 Laura Fernández1
Laura Fernández1 Eugenia Carrillo1*
Eugenia Carrillo1* Lucas Sánchez-Sampedro2
Lucas Sánchez-Sampedro2 Carmen Sánchez1
Carmen Sánchez1 Ana Victoria Ibarra-Meneses1
Ana Victoria Ibarra-Meneses1 Ma Angeles Jimenez3
Ma Angeles Jimenez3 Valter dos Anjos Almeida4
Valter dos Anjos Almeida4 Mariano Esteban2
Mariano Esteban2 Javier Moreno1
Javier Moreno1
- 1WHO Collaborating Center for Leishmaniasis, National Center of Microbiology, Instituto de Salud Carlos III, Madrid, Spain
- 2Department of Molecular and Cellular Biology, Centro Nacional de Biotecnología, Consejo Superior de Investigaciones Científicas (CNB-CSIC), Madrid, Spain
- 3Departamento Medicina y Cirugia Animal, Facultad de Veterinaria, Universidad Complutense de Madrid, Madrid, Spain
- 4Instituto Gonçalo Moniz, Fundação Oswaldo Cruz, Salvador, Bahia, Brazil
Leishmania-activated C-kinase antigen (LACK) is a highly conserved protein among Leishmania species and is considered a viable vaccine candidate for human leishmaniasis. In animal models, prime-boost vaccination with LACK-expressing plasmids plus attenuated vaccinia viruses (modified vaccinia Ankara [MVA] and mutant M65) expressing LACK, has been shown to protect against cutaneous leishmaniasis (CL). Further, LACK demonstrated to induce the production of protective cytokines in patients with active CL or cured visceral leishmaniasis, as well as in asymptomatic individuals from endemic areas. However, whether LACK is capable to trigger cytokine release by peripheral blood mononuclear cells from patients cured of CL due to Leishmania infantum (L. infantum) or induce protection in L. infantum-infected hamsters [visceral leishmaniasis (VL) model], has not yet been analyzed. The present work examines the ex vivo immunogenicity of LACK in cured VL and CL patients, and asymptomatic subjects from an L. infantum area. It also evaluates the vaccine potential of LACK against L. infantum infection in hamsters, in a protocol of priming with plasmid pCI-neo-LACK (DNA-LACK) followed by a booster with the poxvirus vectors MVA-LACK or M65-LACK. LACK-stimulated PBMC from both asymptomatic and cured subjects responded by producing IFN-γ, TNF-α, and granzyme B (Th1-type response). Further, 78% of PBMC samples that responded to soluble Leishmania antigen showed IFN-γ secretion following stimulation with LACK. In hamsters, the protocol of DNA-LACK prime/MVA-LACK or M65-LACK virus boost vaccination significantly reduced the amount of Leishmania DNA in the liver and bone marrow, with no differences recorded between the use of MVA or M65 virus vector options. In summary, the Th1-type and cytotoxic responses elicited by LACK in PBMC from human subjects infected with L. infantum, and the parasite protective effect of prime/boost vaccination in hamsters with DNA-LACK/MVA-LACK and DNA-LACK/M65-LACK, revealed the significance of LACK in activating human and hamster immune responses and support LACK to be a valuable candidate for inclusion in a vaccine against human VL.
Introduction
Leishmaniasis is one of the most neglected tropical diseases and has strong links with poverty (1). Visceral leishmaniasis (VL) is the most severe form; its annual incidence is 200,000–400,000 cases worldwide, and without treatment mortality is high (2). In South Asia and East Africa, VL is caused by Leishmania donovani, while in the Mediterranean, the Middle East and Latin America, the causal agent is Leishmania infantum (L. infantum).
Visceral leishmaniasis treatment is based on pentavalent antimonials, oral miltefosine, liposomal amphotericin B, and paramomycin, but these have been associated with severe toxic side effects and increasing parasite resistance (3). The treatment of leishmaniasis is expensive ranging from US$ 30 to 1,500 for medication alone, which may further increase the poverty of affected individuals (1). While vaccination is the most cost-effective way of controlling infectious diseases (4), no vaccine against human VL exists. However, there are reasons for optimism that one or more safe and effective vaccines against leishmaniasis might be developed (3, 5, 6). A number of parasite antigens have been identified as candidates for vaccine development (5), including Leishmania analog receptor for activated C kinase (LACK) (7). Leishmania-activated C-kinase antigen (LACK) is a 36 kDa protein highly conserved across Leishmania species, and is expressed by both promastigotes and amastigotes (8). LACK antigen from Leishmania braziliensis, Leishmania guyanensis, and Leishmania amazonensis induces the production of IFN-γ and IL-10 in peripheral blood mononuclear cells (PBMC) from patients with cutaneous leishmaniasis (CL), and IL-10 in those of naïve individuals (7, 9–12). LACK drives the expansion of IL-4 secreting T cells (13), and for that reason LACK vaccination trials used approaches, like cytokines or DNA vectors, to redirect early IL-4 responses to a protective Th1 response (14, 15). In this sense, vaccinia virus is a strong adjuvant and delivery vector altogether, modifying IL-4 secretion of Vβ4 Vα8 CD4+ T cells (16).
Immunization with LACK-expressing plasmid DNA, with altered LACK peptides, or with the purified protein in the presence of IL-12, results in protection against CL in a mouse model, advancing the candidacy of LACK in the production of a vaccine against leishmaniasis (9). Heterologous immunization, i.e., priming with a DNA vector expressing LACK followed by a boost with non-replicating modified vaccinia virus Ankara (MVA) expressing LACK has proven to be an effective protocol in affording protection against Leishmania major infection in a murine model of CL, and against L. infantum infection in a canine model of VL (17, 18). Further, boosting with LACK-expressing replicative M65 vaccinia virus has been reported to protect against CL in a murine model (17). The protection conferred by heterologous prime/boost immunization involving a DNA vector expressing LACK, plus LACK-expressing M65 virus, however, has not been tested in any experimental model of VL. LACK-expressing M65 and MVA have both been reported to activate polyfunctional CD4+ and CD8+ T cells with effector memory phenotypes. However, prime/boost immunization involving a LACK-expressing pCI-neo plasmid plus LACK-expressing M65 preferentially induced CD4+ T cell responses, while the same plasmid plus LACK-expressing MVA preferentially induced a CD8+ T cell response (17). The replication competence of these viruses has also been described to play an important role in disease prevention (17). The preferential CD4+ T cell response of LACK-expressing M65 plus its replication competence suggest this virus, in combination with a priming LACK-expressing plasmid, might induce potent protection against VL.
With the eventual goal of producing an effective vaccine against human VL caused by L. infantum, the aim of this work was to examine the cellular immune response to the LACK protein induced in humans who have been in contact with L. infantum, and to assess the parasite efficacy of prime/boost immunization involving a LACK-expressing plasmid plus LACK-expressing MVA or M65 viruses in a hamster model of VL.
Materials and Methods
Ethics Statement
Work involving human subjects was approved by the Hospital de Fuenlabrada Ethics and Research Committee (APR 12-65 and APR 14-64). All participants gave their written, informed consent to be included. Work involving animals was approved by the Research Ethics and Animal Welfare Committee of the Instituto de Salud Carlos III, and performed adhering to Spanish legislation on the protection of animals used for experimentation and other scientific purposes (Royal Decree 1201/2005 and Law 32/2007; this law is a transposition of Directive 86/609/EEC). One efficacy trial was performed in this study, and duplication of the animal experiments was not approved for the Research Ethics and Animal Welfare Committee.
Study Subjects
Peripheral blood samples were collected between 2013 and 2015 from 13 patients cured of VL (CVL) and 10 cured of CL (CCL) (with cure confirmed at 6 months after the end of treatment). The patients with VL had been treated with liposomal amphotericin B, and those with CL with meglumine antimoniate. All were attended to at the Hospital de Fuenlabrada, Madrid. Blood samples were also collected from 90 healthy blood donors at the hospital blood bank. Among these, 18 showed a Leishmania-specific cell proliferative response when PBMC were stimulated in vitro with soluble Leishmania antigen (SLA) (SI > 2.1); these were defined as asymptomatic (ASYMP) (19) and selected for inclusion. Another 14 donors who showed no Leishmania-specific cell response were randomly selected for inclusion as endemic-area healthy controls (EC). Other classical assays, such as the leishmanin skin test (LST; no GMP commercial production approved in EU) and the SLA-based ELISA (very low sensitivity for L. infantum asymptomatic subjects), were not considered in this study for identification of asymptomatic infections. All patients plus the ASYMP and EC subjects lived in Fuenlabrada, a L. infantum post-outbreak area in Madrid (Spain).
Animals and Parasite Strain
Thirty two, 12-week-old, male golden hamsters (Mesocricetus auratus) were purchased from Janvier (France). All were housed in the animal facilities of the National Centre for Microbiology and randomly assigned to one of four experimental groups (see below).
Leishmania infantum promastigotes (MCAN/ES/98/LLM-724, JPC strain) were grown for 2 weeks in Novy-MacNeal-Nicolle (NNN) medium and RPMI medium (Gibco, UK) supplemented with 100 UI/ml of penicillin, 100 mg/ml of streptomycin, 2 mM l-glutamine, 2-mercaptoethanol, and 10% heat inactivated fetal calf serum (FCS) (Lonza, Spain).
Cells, Plasmids, and Viruses
The mammalian expression plasmid vector pCI-neo-LACK has been previously described (8). The empty pCI-neo plasmid (Promega, USA) was used as a control (plasmid-φ).
The viruses used in this study included the MVA strain used as a control (MVA-φ) and recombinant MVA and M65 vaccinia virus strains expressing the L. infantum LACK antigen at the viral HA locus (MVA-LACK and M65-LACK, respectively). MVA-LACK and M65-LACK were obtained by transfection with the pHLZ-LACK plasmid, produced by cloning the L. infantum LACK gene into the SmaI site of the pHLZ VACV insertion plasmid under the control of the synthetic early/late pE/L viral promoter and the hemagglutinin (HA) flanking sites. Transfection was achieved by infecting pHLZ-LACK plasmid-containing BSC-40 cells (for M65-LACK) or chick cells (for MVA-LACK) with MVA or M65 viruses and harvesting 48–72 h post-infection. Since pHLZ-LACK contains the Escherichia coli (E. coli) β-glucuronidase gene under the control of the p7.5 early/late viral promoter, β-glucuronidase-producing plaques were identified after the addition of X-Gluc to the agar. Recombinant viruses were plaque purified several times as previously described (20) (checked by PCR), grown in primary chicken embryo fibroblasts (CEF) (from pathogen-free 11-day-old eggs [Intervet, Spain]) in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% FCS in a humidified 5% CO2 atmosphere at 37°C and purified by sedimentation after two sucrose-cushions. Viruses were titrated by immunostaining in CEF cells.
For inoculation in animals, pCI-neo-LACK plasmids and LACK-expressing viruses were diluted in endotoxin-free PBS.
Soluble Leishmania and LACK Antigens
Soluble Leishmania antigen was prepared from L. infantum promastigotes in the stationary phase of growth (MCAN/ES/98/LLM-724, JPC strain). Parasite cultures were centrifuged at 1,000 g for 20 min at 4°C. The pellet was then resuspended in lysis buffer (50 mM Tris/5 mM EDTA/HCl, pH 7), subjected to three cycles of freezing–thawing, sonicated, and further centrifuged at 27,000 g for 4 h at 4°C. The resulting supernatant was divided into aliquots and stored at −20°C. The protein concentration was determined using the Pierce BCA Protein Assay Kit (Thermo Scientific, USA) following the manufacturer’s recommendations.
Recombinant L. infantum LACK was expressed in E. coli BL21 pLysS (transformed with plasmid pRSET-B-LACK) as a fusion protein with an N-terminal histidine tag, and purified by affinity chromatography in a Ni2+ column. All bacteria were grown in the presence of ampicillin and LACK expression induced with isopropyl thio-β-D-galactoside (final concentration 0.5 mM). Cultures were centrifuged at 3,600 g for 15 min, resuspended in lysis buffer (8 M urea, 50 mM Tris-HCL pH 8, 500 mM NaCl), and incubated for 1 h with end-over-end mixing in a tube rotator at 4°C. After a further 30 min centrifugation at 14,000 g, the supernatant was incubated with 2 ml of equilibrated ProBond Nickel-Chelating Resin (Invitrogen, USA). The beads were then washed three times with wash buffer (8 M urea, 50 mM Tris-HCL pH 8, 500 mM NaCl, 10 mM imidazol) and elution performed using an increasing imidazol series (final concentration 200 mM). LACK protein for in vitro studies was kept in saline in PD10 Desalting Columns (GE Healthcare, USA). LPS levels were determined by the Limulus Amebocyte Lysate kit (Sigma-Aldrich, Heidelberg, Germany), and levels were equal to 0.2 endotoxin units per mg protein.
Quantification of Cytokines and Granzyme B in Supernatants From PBMC Stimulated With SLA and LACK
Peripheral blood mononuclear cells from study subjects were isolated from heparinized blood in a Ficoll–Hypaque gradient (Rafer, UK) and resuspended in RPMI 1640 supplemented with 10% FCS and 100 U/ml penicillin/streptomycin (Lonza, Sweden) as previously described (21). 2 × 105 cells/well were then distributed into 96-well plates and cultured for 5 days with supplemented RPMI 1640 medium either alone or with 10 µg/ml SLA or 25 µg/ml LACK at 37°C in a 5% CO2 atmosphere. The supernatants were then collected and stored at −20°C until analyzed. IFN-γ, TNF-α, granzyme B, and IL-10 were quantified by flow cytometry using the BD Cytometric Bead Array Human Flex Kit (Beckton & Dickinson Bioscience, USA) as previously described (21). Data were analyzed using FCAP Array software v.3.0 (Beckton & Dickinson Bioscience, USA).
Immunization and Experimental Infection of Hamsters
Hamsters were randomly distributed into four experimental groups of eight animals each: (1) C-PBS—non-vaccinated controls; (2) C-DNA—prime/boost vaccinated with plasmid-φ and MVA-φ; (3) MVA-LACK—prime/boost vaccinated with pCI-neo-LACK and MVA-LACK; (4) M65-LACK—prime/boost vaccinated with pCI-neo-LACK and M65-LACK.
The above priming injections were performed intramuscularly using 100 µg of plasmid-φ or pCI-neo-LACK as required in 100 µl final volumes of PBS. Four weeks later, the animals were boosted intramuscularly (2 × 107 PFU/hamster) with MVA-φ, MVA-LACK, or M65-LACK as required. All immunization protocols were repeated 4 weeks after the initial boost. The C-PBS control group was injected with 100 µl of PBS at each immunization point. Four weeks after the last reinforcement, all hamsters were inoculated with 2 × 107 promastigotes via the intracardiac route (22), and 4 months later anesthetized with isoflurane and sacrificed by cardiac puncture.
PBMC Isolation and Proliferation Assay in Hamsters
Blood was drawn from the hamsters at the time of sacrifice, and PBMC isolated using a Ficoll–Hypaque density gradient. 1 × 105 cells/well were plated on RPMI medium either alone or with 10 µg/ml SLA or 25 µg/ml LACK for 5 days. 5-bromo-2 deoxyuridine (BrdU) (25 µl, 10 µM) was added to each well for the last 18 h to examine lymphocyte proliferation using the BrdU Cell Proliferation Assay Kit (GE Healthcare Life Sciences, UK) according to the manufacturer’s instructions. Results were expressed as stimulation indices, which represent the ratio between the mean absorbance of stimulated cells and that of unstimulated cells (22).
Immunoenzyme Assay
An aliquot of blood (with heparin) was also taken at the time of sacrifice to provide plasma samples via centrifugation at 2,000 g for 10 min. The plasma collected was then stored at −20°C until use. Maxisorp microtiter plates (Nunc, Denmark) were antigen-coated overnight with SLA (10 µg/ml) or LACK (25 µg/ml) in carbonate buffer (1 mM Na2CO3, 28 mM NaHCO3, pH9.6) and blocked for 1 h at 37°C with 200 mL of 1% BSA and 0.1% Tween 20 in PBS. PBS containing 0.01% Tween 20 was used to wash the plates three times, which were then incubated for 30 min with 100 µl of plasma diluted 1:100 in buffer (0.1% BSA and 0.1% Tween 20 in PBS). Plates were washed and incubated for 30 min with 1:2,000 horseradish peroxidase-conjugated goat anti-hamster IgG (Abd Serotec, UK) for LACK ELISA, and 1:5,000 for SLA ELISA. o-Phenylenediamine dihydrochloride tablets (Sigma, Spain) were used as the enzyme’s substrate; the reaction was stopped with 50 µl of 1 M H2SO4. The absorbance was measured at 492 nm in a Multiskan FC microplate photometer (Thermo Scientific, USA).
Histopathology
All 32 hamsters underwent a complete necropsy and samples of liver, spleen, kidney, and bone marrow were collected from each. Tissues were fixed in 10% buffered formalin, trimmed, processed, and embedded in paraffin wax following routine laboratory procedures, sectioned at 4 µm, and stained with hematoxylin-eosin for histopathological examination under the light microscope. Two samples of liver from the middle and lateral were processed. Two step sections 20 µm apart were obtained for examination from each liver sample. Similar step sections 20 µm apart from spleen, bone marrow, and kidneys were examined for each animal by a trained pathologist blinded for the experiment. Inflammatory and degenerative lesions in each tissue were qualitatively described and scored semi-quantitatively according to their severity as either non-existent (0), mild (+), moderate (++), or severe (+++). Mild was established when inflammatory infiltrates occupied less than 20% of the sections, moderate when inflammatory infiltrates occupied between 20 and 40% of the sections, and severe when inflammatory infiltrates occupied over 40% of the examined parenchyma. Additional specific features such as giant multinucleated cells and Schauman bodies were descriptively noted and recorded for each case.
DNA Isolation and Quantitative Real Time PCR
During necropsy, liver and spleen samples were homogenized in RPMI medium using a 40 µm stainless steel tissue grinder, and 1 × 106 cells used for total DNA isolation via traditional phenol/chloroform extraction and ethanol precipitation. Total DNA was resuspended in 100 µl of distilled water and quantified using a ND-1000 UV-V spectrophotometer (NanoDrop Technology, USA). Leishmania DNA was quantified by quantitative real-time PCR (qPCR) using a LightCycler high speed thermocycler and the LightCycler FastStart DNA Master SYBR Green I kit (Roche Diagnostics, Spain), as previously described (22).
Statistical Analysis
Data were analyzed using the Mann–Whitney U-test. Significance was set at p ≤ 0.05. The cut-off values for IFN-γ production after incubation with SLA or LACK were determined by calculating the area under the receiver operating characteristic (ROC) curve (AUC) along with the 95% confidence interval (CI). In the figures of the efficacy trial, each dot represents a single hamster, and the black line shows the group geometric mean. All calculations were performed using GraphPad Prism 7.0 software (GraphPad Software, USA).
Results
LACK Induced a Significant Increase in TNF-α, IFN-γ, and Granzyme B Production in PBMC From L. infantum Cured and Asymptomatic Human Subjects
Leishmania-activated C-kinase antigen induced a significant increase in the secretion of IFN-γ and granzyme B by the PBMC from the CVL (p ≤ 0.001), CCL (p ≤ 0.01), and ASYMP subjects (p ≤ 0.0001) over that produced by the EC subjects (Figures 1A,C). TNF-α production induced by LACK was significantly increased in the CCL (p ≤ 0.001) and ASYMP subjects (p < 0.01) (Figure 1B). SLA stimulation led to significant increases in the production of IFN-γ, granzyme B, and TNF-α in the CVL, CCL, and ASYMP subjects (p ≤ 0.0001 in all cases) (Figures 1E–G). IL-10 production was detected in all groups, including the controls, after LACK stimulation (Figure 1D). After SLA stimulation, PBMC from the CVL and asymptomatic subjects showed significantly greater IL-10 production compared to the EC subjects’ cells (although levels were low) (Figure 1H).
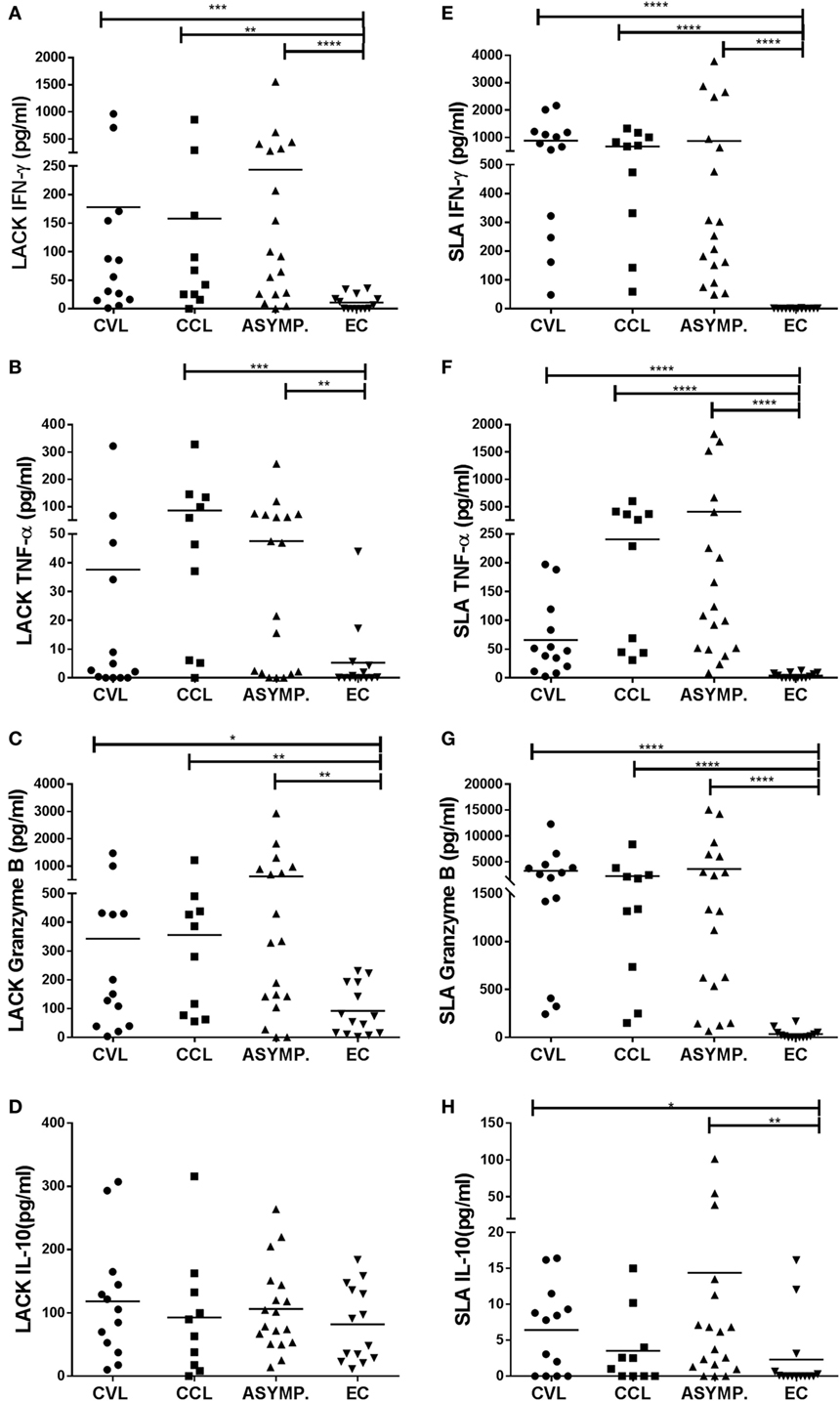
Figure 1. Production of IFN-γ, TNF-α, granzyme B, and IL-10 in peripheral blood mononuclear cells stimulated with Leishmania-activated C-kinase antigen [(A–D) respectively], and with soluble Leishmania antigen [(E–H) respectively], in healthy controls (EC, n = 14), asymptomatic subjects (ASYMP, n = 18), patients cured of cutaneous leishmaniasis (CCL, n = 10), and patients cured of visceral leishmaniasis (CVL, n = 13). *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001.
LACK Was Recognized by a High Percentage of CVL, CCL, and Asymptomatic Subjects
Following SLA stimulation, the PBMC of all CVL, CCL, and ASYMP subjects showed increased production of IFN-γ compared to the EC subjects’ cells. This cytokine was, therefore, chosen to determine the percentage of individuals that recognized LACK. IFN-γ was produced—and, therefore, LACK recognized—by 69% (9/13) of the CVL subjects, 80% of the CCL subjects (8/10), and 83% of the ASYMP subjects (15/18) (Table 1).
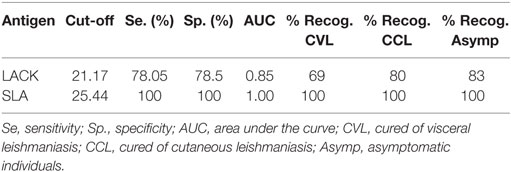
Table 1. Percentage of individuals with immune cellular memory against L. infantum who recognized Leishmania-activated C-kinase antigen and soluble Leishmania antigen.
Hamsters Vaccinated With pCI-neo-LACK/MVA-LACK Had the Highest Anti-LACK IgG Titers
The MVA-LACK and M65-LACK groups produced greater amounts of anti-IgG antibodies against LACK than did the C-DNA control group (p ≤ 0.01 and p ≤ 0.05, respectively) (Figure 2A). The C-PBS control group also produced greater amounts (p ≤ 0.05) of anti-LACK IgG antibodies than the C-DNA control group.
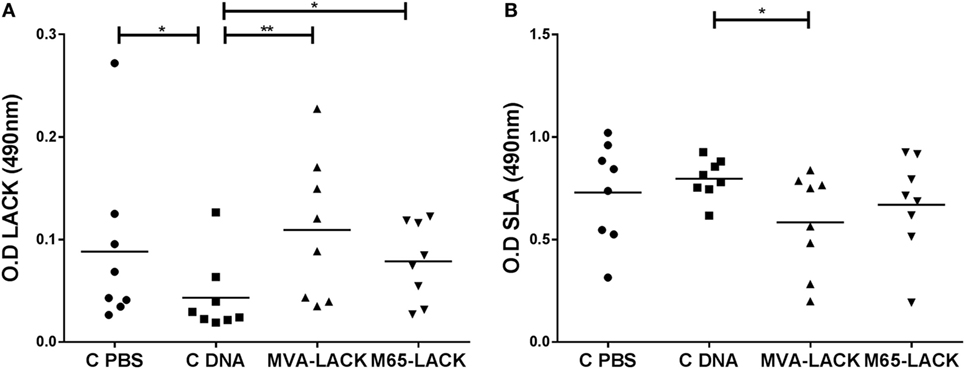
Figure 2. IgG levels against Leishmania-activated C-kinase antigen (LACK) (A) and soluble Leishmania antigen (SLA) (B) in the plasma of prime-boost immunized, C-DNA, and C-PBS group hamsters later infected with Leshmania. infantum. *p ≤ 0.05, **p ≤ 0.01.
The MVA-LACK animals produced smaller amounts of anti-IgG antibodies against SLA than did the C-DNA control animals (p ≤ 0.05) (Figure 2B).
After Stimulation With SLA, Both Vaccinated Groups of Hamsters Showed Increased PBMC Proliferation
Following stimulation with LACK, no differences were seen between any of the groups in terms of PBMC proliferation (Figure 3A). Lymphoproliferation after SLA stimulation, however, was significantly greater in the MVA-LACK group compared to the C-PBS and C-DNA controls (p ≤ 0.05). Proliferation was also greater in the M65-LACK group compared to the C-PBS group (p ≤ 0.05) (Figure 3B).
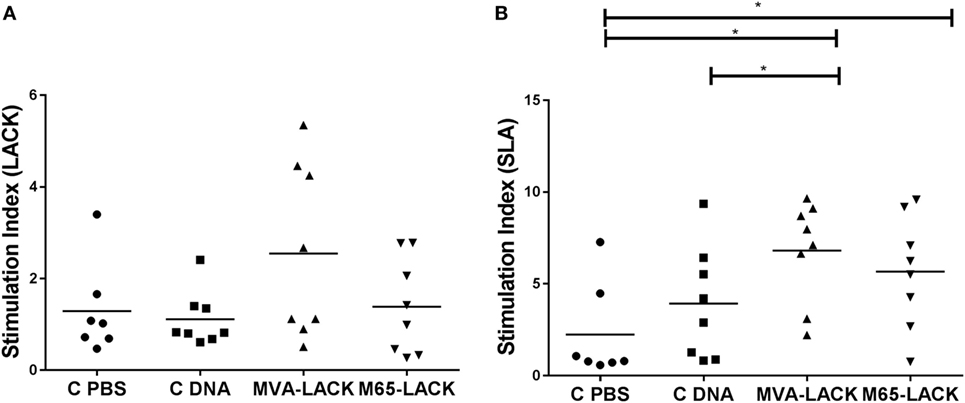
Figure 3. Post-infection peripheral blood mononuclear cells stimulation indices for C-PBS, C-DNA, and prime-boost immunized group hamsters in response to Leishmania-activated C-kinase antigen (A) and soluble Leishmania antigen (B). *p ≤ 0.05.
Vaccinated Animals Showed Less Tissue Damage and Inflammation Than Control Group Animals
Inflammatory infiltrates were observed in the four groups and in all the examined organs with the exception of the kidneys. In the liver, inflammatory infiltrates were randomly distributed and varied in extension. The infiltrates were located surrounding portal triads and central veins. Inflammation was granulomatous in all cases, was frequently organized in granulomas, and consisted of macrophages, lymphocytes, plasma cells and neutrophils (Figure 4). Giant multinucleated cells and giant Langhans cells, many containing mineral concretions (Schauman bodies), were variably present regardless of the experimental group. Giant multinucleated cells, Langhans cells, and Schauman bodies were predominantly, though not exclusively, found in discrete granulomas (Figure 4). Amastigotes were very rarely observed within macrophages.
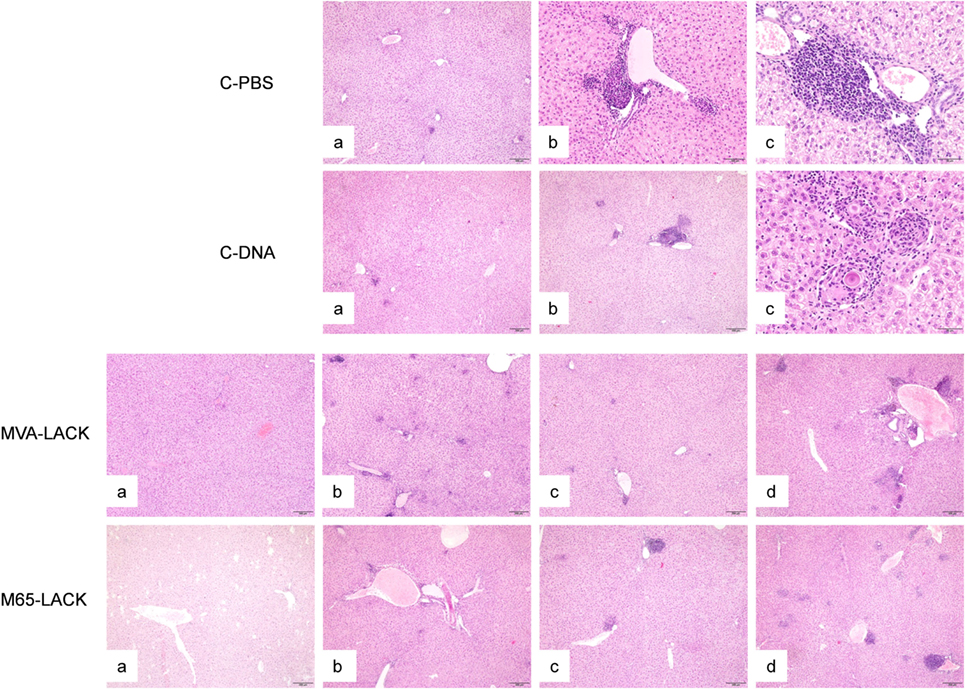
Figure 4. Liver histopathology of hamsters in the C-PBS, C-DNA, MVA-LACK, and M65-LACK groups. C-PBS group— mild, multifocal granulomatous hepatitis (a), moderate (b), and severe, multifocal, periportal, granulomatous hepatitis with intralesional Shauman bodies (c). C-DNA group—minimal hepatic inflammation (a), moderate multifocal, periportal granulomatous hepatitis (b), and severe, multifocal to coalescing granulomatous hepatitis with intralesional Shauman bodies (c). MVA-LACK group—minimal hepatic inflammation (a), mild, multifocal granulomatous hepatitis (b), moderate, multifocal, granulomatous hepatitis (c), and severe, multifocal to coalescing granulomatous hepatitis (d). M65-LACK group—minimal hepatic inflammation (a), mild, multifocal granulomatous hepatitis (b), moderate, multifocal granulomatous hepatitis (c), and severe, multifocal to coalescing, periportal, granulomatous hepatitis (d).
The spleen and bone marrow also contained variable infiltrates of granulomatous inflammation. No giant cells or Schauman bodies were observed in either organ in any of the groups.
The severity of the inflammation varied among the infected animals (Table 2). Animals in the PBS group had more severe granulomatous hepatitis than animals in the MVA-LACK group (Figure 4). The MVA-LACK group showed the least liver damage, followed by the M65-LACK group. A tendency to show inflammation in two (liver and spleen) or more organs was observed in both control groups.
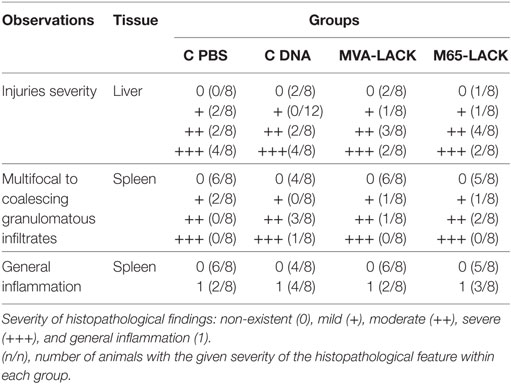
Table 2. Histopathological findings.
Vaccination With pCI-neo-LACK/MVA-LACK and pCI-neo-LACK/M65-LACK Protected Hamsters Against L. infantum
The animals of the MVA-LACK group showed a significantly smaller liver amount of Leishmania DNA than the C-PBS and C-DNA control groups (p ≤ 0.01), as did the M65-LACK animals (p ≤ 0.05) (Figure 5A). The MVA-LACK and M65-LACK treatments also led to significant reductions in the bone marrow amount of Leishmania DNA compared to the C-PBS (both p ≤ 0.01) and C-DNA control groups (p ≤ 0.001 and p ≤ 0.01, respectively) (Figure 5B). qPCR on spleen samples showed similar amount of Leishmania DNA among groups (data not shown).
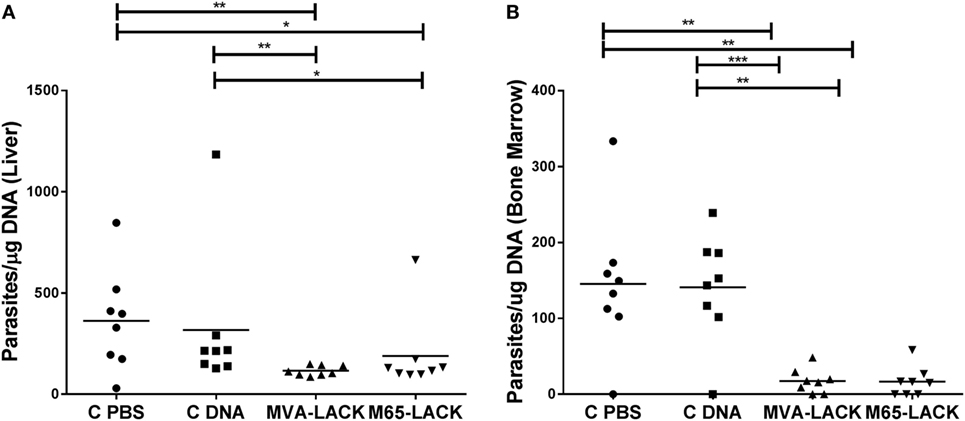
Figure 5. Liver and bone marrow amount of Leishmania DNA in the liver (A) and bone marrow (B) as determined by qPCR. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001.
Discussion
The present results show that LACK recombinant protein is recognized by a high percentage of individuals with cellular immunity against the parasite, leading to the production of the Th1 cytokines (IFN-γ and TNF-α) and granzyme B. LACK protein is an immunodominant antigen of the parasite (12), that has previously been shown to induce the production of IFN-γ in lymphocytes from patients with active CL from L. amazonensis, L. guyanensis, and L. braziliensis (7, 12), as well as IFN-γ and TNF-α in asymptomatic individuals and patients cured of VL (23, 24). This is the first study in which PBMC from subjects cured of CL caused by L. infantum have been exposed to LACK. As in other groups of infected individuals, exposure to SLA and LACK elicited a Th1 response (production of IFN-γ and TNF-α) plus the production granzyme B. Although levels of LPS were residual (equal to 0.2 endotoxin units per mg protein), it is known that LPS-triggered cytokines produced by antigen-presenting myeloid cells contribute to the induction of T cell responses (25). For that reason, we cannot discard its potential adjuvant-like phenomenon in the present results (26).
Several studies have examined the capacity of Leishmania vaccine candidates to induce cytotoxic responses or to present epitopes restricted to HLA class I (9, 27–32). These showed that LACK-stimulation of PBMC induces the secretion of IFN-γ by CD8+ T cells in exposed individuals and patients with active CL (9, 31, 32). To our knowledge, this study is the first in which the LACK-induced production of granzyme B has been quantified in cured patients of CL and VL, and asymptomatic individuals from an L. infantum-endemic area. The cellular response seen in these subjects confirms that LACK induces specific cytotoxic responses in subjects with a cellular memory response against L. infantum.
Following LACK-stimulation, the PBMC of all individuals—including those of the control groups—produced IL-10, as previously reported (7, 12, 23, 24). This production has been linked to a dominant Th2-type epitope lying between amino acids 157 and 173 (FSPSLEHPIVVSGSWDN) (12) of the antigen.
The significant recognition of LACK by the T receptors of the present cured and asymptomatic subjects, plus the ability of this protein to induce IFN-γ, TNF-α, and granzyme B (CD4+ and CD8+ T cell responses), indicates this antigen to be of potential use in vaccines against human leishmaniasis. However, the intrinsic ability of LACK to also induce IL-10 production and to expand IL-4-secreting T cells (13) illustrates the need to further optimize a LACK-based vaccine for the induction of a protective Th1 response. DNA vaccines involving the LACK antigen have returned promising results in murine models of L. major and L. amazonensis infection, as well as in a canine model of L. infantum infection (8, 17, 18, 33). In these earlier studies, the vaccines were applied under a heterologous immunization protocol, stimulating humoral, Th1-type, and cytotoxic immune responses.
The relevant effect observed in human PBMC from asymptomatic and cured patients of increase secretion of IFN-gamma and granzyme B in response to LACK, provided the rational to explore the significance of these findings in the hamster model of VL. Both the DNA-LACK/MVA-LACK and DNA-LACK/M65-LACK vaccines induced the production of anti-LACK antibodies beyond that seen in the C-DNA group. Immunization with both vaccines also induced a potent cellular response, as shown by the higher mean stimulation index to SLA observed in the immunized groups after experimental infection with L. infantum promastigotes. Vaccinia viruses are capable of inducing CD4+ and CD8+ T cell responses. In a mouse CL model, heterologous immunization with DNA-LACK/MVA-LACK has been reported to preferentially promote a CD8+ T cell-type response, whereas that generated by DNA-LACK/M65-LACK immunization is preferably of the CD4+ T cell type (17). In the present efficacy trial in hamsters, a similar immune response was observed in both the MVA-LACK and M65-LACK groups. As seen for the present response against VL in hamsters, Ramos et al. reported that dogs vaccinated with DNA-LACK/MVA-LACK showed greater lymphoproliferation after SLA-stimulation than did control animals. This specific immune response was related to the protection conferred by the vaccine in the canine model of L. infantum infection (18). In our study, the patent cellular response induced by both DNA-LACK/MVA-LACK and DNA-LACK/M65-LACK was associated with the control of the amount of Leishmania DNA observed in the liver and bone marrow. The extension and severity of the inflammatory infiltrates in the liver, observed histologically were not significant between groups, however. This could in part be explained because MVA-LACK and M65-LACK induce a more effective cellular response, as suggested by the higher lymphoproliferation values and lower parasite burdens, rather than a reduction in the quantity of the cellular response to the infection.
However, other authors report that immunization with plasmid DNA expressing the LACK antigen fails to induce any protection in mice infected with L. infantum or L. donovani (34, 35). The discrepancy with the present results is likely due to the fact that these earlier studies involved only homologous immunization with a LACK-expressing plasmid, while our studies were performed with heterologous combination of vectors, a protocol known to activate strong B and T cell immune responses to LACK antigen (31).
In conclusion, LACK is well recognized by the T cells from individuals cured of leishmaniasis, and by those of asymptomatic subjects living in a L. infantum-endemic area. Prime-boost vaccination with pCI-neo-LACK/MVA-LACK, and pCI-neo-LACK/M65-LACK, protected against L. infantum infection in a hamster model of VL. These results highlight the significance of LACK as immune activator and suggest that LACK should be considered in the formulation of a vaccine designed to induce a cellular response against VL in humans.
Ethics Statement
Work involving human subjects was approved by the Hospital de Fuenlabrada Ethics and Research Committee (APR 12-65 and APR 14-64). All participants gave their written, informed consent to be included. Work involving animals was approved by the Research Ethics and Animal Welfare Committee of the Instituto de Salud Carlos III, and performed adhering to Spanish legislation on the protection of animals used for experimentation and other scientific purposes (Royal Decree 1201/2005 and Law 32/2007; this law is a transposition of Directive 86/609/EEC).
Author Contributions
LF, LS-S, CS, AI-M, MJ, and VA conducted the experiments. LF, CS, and AI-M acquired the data. LF, EC, MJ, and VA analyzed the data. EC, LS-S, ME, and JM designed the research studies. EC, LS-S, ME, and JM provided reagents. LF, EC, and JM wrote the manuscript. All authors have read and approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Juan Víctor San Martín from the Hospital de Fuenlabrada (Madrid, Spain) and the staff of the Blood Bank and Haemotherapy Department of the same hospital, for the collection of blood samples. We also want to thank Dr. Sydney Almeida for his help in the animal experiments included in this work.
Funding
EC was supported by a contract from RD16CIII/0003/0002 Red de Enfermedades Tropicales, Subprograma RETICS del Plan Estatal de I + D + I 2013-2016, co-funded by FEDER “Una manera de hacer Europa” funds. ME was supported by AEI MINECO/FEDER grant SAF2013-45232-R.
References
1. Okwor I, Uzonna J. Social and economic burden of human leishmaniasis. Am J Trop Med Hyg (2016) 94(3):489–93. doi:10.4269/ajtmh.15-0408
2. Coler RN, Duthie MS, Hofmeyer KA, Guderian J, Jayashankar L, Vergara J, et al. From mouse to man: safety, immunogenicity and efficacy of a candidate leishmaniasis vaccine LEISH-F3+GLA-SE. Clin Transl Immunol (2015) 4(4):e35. doi:10.1038/cti.2015.6
3. Kumar R, Engwerda C. Vaccines to prevent leishmaniasis. Clin Transl Immunol (2014) 3(3):e13. doi:10.1038/cti.2014.4
4. Gillespie PM, Beaumier CM, Strych U, Hayward T, Hotez PJ, Bottazzi ME. Status of vaccine research and development of vaccines for leishmaniasis. Vaccine (2016) 34(26):2992–5. doi:10.1016/j.vaccine.2015.12.071
5. Alvar J, Croft SL, Kaye P, Khamesipour A, Sundar S, Reed SG. Case study for a vaccine against leishmaniasis. Vaccine (2013) 31(Suppl 2):B244–9. doi:10.1016/j.vaccine.2012.11.080
6. Sinha S, Kumar A, Sundaram S. A comprehensive analysis of LACK (Leishmania homologue of receptors for activated C kinase) in the context of visceral leishmaniasis. Bioinformation (2013) 9(16):832–7. doi:10.6026/97320630009832
7. Azeredo-Coutinho RB, Matos DC, Armoa GG, Maia RM, Schubach A, Mayrink W, et al. Contrasting human cytokine responses to promastigote whole-cell extract and the Leishmania analogue receptor for activated C kinase antigen of L. amazonensis in natural infection versus immunization. Clin Exp Immunol (2008) 153(3):369–75. doi:10.1111/j.1365-2249.2008.03705.x
8. Perez-Jimenez E, Kochan G, Gherardi MM, Esteban M. MVA-LACK as a safe and efficient vector for vaccination against leishmaniasis. Microbes Infect (2006) 8(3):810–22. doi:10.1016/j.micinf.2005.10.004
9. Bourreau E, Pascalis H, Prevot G, Kariminia A, Jolly N, Milon G, et al. Increased production of interferon-gamma by Leishmania homologue of the mammalian receptor for activated C kinase-reactive CD4+ T cells among human blood mononuclear cells: an early marker of exposure to Leishmania? Scand J Immunol (2003) 58(2):201–10. doi:10.1046/j.1365-3083.2003.01280.x
10. Maasho K, Satti I, Nylen S, Guzman G, Koning F, Akuffo H. A Leishmania homologue of receptors for activated C-kinase (LACK) induces both interferon-gamma and interleukin-10 in natural killer cells of healthy blood donors. J Infect Dis (2000) 182(2):570–8. doi:10.1086/315725
11. Carvalho LP, Passos S, Dutra WO, Soto M, Alonso C, Gollob KJ, et al. Effect of LACK and KMP11 on IFN-gamma production by peripheral blood mononuclear cells from cutaneous and mucosal leishmaniasis patients. Scand J Immunol (2005) 61(4):337–42. doi:10.1111/j.1365-3083.2005.01581.x
12. Bourreau E, Prevot G, Gardon J, Pradinaud R, Hasagewa H, Milon G, et al. LACK-specific CD4(+) T cells that induce gamma interferon production in patients with localized cutaneous leishmaniasis during an early stage of infection. Infect Immun (2002) 70(6):3122–9. doi:10.1128/IAI.70.6.3122-3129.2002
13. Julia V, Glaichenhaus N. CD4(+) T cells which react to the Leishmania major LACK antigen rapidly secrete interleukin-4 and are detrimental to the host in resistant B10.D2 mice. Infect Immun (1999) 67(7):3641–4.
14. Mougneau E, Altare F, Wakil AE, Zheng S, Coppola T, Wang ZE, et al. Expression cloning of a protective Leishmania antigen. Science (1995) 268(5210):563–6. doi:10.1126/science.7725103
15. Afonso LC, Scharton TM, Vieira LQ, Wysocka M, Trinchieri G, Scott P. The adjuvant effect of interleukin-12 in a vaccine against Leishmania major. Science (1994) 263(5144):235–7. doi:10.1126/science.7904381
16. Launois P, Maillard I, Pingel S, Swihart KG, Xenarios I, Acha-Orbea H, et al. IL-4 rapidly produced by V beta 4 V alpha 8 CD4+ T cells instructs Th2 development and susceptibility to Leishmania major in BALB/c mice. Immunity (1997) 6(5):541–9. doi:10.1016/S1074-7613(00)80342-8
17. Sanchez-Sampedro L, Gomez CE, Mejias-Perez E, Perez-Jimenez E, Oliveros JC, Esteban M. Attenuated and replication-competent vaccinia virus strains M65 and M101 with distinct biology and immunogenicity as potential vaccine candidates against pathogens. J Virol (2013) 87(12):6955–74. doi:10.1128/JVI.03013-12
18. Ramos I, Alonso A, Marcen JM, Peris A, Castillo JA, Colmenares M, et al. Heterologous prime-boost vaccination with a non-replicative vaccinia recombinant vector expressing LACK confers protection against canine visceral leishmaniasis with a predominant Th1-specific immune response. Vaccine (2008) 26(3):333–44. doi:10.1016/j.vaccine.2007.11.021
19. Ibarra-Meneses AV, Carrillo E, Sanchez C, Garcia-Martinez J, Lopez Lacomba D, San Martin JV, et al. Interleukin-2 as a marker for detecting asymptomatic individuals in areas where Leishmania infantum is endemic. Clin Microbiol Infect (2016) 22(8):739.e1–4. doi:10.1016/j.cmi.2016.05.021
20. Didierlaurent A, Ramirez JC, Gherardi M, Zimmerli SC, Graf M, Orbea HA, et al. Attenuated poxviruses expressing a synthetic HIV protein stimulate HLA-A2-restricted cytotoxic T-cell responses. Vaccine (2004) 22(25–26):3395–403. doi:10.1016/j.vaccine.2004.02.025
21. Carrillo E, Carrasco-Anton N, Lopez-Medrano F, Salto E, Fernandez L, San Martin JV, et al. Cytokine release assays as tests for exposure to Leishmania, and for confirming cure from leishmaniasis, in solid organ transplant recipients. PLoS Negl Trop Dis (2015) 9(10):e0004179. doi:10.1371/journal.pntd.0004179
22. Carrillo E, Jimenez MA, Sanchez C, Cunha J, Martins CM, da Paixao Seva A, et al. Protein malnutrition impairs the immune response and influences the severity of infection in a hamster model of chronic visceral leishmaniasis. PLoS One (2014) 9(2):e89412. doi:10.1371/journal.pone.0089412
23. Singh OP, Stober CB, Singh AK, Blackwell JM, Sundar S. Cytokine responses to novel antigens in an Indian population living in an area endemic for visceral leishmaniasis. PLoS Negl Trop Dis (2012) 6(10):e1874. doi:10.1371/journal.pntd.0001874
24. Stober CB, Jeronimo SM, Pontes NN, Miller EN, Blackwell JM. Cytokine responses to novel antigens in a peri-urban population in Brazil exposed to Leishmania infantum chagasi. Am J Trop Med Hyg (2012) 87(4):663–70. doi:10.4269/ajtmh.2012.12-0180
25. Morelli AE, Zahorchak AF, Larregina AT, Colvin BL, Logar AJ, Takayama T, et al. Cytokine production by mouse myeloid dendritic cells in relation to differentiation and terminal maturation induced by lipopolysaccharide or CD40 ligation. Blood (2001) 98(5):1512–23. doi:10.1182/blood.V98.5.1512
26. Gayed PM. Toward a modern synthesis of immunity: Charles A. Janeway Jr. and the immunologist’s dirty little secret. Yale J Biol Med (2011) 84(2):131–8.
27. Basu R, Roy S, Walden P. HLA class I-restricted T cell epitopes of the kinetoplastid membrane protein-11 presented by Leishmania donovani-infected human macrophages. J Infect Dis (2007) 195(9):1373–80. doi:10.1086/513439
28. Seyed N, Zahedifard F, Safaiyan S, Gholami E, Doustdari F, Azadmanesh K, et al. In silico analysis of six known Leishmania major antigens and in vitro evaluation of specific epitopes eliciting HLA-A2 restricted CD8 T cell response. PLoS Negl Trop Dis (2011) 5(9):e1295. doi:10.1371/journal.pntd.0001295
29. Naouar I, Boussoffara T, Chenik M, Gritli S, Ben Ahmed M, Belhadj Hmida N, et al. Prediction of T cell epitopes from Leishmania major potentially excreted/secreted proteins inducing granzyme B production. PLoS One (2016) 11(1):e0147076. doi:10.1371/journal.pone.0147076
30. Carrillo E, Fernandez L, Ibarra-Meneses AV, Santos MLB, Nico D, de Luca PM, et al. F1 domain of the Leishmania (Leishmania) donovani nucleoside hydrolase promotes a Th1 response in Leishmania (Leishmania) infantum cured patients and in asymptomatic individuals living in an endemic area of leishmaniasis. Front Immunol (2017) 8:750. doi:10.3389/fimmu.2017.00750
31. Bottrel RL, Dutra WO, Martins FA, Gontijo B, Carvalho E, Barral-Netto M, et al. Flow cytometric determination of cellular sources and frequencies of key cytokine-producing lymphocytes directed against recombinant LACK and soluble Leishmania antigen in human cutaneous leishmaniasis. Infect Immun (2001) 69(5):3232–9. doi:10.1128/IAI.69.5.3232-3239.2001
32. Bourreau E, Collet M, Prevot G, Milon G, Ashimoff D, Hasagewa H, et al. IFN-gamma-producing CD45RA+CD8+ and IL-10-producing CD45RA-CD4+ T cells generated in response to LACK in naive subjects never exposed to Leishmania. Eur J Immunol (2002) 32(2):510–20. doi:10.1002/1521-4141(200202)32:2<510::AID-IMMU510>3.0.CO;2-4
33. Sanchez-Sampedro L, Gomez CE, Mejias-Perez E, Sorzano CO, Esteban M. High quality long-term CD4+ and CD8+ effector memory populations stimulated by DNA-LACK/MVA-LACK regimen in Leishmania major BALB/c model of infection. PLoS One (2012) 7(6):e38859. doi:10.1371/journal.pone.0038859
34. Marques-da-Silva EA, Coelho EA, Gomes DC, Vilela MC, Masioli CZ, Tavares CA, et al. Intramuscular immunization with p36(LACK) DNA vaccine induces IFN-gamma production but does not protect BALB/c mice against Leishmania chagasi intravenous challenge. Parasitol Res (2005) 98(1):67–74. doi:10.1007/s00436-005-0008-8
Keywords: vaccine, visceral leishmaniasis, Leishmania-activated C-kinase antigen, antigenicity, cytokines, hamster
Citation: Fernández L, Carrillo E, Sánchez-Sampedro L, Sánchez C, Ibarra-Meneses AV, Jimenez MA, Almeida VA, Esteban M and Moreno J (2018) Antigenicity of Leishmania-Activated C-Kinase Antigen (LACK) in Human Peripheral Blood Mononuclear Cells, and Protective Effect of Prime-Boost Vaccination With pCI-neo-LACK Plus Attenuated LACK-Expressing Vaccinia Viruses in Hamsters. Front. Immunol. 9:843. doi: 10.3389/fimmu.2018.00843
Received: 20 November 2017; Accepted: 05 April 2018;
Published: 23 April 2018
Edited by:
Uwe Ritter, University of Regensburg, GermanyReviewed by:
Ricardo Silvestre, Instituto de Pesquisa em Ciências da Vida e da Saúde (ICVS), PortugalChristian Bogdan, Friedrich-Alexander-Universität Erlangen-Nürnberg, Germany
Copyright: © 2018 Fernández, Carrillo, Sánchez-Sampedro, Sánchez, Ibarra-Meneses, Jimenez, Almeida, Esteban and Moreno. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eugenia Carrillo, ZWNhcnJpbGxvQGlzY2lpaS5lcw==