 Stephanie C. M. van Dalen1
Stephanie C. M. van Dalen1 Arjen B. Blom1
Arjen B. Blom1 Birgitte Walgreen1Annet W. Slöetjes1Monique M. A. Helsen1Edwin J. W. Geven1Menno ter Huurne1
Birgitte Walgreen1Annet W. Slöetjes1Monique M. A. Helsen1Edwin J. W. Geven1Menno ter Huurne1 Thomas Vogl2
Thomas Vogl2 Johannes Roth2
Johannes Roth2 Fons A. J. van de Loo1
Fons A. J. van de Loo1 Marije I. Koenders1
Marije I. Koenders1 Louis Casteilla3Peter M. van der Kraan1
Louis Casteilla3Peter M. van der Kraan1 Martijn H. J. van den Bosch1
Martijn H. J. van den Bosch1 Peter L. E. M. van Lent1*
Peter L. E. M. van Lent1*- 1Experimental Rheumatology, Department of Rheumatology, Radboud University Medical Center, Nijmegen, Netherlands
- 2Institute of Immunology, University of Münster, Münster, Germany
- 3STROMALab, University of Toulouse, Toulouse, France
Background: Injection of adipose-derived mesenchymal stromal cells (ASCs) into murine knee joints after induction of inflammatory collagenase-induced osteoarthritis (CiOA) reduces development of joint pathology. This protection is only achieved when ASCs are applied in early CiOA, which is characterized by synovitis and high S100A8/A9 and IL-1β levels, suggesting that inflammation is a prerequisite for the protective effect of ASCs. Our objective was to gain more insight into the interplay between synovitis and ASC-mediated amelioration of CiOA pathology.
Methods: CiOA was induced by intra-articular collagenase injection. Knee joint sections were stained with hematoxylin/eosin and immunolocalization of polymorphonuclear cells (PMNs) and ASCs was performed using antibodies for NIMP-R14 and CD271, respectively. Chemokine expression induced by IL-1β or S100A8/A9 was assessed with qPCR and Luminex. ASC-PMN co-cultures were analyzed microscopically and with Luminex for inflammatory mediators. Migration of PMNs through transwell membranes toward conditioned medium of non-stimulated ASCs (ASCNS-CM) or IL-1β-stimulated ASCs (ASCIL-1β-CM) was examined using flow cytometry. Phagocytic capacity of PMNs was measured with labeled zymosan particles.
Results: Intra-articular saline injection on day 7 of CiOA increased synovitis after 6 h, characterized by PMNs scattered throughout the joint cavity and the synovium. ASC injection resulted in comparable numbers of PMNs which clustered around ASCs in close interaction with the synovial lining. IL-1β-stimulation of ASCs in vitro strongly increased expression of PMN-attracting chemokines CXCL5, CXCL7, and KC, whereas S100A8/A9-stimulation did not. In agreement, the number of clustered PMNs per ASC was significantly increased after 6 h of co-culturing with IL-1β-stimulated ASCs. Also migration of PMNs toward ASCIL-1β-CM was significantly enhanced (287%) when compared to ASCNS-CM. Interestingly, association of PMNs with ASCs significantly diminished KC protein release by ASCs (69% lower after 24 h), accompanied by reduced release of S100A8/A9 protein by the PMNs. Moreover, phagocytic capacity of PMNs was strongly enhanced after priming with ASCIL-1β-CM.
Conclusions: Local application of ASCs in inflamed CiOA knee joints results in clustering of attracted PMNs with ASCs in the synovium, which is likely mediated by IL-1β-induced up-regulation of chemokine release by ASCs. This results in enhanced phagocytic capacity of PMNs, enabling the clearance of debris to attenuate synovitis.
Background
Osteoarthritis (OA) affects various tissues within the joint and is characterized by destruction of cartilage, formation of osteophytes and eventually disability. Additionally, in joints of many OA patients synovial inflammation is evident (1, 2), which could be ignited by several factors including damage-associated molecular patterns (DAMPs) (3), crystals (4), or cartilage components (5). Synovitis is likely important for removal of this tissue debris, thereby promoting repair processes (6), but on the other hand, synovial inflammation may also aggravate joint destruction via prolonged release of pro-inflammatory factors and cartilage-degrading enzymes (7).
Bone marrow (BM)-derived mesenchymal stromal cells (MSCs) exhibit immunosuppressive effects, and are therefore seen as a potential therapeutic tool for various inflammatory diseases (8, 9). Because adipose-derived mesenchymal stromal cells (ASCs) share multiple properties with BM-derived MSCs, are more abundant (5% of nucleated cells in adipose tissue vs. 0.0001–0.001% in BM), and can more easily be obtained, adipose tissue is an attractive alternative source of multipotent MSCs (10).
Over the recent years, several clinical trials have been conducted to treat OA patients with ASCs (11). In multiple proof-of-concept trials, including several dose-escalation studies, no severe adverse events or complications were found (12–15). The first phase I dose-escalating clinical trial, ADIPOA, showed that ASC injection is safe and well-tolerated in patients with knee osteoarthritis, and provided encouraging preliminary evidence of efficacy (16). The mechanism by which ASCs reduce joint pathology remains largely unknown. Recently we described that local application of ASCs in murine knee joints with early collagenase-induced OA (CiOA) ameliorated end stage joint destruction (17). Moreover, ASCs rapidly suppressed synovial thickening in early CiOA, and already 6 h after intra-articular injection, synovial release of key pro-inflammatory factors S100A8/A9 and interleukin (IL-)1β had significantly diminished. Cytokine levels remained low throughout the course of the disease measured up to day 42 after induction of the CiOA model (18).
This suppressive effect was only found when ASCs were applied early after induction of CiOA when synovitis was substantial, but not when administered at a later phase when synovitis was much lower (17). Cartilage protection after early ASC injection was confirmed in the mild medial meniscal destabilization model of OA in rabbits (19). Correspondingly with these studies, intra-articular application of ASCs early after induction of the destabilization of the medial meniscus (DMM) model, a surgical model of OA in which synovial inflammation is scant, had no effect on development of joint destruction (18). These findings indicate that the protective effect of ASCs is promoted by synovitis.
Many studies have suggested that ASCs directly suppress inflammation and tissue pathology solely by producing anti-inflammatory factors (20). However, previous results from our lab suggest that other mechanisms may also be relevant. We found that ASCs associate with the inflamed synovium within 24 h after intra-articular injection, but no more ASCs were detected 5 days after injection (17). Furthermore, a recent phase I clinical trial reported that a low dose of intra-articularly injected ASCs reduced pain levels and improved joint function (16). Based on these findings we hypothesized that early application of ASCs in inflammatory OA joints results in a rapid, potent protection, which remains effective up till later stages.
The aim of the present study was to investigate the underlying mechanism of ASC-mediated amelioration of joint destruction during CiOA. We started off with a histological analysis of CiOA joints shortly after intra-articular injection of ASCs. These in vivo observations provided the basis for further examination in vitro. We analyzed the result of ASC stimulation with inflammatory factors which are present in early CiOA on their chemokine production, as well as their physical interactions with immune cells. Additionally, to identify a possible mechanism by which ASCs may interrupt the vicious cycle of inflammation and joint destruction in CiOA, the immunomodulatory effects of ASCs on the attracted immune cells were determined.
Methods
Animals
Female C57BL/6NRj mice were obtained from Janvier. Mice were used at the age of 12 weeks old and were housed in filter-top cages with corncob bedding under standard conditions. A standard diet and tap water was available ad libitum. Group sizes were based on expected variation and a power of 80%. All animal studies were according to the Dutch law and were approved by the local Animal Experimentation Committee (RU-DEC 2013-215).
Induction of CiOA
CiOA was induced by two unilateral, intra-articular injections of 1 U collagenase type VII from Clostridium histolyticum (Sigma-Aldrich) into knee joints of 16 C57BL/6NRj mice on days 0 and 2, causing damage to collateral and cruciate ligaments leading to local instability of the knee joint. This resulted in an OA-like phenotype with chronic synovial activation and cartilage destruction as was presented before in the article that initially described this OA-model (21). Day 42 was taken to model end point of the disease. Contralateral saline-injected knee joints were used as controls.
ASC Isolation and Culture
Murine ASCs were isolated from the heterogeneous crude stromal fraction of adipose tissue surrounding the inguinal lymph nodes of C57BL/6NRj donor mice, by digestion of the fat tissue with Collagenase A (Roche) and selection by adhesion onto plastic overnight. ASCs were cultured according to standard procedures in DMEM/F12 (Gibco) supplemented with 10% newborn calf serum (Sigma-Aldrich), 1% penicillin/streptomycin (Invitrogen), 0.5% amphotericin B (Invitrogen), 16 μM biotin (Sigma-Aldrich), 18 μM pantothenic acid (Sigma-Aldrich), and 100 μM ascorbic acid (Sigma-Aldrich). Purity of the population was checked by flow cytometry analysis of presence (>80%) of cell markers Sca-1, CD44, CD105, and absence (<5%) of CD11b, c-Kit, and CD34, as described earlier (17).
Human ASCs were isolated from the stromal vascular fraction from liposuctions as previously described (16). Briefly, after collagenase digestion of the fat tissue (NB6, Coger), the stromal vascular fraction was seeded and selected by adhesion onto plastic overnight. Cells were cultured according to standard procedures in MEM (MacoPharma, Tourcoing) supplemented with human platelet growth factor-enriched plasma, 10 μg/mL ciprofloxacin, and 1 U/mL heparin. Purity of the population was checked by flow cytometry analysis for presence of cell markers CD73 (>90%), CD90 (>90%), and CD105 (>80%) and absence of cell markers CD14 (<2%), CD34 (<10%), and CD45 (<2%). All cells used in experiments were <passage 3.
Intra-articular ASC Injection
On day 7 after induction of CiOA, 20,000 ASCs in 6 μL saline supplemented with 1% bovine serum albumin (BSA) fraction V (Sigma-Aldrich), were injected intra-articularly in both knees of 8 mice. Eight mice with CiOA were injected bilaterally with only saline supplemented with 1% BSA as control. In a separate experiment, three groups of 22 mice with CiOA were injected intra-articularly with either 20,000 ASCs or 100,000 ASCs in 6 μL saline supplemented with 1% BSA fraction V, or only saline supplemented with 1% BSA as control, on day 7 after induction of the model. Before injection, the viability of the ASCs was determined using trypan blue (Sigma-Aldrich) uptake and no cell death was observed.
Immunohistochemistry
Both left and right total knee joints of 16 mice were collected 6 h after ASC injection, as well as the right joints with CiOA of 33 mice on days 14 and 42 after induction of the model. Joints were fixed in 4% formalin, decalcified in 4% formic acid buffered in phosphate-buffered saline (PBS) and embedded in paraffin. Coronal sections (7 μm) were stained with hematoxylin/eosin (HE). Immunostaining of total knee joints was performed to visualize the presence of polymorphonuclear cells (PMNs) with the specific antibody NIMP-R14 (kind gift from M. Strath, National Institute for Medical Research, London, UK). Briefly, after deparaffinization and antigen retrieval with trypsin for 5 min, sections were stained for 1 h with NIMP-R14 antibody, followed by horseradish peroxidase-conjugated rabbit anti-rat antibody for 1 h. PMN influx was quantified separately as infiltrate in the synovium and exudate in the joint cavity, which was then combined to an average total PMN influx (arbitrary score, 0–3). The localization of ASCs was visualized with antibodies against CD271 (AP07713PU-N, OriGene). Briefly, after deparaffinization and antigen retrieval in citrate buffer, sections were stained for 1 h with CD271 antibody, followed by biotinylated anti-rabbit antibody for 30 min and horseradish peroxidase-conjugated avidin–biotin complexes for 30 min. All sections were developed with diaminobenzidine (Sigma-Aldrich) and counterstained with hematoxylin.
In vitro ASC Stimulation
Murine and human ASCs were plated in a 24 wells plate (80% confluency) and after adhesion overnight, cells were stimulated for 24 h with 1 μg/ml S100A8 or S100A9 [produced in our facilities (22)], various concentrations of IL-1β (0.1, 1, and 10 ng/ml) [murine: kind gift from I. G. Otterness (Pfizer Central Research, Groton, CT); human: R&D Systems], or a combination. 100 ng/ml LPS was used as a control for the presence of Toll-like receptor (TLR)4 on the cell membrane of ASCs.
Preparation of RNA and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Stimulated cells were lysed in TRI-reagent (Sigma-Aldrich) followed by RNA isolation according to the manufacturer's protocol. The RNA was reverse transcribed into cDNA as previously described (23). mRNA levels of chemokines were detected using a StepOnePlus qRT-PCR system (Applied Biosystems) using SYBRgreen master mix (Applied Biosystems) and specific primers (Biolegio; Table 1). Relative quantification of the qRT-PCR signals was performed by correcting the Ct value of the gene of interest for glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression (–ΔCt).

Table 1. Primers used for qRT-PCR.
Measurement of Protein Levels in Culture Supernatants
Culture supernatants were collected to analyze protein levels. The release of keratinocyte chemoattractant (KC) and IL-8 were measured using Luminex multianalyte technology, using the Bio-Rad Bio-Plex™ 200 System with specific magnetic beads. Protein levels of S100A8/A9 in these supernatants was quantified by a sandwich enzyme-linked immunosorbent assay (ELISA) specific for murine S100A8/A9 as described previously (24).
Clustering Assay
Murine primary bone marrow (BM)-PMNs were freshly isolated from femurs of C57BL/6NRj mice with MACS microbeads specific for Ly6B.2 (Miltenyi Biotec) according to the manufacturer's protocol. Murine ASCs were plated in a 24 wells plate (50% confluency), and after adhesion overnight, freshly isolated BM-PMNs were added in a 1:10 ratio and cultured with or without 1 ng/ml IL-1β in 50% ASC medium and 50% PMN medium [RPMI (Gibco) supplemented with 10% fetal calf serum (Sigma-Aldrich) and 1% penicillin/streptomycin (Invitrogen)]. After 3, 6, 24, and 48 h supernatants were collected for measurement of protein release upon clustering. Moreover, after 6 h, the culture medium was removed and non-adherent cells were vigorously washed away during the several steps of fixing and staining the attached cells in the plate with May-Grünwald Giemsa (MGG). The number of clustering cells was quantified in pictures taken at five separate areas in the well using ImageJ 1.46r software. A cluster was defined as an adherent ASC with at least one PMN attached on its surface.
Preparation of Conditioned Medium
Conditioned medium (CM) from ASCs was collected after 24 h culture without stimulation (ASCNS-CM), or after IL-1β stimulation (ASCIL-1β-CM). In the latter case ASCs were stimulated for 24 h with 1 ng/ml IL-1β, washed twice with saline to remove all exogenous IL-1β, and cultured for another 24 h with fresh ASC culture medium before collection. CM was filtered before use in the migration and phagocytosis assays described below and stored at −20°C when not used immediately.
Migration Assay
ASCNS-CM was supplemented with 100 ng/ml KC (R&D Systems) as positive control, and with 25 ng/ml monocyte chemotactic protein 1 (MCP-1) or 1 ng/ml IL-1β as negative controls. CMs were added to the lower compartment of 3 μm transwell inserts (Costar). 500,000 freshly isolated primary BM-PMNs in ASCNS-CM were placed in the top compartment of the transwell inserts and incubated for 2 h. Migrated cells in the lower compartment were collected and quantified with flow cytometry after addition of 123count eBeads™ Counting Beads (Thermo Fisher Scientific).
Phagocytosis Assay
Freshly MACS-isolated primary murine BM-PMNs and human blood PMNs (MACSxpress® Whole Blood Neutrophil Isolation Kit, Miltenyi Biotec) were incubated at 37°C for 1 h with ASCNS-CM, ASCIL-1β-CM (without exogenous IL-1β), or fresh ASC culture medium as negative control. After washing with PBS, cells were incubated with pHrodo™ Red Zymosan BioParticles® (Life Technologies) at 37°C for 1 h. Next, the phagocytic capacity was assessed by measuring fluorescence intensity (FI) using a Clariostar Monochromator Microplate Reader (BMG LABTECH). The FI was corrected for the average background fluorescence measured in triplicate in samples containing no cells.
Statistical Analysis
Statistical analyses were performed using Graphpad Prism version 5.03. Differences between groups were tested using a Student's t-test or a one-way analysis of variance (ANOVA), followed by a Dunnett's posttest or a Bonferroni Multiple Comparison posttest. P-values lower than 0.05 were considered significant. Results are expressed as mean values ± standard deviation (SD).
Results
PMNs Reallocate and Cluster With ASCs in Knee Joints With Early CiOA After Intra-articular ASC Injection
To investigate the acute ameliorating effects of ASCs, we injected 20,000 ASCs in murine knee joints on day 7 after induction of CiOA, when synovitis is high. An increased number of immune cells was observed within the joint cavity 6 h after intra-articular ASC injection (Figure 1B). Interestingly, this cell influx was also found in the saline-injected control CiOA knee joints (Figure 1A). Morphological screening of HE-stained sections showed that the majority of the attracted cells in both saline- and ASC-injected CiOA knee joints had a PMN phenotype (Figures 1C,D) which was confirmed by immune staining using the PMN-specific antibody NIMP-R14. We observed that injection of saline caused an influx of PMNs scattered throughout the joint (Figure 1E). Interestingly, after injection of ASCs the attracted PMNs were found in aggregates along the synovial lining layer (Figure 1F). Twenty four hours after ASC injection, the number of PMNs was greatly reduced in both the saline- and ASC-injected CiOA joints, suggesting that the reallocation of PMNs is short-lasting (Figure 1G). ASC injection in naïve joints resulted in less PMN influx and no accumulation of cells nearby the synovium (Figure 1H). Quantification of the attraction of PMNs showed that both ASC and control injections result in a significantly elevated influx of PMNs in CiOA knees compared to naïve joints (Figure 1I). This indicates that an injection in an already inflamed OA knee joint in itself causes a rapid flare leading to a transient attraction of immune cells. Interestingly, at higher magnification we observed a clustering of PMNs around cells we supposed to be ASCs based on their size (arrow in Figure 1J), which was confirmed with immunolocalization of ASCs using an antibody against the specific marker CD271 (Figure 1K). These in vivo results possibly designate an underlying mechanism for ASC-mediated joint protection. Therefore they were a prelude for the following in vitro studies in which we investigated the clustering of PMNs with ASCs in more detail to gain insight into this possible mechanism.
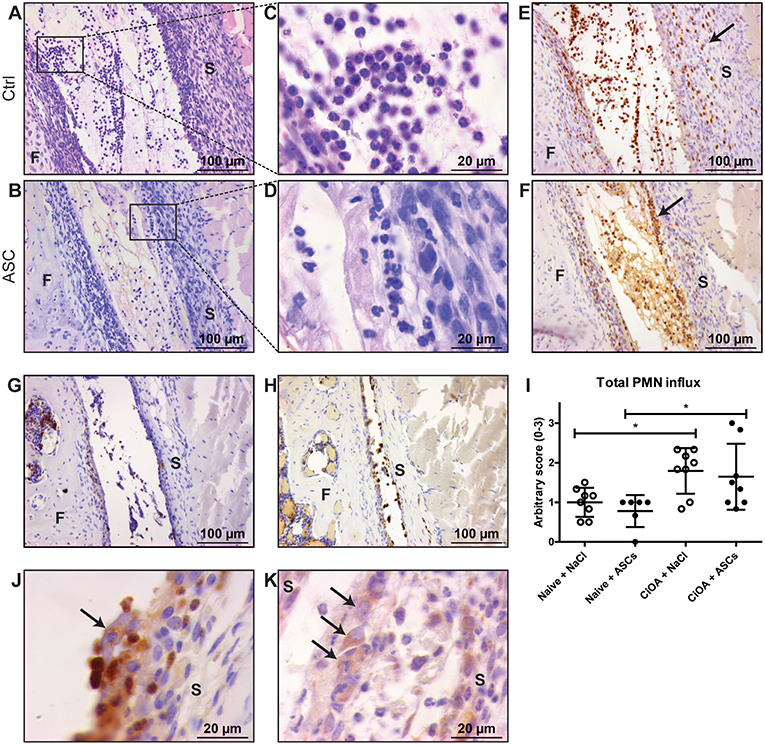
Figure 1. PMNs reallocate and cluster with ASCs in knees with early CiOA after intra-articular ASC injection. (A,B) Intra-articular injection on day 7 of CiOA resulted in attraction of immune cells within 6 h, both in the control (A) and ASC-injected (B) joints, as was shown in HE-stained total knee joint sections. (C,D) Higher magnifications showed that the immune cells in both saline- (C) and ASC-injected (D) CiOA knee joints had a polymorphonuclear (PMN) phenotype. (E,F) Immunohistochemistry with the specific antibody NIMP-R14 confirmed that a large number of PMNs is attracted to the joints. They were equally spread throughout the synovium and the joint cavity in the control-injected joints (E), but in the ASC-injected joints a remarkable accumulation of PMNs along the lining was found (F) as indicated by arrows. (G) NIMP-R14 staining showed that 24 h after ASC injection the PMN influx had largely disappeared. (H) ASC injection in naïve joints resulted in NIMP-R14 positive cell influx, albeit lower than in CiOA joints. (I) Quantification of attracted PMNs after intra-articular injection confirmed a significantly increased number of PMNs in CiOA joints compared to naïve joints. (J) At higher magnification we observed a clustering of PMNs around cells we supposed to be ASCs (arrow), which was confirmed with a CD271 antibody (K) which specifically stains ASCs (arrows). Control joints were CiOA knees injected with saline supplemented with 1% BSA. Images shown are representative for the treatment groups (n = 8 per group). Original magnification × 200 (A,B,E,F,G,H) or × 1,000 (C,D,J,K). Differences between groups were tested using a one-way ANOVA followed by a Bonferroni Multiple Comparison posttest. F, femur; S, synovium. Bars show mean values ± SD. *P < 0.05.
Expression of PMN-Attracting Chemokines by ASCs Is Elevated by IL-1β in vitro
First, we investigated whether the apparent preference of PMNs to co-localize with the ASCs is induced by inflammatory mediators that are present during inflammatory OA. Therefore, we determined if S100A8 and IL-1β, which are predominantly elevated during the early stage of CiOA, enhanced the production of the PMN-attracting chemokines C-X-C motif chemokine ligand 5 (CXCL5), CXCL7, and KC by murine ASCs. A significant elevation in gene expression of these chemokines was found after 24 h stimulation with IL-1β (Figure 2A), with a maximal increase after stimulation with only 1 ng/ml IL-1β (70, 38, and 107, fold increase for CXCL5, CXCL7, and KC, respectively, P < 0.001). KC showed the strongest up-regulation of gene expression and is generally accepted as the main PMN-attracting chemokine, which typically shows a very specific and potent attraction of PMNs. Therefore, we determined protein levels of KC as a representative PMN-attracting chemokine in culture supernatants and found a 22-fold increase after stimulation with only 1 ng/ml IL-1β (Figure 2B). Unexpectedly, stimulation with high levels of S100A8 (1 μg/ml) did not enhance chemokine gene expression or protein levels. Moreover, a combination of S100A8 and IL-1β did not alter the effect of IL-1β.
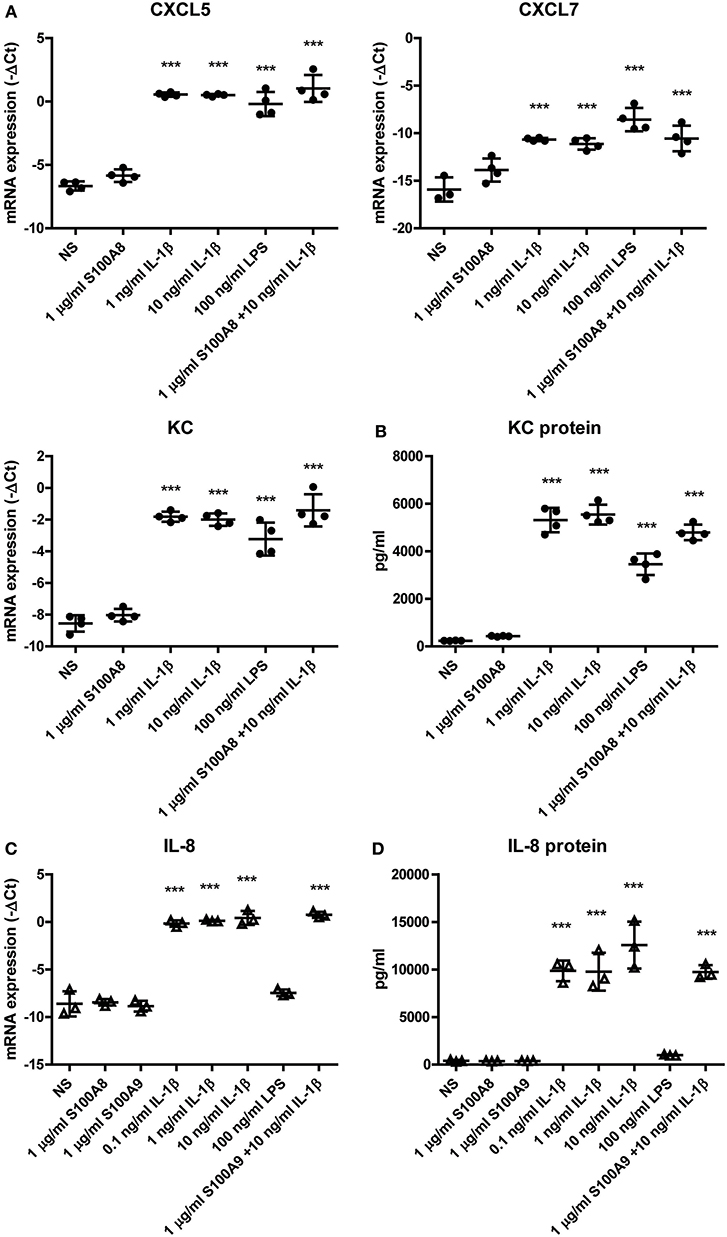
Figure 2. Expression of PMN-attracting chemokines by ASCs is elevated by IL-1β in vitro. The effect of an inflammatory milieu on gene expression in ASCs was assessed by 24 h stimulation with different pro-inflammatory mediators. (A) Murine ASCs showed a significantly increased gene expression of PMN-attracting chemokines CXCL5, CXCL7, and KC after stimulation with IL-1β. (B) Also on protein level in the supernatant, KC was significantly up-regulated after IL-1β stimulation. Human ASCs show a comparable elevated level of the functional KC-homolog IL-8 after IL-1β stimulation on both gene expression (C) and protein level (D). Gene expression levels are presented as –ΔCt compared to GAPDH. Closed circles represent murine samples, open triangles represent human samples [n = 4 (A,B) or 3 (C,D) per group]. Differences between non-stimulated (NS) ASCs and the several stimulations were tested using a one-way ANOVA followed by a Dunnett's posttest. Bars show mean values ± SD. ***P < 0.001 vs. NS ASCs.
To extrapolate our findings in murine ASCs to the human situation, we next stimulated ASCs obtained from human adipose tissue for 24 h with S100A8, S100A9, IL-1β, or a combination of S100A9 and IL-1β, to confirm that ASC-mediated attraction of PMNs is also plausible to be present in ASC-injected OA patients. Like in murine ASCs, low levels of IL-1β (0.1 ng/ml) already gave a significant 355-fold increase in gene expression (Figure 2C) and a 26-fold increase in protein levels (Figure 2D) of IL-8, the human functional homolog of KC (P < 0.001). In contrast, high levels of S100A8 and S100A9 (1 μg/ml) did not elevate IL-8 levels.
Clustering of PMNs With ASCs Is Enhanced by IL-1β in vitro
To further study the tendency of the attracted PMNs to cluster with ASCs, murine primary PMNs and adherent murine ASCs were co-cultured without stimulation (NS) or with 1 ng/ml IL-1β. After 6 h, PMNs (small cells in Figure 3A) were clearly clustered with ASCs (large cells in Figure 3A). Analysis of the number of ASCs with clustered PMNs showed a significant increase of 99% after co-culture in the presence of IL-1β (P < 0.01) (Figure 3B). Moreover, the average number of PMNs clustering per individual ASC was significantly increased with 45% after 6 h of co-culture with IL-1β (P < 0.05). These data demonstrate that IL-1β promotes an interaction between ASCs and PMNs (Figure 3C).
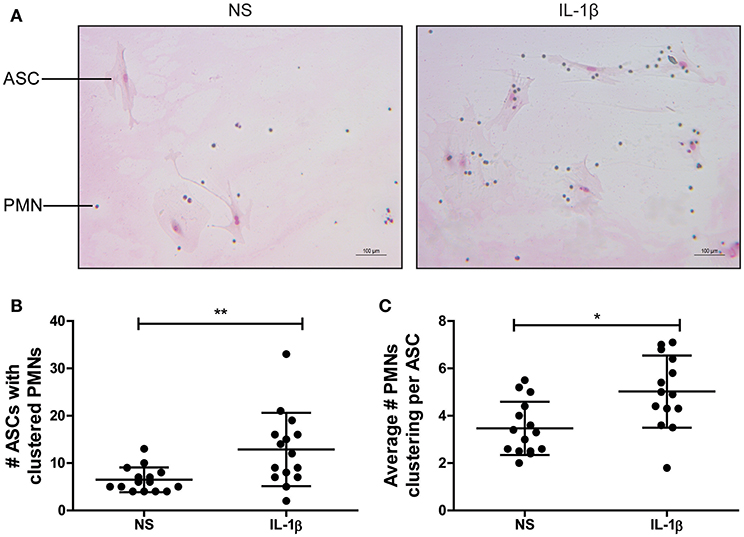
Figure 3. Clustering of murine PMNs with ASCs is enhanced by IL-1β in vitro. The association of freshly isolated murine PMNs with adherent ASCs driven by IL-1β was assessed in a co-culture experiment. (A) After 6 h, clustering cells were stained with May-Grünwald Giemsa and quantified. Large cells are ASCs, small cells are PMNs. Both the number of ASCs that formed clusters with PMNs (B) and the average number of PMNs clustered per ASC (C) were significantly increased in presence of IL-1β when compared to the non-stimulated co-cultures (NS). N = 3 per group, quantified at five separate areas per sample. Differences between the groups were tested using a Student's t-test. Bars show mean values ± SD. *P < 0.05, **P < 0.01.
PMN Attraction by ASCs Is Increased by IL-1β in vitro
In vivo the PMNs need to be actively attracted to the ASCs, before they are able to cluster. Because IL-1β strongly enhanced KC expression by ASCs, we investigated the attraction of PMNs in vitro with conditioned medium of non-stimulated ASCs (ASCNS-CM) and ASCs that were stimulated with IL-1β for 24 h (ASCIL-1β-CM). Luminex measurements confirmed that protein levels of KC in ASCIL-1β-CM were strongly increased when compared to ASCNS-CM (5,576 vs. 5.33 pg/ml) (Figure 4A). Using transwell inserts we found that CM of IL-1β-stimulated ASCs significantly increased the migration of PMNs by 287% (from 3.7 to 10.6%), confirming that ASCs attract PMNs upon IL-1β-stimulation (Figure 4B). Likewise, ASCNS-CM supplemented with KC as positive control showed an increase of PMN migration by 268% (from 3.7 to 9.9%). In contrast, the migration of PMNs toward ASCNS-CM was low (<5%), comparable to the negative controls MCP-1 and IL-1β alone.
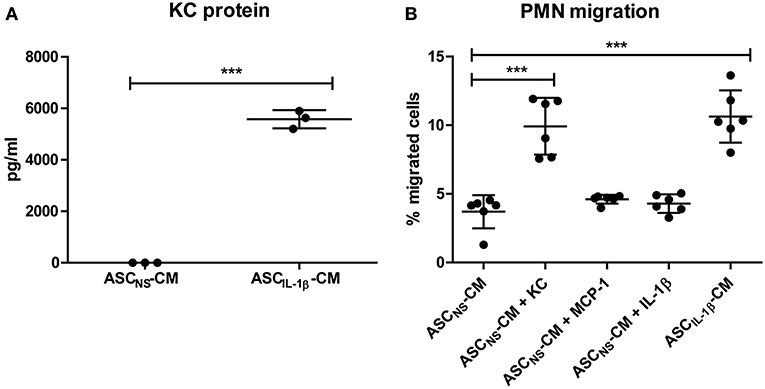
Figure 4. PMN attraction by ASCs is increased by IL-1β in vitro. The potential of IL-1β-stimulated ASCs to attract PMNs was investigated using transwell inserts. (A) KC levels in conditioned medium (CM) of IL-1β-stimulated ASCs (ASCIL-1β-CM, no exogenous IL-1β present) were significantly elevated compared to CM of non-stimulated ASCs (ASCNS-CM). (B) Migration of freshly isolated PMNs toward ASCIL-1β-CM as well as toward the positive control KC was significantly increased compared to ASCNS-CM. No enhanced migration of PMNs was found in the negative controls containing MCP-1 or IL-1β. The number of migrated PMNs was expressed as percentage of total cells added to the top compartment [n = 3 (A) or 6 (B) per group]. Differences between groups were tested using a Student's t-test (A) or a one-way ANOVA followed by a Dunnett's posttest (B). Bars show mean values ± SD. ***P < 0.001 vs. ASCNS-CM.
IL-1β-Mediated Release of Inflammatory Factors Is Significantly Lowered After Clustering of PMNs With ASCs
As attraction of PMNs and clustering with ASCs in presence of IL-1β was evident, we next determined the effect on the inflammatory activity of these cells by analyzing the release of KC protein after co-culture. Only minimal amounts of KC were released by PMNs cultured alone in the presence of IL-1β, indicating that ASCs are the main source of KC production. When no IL-1β was added, no KC was released in all co-cultures (Figure 5A), underlining that IL-1β is a potent inducer of PMN-attracting chemokine release by ASCs. In the presence of IL-1β, protein levels of KC in the supernatant of ASCs were already significantly elevated after 3 h (P < 0.001). KC levels strongly increased to maximal elevation (up to 65 ng/ml) after 24 h of stimulation with IL-1β which remained at the same level after 48 h. Interestingly, 24 h of co-culturing PMNs with ASCs in presence of IL-1β resulted in significantly lower KC release by ASCs (69 and 76% lower after 24 and 48 h incubation, respectively) (Figure 5A), suggesting that IL-1β-driven chemokine production by ASC is rapidly inhibited after clustering of PMNs. Furthermore, S100A8/A9 release by PMNs was not altered in the presence of IL-1β or in co-culture with ASCs after 3 and 6 h (Figure 5B). However, after 24 and 48 h we found strongly elevated S100A8/A9 release. S100A8/A9 release by a monoculture of ASCs with or without IL-1β stimulation was undetectable. Interestingly, in the presence of IL-1β-stimulated ASCs PMNs released lower levels of S100A8/A9 when compared to non-stimulated ASCs.
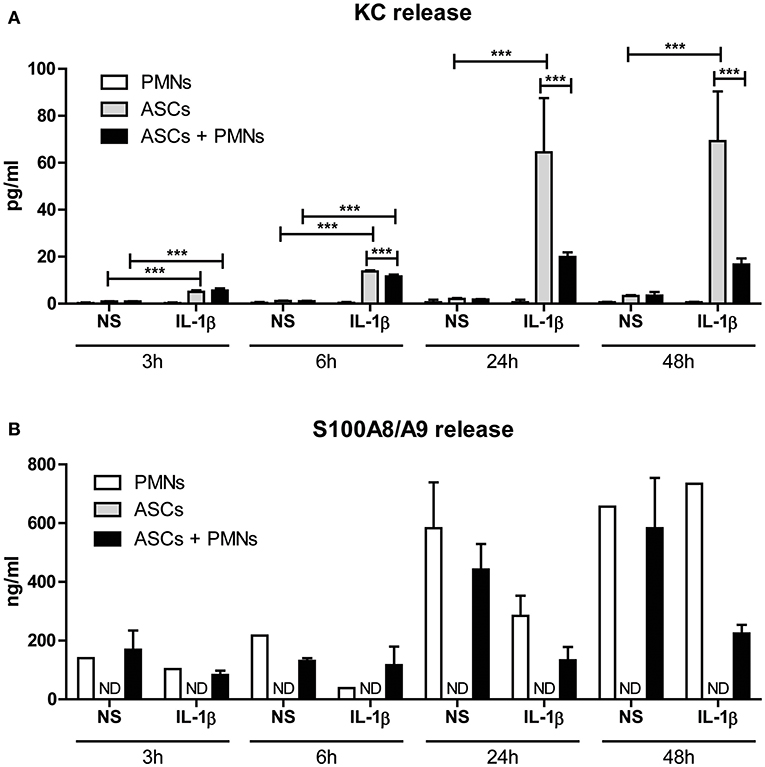
Figure 5. IL-1β-mediated release of inflammatory factors is significantly lowered after clustering of PMNs with ASCs. The effect of clustering of PMNs with ASCs on protein release was determined. (A) KC release by ASCs was already significantly increased after 3 and 6 h of co-culture with PMNs in presence of IL-1β. After 24 and 48 h of co-culture with IL-1β, clustering of PMNs with ASCs resulted in a significantly lowered KC release when compared to ASCs cultured alone. (B) S100A8/A9 release was not detectable in a monoculture of ASCs, but PMNs released basal levels of S100A8/A9. These levels increased over time, but remained lower in co-cultures of PMNs with ASCs in presence of IL-1β. N = 3 per group. Differences between groups were tested using a one-way ANOVA followed by a Bonferroni Multiple Comparison posttest for each time point separately. Bars show mean values ± SD. ***P < 0.001. ND, not detectable.
Phagocytosis by PMNs Is Enhanced After Priming With Conditioned Medium of IL-1β-Stimulated ASCs
To gain insight in the possible way in which the attraction and clustering of PMNs contributes to the amelioration of OA pathology, we finally investigated whether IL-1β-activated ASCs could enhance removal of tissue debris by inducing phagocytosis. Already after 1 h of priming murine PMNs with ASCNS-CM, ingestion of fluorescent zymosan particles was significantly increased to 198% (P < 0.001) when compared to the negative control containing only culture medium. Interestingly, priming PMNs with ASCIL-1β-CM resulted in an even further enhanced phagocytic capacity of 34% (P < 0.05) compared to ASCNS-CM (Figure 6A). We next set out to demonstrate that these results can be translated from the murine to the human situation by using human PMNs which were primed with CM of human ASCs. Again, we found a significant up-regulation of 84% (P < 0.01) in the phagocytic capacity of PMNs primed with ASCNS-CM, which was further increased with an additional 42% (P < 0.01) after priming with ASCIL-1β-CM (Figure 6B). Our findings show that IL-1β-stimulated ASCs not only attract PMNs, but also increase their phagocytic activity thereby possibly promoting tissue repair.
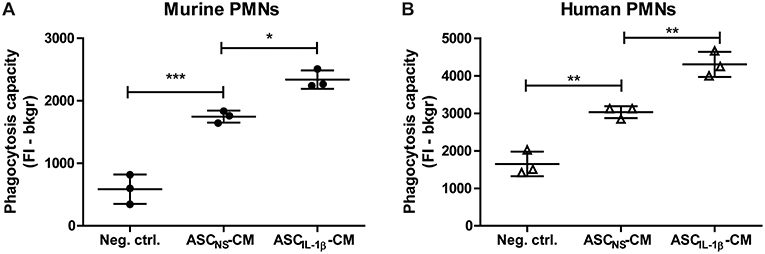
Figure 6. Phagocytosis by PMNs is enhanced after priming with conditioned medium of IL-1β-stimulated ASCs. The immunomodulatory effect of ASCs on PMNs was investigated by assessment of the phagocytic capacity after priming with ASC-conditioned medium. (A) The uptake of labeled zymosan particles by murine PMNs primed with conditioned medium (CM) of non-stimulated ASCs (ASCNS-CM) was significantly enhanced when compared to the negative control containing fresh culture medium. Incubation of PMNs with CM of ASCs which were stimulated with IL-1β (ASCIL-1β-CM, no exogenous IL-1β is present) further enhanced phagocytosis when compared to ASCNS-CM. (B) Human PMNs primed with ASCNS-CM showed a similar increase in phagocytic capacity when compared to culture medium, which was even further enhanced after priming with ASCIL-1β-CM. Closed circles represent murine samples, open triangles represent human samples (n = 3 per group). Differences between groups were tested using a one-way ANOVA followed by a Bonferroni Multiple Comparison posttest. Bars show mean values ± SD. *P < 0.05, **P < 0.01, ***P < 0.001.
Discussion
In the present study we find that local application of ASCs in inflamed CiOA knee joints leads to rapid clustering of PMNs around ASCs, which may designate a mechanism by which ASCs reduce OA pathology. In vitro we uncover that IL-1β stimulates this clustering which reduces the pro-inflammatory activity of the PMNs. At the same time the phagocytic capacity of PMNs is enhanced, which may substantially resolve inflammation and potentially accelerates joint repair processes.
ASCs are known for their regenerative capacities that enable the repair of damaged tissues such as cartilage (25), but they also possess immunosuppressive characteristics (20, 26–28). Previously, we found that ASCs rapidly suppressed synovitis and development of joint destruction when locally administered to inflamed OA joints (17, 18). However, the exact working mechanism of ASCs remained unknown, albeit several noteworthy observations have been published. Firstly, a previous study demonstrated that human ASCs applied in knee joints of normal SCID mice may reside at the site of injection up to 6 months (29). Contrarily, we showed that murine ASCs injected into an inflamed CiOA joint first home to the synovium within 24 h but disappear quickly thereafter (17). The presence of inflammation may explain the enhanced ASC clearance in CiOA knee joints. Pro-inflammatory mediators like IL-1β promote the migratory behavior of ASCs by elevating chemokine release and expression of adhesion molecules such as CD49d (integrin α4), and CD54 (ICAM-1) (30–32). Secondly, injection of numbers as low as only 20,000 ASCs more effectively protected against synovial inflammation in CiOA joints than a 5 times higher dose as shown in Supplementary Figure 1. This finding was confirmed in a recent phase I clinical trial that reported reduced pain levels and improved joint function after an intra-articular injection of a low dose of ASCs (16). Finally, ASCs only suppressed joint damage when applied in an inflammatory environment (17, 18). These three crucial observations make it unlikely that ASCs exclusively protect directly against joint damage in our murine OA model via production of anti-inflammatory mediators. This led to our hypothesis that early application of ASCs in inflammatory OA joints results in a rapid, potent protection, which remains effective up till later stages. This protection can be achieved by interactions of ASCs with the immune system, causing a powerful and long-lasting amplification of their immunosuppressive capacity.
Our hypothesis is supported by the strikingly different localization of PMNs 6 h after intra-articular injection. The saline injection flared the smoldering synovial inflammation, resulting in recruitment of PMNs that scattered throughout the joint, whereas application of ASCs induced association of PMNs with ASCs into aggregates that resided particularly on top of the inflamed synovial lining layer. That this interaction indeed has the potency to shift the inflammatory activity of ASCs and PMNs was reflected by the reduced KC and S100A8/A9 release, respectively, in our in vitro co-culture experiments. Previous studies showed that synovial lining macrophages are crucial in mediating S100A8/A9-driven synovitis during experimental OA (33, 34). The physical interaction of immunomodulatory ASC-PMN aggregates with the synovial lining may explain why synovial S100A8/A9 protein levels were already significantly reduced between 6 and 48 h after intra-articular ASC injection on day 7 of CiOA (18).
Next to the altered release of inflammatory mediators, ASCs can simultaneously direct PMNs to effectively ameliorate ongoing inflammation in OA joints via enhanced phagocytic activity. Apart from the clearance of debris, phagocytosis also causes PMNs to become apoptotic. This leads to ingestion by macrophages, which in turn start to release anti-inflammatory and reparative cytokines thereby inducing tissue repair (35). CM from non-stimulated ASCs already augmented phagocytosis by PMNs, which was further increased by IL-1β-stimulation of ASCs. The potential effect of ASCs on PMNs was underlined in a murine sepsis model, where ASC-induced phagocytosis by PMNs resulted in enhanced bacterial clearance and lowered mortality (36). In a different study, microarray analysis demonstrated that MSC treatment of mice with the cecal ligation and puncture model of sepsis resulted in enriched pathways for Fc receptor–mediated phagocytosis, natural killer cell signaling, and antigen presentation (37).
Phagocytosis is likely to be a crucial process in our CiOA model as collagenase injections damage collateral ligaments leading to mechanical instability of the joint (38). Insufficient removal of tissue debris prolongs synovitis thereby contributing to cartilage degradation and ectopic bone formation (21). Enhanced removal of tissue debris by ASC-stimulated PMNs may reduce the inflammatory response and simultaneously clear the area for more effective repair of ligaments. This would explain why we previously found less joint destruction in end stage CiOA after ASC injection (17). PMN attraction alone is not sufficient to reduce joint damage, since control injections without ASCs induced similar PMN influx but did not protect the joint from developing cartilage and bone destruction later on (17, 18). This supports our idea that ASCs applied in an inflammatory environment not only reallocate but also activate PMNs to protect the joint from further destruction.
In search of soluble factors that stimulate ASCs to attract PMNs, we focused on two pro-inflammatory mediators that characterize synovitis in early CiOA. Firstly, the contribution of the important TLR4 ligand S100A8/A9 to joint destruction during CiOA is well-described (34, 39). Elevated levels of S100A8/A9 have also been measured in OA patients (34, 40). A previous study showed that after stimulation with the TLR4 ligand LPS MSCs produce inflammatory mediators such as IL-1β, IL-6, and IL-8, indicating the expression of TLR4 on the cell membrane of ASCs (41, 42). Unexpectedly, S100A8/A9 did not induce the expression of KC or IL-8 by ASCs in vitro. This difference could be explained by the observation that TLR4 signaling by S100A8/A9 is different from that of LPS since S100A8/A9 signaling is independent of CD14 (24).
Secondly, we focused on IL-1β that is elevated in synovium and cartilage in OA patients (2, 43). Although the role of IL-1β in human OA is still a matter of debate (44), we recently showed that IL-1β does not contribute to CiOA pathology (45, 46). IL-1β directly induces the immunosuppressive phenotype of ASCs via activation of NF-κB and JNK signaling pathways (47) thereby stimulating several paracrine effects (48, 49). In the current study we found that very low levels of IL-1β strongly enhance gene and protein expression of PMN-attracting chemokines KC in murine cells and IL-8 in human cells, enabling a powerful amplification of PMN reallocation and clustering with ASCs. This in turn enables the ASC-mediated increase in phagocytosis in vitro, and possibly also in vivo. These results emphasize that although IL-1β does not aggravate joint destruction in CiOA, it may be a crucial factor for the ASC-mediated protection against joint destruction that was published before (16–18, 50). It also explains why ASC treatment is only effective when applied early in inflammatory CiOA (17), as elevated IL-1β levels are only present during early stages (34). ASC treatment after induction of CiOA in IL-1β−/− mice would be an elegant experiment to confirm the crucial role of IL-1β.
The ultimate goal is to treat human OA patients with ASCs. In line with previous mouse studies (17, 18, 27), a phase 1 human trial showed that a single injection of a low number of ASCs into knee joints of OA patients ameliorated pain and MRI analysis of ASC-injected joints indicates reduced development of joint destruction (16). Currently, a double blind phase IIb study called ADIPOA2 is performed in which 150 OA patients will be treated intra-articularly with autologous ASCs.
Conclusions
In our study we demonstrate that IL-1β-stimulated ASCs reallocate inflammatory PMNs thereby inducing cluster formation with ASCs, resulting in increased phagocytic capacity. We found this not only with murine, but also with human cells. These results may explain the previously published reduction in OA pathology after ASC injection into an inflamed joint. This more extensive knowledge of how intra-articular injection of ASCs initiates the protective effect, and particularly which factors drive this process, enables the prediction of the efficacy of ASC treatment. This paves the way for the development of well-defined inclusion criteria for OA patients.
Ethics Statement
All animal studies were according to the Dutch law and were approved by the local Animal Experimentation Committee (RU-DEC 2013-215).
Author Contributions
SvD, AB, FvdL, MK, PvdK, MvdB, and PvL: conception and design of study. SvD, BW, AS, MH, EG, MtH, TV, and JR: acquisition of data. SvD, AB, MtH, MvdB, and PvL: analysis and interpretation of data. TV, JR, and LC: contribution of reagents, materials, analysis tools. SvD, AB, MvdB, and PvL: drafting the article. SvD, AB, BW, AS, MH, EG, MtH, TV, JR, FvdL, MK, LC, PvdK, MvdB, and PvL: final approval of the submitted manuscript.
Funding
This study was financially supported by the Dutch Arthritis Foundation (Reumafonds) grant numbers 12-2-405 and 2018-1-251060, and by the European Union's Horizon 2020 research and innovation programme under grant agreement No 643809. The funding sources had no role in study design, collection, analysis or interpretation of the data, or in writing the manuscript and decision to submit the manuscript. The materials presented and views expressed here are the responsibility of the authors(s) only. The EC Commissions takes no responsibility for any use made of the information set out.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2019.01075/full#supplementary-material
Supplementary Figure 1. 20,000 ASCs protect more efficiently against synovial inflammation in CiOA than 100,000 ASCs. On day 7 after induction of CiOA, two groups of 22 mice were injected intra-articularly with either 20,000 ASCs or 100,000 ASCs in 6 μL saline supplemented with 1% bovine serum albumin (BSA) fraction V. Twenty-two mice with CiOA were injected with only saline supplemented with 1% BSA as control. On day 14 and 42, total knee joints with CiOA were collected and stained with hematoxylin/eosin (HE) to score synovial inflammation which was identified as thickening of the synovium (arbitrary score from 0 to 3, three sections per knee joint). Intra-articular injection of 20,000 ASCs resulted in significantly lower synovial inflammation in CiOA than injection of 100,000 ASCs on both day 14 and day 42. N = 11 per group. Differences between groups were tested using a one-way ANOVA followed by a Bonferroni Multiple Comparison posttest. Bars show mean values ± SD. *P < 0.05.
Abbreviations
ANOVA, one-way analysis of variance; ASC, adipose-derived mesenchymal stromal cell; BM, bone marrow; BSA, bovine serum albumin; CD, cluster of differentiation; cDNA, complementary DNA; CiOA, collagenase-induced OA; CM, conditioned medium; Ct, cycle threshold; CXCL, C-X-C motif chemokine ligand; DAMP, damage-associated molecular pattern; DMM, destabilization of the medial meniscus; ELISA, enzyme-linked immunosorbent assay; FI, fluorescence intensity; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; HE, hematoxylin/eosin; ICAM-1, intercellular adhesion molecule-1; IL, interleukin; JNK, c-Jun N-terminal kinase; KC, keratinocyte chemoattractant; LPS, lipopolysaccharide; MCP-1, monocyte chemotactic protein 1; MGG, May-Grünwald Giemsa; mRNA, messenger RNA; MSC, mesenchymal stromal cell; NF-κB, nuclear factor κB; ND, not detectable; NS, no stimulation; OA, osteoarthritis; PBS, phosphate-buffered saline; PMN, polymorphonuclear cell; qRT-PCR, quantitative real-time polymerase chain reaction; SD, standard deviation; TLR, Toll-like receptor; U, unit.
References
1. Haywood L, McWilliams DF, Pearson CI, Gill SE, Ganesan A, Wilson D, et al. Inflammation and angiogenesis in osteoarthritis. Arthritis Rheum. (2003) 48:2173–7. doi: 10.1002/art.11094
2. Benito MJ, Veale DJ, FitzGerald O, van den Berg WB, Bresnihan B. Synovial tissue inflammation in early and late osteoarthritis. Ann Rheum Dis. (2005) 64:1263–7. doi: 10.1136/ard.2004.025270
3. Liu-Bryan R. Synovium and the innate inflammatory network in osteoarthritis progression. Curr Rheumatol Rep. (2013) 15:323. doi: 10.1007/s11926-013-0323-5
4. Sun Y, Mauerhan DR, Franklin AM, Zinchenko N, Norton HJ, Hanley EN Jr, et al. Fibroblast-like synoviocytes induce calcium mineral formation and deposition. Arthritis. (2014) 2014:812678. doi: 10.1155/2014/812678
5. Fernandez-Madrid F, Karvonen RL, Teitge RA, Miller PR, An T, Negendank WG. Synovial thickening detected by MR imaging in osteoarthritis of the knee confirmed by biopsy as synovitis. Magn Reson Imag. (1995) 13:177–83. doi: 10.1016/0730-725X(94)00119-N
6. Murray PJ, Wynn TA. Protective and pathogenic functions of macrophage subsets. Nat Rev Immunol. (2011) 11:723–37. doi: 10.1038/nri3073
7. Sellam J, Berenbaum F. The role of synovitis in pathophysiology and clinical symptoms of osteoarthritis. Nat Rev Rheumatol. (2010) 6:625–35. doi: 10.1038/nrrheum.2010.159
8. Maumus M, Guerit D, Toupet K, Jorgensen C, Noel D. Mesenchymal stem cell-based therapies in regenerative medicine: applications in rheumatology. Stem Cell Res Ther. (2011) 2:14. doi: 10.1186/scrt55
9. Pak J, Lee JH, Park KS, Park M, Kang LW, Lee SH. Current use of autologous adipose tissue-derived stromal vascular fraction cells for orthopedic applications. J Biomed Sci. (2017) 24:9. doi: 10.1186/s12929-017-0318-z
10. Puissant B, Barreau C, Bourin P, Clavel C, Corre J, Bousquet C, et al. Immunomodulatory effect of human adipose tissue-derived adult stem cells: comparison with bone marrow mesenchymal stem cells. Br J Haematol. (2005) 129:118–29. doi: 10.1111/j.1365-2141.2005.05409.x
11. Vadala G, Papalia R, La Verde L, Russo F, Denaro V, Rosa MA. Bone marrow concentrated cells and stromal vascular fraction cells injections for osteoarthritis treatment: a systematic review. J Biol Regul Homeostat Agents. (2016) 30(4 Suppl 1):69–76.
12. Jo CH, Lee YG, Shin WH, Kim H, Chai JW, Jeong EC, et al. Intra-articular injection of mesenchymal stem cells for the treatment of osteoarthritis of the knee: a proof-of-concept clinical trial. Stem Cells. (2014) 32:1254–66. doi: 10.1002/stem.1634
13. Fodor PB, Paulseth SG. Adipose derived stromal cell (ADSC) injections for pain management of osteoarthritis in the human knee joint. Aesthet Surg J. (2016) 36:229–36. doi: 10.1093/asj/sjv135
14. Koh YG, Choi YJ, Kwon SK, Kim YS, Yeo JE. Clinical results and second-look arthroscopic findings after treatment with adipose-derived stem cells for knee osteoarthritis. Knee Surg Sports Traumatol Arthrosc. (2015) 23:1308–16. doi: 10.1007/s00167-013-2807-2
15. Bansal H, Comella K, Leon J, Verma P, Agrawal D, Koka P, et al. Intra-articular injection in the knee of adipose derived stromal cells (stromal vascular fraction) and platelet rich plasma for osteoarthritis. J Transl Med. (2017) 15:141. doi: 10.1186/s12967-017-1242-4
16. Pers YM, Rackwitz L, Ferreira R, Pullig O, Delfour C, Barry F, et al. Adipose mesenchymal stromal cell-based therapy for severe osteoarthritis of the knee: a phase I dose-escalation trial. Stem Cells Transl Med. (2016) 5:847–56. doi: 10.5966/sctm.2015-0245
17. ter Huurne M, Schelbergen R, Blattes R, Blom A, de Munter W, Grevers LC, et al. Antiinflammatory and chondroprotective effects of intraarticular injection of adipose-derived stem cells in experimental osteoarthritis. Arthritis Rheum. (2012) 64:3604–13. doi: 10.1002/art.34626
18. Schelbergen RF, van Dalen S, ter Huurne M, Roth J, Vogl T, Noel D, et al. Treatment efficacy of adipose-derived stem cells in experimental osteoarthritis is driven by high synovial activation and reflected by S100A8/A9 serum levels. Osteoarthritis Cartilage. (2014) 22:1158–66. doi: 10.1016/j.joca.2014.05.022
19. Saulnier N, Viguier E, Perrier-Groult E, Chenu C, Pillet E, Roger T, et al. Intra-articular administration of xenogeneic neonatal Mesenchymal Stromal Cells early after meniscal injury down-regulates metalloproteinase gene expression in synovium and prevents cartilage degradation in a rabbit model of osteoarthritis. Osteoarthritis Cartilage. (2015) 23:122–33. doi: 10.1016/j.joca.2014.09.007
20. Uccelli A, Moretta L, Pistoia V. Mesenchymal stem cells in health and disease. Nat Rev Immunol. (2008) 8:726–36. doi: 10.1038/nri2395
21. van der Kraan PM, Vitters EL, van Beuningen HM, van de Putte LB, van den Berg WB. Degenerative knee joint lesions in mice after a single intra-articular collagenase injection. A new model of osteoarthritis. J Exp Pathol. (1990) 71:19–31.
22. Fassl SK, Austermann J, Papantonopoulou O, Riemenschneider M, Xue J, Bertheloot D, et al. Transcriptome assessment reveals a dominant role for TLR4 in the activation of human monocytes by the alarmin MRP8. J Immunol. (2015) 194:575–83. doi: 10.4049/jimmunol.1401085
23. Schelbergen RF, Blom AB, van den Bosch MH, Sloetjes A, Abdollahi-Roodsaz S, Schreurs BW, et al. Alarmins S100A8 and S100A9 elicit a catabolic effect in human osteoarthritic chondrocytes that is dependent on Toll-like receptor 4. Arthritis Rheum. (2012) 64:1477–87. doi: 10.1002/art.33495
24. Vogl T, Tenbrock K, Ludwig S, Leukert N, Ehrhardt C, van Zoelen MA, et al. Mrp8 and Mrp14 are endogenous activators of Toll-like receptor 4, promoting lethal, endotoxin-induced shock. Nat Med. (2007) 13:1042–9. doi: 10.1038/nm1638
25. Gu X, Li C, Yin F, Yang G. Adipose-derived stem cells in articular cartilage regeneration: current concepts and optimization strategies. Histol Histopathol. (2017) 33:11955. doi: 10.14670/HH-11-955
26. Gonzalez-Rey E, Anderson P, Gonzalez MA, Rico L, Buscher D, Delgado M. Human adult stem cells derived from adipose tissue protect against experimental colitis and sepsis. Gut. (2009) 58:929–39. doi: 10.1136/gut.2008.168534
27. Garimella MG, Kour S, Piprode V, Mittal M, Kumar A, Rani L, et al. Adipose-derived mesenchymal stem cells prevent systemic bone loss in collagen-induced arthritis. J Immunol. (2015) 195:5136–48. doi: 10.4049/jimmunol.1500332
28. Manferdini C, Maumus M, Gabusi E, Piacentini A, Filardo G, Peyrafitte JA, et al. Adipose-derived mesenchymal stem cells exert antiinflammatory effects on chondrocytes and synoviocytes from osteoarthritis patients through prostaglandin E2. Arthritis Rheum. (2013) 65:1271–81. doi: 10.1002/art.37908
29. Toupet K, Maumus M, Peyrafitte JA, Bourin P, van Lent PL, Ferreira R, et al. Long-term detection of human adipose-derived mesenchymal stem cells after intraarticular injection in SCID mice. Arthritis Rheum. (2013) 65:1786–94. doi: 10.1002/art.37960
30. De Ugarte DA, Alfonso Z, Zuk PA, Elbarbary A, Zhu M, Ashjian P, et al. Differential expression of stem cell mobilization-associated molecules on multi-lineage cells from adipose tissue and bone marrow. Immunol Lett. (2003) 89:267–70. doi: 10.1016/S0165-2478(03)00108-1
31. Ren G, Zhao X, Zhang L, Zhang J, L'Huillier A, Ling W, et al. Inflammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 in mesenchymal stem cells are critical for immunosuppression. J Immunol. (2010) 184:2321–8. doi: 10.4049/jimmunol.0902023
32. Carrero R, Cerrada I, Lledo E, Dopazo J, Garcia-Garcia F, Rubio MP, et al. IL1beta induces mesenchymal stem cells migration and leucocyte chemotaxis through NF-kappaB. Stem Cell Rev. (2012) 8:905–16. doi: 10.1007/s12015-012-9364-9
33. Blom AB, van Lent PL, Holthuysen AE, van der Kraan PM, Roth J, van Rooijen N, et al. Synovial lining macrophages mediate osteophyte formation during experimental osteoarthritis. Osteoarthritis Cartilage. (2004) 12:627–35. doi: 10.1016/j.joca.2004.03.003
34. van Lent PL, Blom AB, Schelbergen RF, Sloetjes A, Lafeber FP, Lems WF, et al. Active involvement of alarmins S100A8 and S100A9 in the regulation of synovial activation and joint destruction during mouse and human osteoarthritis. Arthritis Rheum. (2012) 64:1466–76. doi: 10.1002/art.34315
35. Wang J. Neutrophils in tissue injury and repair. Cell Tissue Res. (2018) 371:531–9. doi: 10.1007/s00441-017-2785-7
36. Hall SR, Tsoyi K, Ith B, Padera RF Jr, Lederer JA, Wang Z, et al. Mesenchymal stromal cells improve survival during sepsis in the absence of heme oxygenase-1: the importance of neutrophils. Stem Cells. (2013) 31:397–407. doi: 10.1002/stem.1270
37. Mei SH, Haitsma JJ, Dos Santos CC, Deng Y, Lai PF, Slutsky AS, et al. Mesenchymal stem cells reduce inflammation while enhancing bacterial clearance and improving survival in sepsis. Am J Respir Crit Care Med. (2010) 182:1047–57. doi: 10.1164/rccm.201001-0010OC
38. Blalock D, Miller A, Tilley M, Wang J. Joint instability and osteoarthritis. Clin Med Insights Arthritis Musculoskelet Disord. (2015) 8:15–23. doi: 10.4137/CMAMD.S22147
39. Schelbergen RF, Geven EJ, van den Bosch MH, Eriksson H, Leanderson T, Vogl T, et al. Prophylactic treatment with S100A9 inhibitor paquinimod reduces pathology in experimental collagenase-induced osteoarthritis. Ann Rheum Dis. (2015) 74:2254–8. doi: 10.1136/annrheumdis-2014-206517
40. Schelbergen RF, de Munter W, van den Bosch MH, Lafeber FP, Sloetjes A, Vogl T, et al. Alarmins S100A8/S100A9 aggravate osteophyte formation in experimental osteoarthritis and predict osteophyte progression in early human symptomatic osteoarthritis. Ann Rheum Dis. (2016) 75:218–25. doi: 10.1136/annrheumdis-2014-205480
41. Romieu-Mourez R, Francois M, Boivin MN, Bouchentouf M, Spaner DE, Galipeau J. Cytokine modulation of TLR expression and activation in mesenchymal stromal cells leads to a proinflammatory phenotype. J Immunol. (2009) 182:7963–73. doi: 10.4049/jimmunol.0803864
42. Hwa Cho H, Bae YC, Jung JS. Role of toll-like receptors on human adipose-derived stromal cells. Stem Cells. (2006) 24:2744–52. doi: 10.1634/stemcells.2006-0189
43. Towle CA, Hung HH, Bonassar LJ, Treadwell BV, Mangham DC. Detection of interleukin-1 in the cartilage of patients with osteoarthritis: a possible autocrine/paracrine role in pathogenesis. Osteoarthritis Cartilage. (1997) 5:293–300. doi: 10.1016/S1063-4584(97)80008-8
44. Chevalier X, Goupille P, Beaulieu AD, Burch FX, Bensen WG, Conrozier T, et al. Intraarticular injection of anakinra in osteoarthritis of the knee: a multicenter, randomized, double-blind, placebo-controlled study. Arthritis Rheum. (2009) 61:344–52. doi: 10.1002/art.24096
45. van Dalen SC, Blom AB, Sloetjes AW, Helsen MM, Roth J, Vogl T, et al. Interleukin-1 is not involved in synovial inflammation and cartilage destruction in collagenase-induced osteoarthritis. Osteoarthritis Cartilage. (2017) 25:385–96. doi: 10.1016/j.joca.2016.09.009
46. Nasi S, Ea HK, So A, Busso N. Revisiting the role of interleukin-1 pathway in osteoarthritis: interleukin-1alpha and−1beta, and NLRP3 inflammasome are not involved in the pathological features of the murine menisectomy model of osteoarthritis. Front Pharmacol. (2017) 8:282. doi: 10.3389/fphar.2017.00282
47. Hu E, Ding L, Miao H, Liu F, Liu D, Dou H, et al. MiR-30a attenuates immunosuppressive functions of IL-1beta-elicited mesenchymal stem cells via targeting TAB3. FEBS Lett. (2015) 589(24 Pt B):3899–907. doi: 10.1016/j.febslet.2015.11.001
48. Luo Y, Wang Y, Poynter JA, Manukyan MC, Herrmann JL, Abarbanell AM, et al. Pretreating mesenchymal stem cells with interleukin-1beta and transforming growth factor-beta synergistically increases vascular endothelial growth factor production and improves mesenchymal stem cell-mediated myocardial protection after acute ischemia. Surgery. (2012) 151:353–63. doi: 10.1016/j.surg.2011.09.033
49. Chen H, Min XH, Wang QY, Leung FW, Shi L, Zhou Y, et al. Pre-activation of mesenchymal stem cells with TNF-alpha, IL-1beta and nitric oxide enhances its paracrine effects on radiation-induced intestinal injury. Sci Rep. (2015) 5:8718. doi: 10.1038/srep08718
Keywords: CiOA, synovitis, adipose-derived mesenchymal stromal cells, interleukin-1β, chemokines, PMNs, phagocytosis
Citation: van Dalen SCM, Blom AB, Walgreen B, Slöetjes AW, Helsen MMA, Geven EJW, ter Huurne M, Vogl T, Roth J, van de Loo FAJ, Koenders MI, Casteilla L, van der Kraan PM, van den Bosch MHJ and van Lent PLEM (2019) IL-1β-Mediated Activation of Adipose-Derived Mesenchymal Stromal Cells Results in PMN Reallocation and Enhanced Phagocytosis: A Possible Mechanism for the Reduction of Osteoarthritis Pathology. Front. Immunol. 10:1075. doi: 10.3389/fimmu.2019.01075
Received: 03 December 2018; Accepted: 26 April 2019;
Published: 27 May 2019.
Edited by:
Manuela Mengozzi, Brighton and Sussex Medical School, United KingdomReviewed by:
Nicolai Miosge, University of Göttingen, GermanyPeter Watt, University of Brighton, United Kingdom
Copyright © 2019 van Dalen, Blom, Walgreen, Slöetjes, Helsen, Geven, ter Huurne, Vogl, Roth, van de Loo, Koenders, Casteilla, van der Kraan, van den Bosch and van Lent. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter L. E. M. van Lent, cGV0ZXIudmFubGVudEByYWRib3VkdW1jLm5s