 Yean K. Yong1,2,3*†
Yean K. Yong1,2,3*† Alireza Saeidi2†Hong Y. Tan1,2,3,4Mohamed Rosmawati2Philip F. Enström5
Alireza Saeidi2†Hong Y. Tan1,2,3,4Mohamed Rosmawati2Philip F. Enström5 Rami Al Batran6
Rami Al Batran6 V. Vasuki7Indranil Chattopadhyay8
V. Vasuki7Indranil Chattopadhyay8 Amudhan Murugesan9
Amudhan Murugesan9 Ramachandran Vignesh10Adeeba Kamarulzaman2,11Jayakumar Rajarajeswaran12
Ramachandran Vignesh10Adeeba Kamarulzaman2,11Jayakumar Rajarajeswaran12 Abdul W. Ansari2
Abdul W. Ansari2 Jamuna Vadivelu6
Jamuna Vadivelu6 James E. Ussher13
James E. Ussher13 Vijayakumar Velu14
Vijayakumar Velu14 Marie Larsson5
Marie Larsson5 Esaki M. Shankar11,15,16*†
Esaki M. Shankar11,15,16*†
- 1Laboratory Center, Xiamen University Malaysia, Sepang, Malaysia
- 2Department of Medicine, University of Malaya Medical Centre, Kuala Lumpur, Malaysia
- 3China-ASEAN Institute of Marine Science (CAMS), Xiamen University Malaysia, Sepang, Malaysia
- 4Department of Traditional Chinese Medicine, Xiamen University Malaysia, Sepang, Malaysia
- 5Division of Molecular Virology, Department of Clinical and Experimental Medicine, Linköping University, Linköping, Sweden
- 6Department of Medical Microbiology, Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia
- 7Department of Microbiology, The Government Thiruvarur Medical College and Hospital, Thiruvarur, India
- 8Division of Molecular Cancer Biology, Department of Life Sciences, Central University of Tamil Nadu, Thiruvarur, India
- 9The Government Theni Medical College and Hospital, Theni, India
- 10Laboratory-Based Department, Universiti Kuala Lumpur, Ipoh, Malaysia
- 11Center of Excellence for Research in AIDS, University of Malaya, Kuala Lumpur, Malaysia
- 12Department of Molecular Medicine, Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia
- 13Department of Microbiology and Immunology, University of Otago, Dunedin, New Zealand
- 14Department of Microbiology and Immunology, Emory Vaccine Center, Atlanta, GA, United States
- 15Division of Infection Biology, Department of Life Sciences, Central University of Tamil Nadu, Thiruvarur, India
- 16Department of Microbiology, Central University of Tamil Nadu, Thiruvarur, India
Mucosal-associated invariant T (MAIT) cells, defined as CD161++TCR iVα7.2+ T cells, play an important role in the innate defense against bacterial infections, and their functionality is impaired in chronic viral infections. Here, we investigated the frequency and functional role of MAIT cells in chronic hepatitis B virus (HBV) infection. The peripheral CD3+CD161++TCR iVα7.2+ MAIT cells in chronic HBV-infected patients and healthy controls were phenotypically characterized based on CD57, PD-1, TIM-3, and CTLA-4, as well as HLA-DR and CD38 expression. The frequency of MAIT cells was significantly decreased among chronic HBV-infected individuals as compared to controls. Expression of CD57, PD-1, CTLA-4, as well as HLA-DR and CD38 on MAIT cells was significantly elevated in chronic HBV-infected individuals relative to controls. The percentage of T cell receptor (TCR) iVα7.2+ CD161+ MAIT cells did not correlate with HBV viral load but inversely with HLA-DR on CD4+ T cells and MAIT cells and with CD57 on CD8+ T cells suggesting that decrease of MAIT cells may not be attributed to direct infection by HBV but driven by HBV-induced chronic immune activation. The percentage and expression levels of PD-1 as well as CTLA-4 on MAIT cells inversely correlated with plasma HBV-DNA levels, which may suggest either a role for MAIT cells in the control of HBV infection or the effect of HBV replication in the liver on MAIT cell phenotype. We report that decrease of TCR iVα7.2+ MAIT cells in the peripheral blood and their functions were seemingly impaired in chronic HBV-infected patients likely because of the increased expression of PD-1.
Introduction
Hepatitis B virus (HBV) infection remains a major threat worldwide with ~240 million people chronically infected, and estimates suggest that ~780,000 die annually due to complications including cirrhosis, hepatocellular carcinoma (HCC), and chronic liver failure (1). Therefore, reducing the pool of HBV-infected individuals is of paramount importance for public health. Despite the availability of an effective prophylactic vaccine, a smaller fraction of vaccinated individuals (~2%) in endemic regions can still be chronically infected with HBV (2). Furthermore, prophylactic vaccines have little or no effect on those who have already been chronically infected with HBV (3, 4).
The therapeutic goal in chronic HBV infection is to achieve sustained suppression of viral replication and thereby reduce liver disease progression and the risk of clinical end-points. Despite potent antiviral drugs that inhibit HBV reverse transcriptase, “complete” virological response, defined as hepatitis B surface antigen (HBsAg) clearance, is rarely achieved (5–7). The HBV genome is present in the hepatocytes in a mono-chromosomal form, i.e., cccDNA, and remains intact in spite of the use of these anti-HBV drugs (8). Therefore, virus eradication is seldom achieved and patients experience viral rebound after withdrawal of antiviral therapy (2).
In the absence of treatment, failure to achieve sustained viral control is associated with sub-optimal antiviral CD4+ and CD8+ T-cell responses against HBV core, polymerase protein (9, 10), and antibody responses to envelope (11). Consequently, failure to elicit effective immune responses may be due to clonal deletion of HBV-specific T cell responses (12) and functional exhaustion resulting from the upregulation of co-inhibitory molecules, for instance PD-1 (13), CTLA-4 (14), and TIM-3 (15, 16). Hence, there is a pressing need to better understand the virus–host interactions and the immunological mechanisms in chronic HBV infection, which allows for the designing of novel immunotherapeutic approaches to boost antiviral immune responses.
Mucosal-associated invariant T (MAIT) cells are a recently described subset of innate-like T cells that comprise ~5% of the T-cell pool of adult peripheral blood and are enriched to ~40% in mucosal tissues, liver, and lung in healthy individual (17–19). MAIT cells express a semi-invariant T cell receptor (TCR), including Vα7.2 coupled with restricted Jα segments (Jα33, Jα12, or Jα20), and limited Vβ repertoires (20, 21) as well as their high expression of CD161 (17, 22). MAIT cells play an important role in innate host defense against bacterial infections and are either depleted and/or exhausted in chronic viral infections, including HIV (23–25), chronic HBV (26), and HCV infection (22, 27, 28), and TB infection (29, 30). Here, we investigated the frequency and functional role of MAIT cells along with those of CD4+ and CD8+ T cells in chronic HBV infection. We characterized the phenotypes of peripheral CD3+CD161++TCR iVα7.2+ MAIT cells in chronic HBV-infected patients with and without HBV-DNAemia as compared to healthy controls (HCs) based on the expression of co-inhibitory and senescence markers, CD57, PD-1, TIM-3, and CTLA-4, as well as, HLA-DR and CD38.
Materials and Methods
Ethics Statement
The protocols involving human subjects were approved by the Medical Ethics Committee (MEC) of Universiti Malaya Medical Centre (UMMC), Kuala Lumpur, Malaysia (MEC201311-0496), and conducted as per the guidelines of the International Conference on Harmonization Guidelines and Declaration of Helsinki. All human subjects were adults and provided written informed consent. The written consent form was approved by the ethics committee and signed by the subject or the subject’s legally authorized representative. A copy of the document was given to the person signing the form. The entire consent process was approved by the MEC for conduct of the research.
Study Subjects
A total of 21 individuals with chronic HBV (CHB) infection with HBV-DNAemia positive (limit of detection, 19 IU equivalent to 120 copies/ml) (CHB DNA +ve, n = 12) and HBV-DNAemia negative (CHB DNA −ve, n = 9) as well as HCs (n = 13) were recruited for a cross-sectional analysis. Peripheral blood was obtained from HBV-infected subjects at the Hepatology Unit of the UMMC, Malaysia. HCs were defined as individuals free from chronic viral (HBV, HCV, and HIV) and Mycobacterium tuberculosis infections. HBV plasma viral loads were measured using a commercial COBAS AmpliPrep-COBAS TaqMan HBV test (CAP-CTM; Roche Molecular Systems, Inc., Branchburg, NJ, USA). Plasma alanine transaminase (ALT) level in the patients was measured on a Hitachi7050 Automatic Analyzer (Hitachi Corp., Tokyo, Japan) using a commercial ALT assay kit (Wako Pure Chemicals, Osaka, Japan). The cutoff values were set at 20 ng/ml.
Peripheral Blood Mononuclear Cells (PBMCs)
Ten millilitres of venous blood was collected by venipuncture in lithium heparin BD Vacutainer (BD Biosciences, Franklin Lakes, NJ, USA) tubes and was kept at room temperature. PBMCs were extracted by density-gradient centrifugation using Ficoll Paque Plus (Sigma-Aldrich, St. Louis, MO, USA) overlay within 4 h post-collection. Cell viability was assessed by 0.4% trypan blue vital staining. Purified PBMCs were subsequently used in the immunophenotyping and cell culture experiments.
MAIT Cell Activation and Intracellular Staining
For intracellular cytokine staining, the cells were stimulated with PMA (100 ng/ml) and ionomycin (0.67 μM) for 5 h at 37°C and 5% CO2 prior to immunostaining. Brefeldin A (10 g/ml) was added for the last 4 h of stimulation. The immunostained samples were washed twice prior to acquisition on a FACS Canto II Immunocytometry system (BD Biosciences).
Multicolor Flow Cytometry
All antibodies were pre-titrated to determine appropriate working concentrations. All antibodies were purchased from BD Pharmingen™ (BD Biosciences) unless otherwise specified. Immunostaining was performed with two panels for surface markers, where the first one included fluorescein isothiocyanate (FITC)-conjugated anti-CD57, phycoerythrin (PE)-conjugated anti-TCR-Va7.2 (MiltenyiBiotec), peridinin chlorophyll protein (PerCp)-Cy5.5-conjugated anti-CD3, PE-Cy7-conjugated anti-TIM3 (eBioscience), allophycocynanin (APC)-conjugated anti-CD161, APC-H7-conjugated anti-CD8, V500-conjugated CD4, and brilliant violet 421 (BV421)-conjugated anti-CD279 (PD-1). The second panel was performed with FITC-conjugated anti-HLA-DR, PE-conjugated anti-CD38, PerCP-Cy5.5-conjugated anti-CD3, PE-conjugated anti-TCR-Va7.2-Vio770 (MiltenyiBiotec), APC-conjugated anti-CD161, APC-H7-conjugated anti-CD8, V500-conjugated CD4, and BV-421-conjugated anti-CTLA-4. The functional assays were stained using two panels; one with FITC-conjugated anti-IFN-γ, PerCp-Cy5.5-conjugated anti-CD3, APC-conjugated anti-CD161, APC-H7-conjugated anti-CD8, PE-conjugated anti-perforin (eBioscience), BV421-conjugated anti-Granzyme-B, and the other with FITC-conjugated anti-IFN-γ, PE, PerCp-Cy5.5-conjugated anti-CD3, APC-conjugated anti-CD161, APC-H7-conjugated anti-CD8, PE-conjugated anti-TNF-alpha (R&D), and PE-Vio770-conjugated anti-TCRVα7.2 (MiltenyiBiotec). Unstained PBMCs and control samples incubated with isotype-matched antibodies of irrelevant specificity were used as controls. After adding the antibodies, the cells were incubated at 4°C in the dark for 30 min and washed twice with washing buffer at 4°C. Finally, 350 μl of washing buffer (PBS, 1% BSA or 10% FBS, 0.1% sodium azide) was added to each tube. The sample tubes were analyzed using a BD FACS Canto II flow cytometer within 1 h post-staining. Flow cytometry analysis was made using FlowJo for Windows, version 10.0.8 (FlowJo LLC, Ashland, OR, USA).
Statistical Analysis
The primary analysis was to compare the percentages and expression levels (mean fluorescence intensity, MFI) of biomarkers on different subsets of T cells and MAIT cells, and compare between the three study groups. Difference between categorical variables were tested using chi-square test or Fisher’s exact test, whereas continuous variables were tested using the non-parametric Kruskal–Wallis test for multiple group comparisons. If P-values were <0.05, 3-way comparisons were subsequently performed separately using Mann–Whitney U tests between the three patient groups applying the Benjamini–Hochberg correction for multiple comparisons. Correlation between two continuous variables was compared using the Spearman’s rank correlation. Differences were considered significant with *P < 0.05, ** < 0.01, *** < 0.001, and **** < 0.0001. All analyses and graphs were performed using GraphPad Prism 6 software (GraphPad, La Jolla, CA, USA).
Results
Patient Cohort Characteristics
The three groups, non-randomized study design consisted of samples from 21 subjects with chronic HBV infection: Group 1 (G1) 12 CHB DNAemia positive, G2 with 9 CHB DNAemia negative, and a control group (G3) of 13 HCs. The specimens were collected between April and August 2015. CHB infection was defined as subjects who tested positive for HBsAg for at least 6 months after the onset of acute infection (1). There were no significant differences for demographic, laboratory, and clinical data among the three study groups (Table 1).
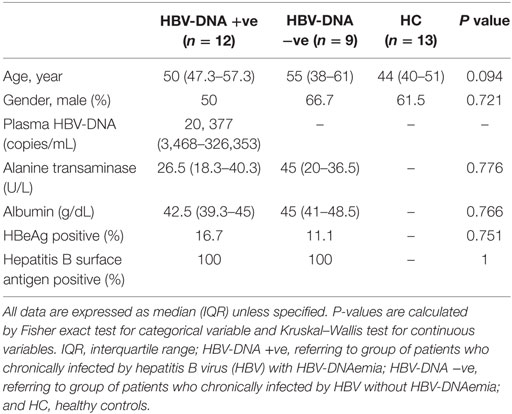
Table 1. Demography, laboratory, and clinical characteristics of study participants.
Chronic HBV Infection Is Associated with Increased Immunosenescence and Exhaustion in CD4+ and CD8+ T Cells
T-cell co-inhibition plays a critical role in T-cell dysfunction during chronic viral infections (31). In order to characterize T-cell co-inhibition during chronic HBV infection, we measured the percentages and expression levels of markers of immunosenescence, i.e., CD57, immune exhaustion, i.e., PD-1, TIM-3, and CTLA-4, as well as chronic immune activation, i.e., CD38, and HLA-DR, in both CD4+ and CD8+ T cells, and compared between the three study groups (Figure 1A). We found that there was ~twofold higher frequencies of CD4+ T cells that expressed CD57 (median = 27.5%) in the HBV-DNA +ve group as compared to HCs (14%) (Figure 1B). The percentage of PD-1+CD8+ T cells was higher in both groups of CHB patients (median = 38.5 and 36.5%, respectively) as compared to HCs (median = 25%). This result was consistent with the expression levels of PD-1 (measured as MFI) where the MFI of PD-1 were also significantly higher in CD8+ T cells in both CHB groups (1.4- and 1.27-fold higher in CHB DNA +ve and DNA −ve, respectively) as compared to HCs. The expression of PD-1 on CD4+ T cells was not significantly different among the three study groups. In addition, the percentages and expression of CTLA-4 was higher on both CD4+ and CD8+ T cells of CHB patients compared to HCs. The percentage and expression of CD38 was lower in CD8+ T cells of CHB patients relative to HCs; while the percentages of HLA-DR were significantly higher in CHB patients as compared to HCs. Collectively, our data suggest that there is an increase in immune senescence and exhaustion across both CD4+ and CD8+ T cells at varying levels in CHB patients (Figure 1B).
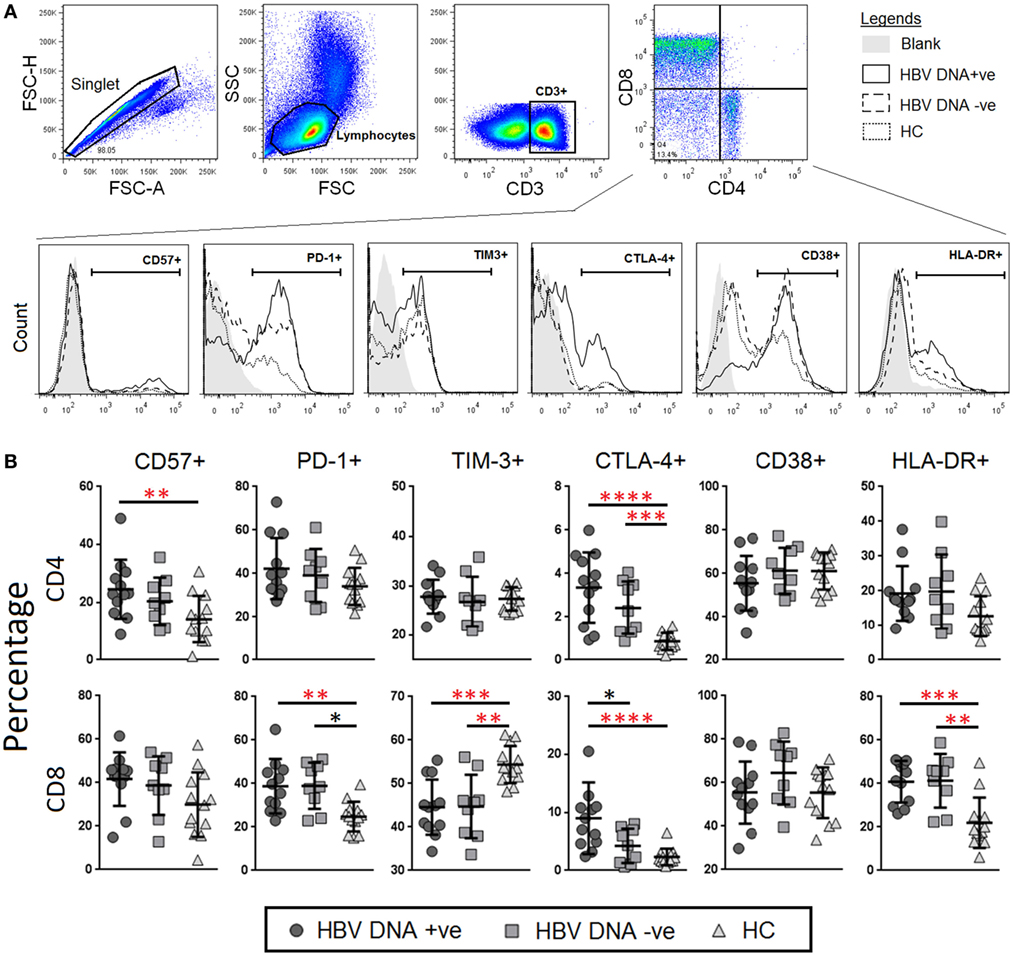
Figure 1. Frequencies and levels of immune senescence, exhaustion, and activation markers expressed on CD4+ and CD8+ T-cells between chronic hepatitis B virus (HBV)-infected patients with (circle) and without (square) HBV-DNAemia and healthy control (HC) (triangle). (A) The gating strategy to identify (B) expression levels of CD57, PD-1, TIM-3, CTLA-4 CD38, and HLA-DR on CD4+ and CD8+ T-cells. Levels of surface markers were compared across the three patient groups and post hoc Mann–Whitney U tests were then performed for those biomarkers with a Kruskal–Wallis test P value of <0.05 (*P < 0.05, **<0.01, ***<0.001, and ****<0.0001). P-values remained significant after Benjamini–Hochberg correction of multiple comparisons (marked in red*).
Chronic HBV Infection Was Associated with Depletion of TCR iVα7.2+ MAIT Cells Together with Increased Frequencies and Expression of PD-1 and CTLA-4 on TCR iVα7.2+ MAIT Cells
To understand the response of MAIT cells and their potential role in chronic HBV infection, we examined the profile of co-expression of TCR iVα7.2 and CD161 on CD3+ lymphocytes (Figure 2A). Unlike CD4+ and CD8+ T cells (Figure S1 in Sup-plementary Material), the frequencies of TCR iVα7.2+ CD161+ subset were significantly reduced in the peripheral blood of chronic HBV-infected groups as compared to HCs (P < 0.05) (Figure 2B). In addition, there was an increase in the frequencies of CD57, PD-1 and CTLA-4 as well as HLA-DR expression in both HBV-DNA +ve and DNA −ve patient groups as compared to HCs. Of note, the HBV patients generally had a ~30-fold higher percentage of CD57+ MAIT cells as compared to HCs. Furthermore, the expression of PD-1 and CTLA-4 was significantly higher in HBV-DNA +ve patients as compared to HBV-DNA −ve patients (P < 0.00001 and P < 0.05, respectively). The percentage of PD-1 and CTLA-4 on MAIT cells were significantly higher in HBV-DNA +ve patients compared to HBV-DNA −ve patients (Figure 2C). In addition, the percentage and expression of CD38 and HLA-DR were generally higher among HBV patient groups than HCs, and there were an enhanced in the expression of CTLA-4 and CD38 (Figure 2D). We also found that the expression of PD-1 and TIM-3 were directly correlated in CD4+, CD8+ T cells, and MAIT cells (Figure S2 in Supplementary Material).
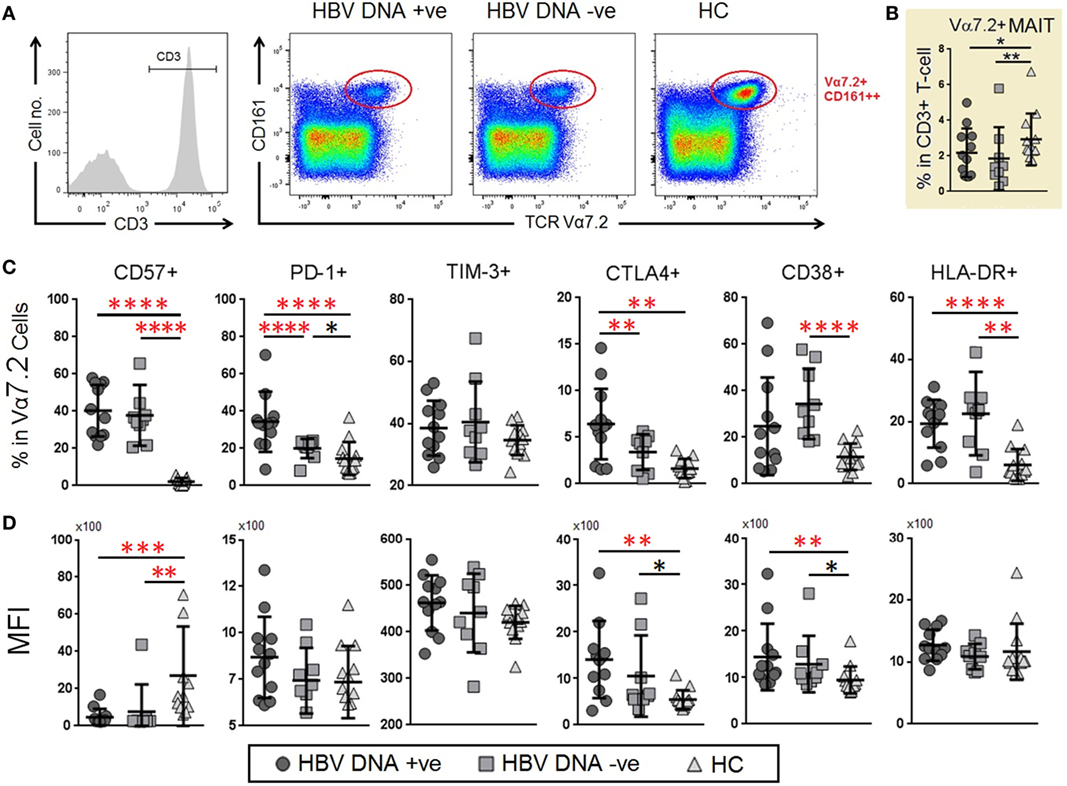
Figure 2. Frequencies and levels of immune senescence, exhaustion, and activation markers expressed on mucosal-associated invariant T (MAIT) cells between chronic hepatitis B virus (HBV)-infected patients with (G1, circle) and without (G2, square) HBV-DNAemia and healthy control (HC) (G3, triangle). (A) The gating strategy to identify CD3+TCR Vα7.2+ MAIT cells and Vα7.2+CD161− T cells. (B) Comparison of T cell receptor (TCR) Vα7.2+ cells frequencies across the three groups comparison of percentage. (C) Expression levels [mean fluorescence intensity (MFI)] (D) of CD57, PD-1, TIM-3, CTLA-4 CD38 and HLA-DR on TCR Vα7.2+ MAIT cells between the three patient groups. Levels of surface markers were compared across the three patient groups and post hoc Mann–Whitney U tests were then performed for those biomarkers with a Kruskal–Wallis test P value of <0.05 (*P < 0.05, **<0.01, ***<0.001, and ****<0.0001). P-values remained significant after Benjamini–Hochberg correction of multiple comparisons (marked in red*).
Decrease of TCR iVα7.2+ MAIT Cells Was Associated with Chronic Immune Activation and Immunosenescence
Given that the frequency of MAIT cells was decreased in chronic HBV infection, we next asked whether the degree of reduction was associated with the levels of viral replication or with markers of immunosenescence or chronic immune activation. We compared the frequencies of MAIT cells with levels of HBV-DNAemia, CD57, PD-1, CTLA-4, and HLA-DR using Spearman’s rank test. The frequency of MAIT cells did not correlate with the levels of HBV plasma DNAemia, but inversely correlated with expression (MFI) levels of HLA-DR on CD4+ T-cells and MAIT cells as well as with expression levels of CD57 on CD8+ T-cells (Figure 3). These data indicate that the depletion of MAIT cells may not directly due to HBV infection but rather due to persistent immune activation and immune senescence induced by chronic HBV infection.
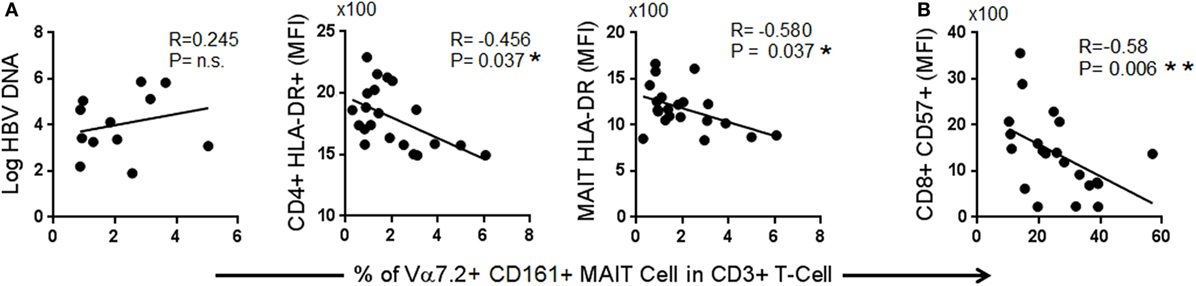
Figure 3. Spearman correlation between frequencies of T cell receptor Vα7.2+ CD161+ mucosal-associated invariant T (MAIT) with (A) markers of immune activation and (B) markers of immune senescence.
Expression of PD-1 and CTLA-4 on MAIT Cells Correlated Positively with the Plasma Levels of HBV-DNA
In the HBV-DNA +ve patients, we examined the immune correlate that could be associated with the plasma levels of HBV-DNAemia. In order to answer this question, using Spearman’s rank test, we compared the plasma levels of HBV-DNA (copies/μL) with percentages and expression (MFI) of CD57, PD-1, TIM-3, CTLA-4, CD38, and HLA-DR on CD4+, CD8+ T-cells and TCR Vα7.2+ CD161+ MAIT cells. Our analyses showed that the plasma levels of HBV-DNA positively correlated with the MFI of PD-1 (r = 0.61, P = 0.036) and frequency of HLA-DR expression on CD4+ T-cells (r = 0.00601, P = 0.039); MFI of CTLA-4 (r = 0.587, P = 0.045) and frequency of HLA-DR expression (r = 0.601, P = 0.039) on CD8+ T-cells and well as frequency of PD-1 expression (r = 0.601, P = 0.039), MFI of PD-1 (r = 0.608, P = 0.036), and MFI of CTLA-4 (r = 0.699, P = 0.011) on TCR Vα7.2+ CD161+ MAIT cells. Interestingly, we also found that the percentage of CD8+CD38+ T-cells inversely correlated with plasma levels of HBV-DNA (r = −0.678, P = 0.015) (Figure 4A).
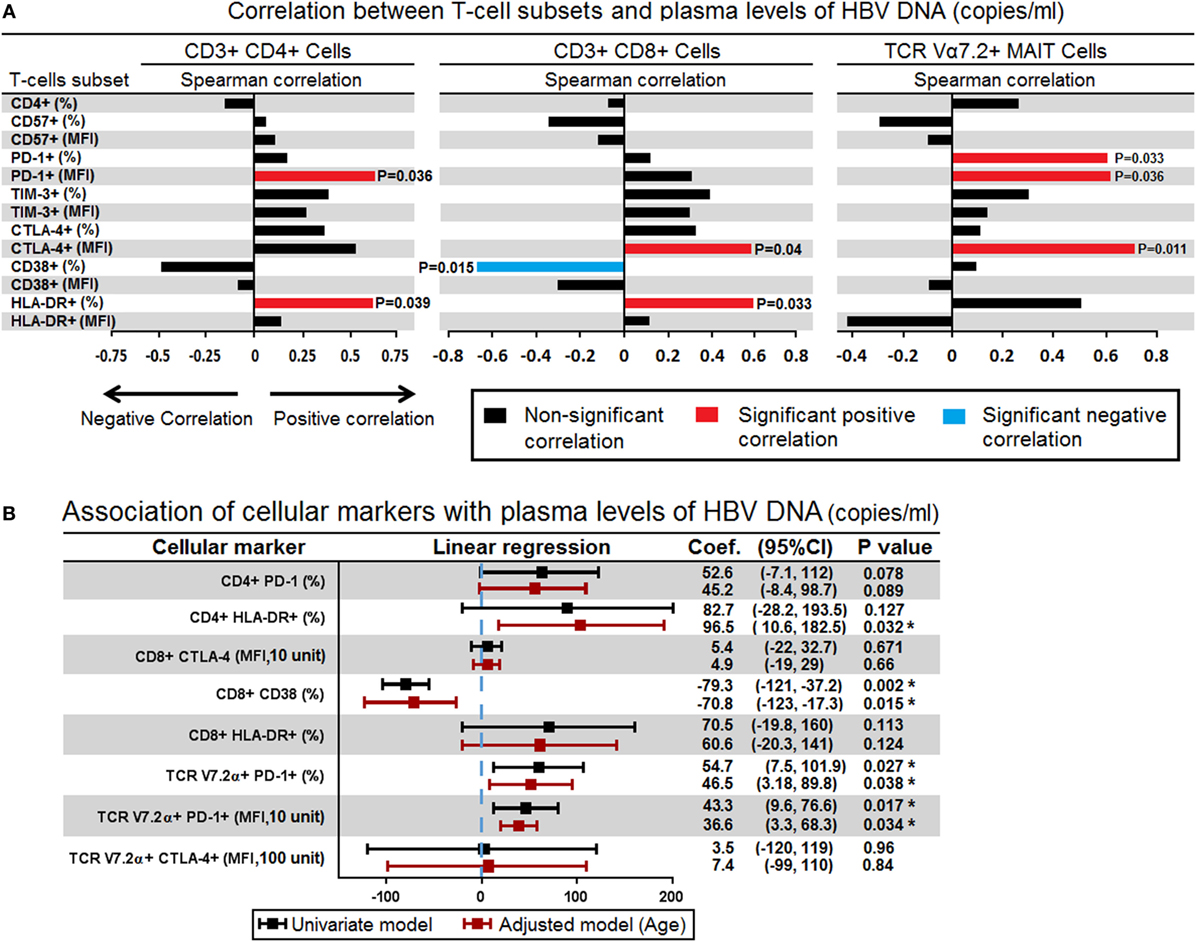
Figure 4. (A) Spearman correlations between surface markers in CD4+ (left panel), CD8+ (middle panel), and mucosal-associated invariant T (MAIT) cells (right panel) with level of plasma hepatitis B virus (HBV)-DNA. The bar represents the strength of association (r values) where red bar represents significant positive correlation, blue bar represent significant negative association and black bar represents P value > 0.05 (non-significant association) (<0.05, **<0.01, ***<0.001, and ****<0.0001). (B) Association of all surface markers that showed significant correlation with plasma HBV-DNA levels were assessed in simple logistic regression model and adjusted for age. Coefficient values below or above threshold levels were displayed in a forest plot; median and 95% CI were calculated. CI, confidence interval (*P < 0.05).
Given that these surface markers may also be influenced by age, next we investigated the association between these surface markers and plasma levels of HBV-DNA using linear regression model and adjusted by age. We found that frequency and expression (MFI) of PD-1 on TCR Vα7.2+ CD161+ MAIT cells were associated with plasma levels of HBV-DNA; every 1% increase in the frequency of PD-1 expressing cells and 10 unit increase in the MFI of PD-1 were associated with increased plasma HBV-DNA levels by an average of 46.5 (P = 0.038) and 36.2 copies/μL (P = 0.034), respectively. In contrast every 1% increase in the frequency of CD8+CD38+ T-cells was associated with decreased plasma HBV-DNA levels by an average of 70.8 copies/μL (P = 0.015) (Figure 4B). These results indicated that the expression PD-1 on TCR iVα7.2 MAIT cells and CD38 on CD8 cells played significant role in controlling the chronic HBV infection.
Expression of PD-1 on TCR Vα7.2 MAIT Cells Correlated Inversely with Levels of Plasma ALT
It has previously been shown that the levels of serum alanine aminotransferase (ALT) are linked to a more robust immune response that can lead to seroclearance of HBeAg as well as decreased plasma HBV-DNA loads (32, 33). Here, we failed to find a correlation between ALT levels and plasma levels of HBV-DNA (Figure 5A). The ALT levels were also not correlated with frequencies of TCR Vα7.2+ MAIT cells (Figure 5B). However, the frequencies and MFI of several inhibitory molecules, especially TIM-3 and PD-1 expressed on CD4+, CD8+ T cells, and TCR Vα7.2+ MAIT cells, were found to inversely correlate with the levels of ALT. These results further supported our finding that PD-1 plays an important role in persistent HBV infection (Figure 5C).
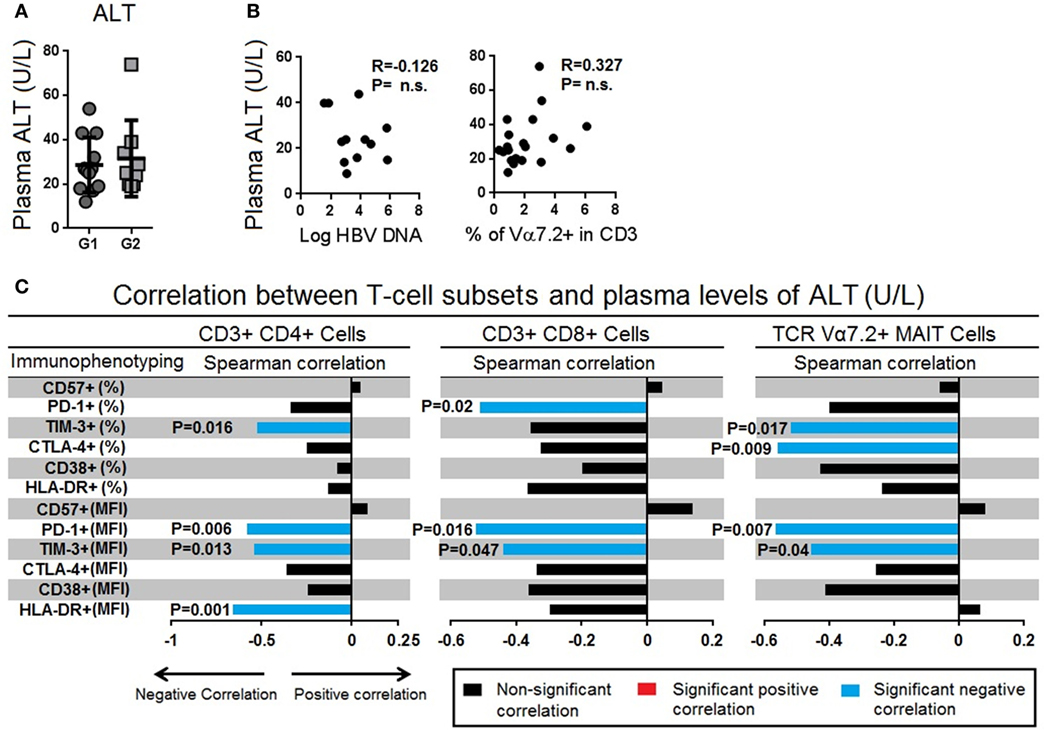
Figure 5. (A) Comparison of serum alanine transaminase (ALT) levels between chronic hepatitis B virus (HBV)-infected patients with HBV-DNAemia (circle) and without HBV-DNAemia (square). (B) Spearman correlation between serum ALT levels and frequencies of T cell receptor (TCR) Vα7.2+ CD161+ mucosal-associated invariant T (MAIT) cells. (C) Spearman correlation between frequencies and expression levels of CD57, PD-1, TIM-3, CTLA-4 CD38, and HLA-DR with serum level of ALT. The bar represents the strength of association (r values) where red bar represents significant positive correlation, blue bar represent significant negative association and black bar represents P value > 0.05 (non-significant association) (<0.05, **<0.01, ***<0.001, and ****<0.0001).
TCR iVα7.2+ MAIT Cells of Chronic HBV-Infected Patients Were Functionally Impaired in Granzyme-B and IFN-γ Production
Given that the frequency of TCR Vα7.2+ MAIT cells was reduced and expressed higher levels of PD-1 and CTLA-4 in chronic HBV-infected patients, we examined if this phenotype was associated with functional impairment by performing intracellular staining for perforin, granzyme-B, IFN-γ, and TNF-α (Figure 6A). Our results showed that the frequencies of cells producing IFN-γ, TNF-α were lower (Figure 6B), while expression (MFI) of perforin and granzyme-B were higher in chronic HBV-infected patients as compared to HCs (Figure 6C). Furthermore, results from ex vivo PMA stimulation experiments showed that the TCR Vα7.2+ MAIT cells of chronic HBV-infected patients had lower expressions of perforin, granzyme-B, and IFN-γ compared to HCs (Figure 6C). Analyses using Wilcoxon signed rank test for the samples before and after PMA stimulation showed that the production of cytokines were generally increased in all patients groups after PMA stimulation except for granzyme-B in HBV-infected patients where changes were not significantly altered after PMA stimulation (Figure S3 in Supplementary Material). Further analyses also showed that levels of perforin, granzyme-B and IFN-γ were ~4-, ~25-, and ~5-fold lower, respectively, in HBV-infected patients compared to HCs (Figure 6D). Together, these results indicate that there was a functional impairment in TCR Vα7.2+ MAIT cells.
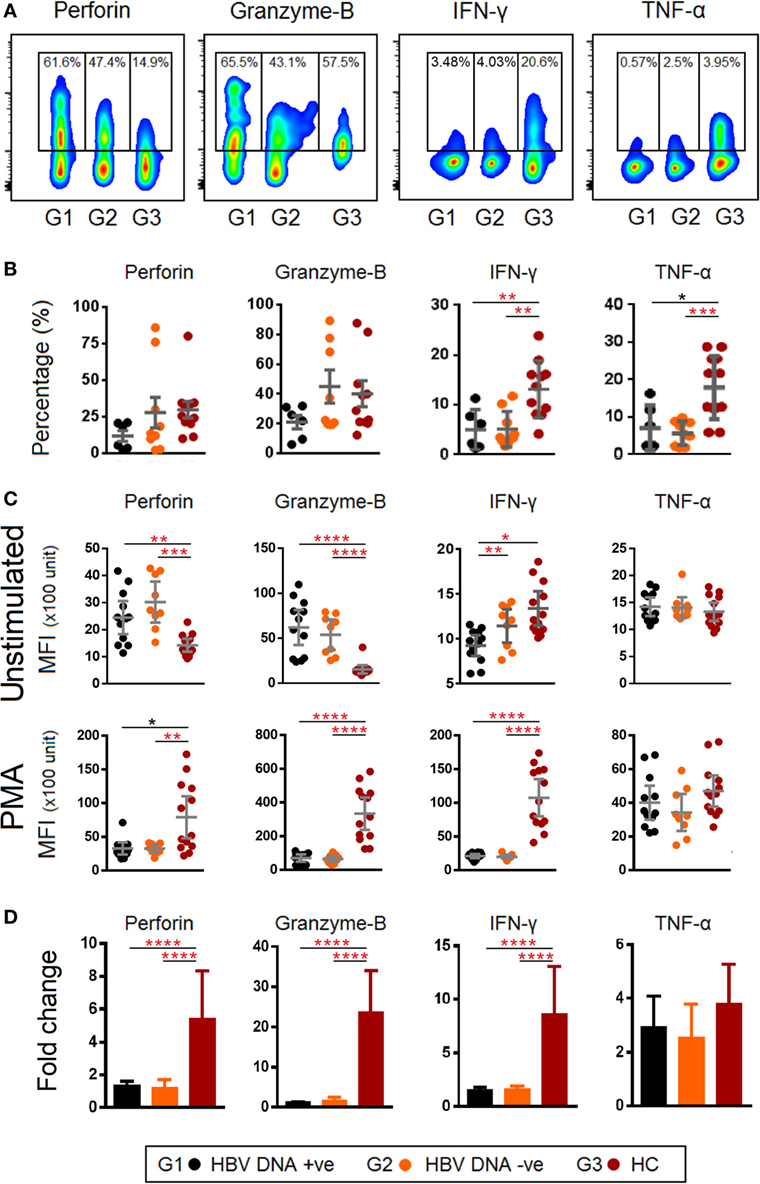
Figure 6. (A) Representative density dot plots for perforin, granzyme-B, IFN-γ, and TNF-α (without stimulation) in T cell receptor (TCR) Vα7.2+ mucosal-associated invariant T (MAIT) cells in chronic hepatitis B virus (HBV)-infected patients with HBV-DNAemia (G1) and without HBV-DNAemia (G2) and healthy control (HC) (G3). (B) Comparisons of percentages of perforin, granzyme-B, IFN-γ, and TNF-α in TCR Vα7.2+ across the three groups. (C) Comparisons of expression levels [mean fluorescence intensity (MFI)] of perforin, granzyme-B, IFN-γ, and TNF-α in TCR Vα7.2+ before and after PMA stimulation. (D) Expression (measured as fold change of MFI) of perforin, granzyme-B, IFN-γ, and TNF-α in TCR Vα7.2+ after PMA stimulation. Levels of cytokine expression were compared across the three patient groups and post hoc Mann–Whitney U tests were subsequently performed for those biomarkers using a Kruskal–Wallis test (level of significance P < 0.05) (*P < 0.05, **<0.01, ***<0.001, and ****<0.0001). P-values remained significant after Benjamini–Hochberg correction of multiple comparisons (marked in red*).
Discussion
Upregulation of co-inhibitory molecules on CD8+ and CD4+ T-cells has been shown to be associated with persistent viral replication in the setting of chronic viral infections (34–39), however the role of immune exhaustion in MAIT cells remain less well understood. Given that the TCR Vα7.2+ MAIT cells are found in circulation and enriched in mucosal compartment and liver, we postulate that the levels of immune exhaustion on MAIT cells are secondary to ongoing HBV infection and may hamper HBV viral clearance. In our study, we found that the frequencies of MAIT cells were reduced among chronic HBV-infected patients. More importantly, the immune checkpoint PD-1 was over expressed in chronic HBV-infected patients with HBV-DNAemia compared to other groups and the frequency and expression levels of PD-1 on MAIT cells were predictive of the levels of HBV-DNA in plasma. MAIT cells from chronic HBV-infected patients were producing low levels of granzyme-B and IFN-γ after stimulation.
Mucosal-associated invariant T cells are activated either by recognizing riboflavin metabolites produced by a wide range of microbes (including bacteria and fungi) presented on MHC-Ib-related protein (MR1) (40, 41), or independently of the TCR by stimulation by pro-inflammatory cytokine such as IL-12 and IL-18 (42, 43), which could occur during viral infections (23, 27). Indeed, TLR8 agonists have been shown to activate intrahepatic MAIT cells by inducing the expression of IL-12 and IL-18 by intrahepatic monocytes (42). Upon activation, MAIT cells respond rapidly with cellular cytotoxicity (44) as well as secretion of pro-inflammatory and antiviral cytokines such as IFN-γ, and TNF-α (19, 45). Given the important role of MAIT cell responses in the mucosa, we also found a decreased frequency of MAIT cells in the blood of patients chronically infected with HBV.
The frequency of MAIT cells was not correlated with the levels of HBV plasma DNAemia, but correlated with expression of immune activation and immune senescence markers on CD4+ and CD8+ T cells, suggesting the depletion of MAIT cell could likely be due to immune activation and senescence driven by chronic HBV infection. PD-1 was upregulated on MAIT cells, suggesting that they may be functionally exhausted in chronic HBV infection. This was also evident from intracellular staining experiments that indicated a marked reduction in the induction of granzyme-B and IFN-γ among HBV-infected patients. Furthermore, the MFI of IFN-γ expressing MAIT cells in HBV-DNA −ve patients were higher compared to HBV-DNA +ve patients indicating that the MAIT cells’ functions may be partially normalized in HBV-DNA −ve patients. These findings highlight the importance of IFN-γ in seroclearance of HBV-DNA.
PD-1 on the other hand, is a negative regulator of immune responses and is predominantly expressed on activated T cells (46, 47), while the ligand of PD-1, PD-1L is expressed on activated antigen presenting cells such as DCs (46, 47). The interaction between PD-1 and PD-L1 transmits a bidirectional inhibitory signal that limits downstream activation, proliferation, and cytokine production (48, 49). Though expression of PD-1 has been associated with tissue protection against severe injury by exacerbated immune responses (48, 49), the same mechanism has also been implicated in facilitating residual viral replication and persistent viral infections as the blockade of PD-1 restores T-cells functions against chronic viral infections including lymphocytic choriomeningitis virus (50), Epstein–Barr virus (51), and HIV, where the viral load is reduced in infected humanized mice (52). In this study, despite the elevated expression of several markers of T-cell dysfunction on CD4+ and CD8+ T cells and MAIT cells in patients with chronic HBV infection, the frequency of PD-1 expressing MAIT cells was the only marker that clearly differentiated between HBV-infected patients with HBV-DNAemia, those without HBV-DNAemia, and HC. Furthermore, there was a positive correlation of both frequency and level of expression of PD-1 on MAIT cells with plasma levels of HBV-DNA and a negative correlation with serum ALT. Taken together, our results suggest that MAIT cells may play a role in controlling HBV replication, and that elevated HBV-DNA levels results in MAIT cell depletion and exhaustion; while suppression of HBV replication in the liver improves MAIT cell function the numbers do not appear to recover in the absence of DNAemia.
Plasma HBV-DNA levels are an important predictor for HBV disease progression to clinical end-point. Both case–control and cohort studies have shown a significant, level-dependent association between HBV-DNA and the subsequent risk of cirrhosis (53–55) and HCC (56–58). Intervention studies also showed the same association between levels of serum HBV-DNA and disease progression seen on liver biopsy (59, 60). Our results showed that the level of PD-1 expression on MAIT cells was predictive of the plasma HBV-DNA levels, which suggests that MAIT cell functions may be restored by blocking PD-1 expression. It remains to be seen whether such a therapy would have any effect on the risk of HBV clinical end-point disease. Further studies by blocking co-inhibitory molecule expression in HBV-infected patients may be warranted.
In future studies, it will be important to study MAIT cells in the hepatic compartment. While peripheral blood MAIT cells were found to be depleted in this study, the intrahepatic population appears to be preserved in chronic hepatitis B infection, although not in end-stage disease (37). It remains to be determined whether intrahepatic MAIT cells in chronic hepatitis B infection are functionally exhausted, as they are in the periphery, and whether they can still respond normally to TCR and cytokine-mediated stimulation.
In summary, we have characterized for the first time the effect of chronic HBV infection on peripheral blood MAIT cells by investigating the expression of co-inhibitory molecules and cytotoxicity molecules, and level of cytokines production. These findings suggest a possible role for MAIT cells in the control of HBV replication, and that failure to control infection results in MAIT cell functional exhaustion.
Ethics Statement
This study was carried out in accordance with the recommendations of “name of guidelines, name of committee” with written informed consent from all subjects. All subjects gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by the Medical Ethics Committee of the UMMC Kuala Lumpur, Malaysia (MEC201311-0496).
Author Contributions
YY, AS, HT, RB, PE, and ES conceived and designed the experiments; TY, AS, RB, and PE carried out experiments and data analysis; YY, ES, MR, JR, and ML documented the findings; AK, MR, JU, and VaV selected patient samples and collected the clinical data for patients. RV, IC, AM, JU, VVe, JR, ML, and ES wrote the manuscript; VVe, AA, AM, JU, and ML contributed reagents and analysis tools; AA, RV, JU, IC, VVe, VaV, and ML provided critical inputs to the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to all the participants, clinical, para-clinical and laboratory staff of University of Malaya Medical Centre for assistance with patient recruitment, specimen collection and cooperation. The authors are grateful to Dr. Vetri V. Sakkaravarthi for providing valuable inputs to the contents of the manuscript.
Funding
This work was supported by a grant from the Frontier Research Grant (FRG), FG019-17AFR to Mohamed Rosmawati. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at https://www.frontiersin.org/articles/10.3389/fimmu.2018.00472/full#supplementary-material.
Figure S1. Comparison of percentages of CD4+ and CD8+ T-cells between chronic hepatitis B virus (HBV)-infected patients with (circle) and without (square) HBV-DNAemia and healthy control (triangle).
Figure S2. Spearman correlation between percentage and MFI of TIM-3 and PD-1 in CD4+, CD8+ and MAIT cells.
Figure S3. Changes in perforin, granzyme-B, IFN-γ and TNF-α expression following stimulation by PMA. Data were analysed using Wilcoxon matched-pairs test for paired analyses within each study group. *P < 0.05, **P < 0.01, ***P < 0.001.
References
1. Lozano R, Naghavi M, Foreman K, Lim S, Shibuya K, Aboyans V, et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet (2012) 380(9859):2095–128. doi:10.1016/S0140-6736(12)61728-0
2. Bertoletti A, Gehring AJ. Immune therapeutic strategies in chronic hepatitis B virus infection: virus or inflammation control? PLoS Pathog (2013) 9(12):e1003784. doi:10.1371/journal.ppat.1003784
3. Pol S, Nalpas B, Driss F, Michel ML, Tiollais P, Denis J, et al. Efficacy and limitations of a specific immunotherapy in chronic hepatitis B. J Hepatol (2001) 34(6):917–21. doi:10.1016/S0168-8278(01)00028-9
4. Vandepapeliere P, Lau GK, Leroux-Roels G, Horsmans Y, Gane E, Tawandee T, et al. Therapeutic vaccination of chronic hepatitis B patients with virus suppression by antiviral therapy: a randomized, controlled study of co-administration of HBsAg/AS02 candidate vaccine and lamivudine. Vaccine (2007) 25(51):8585–97. doi:10.1016/j.vaccine.2007.09.072
5. Perrillo R. Benefits and risks of interferon therapy for hepatitis B. Hepatology (2009) 49(5 Suppl):S103–11. doi:10.1002/hep.22956
6. Lau GK, Piratvisuth T, Luo KX, Marcellin P, Thongsawat S, Cooksley G, et al. Peginterferon Alfa-2a, lamivudine, and the combination for HBeAg-positive chronic hepatitis B. N Engl J Med (2005) 352(26):2682–95. doi:10.1056/NEJMoa043470
7. Chang J, Guo F, Zhao X, Guo JT. Therapeutic strategies for a functional cure of chronic hepatitis B virus infection. Acta Pharm Sin B (2014) 4(4):248–57. doi:10.1016/j.apsb.2014.05.002
8. Bertoletti A, Ferrari C. Innate and adaptive immune responses in chronic hepatitis B virus infections: towards restoration of immune control of viral infection. Gut (2012) 61(12):1754–64. doi:10.1136/gutjnl-2011-301073
9. Said ZN, Abdelwahab KS. Induced immunity against hepatitis B virus. World J Hepatol (2015) 7(12):1660–70. doi:10.4254/wjh.v7.i12.1660
10. Ganem D, Prince AM. Hepatitis B virus infection – natural history and clinical consequences. N Engl J Med (2004) 350(11):1118–29. doi:10.1056/NEJMra031087
11. Loggi E, Bihl F, Chisholm JV III, Biselli M, Bontadini A, Vitale G, et al. Anti-HBs re-seroconversion after liver transplantation in a patient with past HBV infection receiving a HBsAg positive graft. J Hepatol (2009) 50(3):625–30. doi:10.1016/j.jhep.2008.08.026
12. Chen MT, Billaud JN, Sallberg M, Guidotti LG, Chisari FV, Jones J, et al. A function of the hepatitis B virus precore protein is to regulate the immune response to the core antigen. Proc Natl Acad Sci U S A (2004) 101(41):14913–8. doi:10.1073/pnas.0406282101
13. Boni C, Fisicaro P, Valdatta C, Amadei B, Di Vincenzo P, Giuberti T, et al. Characterization of hepatitis B virus (HBV)-specific T-cell dysfunction in chronic HBV infection. J Virol (2007) 81(8):4215–25. doi:10.1128/JVI.02844-06
14. Schurich A, Khanna P, Lopes AR, Han KJ, Peppa D, Micco L, et al. Role of the coinhibitory receptor cytotoxic T lymphocyte antigen-4 on apoptosis-Prone CD8 T cells in persistent hepatitis B virus infection. Hepatology (2011) 53(5):1494–503. doi:10.1002/hep.24249
15. Nebbia G, Peppa D, Schurich A, Khanna P, Singh HD, Cheng Y, et al. Upregulation of the Tim-3/galectin-9 pathway of T cell exhaustion in chronic hepatitis B virus infection. PLoS One (2012) 7(10):e47648. doi:10.1371/journal.pone.0047648
16. Wu W, Shi Y, Li S, Zhang Y, Liu Y, Wu Y, et al. Blockade of Tim-3 signaling restores the virus-specific CD8(+) T-cell response in patients with chronic hepatitis B. Eur J Immunol (2012) 42(5):1180–91. doi:10.1002/eji.201141852
17. Dusseaux M, Martin E, Serriari N, Peguillet I, Premel V, Louis D, et al. Human MAIT cells are xenobiotic-resistant, tissue-targeted, CD161hi IL-17-secreting T cells. Blood (2011) 117(4):1250–9. doi:10.1182/blood-2010-08-303339
18. Huang S, Martin E, Kim S, Yu L, Soudais C, Fremont DH, et al. MR1 antigen presentation to mucosal-associated invariant T cells was highly conserved in evolution. Proc Natl Acad Sci U S A (2009) 106(20):8290–5. doi:10.1073/pnas.0903196106
19. Le Bourhis L, Mburu YK, Lantz O. MAIT cells, surveyors of a new class of antigen: development and functions. Curr Opin Immunol (2013) 25(2):174–80. doi:10.1016/j.coi.2013.01.005
20. Reantragoon R, Kjer-Nielsen L, Patel O, Chen Z, Illing PT, Bhati M, et al. Structural insight into MR1-mediated recognition of the mucosal associated invariant T cell receptor. J Exp Med (2012) 209(4):761–74. doi:10.1084/jem.20112095
21. Lepore M, Kalinichenko A, Colone A, Paleja B, Singhal A, Tschumi A, et al. Parallel T-cell cloning and deep sequencing of human MAIT cells reveal stable oligoclonal TCRbeta repertoire. Nat Commun (2014) 5:3866. doi:10.1038/ncomms4866
22. Billerbeck E, Kang YH, Walker L, Lockstone H, Grafmueller S, Fleming V, et al. Analysis of CD161 expression on human CD8+ T cells defines a distinct functional subset with tissue-homing properties. Proc Natl Acad Sci U S A (2010) 107(7):3006–11. doi:10.1073/pnas.0914839107
23. Cosgrove C, Ussher JE, Rauch A, Gartner K, Kurioka A, Huhn MH, et al. Early and nonreversible decrease of CD161++/MAIT cells in HIV infection. Blood (2013) 121(6):951–61. doi:10.1182/blood-2012-06-436436
24. Ussher JE, Phalora P, Cosgrove C, Hannaway RF, Rauch A, Gunthard HF, et al. Molecular analyses define Valpha7.2-Jalpha33+ MAIT cell depletion in HIV infection: a case-control study. Medicine (2015) 94(29):e1134. doi:10.1097/MD.0000000000001134
25. Saeidi A, Buggert M, Che KF, Kong YY, Velu V, Larsson M, et al. Regulation of CD8+ T-cell cytotoxicity in HIV-1 infection. Cell Immunol (2015) 298(1–2):126–33. doi:10.1016/j.cellimm.2015.10.009
26. Yong YK, Tan HY, Saeidi A, Rosmawati M, Atiya N, Ansari AW, et al. Decrease of CD69 levels on TCR Valpha7.2(+)CD4(+) innate-like lymphocytes is associated with impaired cytotoxic functions in chronic hepatitis B virus-infected patients. Innate Immun (2017) 23(5):459–67. doi:10.1177/1753425917714854
27. Barathan M, Mohamed R, Vadivelu J, Chang LY, Saeidi A, Yong YK, et al. Peripheral loss of CD8(+) CD161(++) TCRValpha7.2(+) mucosal-associated invariant T cells in chronic hepatitis C virus-infected patients. Eur J Clin Invest (2016) 46(2):170–80. doi:10.1111/eci.12581
28. McDonald JH. Handbook of Biological Statistics 2014 (2016). Available from: http://www.biostathandbook.com/multiplecomparisons.html
29. Saeidi A, Tien Tien VL, Al-Batran R, Al-Darraji HA, Tan HY, Yong YK, et al. Attrition of TCR Valpha7.2+ CD161++ MAIT cells in HIV-tuberculosis co-infection is associated with elevated levels of PD-1 expression. PLoS One (2015) 10(4):e0124659. doi:10.1371/journal.pone.0124659
30. Saeidi A, Chong YK, Yong YK, Tan HY, Barathan M, Rajarajeswaran J, et al. Concurrent loss of co-stimulatory molecules and functional cytokine secretion attributes leads to proliferative senescence of CD8(+) T cells in HIV/TB co-infection. Cell Immunol (2015) 297(1):19–32. doi:10.1016/j.cellimm.2015.05.005
31. Kahan SM, Wherry EJ, Zajac AJ. T cell exhaustion during persistent viral infections. Virology (2015) 47(9–480):180–93. doi:10.1016/j.virol.2014.12.033
32. Liaw YF. Hepatitis flares and hepatitis B e antigen seroconversion: implication in anti-hepatitis B virus therapy. J Gastroenterol Hepatol (2003) 18(3):246–52. doi:10.1046/j.1440-1746.2003.02976.x
33. Wenjin Z, Chuanhui P, Yunle W, Lateef SA, Shusen Z. Longitudinal fluctuations in PD1 and PD-L1 expression in association with changes in anti-viral immune response in chronic hepatitis B. BMC Gastroenterol (2012) 12:109. doi:10.1186/1471-230X-12-109
34. Raziorrouh B, Schraut W, Gerlach T, Nowack D, Gruner NH, Ulsenheimer A, et al. The immunoregulatory role of CD244 in chronic hepatitis B infection and its inhibitory potential on virus-specific CD8+ T-cell function. Hepatology (2010) 52(6):1934–47. doi:10.1002/hep.23936
35. Blackburn SD, Shin H, Haining WN, Zou T, Workman CJ, Polley A, et al. Coregulation of CD8+ T cell exhaustion by multiple inhibitory receptors during chronic viral infection. Nat Immunol (2009) 10(1):29–37. doi:10.1038/ni.1679
36. Kaufmann DE, Kavanagh DG, Pereyra F, Zaunders JJ, Mackey EW, Miura T, et al. Upregulation of CTLA-4 by HIV-specific CD4+ T cells correlates with disease progression and defines a reversible immune dysfunction. Nat Immunol (2007) 8(11):1246–54. doi:10.1038/ni1515
37. Yamamoto T, Price DA, Casazza JP, Ferrari G, Nason M, Chattopadhyay PK, et al. Surface expression patterns of negative regulatory molecules identify determinants of virus-specific CD8+ T-cell exhaustion in HIV infection. Blood (2011) 117(18):4805–15. doi:10.1182/blood-2010-11-317297
38. Raziorrouh B, Heeg M, Kurktschiev P, Schraut W, Zachoval R, Wendtner C, et al. Inhibitory phenotype of HBV-specific CD4+ T-cells is characterized by high PD-1 expression but absent coregulation of multiple inhibitory molecules. PLoS One (2014) 9(8):e105703. doi:10.1371/journal.pone.0105703
39. Saeidi A, Ellegard R, Yong YK, Tan HY, Velu V, Ussher JE, et al. Functional role of mucosal-associated invariant T cells in HIV infection. J Leukoc Biol (2016) 100(2):305–14. doi:10.1189/jlb.4RU0216-084R
40. Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, et al. MR1 presents microbial vitamin B metabolites to MAIT cells. Nature (2012) 491(7426):717–23. doi:10.1038/nature11605
41. Corbett AJ, Eckle SB, Birkinshaw RW, Liu L, Patel O, Mahony J, et al. T-cell activation by transitory neo-antigens derived from distinct microbial pathways. Nature (2014) 509(7500):361–5. doi:10.1038/nature13160
42. Jo J, Tan AT, Ussher JE, Sandalova E, Tang XZ, Tan-Garcia A, et al. Toll-like receptor 8 agonist and bacteria trigger potent activation of innate immune cells in human liver. PLoS Pathog (2014) 10(6):e1004210. doi:10.1371/journal.ppat.1004210
43. Ussher JE, Bilton M, Attwod E, Shadwell J, Richardson R, de Lara C, et al. CD161++ CD8+ T cells, including the MAIT cell subset, are specifically activated by IL-12+IL-18 in a TCR-independent manner. Eur J Immunol (2014) 44(1):195–203. doi:10.1002/eji.201343509
44. Kurioka A, Ussher JE, Cosgrove C, Clough C, Fergusson JR, Smith K, et al. MAIT cells are licensed through granzyme exchange to kill bacterially sensitized targets. Mucosal Immunol (2015) 8(2):429–40. doi:10.1038/mi.2014.81
45. Godfrey DI, Uldrich AP, McCluskey J, Rossjohn J, Moody DB. The burgeoning family of unconventional T cells. Nat Immunol (2015) 16(11):1114–23. doi:10.1038/ni.3298
46. Iwai Y, Terawaki S, Ikegawa M, Okazaki T, Honjo T. PD-1 inhibits antiviral immunity at the effector phase in the liver. J Exp Med (2003) 198(1):39–50. doi:10.1084/jem.20022235
47. Karrar A, Broome U, Uzunel M, Qureshi AR, Sumitran-Holgersson S. Human liver sinusoidal endothelial cells induce apoptosis in activated T cells: a role in tolerance induction. Gut (2007) 56(2):243–52. doi:10.1136/gut.2006.093906
48. Chen L, Zhang Z, Chen W, Zhang Z, Li Y, Shi M, et al. B7-H1 up-regulation on myeloid dendritic cells significantly suppresses T cell immune function in patients with chronic hepatitis B. J Immunol (2007) 178(10):6634–41. doi:10.4049/jimmunol.178.10.6634
49. Kassel R, Cruise MW, Iezzoni JC, Taylor NA, Pruett TL, Hahn YS. Chronically inflamed livers up-regulate expression of inhibitory B7 family members. Hepatology (2009) 50(5):1625–37. doi:10.1002/hep.23173
50. Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, et al. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature (2006) 439(7077):682–7. doi:10.1038/nature04444
51. Quan L, Chen X, Liu A, Zhang Y, Guo X, Yan S, et al. PD-1 blockade can restore functions of T-cells in Epstein-Barr virus-positive diffuse large B-cell lymphoma in vitro. PLoS One (2015) 10(9):e0136476. doi:10.1371/journal.pone.0136476
52. Seung E, Dudek TE, Allen TM, Freeman GJ, Luster AD, Tager AM. PD-1 blockade in chronically HIV-1-infected humanized mice suppresses viral loads. PLoS One (2013) 8(10):e77780. doi:10.1371/journal.pone.0077780
53. Yuan HJ, Yuen MF, Ka-Ho Wong D, Sablon E, Lai CL. The relationship between HBV-DNA levels and cirrhosis-related complications in Chinese with chronic hepatitis B. J Viral Hepat (2005) 12(4):373–9. doi:10.1111/j.1365-2893.2005.00603.x
54. Chen G, Lin W, Shen F, Iloeje UH, London WT, Evans AA. Past HBV viral load as predictor of mortality and morbidity from HCC and chronic liver disease in a prospective study. Am J Gastroenterol (2006) 101(8):1797–803. doi:10.1111/j.1572-0241.2006.00647.x
55. Iloeje UH, Yang HI, Su J, Jen CL, You SL, Chen CJ, et al. Predicting cirrhosis risk based on the level of circulating hepatitis B viral load. Gastroenterology (2006) 130(3):678–86. doi:10.1053/j.gastro.2005.11.016
56. Wu CF, Yu MW, Lin CL, Liu CJ, Shih WL, Tsai KS, et al. Long-term tracking of hepatitis B viral load and the relationship with risk for hepatocellular carcinoma in men. Carcinogenesis (2008) 29(1):106–12. doi:10.1093/carcin/bgm252
57. Liu TT, Fang Y, Xiong H, Chen TY, Ni ZP, Luo JF, et al. A case-control study of the relationship between hepatitis B virus DNA level and risk of hepatocellular carcinoma in Qidong, China. World J Gastroenterol (2008) 14(19):3059–63. doi:10.3748/wjg.14.3059
58. Yang HI, Yeh SH, Chen PJ, Iloeje UH, Jen CL, Su J, et al. Associations between hepatitis B virus genotype and mutants and the risk of hepatocellular carcinoma. J Natl Cancer Inst (2008) 100(16):1134–43. doi:10.1093/jnci/djn243
59. Mommeja-Marin H, Mondou E, Blum MR, Rousseau F. Serum HBV DNA as a marker of efficacy during therapy for chronic HBV infection: analysis and review of the literature. Hepatology (2003) 37(6):1309–19. doi:10.1053/jhep.2003.50208
Keywords: HBV infection, HLA-DR, immune exhaustion, immunosenescence, mucosal-associated invariant T cells, PD-1, CTLA-4
Citation: Yong YK, Saeidi A, Tan HY, Rosmawati M, Enström P, Batran RA, Vasuki V, Chattopadhyay I, Murugesan A, Vignesh R, Kamarulzaman A, Rajarajeswaran J, Ansari AW, Vadivelu J, Ussher JE, Velu V, Larsson M and Shankar EM (2018) Hyper-Expression of PD-1 Is Associated with the Levels of Exhausted and Dysfunctional Phenotypes of Circulating CD161++TCR iVα7.2+ Mucosal-Associated Invariant T Cells in Chronic Hepatitis B Virus Infection. Front. Immunol. 9:472. doi: 10.3389/fimmu.2018.00472
Received: 02 November 2017; Accepted: 22 February 2018;
Published: 19 March 2018
Edited by:
Jason Paul Gigley, University of Wyoming, United StatesReviewed by:
Masaaki Miyazawa, Kindai University, JapanHridayesh Prakash, All India Institute of Medical Sciences, India
Copyright: © 2018 Yong, Saeidi, Tan, Rosmawati, Enström, Batran, Vasuki, Chattopadhyay, Murugesan, Vignesh, Kamarulzaman, Rajarajeswaran, Ansari, Vadivelu, Ussher, Velu, Larsson and Shankar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yean K. Yong, eW9uZy55ZWFua29uZ0BnbWFpbC5jb20=;
Esaki M. Shankar, c2hhbmthcmVtQGN1dG4uYWMuaW4=
†These authors have contributed equally to the experimental work and writing of the manuscript.