 Anicet Christel Maloupazoa Siawaya1,2
Anicet Christel Maloupazoa Siawaya1,2 Ofilia Mvoundza Ndjindji1,2
Ofilia Mvoundza Ndjindji1,2 Eliane Kuissi Kamgaing3,4
Eliane Kuissi Kamgaing3,4 Amandine Mveang-Nzoghe1,2
Amandine Mveang-Nzoghe1,2 Chérone Nancy Mbani Mpega5
Chérone Nancy Mbani Mpega5 Marielle Leboueny1,2Roselyne Kengue Boussougou6Armel Mintsa Ndong6
Marielle Leboueny1,2Roselyne Kengue Boussougou6Armel Mintsa Ndong6 Paulin N. Essone2,7,8
Paulin N. Essone2,7,8 Joel Fleury Djoba Siawaya1,2*
Joel Fleury Djoba Siawaya1,2*
- 1Centre Hospitalier Universitaire Mère-Enfant Fondation Jeanne Ebori (CHUMEFJE), Libreville, Gabon
- 2Unités de Recherche et de Diagnostics Spécialisés, Laboratoire National de Santé Publique à Libreville (LNSP), Libreville, Gabon
- 3Département de Pédiatrie, Université des Sciences de la Santé d’Owendo (USS), Owendo, Gabon
- 4Service de Néonatologie, Centre Hospitalier Universitaire de Libreville (CHUL), Libreville, Gabon
- 5Département de Chimie, Faculté des Sciences, Université des sciences et techniques de Masuku, Franceville, Gabon
- 6Unité de Virologie, Laboratoire National de Santé Publique à Libreville (LNSP), Libreville, Gabon
- 7Centre de Recherche Médicales de Lambaréné, Lambaréné, Gabon
- 8Institut für Tropenmedizin, Universitätsklinikum Tübingen, Tübingen, Germany
Pathogen sensing and recognition through pattern recognition receptors, and subsequent production of pro-inflammatory cytokines, is the cornerstone of the innate immune system. Despite the fact that HIV-exposed uninfected (HEU) infants are prone to serious bacterial infections, no study has focused on the functionality of their bacteria recognition system. This is the first study to investigate baseline levels of three critically important immune response molecules in this population: complement component (C)-3, toll-like receptor (TLR)-4, and C-reactive protein (CRP). We enrolled 16 HEU and 6 HIV-unexposed (HU) infants. TLR4 function was investigated by stimulating whole blood with increasing concentrations of TLR4-agonist ultrapure lipopolysaccharides. TLR4/TLR4-agonist dose response were assessed by measuring IL-6 secretion. Complement C3 and CRP were measured by photo spectrometry. Data showed no significant differences in baseline concentration of CRP between HEU and HU infants. Complement C3 was significantly higher in HEU infants than HU infants. TLR4 anergy was observed in 7 of 12 HEU infants, whereas the rest of HEU infants (n = 4) and the control HU infants tested (n = 3) showed responsive TLR4. None of the HEU infants investigated in this study had severe infections in the year after their birth. In conclusion, TLR4 anergy can occur in HEU infants without necessarily translating to increased vulnerability to infectious diseases.
Introduction
Progress in the prevention of mother-to-child transmission of HIV has led to a significant reduction of mother-to-child HIV transmission rate, increasing the population of HIV-exposed uninfected (HEU) infants (1–3). Indeed, HEU infants are a growing population with over a million infants born every year from HIV-infected mothers (2). Studies have shown that HEU infants are more vulnerable to diseases than infants born from HIV free mothers (4–8). The global efforts to understand HEU infants’ increased susceptibility to infections during the first months of their life has shed some light on the immune determinants of their susceptibility. We recently showed impaired oxidative burst in neutrophils from a number of HEU infants (9). Others have reported impaired humoral response (10, 11), altered chemokine receptor expression by CD4+ T cells (12), altered natural killer cell function, and reduced thymic output of naive CD4+ cells (13).
The innate immune sensing and recognition of microbial elements is critical for the host defense against infection (14). Pattern recognition receptors such as toll-like receptors (TLRs) that detect pathogen-associated molecular patterns (PAMPs) are the hallmark of pathogen sensing, which triggers both innate and adaptive immune response (14). TLRs are type 1 transmembrane receptors. So far 10 functional TLRs have been described in humans including TLRs 1, 2, 4, 5, 6, 7, 8, 9, and 10, each with distinct functions. TLR4 is activated by lipopolysaccharides (LPS) found in Gram-negative bacteria (e.g., Klebsiella pneumoniae, Haemophilus influenza, etc.) (15, 16), which are a significant cause of morbidity and mortality among infants (17). TLR4 plays a central role in defense against Gram-negative bacteria; however, mutations on the TLR4 gene or TLR4-deficient mice have been linked to hypo-responsiveness to LPS and high susceptibility to Gram-negative bacterial infections (15, 18, 19).
In addition to TLR 4, complement component (C)-3 and C-reactive protein (CRP) are also important in innate immune recognition and killing of invading pathogens. The complement C3 is the most abundant complement component in plasma, plays a key role in the activation of complement system, and is required for both classical and alternative complement activation pathways (20). Proteolytic cleavage of the C3 α-chain leads to C3a (a key inflammatory mediator) and C3b (a major opsonin) (20). CRP plays a major role in host defense against infection and in inflammatory processes (21, 22); it is a pro-inflammatory mediator, a pathogen surveillance molecule, an opsonin for diverse pathogens, and an activator of complement that provides early defense against invading pathogens (23).
Because of the critical role of these three molecules, we investigated the function of TLR4 and assessed the baseline levels of complement C3 and CRP in HEU infants. Our results contribute to a global effort to understand the immunity of this infant population. Understanding HEU infants’ immunity is essential to the development of new health-care policies (therapeutic, vaccines, etc.) to combat infectious agents.
Materials and Methods
Subjects
The present prospective study included 16 HEU and 6 HIV-unexposed (HU) infants from the National Laboratory of Public Health in Libreville (Gabon). Whole blood was collected in 5 ml EDTA tubes and transported to the laboratory for processing. The national laboratory of public health ethical committee approved this study. Written informed consent was obtained from parents for individual participants. Mothers’ and infants’ information on preventive treatment, breastfeeding, and time of antiretroviral therapy initiation were also collected. All mothers came from modest background.
HIV perinatal infection (RT-PCR, NucliSENS, Biomerieux, France) was checked from each HEU infant in peripheral blood at 1.5, 3, 6, and 9 months after birth as part of their scheduled check up in the mother-to-child transmission prevention programs reference laboratory. An additional sample was taken to determine anti-HIV-1 antibodies seroconversion by ELISA at 18 months old. During each scheduled visit, infants’ medical records were checked and mothers were asked if infants had experienced any severe infection or if they were hospitalized. All HIV-positive infants and infants with any illness or infection at the time of recruitment were excluded from the study.
Baseline Complement C3 and CRP Levels (Circulating Levels of Complement C3 and CRP in Absence of Any Stimulus)
Circulating levels of complement C3 and CRP in HEU infants (N = 11) and HU infants (N = 6) were assessed using MD Pacific kits (C3: Ref MD015.SL and CRP: Ref MD021.SL) and protein analyzer (MD Pacific Biotechnology, Tianjin, China). The protocol was performed according to the manufacturer’s instructions. Briefly, whole blood samples were centrifuged before plasma collection. 20 µl of the collected plasma was diluted 1:10 in sample diluent and read on the MD Pacific analyzer to measure the quantity of complement C3 and CRP.
TLR4 Activation Profile Assay
A volume of 200 µl of freshly drawn whole blood was directly transferred into TLR4 test strips pre-coated with a TLR4 agonist at different concentrations (ultrapure LPS at: 0, 0.001, 0.01, 0.1, 1, 10, 100, and 1,000 ng) (Ref tlrs-tlr4, Invivogen, San Diego, CA, USA). All study participants with less than 1.6 ml of whole blood were excluded from this experiment. The TLR4 test strips containing blood samples from infants were incubated overnight at 37°C in a humidified 5% CO2 incubator. The following day, blood supernatants were collected and stored at −40°C for 18 months. We were able to harvest enough plasma from 12 HEU and 3 HU infants after whole blood stimulation with LPS.
IL-6 Expression
Frozen whole blood supernatants from the TLR4/TLR4-agonist stimulation assay were thawed and diluted 1/10 in assay diluent. IL-6 concentration was measured using Elecsys cartridge (Ref 05109442190, Roche Diagnostics, Germany) on the Cobas e411 analyzer (Roche Diagnostics, Germany). The test measuring range was 1.5–5.000 pg/ml. Samples with an IL-6 concentration above the test limit of 5,000 pg/ml were further diluted (in assay diluent) and reanalyzed.
Statistical Analyses
The levels of complement C3 and CRP between the two groups of infants were analyzed using the Mann–Whitney U-test with an alpha value of 0.05. For each infant, a TLR4/TLR4-agonist dose–response curve was generated and analyzed by linear regression. All statistical analyses were done on GraphPad Prism 6 (GraphPad Software, Inc., USA).
Results
Information on mothers’ and infants’ preventive treatment, breastfeeding, and time of antiretroviral therapy initiation are presented in Table 1. All HIV-positive mothers followed the mother-to-child preventive therapy. All infants except one followed the nevirapine and co-trimoxazole preventive therapy. 92% of infants were not breastfed.
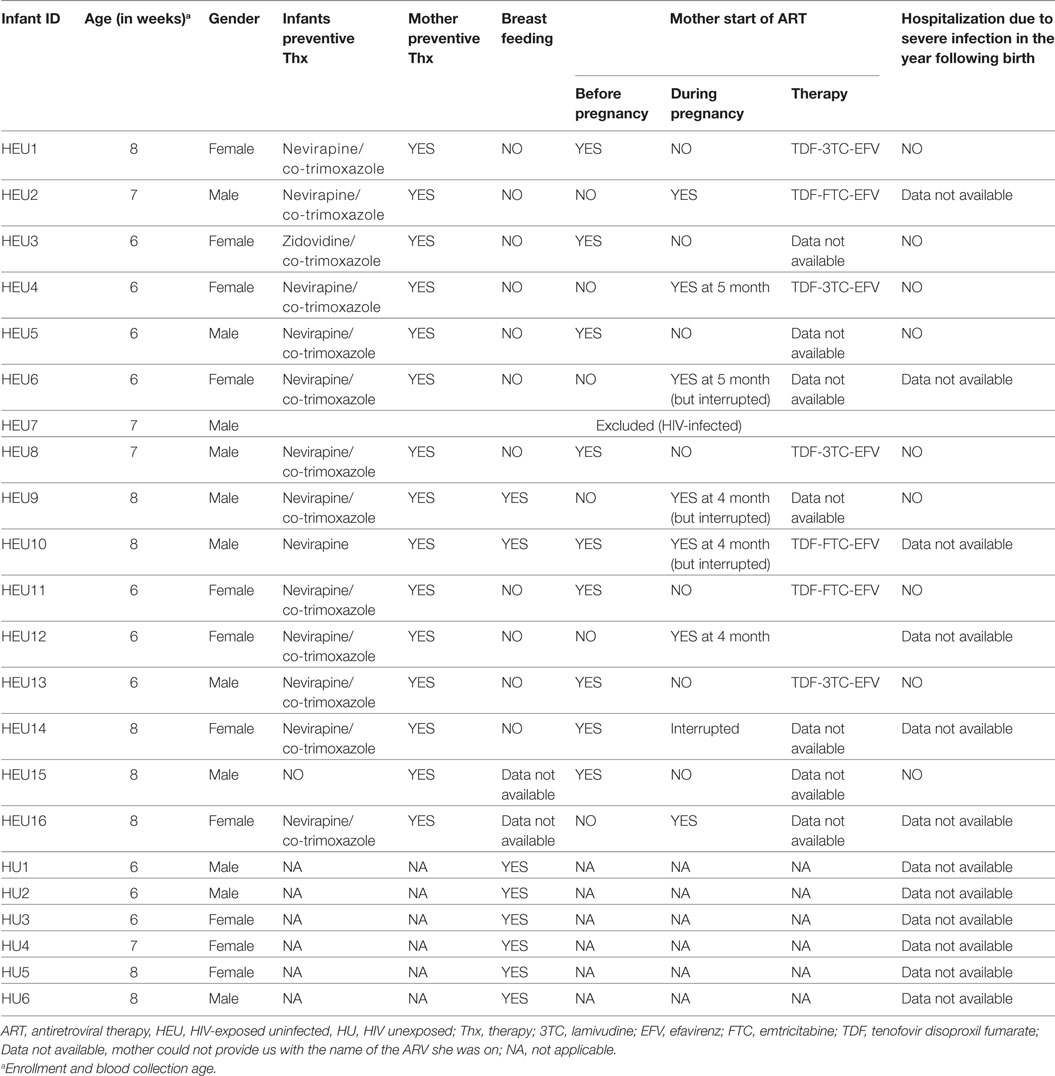
Table 1. Participants anthropometric and clinical information.
High Level of Complement C3 in HEU Infants
To assess the baseline inflammatory status of infants (HEU: N = 11; HU: N = 6), we investigated the circulating levels of complement C3 and CRP. The level of complement C3 was significantly higher in HEU infants compared with HU infants (median of 780 vs. 595; p = 0.03). No significant difference was observed between infants’ groups for CRP (Figure 1).
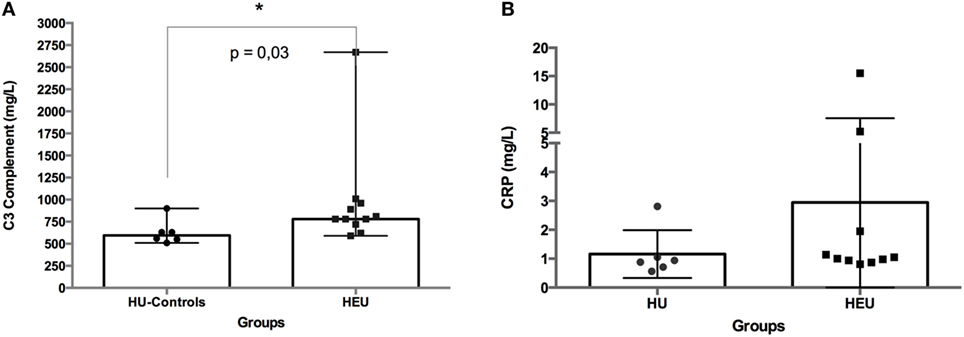
Figure 1. Circulating levels of C3 complement (A) and C-reactive protein (CRP) (B) in whole blood from HIV-exposed and HIV-unexposed (HU) infants. C3 complement and CRP were measured in infants’ serum samples. The difference between HIV-exposed and HU infants was analyzed using the Mann–Whitney U-test. The differences were considered significant for a p-value <0.05. In the figures, bars indicate medians and ranges.
Response to TLR4-Agonist
HIV-exposed uninfected infants showed two distinct TLR4 response profiles when stimulated for 24 h with ultrapure LPS (TLR4-agonist) (Figure 2). More than half of HEU infants (7 of 12) did not respond to LPS stimulation as shown by their IL-6 secretion profiles. Despite stimulation with increasing concentration of TLR4 agonist (0.0001–1,000 ng/ml), in the hyporesponsive group, IL-6 secretions did not exceed 41 pg/ml even for the top agonist concentration. The remaining HEU and HU infants showed enhanced IL-6 secretions to TLR4-agonist stimulations (Figure 2). We observed a positive correlation between IL-6 secretion and LPS concentration. This correlation was similar in all LPS responders independently of their HIV exposure status.
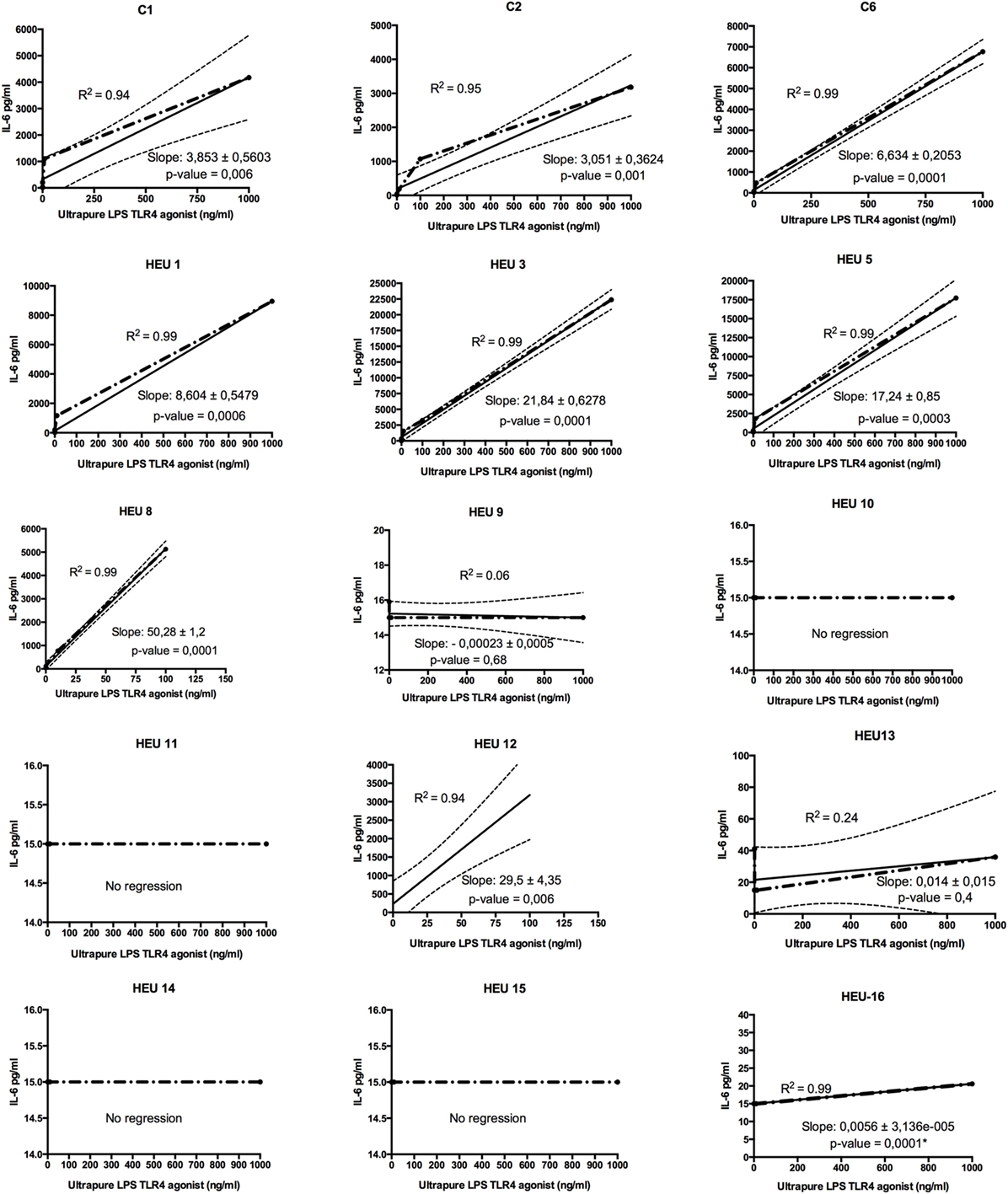
Figure 2. Regression analysis of toll-like receptor (TLR)-4/ultrapure lipopolysaccharides (LPS) dose–response of HIV-exposed uninfected (HEU) infants and HIV-unexposed infants (C). Whole blood from individual participant was stimulated at increasing concentration of LPS (from 0, 0.001 to 1,000 ng), and IL-6 was measured. The slope indicator measures the rise over the run for a linear regression. The p-value simply tests whether the slope is exactly 0 or ≠0. The coefficient of determination means that R2 × 100% of the variation in IL-6 can be explained by linear relationship with TLR4-agonist stimulation. (*) Only the one point deviated significantly from 0 (corresponding to the top LPS stimulation).
Linear regression analysis of the TLR4 dose–response assay confirmed that selected HEU infants had hyporesponsive TLR4. Indeed, six healthy infants born from HIV-positive women (HEU 9, 10, 11, 13, 14, and 15) showed no significant or no deviation of their dose–response slopes. Further, one HEU infant (HEU-16) showed a dose–response slope deviating significantly from zero. However, the slope, which represents the rise across an increasing concentration of TLR4-agonist, was very weak (0.0056), indicating a very weak response to TLR4-agonist. The remaining five HEU infants showed a linearity between TLR4-agonist concentration and IL-6 secretion.
Discussion
There is evidence that in utero exposure to HIV and antiretroviral therapy (ART) have knock-on health effects on infants, including an increased susceptibility to bacterial infection (4–7, 24–27). Research on the etiology of HEU infants’ susceptibility has highlighted a number of immune abnormalities in these infants (9, 10, 25, 28–30). We believe that the benefits of ART in preventing HIV vertical transmission and improving mother and child health are undeniable (31). Although these benefits certainly outweigh the potential adverse effects of ART exposure to infants, these adverse effects should not be neglected, as they can be detrimental to infants’ health (25).
In this study, half of the mothers started therapy during pregnancy, two of them interrupted their treatment, and 92% of infants were not breastfed. This is not surprising as, in Gabon, despite free antenatal care, early access, and adherence to components of mother to child, HIV transmission preventive care remains unsatisfactory (3). Therefore, mothers are often advised not to breastfeed if they are not fully compliant to the ARV therapy.
Complement C3, plays a key role in both classical and alternative complement activation pathways (20). Here, we showed that HEU infants had significantly higher concentrations of complement C3 compared to the control group of HU infants. Hemolytic activity and components of the complement system (32) are low in newborn; therefore, the low levels of complement C3 in our group of HU infants are not surprising. Viral particles including HIV are known to induce the production and activate complement C3 (33–35). Also, studies showed that fetal exposure to HIV and ART skews newborns infant’s physiological immune responses toward inflammation (36). Therefore, HEU infants in utero exposure to HIV particles and pro-inflammatory environment (37) could explain the higher level of complement C3 found in HEU infants.
However, our results need to be interpreted with caution due to the limited sample size of the present study, and note further studies are required to validate this observation. No significant differences between HEU and HU infants were seen in the baseline level of the inflammatory factor CRP. This result is different from Prendergast et al. (38) who showed that at 6 weeks of age, HEUinfants had significantly higher CRP than HU infants. The difference in CRP results between Prendergast et al. studies and our study could be explained by the fact that, contrary to Prendergast et al. study, almost all HEU infants in our study were not breastfed, limiting the transfer of breastmilk associated HIV-antigen or pro-inflammatory factors. Indeed, Adair et al. (39) demonstrated that in exclusively breastfed infants born to HIV-infected mothers, maternal factors (inflammatory factors included) correlated to infants CRP levels. Another possible explanation would be the difference in studies’ sizes and power.
Our data indicate that a number of HEU infants have impaired TLR4 function. More than half of HEU infants did not respond to TLR4-agonist (LPS) stimulation. The mechanisms explaining or leading the observed TLR4 anergy are unknown. Further studies should be conducted to confirm our observation and uncover the mechanism involved. Environmental factors in utero have been shown to modulate early life immune responses in different ways (40). Without affirming that it is the case here, it has been demonstrated by Piccinini et al. (41) that fetal antiretroviral exposure induced a blockade of NF-κB activation and degradation of its inhibitor IκBα. Furthermore, Equils and colleagues demonstrated that HIV-1 protease inhibitors block TLR4-induced NF-κB activation as well as LPS-induced IL-6 promoter transactivation (42). It might be possible that fetal exposure to other antiretroviral drugs has the same effect.
Accumulated data on HEU infants’ immune system suggest heterogeneity in their immune response. Some are able to mount strong responses to specific antigens; whereas, others showed a weak or absent to them (9–11, 28). It seems like any part or pathway of the immune system can be affected. Our study, although small, shows that selected HEU infants are characterized by a TLR4 anergy, whereas others respond to LPS, confirming the heterogeneity of HEU infants’ immune response. HEU infants enhanced response to PAMPs in their early life have also been reported by Reikie et al. (43).
Our findings would suggest that selected HEU infants (those not responding to LPS stimulation) would be particularly susceptible to Gram-negative bacteria infections. However, none of the HEU infants investigated in this study had severe infections in the year after their birth suggesting an alternative or compensatory mechanism to TLR4 function in infants non-responsive to LPS stimulation. It could also be that infants were well cared for after counseling of their mothers by our health-care workers.
In conclusion, HEU infants are characterized by a relative enhanced complement C3 expression and an impaired TLR4 response. TLR4 anergy can occur in HEU infants without necessarily translating to increased vulnerability to infection. The limited amount of whole blood obtained from enrolled infants made it difficult to further compare HEU and HU infants. This present pilot study needs to be validated with a larger population to confirm these observations (especially for complement C3) as larger your sample is, the more accurately it reflects the population.
Ethics Statement
The National Laboratory of Public Health board approved the study. Parental consent was obtained for all infants.
Author Contributions
JFDS is the principal investigators who conceived and designed the study. ACMS, OMN, and CNMM recruited participants and acquired the samples. ACMS, OMN, and AMN1 (Amandine) helped in designing the TLR4 function experiment. ACMS, OMN, and AMN1. did the experiments and all supervised by JFDS ML, RKB, and AMN2 (Armel) helped in recruited participants and acquired the samples; RKB and AMN2 did all the HIV testing. PNE helped writing the manuscript. JFDS did the analyses and wrote the manuscript with input from all authors supervised. All authors approved the final manuscript revisions.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge Dr. Victor Schmidt and Dr. Matthew McCall for the critical reading of this manuscript. Also, we acknowledge our trainees Miss. Laurena Schelly Otere Nkiessouma and Mr. Davy Ulrich L. Mouangala who assisted in collecting infants’ data. Thanks to Gabonese Government for its financial contribution.
Funding
The present study was funded by a small grant from the Gabonese Government.
References
1. Padian NS, McCoy SI, Manian S, Wilson D, Schwartlander B, Bertozzi SM. Evaluation of large-scale combination HIV prevention programs: essential issues. J Acquir Immune Defic Syndr (2011) 58(2):e23–8. doi:10.1097/QAI.0b013e318227af37
2. Sugandhi N, Rodrigues J, Kim M, Ahmed S, Amzel A, Tolle M, et al. HIV-exposed infants: rethinking care for a lifelong condition. AIDS (2013) 27(Suppl 2):S187–95. doi:10.1097/QAD.0000000000000090
3. Mintsa-Ndong A, Ndong-Ella C, Boussougou RK, Busugu LM, Mba A, Agwambouet FA, et al. Mother-to-child HIV-transmission prevention programs in a sub-Saharan African setting: the Gabonese experience. Int J STD AIDS (2018) 29(3):221–6. doi:10.1177/0956462417722132
4. Marinda E, Humphrey JH, Iliff PJ, Mutasa K, Nathoo KJ, Piwoz EG, et al. Child mortality according to maternal and infant HIV status in Zimbabwe. Pediatr Infect Dis J (2007) 26(6):519–26. doi:10.1097/01.inf.0000264527.69954.4c
5. Brahmbhatt H, Kigozi G, Wabwire-Mangen F, Serwadda D, Lutalo T, Nalugoda F, et al. Mortality in HIV-infected and uninfected children of HIV-infected and uninfected mothers in rural Uganda. J Acquir Immune Defic Syndr (2006) 41(4):504–8. doi:10.1097/01.qai.0000188122.15493.0a
6. Slogrove A, Reikie B, Naidoo S, De Beer C, Ho K, Cotton M, et al. HIV-exposed uninfected infants are at increased risk for severe infections in the first year of life. J Trop Pediatr (2012) 58(6):505–8. doi:10.1093/tropej/fms019
7. Slogrove AL, Cotton MF, Esser MM. Severe infections in HIV-exposed uninfected infants: clinical evidence of immunodeficiency. J Trop Pediatr (2010) 56(2):75–81. doi:10.1093/tropej/fmp057
8. Evans C, Humphrey JH, Ntozini R, Prendergast AJ. HIV-exposed uninfected infants in Zimbabwe: insights into health outcomes in the pre-antiretroviral therapy era. Front Immunol (2016) 7:190. doi:10.3389/fimmu.2016.00190
9. Maloupazoa Siawaya AC, Mveang-Nzoghe A, Mvoundza Ndjindji O, Mintsa Ndong A, Essone PN, Djoba Siawaya JF. Cases of impaired oxidative burst in HIV-exposed uninfected infants’ neutrophils – a pilot study. Front Immunol (2017) 8:262. doi:10.3389/fimmu.2017.00262
10. Abramczuk BM, Mazzola TN, Moreno YM, Zorzeto TQ, Quintilio W, Wolf PS, et al. Impaired humoral response to vaccines among HIV-exposed uninfected infants. Clin Vaccine Immunol (2011) 18(9):1406–9. doi:10.1128/CVI.05065-11
11. Bunders M, Pembrey L, Kuijpers T, Newell ML. Evidence of impact of maternal HIV infection on immunoglobulin levels in HIV-exposed uninfected children. AIDS Res Hum Retroviruses (2010) 26(9):967–75. doi:10.1089/aid.2009.0241
12. Bunders MJ, van Hamme JL, Jansen MH, Boer K, Kootstra NA, Kuijpers TW. Fetal exposure to HIV-1 alters chemokine receptor expression by CD4+T cells and increases susceptibility to HIV-1. Sci Rep (2014) 4:6690. doi:10.1038/srep06690
13. Smith C, Jalbert E, de Almeida V, Canniff J, Lenz LL, Mussi-Pinhata MM, et al. Altered natural killer cell function in HIV-exposed uninfected infants. Front Immunol (2017) 8:470. doi:10.3389/fimmu.2017.00470
14. Pandey S, Agrawal DK. Immunobiology of toll-like receptors: emerging trends. Immunol Cell Biol (2006) 84(4):333–41. doi:10.1111/j.1440-1711.2006.01444.x
15. Elson G, Dunn-Siegrist I, Daubeuf B, Pugin J. Contribution of toll-like receptors to the innate immune response to Gram-negative and Gram-positive bacteria. Blood (2007) 109(4):1574–83. doi:10.1182/blood-2006-06-032961
16. Poltorak A, He X, Smirnova I, Liu MY, Van Huffel C, Du X, et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science (1998) 282(5396):2085–8. doi:10.1126/science.282.5396.2085
17. Berezin EN, Solorzano F, Latin America Working Group on Bacterial Resistance. Gram-negative infections in pediatric and neonatal intensive care units of Latin America. J Infect Dev Ctries (2014) 8(8):942–53. doi:10.3855/jidc.4590
18. Branger J, Knapp S, Weijer S, Leemans JC, Pater JM, Speelman P, et al. Role of toll-like receptor 4 in Gram-positive and Gram-negative pneumonia in mice. Infect Immun (2004) 72(2):788–94. doi:10.1128/IAI.72.2.788-794.2004
19. Roger T, Froidevaux C, Le Roy D, Reymond MK, Chanson AL, Mauri D, et al. Protection from lethal Gram-negative bacterial sepsis by targeting toll-like receptor 4. Proc Natl Acad Sci U S A (2009) 106(7):2348–52. doi:10.1073/pnas.0808146106
20. Elvington M, Liszewski MK, Atkinson JP. Evolution of the complement system: from defense of the single cell to guardian of the intravascular space. Immunol Rev (2016) 274(1):9–15. doi:10.1111/imr.12474
21. Ahmed AE, James PB. Clinical utility of complement assessment. Clin Diagn Lab Immunol (1995) 2(5):509–17.
22. Ansar W, Ghosh S. C-reactive protein and the biology of disease. Immunol Res (2013) 56(1):131–42. doi:10.1007/s12026-013-8384-0
23. Du Clos TW. Function of C-reactive protein. Ann Med (2000) 32(4):274–8. doi:10.3109/07853890009011772
24. Evans C, Jones CE, Prendergast AJ. HIV-exposed, uninfected infants: new global challenges in the era of paediatric HIV elimination. Lancet Infect Dis (2016) 16(6):e92–107. doi:10.1016/S1473-3099(16)00055-4
25. Tobin NH, Aldrovandi GM. Immunology of pediatric HIV infection. Immunol Rev (2013) 254(1):143–69. doi:10.1111/imr.12074
26. Kuhn L, Thea DM, Aldrovandi GM. Bystander effects: children who escape infection but not harm. J Acquir Immune Defic Syndr (2007) 46(5):517–8. doi:10.1097/QAI.0b013e31814d6600
27. Denneman L, Cohen S, Godfried MH, van Leeuwen E, Nellen JF, Kuijpers TW, et al. In-utero exposure to tenofovir is associated with impaired fetal and infant growth: need for follow-up studies in combination antiretroviral therapy/HIV-exposed infants. AIDS (2016) 30(13):2135–7. doi:10.1097/QAD.0000000000001156
28. Nielsen SD, Jeppesen DL, Kolte L, Clark DR, Sorensen TU, Dreves AM, et al. Impaired progenitor cell function in HIV-negative infants of HIV-positive mothers results in decreased thymic output and low CD4 counts. Blood (2001) 98(2):398–404. doi:10.1182/blood.V98.2.398
29. Afran L, Garcia Knight M, Nduati E, Urban BC, Heyderman RS, Rowland-Jones SL. HIV-exposed uninfected children: a growing population with a vulnerable immune system? Clin Exp Immunol (2014) 176(1):11–22. doi:10.1111/cei.12251
30. Ruck C, Reikie BA, Marchant A, Kollmann TR, Kakkar F. Linking susceptibility to infectious diseases to immune system abnormalities among HIV-exposed uninfected infants. Front Immunol (2016) 7:310. doi:10.3389/fimmu.2016.00310
31. Siegfried N, van der Merwe L, Brocklehurst P, Sint TT. Antiretrovirals for reducing the risk of mother-to-child transmission of HIV infection. Cochrane Database Syst Rev (2011) (7):CD003510. doi:10.1002/14651858.CD003510.pub3
32. Drew JH, Arroyave CM. The complement system of the newborn infant. Biol Neonate (1980) 37(3–4):209–17. doi:10.1159/000241276
33. Blue CE, Spiller OB, Blackbourn DJ. The relevance of complement to virus biology. Virology (2004) 319(2):176–84. doi:10.1016/j.virol.2003.11.029
34. Bruder C, Hagleitner M, Darlington G, Mohsenipour I, Wurzner R, Hollmuller I, et al. HIV-1 induces complement factor C3 synthesis in astrocytes and neurons by modulation of promoter activity. Mol Immunol (2004) 40(13):949–61. doi:10.1016/j.molimm.2003.10.016
35. Nitkiewicz J, Borjabad A, Morgello S, Murray J, Chao W, Emdad L, et al. HIV induces expression of complement component C3 in astrocytes by NF-kappaB-dependent activation of interleukin-6 synthesis. J Neuroinflammation (2017) 14(1):23. doi:10.1186/s12974-017-0794-9
36. Schoeman JC, Moutloatse GP, Harms AC, Vreeken RJ, Scherpbier HJ, Van Leeuwen L, et al. Fetal metabolic stress disrupts immune homeostasis and induces proinflammatory responses in human immunodeficiency virus type 1- and combination antiretroviral therapy-exposed infants. J Infect Dis (2017) 216(4):436–46. doi:10.1093/infdis/jix291
37. Faye A, Pornprasert S, Mary JY, Dolcini G, Derrien M, Barre-Sinoussi F, et al. Characterization of the main placental cytokine profiles from HIV-1-infected pregnant women treated with anti-retroviral drugs in France. Clin Exp Immunol (2007) 149(3):430–9. doi:10.1111/j.1365-2249.2007.03411.x
38. Prendergast AJ, Chasekwa B, Rukobo S, Govha M, Mutasa K, Ntozini R, et al. Intestinal damage and inflammatory biomarkers in human immunodeficiency virus (HIV)-exposed and HIV-infected Zimbabwean infants. J Infect Dis (2017) 216(6):651–61. doi:10.1093/infdis/jix367
39. Adair LS, Thompson AL, Kayira D, Chasela C, Kacheche Z, Kamwendo D, et al. Maternal and infant predictors of CRP in exclusively breastfed infants born to HIV-infected Malawian mothers. FASEB J (2012) 26(1 Suppl):43–6.
40. MacGillivray DM, Kollmann TR. The role of environmental factors in modulating immune responses in early life. Front Immunol (2014) 5:434. doi:10.3389/fimmu.2014.00434
41. Piccinini M, Rinaudo MT, Chiapello N, Ricotti E, Baldovino S, Mostert M, et al. The human 26S proteasome is a target of antiretroviral agents. AIDS (2002) 16(5):693–700. doi:10.1097/00002030-200203290-00004
42. Equils O, Shapiro A, Madak Z, Liu C, Lu D. Human immunodeficiency virus type 1 protease inhibitors block toll-like receptor 2 (TLR2)- and TLR4-induced NF-kappaB activation. Antimicrob Agents Chemother (2004) 48(10):3905–11. doi:10.1128/AAC.48.10.3905-3911.2004
Keywords: infants, HIV-1, toll-like receptor-4, complement component-3, C-reactive protein
Citation: Maloupazoa Siawaya AC, Mvoundza Ndjindji O, Kuissi Kamgaing E, Mveang-Nzoghe A, Mbani Mpega CN, Leboueny M, Kengue Boussougou R, Mintsa Ndong A, Essone PN and Djoba Siawaya JF (2018) Altered Toll-Like Receptor-4 Response to Lipopolysaccharides in Infants Exposed to HIV-1 and Its Preventive Therapy. Front. Immunol. 9:222. doi: 10.3389/fimmu.2018.00222
Received: 30 November 2017; Accepted: 26 January 2018;
Published: 14 February 2018
Edited by:
Alexandre Corthay, Oslo University Hospital, NorwayReviewed by:
Nelson Silva Filho, Universidade Estadual Paulista Júlio de Mesquita Filho (UNESP), BrazilChristiana Elizabeth Smith, University of Colorado Denver, United States
Copyright: © 2018 Maloupazoa Siawaya, Mvoundza Ndjindji, Kuissi Kamgaing, Mveang-Nzoghe, Mbani Mpega, Leboueny, Kengue Boussougou, Mintsa Ndong, Essone and Djoba Siawaya. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joel Fleury Djoba Siawaya, am9lbC5kam9iYUBnbWFpbC5jb20=