 Sara Nunes1,2
Sara Nunes1,2 Icaro Bonyek Silva1,2
Icaro Bonyek Silva1,2 Mariana Rosa Ampuero1,2
Mariana Rosa Ampuero1,2 Almério Libório Lopes de Noronha3Lígia Correia Lima de Souza2Thaizza Cavalcante Correia1
Almério Libório Lopes de Noronha3Lígia Correia Lima de Souza2Thaizza Cavalcante Correia1 Ricardo Khouri1,2
Ricardo Khouri1,2 Viviane Sampaio Boaventura1,2Aldina Barral1,2
Viviane Sampaio Boaventura1,2Aldina Barral1,2 Pablo Ivan Pereira Ramos1,4
Pablo Ivan Pereira Ramos1,4 Cláudia Brodskyn1,2Pablo Rafael Silveira Oliveira2,4
Cláudia Brodskyn1,2Pablo Rafael Silveira Oliveira2,4 Natalia Machado Tavares1,2*
Natalia Machado Tavares1,2*
- 1Oswaldo Cruz Foundation, Gonçalo Moniz Institute, FIOCRUZ, Salvador, Brazil
- 2Federal University of Bahia, Salvador, Brazil
- 3Laboratório de Anatomia Patológica, Feira de Santana, Brazil
- 4Centre for Data and Knowledge Integration for Health (CIDACS), FIOCRUZ, Salvador, Brazil
Localized cutaneous leishmaniasis (LCL) is a chronic disease characterized by ulcerated skin lesion(s) and uncontrolled inflammation. The mechanisms underlying the pathogenesis of LCL are not completely understood, and little is known about posttranscriptional regulation during LCL. MicroRNAs (miRNAs) are non-coding small RNAs that regulate gene expression and can be implicated in the pathogenesis of LCL. We investigated the involvement of miRNAs and their targets genes in human LCL using publicly available transcriptome data sets followed by ex vivo validation. Initial analysis highlighted that miRNA expression is altered during LCL, as patients clustered separately from controls. Joint analysis identified eight high confidence miRNAs that had altered expression (−1.5 ≤ fold change ≥ 1.5; p < 0.05) between cutaneous ulcers and uninfected skin. We found that the expression of miR-193b and miR-671 are greatly associated with their target genes, CD40 and TNFR, indicating the important role of these miRNAs in the expression of genes related to the inflammatory response observed in LCL. In addition, network analysis revealed that miR-193b, miR-671, and TREM1 correlate only in patients who show faster wound healing (up to 59 days) and not in patients who require longer cure times (more than 60 days). Given that these miRNAs are associated with control of inflammation and healing time, our findings reveal that they might influence the pathogenesis and prognosis of LCL.
Introduction
Leishmaniasis is a group of chronic diseases caused by intracellular protozoan parasites from the Leishmania genus that is transmitted by infected sandflies bites (1). Localized cutaneous leishmaniasis (LCL) is the most frequent form of these diseases and is characterized by ulcerated skin lesion(s) that can take a long time to heal (2). Human LCL caused by Leishmania braziliensis is associated with a chronic inflammation that is critical for parasite clearance but also for tissue injury and disease pathogenesis. The notion that the severity of LCL is mainly the result of an exacerbated inflammatory response than a consequence of high parasite burden is now supported by multiple lines of evidence (1–4).
The resolution of inflammation is a tightly regulated and active process that requires a switch in gene expression, leading to a downregulation of inflammatory mediators (5, 6). Therefore, we hypothesized that regulators of gene expression could be altered during L. braziliensis infection, allowing a hyperactive inflammatory response even when the parasite burden is controlled. MicroRNAs (miRNAs) have emerged as key players in the regulation of gene products in recent years. These molecules are non-coding, endogenous, small RNAs (19 to 25 nucleotides) that modulate expression through the repression of translation and degradation of target messenger RNAs (mRNAs) (7, 8). miRNAs mediate posttranscriptional gene silencing or downregulation by binding to mRNAs through complete or partially complementary sequences (9). Thus, miRNAs regulate multiple biological processes through the regulation of protein translation and affect the immune response. Due to these features, miRNAs have become attractive targets for diagnosis, therapy, and biomarkers in a number of human diseases (9–12).
Altered miRNA expression has been associated with many human disorders (13–15). Regarding inflammatory skin diseases, miRNA expression profiles seem to be specific for each condition (16, 17). The upregulation of miR-203 has been demonstrated in human psoriasis lesions compared with atopic eczema skin. This miRNA inhibits the expression of SOCS-3, contributing to the dysfunction of infiltrating cells and to the pathogenesis of psoriasis (16).
The evaluation of host miRNA expression profiles in humans affected by pathogen-borne diseases can drive new strategies for diagnosis and treatment. Indeed, patients with chronic Hepatitis C Virus infection treated with Miravirsen, an antisense miRNA complementary to miR-122, showed prolonged dose-dependent reduction in viral load (18). In addition, miRNAs are promising candidates as biomarkers for sepsis (19, 20). Nevertheless, few studies have evaluated the role of miRNAs in human parasitic infections, such as Cryptosporidium parvum (etiological agent of cryptosporidiosis) (21–23), Plasmodium spp. (causative agent of malaria) (24, 25), and Trypanosoma ssp. (causing sleeping sickness disease) (26–29). To the best of our knowledge, this is the first report dedicated to evaluating the role of miRNAs from lesions of L. braziliensis-infected patients. We employed an integrative, dual-phase approach and first aimed to identify miRNAs and their target genes involved in the pathogenesis of human LCL by a joint analysis of previously published expression datasets. In a second phase, we performed ex vivo validation of the expression of some implicated miRNAs and correlated them with clinical features of the patients.
Materials and Methods
Acquisition of Microarray Datasets
Expression data sets were obtained from the NCBI1 Gene Expression Omnibus (GEO) database using the search terms “human,” “biopsies,” “skin,” and “Leishmania braziliensis.” Two microarray datasets matching these criteria were selected. The first dataset (GEO accession number GSE55664), published by Novais et al. (30), applied the Illumina HT12 v4 platform (GPL10558) to analyze 10 healthy control (HC) skin samples and 25 skin biopsies from lesions of patients with LCL from Corte de Pedra, Bahia (Brazil). The second dataset (GEO accession number GSE63931), published by Oliveira et al. (31), used the Agilent Sure Print GE Human G3v2 platform (GPL17077) and compared eight control skin samples and eight skin biopsies from patients with LCL from Itabuna, Bahia (Brazil). The Ensembl/BioMart interface2 was used to screen each platform for miRNA genes, which were further classified using miRBase (see Identification of High Confidence MicroRNAs). Approximately 50,000 probes were analyzed in each dataset (Figure S1 in Supplementary Material).
Identification of High Confidence MicroRNAs
Release 21 of miRBase was used throughout our analysis. Within miRBase miRNAs are classified either as high or low confidence annotations based on standard mapped reads and deep-sequencing data (32–34). The total number of human high confidence miRNA precursors is 296, whereas 1,585 are classified as low confidence (Figure S2A in Supplementary Material). Therefore, we analyzed our data according to this miRBase standard, where miRNAs were classified as high confidence, low confidence or non-miRBase 21 miRNAs (miRNA probes were absent in miRBase release 21) in both datasets, GSE55664 and GSE63931 (Figure S2B in Supplementary Material).
MicroRNA Expression Profiles
The expression data from both platforms were analyzed using the microarray data analysis tool Multi Experiment Viewer.3 Using the log2-transformed miRNA expression values as input, unsupervised hierarchical clustering was performed using average-linkage and Euclidean distance as metrics. Heat maps were used to represent the expression of miRNA probes through signal strength, and principal component analysis (PCA) enabled the grouping of samples based on miRNAs expression. An absolute −1.5≤ fold change of ≥1.5 with FDR adjusted p-value < 0.05 were used as criteria to identify differentially expressed miRNAs.
Pathway Enrichment Analysis
Quantile-normalized data from both platforms were analyzed by Ingenuity Pathway Analysis4 to identify Canonical Pathways in Leishmaniasis. The “Core Analysis” was performed to relate pathways with “skin, dermis, epidermis, immune cell lines, and innate immune response” common to both Leishmaniasis datasets. After this analysis, the molecules within these pathways were searched at Target Scan5 as possible targets for the high confidence miRNAs differentially expressed in LCL (Figure S3 in Supplementary Material).
Ethics Statement
This study was conducted according to the principles of the Declaration of Helsinki and under local ethical guidelines. This study was approved by the Ethical Committee of the Gonçalo Moniz Institute (Salvador, Bahia, Brazil—CAAE: 47120215.8.0000.0040). All patients provided written informed consent for the collection of samples and subsequent analysis.
Patients and Biopsies
The patients were examined at the health post in the city of Jiquiriçá (State of Bahia, Brazil), which is a well-known area of L. braziliensis transmission. The criteria for diagnosis were clinical symptoms typical of LCL, histological characteristics and a positive delayed-type hypersensitivity response to L. braziliensis antigen or serology. Prior to therapy, biopsies were collected at the border of the lesion with a 4-mm punch from 12 patients (clinical information is available in Table S1 in Supplementary Material). Healthy skin control samples were collected from plastic surgery of seven donors living in a non-endemic area without history of LCL.
MicroRNA Isolation
The samples were placed in TRIzol reagent (Invitrogen). Total RNA was extracted using the miRNeasy Mini Kit (Qiagen) following the manufacturer’s instructions. RNA concentrations were measured using the NanoDrop 2000 UV-Vis Spectrophotometer (Thermo Scientific). cDNA synthesis was performed using the miScript II RT Kit with HiFlex buffer (Qiagen) following the manufacturer’s instructions.
Real-Time Quantitative PCR (RT-qPCR) Validation Assays
The relative expression of miRNAs was measured by RT-qPCR in the Jiquiriçá (Bahia, Brazil) cohort, which was composed of 12 LCL patients and 7 skin controls, as described above (see Patients and Biopsies). The assays were completed in duplicate using an ABI 7500 real-time PCR instrument (Applied Biosystems) following the manufacturer’s instructions. The expression levels of hsa-miR-155, hsa-miR-503, hsa-miR-193b, hsa-miR-99a, and hsa-miR-221 were normalized with SNORD8, SNORA70, and SNORD46 used as endogenous controls. All primers were synthesized by Integrated DNA Technologies.
Statistical Analysis
Spearman correlation analysis tests were applied using R (for the correlation matrix) and GraphPad Prism 5 software; a p-value < 0.05 was considered significant.
Results
MicroRNA Expression Profiling Is Able to Distinguish LCL Lesions From Uninfected Skin
Two publicly available transcriptome data sets of human LCL lesions caused by L. braziliensis were analyzed (Figure S1 in Supplementary Material). The first (published under accession code number GSE55664) applied the Illumina HT12 v4 platform to compare global gene expression patterns between 25 skin biopsies samples from LCL patients and 10 HC samples from uninfected skin (30). From this gene array, 792 probes (from a total of 47,305) amplify miRNAs (Table S2 in Supplementary Material). The miRNA expression profile was able to separately cluster cutaneous ulcers from healthy skin biopsies (Figure S4A in Supplementary Material). For further analysis, we used 33 miRNA probes considered to be differentially regulated (p < 0.05; ≥1.5-fold increase or decrease in expression). The expression patterns retained the ability to segregate LCL and healthy skin samples into two distinct clusters (Figure 1A). PCA confirmed this result (Figure 1B). From these miRNA probes, 17 were upregulated (red) and 16 were downregulated (green) in LCL relative to control skin (Figure 1C).
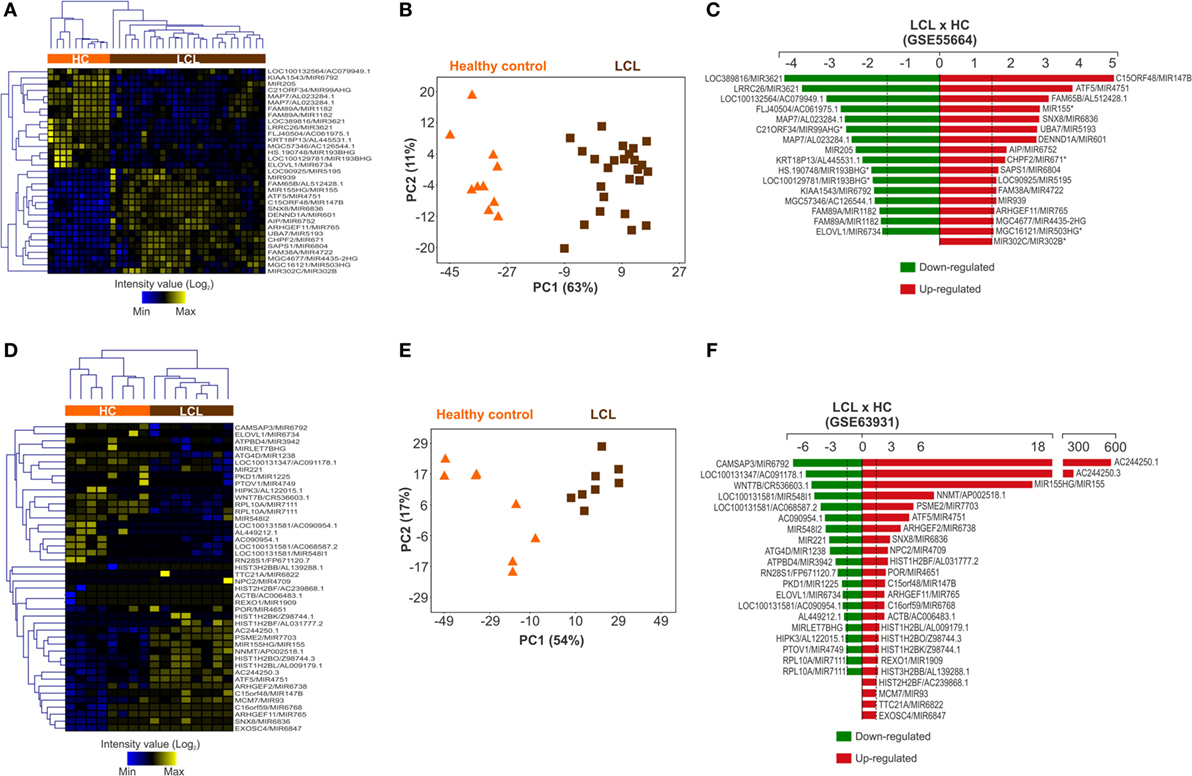
Figure 1. Expression profile of microRNAs is altered during cutaneous leishmaniasis caused by L. braziliensis. MicroRNA probes that were differentially expressed (p < 0.05 and −1.5< fold change >1.5) between healthy control (HC; orange bar) and localized cutaneous leishmaniasis (LCL; brown bar) skin samples were included in this analysis. Heat maps showing the expression profile of (A) 33 microRNA probes from GSE55664 and (B) 44 microRNA probes from GSE63931. Rows represent microRNA probes and columns represent samples. Unsupervised hierarchical clustering of the samples was performed using the Euclidean distance method; Principal Component Analysis using microRNA probes that were differentially expressed between the HC and LCL skin samples were conducted for (C) GSE55664 or (D) GSE63931 data sets. Each symbol represents one sample; fold changes of microRNA probes modulated in LCL relative to HC samples are shown for (E) GSE55664 and (F) GSE63931 data sets.
Similar results in the same direction were obtained with the second data set (published under accession code number GSE63931) that applied the Agilent Sure Print GE Human G3v2 platform to compare eight lesion biopsies from LCL patients with eight HC skin samples (31). This gene array comprised 50,737 gene probes, with only 141 probes among these covering miRNAs regions (Table S3 in Supplementary Material). Despite this small number of miRNA probes, cluster analysis was also able to segregate ulcers from normal skin samples (Figure S4B in Supplementary Material). This clustering pattern was maintained after we restricted our analysis to the 44 miRNA probes that were differentially expressed (p < 0.05; fold change ≥1.5 increase or decrease in expression) between lesions and controls (Figure 1D). This pattern was also confirmed by PCA (Figure 1E). From the 44 microRNA probes, 24 miRNAs were upregulated (red) and 20 downregulated (green) in ulcers from LCL patients compared with control skin (Figure 1F). Taken together, these data show that the miRNA expression profile is broadly modulated in skin lesions caused by L. braziliensis.
The Expression of 8 High Confidence MicroRNAs Are Modulated in L. braziliensis Lesions in Patients From Different Endemic Areas
A subset of high confidence miRNAs was identified based on the confidence criteria established by the miRBase database (32, 34–37). Considering the two datasets evaluated, we identified a total of 77 miRNA probes (covering 49 miRNAs) whose expression levels were modulated in LCL lesions (Table S4 in Supplementary Material); this subset included 23 miRNAs from the GSE55664 platform and 26 from the GSE63931 platform (Figure 2A). Of these, only eight were high confidence miRNAs (according to miRBase), and, among these, only miR-155 was common to both data sets. To technically validate these findings, we performed real-time RT-PCR of selected miRNAs using commercially validated assays (available for miR-155, miR-503, miR-193b, miR-99a, and miR-221) and fresh skin biopsies of cutaneous lesions from 12 patients living in another endemic area who had clinically diagnosed with LCL caused by L. braziliensis and seven healthy skin samples. When the fold change values between microarray data and PCR results were compared, a positive correlation (r = 0.7) between these platforms for the miRNAs tested was observed (Figure 2B). The differences in scales observed for each technique may be due to the sensitivity of the assays, the PCR primers used or the variable genetic backgrounds of the patients from different cohorts. Nevertheless, this result validated the altered expression of miRNAs in three different cohorts of LCL patients, and the RT-qPCR results are in accordance with our integrated analysis, which indicated that this approach is a useful tool to identify miRNA signatures.
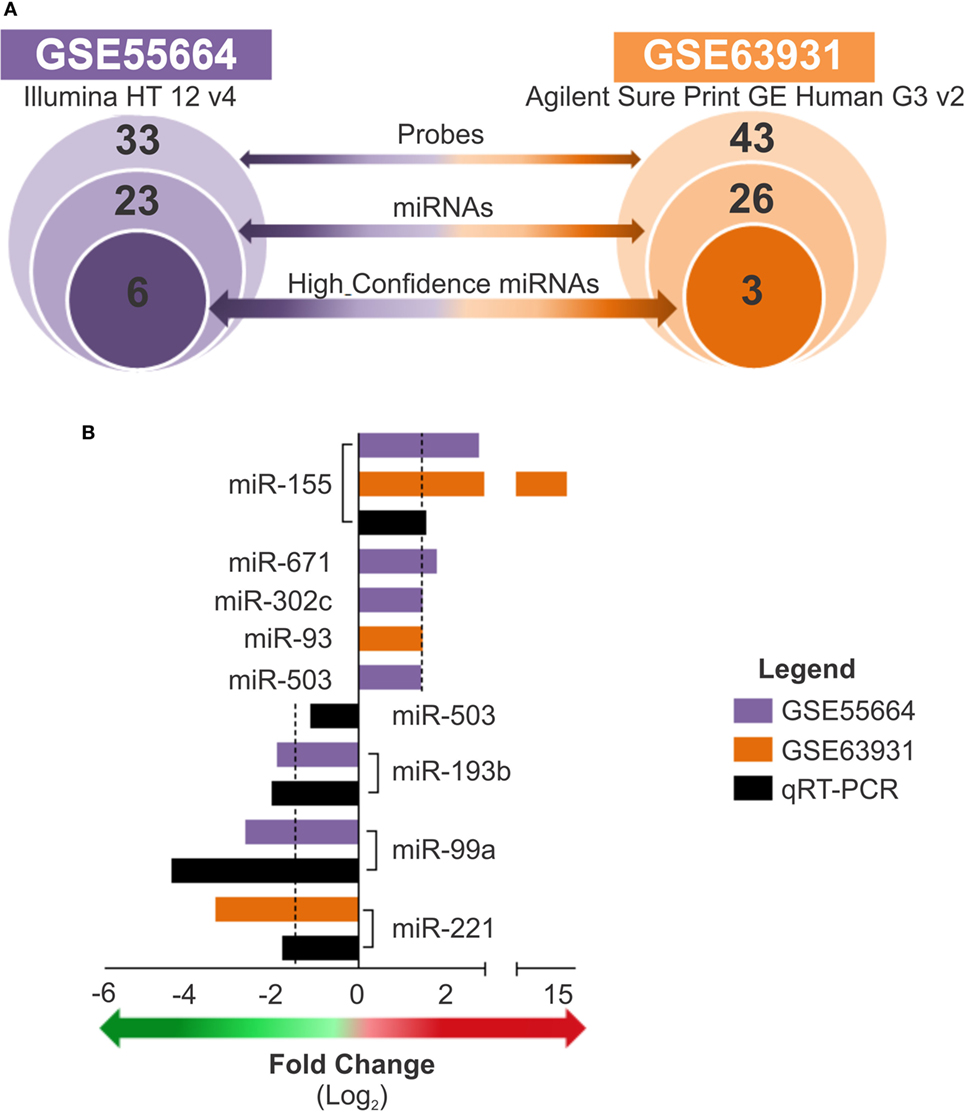
Figure 2. Expression of high confidence microRNAs is modulated in cutaneous lesions caused by Leishmania braziliensis. (A) The total number of probes, sequences, or high confidence microRNAs is presented for each data set, GSE55664 (purple) and GSE63931 (orange), from the outer to the inner circles, respectively (p < 0.05). (B) Bars graph shows the fold changes of high confidence microRNAs that differentially expressed from both datasets, GSE55664 (purple bars) and GSE63931 (orange bars), as well as real-time quantitative PCR to measure the relative expression of high confidence microRNAs in new samples from active lesions of patients with localized cutaneous leishmaniasis compared with healthy control skin for validation of the identified microRNAs in another cohort (black bars). The arrows indicate upregulated (red) or downregulated (green) expression (fold change ≥1.5).
Identification of MicroRNA Targets in Pathways Associated With Innate Immune Response During LCL
The innate immune response is crucial for host defense against Leishmania, which needs to subvert these mechanisms to establish infection (38). Therefore, modulation of innate immune response via miRNAs could represent an important aspect of LCL immunopathogenesis. To identify potential genes whose mRNAs might be targeted by miRNAs during LCL, we completed pathway enrichment analysis for both datasets using Ingenuity Pathways Analysis software (Ingenuity Pathway Analysis) (Table S5 in Supplementary Material) and focusing on canonical pathways associated with innate immune response. We found 13 different inflammatory pathways (Table S6 in Supplementary Material) associated (p < 0.05) with LCL between both datasets (Figure 3A). However, only six of these pathways remained significantly associated with the disease after Bonferroni correction (cutoff p = 0.00192308). The most significantly enriched pathways in GSE55664 data set (Figure 3B) were the same as those in the GSE63931 data set (Figure 3C), reinforcing the involvement of these pathways in the pathogenesis of LCL.
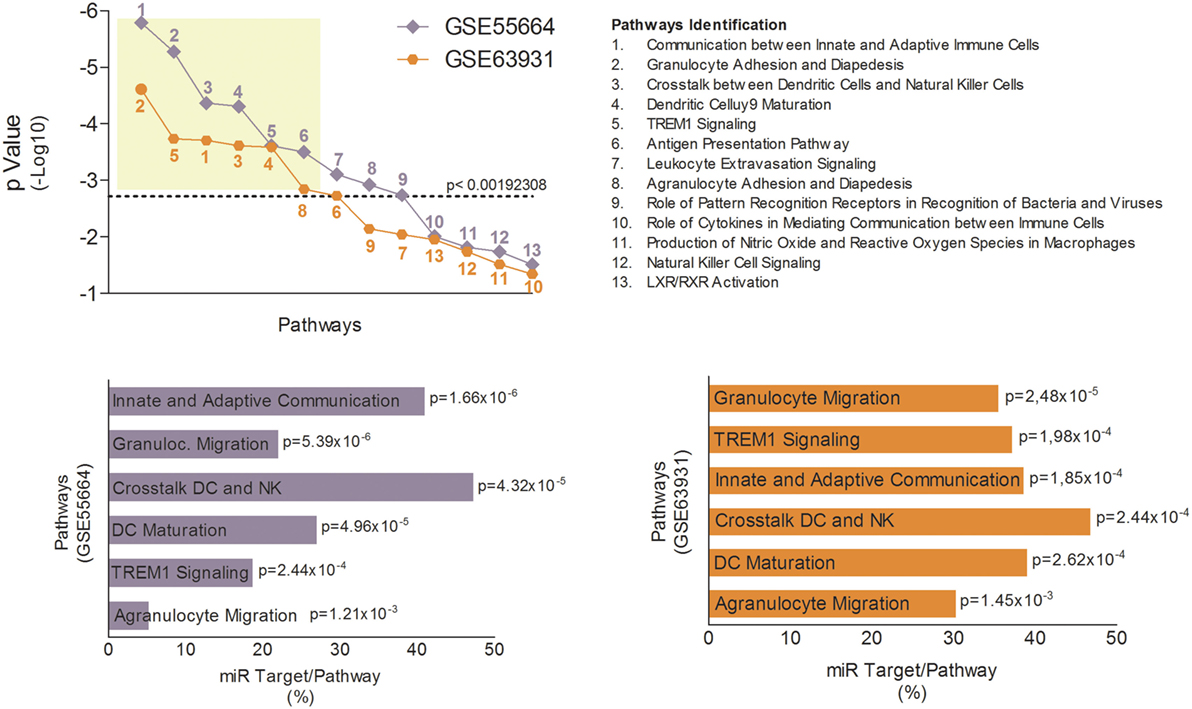
Figure 3. Canonical pathways associated with innate immune response identified in localized cutaneous leishmaniasis (LCL). (A) p-Value of 13 canonical pathways common to both data sets, GSE55664 (purple) and GSE63931 (orange), and related to the innate immune response. Each symbol represents one pathway identified by numbers and listed on the right. The most significant pathways in GSE66554 (B) and GSE63931 (C) are identified by p-value and percentage of molecules that are targets for LCL microRNAs.
Subsequently, we used the TargetScan algorithm (release 7.1) to identify genes in these pathways whose mRNAs showed consensus sequences for one or more of the eight high confidence miRNAs modulated during LCL (Figure S3 in Supplementary Material). We found, on average, that 30–40% of the molecules involved in the innate immune response against L. braziliensis are potential targets for the miRNAs identified here (Figure 3B,C)). Together, these finding suggest that the uncontrolled inflammatory response observed in LCL could also be due to an imbalance in the regulatory mechanisms mediated by miRNAs.
MicroRNA-193b and -671 and Their Targets Genes Correlate With LCL Outcome
Specific target genes for the miRNAs were defined by expression pairing analysis performed at the probe level (Figure S3 in Supplementary Material), in addition to displaying a consensus sequence for a specific miRNA. This approach compares the expression of a miRNA and its predicted mRNA target in the same corresponding sample, which allows the identification of a potential mRNA target when its expression is inversely correlated with the expression of a given miRNA (39). Thus, only a small subset of the total genes predicted by TargetScan were defined as targets for the miRNAs (Figure 4A). In fact, six out of eight high confidence miRNAs did not have mRNA targets matching those criteria. We found only STAT3 as a potential target for miR-671 and 11 target genes for miR-193b considering both data sets (Figure 4A).
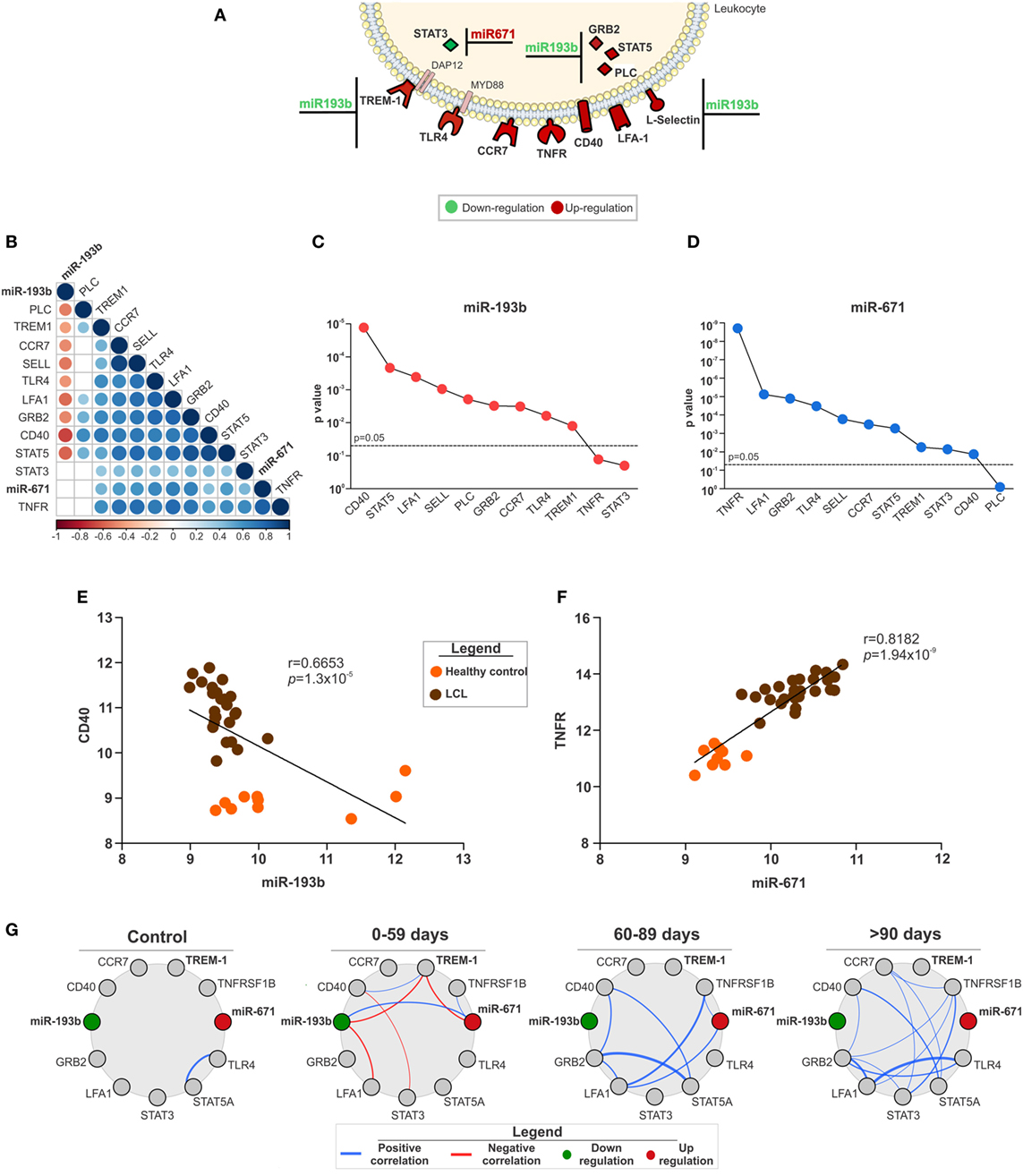
Figure 4. The expression of miR-193b and -671 and their target genes as key in localized cutaneous leishmaniasis (LCL) outcome. (A) Representative image of molecules from innate immune response pathways and their respective microRNAs with opposite expression (↑miR ↓target or ↓miR ↑target). (B) Correlation matrix between the expression of microRNAs and their target genes. Circles and corresponding sizes represent significance (p < 0.05) and r-values for each Spearman correlation, respectively. Colors represent the directionality of the correlation (blue infers positive correlation, whereas red indicates negative correlations). (C) p-Values of negative Spearman correlations between miR-193b and the indicated target genes are shown. (D) p-Values of positive Spearman correlations between miR-671 and the indicated target genes are described. Spearman correlations between (E) miR-193b/CD40 and (F) miR-671/TNFR. (G) Network analysis shows Spearman correlations (−0.6 < r > 0.6) between microRNAs and target genes according to the response to treatment (cured by 59, 60–89 or >90 days). Colors represent the directionality of the correlation (red line, negative correlation and blue line, positive correlation; red circles, upregulated and green circles, downregulated genes).
Considering that these innate immunity pathways could be associated with tissue injury and LCL outcome, we assessed publicly available clinical information for patients in the GSE55664 data set (30). First, we correlated the expression levels of miRNAs and their target genes. The correlation matrix presented in Figure 4B) summarizes these results. Almost every gene evaluated correlated with its respective miRNA. Interestingly, only miR-193b had a negative correlation with its target genes (Figure 4B), suggesting that miR-193b could play a significant role in the amplification of the inflammatory response observed in LCL. The statistical significance (p-value) of the correlations between miR-193b (Figure 4C) and miR-671 (Figure 4D) with their target genes reinforced the relevance of miRNAs in the regulation of inflammation. The most significant genes (CD40 and TNFR) were selected for detailed correlation analysis with miR-193b (Figure 4E) and miR-671 (Figure 4F). As expected, there was a negative correlation between miR-193b and CD40, meaning that increased levels of CD40 are frequently related with reduced miR-193b (Figure 4E), which contributes to the inflammatory response. On the other hand, miR-671 showed a strong positive correlation with TNFR (Figure 4F), suggesting that miR-671 could indirectly enhance the expression of TNFR.
We also examined the correlations between miRNAs and their target genes within groups of patients who had different responses to treatment including patients who were cured in less than 59 days, between 60 and 89 days and more than 90 days. The correlation profile was distinct for each group of patients (Figure 4G), especially for those with reduced cure times (0–59 days). Only patients with a good response to treatment (0–59 days) presented negative correlations (red lines) among miR-193b, TREM-1, miR-671, and LFA-1. On the other hand, patients who needed more than 60 days to be cured presented only positive correlations (blue lines) between different molecules, such as GRB2 and STAT5. Moreover, the total number of correlations was distinct among these groups, increasing in groups of patients with the longest healing times (Figure S5 in Supplementary Material). In addition, there was no correlation among miR-193b, miR-671, and TREM-1 in patients who required more than 90 days of treatment (Figure 4G), suggesting a role for miR-193b, miR-671, and TREM1 in better prognosis for LCL.
Discussion
Integrative analysis of data sets from different sources improves the understanding of complex interactions and networks involved in disease pathogenesis (40–42). This approach allowed us to identify the expression profile of miRNAs and their target genes in lesion samples of patients with LCL caused by L. braziliensis from different endemic areas.
Differential miRNA expression has been investigated in different contexts of human diseases, such as biomarkers of allergic asthma (43), treatment for Hepatitis C Virus infection (18), prognostic markers of different types of cancer (44–46) and many others. However, little is known about miRNA expression profiles in human parasitic diseases. Some studies have identified profiles of circulating miRNAs in murine models of infection by Schistosoma japonicum (47), Schistosoma mansoni (48), and Toxoplasma gondii (49) as potential targets for disease progression or diagnosis. These miRNA expression profiles in the plasma are specific for each disease evaluated, but these findings need to be validated in human samples. On the other hand, miR-451 is significantly enriched in human red blood cells infected by Plasmodium falciparum (50), and, more importantly, miR-451 translocates to the parasite and protects the host, thus reducing parasite survival (51). Those findings provide the first evidence of the protective role from a specific miRNA in human malaria. Recently, it was shown that circulating levels of miR-451 and miR-16 were downregulated in patients infected with Plasmodium vivax at parasitic stages or with multi-organ failure involvement (25, 52). This association of miRNAs profile with clinical severity of malaria infection emphasize that miRNAs are potential tools for diagnosis and therapeutics for parasitic diseases. Moreover, 25 miRNAs released by S. mansoni exosomes are present at high levels in the circulation of infected mice. These results provide evidence of schistosome-derived exosomes could play important roles in host-parasite interactions and could be a useful tool in the development of vaccines and therapeutics (53).
Few studies have evaluated the modulation of miRNA expression during Leishmania infection in cultured cells or experimental models of infection by species causative of Visceral Leishmaniasis (54–58), species from the Old World that causes LCL (59–61) or both (62). Their findings indicate that the expression of miRNAs is specific for each condition. Only a recent study evaluated the expression of 84 miRNAs in mouse macrophages infected by L. amazonensis at different time points after infection (63). These authors showed that miR-294 and miR-721 are induced by L. amazonensis infection and target NOS2 and l-arginine metabolism, thus favoring parasite establishment. However, altered expression of these miRNAs was not observed in any of the other studies of Leishmania infection. Together, these studies underscore the specificity of each model, since the results obtained were not observed in any different condition tested. In fact, this reinforces the importance of evaluating alterations directly from naturally infected patients where the complexity of the human organism is taken into account and potential artifacts from experimental models are avoided.
The approach we applied in this study also allowed the identification of enriched canonical pathways among the genes modulated in each data set. Since miRNAs are important regulators of innate and adaptive immune responses in different pathological conditions (10, 64), such as autoimmune diseases (65), diabetes (66), toxoplasmosis (67), and infections (68, 69), we correlated the molecules within each pathway with the corresponding miRNA by expression pairing.
The analysis of expression pairing results in a more focused set of miRNAs since the prediction of miRNA targets identifies a great number of potential mRNAs. This approach identifies an inverse correlation between a given miRNA and its mRNA targets, which significantly reduces the number of potential mRNA candidates and has been applied in cancer studies (70). From this analysis, only miR-193b and miR-671 had significant mRNA targets that matched these criteria. The involvement of miR-193b in the regulation of relevant target genes has already been described for different types of cancers (71–73) and for hepatitis B infection in humans (74). However, few studies have evaluated the role of miR-671. This analysis implicated this miRNA in neuronal apoptosis (75), in the inflammatory response in colonic epithelial cells (76) and in cancers (77, 78). Furthermore, no reports in the literature have described altered expression of miR-193b and miR-671 in the same context. In this study, we identified both miR-193b and miR-671 as relevant regulators of the inflammatory response observed in human LCL patients.
Our findings revealed six innate immune pathways common to both data sets: communication between innate and adaptive immune cells; granulocyte adhesion and diapedesis; crosstalk between dendritic cells and natural killer cells; dendritic cell maturation; TREM1 signaling; and agranulocyte adhesion and diapedesis. Most of these pathways have been described in the literature as important to the pathogenesis of human LCL (79–81), except for TREM1 signaling (Triggering Receptor Expressed on Myeloid cells). This pathway is a potent amplifier of the pro-inflammatory innate immune response, which should improve pathogen detection and killing. However, the excessive production of inflammatory mediators can also injure host tissues, leading to immunopathology (82). In this context, mice deficient in TREM1 exhibit reduced inflammation and lesion size upon infection by L. major (83), suggesting a pathogenic impact of TREM-1 signaling. Similar results were observed for mice infected with influenza virus or Legionella pneumophila (83). Furthermore, it has been demonstrated that, after in vitro stimulation with L. braziliensis, TREM1 signaling is modulated only in cells from LCL patients who are high producers of IFN-γ (6). Together, these findings reveal the pathogenic potential of TREM-1 signaling but also indicate that its regulation by endogenous miRNAs could be promising in preventing excessive inflammation while preserving pathogen control. The potential of TREM-1 as a therapeutic target was already demonstrated for experimental colitis (84), influenza infection and LCL caused by L. major (83). However, the identity of endogenous agonists/antagonists of TREM-1 has not been completely resolved. The present study identifies miR-193b as a candidate to antagonize TREM-1 expression under inflammatory conditions in an endogenous manner. Ongoing experiments from our group will define the pathogenic role of TREM-1 in patients with LCL caused by L. braziliensis that could ultimately determine disease outcome and prognosis. In addition, manipulation of the miR-193b and TREM-1 axis could also be useful for other inflammatory diseases, since the mechanism by which TREM-1 promotes inflammation seems to be common for distinct models of immune-mediated illness.
Several molecules are involved in inflammatory processes, such as different surface receptors, cytokines, and chemokines, which can also be targeted by miRNAs (85–87). In addition to TREM-1, we observed a strong correlation between miR-193b/CD40 and miR-671/TNFR, and both receptors are considered essential in mediating a variety of immune and inflammatory responses (88–94). Signaling through CD40 and TNFR cooperate to induce anti-Leishmania activity and inflammation (95). However, excessive expression of these molecules can lead to tissue injury and exacerbate immunopathology (96–98). Anti-TNF therapy, which limits TNFR signaling, has been tested in mouse models for different inflammatory diseases, such as rheumatoid arthritis, psoriasis, and bowel diseases (99–101). In this way, the search for new strategies to regulate TNFR expression should include miRNAs. Promising data were published in a mouse model of inflammatory shock where miR-511 was induced to downregulate TNFR protein expression, whereas its target mRNA remained unaltered (98). Silencing the CD40 pathway has also become an attractive target to reduce inflammation in different disease models (97, 102–104). Under inflammatory conditions, endothelial cells downregulate the expression of miR-424 and miR-503, which are both related with the upregulation of CD40. However, treatment of these cells with pioglitazone, a PPARg agonist, activates the transcription of miR-424 and miR-503 and leads to suppression of CD40 expression, suggesting that miRNAs may act as key modulators of the endothelial inflammatory response (97). Together, these findings indicate that a similar approach might be applied in the context of Cutaneous Leishmaniasis, where the altered expression of miR-193b and miR-671 could be reverted in order to downregulate TNFR expression. Further experiments are necessary to clarify if this could help to balance the inflammatory response and minimize tissue injury.
Molecules with significantly altered expression could also be used as markers for diagnosis, disease progression, response to treatment or clinical outcome, contributing to clinical management and improving patient quality of life (105–108). Several studies have shown that the use of biomarkers, including miRNAs, improves diagnostic efficiency (109–112). Since miRNA expression has been shown to be stable and tissue-specific, miRNAs are potential biomarkers in the analysis of tissue and body fluids (113–115). Prediction analysis revealed that the expression profile of miR-200b and miR-200c is able to discriminate patients with prostate cancer from HCs and define the risk of disease in predisposed individuals. These results were validated in serum samples and showed higher specificity and sensibility then PSA (Prostate-specific antigen) testing (112). Similar results were also obtained for different types of cancer, where circulating miRNAs are potential markers of disease progression and response to treatment, as well as a promising screening test for cancer detection (12, 116–118). However, there is a lack of studies suggesting the potential of miRNAs as markers of infectious/parasitic diseases in humans. The majority of these studies have focused on tuberculosis, sepsis, and viral infections (such as HCV, HBV, and HIV) and compared the profiles of circulating miRNAs between patients and HCs (20, 45, 119, 120). Few studies have suggested that S. mansoni-derived miR-277, miR-3479, and bantam (a specific Schistosoma sp. miRNA) are potential markers for parasitic infection, in mouse and human serum, in order to evaluate the response to treatment (48). Furthermore, in the mouse model of T. gondii infection, miR-712, miR-511, and miR-217 have been indicated as a potential miRNA signature that can be useful for early detection of infection (49). In addition, a profile of deregulated miRNAs in malaria patients with multiple organ failure indicated that miRNAs can be used as biomarkers of disease severity (52). Together, these studies reveal that miRNAs can also be useful tools in the management of infectious diseases.
Regarding Leishmania infection in humans, our study provides evidence that the healing time of patients with LCL can be associated with the correlation between miRNAs and theirs target genes. We observed that the axis of miR-193b, miR-671, and TREM-1 could be used as a predictive marker of prognosis for human LCL. Further studies are necessary to define whether this miRNA expression profile can be applied as a biosignature for better prognosis, allowing an early definition of the most effective therapy of human LCL. In summary, we revealed in this study a previously unknown expression profile of miRNAs involved in the regulation of pathways that are important for the immunopathogenesis of human LCL, shedding new light into the mechanisms of disease severity and prognosis. It is tempting to speculate that the axis of miR-193b, miR-671, and TREM-1 is a potential candidate to the development of new tools to manage this disease and other inflammatory skin disorders.
Ethics Statement
This study was carried out in accordance with the recommendations of Institutional Review Board of the Gonçalo Moniz Institute, Bahia-Brazil with written informed consent from all subjects. All subjects gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by the Institutional Review Board of the Gonçalo Moniz Institute, Bahia-Brazil (license number: 47120215.8.0000.0040).
Author Contributions
SN contributed to the design and conception of the experiments, conducted molecular biology experiments, analyzed and interpreted data, and drafted the manuscript. SN, IS, and MA performed RT-qPCR assays and helped with quality control and analysis. SN, PR, CB, and PO normalized the data and performed all statistical analyses. AN, LS, TC, RK, VB, and AB provided biopsies skin samples. SN, PO and NT conceived the study and participated in its design, helped to analyze and interpret the data, and drafted the manuscript.
Conflict of Interest Statement
The authors declare that they have no commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Dr. Bruno Bezerril, Dr. Arthur Queiroz, Dr. Leonardo Farias, Dr. Henrique Serezani, Dr. Johan Van Weyenbergh, Dr. Camila Indiani, Dr. Adriano Alcântara, and Rafael Tibúrcio for their suggestions and comments on this work. The authors thank Andrezza Kariny and Juliana Oliveira for secretarial assistance. The authors thank Zaira Onofre from RT-PCR facility—PDTIS (FIOCRUZ).
Funding
This study was financed by FAPESB (Grant number: 460/2015). AB and CB are senior investigators of the National Council for Research (CNPq). PO, SN, and MA are fellows from CNPq, IS from CAPES.
Supplementary Material
The Supplementary Material for this article can be found online at https://www.frontiersin.org/articles/10.3389/fimmu.2018.00640/full#supplementary-material.
Table S1. Demographic and clinical metadata from Leishmania braziliensis patients.
Table S2. MicroRNA probes identified on the Illumina HT 12 v4 platform (GSE55664 data set).
Table S3. MicroRNA probes identified on the Agilent SurePrint GE Human G3v2 platform (GSE63931 data set).
Table S4. Identification of known microRNAs after data refinement.
Table S5. Raw datasets of identified canonical pathways related to LCL found on Ingenuity Pathways Analysis.
Table S6. Canonical pathways of the innate immune response that are significantly modulated in leishmaniasis as identified by Ingenuity Pathways Analysis.
Figure S1. Pipeline of the search for public transcriptome data sets of human cutaneous leishmaniasis. Microarray data sets for experiments performed on human skin biopsies of cutaneous leishmaniasis caused by Leishmania braziliensis were searched at the Gene Expression Omnibus database (www.ncbi.nlm.nih.gov/geo/). Two data sets were found [GSE55664 (Illumina HT 12 v4 platform) and GSE63931 (Agilent SurePrint GE Human G3v2 platform)]. The Ensembl/BioMart tool was applied for data mining, and the numbers of microRNA probes found were 792 and 141 for GSE55664 and GSE63931, respectively.
Figure S2. Unsupervised clustering of healthy and cutaneous leishmaniasis skin biopsies based on microRNA expressions. Hierarchical clustering was performed using the Euclidean distance method for sets of miRNA probes expressed in healthy control (HC; orange bar) and in localized cutaneous leishmaniasis (LCL; brown bar) skin samples. (A) Heat map showing the expression profiles of 792 microRNA probes from the GSE55664 (Illumina HT12 v4 platform) data set in 10 HC and 25 LCL samples. (B) Heat map showing the expression profiles of 141 microRNA probes from the GSE63931 (Agilent Sure Print GE Human G3v2 platform) data set in eight HC and eight LCL samples. Rows represent microRNA probes and columns represent samples. The scale bar symbolizes the intensity of expression for each microRNA probe (Log2).
Figure S3. Pipeline for the identification of pathways related to immune response in LCL. Both LCL datasets, GSE55664 and GSE63931, were analyzed by Ingenuity Pathways Analysis to identify the canonical pathways related to innate immune response during this disease. Molecules composing these pathways with differential expression (−2< FC >2 and p-value < 0.05) were selected. Using the Targetscan tool, these molecules were searched as targets for the microRNAs found in LCL.
Figure S4. High confidence microRNAs deposited in miRBase 21 database and available in each platform. (A) Total number of Homo sapiens microRNA precursors considered high or low confidence, as described in miRBase 21 (www.mirbase.org/). (B) Absolute number (#) of high (black bars) or low (gray bars) confidence microRNA probes. Some probes covered putative microRNAs not deposited in miRBase 21 (white bars).
Figure S5. Number of correlations between microRNAs and their target genes according to different response to treatment. Total number of correlations between miRNAs and their target genes within groups defined based on the response to treatment. This analysis quantifies the number of correlations and compares among healthy control individuals (CTR), LCL patients cured up to 59 days (0–59), 60–89 days (60–89), or more than 90 days (>90).
Footnotes
References
1. Kaye P, Scott P. Leishmaniasis: complexity at the host-pathogen interface. Nat Rev Microbiol (2011) 9:604–15. doi:10.1038/nrmicro2608
2. Scott P, Novais FO. Cutaneous leishmaniasis: immune responses in protection and pathogenesis. Nat Rev Immunol (2016) 16(9):581–92. doi:10.1038/nri.2016.72
3. Hu SC-S, Yu H-S, Yen F-L, Lin C-L, Chen G-S, Lan C-CE. Neutrophil extracellular trap formation is increased in psoriasis and induces human β-defensin-2 production in epidermal keratinocytes. Sci Rep (2016) 6:31119. doi:10.1038/srep31119
4. Cardoso TM, Machado Á, Costa DL, Carvalho LP, Queiroz A, Machado P, et al. Protective and pathological functions of CD8+ T cells in Leishmania braziliensis infection. Infect Immun (2015) 83:898–906. doi:10.1128/IAI.02404-14
5. Alves-Ferreira EVC, Toledo JS, De Oliveira AHC, Ferreira TR, Ruy PC, Pinzan CF, et al. Differential gene expression and infection profiles of cutaneous and mucosal Leishmania braziliensis isolates from the same patient. PLoS Negl Trop Dis (2015) 9:e0004018. doi:10.1371/journal.pntd.0004018
6. Carneiro MW, Fukutani KF, Andrade BB, Curvelo RP, Cristal JR, Carvalho AM, et al. Gene expression profile of high IFN-γ producers stimulated with Leishmania braziliensis identifies genes associated with cutaneous leishmaniasis. PLoS Negl Trop Dis (2016) 10:e0005116. doi:10.1371/journal.pntd.0005116
7. Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol (2014) 15:509–24. doi:10.1038/nrm3838
8. He L, Hannon GJ, Liu M, Li R, Tian B, Zhang X, et al. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet (2004) 5:522–31. doi:10.1038/nrg1379
9. Mehta A, Baltimore D. MicroRNAs as regulatory elements in immune system logic. Nat Rev Immunol (2016) 16:279–94. doi:10.1038/nri.2016.40
10. O’Connell RM, Rao DS, Baltimore D. MicroRNA regulation of inflammatory responses. Annu Rev Immunol (2012) 30:295–312. doi:10.1146/annurev-immunol-020711-075013
11. Varamo C, Occelli M, Vivenza D, Merlano M, Lo Nigro C. MicroRNAs role as potential biomarkers and key regulators in melanoma. Genes Chromosomes Cancer (2016) 56(1):3–10. doi:10.1002/gcc.22402
12. Cortez MA, Welsh JW, Calin GA. Circulating microRNAs as noninvasive biomarkers in breast cancer. Recent Results Cancer Res (2012) 195:151–61. doi:10.1007/978-3-642-28160-0_13
13. Asangani IA, Rasheed SA, Nikolova DA, Leupold JH, Colburn NH, Post S, et al. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene (2008) 27:2128–36. doi:10.1038/sj.onc.1210856
14. Gabriely G, Wurdinger T, Kesari S, Esau CC, Burchard J, Linsley PS, et al. MicroRNA 21 promotes glioma invasion by targeting matrix metalloproteinase regulators. Mol Cell Biol (2008) 28:5369–80. doi:10.1128/MCB.00479-08
15. Meng F, Henson R, Wehbe-Janek H, Ghoshal K, Jacob ST, Patel T. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology (2007) 133:647–58.
16. Sonkoly E, Wei T, Janson PCJ, Sääf A, Lundeberg L, Tengvall-Linder M, et al. MicroRNAs: novel regulators involved in the pathogenesis of psoriasis? PLoS One (2007) 2:e610. doi:10.1371/journal.pone.0000610
17. Zibert JR, Løvendorf MB, Litman T, Olsen J, Kaczkowski B, Skov L. MicroRNAs and potential target interactions in psoriasis. J Dermatol Sci (2010) 58:177–85. doi:10.1016/j.jdermsci.2010.03.004
18. Janssen HLA, Reesink HW, Lawitz EJ, Zeuzem S, Rodriguez-Torres M, Patel K, et al. Treatment of HCV infection by targeting microRNA. N Engl J Med (2013) 368:1685–94. doi:10.1056/NEJMoa1209026
19. Wang H, Zhang P, Chen W, Feng D, Jia Y, Xie L. Four serum microRNAs identified as diagnostic biomarkers of sepsis. J Trauma Acute Care Surg (2012) 73:850–4. doi:10.1097/TA.0b013e31825a7560
20. Benz F, Roy S, Trautwein C, Roderburg C, Luedde T. Circulating microRNAs as biomarkers for sepsis. Int J Mol Sci (2016) 17:78. doi:10.3390/ijms17010078
21. Ming Z, Zhou R, Chen XM. Regulation of host epithelial responses to Cryptosporidium infection by microRNAs. Parasite Immunol (2017) 39:1–9. doi:10.1111/pim.12408
22. Xie H, Lei N, Gong AY, Chen XM, Hu G. Cryptosporidium parvum induces SIRT1 expression in host epithelial cells through downregulating let-7i. Hum Immunol (2014) 75:760–5. doi:10.1016/j.humimm.2014.05.007
23. Zhou R, Feng Y, Chen X-M. Non-coding RNAs in epithelial immunity to Cryptosporidium infection. Parasitology (2014) 141:1233–43. doi:10.1017/S0031182014000614
24. Barker KR, Lu Z, Kim H, Zheng Y, Chen J, Conroy AL, et al. miR-155 modifies inflammation, endothelial activation and blood-brain barrier dysfunction in cerebral malaria. Mol Med (2017) 23:1. doi:10.2119/molmed.2016.00139
25. Chamnanchanunt S, Kuroki C, Desakorn V, Enomoto M, Thanachartwet V, Sahassananda D, et al. Downregulation of plasma miR-451 and miR-16 in Plasmodium vivax infection. Exp Parasitol (2015) 155:19–25. doi:10.1016/j.exppara.2015.04.013
26. Monteiro CJ, Mota SLA, Diniz LDF, Bahia MT, Moraes KCM. miR-190b negatively contributes to the trypanosoma cruzi-infected cell survival by repressing PTEN protein expression. Mem Inst Oswaldo Cruz (2015) 110:996–1002. doi:10.1590/0074-02760150184
27. Linhares-Lacerda L, Palu CC, Ribeiro-Alves M, Paredes BD, Morrot A, Garcia-Silva MR, et al. Differential expression of microRNAs in thymic epithelial cells from Trypanosoma cruzi acutely infected mice: putative role in thymic atrophy. Front Immunol (2015) 6:1–12. doi:10.3389/fimmu.2015.00428
28. Navarro IC, Ferreira FM, Nakaya HI, Baron MA, Vilar-Pereira G, Pereira IR, et al. MicroRNA transcriptome profiling in heart of Trypanosoma cruzi-infected mice: parasitological and cardiological outcomes. PLoS Negl Trop Dis (2015) 9:e0003828. doi:10.1371/journal.pntd.0003828
29. Ferreira LRP, Frade AF, Santos RHB, Teixeira PC, Baron MA, Navarro IC, et al. MicroRNAs miR-1, miR-133a, miR-133b, miR-208a and miR-208b are dysregulated in chronic chagas disease cardiomyopathy. Int J Cardiol (2014) 175:409–17. doi:10.1016/j.ijcard.2014.05.019
30. Novais FO, Carvalho LP, Passos S, Roos DS, Carvalho EM, Scott P, et al. Genomic profiling of human Leishmania braziliensis lesions identifies transcriptional modules associated with cutaneous immunopathology. J Invest Dermatol (2015) 135:94–101. doi:10.1038/jid.2014.305
31. Oliveira PRS, Dessein H, Romano A, Cabantous S, De Brito MEF, Santoro F, et al. IL2RA genetic variants reduce IL-2-dependent responses and aggravate human cutaneous leishmaniasis. J Immunol (2015) 194(6):2664–72. doi:10.4049/jimmunol.1402047
32. Kozomara A, Griffiths-Jones S. miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res (2014) 42:D68–73. doi:10.1093/nar/gkt1181
33. Kolbert CP, Feddersen RM, Rakhshan F, Grill DE, Simon G, Middha S, et al. Multi-platform analysis of microRNA expression measurements in RNA from fresh frozen and FFPE tissues. PLoS One (2013) 8:e52517. doi:10.1371/journal.pone.0052517
34. Kozomara A, Griffiths-Jones S. miRBase: integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res (2011) 39:152–7. doi:10.1093/nar/gkq1027
35. Griffiths-Jones S, Saini HK, Van Dongen S, Enright AJ. miRBase: tools for microRNA genomics. Nucleic Acids Res (2008) 36:154–8. doi:10.1093/nar/gkm952
36. Ambros V, Bartel B, Bartel DP, Burge CB, Carrington JC, Chen X, et al. A uniform system for microRNA annotation. RNA (2003) 9:277–9. doi:10.1261/rna.2183803
37. Griffiths-Jones S. miRBase: microRNA sequences and annotation. Curr Protoc Bioinformatics (2010) 34:1–12. doi:10.1002/0471250953.bi1209s29
38. Geiger A, Bossard G, Sereno D, Pissarra J, Lemesre JL, Vincendeau P, et al. Escaping deleterious immune response in their hosts: lessons from trypanosomatids. Front Immunol (2016) 7:1–212. doi:10.3389/fimmu.2016.00212
39. Huang JC, Babak T, Corson TW, Chua G, Khan S, Gallie BL, et al. Using expression profiling data to identify human microRNA targets. Nat Methods (2007) 4:1045–9. doi:10.1038/nmeth1130
40. Andres-Terre M, McGuire HM, Pouliot Y, Bongen E, Sweeney TE, Tato CM, et al. Integrated, multi-cohort analysis identifies conserved transcriptional signatures across multiple respiratory viruses. Immunity (2015) 43:1199–211. doi:10.1016/j.immuni.2015.11.003
41. Wang T, Birsoy K, Hughes NW, Krupczak KM, Post Y, Wei JJ, et al. Identification and characterization of essential genes in the human genome. Science (2015) 350:1096–101. doi:10.1126/science.aac7041
42. Zhu M, Yi M, Kim CH, Deng C, Li Y, Medina D, et al. Integrated miRNA and mRNA expression profiling of mouse mammary tumor models identifies miRNA signatures associated with mammary tumor lineage. Genome Biol (2011) 12:R77. doi:10.1186/gb-2011-12-8-r77
43. Milger K, Gotschke J, Krause L, Nathan P, Alessandrini F, Tufman A, et al. Identification of a plasma miRNA biomarker-signature for allergic asthma: a translational approach. Int J Lab Hematol (2016) 38:42–9. doi:10.1111/ijlh.12426
44. Miao Y, Zheng W, Li N, Su Z, Zhao L, Zhou H, et al. MicroRNA-130b targets PTEN to mediate drug resistance and proliferation of breast cancer cells via the PI3K/Akt signaling pathway. Sci Rep (2017) 7:41942. doi:10.1038/srep41942
45. Zhu HT, Liu RB, Liang YY, Hasan AME, Wang HY, Shao Q, et al. Serum microRNA profiles as diagnostic biomarkers for HBV-positive hepatocellular carcinoma. Liver Int (2017) 37(6):888–96. doi:10.1111/liv.13356
46. Liu C, Liu R, Zhang D, Deng Q, Liu B, Chao H-P, et al. MicroRNA-141 suppresses prostate cancer stem cells and metastasis by targeting a cohort of pro-metastasis genes. Nat Commun (2017) 8:14270. doi:10.1038/ncomms14270
47. Cai P, Gobert GN, You H, Duke M, McManus DP. Circulating miRNAs: potential novel biomarkers for hepatopathology progression and diagnosis of Schistosomiasis japonica in two murine models. PLoS Negl Trop Dis (2015) 9:e0003965. doi:10.1371/journal.pntd.0003965
48. Hoy AM, Lundie RJ, Ivens A, Quintana JF, Nausch N, Forster T, et al. Parasite-derived microRNAs in host serum as novel biomarkers of helminth infection. PLoS Negl Trop Dis (2014) 8:e2701. doi:10.1371/journal.pntd.0002701
49. Jia B, Chang Z, Wei X, Lu H, Yin J, Jiang N, et al. Plasma microRNAs are promising novel biomarkers for the early detection of Toxoplasma gondii infection. Parasit Vectors (2014) 7:433. doi:10.1186/1756-3305-7-433
50. Xue X, Zhang Q, Huang Y, Feng L, Pan W. No miRNA were found in Plasmodium and the ones identified in erythrocytes could not be correlated with infection. Malar J (2008) 7:47. doi:10.1186/1475-2875-7-47
51. LaMonte G, Philip N, Reardon J, Lacsina JR, Majoros W, Chapman L, et al. Translocation of sickle cell erythrocyte microRNAs into Plasmodium falciparum inhibits parasite translation and contributes to malaria resistance. Cell Host Microbe (2012) 12:187–99. doi:10.1016/j.chom.2012.06.007
52. Chamnanchanunt S, Fucharoen S, Umemura T. Circulating microRNAs in malaria infection: bench to bedside. Malar J (2017) 16:334. doi:10.1186/s12936-017-1990-x
53. Samoil V, Dagenais M, Ganapathy V, Aldridge J, Glebov A, Jardim A, et al. Vesicle-based secretion in schistosomes: analysis of protein and microRNA (miRNA) content of exosome-like vesicles derived from Schistosoma mansoni. Sci Rep (2018) 8:3286. doi:10.1038/s41598-018-21587-4
54. Ghosh J, Bose M, Roy S, Bhattacharyya SN. Leishmania donovani targets dicer1 to downregulate miR-122, lower serum cholesterol, and facilitate murine liver infection. Cell Host Microbe (2013) 13:277–88. doi:10.1016/j.chom.2013.02.005
55. Mukherjee B, Paul J, Mukherjee S, Mukhopadhyay R, Das S, Naskar K, et al. Antimony-resistant Leishmania donovani exploits miR-466i to deactivate host MyD88 for regulating IL-10/IL-12 levels during early hours of infection. J Immunol (2015) 195:2731–42. doi:10.4049/jimmunol.1402585
56. Pandey RK, Sundar S, Prajapati VK. Differential expression of miRNA regulates T cell differentiation and plasticity during visceral leishmaniasis infection. Front Microbiol (2016) 7:206. doi:10.3389/fmicb.2016.00206
57. Singh AK, Pandey RK, Shaha C, Madhubala R. MicroRNA expression profiling of Leishmania donovani-infected host cells uncovers the regulatory role of MIR30A-3p in host autophagy. Autophagy (2016) 12:1817–31. doi:10.1080/15548627.2016.1203500
58. Chakrabarty Y, Bhattacharyya SN. Leishmania donovani restricts mitochondrial dynamics to enhance miRNP stability and target RNA repression in host macrophages. Mol Biol Cell (2017) 28:2091–105. doi:10.1091/mbc.E16-06-0388
59. Kelada S, Sethupathy P, Okoye IS, Kistasis E, Czieso S, White SD, et al. miR-182 and miR-10a are key regulators of Treg specialisation and stability during schistosome and Leishmania-associated inflammation. PLoS Pathog (2013) 9:e1003451. doi:10.1371/journal.ppat.1003451
60. Lemaire J, Mkannez G, Guerfali FZ, Gustin C, Attia H, Sghaier RM, et al. MicroRNA expression profile in human macrophages in response to Leishmania major infection. PLoS Negl Trop Dis (2013) 7:e2478. doi:10.1371/journal.pntd.0002478
61. Frank B, Marcu A, de Oliveira Almeida Petersen AL, Weber H, Stigloher C, Mottram JC, et al. Autophagic digestion of Leishmania major by host macrophages is associated with differential expression of BNIP3, CTSE, and the miRNAs miR-101c, miR-129, and miR-210. Parasit Vectors (2015) 8:404. doi:10.1186/s13071-015-0974-3
62. Geraci NS, Tan JC, McDowell MA. Characterization of microRNA expression profiles in Leishmania -infected human phagocytes. Parasite Immunol (2015) 37:43–51. doi:10.1111/pim.12156
63. Muxel SM, Silva MFL, Zampieri RA, Floeter-Winter LM. Leishmania (Leishmania) amazonensis induces macrophage miR-294 and miR-721 expression and modulates infection by targeting NOS2 and L-arginine metabolism. Sci Rep (2017) 7:44141. doi:10.1038/srep44141
64. Baltimore D, Boldin MP, Connell RMO, Rao DS, Taganov KD. MicroRNAs: new regulators of immune cell development and function. Nat Immunol (2008) 9:839–45. doi:10.1038/ni.f.209
65. Yan S, Xu Z, Lou F, Zhang L, Ke F, Bai J, et al. NF-κB-induced microRNA-31 promotes epidermal hyperplasia by repressing protein phosphatase 6 in psoriasis. Nat Commun (2015) 6:7652. doi:10.1038/ncomms8652
66. Regazzi R, Guay C. Circulating microRNAs as novel biomarkers for diabetes mellitus. Nat Rev Endocrinol (2013) 9:513–21. doi:10.1038/nrendo.2013.86
67. Zeiner GM, Norman KL, Thomson JM, Hammond SM, Boothroyd JC. Toxoplasma gondii infection specifically increases the levels of key host microRNAs. PLoS One (2010) 5:e8742. doi:10.1371/journal.pone.0008742
68. Kumar M, Sahu SK, Kumar R, Subuddhi A, Maji RK, Jana K, et al. MicroRNA let-7 modulates the immune response to Mycobacterium tuberculosis infection via control of A20, an inhibitor of the NF-kB pathway. Cell Host Microbe (2015) 17(3):345–56. doi:10.1016/j.chom.2015.01.007
69. Jopling CL, Yi M, Lancaster AM, Lemon SM, Sarnow P. Modulation of hepatitis C virus RNA abundance by a. Science (2005) 1:1577–81. doi:10.1126/science.1113329
70. Zhu W, Liu X, He J, Chen D, Hunag Y, Zhang YK. Overexpression of members of the microRNA-183 family is a risk factor for lung cancer: a case control study. BMC Cancer (2011) 11:393. doi:10.1186/1471-2407-11-393
71. Moore C, Parrish JK, Jedlicka P. miR-193b, downregulated in Ewing sarcoma, targets the ErbB4 oncogene to inhibit anchorage-independent growth. PLoS One (2017) 12:e0178028. doi:10.1371/journal.pone.0178028
72. Lin S-R, Yeh H-C, Wang W-J, Ke H-L, Lin H-H, Hsu W-C, et al. miR-193b mediates CEBPD-induced cisplatin sensitization through targeting ETS1 and cyclin D1 in human urothelial carcinoma cells. J Cell Biochem (2017) 118:1563–73. doi:10.1002/jcb.25818
73. Torres-Ferreira J, Ramalho-Carvalho J, Gomez A, Menezes FD, Freitas R, Oliveira J, et al. miR-193b promoter methylation accurately detects prostate cancer in urine sediments and miR-34b/c or miR-129-2 promoter methylation define subsets of clinically aggressive tumors. Mol Cancer (2017) 16:26. doi:10.1186/s12943-017-0604-0
74. van der Ree MH, Jansen L, Kruize Z, van Nuenen AC, van Dort KA, Takkenberg RB, et al. Plasma microRNA levels are associated with HBeAg status and treatment response in chronic hepatitis B patients. J Infect Dis (2017) 215(9):1421–9. doi:10.1093/infdis/jix140
75. Nan A, Chen L, Zhang N, Liu Z, Yang T, Wang Z, et al. A novel regulatory network among LncRpa, CircRar1, miR-671 and apoptotic genes promotes lead-induced neuronal cell apoptosis. Arch Toxicol (2017) 91:1671–84. doi:10.1007/s00204-016-1837-1
76. Chuang AY, Chuang JC, Zhai Z, Wu F, Kwon JH. NOD2 expression is regulated by microRNAs in colonic epithelial HCT116 cells. Inflamm Bowel Dis (2016) 20(1):126–35. doi:10.1097/01.MIB.0000436954.70596.9b
77. Chen Z, Liu H, Jin W, Ding Z, Zheng S, Yu Y. Tissue microRNA-21 expression predicted recurrence and poor survival in patients with colorectal cancer – a meta-analysis. Onco Targets Ther (2016) 9:2615–24. doi:10.2147/OTT.S103893
78. Rutnam ZJ, Yang BB. The involvement of microRNAs in malignant transformation. Histol Histopathol (2012) 27:1263–70. doi:10.14670/HH-27.1263
79. Bogdan C. Natural killer cells in experimental and human leishmaniasis. Front Cell Infect Microbiol (2012) 2:69. doi:10.3389/fcimb.2012.00069
80. Feijó D, Tibúrcio R, Ampuero M, Brodskyn C, Tavares N. Dendritic cells and Leishmania infection: adding layers of complexity to a complex disease. J Immunol Res (2016) 2016:9. doi:10.1155/2016/3967436
81. Falcão Sde A, Jaramillo TM, Ferreira LG, Bernardes DM, Santana JM, Favali CB. Leishmania infantum and Leishmania braziliensis: differences and similarities to evade the innate immune system. Front Immunol (2016) 7:287. doi:10.3389/fimmu.2016.00287
82. Yasuda T, Takeyama Y, Ueda T, Shinzeki M, Sawa H, Takahiro N, et al. Increased levels of soluble triggering receptor expressed on myeloid cells-1 in patients with acute pancreatitis*. Crit Care Med (2008) 36:2048–53. doi:10.1097/CCM.0b013e31817b8824
83. Weber B, Schuster S, Zysset D, Rihs S, Dickgreber N, Schürch C, et al. TREM-1 deficiency can attenuate disease severity without affecting pathogen clearance. PLoS Pathog (2014) 10:e1003900. doi:10.1371/journal.ppat.1003900
84. Schenk M, Bouchon A, Seibold F, Mueller C. TREM-1–expressing intestinal macrophages crucially amplify chronic inflammation in experimental colitis and inflammatory bowel diseases. J Clin Invest (2007) 117:3097–106. doi:10.1172/JCI30602
85. Schulte LN, Westermann AJ, Vogel J. Differential activation and functional specialization of miR-146 and miR-155 in innate immune sensing. Nucleic Acids Res (2013) 41:542–53. doi:10.1093/nar/gks1030
86. Sonkoly E, Pivarcsi A. microRNAs in inflammation. Int Rev Immunol (2009) 28:535–61. doi:10.3109/08830180903208303
87. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell (2009) 136:215–33. doi:10.1016/j.cell.2009.01.002
88. Sedger LM, McDermott MF. TNF and TNF-receptors: from mediators of cell death and inflammation to therapeutic giants – past, present and future. Cytokine Growth Factor Rev (2014) 25:453–72. doi:10.1016/j.cytogfr.2014.07.016
89. Senhaji N, Kojok K, Darif Y, Fadainia C, Zaid Y. The contribution of CD40/CD40L axis in inflammatory bowel disease: an update. Front Immunol (2015) 6:1–6. doi:10.3389/fimmu.2015.00529
90. Croft M, Siegel RM. Beyond TNF: TNF superfamily cytokines as targets for the treatment of rheumatic diseases. Nat Rev Rheumatol (2017) 13:217–33. doi:10.1038/nrrheum.2017.22
91. Yan S, Yim LY, Tam RCY, Chan A, Lu L, Lau CS, et al. MicroRNA-155 mediates augmented CD40 expression in bone marrow derived plasmacytoid dendritic cells in symptomatic lupus-prone NZB/W F1 mice. Int J Mol Sci (2016) 17:1282. doi:10.3390/ijms17081282
92. Terrazzano G, Pisanti S, Grimaldi S, Sica M, Fontana S, Carbone E, et al. Interaction between natural killer and dendritic cells: the role of CD40, CD80 and major histocompatibility complex class i molecules in cytotoxicity induction and interferon-gamma production. Scand J Immunol (2004) 59:356–62. doi:10.1111/j.0300-9475.2003.01387.x
93. Srivastava S, Tsongalis GJ, Kaur P. Role of microRNAs in regulation of the TNF/TNFR gene superfamily in chronic lymphocytic leukemia. Clin Biochem (2016) 49:1307–10. doi:10.1016/j.clinbiochem.2016.08.010
94. Yuan M, Zhang L, You F, Zhou J, Ma Y, Yang F, et al. miR-145-5p regulates hypoxia-induced inflammatory response and apoptosis in cardiomyocytes by targeting CD40. Mol Cell Biochem (2017) 431:123–31. doi:10.1007/s11010-017-2982-4
95. Portillo J-AC, Feliciano LM, Okenka G, Heinzel F, Subauste MC, Subauste CS. CD40 and tumour necrosis factor-α co-operate to up-regulate inducuble nitric oxide synthase expression in macrophages. Immunology (2012) 135:140–50. doi:10.1111/j.1365-2567.2011.03519.x
96. Mach F, Schönbeck U, Sukhova GK, Bourcier T, Bonnefoy JY, Pober JS, et al. Functional CD40 ligand is expressed on human vascular endothelial cells, smooth muscle cells, and macrophages: implications for CD40-CD40 ligand signaling in atherosclerosis. Proc Natl Acad Sci U S A (1997) 94:1931–6. doi:10.1073/pnas.94.5.1931
97. Lee A, Papangeli I, Park Y, Jeong H, Choi J, Kang H, et al. A PPARγ-dependent miR-424/503-CD40 axis regulates inflammation mediated angiogenesis. Sci Rep (2017) 7:2528. doi:10.1038/s41598-017-02852-4
98. Puimege L, Van Hauwermeiren F, Steeland S, Van Ryckeghem S, Vandewalle J, Lodens S, et al. Glucocorticoid-induced microRNA-511 protects against TNF by down-regulating TNFR1. EMBO Mol Med (2015) 7:1004–17. doi:10.15252/emmm.201405010
99. Chadwick W, Magnus T, Martin B, Keselman A, Mattson MP, Maudsley S. Targeting TNF-alpha receptors for neurotherapeutics. Trends Neurosci (2008) 31:504–11. doi:10.1016/j.tins.2008.07.005
100. Van Hauwermeiren F, Vandenbroucke RE, Libert C. Treatment of TNF mediated diseases by selective inhibition of soluble TNF or TNFR1. Cytokine Growth Factor Rev (2011) 22:311–9. doi:10.1016/J.CYTOGFR.2011.09.004
101. Vandenbroucke RE, Vanlaere I, Van Hauwermeiren F, Van Wonterghem E, Wilson C, Libert C. Pro-inflammatory effects of matrix metalloproteinase 7 in acute inflammation. Mucosal Immunol (2014) 7:579–88. doi:10.1038/mi.2013.76
102. Hueso M, De Ramon L, Navarro E, Ripoll E, Cruzado JM, Grinyo JM, et al. Silencing of CD40 in vivo reduces progression of experimental atherogenesis through an NF-κB/miR-125b axis and reveals new potential mediators in the pathogenesis of atherosclerosis. Atherosclerosis (2016) 255:80–9. doi:10.1016/j.atherosclerosis.2016.11.002
103. Lin Q, Geng Y, Zhao M, Lin S, Zhu Q, Tian Z. miR-21 regulates TNF-α-induced CD40 expression via the SIRT1-NF-κB pathway in renal inner medullary collecting duct cells. Cell Physiol Biochem (2017) 41:124–36. doi:10.1159/000455981
104. Lutgens E, Lievens D, Beckers L, Wijnands E, Soehnlein O, Zernecke A, et al. Deficient CD40-TRAF6 signaling in leukocytes prevents atherosclerosis by skewing the immune response toward an antiinflammatory profile. J Exp Med (2010) 207:391–404. doi:10.1084/jem.20091293
105. Gurvits N, Löyttyniemi E, Nykänen M, Kuopio T, Kronqvist P, Talvinen K. Separase is a marker for prognosis and mitotic activity in breast cancer. Br J Cancer (2017) 1:1383–91. doi:10.1038/bjc.2017.301
106. Letellier E, Schmitz M, Ginolhac A, Rodriguez F, Ullmann P, Qureshi-Baig K, et al. Loss of myosin Vb in colorectal cancer is a strong prognostic factor for disease recurrence. Br J Cancer (2017) 117(11):1689–701. doi:10.1038/bjc.2017.352
107. Costa AFO, Menezes DL, Pinheiro LHS, Sandes AF, Nunes MAP, Lyra Junior DP, et al. Role of new immunophenotypic markers on prognostic and overall survival of acute myeloid leukemia: a systematic review and meta-analysis. Sci Rep (2017) 7:4138. doi:10.1038/s41598-017-00816-2
108. Romanelli RM, Magny JF, Jacquemard F. Prognostic markers of symptomatic congenital cytomegalovirus infection. Braz J Infect Dis (2008) 12:38–43. doi:10.1590/S1413-86702008000100009
109. Ortiz-Quintero B. Cell-free microRNAs in blood and other body fluids, as cancer biomarkers. Cell Prolif (2016) 49:281–303. doi:10.1111/cpr.12262
110. Lan H, Lu H, Wang X, Jin H. MicroRNAs as potential biomarkers in cancer: opportunities and challenges. Biomed Res Int (2015) 2015:125094. doi:10.1155/2015/125094
111. Macha M, Seshacharyulu P, Krishn S, Pai P, Rachagani S, Jain M, et al. MicroRNAs (miRNAs) as biomarker(s) for prognosis and diagnosis of gastrointestinal (GI) cancers. Curr Pharm Des (2014) 20:5287–97. doi:10.1016/j.biotechadv.2011.08.021
112. De Souza MF, Kuasne H, Barros-Filho MDC, Cilião HL, Marchi FA, Fuganti PE, et al. Circulating mRNAs and miRNAs as candidate markers for the diagnosis and prognosis of prostate cancer. PLoS One (2017) 12:e0184094. doi:10.1371/journal.pone.0184094
113. Takamizawa J, Konishi H, Yanagisawa K, Tomida S, Osada H, Endoh H, et al. Advances in brief reduced expression of the let-7 microRNAs in human lung cancers in association with shortened postoperative survival. Cancer Res (2004) 64:3753–6. doi:10.1158/0008-5472.CAN-04-0637
114. Almeida MI, Reis RM, Calin GA. MicroRNA history: discovery, recent applications, and next frontiers. Mutat Res (2011) 717:1–8. doi:10.1016/j.mrfmmm.2011.03.009
115. Bertoli G, Cava C, Castiglioni I. MicroRNAs: new biomarkers for diagnosis, prognosis, therapy prediction and therapeutic tools for breast cancer. Theranostics (2015) 5:1122–43. doi:10.7150/thno.11543
116. Hamam R, Ali AM, Alsaleh KA, Kassem M, Alfayez M, Aldahmash A, et al. MicroRNA expression profiling on individual breast cancer patients identifies novel panel of circulating microRNA for early detection. Sci Rep (2016) 6:25997. doi:10.1038/srep25997
117. Fayyad-Kazan H, Bitar N, Najar M, Lewalle P, Fayyad-Kazan M, Badran R, et al. Circulating miR-150 and miR-342 in plasma are novel potential biomarkers for acute myeloid leukemia. J Transl Med (2013) 11:31. doi:10.1186/1479-5876-11-31
118. Jayaraman M, Radhakrishnan R, Mathews CA, Yan M, Husain S, Moxley KM, et al. Identification of novel diagnostic and prognostic miRNA signatures in endometrial cancer. Genes Cancer (2017) 8:566–76. doi:10.18632/genesandcancer.144
119. Murray DD, Suzuki K, Law M, Trebicka J, Ne J, Joh M, et al. Circulating miR-122 and miR-200a as biomarkers for fatal liver disease individuals. Sci Rep (2017) 7(1):1–11. doi:10.1038/s41598-017-11405-8
Keywords: Leishmania braziliensis, microRNA, skin, transcriptome, TREM-1, human leishmaniasis
Citation: Nunes S, Silva IB, Ampuero MR, Noronha ALLd, Souza LCLd, Correia TC, Khouri R, Boaventura VS, Barral A, Ramos PIP, Brodskyn C, Oliveira PRS and Tavares NM (2018) Integrated Analysis Reveals That miR-193b, miR-671, and TREM-1 Correlate With a Good Response to Treatment of Human Localized Cutaneous Leishmaniasis Caused by Leishmania braziliensis. Front. Immunol. 9:640. doi: 10.3389/fimmu.2018.00640
Received: 11 December 2017; Accepted: 14 March 2018;
Published: 04 April 2018
Edited by:
José Roberto Mineo, Federal University of Uberlandia, BrazilReviewed by:
Hira Nakhasi, Center for Biologics Evaluation and Research (FDA), United StatesMaryam Dadar, Razi Vaccine and Serum Research Institute, Iran
Copyright: © 2018 Nunes, Silva, Ampuero, Noronha, Souza, Correia, Khouri, Boaventura, Barral, Ramos, Brodskyn, Oliveira and Tavares. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Natalia Machado Tavares, bmF0YWxpYS50YXZhcmVzQGJhaGlhLmZpb2NydXouYnI=