isolates (MHH-19 and MHH-24) was tested using an ELISA-based method. After immobilization of the heat-inactivated bacteria on the ELISA plate and incubation with the respective CLR–hFc fusion proteins, their interaction with the C. jejuni isolates was determined by colorimetric detection (Figure 2A). Several controls were included, such as the incubation of the CLR–hFc fusion proteins on non-coated wells (data not shown) or with the hFc protein alone to exclude unspecific binding of the Fc fragment to C. jejuni. In general, the CLR–hFc fusion proteins exhibited a similar binding pattern to both C. jejuni isolates (Figure 2B). No binding was observed for DCAR–hFc and L-SIGN–hFc. All other CLR–hFc fusion proteins displayed weak to strong binding to C. jejuni in the ELISA-based assay and were considered as potential receptors for the C. jejuni isolates MHH-19 and MHH-24.
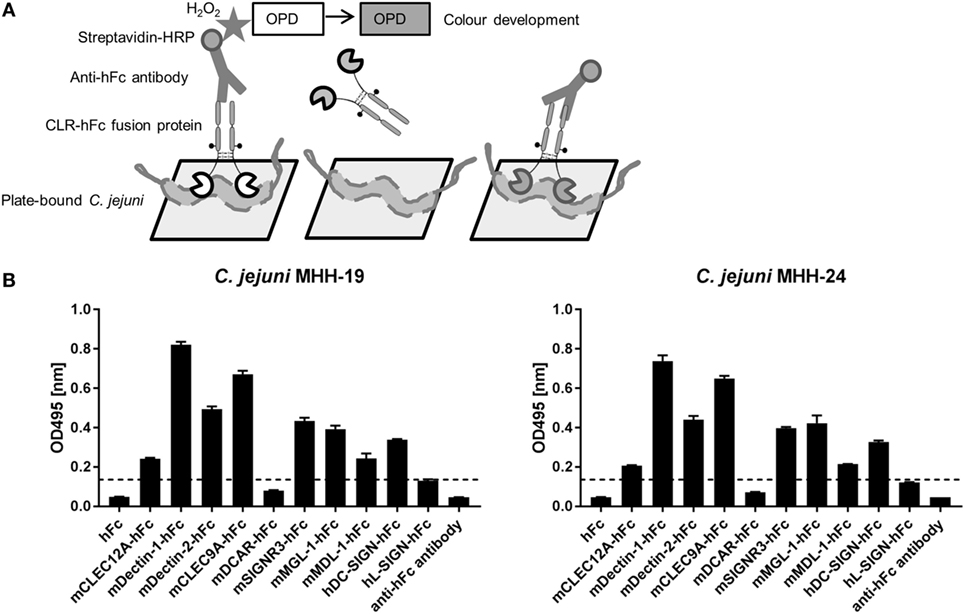
Figure 2. (A) Schematic representation of the ELISA-based-binding study using the C-type lectin receptor (CLR)–hFc fusion proteins. Immobilized C. jejuni was incubated with the respective CLR–hFc fusion proteins. The detection of bound fusion proteins was performed using a horseradish peroxidase (HRP)-conjugated anti-hFc antibody and subsequent colorimetric detection (details are given in Section “Materials and Methods”). (B) Several CLR–hFc fusion proteins were analyzed for their interaction with heat-inactivated immobilized C. jejuni. This prescreening showed no or only marginal binding of DCAR–hFc and L-SIGN–hFc. The CLR–hFc fusion proteins CLEC9A, DC-SIGN, Dectin-1, Dectin-2, MDL-1, MGL-1, CLEC12A, and SIGNR3 exhibited weak to marked binding to both C. jejuni isolates. Representative data from one out of four independent experiments are shown (technical triplicates for each condition). The dashed line represents the defined threshold for CLR/bacteria interactions in the ELISA-based assay. Student’s t-test was performed to compare all CLR–hFc fusion proteins with absorbance above the threshold to the hFc control alone. For both C. jejuni isolates, a highly significant binding (****p ≤ 0.0001) was observed for the CLR–hFc fusion proteins mCLEC12A–hFc, mDectin-1–hFc, mDectin-2–hFc, mCLEC9A–hFC, mSIGNR3–hFC, mMGL-1–hFc, mMDL-1–hFc, and hDC-SIGN–hFc.
Confirmatory Test Using a Flow Cytometry-Based Assay
To verify and extend the results from the ELISA-based detection method, a flow cytometry-based protocol to screen for CLR/bacteria interactions in solution was established. To this end, C. jejuni was incubated with CLR–hFc fusion proteins, and subsequent binding was detected upon staining with a PE-conjugated anti-hFc antibody (Figure 3A). The gating strategy is displayed in Figure 3B and is based on the gating of bacteria in the forward-scatter/side-scatter plot, followed by gating on Syto61-positive events. Incubation with the Dectin-1–hFc fusion protein led to a marked shift in the fluorescence intensity, indicating the binding of Dectin-1–hFc to C. jejuni. For both C. jejuni isolates, no binding was observed for staining with the hFc fragment or with the secondary antibody alone. The lack of binding of the hFc fragment to C. jejuni illustrates the specificity of the recognition of the C. jejuni isolates by Dectin-1–hFc. The analysis of the binding studies shows that both tested C. jejuni isolates were significantly recognized by Dectin-1–hFc and, to a lesser extent, by CLEC12A–hFc (Figure 3C). All other CLR–hFc fusion proteins included in the flow cytometry-based assay exhibited no or only marginal binding to both C. jejuni isolates. Interestingly, no binding of CLEC9A–hFc to both C. jejuni isolates was detected using the flow cytometry-based assay. This finding may either suggest a false-positive result in the ELISA-based assay or may be due to internal C. jejuni ligands that are not accessible in the flow cytometry-based assay. In summary, the CLR–hFc fusion proteins exhibited a similar binding profile to both C. jejuni isolates in the flow cytometry-based assays and revealed Dectin-1 as a novel candidate receptor for C. jejuni recognition.
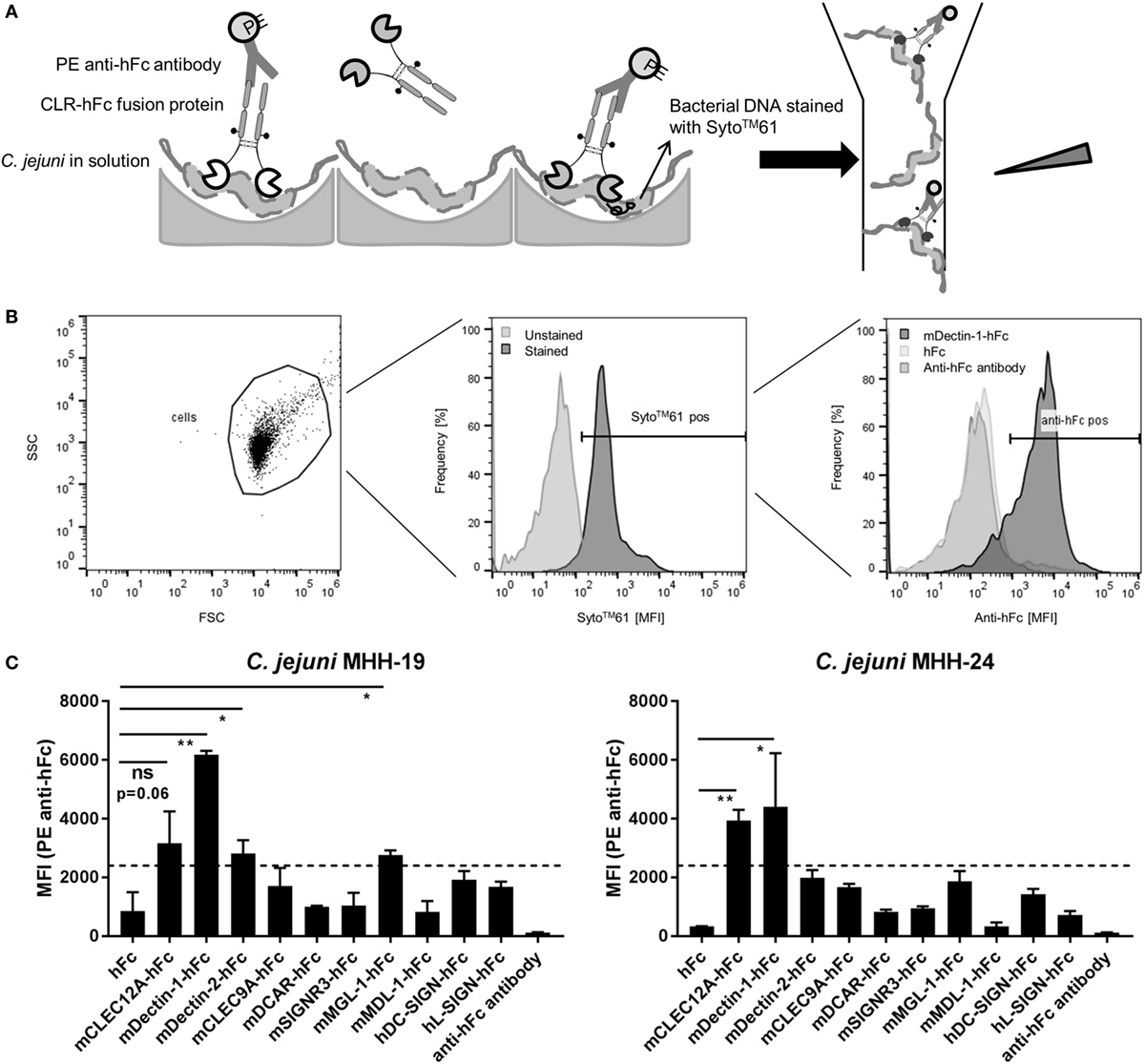
Figure 3. (A) Overview of the flow cytometry-based protocol to detect bacterial recognition by C-type lectin receptor (CLR)–hFc fusion proteins. C. jejuni was incubated with the respective CLR–hFc fusion proteins in solution. Binding was detected using an anti-hFc antibody. Subsequently, samples were analyzed by flow cytometry based on their morphological and fluorescence properties. (B) Histoplots show the gating strategy to detect the binding of CLR–hFc fusion proteins to C. jejuni in a representative experiment. Cells were discriminated from debris by gating on forward scatter (FSC) and side scatter (SSC). C. jejuni was further gated on DNA staining, indicated by Syto61 positivity (pos.). Finally, Syto61-positive bacteria were analyzed for Dectin-1–hFc binding. hFc and the secondary antibody alone were used as controls. (C) Representative data from one out of three independent experiments of the flow cytometry-based CLR–hFc-binding studies to the C. jejuni isolates MHH-19 and MHH-24 (technical duplicates each). Similar binding profiles for both isolates were observed. While Dectin-1–hFc and CLEC12A–hFc exhibited binding to C. jejuni, all other tested CLR–Fc fusion proteins displayed marginal to no binding. The dashed line represents the defined threshold for CLR/bacteria interactions in the flow cytometry-based assay. Data are expressed as mean ± SD. Student’s t-test was performed to compare the binding of CLR–Fc fusion proteins above the threshold to hFc alone. For all statistical analyses, p-values of <0.05 were considered to be significant: ns = not significant, *p ≤ 0.05, **p ≤ 0.01.
Visualization of CLR/Bacteria Interactions Using Confocal Microscopy
To visualize the identified Dectin-1–hFc interaction with C. jejuni in situ, confocal microscopy after immunofluorescent labeling was applied. The incubation of C. jejuni with Dectin-1–hFc and hFc control protein was performed in solution. Subsequently, bacteria were immobilized on poly-L-lysine-coated cover slides, and individual CLR/C. jejuni interactions were visualized using a confocal laser-scanning microscope (Figure 4A). The results confirmed Dectin-1 binding to C. jejuni, whereas no binding was observed for hFc (Figure 4B) and the secondary antibody alone (data not shown). Furthermore, merging the fluorescence channels and the differential interference contrast showed that the Dectin-1–hFc signal colocalized with patches on the bacterial cell periphery. This finding suggests that Dectin-1–hFc recognizes a cell-envelope component of C. jejuni. In conclusion, the combination of ELISA-, flow cytometry-, and confocal microscopy-based methods highlights the utility of CLR–Fc fusion proteins to identify novel CLR/bacteria interactions as demonstrated here using C. jejuni as an example. The functional role of the C. jejuni recognition by Dectin-1 can now be further elucidated in future studies.
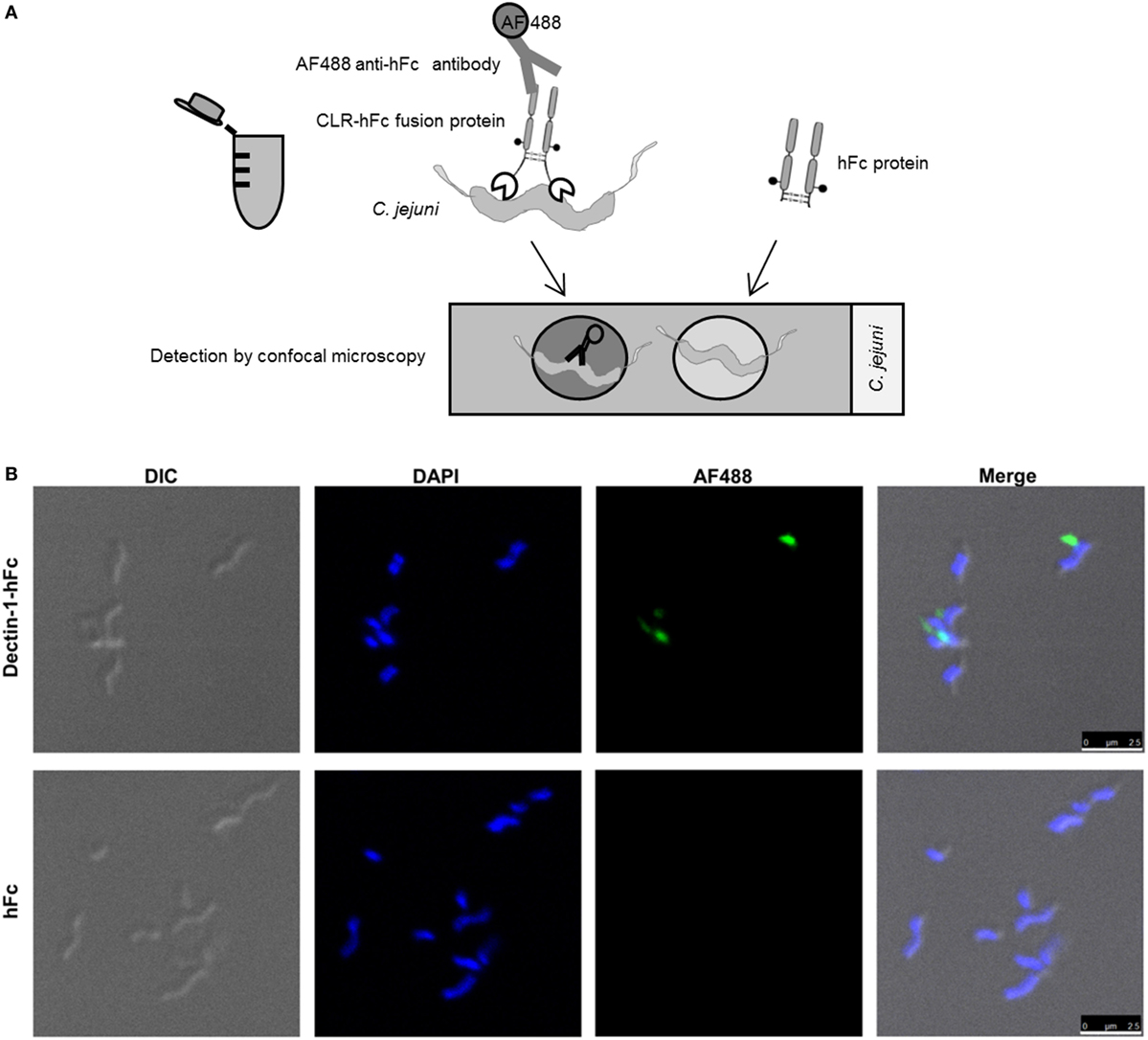
Figure 4. (A) Schematic representation of the C-type lectin receptor (CLR)–hFc fusion protein-binding studies using confocal microscopy. Bacteria were incubated with the CLR–hFc fusion proteins. Bound CLR–hFc fusion proteins were detected by an Alexa Fluor (AF) 488-conjugated anti-hFc antibody. To exclude unspecific binding, bacteria were also incubated with hFc as control protein or with the secondary antibody alone. Bacteria were transferred onto cover slips, mounted onto microscopic slides, and visualized using confocal microscopy. (B) Bacteria were incubated with Dectin-1–hFc (upper panels) or hFc protein (negative control, lower panels). Differential interference contrast (DIC) was included to visualize the bacterial cell wall. Bacterial DNA was stained with DAPI (shown in blue). The binding of the Dectin-1–Fc fusion protein to C. jejuni was detected using an AF488-conjugated secondary antibody (shown in green). While Dectin-1–hFc recognizes C. jejuni, the hFc control and the secondary antibody alone showed no signal in the AF488 channel. The experiment was repeated three times with three randomly selected pictures each, and similar results were revealed. Scale bar indicates 2.5 µm.
Discussion
This article presents three distinct methods to detect and verify novel CLR/bacteria interactions. Often, the identification of novel CLR/pathogen interactions is the first step to unravel the interplay of the host innate immune system with bacterial pathogens. Each of the methods can be applied for different purposes and has certain advantages and drawbacks (presented in Table 2). The ELISA-based method allows for a high-throughput screening of bacteria collections. Due to the possibility of false-positive results caused by protein aggregation on the ELISA plate, this method is mainly suitable for an initial prescreening using the whole CLR–hFc fusion protein library and requires confirmation by additional methods. To confirm initially identified CLR/bacteria interactions, flow-cytometric analysis represents a useful method that has several advantages. First, binding takes place in solution, thus avoiding protein aggregation on the ELISA plate. Second, the flow cytometry-based method offers the possibility to discriminate between bacteria and debris using an appropriate gating strategy. Third, it allows for narrowing down the localization of ligands to the bacterial surface, whereas the ELISA-based method may lead to a partial lysis of bacteria, thus releasing internal ligands. By contrast, confocal (fluorescence) microscopy offers the opportunity of visualizing CLR/bacteria interactions for single-bacteria in situ, thus enabling colocalization studies to further characterize the bacterial ligand. All described methods can be easily applied to other Gram-positive and Gram-negative bacterial species.

Table 2. Advantages and drawbacks of the ELISA-based, flow cytometry-based, and confocal microscopy-based methods to detect novel CLR/bacteria interactions.
Campylobacter jejuni is an interesting candidate for screening glycan-binding factors, since the bacteria are heavily glycosylated with various different glycan species and possess active genes for variable O- and N-glycosylation, providing abilities to glycosylate capsule, LOS, and proteins (34). In addition, strain-specific differences between various C. jejuni strains exist concerning surface determinants and glycosylation (42), based, for instance, on strain-specific genetic differences and phase variation (32). Even in each individual C. jejuni strain, a high-variation potential of the bacterial surface phenotype, for instance, LOS, capsule, or additional LOS glycosylation, exists (33, 43–45), which provides an interesting field of future study. Applying the CLR–hFc fusion protein library to screen for binding to preselected, molecularly typed C. jejuni isolates from two frequent generalist lineages, we identified Dectin-1–hFc as a promising candidate receptor for C. jejuni. Phongsisay et al. screened C. jejuni lysates using a murine CLR–hFc fusion protein library (40). In this previous study, the murine CLR LMIR5 was described to interact with C. jejuni, whereas other tested CLR/C. jejuni interactions remained negative. In bacterial lysates, components such as glycolipids are released and better accessible to potential receptors as in intact live or heat-inactivated bacteria. Thus, the CLR/C. jejuni interactions identified in the respective study may also include internal ligands that are not detected when intact bacteria are immobilized on the plate for ELISA-based detection or used in solution for the flow cytometry-based assay. In addition, the use of different C. jejuni strains or growth under different culture conditions may impact the recognition by CLRs. Since the surface interaction of the bacteria with host lectins might be more relevant for the colonization and infection process in vivo, we employed heat-inactivated intact bacterial cells instead of bacterial lysates for our screening procedures. In our present study, both isolates used in our study, which are genetically related, but not identical, exhibited a similar CLR-binding profile. It will be interesting to compare more C. jejuni strains including distantly related isolates, generalists, and specialists (30, 31) for lectin binding. In addition, phase variation, which is a common genetic mechanism used by C. jejuni to modulate its surface properties (32), may play a role in CLR recognition. Indeed, phase variation between strains and within the population of one C. jejuni isolate (33) may affect cell wall components, as has already been shown for the glycosylation of LOS (43), capsule (44, 45), and for other bacterial properties (32). In this context, it is worth noting that in our confocal microscopy in situ approach, bacterial cells showed an individual variation in Dectin-1 binding or the absence of binding. Phenotypical variation of this trait within the bacterial population has not been revealed in any earlier study on C. jejuni and might be explained by single-cell variation of a Dectin-1-binding surface determinant. This presents a very interesting opportunity for further study of individual bacterial intra-strain variation. In an earlier study, human MGL was shown to recognize C. jejuni through binding to C. jejuni-derived N-glycosylated proteins (39). For murine MGL-1, we observed only weak binding to C. jejuni, which may be due to experimental or strain differences, or to the different binding profiles between murine and human MGL isoforms. While two different orthologs (mMGL-1 and mMGL-2) are found in mice, humans only express one MGL isoform (hMGL). It is known that mMGL-2 displays a similar binding profile as hMGL which may account for the marginal binding observed for the mMGL-1 ortholog in our study (46, 47).
In this study, we have identified mouse Dectin-1 as a candidate receptor for the innate recognition of C. jejuni. To date, Dectin-1 has mainly been described as CLR-recognizing fungal pathogens. For instance, Dectin-1 binds to C. albicans, A. fumigatus, and C. neoformans (15). The Dectin-1 ligand recognized in the context of fungal infection is β-1,3-glucan (48), present in the cell wall of several fungi. Nevertheless, also parasites such as Leishmania infantum (49, 50) and P. carinii (51) were described to be sensed by Dectin-1. Interestingly, also an interaction of Dectin-1 in cooperation with TLR2 was shown for several Mycobacterium species (52). In C. jejuni, α-1,4-glucan has been reported as a capsule component (53).
The identification of candidate CLRs that play a role in bacterial recognition presents the first step to identify a distinct bacterial ligand for the respective receptor and may help to understand the interaction of bacteria with the host innate immune system. Identified CLR candidates can be further investigated for their relevance in vitro and in vivo. Using a comprehensive CLR–hFc library, Rabes et al. demonstrated that Mincle recognizes S. pneumoniae in a Ca2+-dependent manner (17). This work was extended by a recent study showing that Mincle recognizes S. pneumoniae-derived glucosyl-diacylglycerol in a serotype-specific fashion (54). To date, several CLR ligands have been identified by the use of CLR–Fc fusion proteins. For instance, one study revealed Mincle–hFc as a receptor sensing mannose and glucose-rich glycolipids extracted from Malassezia pachydermatis (55). In addition, the identification of distinct glycan ligands of CLRs offers the possibility for glycan-based CLR targeting to deliver vaccine antigens into antigen-presenting cells and to induce subsequent adaptive immune responses (22, 56–58). This approach has already been applied successfully to vaccine design using carbohydrate-based adjuvants (22, 59, 60). Besides CLR–Fc fusion protein libraries, reporter cell lines expressing the respective CLR are used to identify novel CLR–pathogen interactions and CLR ligands (61, 62). In addition, such reporter cell lines also allow for investigating if the identified CLR ligands act as potential agonists or antagonists. In conclusion, this “Methods” paper combines three different screening and confirmatory methods for the detection of CLR–hFc fusion protein binding by pathogens. It also highlights the utility of CLR–hFc fusion proteins to screen for novel CLR/bacteria interactions as a first step toward the identification of distinct bacterial CLR ligands and characterization of their biological functions.
Author Contributions
SM, CJ and BL designed the research; SM and RM performed the research; JM and KE contributed to new reagents/analytical tools; SM, RM, CJ, and BL analyzed data; and SM, CJ, and BL wrote the paper with the help of the other authors.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
BL acknowledges funding from the European Union’s Horizon 2020 Research and Innovation Program (Marie Sklodowska-Curie grant agreement No. 642870, ETN-Immunoshape) and the Niedersachsen-Research Network on Neuroinfectiology (N-RENNT-2). Previously, funding from the Collaborative Research Center (SFB) 765 was crucial to the research program of BL. The work in the laboratory of CJ was supported by project grant B6 within the CRC 900 Collaborative Research Center grant funded by the German Research Foundation, and by the German Center for Infection Research (DZIF), project 01.806. Silke Schöneberg, Birgit Brenneke, and Friederike Kops are acknowledged for technical assistance.
References
1. Hou H, Guo Y, Chang Q, Luo T, Wu X, Zhao X. C-type lectin receptor: old friend and new player. Med Chem (2017) 13(6):536–43. doi:10.2174/1573406413666170510103030
2. Mayer S, Raulf M-K, Lepenies B. C-type lectins: their network and roles in pathogen recognition and immunity. Histochem Cell Biol (2017) 147(2):223–37. doi:10.1007/s00418-016-1523-7
3. Ishikawa E, Ishikawa T, Morita YS, Toyonaga K, Yamada H, Takeuchi O, et al. Direct recognition of the mycobacterial glycolipid, trehalose dimycolate, by C-type lectin Mincle. J Exp Med (2009) 206(13):2879–88. doi:10.1084/jem.20091750
4. Schoenen H, Bodendorfer B, Hitchens K, Manzanero S, Werninghaus K, Nimmerjahn F, et al. Cutting edge: Mincle is essential for recognition and adjuvanticity of the mycobacterial cord factor and its synthetic analog trehalose-dibehenate. J Immunol (2010) 184(6):2756–60. doi:10.4049/jimmunol.0904013
5. Feinberg H, Jegouzo SA, Rowntree TJ, Guan Y, Brash MA, Taylor ME, et al. Mechanism for recognition of an unusual mycobacterial glycolipid by the macrophage receptor Mincle. J Biol Chem (2013) 288(40):28457–65. doi:10.1074/jbc.M113.497149
6. Furukawa A, Kamishikiryo J, Mori D, Toyonaga K, Okabe Y, Toji A, et al. Structural analysis for glycolipid recognition by the C-type lectins Mincle and MCL. Proc Natl Acad Sci U S A (2013) 110(43):17438–43. doi:10.1073/pnas.1312649110
7. Schlesinger LS, Hull SR, Kaufman TM. Binding of the terminal mannosyl units of lipoarabinomannan from a virulent strain of Mycobacterium tuberculosis to human macrophages. J Immunol (1994) 152(8):4070–9.
8. Gringhuis SI, Wevers BA, Kaptein TM, van Capel TM, Theelen B, Boekhout T, et al. Selective C-Rel activation via malt1 controls anti-fungal T(H)-17 immunity by Dectin-1 and Dectin-2. PLoS Pathog (2011) 7(1):e1001259. doi:10.1371/journal.ppat.1001259
9. Monteiro JT, Lepenies B. Myeloid C-type lectin receptors in viral recognition and antiviral immunity. Viruses (2017) 9(3):1–22. doi:10.3390/v9030059
10. Yamasaki S, Ishikawa E, Sakuma M, Hara H, Ogata K, Saito T. Mincle is an ITAM-coupled activating receptor that senses damaged cells. Nat Immunol (2008) 9(10):1179–88. doi:10.1038/ni.1651
11. Ahrens S, Zelenay S, Sancho D, Hanc P, Kjaer S, Feest C, et al. F-actin is an evolutionarily conserved damage-associated molecular pattern recognized by DNGR-1, a receptor for dead cells. Immunity (2012) 36(4):635–45. doi:10.1016/j.immuni.2012.03.008
12. Zhang JG, Czabotar PE, Policheni AN, Caminschi I, Wan SS, Kitsoulis S, et al. The dendritic cell receptor Clec9A binds damaged cells via exposed actin filaments. Immunity (2012) 36(4):646–57. doi:10.1016/j.immuni.2012.03.009
13. Sancho D, Reis e Sousa C. Sensing of cell death by myeloid C-type lectin receptors. Curr Opin Immunol (2013) 25(1):46–52. doi:10.1016/j.coi.2012.12.007
14. Plato A, Willment JA, Brown GD. C-type lectin-like receptors of the Dectin-1 cluster: ligands and signaling pathways. Int Rev Immunol (2013) 32(2):134–56. doi:10.3109/08830185.2013.777065
15. Drummond RA, Brown GD. The role of Dectin-1 in the host defence against fungal infections. Curr Opin Microbiol (2011) 14(4):392–9. doi:10.1016/j.mib.2011.07.001
16. Kottom TJ, Hebrink DM, Jenson PE, Nandakumar V, Wuthrich M, Wang H, et al. The interaction of Pneumocystis with the C-type lectin receptor Mincle exerts a significant role in host defense against infection. J Immunol (2017) 198(9):3515–25. doi:10.4049/jimmunol.1600744
17. Rabes A, Zimmermann S, Reppe K, Lang R, Seeberger PH, Suttorp N, et al. The C-type lectin receptor Mincle binds to Streptococcus pneumoniae but plays a limited role in the anti-pneumococcal innate immune response. PLoS One (2015) 10(2):e0117022. doi:10.1371/journal.pone.0117022
18. Lightfoot YL, Selle K, Yang T, Goh YJ, Sahay B, Zadeh M, et al. SIGNR3-dependent immune regulation by Lactobacillus acidophilus surface layer protein A in colitis. EMBO J (2015) 34(7):881–95. doi:10.15252/embj.201490296
19. Chen ST, Li FJ, Hsu TY, Liang SM, Yeh YC, Liao WY, et al. CLEC5A is a critical receptor in innate immunity against Listeria infection. Nat Commun (2017) 8(1):299. doi:10.1038/s41467-017-00356-3
20. Cheng AC, Yang KY, Chen NJ, Hsu TL, Jou R, Hsieh SL, et al. CLEC9A modulates macrophage-mediated neutrophil recruitment in response to heat-killed Mycobacterium tuberculosis H37Ra. PLoS One (2017) 12(10):e0186780. doi:10.1371/journal.pone.0186780
21. Lepenies B, Lee J, Sonkaria S. Targeting C-type lectin receptors with multivalent carbohydrate ligands. Adv Drug Deliv Rev (2013) 65(9):1271–81. doi:10.1016/j.addr.2013.05.007
22. van Kooyk Y, Unger WW, Fehres CM, Kalay H, Garcia-Vallejo JJ. Glycan-based DC-SIGN targeting vaccines to enhance antigen cross-presentation. Mol Immunol (2013) 55(2):143–5. doi:10.1016/j.molimm.2012.10.031
23. Maglinao M, Eriksson M, Schlegel MK, Zimmermann S, Johannssen T, Gotze S, et al. A platform to screen for C-type lectin receptor-binding carbohydrates and their potential for cell-specific targeting and immune modulation. J Control Release (2014) 175:36–42. doi:10.1016/j.jconrel.2013.12.011
24. Streng-Ouwehand I, Ho NI, Litjens M, Kalay H, Boks MA, Cornelissen LA, et al. Glycan modification of antigen alters its intracellular routing in dendritic cells, promoting priming of T cells. Elife (2016) 5:e11765. doi:10.7554/eLife.11765
25. Hoving JC, Wilson GJ, Brown GD. Signalling C-type lectin receptors, microbial recognition and immunity. Cell Microbiol (2014) 16(2):185–94. doi:10.1111/cmi.12249
26. Blaser MJ, Berkowitz ID, LaForce FM, Cravens J, Reller LB, Wang WL. Campylobacter enteritis: clinical and epidemiologic features. Ann Intern Med (1979) 91(2):179–85. doi:10.7326/0003-4819-91-2-179
27. Caudie C, Quittard Pinon A, Taravel D, Sivadon-Tardy V, Orlikowski D, Rozenberg F, et al. Preceding infections and anti-ganglioside antibody profiles assessed by a dot immunoassay in 306 French Guillain–Barré syndrome patients. J Neurol (2011) 258(11):1958–64. doi:10.1007/s00415-011-6042-9
28. Gölz G, Rosner B, Hofreuter D, Josenhans C, Kreienbrock L, Löwenstein A, et al. Relevance of Campylobacter to public health—the need for a one health approach. Int J Med Microbiol (2014) 304(7):817–23. doi:10.1016/j.ijmm.2014.08.015
29. Gripp E, Hlahla D, Didelot X, Kops F, Maurischat S, Tedin K, et al. Closely related Campylobacter jejuni strains from different sources reveal a generalist rather than a specialist lifestyle. BMC Genomics (2011) 12:584. doi:10.1186/1471-2164-12-584
30. Dearlove BL, Cody AJ, Pascoe B, Meric G, Wilson DJ, Sheppard SK. Rapid host switching in generalist Campylobacter strains erodes the signal for tracing human infections. ISME J (2016) 10(3):721–9. doi:10.1038/ismej.2015.149
31. Hepworth PJ, Ashelford KE, Hinds J, Gould KA, Witney AA, Williams NJ, et al. Genomic variations define divergence of water/wildlife-associated Campylobacter jejuni niche specialists from common clonal complexes. Environ Microbiol (2011) 13(6):1549–60. doi:10.1111/j.1462-2920.2011.02461.x
32. Jerome JP, Bell JA, Plovanich-Jones AE, Barrick JE, Brown CT, Mansfield LS. Standing genetic variation in contingency loci drives the rapid adaptation of Campylobacter jejuni to a novel host. PLoS One (2011) 6(1):e16399. doi:10.1371/journal.pone.0016399
33. Lango-Scholey L, Aidley J, Woodacre A, Jones MA, Bayliss CD. High throughput method for analysis of repeat number for 28 phase variable loci of Campylobacter jejuni strain NCTC11168. PLoS One (2016) 11(7):e0159634. doi:10.1371/journal.pone.0159634
34. Szymanski CM, Gaynor E. How a sugary bug gets through the day: recent developments in understanding fundamental processes impacting Campylobacter jejuni pathogenesis. Gut Microbes (2012) 3(2):135–44. doi:10.4161/gmic.19488
35. Maue AC, Mohawk KL, Giles DK, Poly F, Ewing CP, Jiao Y, et al. The polysaccharide capsule of Campylobacter jejuni modulates the host immune response. Infect Immun (2013) 81(3):665–72. doi:10.1128/iai.01008-12
36. van der Woude MW, Baumler AJ. Phase and antigenic variation in bacteria. Clin Microbiol Rev (2004) 17(3):581–611. doi:10.1128/cmr.17.3.581-611.2004
37. Szymanski CM, Yao R, Ewing CP, Trust TJ, Guerry P. Evidence for a system of general protein glycosylation in Campylobacter jejuni. Mol Microbiol (1999) 32(5):1022–30. doi:10.1046/j.1365-2958.1999.01415.x
38. Szymanski CM, Logan SM, Linton D, Wren BW. Campylobacter—a tale of two protein glycosylation systems. Trends Microbiol (2003) 11(5):233–8. doi:10.1016/S0966-842X(03)00079-9
39. van Sorge NM, Bleumink NM, van Vliet SJ, Saeland E, van der Pol WL, van Kooyk Y, et al. N-glycosylated proteins and distinct lipooligosaccharide glycoforms of Campylobacter jejuni target the human C-type lectin receptor MGL. Cell Microbiol (2009) 11(12):1768–81. doi:10.1111/j.1462-5822.2009.01370.x
40. Phongsisay V. Campylobacter jejuni targets immunoglobulin-like receptor LMIR5. Mol Immunol (2015) 63(2):574–8. doi:10.1016/j.molimm.2014.07.024
41. Rosner BM, Schielke A, Didelot X, Kops F, Breidenbach J, Willrich N, et al. A combined case–control and molecular source attribution study of human Campylobacter infections in Germany, 2011–2014. Sci Rep (2017) 7:5139. doi:10.1038/s41598-017-05227-x
42. Karlyshev AV, Champion OL, Churcher C, Brisson JR, Jarrell HC, Gilbert M, et al. Analysis of Campylobacter jejuni capsular loci reveals multiple mechanisms for the generation of structural diversity and the ability to form complex heptoses. Mol Microbiol (2005) 55(1):90–103. doi:10.1111/j.1365-2958.2004.04374.x
43. Guerry P, Szymanski CM, Prendergast MM, Hickey TE, Ewing CP, Pattarini DL, et al. Phase variation of Campylobacter jejuni 81-176 lipooligosaccharide affects ganglioside mimicry and invasiveness in vitro. Infect Immun (2002) 70(2):787–93. doi:10.1128/IAI.70.2.787-793.2002
44. Bacon DJ, Szymanski CM, Burr DH, Silver RP, Alm RA, Guerry P. A phase-variable capsule is involved in virulence of Campylobacter jejuni 81-176. Mol Microbiol (2001) 40(3):769–77. doi:10.1046/j.1365-2958.2001.02431.x
45. Pequegnat B, Laird RM, Ewing CP, Hill CL, Omari E, Poly F, et al. Phase-variable changes in the position of o-methyl phosphoramidate modifications on the polysaccharide capsule of Campylobacter jejuni modulate serum resistance. J Bacteriol (2017) 199(14):e00027–17. doi:10.1128/jb.00027-17
46. Singh SK, Streng-Ouwehand I, Litjens M, Weelij DR, Garcia-Vallejo JJ, van Vliet SJ, et al. Characterization of murine MGL1 and MGL2 C-type lectins: distinct glycan specificities and tumor binding properties. Mol Immunol (2009) 46(6):1240–9. doi:10.1016/j.molimm.2008.11.021
47. Artigas G, Monteiro JT, Hinou H, Nishimura SI, Lepenies B, Garcia-Martin F. Glycopeptides as targets for dendritic cells: exploring MUC1 glycopeptides binding profile toward macrophage galactose-type lectin (MGL) orthologs. J Med Chem (2017) 60(21):9012–21. doi:10.1021/acs.jmedchem.7b01242
48. Brown GD, Taylor PR, Reid DM, Willment JA, Williams DL, Martinez-Pomares L, et al. Dectin-1 is a major beta-glucan receptor on macrophages. J Exp Med (2002) 196(3):407–12. doi:10.1084/jem.20020470
49. Lefèvre L, Lugo-Villarino G, Meunier E, Valentin A, Olagnier D, Authier H, et al. The C-type lectin receptors Dectin-1, MR, and SIGNR3 contribute both positively and negatively to the macrophage response to Leishmania infantum. Immunity (2013) 38(5):1038–49. doi:10.1016/j.immuni.2013.04.010
50. Lima-Junior DS, Mineo TWP, Calich VLG, Zamboni DS. Dectin-1 activation during Leishmania amazonensis phagocytosis prompts syk-dependent reactive oxygen species production to trigger inflammasome assembly and restriction of parasite replication. J Immunol (2017) 199(6):2055–68. doi:10.4049/jimmunol.1700258
51. Saijo S, Ikeda S, Yamabe K, Kakuta S, Ishigame H, Akitsu A, et al. Dectin-2 recognition of alpha-mannans and induction of Th17 cell differentiation is essential for host defense against Candida albicans. Immunity (2010) 32(5):681–91. doi:10.1016/j.immuni.2010.05.001
52. Yadav M, Schorey JS. The beta-glucan receptor Dectin-1 functions together with TLR2 to mediate macrophage activation by mycobacteria. Blood (2006) 108(9):3168–75. doi:10.1182/blood-2006-05-024406
53. Papp-Szabo E, Kanipes MI, Guerry P, Monteiro MA. Cell-surface alpha-glucan in Campylobacter jejuni 81-176. Carbohydr Res (2005) 340(13):2218–21. doi:10.1016/j.carres.2005.06.023
54. Behler-Janbeck F, Takano T, Maus R, Stolper J, Jonigk D, Tort Tarres M, et al. C-type lectin Mincle recognizes glucosyl-diacylglycerol of Streptococcus pneumoniae and plays a protective role in pneumococcal pneumonia. PLoS Pathog (2016) 12(12):e1006038. doi:10.1371/journal.ppat.1006038
55. Ishikawa T, Itoh F, Yoshida S, Saijo S, Matsuzawa T, Gonoi T, et al. Identification of distinct ligands for the C-type lectin receptors Mincle and Dectin-2 in the pathogenic fungus Malassezia. Cell Host Microbe (2013) 13(4):477–88. doi:10.1016/j.chom.2013.03.008
56. Unger WWJ, van Beelen AJ, Bruijns SC, Joshi M, Fehres CM, van Bloois L, et al. Glycan-modified liposomes boost CD4+ and CD8+ T-cell responses by targeting DC-SIGN on dendritic cells. J Control Release (2012) 160(1):88–95. doi:10.1016/j.jconrel.2012.02.007
57. Boks MA, Ambrosini M, Bruijns SC, Kalay H, van Bloois L, Storm G, et al. MPLA incorporation into DC-targeting glycoliposomes favours anti-tumour T cell responses. J Control Release (2015) 216(Suppl C):37–46. doi:10.1016/j.jconrel.2015.06.033
58. Boks MA, Bruijns SCM, Ambrosini M, Kalay H, van Bloois L, Storm G, et al. In situ delivery of tumor antigen- and adjuvant-loaded liposomes boosts antigen-specific T-cell responses by human dermal dendritic cells. J Invest Dermatol (2015) 135(11):2697–704. doi:10.1038/jid.2015.226
59. Petrovsky N, Cooper PD. Carbohydrate-based immune adjuvants. Expert Rev Vaccines (2011) 10(4):523–37. doi:10.1586/erv.11.30
60. Johannssen T, Lepenies B. Glycan-based cell targeting to modulate immune responses. Trends Biotechnol (2017) 35(4):334–46. doi:10.1016/j.tibtech.2016.10.002
61. Kiyotake R, Oh-Hora M, Ishikawa E, Miyamoto T, Ishibashi T, Yamasaki S. Human Mincle binds to cholesterol crystals and triggers innate immune responses. J Biol Chem (2015) 290(42):25322–32. doi:10.1074/jbc.M115.645234
Keywords: C-type lectin receptors, Campylobacter jejuni, innate immunity, flow cytometry, confocal microscopy, ELISA assay, screening tools, Dectin-1 receptor
Citation: Mayer S, Moeller R, Monteiro JT, Ellrott K, Josenhans C and Lepenies B (2018) C-Type Lectin Receptor (CLR)–Fc Fusion Proteins As Tools to Screen for Novel CLR/Bacteria Interactions: An Exemplary Study on Preselected Campylobacter jejuni Isolates. Front. Immunol. 9:213. doi: 10.3389/fimmu.2018.00213
Received: 16 November 2017; Accepted: 25 January 2018;
Published: 13 February 2018
Edited by:
Thomas A. Kufer, University of Hohenheim, GermanyReviewed by:
Dirk Werling, Royal Veterinary College, United KingdomAnca Dorhoi, Friedrich Loeffler Institute Greifswald, Germany
Copyright: © 2018 Mayer, Moeller, Monteiro, Ellrott, Josenhans and Lepenies. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christine Josenhans, am9zZW5oYW5zQG12cC51bmktbXVlbmNoZW4uZGU=;
Bernd Lepenies, YmVybmQubGVwZW5pZXNAdGloby1oYW5ub3Zlci5kZQ==
†Present address: Rebecca Moeller, Institute for Experimental Virology, TWINCORE, Centre for Experimental and Clinical Infection Research, A Joint Venture between the Medical School Hannover and the Helmholtz Centre for Infection Research, Hannover, Germany