 William J. Turbitt
William J. Turbitt Wendy Demark-Wahnefried
Wendy Demark-Wahnefried Courtney M. Peterson
Courtney M. Peterson Lyse A. Norian1,2,3*†
Lyse A. Norian1,2,3*†- 1Nutrition Obesity Research Center, University of Alabama at Birmingham, Birmingham, AL, United States
- 2Department of Nutrition Sciences, University of Alabama at Birmingham, Birmingham, AL, United States
- 3O'Neal Comprehensive Cancer Center, University of Alabama at Birmingham, Birmingham, AL, United States
There is growing interest in harnessing lifestyle and pharmaceutical interventions to boost immune function, reduce tumor growth, and improve cancer treatment efficacy while reducing treatment toxicity. Interventions targeting glucose metabolism are particularly promising, as they have the potential to directly inhibit tumor cell proliferation. However, because anti-tumor immune effector cells also rely on glycolysis to sustain their clonal expansion and function, it remains unclear whether glucose-modulating therapies will support or hinder anti-tumor immunity. In this perspective, we summarize a growing body of literature that evaluates the effects of intermittent fasting, calorie restriction mimetics, and anti-hyperglycemic agents on anti-tumor immunity and immunotherapy outcomes. Based on the limited data currently available, we contend that additional pre-clinical studies and clinical trials are warranted to address the effects of co-administration of anti-hyperglycemic agents or glucose-lowering lifestyle modifications on anti-tumor immunity and cancer treatment outcomes. We stress that there is currently insufficient evidence to provide recommendations regarding these interventions to cancer patients undergoing immunotherapy. However, if found to be safe and effective in clinical trials, interventions targeting glucose metabolism could act as low-cost combinatorial adjuvants for cancer patients receiving immune checkpoint blockade or other immunotherapies.
Introduction
Cancer encompasses a broad family of diseases that involve abnormal and unregulated cell proliferation. Hanahan and Weinberg (1) have detailed the underlying characteristics that all cancers possess, including sustained proliferative signals, dysregulated cellular energetics, avoidance of immune-mediated killing, tumor-promoting inflammation, invasion, and metastasis. These factors promote a feed-forward loop favoring an immune-evading microenvironment that supports tumor progression. The balance between protective anti-tumor mechanisms and tumor-promoting/immunosuppressive factors is critical for dictating cancer progression or remission (2).
Can Glucose Metabolism be Targeted to Improve Immunotherapies?
Cancer immunotherapies are designed to enhance the protective immune responses that can eliminate established tumors and are promising treatment options for many cancers. Cancer immunotherapy comprises multiple strategies, including cytokine therapies, targeted antibodies, adoptive cell transfers, genetically engineered chimeric antigen receptor (CAR) T cells, cancer vaccines, genetically engineered oncolytic viruses, and immune checkpoint blockade (ICB). Successes have been observed within each category; however, ICB-based therapies are the most frequently utilized immunotherapy and are currently FDA-approved as treatment options in patients with many types of advanced cancers. ICB uses antibodies to disrupt the receptor/ligand pairs that send inhibitory signals to effector T cells (e.g., Programmed Death-1 [PD-1] and Programmed Death-Ligand 1 [PD-L1]) (3). Despite demonstrated clinical benefit, typically <50% of patients receiving ICB experience objective, durable responses (4, 5). This challenge has led to a major push to improve ICB efficacy by developing novel combinatorial treatment strategies to reduce cancer cell viability and proliferation, increase tumor infiltration by effector T cells, and/or promote T cell effector function in the tumor microenvironment.
One combinatorial approach that has garnered much attention in recent years is the use of glucose-limiting lifestyle changes or anti-diabetic drugs (summarized in Table 1) that can be co-administered with immunotherapy. The rationale underlying this approach is that tumor cells are often dependent on glucose as a primary energy source. This glycolytic dependency arises from the continuous proliferation of tumor cells, which necessitates uninterrupted access to energy and the building blocks of cellular biomass. To meet these requirements, cancer cells utilize glycolysis, even in the presence of oxygen, a process referred to as aerobic glycolysis or the “Warburg effect” (6). Thus, lifestyle and pharmacologic interventions that reduce intra-tumoral glucose levels may slow cancer cell replication and render cancer cells more susceptible to immune-mediated killing, thereby boosting the effectiveness of immunotherapy.
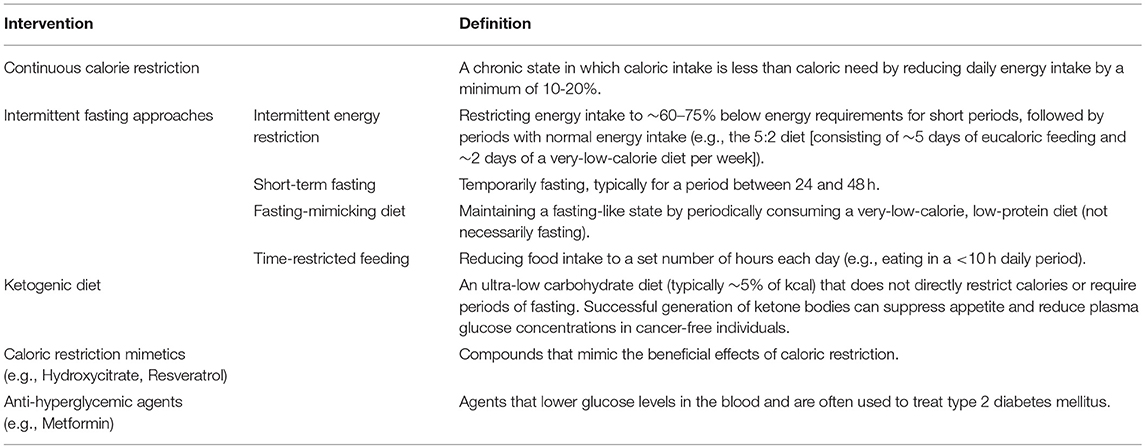
Table 1. Defining lifestyle interventions and pharmaceutical agents.
A major concern with any treatment approach focused on limiting glucose availability is that it may have unintended negative consequences for protective immunity. This is because effector CD8+ T cells also rely on glucose-dependent, Warburg-style metabolism for their clonal expansion and anti-cancer functions, including cytolytic activity and cytokine secretion (7). Prior studies report that dysregulated CD8+ T cell metabolism within the tumor microenvironment impairs T cell effector functions and promotes tumor progression (8, 9). For instance, in treatment-naive human subjects with clear cell renal cell carcinoma, tumor-infiltrating CD8+ T cells exhibit a loss of proliferative capacity due to metabolic defects, including impaired glucose uptake and glycolytic capacity; fragmented and hyperpolarized mitochondria; and increased production of reactive oxygen species (10). These observations lend validity to concerns that further limitations of intra-tumoral glucose will impair both T cell and tumor cell metabolism. However, an elegant study by Chang et al. provided evidence that ICB may selectively protect T cells from reduced glucose availability within the tumor microenvironment (8). In this report, the authors illustrated that ICB administration with either anti-CTLA-4, anti-PD-1, or anti-PD-L1 improved the glycolytic capacity and Interferon-gamma (IFNγ) production of CD8+ tumor-infiltrating T cells (8). The same study determined that anti-PD-L1 inhibited glucose uptake and glycolysis in tumor cells. Therefore, ICB may differentially alter the metabolic programming of tumor cells vs. anti-tumor immune cells to favor cancer regression. This observation makes ICB a particularly attractive type of immunotherapy to combine with glucose-limiting lifestyle interventions or anti-diabetic drugs, as the result may be impaired tumor cell metabolism and viability, with concomitantly improved T cell metabolism and effector function.
However, it remains unclear whether interventions that lower plasma glucose exert a net positive or negative effect on tumor proliferation, anti-tumor immunity, and cancer immunotherapy outcomes, particularly in the context of ICB. Minimal pre-clinical data exists, and no clinical trials have been conducted to determine if glucose-limiting lifestyle interventions or anti-diabetic drugs interact with other immunotherapy platforms, like adoptive cell therapies, cancer vaccines, or CAR T cells. These immunotherapy strategies may drive an immunometabolic profile more susceptible to reductions in glucose availability; therefore, broad-sweeping conclusions cannot be drawn on the applicability and safety of glucose-targeting therapies as an adjuvant to all immunotherapy strategies. Below, we review pre-clinical data regarding the effects of glucose-lowering interventions on tumor cell proliferation and anti-tumor immunity. Several reports have indicated that glucose-regulatory interventions may actually improve the efficacy of ICB and possibly other types of immunotherapy. When available, we also provide information about human subject data or ongoing clinical trials that are investigating these interventions in cancer patients. In light of the growing use of anti-hyperglycemic agents and surging popular interest in intermittent fasting and calorie restriction mimetics, we focus our discussion on this subset of promising interventions. Although other targeted therapies, like tyrosine kinase inhibitors (e.g., PI3K inhibitors), are promising for modulating signaling cascades relevant to glucose metabolism and for impacting immune responses following immunotherapy (11), these interventions were not discussed here because their primary modes of action are not glucose regulation.
Calorie Restriction (CR)
CR is typically defined as a reduction in daily energy intake of at least 10–20% below regular ad libitum feeding, without inducing malnutrition (Table 1). CR has been explored in pre-clinical and clinical studies for its ability to extend lifespan and improve cardiometabolic health and is now being explored for its anti-cancer properties.
Pre-clinical Findings
Abundant evidence from animal models demonstrates that CR reduces cancer incidence and delays cancer progression through multiple mechanisms (12–14). For example, CR can impair cancer cell proliferation by reducing plasma glucose and insulin, which in turn alters expression of cell cycle proteins, modifies tumor suppressor gene function, and disrupts metabolic pathways (15). CR can also reduce insulin-like growth factor-1 (IGF-1), a nutrient-sensing growth factor that is stimulated by glucose (16, 17). IGF-1 activates phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt)/ mammalian target of rapamycin 1 (mTORC1) signaling pathways in cancerous cells to promote glycolysis and tumor cell proliferation, while simultaneously inhibiting apoptosis (17–20). Thus, the pleiotropic effects of CR converge to blunt the proliferative capacity of tumor cells. Pre-clinical data suggest that CR can sensitize cancerous cells to radiotherapy and chemotherapy by negatively regulating anti-apoptotic defense mechanisms (15, 21, 22). Additionally, Farazi et al. reported that chronic CR preserved antigen-specific CD4+ T cell priming and induced a significant survival benefit when combined with anti-OX40 (CD134) immunotherapy in aged tumor-bearing mice (23). Therefore, CR appears to both inhibit tumor cell proliferation and maintain anti-tumor immunity and has the potential to be combined with immunotherapy based on this pre-clinical finding.
Clinical Findings
Despite the potential to enhance immunotherapies, concerns about loss of lean mass and aversion to CR limit therapeutic translation to cancer patients who may already be struggling with cachexia and loss of appetite. Beneficial effects have been observed in an adjuvant setting when combined with targeted therapy or chemotherapy (15); however, to date, there have been no trials examining the effects of CR on ICB in humans (Table 2). Therefore, it is not clear whether CR can be safely combined with ICB or other immunotherapies to improve patient outcomes. Given the possibility for CR to accelerate cachexia in cancer patients, such studies should be approached with caution.
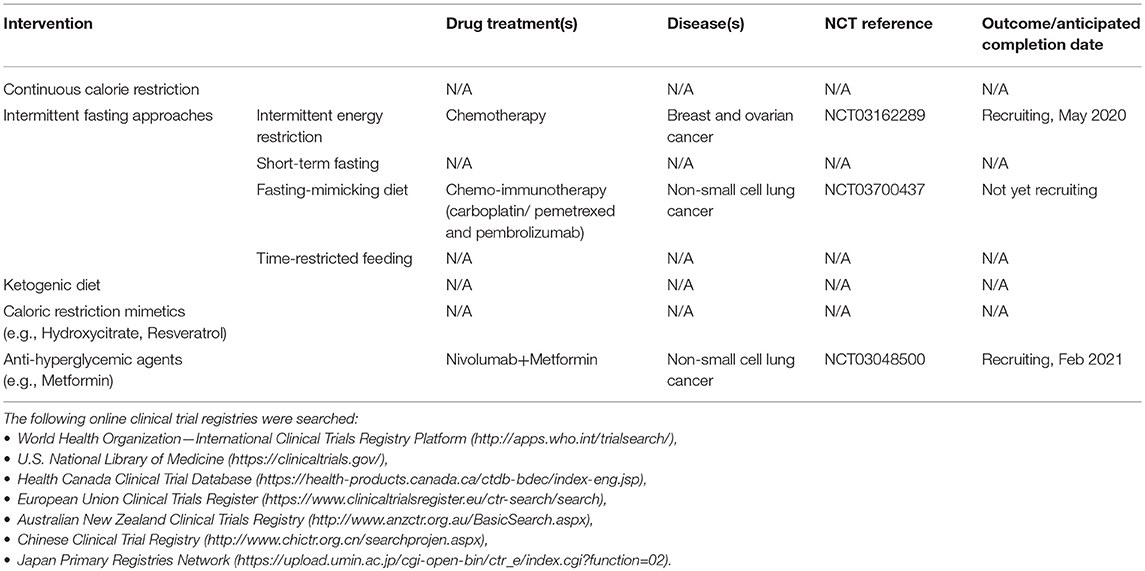
Table 2. Ongoing or completed clinical trials investigating lifestyle or pharmaceutical agents targeting glucose metabolism in combination with immunotherapy.
Intermittent Fasting
Alternatives to CR include intermittent fasting approaches such as short-term fasting, the fasting-mimicking diet (e.g., periodically consuming a very-low-calorie, low-protein diet to mimic a fasting-like state), and time-restricted feeding (eating in a ≤ 10-h daily period) (Table 1).
Pre-clinical Findings
Intermittent fasting interventions (including intermittent energy restriction) have been shown repeatedly to reduce glycemia, improve insulin sensitivity, and reduce whole-body cell proliferation rates in studies of both tumor-free animals and humans (24–29)—suggesting that these approaches target glycemic pathways and have anti-proliferative effects. Studies in tumor-bearing mice show that fasting for 72 h reduces plasma glucose concentrations by ~40% and IGF-1 by ~70% (30). The fasting-mimicking diet can achieve similar reductions in plasma glucose and IGF-1 without the need for a prolonged fast (26). Thus, intermittent fasting may provide several of the same mechanistic benefits as CR, while mitigating adverse outcomes on lean mass.
A recent literature review of pre-clinical models by Lv et al. (13) reports that intermittent fasting approaches were significantly preventive against cancer in ~60% of animal studies. Both the fasting-mimicking diet and time-restricted feeding have been associated with improved cancer treatment outcomes in all animal models of cancer tested thus far, through immunologic and metabolic mechanisms similar to those induced by CR (31–34). In particular, these two interventions have improved responses to anthracycline-based chemotherapies, leading to decreased tumor outgrowth in murine models of sarcoma, lung, colon, melanoma, and breast cancer (31, 32). Delayed tumor growth was dependent upon increased percentages of intra-tumoral cytotoxic CD8+ T cells, with concomitant reductions in regulatory T cells, and elevated expression of the stress-responsive protein heme-oxygenase-1 (HO-1) in the tumor microenvironment. Importantly, key findings, such as increased tumor immunogenicity in mice on the fasting-mimicking diet, could be achieved simply by culturing tumor cells in low glucose culture medium prior to tumor challenge, which mirrors the decreased glucose concentrations achieved in vivo with this dietary intervention (30, 32). These data suggest that limited glucose availability stresses tumor cells and ultimately may promote stronger anti-tumor immunity. Of note, anthracyclines such as mitoxantrone and doxorubicin are known to cause immunogenic tumor cell death (35), explaining their ability to act in concert with fasting-based dietary changes to improve anti-tumor immunity. At this time, no pre-clinical studies have examined the ability of intermittent fasting to enhance the efficacy of ICB or other immunotherapies.
Clinical Findings
Currently, there remains a shortage of clinical data investigating the effects of intermittent fasting on ICB efficacy in humans. To date, only one clinical trial (ISRCTN77916487) (Table 2) has investigated the effects of intermittent fasting (the 5:2 diet consisting of ~5 days of eucaloric feeding and ~2 days of a very-low-calorie diet per week) on cancer-related pathways in individuals at high risk for breast cancer (36). Although the cohort size was limited (n = 24), 55% of women showed evidence of reduced glycolysis, gluconeogenesis, glycogen synthesis, and lipid synthesis, suggesting that intermittent energy restriction altered glucose metabolism and decreased anabolic gene expression in over half of patients. Changes in breast epithelial cell differentiation were observed in some patients, but no changes in peripheral blood lymphocytes were observed. Given that responses were observed in just over half of study participants, it will be important in future studies to identify the factors determining individual responsiveness or resistance to intermittent fasting interventions.
Relatively few clinical trials are investigating the effects of intermittent fasting alone or in combination with chemo-immunotherapy in cancer patients with active disease (Table 2). One trial (NCT03700437) is investigating the ability of a fasting-mimicking diet to improve combined chemotherapy/ICB (carboplatin/pemetrexed and pembrolizumab [anti-PD-1]) outcomes in patients with advanced non-small cell lung cancer. Subjects are being provided Chemolieve® (L-Nutra, Los Angeles, CA), a plant-based, ~300 kcal per day dietary intervention ~72 h prior to and ~24 h post chemo-immunotherapy for the first four cycles of treatment. Another trial (NCT03162289) is investigating the potential interaction between chemotherapy and intermittent energy restriction (fasting with the exception of ~300–400 kcal/day of vegetable juices immediately before and after each chemotherapy cycle) in breast and ovarian cancer patients. Future clinical trials will need to determine not only the best type of intermittent fasting approach to combine with specific cancer therapies, but also the relative timing of intermittent fasting or fasting-mimicking interventions vs. administration of ICB or other immunotherapies.
Ketogenic DIET
There also is interest in using other dietary interventions, such as the ultra-low carbohydrate ketogenic diet (Table 1), to slow cancer progression and/or improve treatment efficacy. Ketogenic diets do not directly restrict calories or require periods of fasting but restrict carbohydrate to typically <5% of energy intake. Given their ability to heavily shift metabolism toward fat oxidation and drive the generation of ketone bodies, which also suppress appetite, they effectively reduce plasma glucose concentrations in cancer-free individuals (37). Thus, there is a growing interest in administering ketogenic diets to cancer patients, with the goal of leveraging reduced systemic glucose as a means of inhibiting tumor cell proliferation (38–40), possibly through the induction of oxidative cellular damage and/or increases in cytokine production and cytolysis via tumor-reactive CD8+ T cells (41, 42).
Pre-clinical Findings
Numerous pre-clinical studies show beneficial anti-cancer effects of the ketogenic diet independent of therapy or in combination with radiotherapy or chemotherapy (41–45); however, to our knowledge, no preclinical study has reported the effects of ketogenic diet on immunotherapy outcomes. Thus, it is not clear at this time whether the degree of glucose limitation that occurs during the use of a ketogenic diet will inhibit or promote anti-tumor immunity and immunotherapeutic outcomes.
Clinical Findings
The beneficial effects of the ketogenic diet observed in pre-clinical models have not translated clinically to all tumor types; however, there are promising results demonstrating the potential of the ketogenic diet to be used as an adjuvant for targeted cancer therapies, as recently reviewed by Klement (46). Ketogenic diet administration in women with ovarian or endometrial cancer, some of whom were treated with chemotherapy, showed beneficial effects on overall physical function and serum insulin, but this study was under-powered to evaluate effects on treatment outcomes or overall survival (40). However, some studies report a decrease in insulin sensitivity and increase in the inflammatory marker high-sensitivity C-reactive protein (hs-CRP) (47), raising some concerns. To our knowledge, the ketogenic diet has not been combined with immunotherapy in any clinical trials (Table 2). Given the growing lay interest in this diet as a weight loss and diabetes intervention, clinical trials designed to determine its impact on cancer treatment outcomes are urgently needed.
Calorie Restriction Mimetics and Anti-Hyperglycemic Agents
Because prolonged CR is difficult to maintain and negatively impacts lean mass, there also is mounting interest in CR “mimetics,” or compounds that produce many of the same benefits as CR without the need to reduce caloric intake (Table 1). Many CR mimetics are anti-hyperglycemic agents themselves, such as the diabetes medications metformin and acarbose, and the natural phenolic acid compound resveratrol (48–51). As summarized below, multiple studies have begun to examine whether repurposing these anti-hyperglycemic agents to target the metabolic pathways used by cancer cells and/or to reverse metabolic defects within anti-tumor immune populations can improve immunotherapy treatment outcomes.
Pre-clinical Findings
In murine models, the CR mimetic hydroxycitrate, a citric acid derivative and inhibitor of ATP citrate lyase, has shown similar efficacy to short-term fasting in mice with tumors. One study demonstrated that hydroxycitrate enhanced the ability of the anthracycline chemotherapeutic mitoxantrone to retard subcutaneous tumor outgrowth in models of fibrosarcoma and breast cancer (31). In that study, improvements in outcomes were dependent upon CD8+ T cells and decreases in regulatory T cell infiltration into tumors, as well as heightened tumor cell autophagy.
Metformin is a commonly-prescribed anti-hyperglycemic agent that has multiple mechanisms of action that include the activation of AMP-activated protein kinase (AMPK), the inhibition of Complex I of the mitochondrial respiratory chain, and the reduction in hepatic gluconeogenesis (52). Several pre-clinical studies have combined metformin (53, 54) or other anti-diabetic drugs such as phenoformin (55) with ICB. These combination therapies reduced myeloid-derived suppressor cell accumulation in tumors while simultaneously increasing proliferation and cytokine secretion in intra-tumoral CD8+ T cells, leading to a net reduction in tumor outgrowth. An explanation for these improvements in T cell function is the finding that metformin treatment of mice with B16 melanoma tumors inhibited tumor cell metabolism (both oxidative phosphorylation and glycolysis) but concurrently enhanced endogenous CD8+ T cell metabolism (oxidative phosphorylation) and cytokine production (53). To our knowledge, only one pre-clinical study has examined the effects of metformin on CD19-CAR T cells. In this study, metformin was found to inhibit CD19-CAR T cell proliferation and cytotoxicity, and induce apoptosis of these cells in vitro; metformin also suppressed the cytotoxicity of CD19-CAR T cells in vivo (56). It remains to be determined if the metabolic programming of genetically engineered, ex vivo expanded T cells differs from that present during endogenous T cell responses. Nevertheless, this is an area of intense research (57, 58) that warrants further investigation, as these results suggest that metformin may impede CD8+ T cell effector functions in some therapeutic settings.
Another popular CR mimetic is resveratrol, a food-derived compound capable of suppressing multiple signaling pathways related to cell proliferation, genome instability, and tumor angiogenesis while enhancing immunosurveillance mechanisms (59, 60). Data from in vitro (61) and in vivo (49, 62) studies demonstrate that resveratrol induces apoptosis in cancerous cells by suppressing the anti-apoptotic B cell lymphoma 2 (Bcl-2) family of regulator proteins and inhibiting nuclear factor kappa-light-chain-enhancer of activated B cells (NF-KB) and activator protein-1 (AP-1). An immunomodulator (59) and chemosensitizing agent (63, 64), resveratrol also improves interleukin (IL)-2-based immunotherapy outcomes in models of melanoma (65) and neuroblastoma (66). These improvements are mediated, in part, by increased immune cell infiltration into the tumor microenvironment, blunted expansion of regulatory T cells, and enhanced in vitro susceptibility of tumor cells to the cytotoxicity of IL-2-activated killer cells. Overall, these pre-clinical studies suggest that resveratrol both improves anti-tumor immunity and directly increases tumor susceptibility to immune-mediated killing.
Clinical Findings
In human studies, epidemiological analyses suggest that chronic administration of some types of anti-hyperglycemic medications (e.g., biguanides such as metformin) or natural polyphenols (such as resveratrol) can reduce cancer risk (50, 67, 68), but few studies have investigated their ability to be combined with immunotherapy. One retrospective cohort study (n = 55 patients) observed trending improvements in overall and progression-free survival in study participants with metastatic malignant melanoma who receive the metformin in combination with ICB (ipilimumab [anti-cytotoxic T-lymphocyte-associated protein 4 or CTLA-4], nivolumab [anti-PD-1], and/or pembrolizumab); however, these changes failed to reach statistical significance compared to ICB alone (69). Another trial is currently investigating metformin in combination with immunotherapy in non-small cell lung cancer (NCT03048500). No clinical studies have reported on the ability of hydroxycitrate or resveratrol to impact ICB efficacy; however, trials in cancer-free (70–72) and tumor-bearing (73–75) subjects suggest that resveratrol can alter systemic metabolites to improve T cell function and favor an anti-cancer response. Clearly, there is a need for larger retrospective analyses and multi-center prospective studies to evaluate the potential benefits of combining anti-hyperglycemic agents or CR mimetics, such as hydroxycitrate, metformin, or resveratrol, with ICB or other types of immunotherapy.
Limitations
As clinical data on glucose-modulating interventions discussed above are sparse or non-existent, there is a pressing need for safety and efficacy data from both animal studies and carefully conducted, prospective clinical trials in cancer patients receiving glucose-modulating interventions in the context of ICB or other immunotherapies. In particular, before proceeding to clinical trials, it is important to better understand the mechanisms of interaction of glucose-modulating interventions with the immune system. Because this information is currently lacking, we caution that there is insufficient data to justify combining glucose-modulating lifestyle-based interventions and/or anti-hyperglycemic agents with immunotherapy in advanced cancer patients. However, this is a promising area of research that warrants further investigation, particularly as several of the lifestyle interventions discussed here, such as intermittent fasting or use of over-the-counter calorie restriction mimetics, are gaining popularity in the lay public and may therefore be adopted by some cancer patients in an attempt to improve their health and treatment outcomes.
Conclusion and Outlook
Here, we have summarized existing pre-clinical murine and human subject data regarding the effects of lowering glucose availability in tumors, either via lifestyle modifications or the use of anti-hyperglycemic agents, on anti-tumor immunity and immunotherapy outcomes. Although the data are preliminary and should be interpreted cautiously, most studies indicate that such glycemic-targeting interventions do not appear to negatively compromise anti-tumor immunity in the context of ICB, and several studies provide evidence of improved T cell function and number, with simultaneous reductions in tumor cell proliferation. It is important to note that these beneficial trends may be reversed in the context of CAR T cell therapies. Future pre-clinical studies should seek to identify the mechanisms by which glycemic pathways both directly and indirectly modulate the metabolism, function, and viability of cancer cells vs. CD8+ effector T cells and other leukocyte subsets. Moreover, since many of these lifestyle and pharmacologic interventions are pleiotropic, it will also be important to determine whether there are additional immunomodulating or anti-proliferative effects induced through glucose-independent mechanisms.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
NIH Grant R01CA181088 and R21CA223126 to LN, NIH Grant R01DK118236 to CP, ACS CRP-14-111-01-CPPB and P30 CA13148-40 to WD-W, and UAB NORC post-doctoral Training Grant T32HL105349 to WT.
Disclaimer
The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. (2011) 144:646–74. doi: 10.1016/j.cell.2011.02.013
2. Hanahan D, Coussens LM. Accessories to the crime: functions of cells recruited to the tumor microenvironment. Cancer Cell. (2012) 21:309–22. doi: 10.1016/j.ccr.2012.02.022
3. Topalian SL, Drake CG, Pardoll DM. Immune checkpoint blockade: a common denominator approach to cancer therapy. Cancer Cell. (2015) 27:450–61. doi: 10.1016/j.ccell.2015.03.001
4. Emens LA, Ascierto PA, Darcy PK, Demaria S, Eggermont AMM, Redmond WL, et al. Cancer immunotherapy: opportunities and challenges in the rapidly evolving clinical landscape. Eur J Cancer. (2017) 81:116–29. doi: 10.1016/j.ejca.2017.01.035
5. Ribas A, Wolchok JD. Cancer immunotherapy using checkpoint blockade. Science. (2018) 359:1350–5. doi: 10.1126/science.aar4060
6. Pavlova NN, Thompson CB. The emerging hallmarks of cancer metabolism. Cell Metab. (2016) 23:27–47. doi: 10.1016/j.cmet.2015.12.006
7. Kishton RJ, Sukumar M, Restifo NP. Metabolic regulation of T cell longevity and function in tumor immunotherapy. Cell Metab. (2017) 26:94–109. doi: 10.1016/j.cmet.2017.06.016
8. Chang CH, Qiu J, O'Sullivan D, Buck MD, Noguchi T, Curtis JD, et al. Metabolic competition in the tumor microenvironment is a driver of cancer progression. Cell. (2015) 162:1229–41. doi: 10.1016/j.cell.2015.08.016
9. Beckermann KE, Dudzinski SO, Rathmell JC. Dysfunctional T cell metabolism in the tumor microenvironment. Cytokine Growth Factor Rev. (2017) 35:7–14. doi: 10.1016/j.cytogfr.2017.04.003
10. Siska PJ, Beckermann KE, Mason FM, Andrejeva G, Greenplate AR, Sendor AB, et al. Mitochondrial dysregulation and glycolytic insufficiency functionally impair CD8 T cells infiltrating human renal cell carcinoma. JCI Insight. (2017) 2:93411. doi: 10.1172/jci.insight.93411
11. O'Donnell JS, Massi D, Teng MWL, Mandala M. PI3K-AKT-mTOR inhibition in cancer immunotherapy, redux. Semin Cancer Biol. (2018) 48:91–103. doi: 10.1016/j.semcancer.2017.04.015
12. Simone BA, Champ CE, Rosenberg AL, Berger AC, Monti DA, Dicker AP, et al. Selectively starving cancer cells through dietary manipulation: methods and clinical implications. Future Oncol. (2013) 9:959–76. doi: 10.2217/fon.13.31
13. Lv M, Zhu X, Wang H, Wang F, Guan W. Roles of caloric restriction, ketogenic diet and intermittent fasting during initiation, progression and metastasis of cancer in animal models: a systematic review and meta-analysis. PLoS ONE. (2014) 9:e115147. doi: 10.1371/journal.pone.0115147
14. Chen Y, Ling L, Su G, Han M, Fan X, Xun P, et al. Effect of intermittent versus chronic calorie restriction on tumor incidence: a systematic review and meta-analysis of animal studies. Sci Rep. (2016) 6:33739. doi: 10.1038/srep33739
15. Kopeina GS, Senichkin VV, Zhivotovsky B. Caloric restriction - A promising anti-cancer approach: from molecular mechanisms to clinical trials. Biochim Biophys Acta. (2017) 1867:29–41. doi: 10.1016/j.bbcan.2016.11.002
16. Longo VD, Fontana L. Calorie restriction and cancer prevention: metabolic and molecular mechanisms. Trends Pharmacol Sci. (2010) 31:89–98. doi: 10.1016/j.tips.2009.11.004
17. Klement R, Fink M. Dietary and pharmacological modification of the insulin/IGF-1 system: exploiting the full repertoire against cancer. Oncogenesis. (2016) 5:e193. doi: 10.1038/oncsis.2016.2
18. Dogan S, Johannsen AC, Grande JP, Cleary MP. Effects of intermittent and chronic calorie restriction on mammalian target of rapamycin (mTOR) and IGF-I signaling pathways in mammary fat pad tissues and mammary tumors. Nutr Cancer. (2011) 63:389–401. doi: 10.1080/01635581.2011.535968
19. Bowers LW, Rossi EL, O'Flanagan CH, deGraffenried LA, Hursting SD. The role of the insulin/IGF system in cancer: lessons learned from clinical trials and the energy balance-cancer link. Front Endocrinol. (2015) 6:77. doi: 10.3389/fendo.2015.00077
20. Lyons A, Coleman M, Riis S, Favre C, O'Flanagan CH, Zhdanov AV, et al. Insulin-like growth factor 1 signaling is essential for mitochondrial biogenesis and mitophagy in cancer cells. J Biol Chem. (2017) 292:16983–98. doi: 10.1074/jbc.M117.792838
21. Nencioni A, Caffa I, Cortellino S, Longo VD. Fasting and cancer: molecular mechanisms and clinical application. Nat Rev Cancer. (2018) 18:707–19. doi: 10.1038/s41568-018-0061-0
22. O'Flanagan CH, Smith LA, McDonell SB, Hursting SD. When less may be more: calorie restriction and response to cancer therapy. BMC Med. (2017) 15:106. doi: 10.1186/s12916-017-0873-x
23. Farazi M, Nguyen J, Goldufsky J, Linnane S, Lukaesko L, Weinberg AD, et al. Caloric restriction maintains OX40 agonist-mediated tumor immunity and CD4 T cell priming during aging. Cancer Immunol Immunother. (2014) 63:615–26. doi: 10.1007/s00262-014-1542-y
24. Varady KA, Roohk DJ, Hellerstein MK. Dose effects of modified alternate-day fasting regimens on in vivo cell proliferation and plasma insulin-like growth factor-1 in mice. J Appl Physiol. (2007) 103:547–51. doi: 10.1152/japplphysiol.00209.2007
25. Varady KA, Roohk DJ, Bruss M, Hellerstein MK. Alternate-day fasting reduces global cell proliferation rates independently of dietary fat content in mice. Nutrition. (2009) 25:486–91. doi: 10.1016/j.nut.2008.10.017
26. Brandhorst S, Choi IY, Wei M, Cheng CW, Sedrakyan S, Navarrete G, et al. A periodic diet that mimics fasting promotes multi-system regeneration, enhanced cognitive performance, and healthspan. Cell Metab. (2015) 22:86–99. doi: 10.1016/j.cmet.2015.05.012
27. Wei M, Brandhorst S, Shelehchi M, Mirzaei H, Cheng CW, Budniak J, et al. Fasting-mimicking diet and markers/risk factors for aging, diabetes, cancer, and cardiovascular disease. Sci Transl Med. (2017) 9:eaai8700. doi: 10.1007/978-94-024-1045-7
28. Patterson RE, Sears DD. Metabolic effects of intermittent fasting. Ann Rev Nutr. (2017) 37:371–93. doi: 10.1146/annurev-nutr-071816-064634
29. Sutton EF, Beyl R, Early KS, Cefalu WT, Ravussin E, Peterson CM. Early time-restricted feeding improves insulin sensitivity, blood pressure, and oxidative stress even without weight loss in men with prediabetes. Cell Metab. (2018) 27:1212–21.e1213. doi: 10.1016/j.cmet.2018.04.010
30. Lee C, Safdie FM, Raffaghello L, Wei M, Madia F, Parrella E, et al. Reduced levels of IGF-I mediate differential protection of normal and cancer cells in response to fasting and improve chemotherapeutic index. Cancer Res. (2010) 70:1564–72. doi: 10.1158/0008-5472.CAN-09-3228
31. Pietrocola F, Pol J, Vacchelli E, Rao S, Enot DP, Baracco EE, et al. Caloric restriction mimetics enhance anticancer immunosurveillance. Cancer Cell. (2016) 30:147–60. doi: 10.1016/j.ccell.2016.05.016
32. Di Biase S, Lee C, Brandhorst S, Manes B, Buono R, Cheng CW, et al. Fasting-mimicking diet reduces HO-1 to promote T cell-mediated tumor cytotoxicity. Cancer Cell. (2016) 30:136–46. doi: 10.1016/j.ccell.2016.06.005
33. Harvie MN, Howell T. Could intermittent energy restriction and intermittent fasting reduce rates of cancer in obese, overweight, and normal-weight subjects? A summary of evidence. Adv Nutr. (2016) 7:690–705. doi: 10.3945/an.115.011767
34. Sundaram S, Yan L. Time-restricted feeding mitigates high-fat diet-enhanced mammary tumorigenesis in MMTV-PyMT mice. Nutr Res. (2018) 59:72–9. doi: 10.1016/j.nutres.2018.07.014
35. Bezu L, Gomes-de-Silva LC, Dewitte H, Breckpot K, Fucikova J, Spisek R, et al. Combinatorial strategies for the induction of immunogenic cell death. Front Immunol. (2015) 6:187. doi: 10.3389/fimmu.2015.00187
36. Harvie MN, Sims AH, Pegington M, Spence K, Mitchell A, Vaughan AA, et al. Intermittent energy restriction induces changes in breast gene expression and systemic metabolism. Breast Cancer Res. (2016) 18:57. doi: 10.1186/s13058-016-0714-4
37. Boden G, Sargrad K, Homko C, Mozzoli M, Stein TP. Effect of a low-carbohydrate diet on appetite, blood glucose levels, and insulin resistance in obese patients with type 2 diabetes. Ann Intern Med. (2005) 142:403–11. doi: 10.7326/0003-4819-142-6-200503150-00006
38. Allen BG, Bhatia SK, Anderson CM, Eichenberger-Gilmore JM, Sibenaller ZA, Mapuskar KA, et al. Ketogenic diets as an adjuvant cancer therapy: History and potential mechanism. Redox Biol. (2014) 2:963–70. doi: 10.1016/j.redox.2014.08.002
39. Wright C, Simone NL. Obesity and tumor growth: inflammation, immunity, and the role of a ketogenic diet. Curr Opin Clin Nutr Metab Care. (2016) 19:294–9. doi: 10.1097/MCO.0000000000000286
40. Cohen CW, Fontaine KR, Arend RC, Alvarez RD, Leath CAIII, Huh WK, et al. A ketogenic diet reduces central obesity and serum insulin in women with ovarian or endometrial cancer. J Nutr. (2018) 148:1253–60. doi: 10.1093/jn/nxy119
41. Allen BG, Bhatia SK, Buatti JM, Brandt KE, Lindholm KE, Button AM, et al. Ketogenic diets enhance oxidative stress and radio-chemo-therapy responses in lung cancer xenografts. Clin Cancer Res. (2013) 19:3905–13. doi: 10.1158/1078-0432.CCR-12-0287
42. Lussier DM, Woolf EC, Johnson JL, Brooks KS, Blattman JN, Scheck AC. Enhanced immunity in a mouse model of malignant glioma is mediated by a therapeutic ketogenic diet. BMC Cancer. (2016) 16:310. doi: 10.1186/s12885-016-2337-7
43. Weber DD, Aminazdeh-Gohari S, Kofler B. Ketogenic diet in cancer therapy. Aging. (2018) 10:164. doi: 10.18632/aging.101382
44. Klement RJ, Champ CE, Otto C, Kammerer U. Anti-tumor effects of ketogenic diets in mice: a meta-analysis. PLoS ONE. (2016) 11:e0155050. doi: 10.1371/journal.pone.0155050
45. Morscher RJ, Aminzadeh-Gohari S, Feichtinger RG, Mayr JA, Lang R, Neureiter D, et al. Inhibition of neuroblastoma tumor growth by ketogenic diet and/or calorie restriction in a CD1-Nu mouse model. PLoS ONE. (2015) 10:e0129802. doi: 10.1371/journal.pone.0129802
46. Klement RJ. The emerging role of ketogenic diets in cancer treatment. Curr Opin Clin Nutr Metab Care. (2019) 22:129–34. doi: 10.1097/MCO.0000000000000540
47. Rosenbaum M, Hall KD, Guo J, Ravussin E, Mayer LS, Reitman ML, et al. Glucose and lipid homeostasis and inflammation in humans following an isocaloric ketogenic diet. Obesity. (2019) 27:971–81. doi: 10.1002/oby.22468
48. Vernieri C, Casola S, Foiani M, Pietrantonio F, de Braud F, Longo V. Targeting cancer metabolism: dietary and pharmacologic interventions. Cancer Discov. (2016) 6:1315–33. doi: 10.1158/2159-8290.CD-16-0615
49. Mohamed SIA, Jantan I, Haque MA. Naturally occurring immunomodulators with antitumor activity: an insight on their mechanisms of action. Int Immunopharmacol. (2017) 50:291–304. doi: 10.1016/j.intimp.2017.07.010
50. Laskar J, Bhattacharjee K, Sengupta M, Choudhury Y. Anti-diabetic drugs: cure or risk factors for cancer? Pathol Oncol Res. (2018) 24:745–55. doi: 10.1007/s12253-018-0402-z
51. Trung LQ, An DTT. Is resveratrol a cancer immunomodulatory molecule? Front Pharmacol. (2018) 9:1255. doi: 10.3389/fphar.2018.01255
52. Rena G, Hardie DG, Pearson ER. The mechanisms of action of metformin. Diabetologia. (2017) 60:1577–85. doi: 10.1007/s00125-017-4342-z
53. Scharping NE, Menk AV, Whetstone RD, Zeng X, Delgoffe GM. Efficacy of PD-1 blockade is potentiated by metformin-induced reduction of tumor hypoxia. Cancer Immunol Res. (2017) 5:9–16. doi: 10.1158/2326-6066.CIR-16-0103
54. Cha JH, Yang WH, Xia W, Wei Y, Chan LC, Lim SO, et al. Metformin promotes antitumor immunity via endoplasmic-reticulum-associated degradation of PD-L1. Mol Cell. (2018) 71:606–20.e607. doi: 10.1016/j.molcel.2018.07.030
55. Kim SH, Li M, Trousil S, Zhang Y, Pasca di Magliano M, Swanson KD, et al. Phenformin inhibits myeloid-derived suppressor cells and enhances the anti-tumor activity of PD-1 blockade in melanoma. J Invest Dermatol. (2017) 137:1740–8. doi: 10.1016/j.jid.2017.03.033
56. Mu Q, Jiang M, Zhang Y, Wu F, Li H, Zhang W, et al. Metformin inhibits proliferation and cytotoxicity and induces apoptosis via AMPK pathway in CD19-chimeric antigen receptor-modified T cells. Onco Targets Ther. (2018) 11:1767–76. doi: 10.2147/OTT.S154853
57. Li X, Wenes M, Romero P, Huang SC, Fendt SM, Ho PC. Navigating metabolic pathways to enhance antitumour immunity and immunotherapy. Nat Rev Clin Oncol. (2019). doi: 10.1038/s41571-019-0203-7. [Epub ahead of print].
58. Schurich A, Magalhaes I, Mattsson J. Metabolic regulation of CAR T cell function by the hypoxic microenvironment in solid tumors. Immunotherapy. (2019) 11:335–45. doi: 10.2217/imt-2018-0141
59. Švajger U, Jeras M. Anti-inflammatory effects of resveratrol and its potential use in therapy of immune-mediated diseases. Int Rev Immunol. (2012) 31:202–22. doi: 10.3109/08830185.2012.665108
60. Varoni EM, Lo Faro AF, Sharifi-Rad J, Iriti M. Anticancer molecular mechanisms of resveratrol. Front Nutr. (2016) 3:8. doi: 10.3389/fnut.2016.00008
61. Hu L, Cao D, Li Y, He Y, Guo K. Resveratrol sensitized leukemia stem cell-like KG-1a cells to cytokine-induced killer cells-mediated cytolysis through NKG2D ligands and TRAIL receptors. Cancer Biol Ther. (2012) 13:516–26. doi: 10.4161/cbt.19601
62. Athar M, Back JH, Tang X, Kim KH, Kopelovich L, Bickers DR, et al. Resveratrol: a review of preclinical studies for human cancer prevention. Toxicol Appl Pharmacol. (2007) 224:274–83. doi: 10.1016/j.taap.2006.12.025
63. Nicolini G, Rigolio R, Miloso M, Bertelli AA, Tredici G. Anti-apoptotic effect of trans-resveratrol on paclitaxel-induced apoptosis in the human neuroblastoma SH-SY5Y cell line. Neurosci Lett. (2001) 302:41–4. doi: 10.1016/S0304-3940(01)01654-8
64. Jazirehi AR, Bonavida B. Resveratrol modifies the expression of apoptotic regulatory proteins and sensitizes non-Hodgkin's lymphoma and multiple myeloma cell lines to paclitaxel-induced apoptosis. Mol Cancer Ther. (2004) 3:71–84.
65. Guan H, Singh NP, Singh UP, Nagarkatti PS, Nagarkatti M. Resveratrol prevents endothelial cells injury in high-dose interleukin-2 therapy against melanoma. PLoS ONE. (2012) 7:e35650. doi: 10.1371/journal.pone.0035650
66. Soto BL, Hank JA, Van De Voort TJ, Subramanian L, Polans AS, Rakhmilevich AL, et al. The anti-tumor effect of resveratrol alone or in combination with immunotherapy in a neuroblastoma model. Cancer Immunol Immunother. (2011) 60:731–8. doi: 10.1007/s00262-011-0971-0
67. Wu L, Zhu J, Prokop LJ, Murad MH. Pharmacologic therapy of diabetes and overall cancer risk and mortality: a meta-analysis of 265 studies. Sci Rep. (2015) 5:10147. doi: 10.1038/srep10147
68. Zhou Y, Zheng J, Li Y, Xu D-P, Li S, Chen Y-M, et al. Natural polyphenols for prevention and treatment of cancer. Nutrients. (2016) 8:515. doi: 10.3390/nu8080515
69. Afzal MZ, Mercado RR, Shirai K. Efficacy of metformin in combination with immune checkpoint inhibitors (anti-PD-1/anti-CTLA-4) in metastatic malignant melanoma. J Immunother Cancer. (2018) 6:64. doi: 10.1186/s40425-018-0375-1
70. Boocock DJ, Faust GE, Patel KR, Schinas AM, Brown VA, Ducharme MP, et al. Phase I dose escalation pharmacokinetic study in healthy volunteers of resveratrol, a potential cancer chemopreventive agent. Cancer Epidemiol Prev Biomarkers. (2007) 16:1246–52. doi: 10.1158/1055-9965.EPI-07-0022
71. Brown VA, Patel KR, Viskaduraki M, Crowell JA, Perloff M, Booth TD, et al. Repeat dose study of the cancer chemopreventive agent resveratrol in healthy volunteers: safety, pharmacokinetics, and effect on the insulin-like growth factor axis. Cancer Res. (2010) 70:9003–11. doi: 10.1158/0008-5472.CAN-10-2364
72. Chow HS, Garland LL, Hsu C-H, Vining DR, Chew WM, Miller JA, et al. Resveratrol modulates drug-and carcinogen-metabolizing enzymes in a healthy volunteer study. Cancer Prev Res. (2010) 3:1168–75. doi: 10.1158/1940-6207.CAPR-09-0155
73. Espinoza JL, Trung LQ, Inaoka PT, Yamada K, An DT, Mizuno S, et al. The repeated administration of resveratrol has measurable effects on circulating T-cell subsets in humans. Oxid Med Cell Longev. (2017) 2017:6781872. doi: 10.1155/2017/6781872
74. Patel KR, Brown VA, Jones DJ, Britton RG, Hemingway D, Miller AS, et al. Clinical pharmacology of resveratrol and its metabolites in colorectal cancer patients. Cancer Res. (2010) 70:7392–9. doi: 10.1158/0008-5472.CAN-10-2027
75. Howells LM, Berry DP, Elliott PJ, Jacobson EW, Hoffmann E, Hegarty B, et al. Phase I randomised double-blind pilot study of micronized resveratrol (SRT501) in patients with hepatic metastases-safety, pharmacokinetics and pharmacodynamics. Cancer Prev Res. (2011) 4:1419–25. doi: 10.1158/1940-6207.CAPR-11-0148
Keywords: immunotherapy, immune checkpoint blockade, tumor immunology, caloric restriction, calorie restriction mimetics, intermittent fasting, fasting-mimicking diet, time-restricted feeding
Citation: Turbitt WJ, Demark-Wahnefried W, Peterson CM and Norian LA (2019) Targeting Glucose Metabolism to Enhance Immunotherapy: Emerging Evidence on Intermittent Fasting and Calorie Restriction Mimetics. Front. Immunol. 10:1402. doi: 10.3389/fimmu.2019.01402
Received: 04 February 2019; Accepted: 03 June 2019;
Published: 25 June 2019.
Edited by:
Giovanna Schiavoni, Istituto Superiore di Sanità (ISS), ItalyReviewed by:
Rodabe N. Amaria, University of Texas MD Anderson Cancer Center, United StatesDavid Allen Reardon, Dana–Farber Cancer Institute, United States
Copyright © 2019 Turbitt, Demark-Wahnefried, Peterson and Norian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Courtney M. Peterson, Y3BldGVyc29AdWFiLmVkdQ==; Lyse A. Norian, bG5vcmlhbkB1YWIuZWR1
†These authors have contributed equally to this work