 Meriem Riani1
Meriem Riani1 Céline Muller1
Céline Muller1 Camille Bour1
Camille Bour1 Philippe Bernard1,2†
Philippe Bernard1,2† Frank Antonicelli1,2,3†
Frank Antonicelli1,2,3† Sébastien Le Jan1*†
Sébastien Le Jan1*†- 1Laboratory of Dermatology, EA7509 IRMAIC, University of Reims-Champagne-Ardenne, Reims, France
- 2Department of Dermatology, University Hospital, University of Reims-Champagne-Ardenne, Reims, France
- 3Department of Biological Sciences, Immunology, UFR Odontology, University of Reims-Champagne-Ardenne, Reims, France
Bullous pemphigoid (BP) is a cutaneous autoimmune disease, characterized by an inflammatory cascade leading to blister formation. Although macrophages were shown to participate in BP pathophysiology, their role in the blister formation process still needs to be investigated. We here addressed the influence of serum and blister fluid (BF) from patients with BP on the polarization status of macrophages with regards to the metalloproteinase-9 (MMP-9) expression. We demonstrated that several markers related to the alternatively activated macrophage phenotype (M2) including IL-10, TARC, arginase, TNFα, and IL-1RA were meaningfully increased in BF of patients with BP. We further showed that BF, but not serum from patients with BP, significantly induced the expression of CD163, CD206, and IL-10 in BP monocyte-derived macrophages (MDMs). Notably IL-10 was the only cytokine to be correlated to the reference clinical score, BP disease activity index (BPDAI), especially to the inflammatory BPDAI subscore evaluating urticarial and erythematous skin lesions (r = 0.57, p = 0.0004). We also found elevated levels of MMP-9 to M2-type macrophages ex vivo and highlighted the presence of CD163+ MMP-9+ macrophages histologically, at skin lesional site. Finally, we showed that methylprednisolone reduced MMP-9 levels in MDMs without modifying the other M2 markers. All together these results strongly support the presence of M2-phenotype macrophages with pro-inflammatory properties susceptible to favor blister formation in BP.
Introduction
Bullous pemphigoid (BP) is the most common skin autoimmune subepidermal blistering disease (1, 2). The disease typically presents in the elderly with a generalized pruriginous, erythematous and bullous eruption. Biologically, BP is characterized by the binding of autoantibodies directed against two components of the hemidesmosome, BP230 and BP180 that generates an inflammatory response critical for blister formation. A pathophysiological mechanism of blister formation was established, which typically involved a variety of cellular types including eosinophils, neutrophils, lymphocytes, mast cells, and macrophages (3, 4). Mast cell activation upon immune complex binding play a major role in neutrophils recruitment, while macrophages were supposed to rather amplify the neutrophil infiltration in a mast cell-dependent fashion (5, 6). However, the role of macrophage polarization has not been taken into consideration in the pathophysiological BP process, and especially regarding the production of MMP-9, a metalloproteinase widely involved in blister formation (7).
Macrophages display a remarkable plasticity and can change their physiology in response to environmental factors with at one extremity the inflammatory or classically activated macrophage M1 phenotype and at the other, the anti-inflammatory or alternatively activated macrophage M2 phenotype (8). The M1 phenotype is classically induced by microbial products or pro-inflammatory cytokines such as IFN-γ and TNFα, whereas molecules such as IL-4, IL-13, M-CSF, immune complexes, IL-10, and glucocorticoids favor the orientation toward the M2-type macrophage phenotype. In addition, M1-type macrophages have been shown to produce high levels of pro-inflammatory molecules such as TNFα, IL-1, IL-6, IL-23, IL-12, type-I IFN, reactive nitrogen intermediate (RNI), reactive oxygen intermediate (ROI), CXCL9, CXCL10, and CXCL11 (9), whereas M2 macrophages express IL-4, IL-10, CD163, and CD206 and promote tissue regeneration and repair (10, 11). Hence, a switch in macrophage phenotype could be of importance in the pathogenesis of autoimmune and inflammatory diseases (12–14).
In line with previous reports on macrophage polarization (15, 16), the presence of immune complexes at the skin lesional site in BP should theorically favor the presence of M2 type macrophages. However, we recently showed that BP was associated with the production of IL-23 and CXCL10, two markers of the M1 macrophage phenotype (17–19), thus further questioning on the status and the role of macrophages in the early, inflammatory process leading to blister formation in BP.
We here aimed at understanding the influence of serum and blister fluids (BF) of patients with BP on macrophage polarization by analyzing the expression of several M1/M2 markers as well as the expression of MMP-9. As corticosteroids, the main treatment of BP, favor M2-macrophage polarization, we also investigated the effect of methylprednisolone on the expression of these macrophage markers and MMP-9.
Methods
Patients and Study Design
This ancillary study is part of a main prospective, single-center study that was conducted in the Department of Dermatology at Reims University Hospital (French Referral Center for Autoimmune Bullous Diseases) between September 2013 and July 2017. Consecutive patients with newly diagnosed BP were included using the following criteria: clinical features typical of BP with presence of at least three out of four well-established criteria by Vaillant et al. (20); subepidermal blister on skin biopsy; and deposits of IgG and/or C3 in a linear pattern along the epidermal basement membrane zone by direct IF. Sera and BFs from patients were collected at time of diagnosis (V1). Serum samples from age- and sex-matched patients without inflammatory and autoimmune diseases admitted to the trauma department of the Reims University Hospital were used as controls. The investigation was conducted under the approval of the Ethic Committee of the University Hospital of Reims (CNIL authorization DR-2013-320), and all of the subjects gave their informed and written consent before participating in the study in accordance with the Helsinki Declaration.
Cell Preparation
THP-1 monocytic cell line, kindly provided by Dr. S. Hart from University of Edinburgh, UK, was cultured in RPMI 1640 medium (Life Technologies), supplemented with 10% fetal bovine serum (FBS), 2 mM glutamine, 25 U/ml penicillin, and 25 U/ml streptomycin at 37°C in a humidified 5% CO2 incubator. THP-1-derived macrophages were generated as described previously (21). Briefly THP-1 cells were seeded at a concentration of 2 × 106 cells per milliliter in FBS-free medium and treated with 320 nM PMA for 24 h. To generate M1-polarized THP-1 macrophages, THP-1 cells were treated with 320 nM PMA for 6 h and then cultured with PMA plus 20 ng/ml IFN-γ and 20 ng/ml TNFα for 18 h. To generate M2-polarized THP-1 macrophages, THP-1 cells were treated with 320 nM PMA for 6 h, and then cultured with PMA plus 20 ng/ml IL10 for another 18 h. Cells and culture media were then harvested and analyzed for gene expression and MMP-9 secretion, respectively. For stimulation, THP-1 cells were treated with 320 nM PMA in FBS-free medium for 6 h and then cultured for 18 h with 320 nM PMA plus 10% BF or 10% serum from control subjects (CTR serum) or BP patients (BP serum). When mentioned methylprednisolone (Sigma) was introduced with serum at a concentration of 10 μM. Cells were then harvested and analyzed for gene expression.
Peripheral blood mononuclear cells (PBMCs) from control subjects or patients with BP were obtained by density-gradient centrifugation from EDTA-treated whole blood using a density gradient medium (Granulosep, Eurobio-Abcys). Monocytes were purified from PBMCs by positive selection using CD14 immunomagnetic beads (MACS; Miltenyi Biotech) according to manufacturer instructions. To generate monocyte-derived macrophages (MDMs), freshly isolated monocytes were cultured at a concentration of 2 × 106 cells per milliliter for 7 days in RPMI 1640 medium (Life Technologies), 2 mM glutamine, 25 U/ml penicillin, 25 U/ml streptomycin and 10% serum from control subjects or BP patients. Culture media was replaced at day 4. When stimulated, MDM were treated for 24 h with 10% BF simultaneously or not with methylprednisolone. Cells were then harvested and analyzed for gene expression.
Gene Expression Analysis
Total RNA from isolated and cultured cells was extracted using TRI-Reagent (Euromedex). Reverse transcription was performed from 1 μg of total RNA using Maxima First Strand cDNA kit with dsDNAse (Life Technologies) according to manufacturer's instructions. cxcl10, il-23, inos, tnfα, CD163, CD206, il-10, il-1ra, and mmp9 gene expressions were analyzed by quantitative real-time PCR using Platinum SYBR Green qPCR SuperMix-UDG kit (Invitrogen) on the LightCycler system (Roche Diagnostics). Relative quantification was performed using the GAPDH as a reference gene.
In situ Protein Expression Analysis
Perilesional skin biopsy specimens performed in patients with BP before introduction of any corticosteroid treatment were obtained from the Department of Anatomo-Pathology at Reims Hospital. Tissues were fixed in paraformaldehyde, embedded in paraffin, and sectioned. To visualize M2 type macrophages, immunostaining with a mouse monoclonal anti-human CD163 antibody (Clone MRQ26, Roche Diagnostics, Meylan, France) was performed by using routine methods with a biotinylated anti-mouse secondary antibody (BA-2000; Vector Laboratories, Burlingame, USA) and the ABC-peroxidase complex (Vector Laboratories) with diaminobenzidine-H2O2 used as the chromogen for detection. In order to identify cells positive for both CD163 and MMP-9, simultaneous staining was performed with mouse monoclonal anti-human CD163 antibody (Clone MRQ26, Roche Diagnostics, Meylan, France) and rabbit anti-human MMP-9 [MMP-9 (E11), Santa Cruz Biotechnologies] and followed by incubation with matched secondary antibodies: chicken anti-goat IgG Alexa Fluor 488 and chicken anti-mouse IgG Alexa Fluor 594, respectively (Invitrogen). Images were captured with confocal microscope (LSM 710; Zeiss) using Zen software.
Determination of Cytokine Levels
M1 type (TNF-α, IFN-γ) and M2 type (arginase, TARC, IL-10, IL-1RA) macrophage marker concentrations in sera and BFs were determined using the bead-based immunoassays LEGENDplex Human Macrophage/Microglia Panel and Human Th Cytokines Panel (BioLegend, San Diego, CA, USA). The assays were performed in 96-well plates following the manufacturer's instructions. For measurements a LSR Fortessa flow cytometer (BD Biosciences) was employed, and data were evaluated with the LEGENDplex™ Data Analysis software.
Zymography
The quantity of MMP-9 was determined on conditioned media by the zymography technique. Briefly, samples were electrophoresed on a 10% SDS polyacrylamide gel impregnated with 0.1% of gelatin as substrate under non-reducing conditions (22). The gels were subsequently fixed and stained with 0.25% Coomassie brilliant blue R-250 to visualize the proteolytic activity bands. Quantification was performed using the ImageJ software (National Institutes of Health, Bethesda, MD).
Statistical Analysis
Descriptive statistics such as means and SEMs were conducted for all quantitative measures. The distribution of the variables was assessed using D'Agostino and Pearson omnibus normality test. As population could not be assumed to be normal and some of the groups examined were small, we used non-parametric testing to compare populations in this study. Comparisons between two groups were performed using the exact Wilcoxon signed-rank test for paired data and the Mann-Whitney test for unpaired data. Correlations were performed using non-parametric Spearman's correlation test. The results were considered significant if p-values were 0.05 or less.
Results
BF Is Associated With a M2 Macrophage Cytokine Profile in BP
First, we measured the concentration of several markers associated with macrophage polarization both in sera and BFs collected from patients with BP at time of diagnosis and before treatment and in control sera (Figure 1). The concentrations of TNF-α but not IFN-γ, both used as M1 type macrophage markers (Figure 1A), and of IL-10, IL-1RA, arginase used as M2 type macrophage markers (Figure 1B) showed significant increase in BFs compared with their concentrations in both control and BP patient sera. Concentrations of TARC, another M2 marker, were significantly increased in BF compared with control sera but not with BP sera. Notably, no variation was evidenced in the serum of BP patients with respect to control sera (Figures 1A,B).
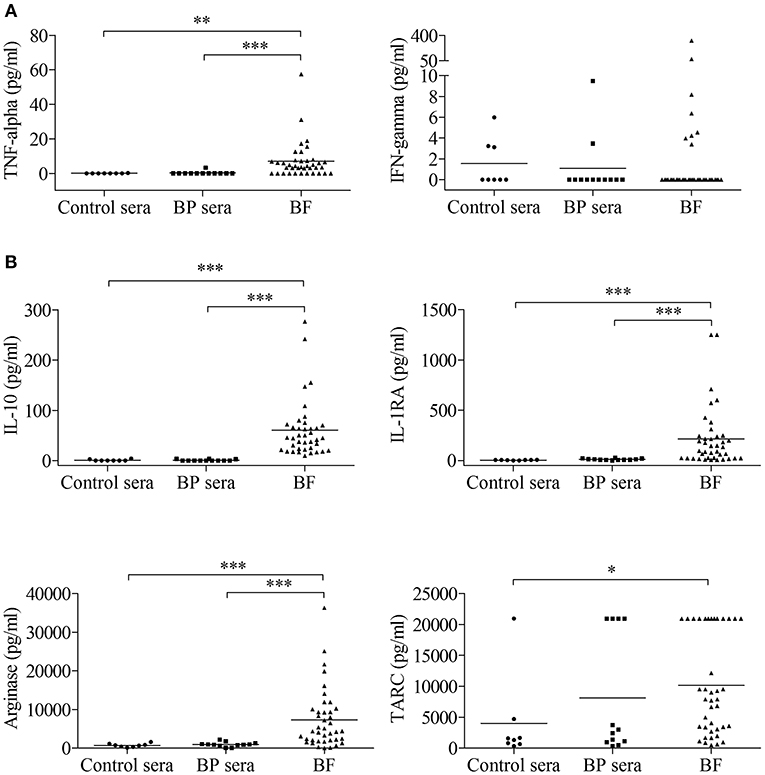
Figure 1. Detection of macrophage-associated cytokines in serum and blister fluid (BF) of BP patients. The concentrations of a panel of M1-type (A) and M2-type (B) macrophage markers were measured in serum from control subjects (Control sera, n = 8) and in serum (BP sera, n = 12) and BF (n = 40) from patients with BP at time of diagnosis using the bead-based immunoassays LEGENDplex Human Macrophage/Microglia Panel and Human Th Cytokines Panel. Non-parametric unpaired Mann–Whitney's test was used to compare populations (*p < 0.05; **p < 0.01; ***p < 0.001).
BF Rather Favored M2 Type Than M1 Type Macrophage Phenotype
To investigate whether those expressions were related to macrophages, we next tested the effects of BF from BP patients on the expression of M1 and M2 macrophage markers by the macrophages derived from PMA-induced THP-1 monocytic cells (Figure 2). Among the M1-associated markers, a significant increase in iNOS expression was observed upon BF stimulation (p < 0.05), whereas a decrease for IL-23 mRNA expression (p < 0.01) and no variation in CXCL10 and TNFα mRNA expression were evidenced (Figure 2A). Analysis of the M2-associated markers showed that BF stimulation induced an increase in CD163, CD206, and IL-10 (p < 0.01, p < 0.01, and p < 0.05, respectively), but a decrease in IL-1RA expression (p < 0.001), suggesting that BF rather favored M2 type than M1 type macrophage phenotype (Figure 2B). We also used the PMA-induced THP-1 macrophages to investigate whether BP serum could favor either an M1 or an M2 phenotype (Figures 3A,B). Compared with control serum, short term stimulation with BP serum of PMA-induced THP-1 macrophages increased CXCL10 and TNFα expression (p < 0.01 and p < 0.05, respectively) and decreased the expression of IL-23, IL-1RA, and CD206 (p < 0.05; Figure 3A). Such variations were not observed when macrophages were differentiated from monocytes (MdM) isolated from control or BP patients through a longer period (7 days) in presence of serum (Figures 3C,D). Based on these results, we next investigated whether the differentiation of BP monocyte into macrophage upon serum stimulation affected the effects of BF on the expression of macrophage markers (Figure 4). Stimulation of BP-MdM with BF significantly increased TNFα, CD163, CD206, and IL-10 (p < 0.05, p < 0.01, p < 0.01, and p < 0.01, respectively) but not CXCL10 expression (Figures 4A,B).
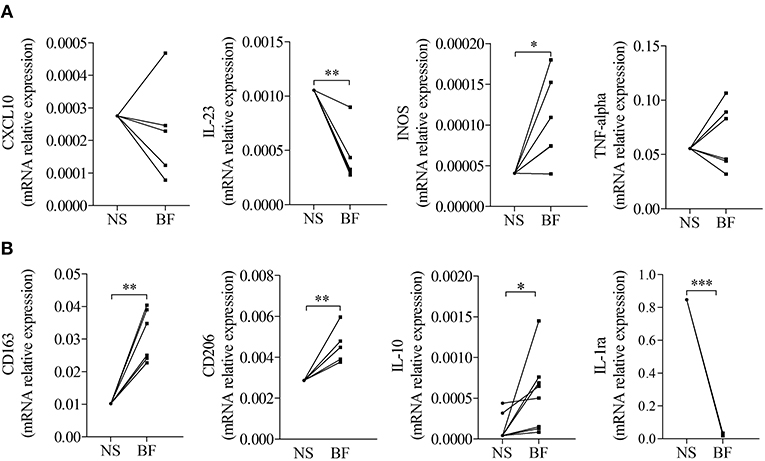
Figure 2. Blister fluid (BF) from patients with BP affected the expression of M1/M2-type macrophage markers in THP-1 derived macrophages. M1-type (A) and M2-type (B) macrophage marker expressions were analyzed by real-time qPCR in THP-1 derived macrophage unstimulated (NS) or stimulated with BF (BF) from patients with BP. A paired T-Test was performed for statistics (*p < 0.05; **p < 0.01; ***p < 0.001).
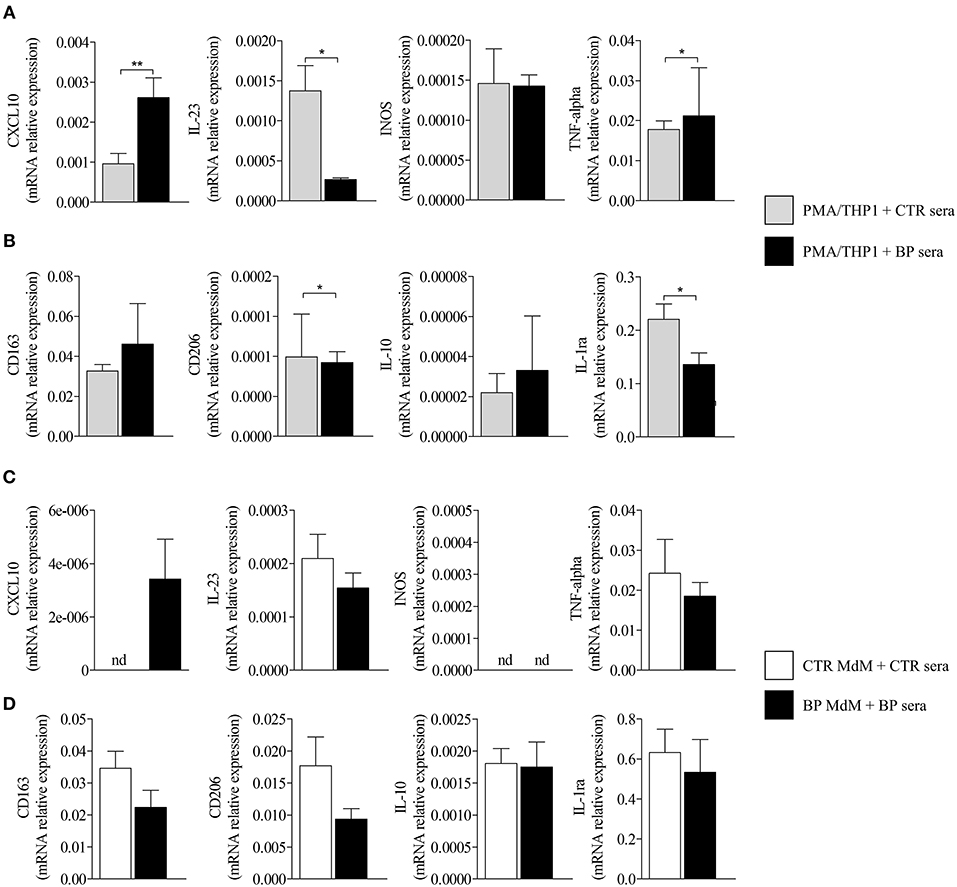
Figure 3. Regulation of M1-type and M2-type macrophage marker expression in THP-1- and monocyte-derived macrophages. M1-type (A,C) and M2-type (B,D) macrophage marker expressions were analyzed by real-time qPCR in CTR or BP serum-stimulated macrophages originated either from PMA-derived THP-1 cells (A,B) or from CTR or BP monocytes differentiated with their autologous serum for 7 days (C,D). The error bars denote the mean ± SEM. Non-parametric unpaired Mann–Whitney's test was used to compare populations (*p < 0.05; **p < 0.01; nd, not detected).
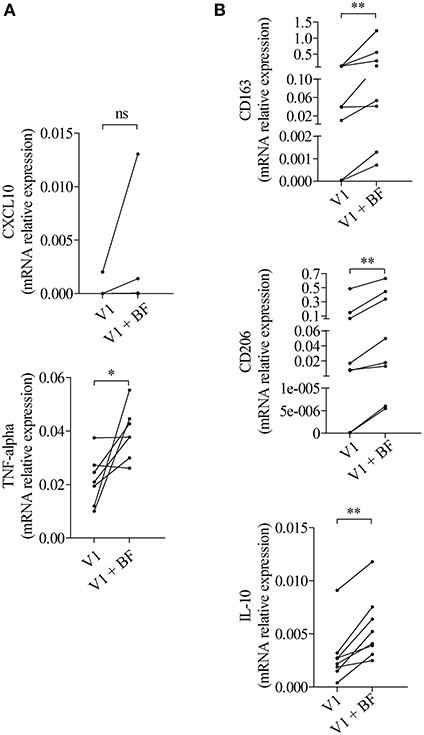
Figure 4. BF essentially increased the expression of M2-type macrophage markers in BP monocyte-derived macrophages. M1-type (A) and M2-type (B) macrophage marker expressions were analyzed by real-time qPCR in macrophages issued from BP monocytes differentiated for 7 days with the autologous serum collected at diagnosis (V1) and stimulated with BF for 24 h (V1 + BF). Non-parametric paired Wilcoxon's test was used to compare populations (ns, not significant; *p < 0.05; **p < 0.01).
M2 Type Macrophage Profile Was Associated With Disease Activity in BP
Within the BF, IL-10 concentrations were correlated to those of arginase (Figure 5A, r = 0.41, p = 0.0086). Also, IL-10 was the only cytokine among all cytokines tested in Figure 1 to be related to the clinical activity score. Indeed IL-10 concentrations in the BF were correlated with disease activity measured by the total BPDAI score (Figure 5B, r = 0.45, p = 0.006), and especially with the inflammatory BPDAI subscore associated with urticarial and erythematous plaques (Figure 5C, r = 0.57, p = 0.0004), but not with the subscore bound to skin lesions (Figure 5D, r = 0.21, p = 0.23).
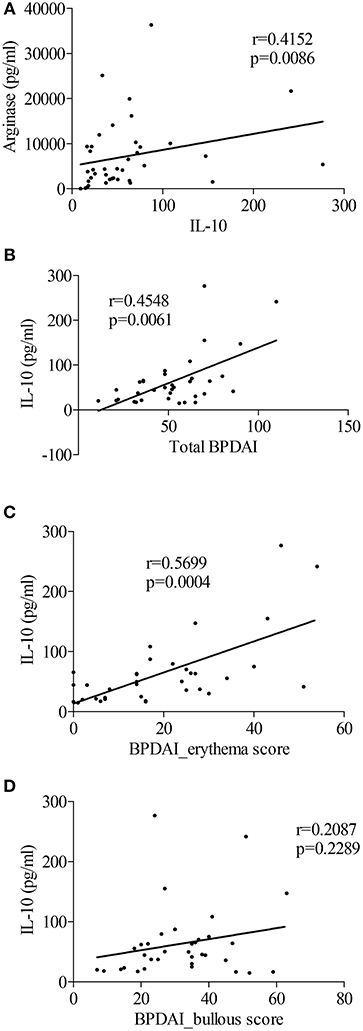
Figure 5. The M2-type macrophage marker IL-10 was related to BP disease activity. (A) Correlation between levels of IL-10 and the M2-type macrophage marker Arginase in blister fluid of patients with BP at diagnosis was analyzed using the nonparametric Spearman's correlation test. (B–D) Correlations between total BPDAI score (B), blister/erosion (C), and erythema (D) BPDAI subscores and IL-10 levels measured in BF were analyzed using the non-parametric Spearman's correlation test (n = 35).
BF Increased the Production of MMP-9 by M2 Type Macrophages in BP
To investigate how a M2 type macrophage profile could be associated with disease activity, we analyzed the influence of macrophage polarization on their capacity to produce the protease MMP-9 (Figure 6). To that purpose, PMA-induced THP-1 macrophages were further polarized into M1 and M2 type macrophages with classical stimuli such as IFN-γ and IL-10, respectively. M1 polarization resulted in significant decrease of MMP-9 expression and secretion, whereas this protease was still strongly produced by M2-differentiated THP-1 macrophages (Figures 6A,B). Stimulation with serum and BF also resulted in sustained MMP-9 expression by THP-1 differentiated macrophages (Figure 6C). No differences were noticed between BP serum and control serum on MMP-9 expression both in differentiated THP-1 and in MdM (Figures 6C,D). However, stimulation with BF further increased MMP-9 expression in MdM originated from patients with BP (Figure 6E, p < 0.05). Immunohistochemistry studies of skin biopsy specimens of BP revealed a substantial number of CD163+ macrophages at lesional site, with double-positive cells CD163+ MMP-9+ in the blister cavity (Figure 6F).
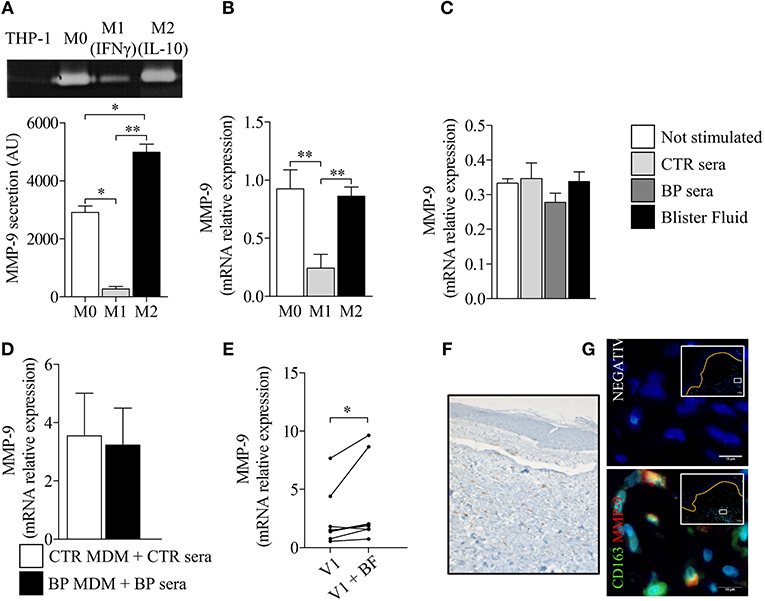
Figure 6. Polarization of BP-MdM influenced their capacity to produce the protease MMP-9. MMP-9 secretion (A) and MMP-9 mRNA expression (B) were analyzed, respectively, by gelatin zymography and real-time qPCR in M0, M1 (+IFNγ), and M2 (+IL-10) THP-1 derived macrophages. The error bars denote the mean ± SEM. Paired Student's T-Test was used for statistics (n = 3; *p < 0.05; **p < 0.01). (C–E) MMP-9 mRNA expression was analyzed by real-time qPCR in both THP-derived (C) and BP monocyte-derived macrophages (D,E) stimulated with either control serum, BP serum or blister fluid from BP patient. The error bars denote the mean ± SEM. Nonparametric unpaired Mann-Whitney's test was used for statistical analysis of (C,D) (n = 8 and n = 5, respectively). Nonparametric paired Wilcoxon's test was used to statically analyze (E) (n = 8; *p < 0.05). (F,G) Punch biopsies of lesional skin from patients with BP were subjected to CD163 immuno-staining alone (F) or double immunofluorescence staining for MMP-9 (red) and CD163 (green) (G) with Hoescht counterstain (blue). Negative CTR, negative control where primary antibodies were not added. White islets show lower magnification images of blister area and orange line designed the dermo-epidermal junction. The boxes in white islets match to the area where the high magnification image has been taken.
Corticosteroids Inhibited the Capacity of M2-macrophages to Produce MMP-9
Finally, treatment with methylprednisolone reduced MMP-9 expression from BP-derived MdM, while enhancing the expression of both M2 type makers CD163 and CD206 (Figure 7), therefore restoring their tissue regeneration and repair profile.
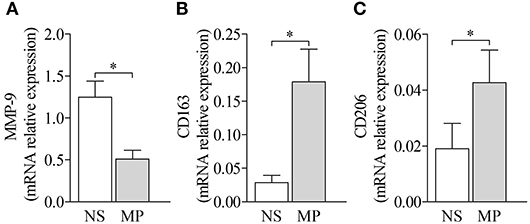
Figure 7. Treatment by methylprednisolone decreased MMP-9 expression and favored M2-type macrophage markers. MMP-9 (A) and M2-type macrophage markers CD163 (B) and CD206 (C) mRNA expressions were analyzed by real-time qPCR in BP MdM treated (MP) or not (NS) with 10 μM methylprednisolone. The error bars denote the mean ± SEM. Nonparametric paired Wilcoxon's test was used (*p < 0.05).
Discussion
In this study, we evidenced that macrophage polarization in BP is modified when cells encounter factors present in the BF. Surprisingly, at the site of lesion, macrophages displayed an alternatively activated (M2)-like phenotype and also a high expression level of MMP-9, therefore questioning on the role of macrophages in BP pathophysiology.
Polarization toward M2-type macrophages was dictated at site of lesion in BP. Indeed BFs alone or following serum-induced macrophage differentiation, enhanced the expression of CD163 and CD206 in THP-1-derived macrophages and BP-MdM, respectively, supporting a recent study showing in situ the presence of M2-type macrophages in the area of BP skin lesion (6). This is also in line with a polarization toward the M2-phenotype induced by immune complexes and complement components present in the BFs of BP patients (15, 16, 23). It has to be noticed that serum has been shown to influence monocyte maturation and differentiation toward macrophage (24, 25). In line with our recent publication showing that CXCL10 was increased in the blood and in the skin of patients with BP (19), we here demonstrated that serum from BP patients increased CXCL10 expression in THP-1-derived macrophages. However, the increase in CXCL10, but not of the other polarization markers with BP patient serum vs. control serum rather suggests that serum initiates macrophage activation than polarization. Recent works established that THP-1 cells can respond to the polarization protocols used for primary macrophages, and can either match or diverge from what is seen in human peripheral-blood MdMs, then clarifying the discrepancies observed in our study (26, 27). In our study, such differences may be linked to the time of stimulation. Indeed, the THP-1 derived macrophages present the advantages to investigate serum acute effects, whereas patients derived macrophages rather show the steady state reached after long term stimulation. Thus, altogether, our results suggest that in BP macrophage polarization arise in two steps. First macrophages differentiate from monocytes. In BP, those macrophages rather display an activated state than a polarization profile. Then, macrophage M2 polarization occurs when reaching the skin lesional site.
In the initial phase of inflammatory diseases, macrophages are frequently associated with the classically activated M1 phenotype, whereas M2 macrophages are generated in the later phase and are associated with the resolution of inflammation and with the repair process (28). However, in asthma and in allergy, excessive M2 macrophages may increase Th2 cell recruitment, eosinophils infiltration and mucus secretion, and result in airway hyper-responsiveness (13, 29). Our results are of interest as they show that during the active phase of the BP, molecules within the BF orientate macrophages toward an anti-inflammatory M2-like phenotype, which may later influence the skin regeneration process characterized by an absence of scarring in BP. However, this also questioned on the role of macrophages in blister formation. Indeed, it had been previously shown through a mouse BP model that macrophages rather amplified the inflammatory response and blister formation by enhancing neutrophil infiltration in a mast cell-dependent fashion (5).
To conciliate these opposed aspects of the M2-macrophages in BP, we also evaluated the inflammatory status of such polarized macrophages. The pro-inflammatory cytokine TNF-α and IL-10, initially described as immuno-modulatory, were increased in BF-stimulated BP MdMs, in accordance with previous works that showed higher concentrations of both cytokines in BF (30–32). Notably IL-10 displays both pro-and anti-inflammatory effects during immune response, and especially, IL-10 enhances IgE-mediated mast cell responses (33), a pathway which fits with a local pruritic type-2 orientated inflammatory reaction in BP (34). Accordingly we showed that IL-10 concentration was correlated with the activity of the disease, specifically with the inflammatory BPDAI score. In BP, IL-10 may be produced by several cell type of which Th-2 or B-regulatory cells for instances. Then, IL-10 could favor M2-macrophage polarization, and therefore lead to further IL-10 production by M2-macrophages at the skin lesional site. In contrast, IL-23 expression was not induced in BF-stimulated BP MdMs suggesting that the increased level of IL-23 previously observed in the BFs of BP patients (17) cannot be attributed to macrophages. All together these results indicate that M2-type macrophages in BP could increase inflammatory cell recruitment and could participate to the inflammatory cascade observed in BP.
Besides cytokines, elevated levels both of secretion and expression of the metalloproteinase MMP-9 were also associated with M2-type macrophages in BP. Of note, this high level of expression was consecutive to the stimulation with autologous BP serum, and further enhanced in presence of BFs. Noteworthy, by mean of the THP-1 cell line, we reproduced results from previous studies that established a positive correlation between macrophage-released MMP-9 and M2-type polarization (35–37). Indeed, we showed here that macrophage differentiation is already associated with high level of MMP-9, and that M2-macrophage polarization may further enhance MMP-9 production, in contrast to M1 polarization. Therefore, according to the level of MMP-9 observed in BF-stimulated BP MdMs and in setting with the expression of the cytokines described above, we confirm that macrophages rather display a M2-type than a pro-inflammatory M1-type phenotype in BP. Furthermore, such M2-polarization may accentuate the MMP-9 local production initiated by other immune infiltrated cells, and subsequently amplify the local inflammatory process leading to blister formation.
Although the role of macrophages still needs further investigation, our study raises the question of both M2 macrophage polarization and of the associated increase in MMP-9 in the pathophysiological process of BP. Indeed, the protease MMP-9 has been shown as a key molecule for blister formation and any increase in its production is thought to feed the auto-inflammatory and auto-immune processes thereof (7). In contrast, MMP-9 inhibition in patients with BP treated by corticosteroids over a period of 6–9 months, and occasionally even longer, suggests that MMP-9 inhibition does not alter the scarless regeneration process in BP. Furthermore, we here demonstrated that inhibition of MMP-9 production by methylprednisolone was not associated with a switch toward a M1 macrophage phenotype, but instead reinforced their M2-type phenotype promoting tissue regeneration and repair (10, 11) (Figure 8). Then, in the initial inflammatory phase following auto-antibody binding, macrophage recruitment may interact with mast cell to further activate the inflammatory process as previously demonstrated (5). Meanwhile, those macrophages release MMP-9, therefore further increasing the local protease load. Such MMP-9 production may be further elicited by IL-10 produced by other infiltrated and resident immune cells gathered at the skin lesional site, therefore leading to an auto-amplification loop and to disease extent. Under treatment, such pro-inflammatory cell cooperation is inhibited reducing the pro-inflammatory cytokine and proteases release, and subsequently disease activity.
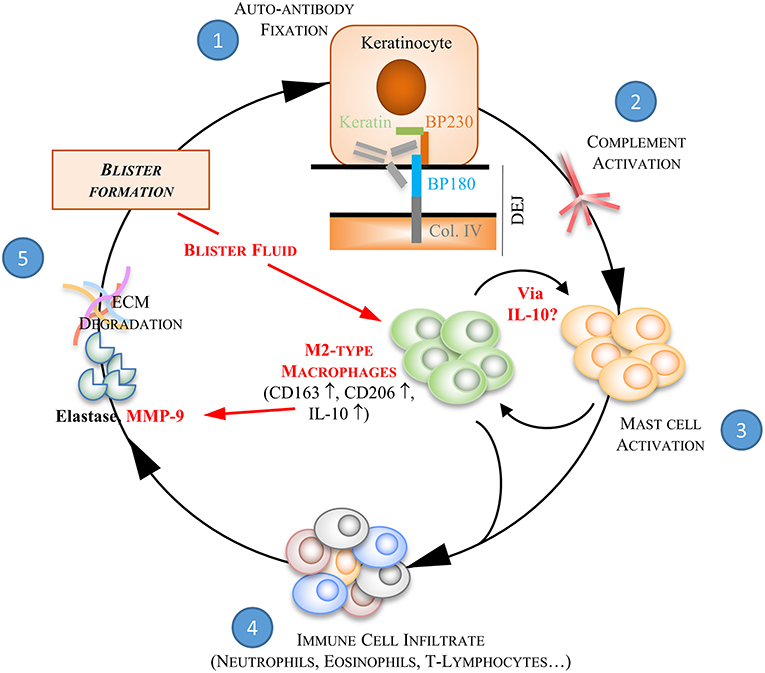
Figure 8. BF-induced M2-type macrophages in the auto-inflammatory response associated to BP. Bullous pemphigoid is characterized by the production of autoantibodies against two proteins of the hemidesmosome structure, BP180 and BP230. The binding of BP autoantibodies ① onto their target initiates the activation of the complement pathway ② which then provokes the release of chemokines and cytokines by mast cells ③. These pro-inflammatory molecules trigger subsequent recruitment of inflammatory cells at the dermal-epidermal junction ④. Inflammatory key mediators secreted by these latter cells induce an overexpression of proteases, such as the matrix-metalloproteinases-9 (MMP9) and the neutrophil elastase, which are involved in extracellular matrix (ECM) degradation and blister formation ⑤. We here further demonstrated the ability of blister fluid to induce M2-type macrophages at site of skin lesion by increasing CD163, CD206, and IL-10 expression. BF-induced M2-type macrophages also show pro-inflammatory function by releasing MMP-9 and then participating to pathophysiological process of BP. Under corticotherapy the auto-amplification process associated to pro-inflammatory cell interaction is dismantled. Indeed, steps ③, ④, and ⑤ are inhibited. In addition, we showed that inhibition of MMP-9 production by methylprednisolone was not associated with a switch toward a M1 macrophage phenotype, but instead reinforced their M2-type phenotype promoting tissue regeneration and repair.
Controlling the amount of MMP-9 locally produced during the active phase of BP disease is clinically of prime importance as MMP-9 along with human elastase are the two main proteases involved in blister formation. In the blood circulation of BP patients, monocytes produce a large amount of MMP-9 (19), which may facilitate their migration into the skin. MMP-9 is secreted in a precursor form (proMMP-9), which must be converted to an active form to exhibit its pathological activity in local lesions. Thus, cytokines such as IL-10 and TNFα may act as a feed forward mechanism creating an auto-amplification loop favoring both M2 polarization and MMP-9 secretion. Other cytokines such as IL-17, CXCL10 released at the site of lesion may further participate to MMP-9 production and activation (17, 19). However, although the use of biologics is of major interest in treatment of autoimmune and auto-inflammatory diseases nowadays, targeting one cytokine or another requires a full understanding of the pathology. Indeed, several case reports highlighted the occurrence of autoimmune bullous skin diseases under anti-TNF therapy (38–42). Based on our results, further preclinical studies are still required to determine whether IL-10 or anti-IL-10 could demonstrate sufficient anti-inflammatory activity in the treatment of BP to warrant further study in a clinical trial.
Data Availability
All datasets generated for this study are included in the manuscript/supplementary files.
Author Contributions
PB, FA, and SL designed the study and wrote the manuscript. MR, CM, CB, and SL performed experiments. All authors critically evaluated the data and approved the final version for publication and analyzed clinical and biological data.
Funding
This work was supported by the French Department of Health's Projet Hospitalier de Recherche Clinique (PHRC) Interregional 2012. The funding source had no role in the design and conduct of the study, collection, management, analysis, and interpretation of data.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Maizières M., Ph.D., for technical support during the study and the Plateforme PICT IBiSA Imagerie Cellulaire et Tissulaire for providing microscopy equipment.
References
1. Liu Z, Diaz LA. Bullous pemphigoid: end of the century overview. J Dermatol. (2001) 28:647–50. doi: 10.1111/j.1346-8138.2001.tb00055.x
2. Bernard P, Antonicelli F. Bullous pemphigoid: a review of its diagnosis, associations and treatment. Am J Clin Dermatol. (2017) 18:513–28. doi: 10.1007/s40257-017-0264-2
3. Liu Z, Diaz LA, Troy JL, Taylor AF, Emery DJ, Fairley JA, et al. A passive transfer model of the organ-specific autoimmune disease, bullous pemphigoid, using antibodies generated against the hemidesmosomal antigen, BP180. J Clin Invest. (1993) 92:2480–8. doi: 10.1172/JCI116856
4. Hirose M, Recke A, Beckmann T, Shimizu A, Ishiko A, Bieber K, et al. Repetitive immunization breaks tolerance to type XVII collagen and leads to bullous pemphigoid in mice. J Immunol. (2011) 187:1176–83. doi: 10.4049/jimmunol.1100596
5. Chen R, Fairley JA, Zhao M-L, Giudice GJ, Zillikens D, Diaz LA, et al. Macrophages, but not T and B lymphocytes, are critical for subepidermal blister formation in experimental bullous pemphigoid: macrophage-mediated neutrophil infiltration depends on mast cell activation. J Immunol. (2002) 169:3987–92. doi: 10.4049/jimmunol.169.7.3987
6. Furudate S, Fujimura T, Kambayashi Y, Kakizaki A, Aiba S. Comparison of CD163+ CD206+ M2 macrophages in the lesional skin of bullous pemphigoid and pemphigus vulgaris: the possible pathogenesis of bullous pemphigoid. Dermatology (Basel). (2014) 229:369–78. doi: 10.1159/000365946
7. Ståhle-Bäckdahl M, Inoue M, Guidice GJ, Parks WC. 92-kD gelatinase is produced by eosinophils at the site of blister formation in bullous pemphigoid and cleaves the extracellular domain of recombinant 180-kD bullous pemphigoid autoantigen. J Clin Invest. (1994) 93:2022–30. doi: 10.1172/JCI117196
8. Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. (2008) 8:958–69. doi: 10.1038/nri2448
9. Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. (2004) 25:677–86. doi: 10.1016/j.it.2004.09.015
10. Martinez FO, Helming L, Gordon S. Alternative activation of macrophages: an immunologic functional perspective. Annu Rev Immunol. (2009) 27:451–83. doi: 10.1146/annurev.immunol.021908.132532
11. Gordon S. Alternative activation of macrophages. Nat Rev Immunol. (2003) 3:23–35. doi: 10.1038/nri978
12. Cassetta L, Cassol E, Poli G. Macrophage polarization in health and disease. Sci World J. (2011) 11:2391–402. doi: 10.1100/2011/213962
13. Liu Y-C, Zou X-B, Chai Y-F, Yao Y-M. Macrophage polarization in inflammatory diseases. Int J Biol Sci. (2014) 10:520–9. doi: 10.7150/ijbs.8879
14. Udalova IA, Mantovani A, Feldmann M. Macrophage heterogeneity in the context of rheumatoid arthritis. Nat Rev Rheumatol. (2016) 12:472–85. doi: 10.1038/nrrheum.2016.91
15. Edwards JP, Zhang X, Frauwirth KA, Mosser DM. Biochemical and functional characterization of three activated macrophage populations. J Leukoc Biol. (2006) 80:1298–307. doi: 10.1189/jlb.0406249
16. Gerber JS, Mosser DM. Reversing lipopolysaccharide toxicity by ligating the macrophage Fc gamma receptors. J Immunol. (2001) 166:6861–8. doi: 10.4049/jimmunol.166.11.6861
17. Le Jan S, Plée J, Vallerand D, Dupont A, Delanez E, Durlach A, et al. Innate immune cell-produced IL-17 sustains inflammation in bullous pemphigoid. J Invest Dermatol. (2014) 134:2908–17. doi: 10.1038/jid.2014.263
18. Plée J, Le Jan S, Giustiniani J, Barbe C, Joly P, Bedane C, et al. Integrating longitudinal serum IL-17 and IL-23 follow-up, along with autoantibodies variation, contributes to predict bullous pemphigoid outcome. Sci Rep. (2015) 5:18001. doi: 10.1038/srep18001
19. Riani M, Le Jan S, Plée J, Durlach A, Le Naour R, Haegeman G, et al. Bullous pemphigoid outcome is associated with CXCL10-induced matrix metalloproteinase 9 secretion from monocytes and neutrophils but not lymphocytes. J Allergy Clin Immunol. (2017) 139:863–72.e3. doi: 10.1016/j.jaci.2016.08.012
20. Vaillant L, Bernard P, Joly P, Prost C, Labeille B, Bedane C, et al. Evaluation of clinical criteria for diagnosis of bullous pemphigoid. French Bullous Study Group. Arch Dermatol. (1998) 134:1075–80. doi: 10.1001/archderm.134.9.1075
21. Tjiu J-W, Chen J-S, Shun C-T, Lin S-J, Liao Y-H, Chu C-Y, et al. Tumor-associated macrophage-induced invasion and angiogenesis of human basal cell carcinoma cells by cyclooxygenase-2 induction. J Invest Dermatol. (2009) 129:1016–25. doi: 10.1038/jid.2008.310
22. Emonard H, Hornebeck W. Binding of 92 kDa and 72 kDa progelatinases to insoluble elastin modulates their proteolytic activation. Biol Chem. (1997) 378:265–71. doi: 10.1515/bchm.1997.378.3-4.265
23. Qiao P, Dang E-L, Fang H, Zhang J-Y, Li B, Shen S-X, et al. Decreased expression levels of complement regulator CD55 contribute to the development of bullous pemphigoid. Oncotarget. (2018) 9:35517–27. doi: 10.18632/oncotarget.21216
24. Musson RA. Human serum induces maturation of human monocytes in vitro. Changes in cytolytic activity, intracellular lysosomal enzymes, and nonspecific esterase activity. Am J Pathol. (1983) 111:331–40.
25. Berer A, Stöckl J, Majdic O, Wagner T, Kollars M, Lechner K, et al. 1,25-Dihydroxyvitamin D(3) inhibits dendritic cell differentiation and maturation in vitro. Exp Hematol. (2000) 28:575–83. doi: 10.1016/S0301-472X(00)00143-0
26. Tedesco S, De Majo F, Kim J, Trenti A, Trevisi L, Fadini GP, et al. Convenience versus biological significance: are PMA-differentiated THP-1 cells a reliable substitute for blood-derived macrophages when studying in vitro polarization? Front Pharmacol. (2018) 9:71. doi: 10.3389/fphar.2018.00071
27. Shiratori H, Feinweber C, Luckhardt S, Linke B, Resch E, Geisslinger G, et al. THP-1 and human peripheral blood mononuclear cell-derived macrophages differ in their capacity to polarize in vitro. Mol Immunol. (2017) 88:58–68. doi: 10.1016/j.molimm.2017.05.027
28. Wang N, Liang H, Zen K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front Immunol. (2014) 5:614. doi: 10.3389/fimmu.2014.00614
29. Jiang Z, Zhu L. Update on the role of alternatively activated macrophages in asthma. J Asthma Allergy. (2016) 9:101–7. doi: 10.2147/JAA.S104508
30. Giacalone B, D'Auria L, Bonifati C, Ferraro C, Riccardi E, Mussi A, et al. Decreased interleukin-7 and transforming growth factor-beta1 levels in blister fluids as compared to the respective serum levels in patients with bullous pemphigoid. Opposite behavior of TNF-alpha, interleukin-4 and interleukin-10. Exp Dermatol. (1998) 7:157–61. doi: 10.1111/j.1600-0625.1998.tb00317.x
31. Ameglio F, D'Auria L, Bonifati C, Ferraro C, Mastroianni A, Giacalone B. Cytokine pattern in blister fluid and serum of patients with bullous pemphigoid: relationships with disease intensity. Br J Dermatol. (1998) 138:611–4. doi: 10.1046/j.1365-2133.1998.02169.x
32. Schmidt E, Bastian B, Dummer R, Tony H-P, Bröcker E-B, Zillikens D. Detection of elevated levels of IL-4, IL-6, and IL-10 in blister fluid of bullous pemphigoid. Arch Dermatol Res. (1996) 288:353–7. doi: 10.1007/BF02507102
33. Polukort SH, Rovatti J, Carlson L, Thompson C, Ser-Dolansky J, Kinney SRM, et al. IL-10 enhances IgE-mediated mast cell responses and is essential for the development of experimental food allergy in IL-10-deficient mice. J Immunol. (2016) 196:4865–76. doi: 10.4049/jimmunol.1600066
34. Cozzani E, Gasparini G, Di Zenzo G, Parodi A. Immunoglobulin E and bullous pemphigoid. Eur J Dermatol. (2018) 28:440–8. doi: 10.1684/ejd.2018.3366
35. Soldano S, Pizzorni C, Paolino S, Trombetta AC, Montagna P, Brizzolara R, et al. Alternatively activated (M2) macrophage phenotype is inducible by Endothelin-1 in cultured human macrophages. PLoS ONE. (2016) 11:e0166433. doi: 10.1371/journal.pone.0166433
36. Mantovani A, Biswas SK, Galdiero MR, Sica A, Locati M. Macrophage plasticity and polarization in tissue repair and remodelling. J Pathol. (2013) 229:176–85. doi: 10.1002/path.4133
37. Biswas SK, Chittezhath M, Shalova IN, Lim J-Y. Macrophage polarization and plasticity in health and disease. Immunol Res. (2012) 53:11–24. doi: 10.1007/s12026-012-8291-9
38. Hoffmann S, Berneburg M, Schreml S. Bullous pemphigoid associated with adalimumab therapy in a patient with ulcerative colitis. Case Rep Dermatol. (2018) 10:145–8. doi: 10.1159/000489163
39. Wessman LL, Blixt EK, Wetter DA, Miest RY. Adalimumab-associated bullous pemphigoid in a patient with ulcerative colitis. JAAD Case Rep. (2017) 3:339–41. doi: 10.1016/j.jdcr.2017.03.008
40. Wilmer EN, Becker N, Chen A, Kroumpouzos G. Etanercept-induced generalization of chronic, localized, anogenital bullous pemphigoid in a psoriatic patient. JAAD Case Rep. (2016) 2:25–7. doi: 10.1016/j.jdcr.2015.12.006
41. Boussemart L, Jacobelli S, Batteux F, Goulvestre C, Grange P, Carlotti A, et al. Autoimmune bullous skin diseases occurring under anti-tumor necrosis factor therapy: two case reports. Dermatology (Basel). (2010) 221:201–5. doi: 10.1159/000318008
Keywords: bullous pemphigoid, autoimmunity, inflammation, macrophage polarization, MMP-9
Citation: Riani M, Muller C, Bour C, Bernard P, Antonicelli F and Le Jan S (2019) Blister Fluid Induces MMP-9-Associated M2-Type Macrophages in Bullous Pemphigoid. Front. Immunol. 10:1858. doi: 10.3389/fimmu.2019.01858
Received: 27 May 2019; Accepted: 23 July 2019;
Published: 07 August 2019.
Edited by:
Ralf J. Ludwig, Universität zu Lübeck, GermanyReviewed by:
Takashi Hashimoto, Osaka City University, JapanKyle T. Amber, University of Illinois at Chicago, United States
Copyright © 2019 Riani, Muller, Bour, Bernard, Antonicelli and Le Jan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sébastien Le Jan, c2ViYXN0aWVuLmxlLWphbkB1bml2LXJlaW1zLmZy
†These authors have contributed equally to this work