 Johanna BödderTasmin Zahan
Johanna BödderTasmin Zahan Rianne van Slooten
Rianne van Slooten Gerty Schreibelt
Gerty Schreibelt I. Jolanda M. de Vries*
I. Jolanda M. de Vries* Georgina Flórez-Grau
Georgina Flórez-Grau- Department of Tumor Immunology, Radboud Institute for Molecular Life Sciences, Radboud University Medical Center, Nijmegen, Netherlands
Immunotherapeutic approaches have revolutionized the treatment of several diseases such as cancer. The main goal of immunotherapy for cancer is to modulate the anti-tumor immune responses by favoring the recognition and destruction of tumor cells. Recently, a better understanding of the suppressive effect of the tumor microenvironment (TME) on immune cells, indicates that restoring the suppressive effect of the TME is crucial for an efficient immunotherapy. Natural killer (NK) cells and dendritic cells (DCs) are cell types that are currently administered to cancer patients. NK cells are used because of their ability to kill tumor cells directly via cytotoxic granzymes. DCs are employed to enhance anti-tumor T cell responses based on their ability to present antigens and induce tumor-antigen specific CD8+ T cell responses. In preclinical models, a particular DC subset, conventional type 1 DCs (cDC1s) is shown to be specialized in cross-presenting extracellular antigens to CD8+ T cells. This feature makes them a promising DC subset for cancer treatment. Within the TME, cDC1s show a bidirectional cross-talk with NK cells, resulting in a higher cDC1 recruitment, differentiation, and maturation as well as activation and stimulation of NK cells. Consequently, the presence of cDC1s and NK cells within the TME might be of utmost importance for the success of immunotherapy. In this review, we discuss the function of cDC1s and NK cells, their bidirectional cross-talk and potential strategies that could improve cancer immunotherapy.
Introduction
Cancer immunotherapy is an approach that aims to activate the immune system to fight cancer. The immune system consists of many different cell types interacting with each other. Natural Killer (NK) cells and dendritic cells (DCs) are two cell types used for immunotherapy that are currently being tested in the clinic.
NK cells are granular innate lymphoid cells that display rapid, contact-dependent cytotoxic activities against viral-infected and cancer cells without prior sensitization. In general, NK cells rapidly accumulate at sites of inflammation, where they recruit other immune cells via cytokine and chemokine secretion. After activation, NK cells induce lysis or apoptosis in mutated cells by releasing granules containing cytotoxic granzymes. Activation occurs in an antigen-independent manner that is regulated by a tight balance of activating and inhibitory germline-encoded surface receptor ligation (1). Activating receptors bind to ligands (e.g., CD155, CD112) upregulated on tumor cells. Inhibitory receptors recognize major histocompatibility complex (MHC) class I, which is expressed by all nucleated cells, and upon binding, suppress NK cell activation. Hence, the highly diverse receptor repertoire on NK cells and the balance of activating and inhibitory receptors determine the magnitude of NK cell-mediated cytotoxicity and allow them to remain tolerant towards healthy cells (1–3). DCs, are a heterogeneous cell population which main function is to initiate an immune response. Immature DCs act as sentinels as they take up antigens, undergo a maturation process, and present these antigens on MHC molecules to naive T cells in lymph nodes. In general, antigen presenting cells present endogenous antigens on MHC class I (MHC-I), and exogenous antigens on MHC class II molecules (MHC-II) and thereby prime and activate CD4+ and CD8+ T cells, respectively (4). However, DCs have the unique capacity to present exogenous antigens on MHC-I molecules to CD8+ T cells, a process known as antigen cross-presentation.
NK cells are exploited as immunotherapeutic tool due to their cytotoxic and immunomodulatory functions and DCs because they are able to antigen-specifically activate T cells. However, both NK cell and DC functions can be restricted by the immunosuppressive tumor microenvironment (TME). In this review, we describe the main features of NK cells and a very rare type of DC, cDC1s, and emphasize the importance of these cell types within the TME. We focus on how to exploit cDC1s and NK cells and their interaction as a potential target to enhance efficacy of cancer immunotherapy.
Dendritic Cells
Both in humans and mice, circulating blood DCs have been classically divided into myeloid or conventional DCs (cDCs), and plasmacytoid DCs (pDCs). The DC subsets are classified by surface marker expression and different functional properties. Human pDCs express CD123, CD303 (BDCA-2) and CD304 (BDCA-4) as distinctive markers and are known for the production of large amounts of type 1 interferon (IFN-I) especially important for strong anti-viral responses (5). Conventional DCs express the common myeloid markers: CD11c, CD11b, CD33, and CD13 and are efficient in antigen presentation and T cell activation (5). They can be subdivided into type 1 conventional DCs (cDC1s) and type 2 cDCs (cDC2s) (6, 7). In humans, cDC1s express CD141 (BDCA-3) and cDC2s express CD1c (BDCA-1) (5). In mice, cDC2s are CD11b+, and cDC1s are characterized by CD8α+ or CD103+ expression (8). Genome-wide association studies of human and mouse cDC1s revealed phenotypic similarities, including expression of nectin-like protein 2 (Necl2), C-type lectin CLEC9a, and the XC chemokine receptor 1 (XCR1) as well as toll-like-receptor 3 (TLR-3) (Figure 1 ) (9–14). Hence, human cDC1s are considered to be the equivalent of the mouse CD8α+ D C subset (10–13, 15, 16) (Figure 1).
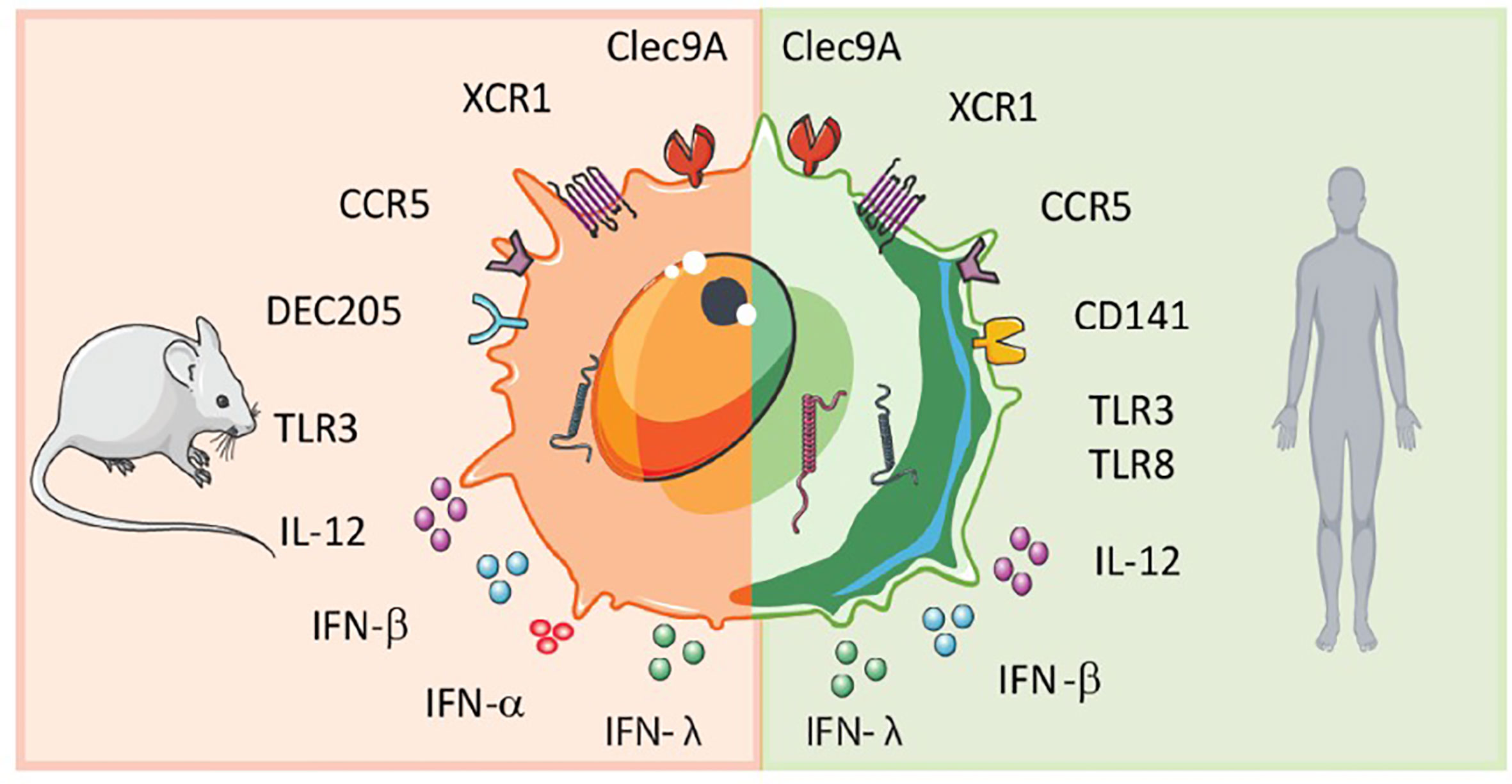
Figure 1 Scheme of murine and human cDC1 features. Human and mouse cDC1s display similarities but also differences in surface receptor expression and cytokine secretion.
cDC1s Phenotypic Characterization
Whereas in mice, cDC1s are the most abundant DC subset, human cDC1s are the rarest with approximately 0.03% of PBMCs and lymphoid and non-lymphoid cells are cDC1s (10, 11). This low occurrence of cDC1s resulted in difficulties to characterize them phenotypically. Initially, CD141 (BDCA-3) was described as a distinctive marker for cDC1s. However, CD141 was also found on cDC2s and other myeloid cells like monocytes (17). Therefore, many research groups sought to redefine subset division and attempted to identify conserved and exclusive markers across species. In this context, single-cell RNA-sequencing data recently demonstrated that cDC1s indeed form a single, separate cluster with CLEC9a as a specific marker for human cDC1s (18). CLEC9a is a receptor for necrotic cell-derived antigens (19). Yet, also CLEC9a is not exclusively expressed on cDC1s (13, 20, 21). The chemokine receptor XCR1 was established to be a specific marker for cDC1s. It is exclusively expressed on both human and mouse cDC1s (12, 13, 22–24). XCR1 mediates chemotaxis of cDC1s towards CD8+ T cells and NK cells, because they are the main producers of the ligand of XCR1, XC chemokine ligand 1 (XCL1), the ligand of XCR1 (12, 23). Besides exclusive expression of XCR1, cDC1s have a higher expression of the TLR-3 compared to other DCs subsets. TLR-3 signaling triggers IRF3/7, leading to IFN-ß secretion, thereby providing cDC1s with the enhanced capability to initiate T helper 1 responses (10, 11). As a result of the expression of CLEC9a and XCR1, cDC1s can take up necrotic cell-derived antigens and can migrate towards CD8+ T cells.
cDC1s Activate T Cells via Cross-Presentation
Cross-presentation of extracellular antigens is essential to activate CD8+ T cells specific for antigens derived from tumor cells (25). In mice, cDC1s are the only subset that cross-present antigens. Depletion of cDC1s, for example using Batf3 knockouts, results in loss of cross-presentation. Mice lacking cDC1s have less tumor-specific CD8+ T cells hence an impaired anti-tumor response (24, 26–29). Of note, depletion of cDC1s leads, besides to diminished cross-presentation also to loss of other functions mediated by cDC1s like attracting CD8+ T cells via XCL1-XCR1 interactions as well as less IL-12 and IFN-β since it is produced by cDC1s upon TLR-3 triggering. These processes also enhance anti-tumor responses and are less efficient upon depletion of cDC1s (6, 24).
In line with data obtained from mouse experiments, human cDC1s are shown to be highly efficient at antigen cross-presentation (10–13). However, in humans, cDC1s are not the only subset capable to cross-present. Plasmacytoid DCs, cDC2s and monocyte-derived DCs are also able to cross-present (30–34). However, data show that cDC1s are the most competent DC subset to cross-present antigens, especially of necrotic cells after uptake via CLEC9A (27, 35). The efficiency of cross-presentation of cDC1 is enhanced after triggering of TLR3 (11). Hence, human cDC1s are also highly specialized to cross-present antigens from necrotic tumor cells and to initiate a tumor antigen specific CD8+ T cell response (36, 37).
Natural Killer Cells
Human blood NK cells are a heterogeneous immune cell population classified by surface receptor expression of CD56 into two main functionally different subsets. CD56high NK cells secrete high levels of IFN-γ and other cytokines important for immunoregulatory functions, but produce low levels of perforin (38). Furthermore, CD56high NK cells express high levels of natural killer group 2A (NKG2A) and C-C chemokine receptor type 7 (CCR7) and low levels of Fcγ receptor IIIa (CD16) and killer cell immunoglobulin-like receptors (KIRs) (38, 39). In contrast, CD56low NK cells display a high expression of KIR and CD16 receptor. The receptor CD16 enables CD56low NK cells to mediate antibody-dependent cellular cytotoxicity (ADCCs). This CD16 expression together with the secretion of perforin and granzyme B provides CD56low NK cells with efficient killing abilities (38).
In general, NK cells display either killer (CD56lowNK cells) or helper (CD56high) functions against virus-infected or tumor cells due to activating and inhibitory receptor signaling and cytokine secretion (Figure 2). Inhibitory KIR and NKG2A receptors recognize MHC-I molecules. In the case of absent, changed, or mismatched MHC-I expression, the inhibitory receptor signal is lost, and NK cells get activated. The diversity of inhibitory receptors expressed on NK cells allows to recognize the high polymorphism of the MHC class I genes resulting in NK cell tolerance for healthy cells (3). Virus-infected or tumor cells often show gradually or complete loss of MHC-I expression to escape cytotoxic T cell recognition, simultaneously leading to a lack of inhibitory signal and NK cell activation (38, 40). Besides NK cell activation by loss of inhibitory signals, ligands overexpressed on mutated cells engaging with activating receptors such as natural cytotoxicity receptors (NKp30, NKp44, and NKp46) and NKG2D also lead to NK cell activation (41). Exposure of NK cells to cytokines such as IL-2 or IL-15 enhances activating receptor upregulation and promotes survival and proliferation (42–44). Activated NK cells can kill by forming a synapse with target cells and releasing perforin and cytotoxic granules. Perforin penetrates the membrane of target cells and granules containing cytotoxic granzymes enter target cells to provoke programmed cell death (45, 46). Another killing mechanism of NK cells is via ADCC. CD16 on NK cells recognize IgG antibody-coated tumor cells upon which granules are released by NK cells and target cells are killed (47). Some anti-tumor therapies based on neutralizing antibodies such as Rituximab or Trastuzumab demonstrate that the clinical benefit is partly mediated by ADCC (47).
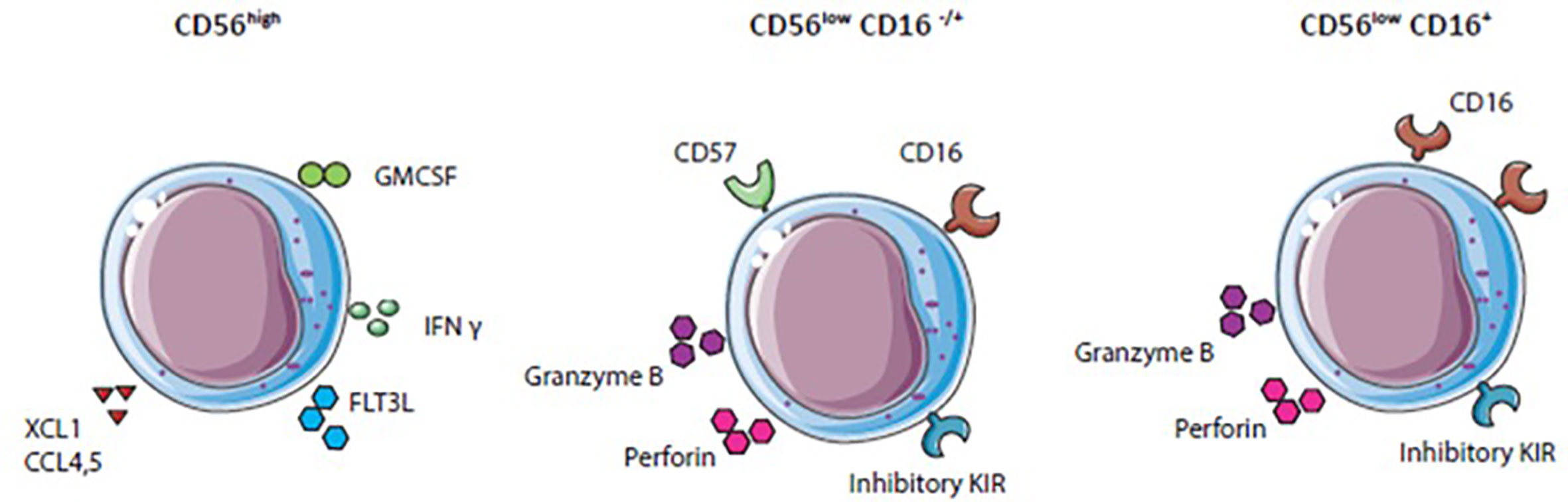
Figure 2 Natural killer (NK) cell subset differentation: NK cell subsets express variouus activating and inhibitory receptors and secrete different cytokines.
Immunomodulatory functions, mainly ascribed to CD56high NK cells are the secretion of different cytokines, chemokines, and growth factors, for example, IFN-γ, TNF-α, fms-like tyrosine kinase 3 ligand (FLT3L), Chemokine (C-C motif) ligand 3 (CCL3), CCL4, CCL5, granulocyte-macrophage colony-stimulating factor (GM-CSF) and XCL1 (41). These factors attract other immune cells like DCs, induce Th1 polarization, and CTL responses against target cells (41, 48). NK cells are essential for anti-tumor immune responses especially for eradication of MHC-I negative tumors. Different strategies to boost NK cell activity, particularly in the suppressive TME, are currently investigated.
Tumor Microenvironment and Immune Escape Mechanisms
Escaping immune surveillance is one of the key hallmarks of cancer (49). Different suppressive mechanisms are employed by cancer cells to bypass immune system attacks. These mechanisms include the upregulation of co-inhibitory ligands (such as programmed death-ligand 1, PD-L1), down-regulation or loss of MHC-I, or secretion of immunosuppressive factors like IL-10, VEGF, and TGF-β (50–54).
The composition of the TME differs between cancer type and patient and plays an essential role in the tumor immune escape. This highly heterogeneous TME is composed of tumor cells, blood vessels, tumor-infiltrating lymphocytes, and other immune cells, fibroblasts, endothelial cells, signaling molecules (cytokines, chemokines), and the extracellular matrix (55). Tumors can be divided into “hot” and “cold” depending on the presence or absence of effector immune cells within the tumor. The occurrence of effector immune cells, for example T cells which directly can eliminate tumor cells, impacts patient outcome (55). In some cancer types, such as epithelial ovarian cancer and colorectal cancer T cell infiltration, specifically in the tumor has been associated with positive clinical effects (56–58). Moreover, pre-existing CD8+ T cells within the tumor are crucial for tumor regression upon PD-1 checkpoint inhibition (59). In contrast to CD8+ T cells, regulatory T cells, myeloid-derived-suppressor cells, or tumor-associated macrophages inhibit the immune response and are therefore associated with tumor growth and a poor clinical outcome (60, 61).
NK Cells and DC Within the TME
Next to the presence of CD8+ T cells, the occurrence and active state of NK cells in the tumor are related to a positive prognostic outcome (62–64). In some types of cancer such as head and neck or prostate cancer, a high ratio of tumor infiltrating NK cells is associated with a positive clinical outcome for patients, whereas in non-small-cell lung cancer, the presence of NK cells within the TME has no clinical impact due to down-regulation of activating receptors on the infiltrated NK cells (63, 65). NK cells isolated from prostate cancer are mainly CD56high and display reduced cytotoxicity because of an increased inhibitory and decreased activating receptor expression pattern. In non- small cell lung carcinoma CTLA-4 expression is upregulated on NK cells in the TME compared with healthy tissue NK cells and negatively effects DC maturation (66). This unfavorable activating and inhibitory receptor expression pattern have been implicated as cause for impaired killing activity of NK cells in the TME (67, 68).
NK cells isolated from tumors differ phenotypically and transcriptomically from NK cells isolated from blood (69). These variations can arise from several cellular and soluble factors present within the TME, which influence phenotype, functionality, and migration characteristics of infiltrated NK cells (63, 66, 70). Soluble factors influencing NK cell cytotoxicity and infiltration into the TME for instance TGF-β and Prostaglandin E2 (PGE2) are secreted by immunosuppressive cells such as regulatory T cells, tumor-associated macrophages, myeloid-derived-suppressor cells (68, 71–73). TGF-β suppresses NK cell cytotoxicity by inhibiting IFN-γ secretion, down-regulating activating receptor expression, and the adapter molecule DAP12 (74–76). PGE2 hampers NK cell cytotoxicity by decreasing IFN-γ secretion, thereby facilitating cancer progression (77, 78). Other factors within the TME, like hypoxia, high expression of checkpoint receptor ligands, and chronic activation of activating receptors, can cause functional exhaustion of NK cells (79–81). Exhausted NK cells show a dampened inflammatory cytokine secretion pattern, a reduced activity due to decreased activating receptor expression and an upregulation of inhibitory receptors and checkpoint receptors such as PD1, TIM3, CD69, and LAG3 (63, 68, 82, 83).
Immunosuppressive factors within the TME also strongly influence the function of DCs within this microenvironment (Figure 3). Tumor-infiltrated DCs are less efficient in antigen presentation and cytokine production upon TLR stimulation than peripheral blood DCs (84, 85). Factors in the TME, including IL-10, IL-6, TGF-ß, and PGE2, modulate DCs, for example, inhibition of DC maturation. Hence, tumor-infiltrated DCs often display an immature phenotype with a low expression of co-stimulatory markers: CD80, CD86, and CD40, while co-inhibitory markers like PD-L1 and TIM3 are upregulated. Consequently, DCs within the TME are often associated with immunosuppressive and impaired functions (86–88). Notably, the infiltration of pDCs is associated with poor prognosis in breast- and ovarian cancer (89, 90). In contrast, cDC1s in the TME have been linked with favorable patient survival (91). Several reports indicate that cDC1s in tumors are associated with a higher number of CD8+ T cells within the TME, better prognosis of cancer patients, and immunotherapeutic success (36, 92–95). In addition to the presence of CD8+ T cells, the occurrence of cDC1s in the TME also correlates with high numbers of NK cells and better overall survival of melanoma patients (14, 94, 96). Positive effects of cDC1s within the TME seem to be partly mediated via their IL-12 secretion which is essential for the induction of T-helper 1 (Th1) responses and CD8+ T cell activation, both of which are crucial for a long-lasting anti-tumor response (97). Moreover, IL-12 effects IFN-γ production by NK cells. The lack of IL-12 secreting cDC1s in mice leads to reduced IFN-γ production by NK cells and growth of metastases (98). In human breast cancer, IL-12 expression correlates with cDC1s and cytotoxic effector molecules like IFNG (99). Nevertheless, positive effects of the presence of cDC1s can be negatively affected by different factors in the TME. For example, in mice, tumor-infiltrating cDC1s show increased PD-L1 expression upon antigen uptake and IFN-γ stimulation. High PD-L1 expression protects cDC1s from killing by CD8+ T cells. However, within the TME, this protective high PD-L1 expression on cDC1s can lead to a diminished anti-tumor immune response (100). Macrophage-derived IL-10 inhibits IL-12 production by cDC1s during chemotherapy, resulting in a lower CD8+ T cell cytotoxicity and, tumor progression in a mammary carcinoma mice model (99). A factor that affects the presence of cDC1s within the tumor is cyclooxygenases (COX), an enzyme crucial for PGE2 production. In mice, increased COX levels lead to diminished cDC1 numbers within melanoma tumors and a dampened activity by suppressed IL-12 production. COX inhibition, together with anti-PD-1 treatment, enhanced eradication of tumors. Changed immune cell infiltration due to COX expression is further confirmed in various tumor types in mice and in human tumors; COX levels correlate negatively with immune cell infiltration (101). Even though, cDC1s in the TME positively affect patient outcome different factors in the TME suppress their functional properties.
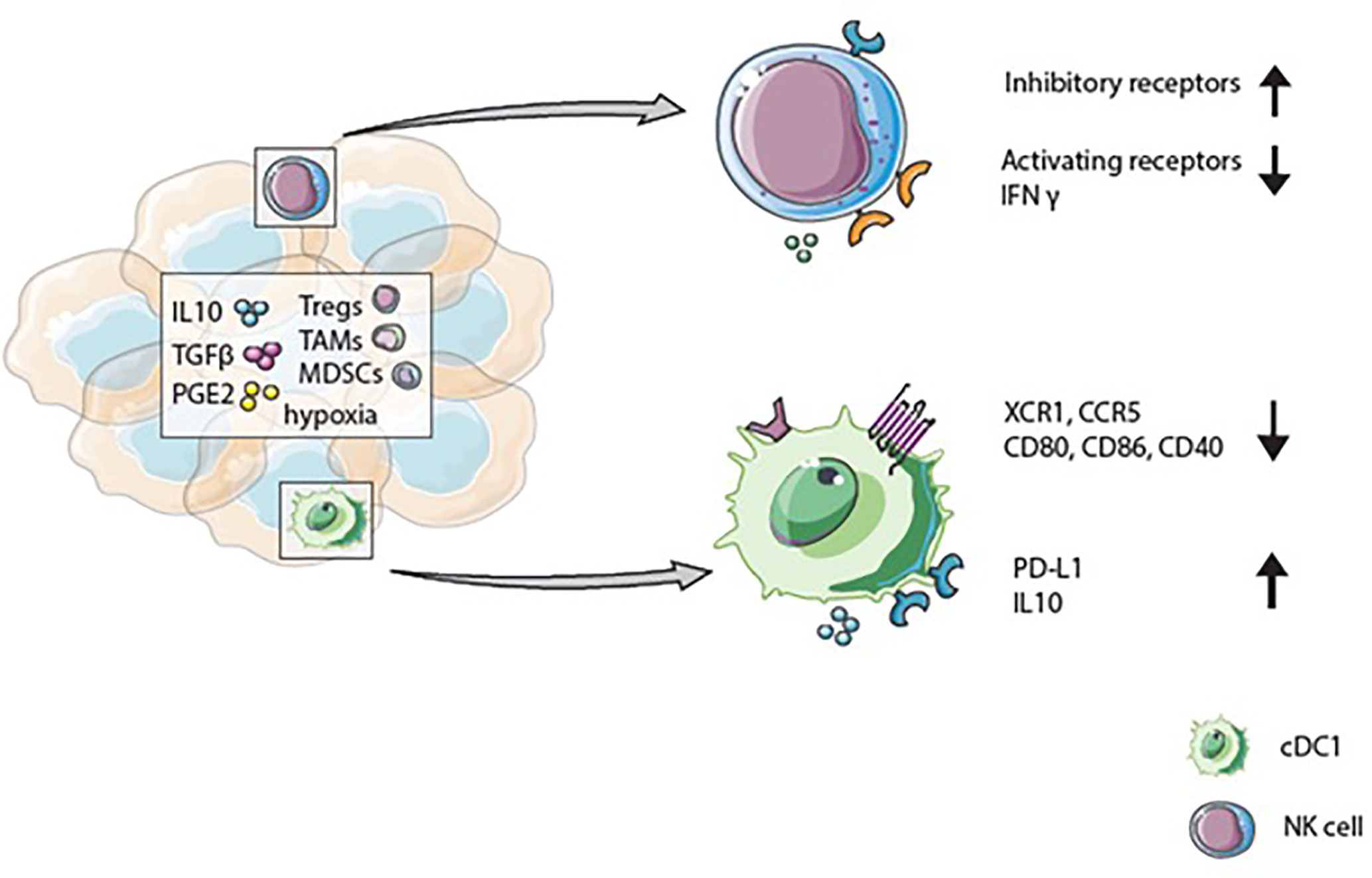
Figure 3 The tumor microenvironment influences cDC1 and natural killer (NK) cell phenotype and function. Immunosupressive factors like IL-10, TGF-β, and PGE2 can be secreted by tumor- and immune cells such as regulatory T cells (Tregs), tumor-associated macrophages (TAMs), or myeloid-derived-suppressor cells (MDSCs) present in the tumor microenvironment (TME). These factors, and hypoxia, can upregulate inhibitory receptors and decrease activating receptors on NK cells. Together with diminished IFN-γ secretion, the changed receptor expression results in reduced cytotoxicity of NK cells. Due to immunosuppressive factors in the TME cDC1s express low levels of XCR1 and CCR5 and display an immature phenotype with reduced CD80, CD86, and CD40 expression. Whereas checkpoint receptors and anti-inflammatory cytokine expression are upregulated.
Migration Patterns of NK Cells and cDC1s
The migration of immune cells towards lymph nodes and solid tumors is essential for proper immune activation and cancer cell elimination (55, 102). In general, the attraction and migration of immune cells occur through chemokine production and the interaction with their receptors (55). DC migration to lymph nodes is mediated by CCL19 and CCL21, the ligands for CCR7, where they present antigens to T cells and induce an antigen-specific immune response (103). The expression of CCR7 on cDC1s is crucial for migration and therefore important for antigen trafficking to lymph nodes. The lack of CCR7 expression in mice results in reduced numbers of cDC1s in the draining lymph node and increased tumor growth compared to wild type mice. In melanoma patients, cDC1s in the TME express high levels of CCR7, which predict T cell infiltration and improves clinical outcome, underscoring the importance of CCR7 expressing cDC1s for antigen trafficking and T cell priming (104).
After antigen specific activation of T cells in the lymph nodes these effector cells should migrate to the tumor to be able to kill tumor cells. Differences in chemokine and cytokine expression between hot and cold tumors impact the infiltration of effector cells (55, 105, 106). Hot tumors express high gene- and protein levels of chemokines CXC ligand (CXCL)9, CXCL10, and CXCL11 (106). All these three chemokines are secreted by DCs and bind to CXCR3 expressed on activated T- and NK cells. The high presence of CXCL9, CXCL10, and CXCL11 in the TME is associated with favorable clinical outcome in some cancers in humans (107). Interestingly, cDC1s are the primary source of those chemokines, underscoring the importance of cDC1s to attract effector cells towards the TME (95). For T cell chemotaxis into the tumor, the CXCR3/CXCL9,10,11 axis seems crucial (108, 109). For NK cells the role of CXCR3 for tumor infiltration is less straight forward. CXCR3 expression differs per NK cell subset, which influences their migration pattern. CD56high NK cells express CCR7 and CXCR3, CD56low NK cells express CXCR1 and CXCR2 (107, 110, 111). In breast and lung cancer, CXCL9 and CXCL10 mediate directed migration of CD56high NK cells, but not of cytotoxic CD56low NK cells (112). Moreover, suppressive factors present in the TME can influence chemokine production leading to altered NK cell migration. TGF-β favors CD56high and dampens CD56low NK cell recruitment. TGF-β decreases chemokine secretion of CXCL2, CX3CL1, and CXCL1 which attract CD56low and increases chemokine production, which recruit CD56high (CXCL9, CXCL10, CXCL11, and CCL5) (107, 110, 113). Even though factors in the TME can influence migration patterns of effector cells, the secretion of chemokines by cDC1s seem to be crucial for the recruitment of T cells and NK cells.
Cross-Talk cDC1s and NK Cells
The cross-talk between NK cells and cDC1s is bidirectional and NK cells as well as T cells can recruit cDC1s into the tumor via chemokine secretion and thereby promote anti-tumor immunity (14). In both preclinical and clinical settings, cDC1s are found enriched in TMEs with a specific chemokine profile. This profile includes XCL1, mainly secreted by tumor resident CD56low NK cells, and CCL4 and CCL5 mostly produced by CD56low and CD56high NK cells and CD8+ T cells (14). cDC1s express the receptors for those chemokines, namely XCR1 and CCR5 (14, 23). Indicating the importance of NK and CD8+ T cells for intratumoral migration of cDC1s due to XCL1, CCL5, and CCL4 (14). NK cells stimulated with IL-18 and IFN-α attract immature DC based on CCR5 expression, and stimulate DCs to increase CXCL9, CXCL10, and CCL5 production, promoting the attraction of effectors cells (114). In vitro, XCR1 and CCR5 expression on cDC1s is downregulated by PGE2, diminishing responsiveness to XCL1 and CCL5. In addition, PGE2 inhibits XCL1 and CCL5 secretion by NK cells, underlining the role of PGE2 as an immunosuppressive mediator to interfere with cDC1s migration to the tumor (14). In human tumors, cDC1 and NK cell gene signatures correlate with CCL5 and XCL1 gene expression and with CD8+ T cell infiltration. Further, NK cell and cDC1 gene signatures in the TME correlate positively with patient survival (14).
The cross-talk between NK cells and cDC1s influences the migration pattern of both cell types in addition to multiple other mechanisms by which DCs and NK cells interact (Figure 4). One mutual process is the maturation of DCs initiated by NK cells. Mature DCs release cytokines (IL-2, IL-12, or IL-18), which provoke NK cells to produce IFN-γ, TNF-α, or GM-CSF. These cytokines promote DC maturation (115, 116). Besides cytokine secretion, NK cells mature DCs via the ligation of CD40/CD40L (117). Upon CD40/CD40L ligation, the membrane bound IL-15 expression on DCs is upregulated promoting proliferation of NK cells (118, 119). The expression of CD40L on NK cells is regulated by IL-12 and IFN-γ and might determine cytotoxicity of NK cells (117, 120). Indeed, in a mouse tumor model, IL-12 and IFN-γ inhibition resulted in the down-regulation of CD40L expression on NK cells and diminished NK cell cytotoxicity (120).
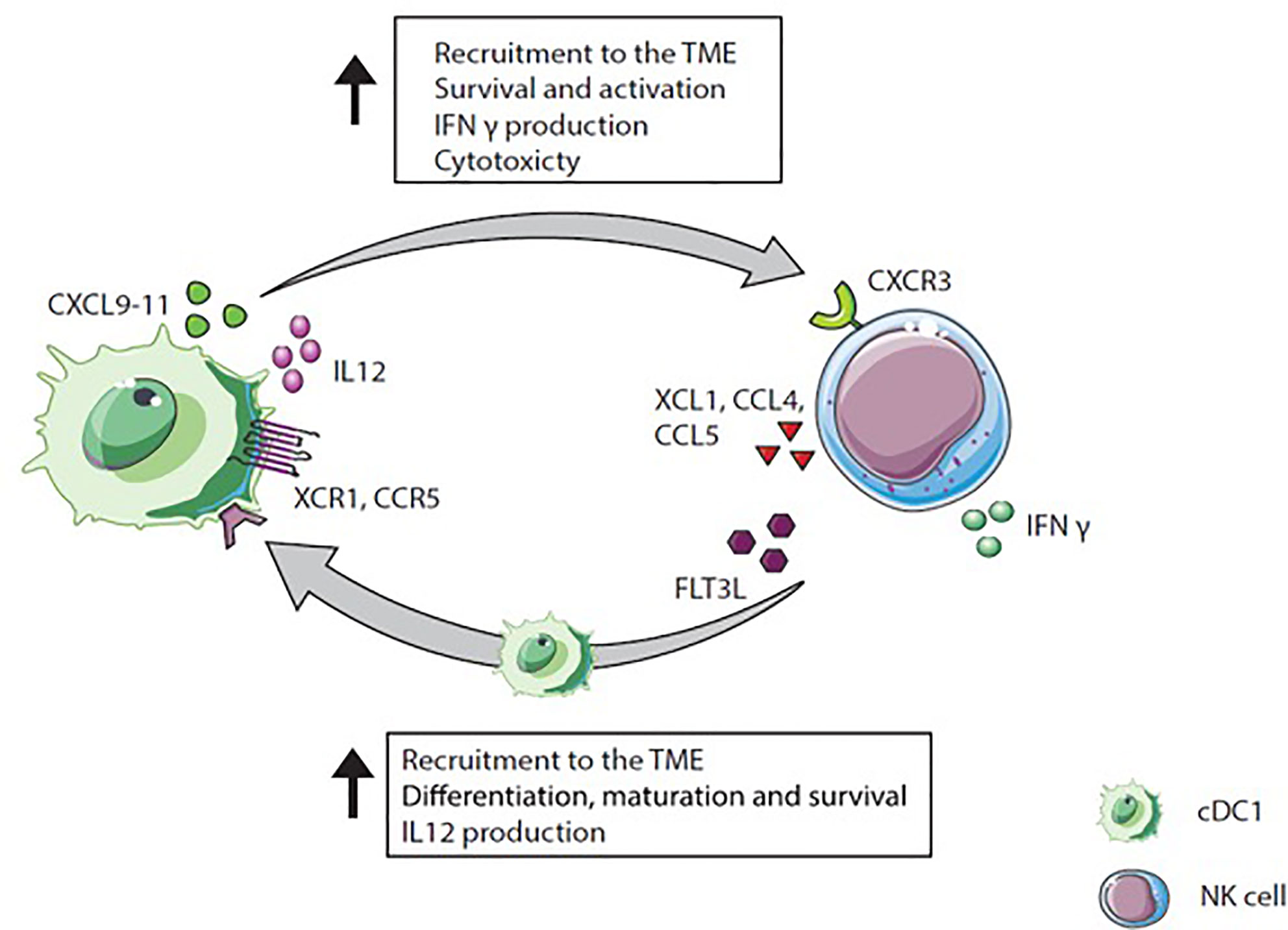
Figure 4 Natural killer (NK) cell and cDC1 cross-talk. NK cells secrete XCL1, CCL4, and CCL5 which attract XCR1 and CCR5 expressing cDC1s. Moreover, NK cells produce FLT3L, a differentiation factor for precursor DCs to cDC1s recruit NK cells via CXCL9-11 secretion into the TME. In a positive feedback loop, IL-12 produced by cDC1s activates IFN-γ extraction by NK cells, which again increases IL-12 secretion by cDC1s.
Besides maturation factors, NK cells produce the differentiation factor FLT3L, which stimulates cDC1 survival, differentiation, and recruitment and thereby positively influence cDC1s in the TME (94, 121). Interestingly, intratumoral NK cells in humans and mice are a major source of FLT3L, which maintains DC viability within the TME (94). The depletion of NK cells in mice reduced the frequency of cDC1s within the tumor, indicating that FLT3L production by NK cells is required for stable cDC1 numbers within the TME. In humans, genes encoding for FLT3LG within the tumor are linked to NK cell presence. In melanoma patients, the presence of cDC1s and NK cells within the tumor correlate with increased survival and responsiveness to anti-PD1 treatment. This underscores the hypothesis that NK cells producing FLT3L are responsible for the abundance of cDC1s within the tumor resulting in improved patient survival (94).
Taken together, chemokines such as XCL1, CCL4, and CCL5 secreted by NK cells and activated T cells within the TME recruit cDC1s into the tumor where they efficiently process antigens and then migrate to lymph nodes while they undergo maturation. In the draining lymph nodes, cDC1s prime naïve CD8+ T cells via cross-presentation. The cytotoxic T cells expand and are attracted to the tumor site, guided by local cDC1s secreting CXCL9 and CXCL10 (Figure 5). Thus, NK cells, cDC1s and their interaction are crucial for anti-tumor activity and, therefore, are promising targets to improve cancer immunotherapy outcome (6, 14, 37, 122).
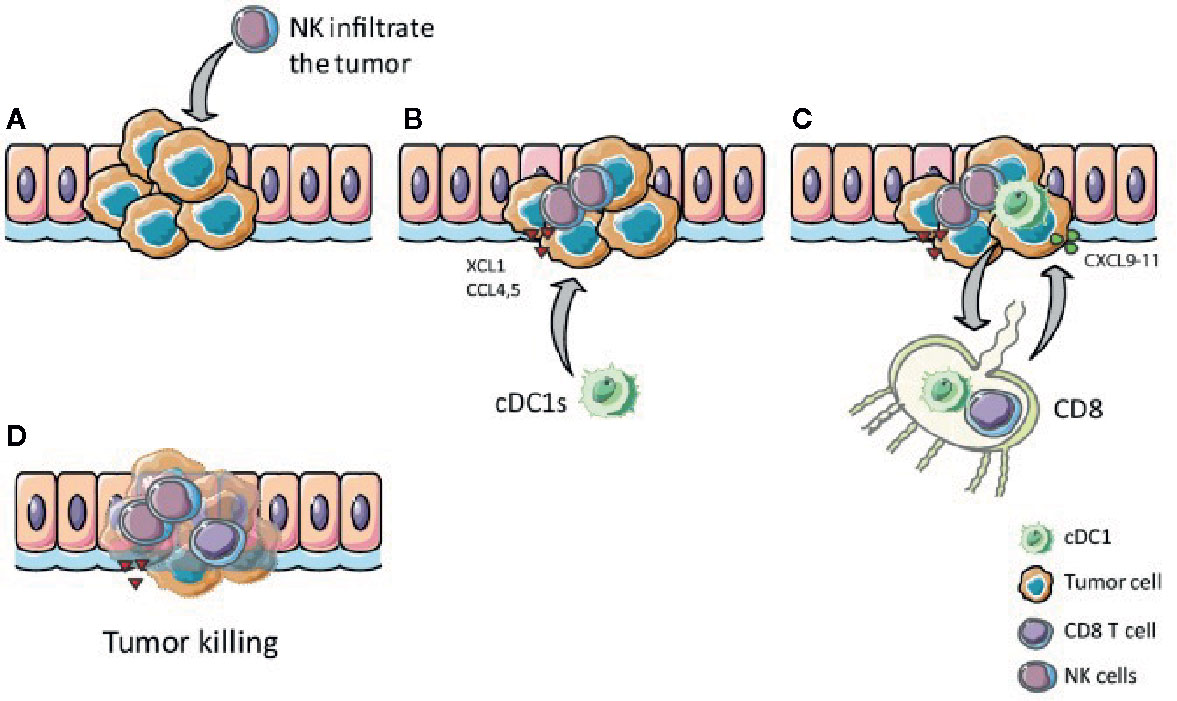
Figure 5 The XCR1 – XCL1 axis plays a role in tumor clearance. (A) NK and T cells, which infiltrate the tumor, produce XCL1 upon stimulation. (B) XCL1 attracts XCR1 expressing cDC1s to the tumor (C) cDC1s internalize, process, and cross-present tumor antigens to CD8+ T cells in the lymph nodes, thereby activate CD8+ T cells, which migrate to the tumor attracted by CXCL9-11 secreted by local cDC1s. (D) Activated CD8+ T cells kill tumor cells antigen-specifically
Manipulation of NK-cDC1 Cross Talk
Targeting cDC1s and XCR1-XCL1 Axis
The presence and interaction of cDC1s and NK cells within the TME are associated with activation and increased cytotoxicity of NK cells, cDC1s infiltration and maturation, and better prognosis for cancer patients (14, 93, 95). Hence, interfering with the XCR1-XCL1 axis could be an approach to increasecDC1s numbers and thereby also increase numbers of T cells and NK cells within the tumor (Figure 6). Activated intratumoral CD8+ T cells and NK cells produce XCL1, attracting XCR1 expressing cDC1s. The intratumoral injection of XCL1 can increase the migration of cDC1s towards the TME (91). In mice, administration of XCL1 linked with antigen (e.g., OVA) targeting cDC1s results in an antigen-specific T cell response and reduced tumor growth (123–125). Nonetheless, there is skepticism about using XCL1 to attract cDC1s because of its unstable structure and relatively weak chemotactic activity (126, 127). For this reason, Kazuhiko et al. engineered a stabilized, more potent agonist form of murine XCL1. Upon injection in mice, this potent version of XCL1 showed increased recruitment of XCR1+ DCs, compared to wild-type XCL1 (126).
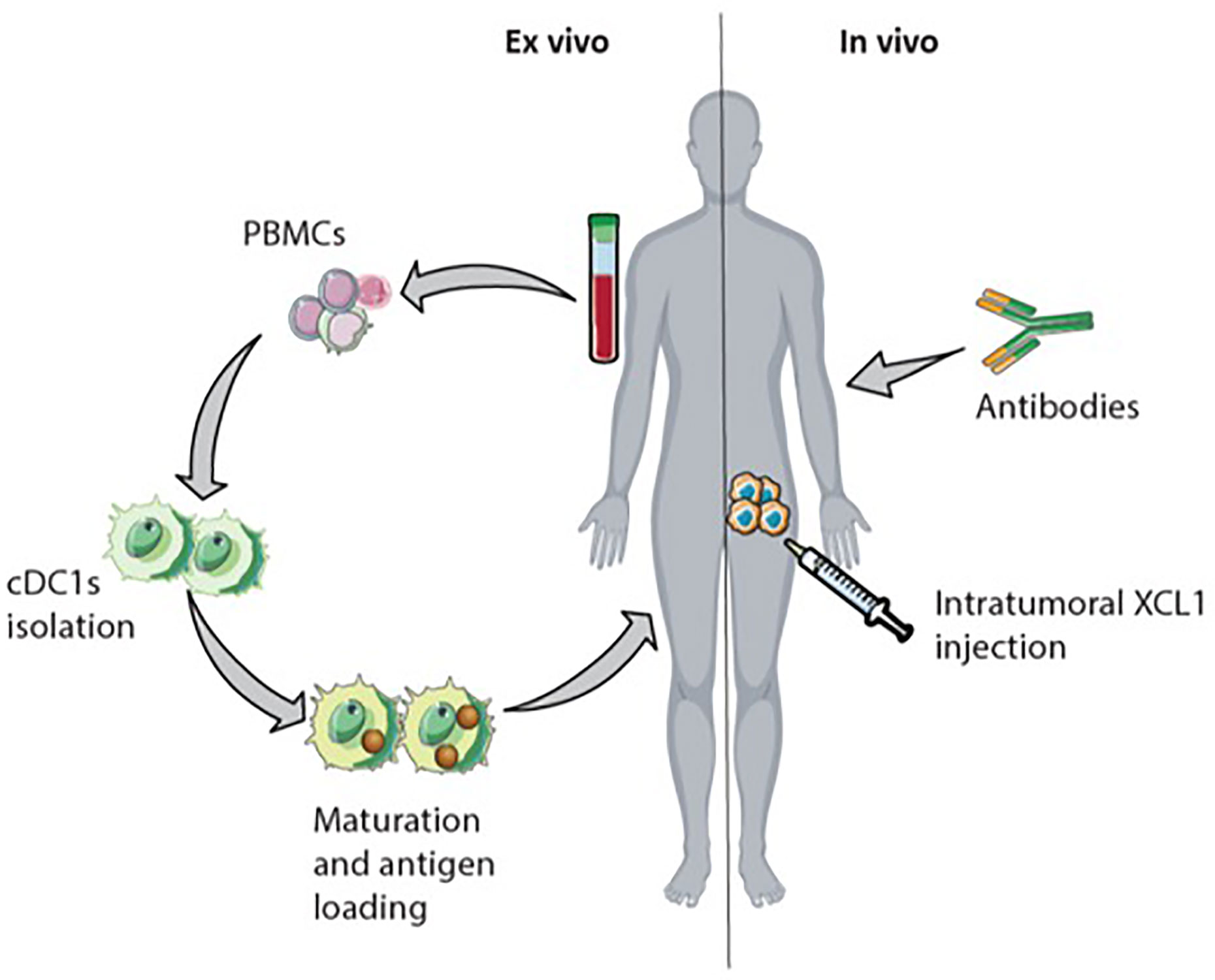
Figure 6 Possible immunotherapeutic approach targeting natural killer (NK) cells and cDC1s. Interfering with the XCR1-XCL1 axis in the tumor could increase antigen-specific CD8+ T cell infiltration. The intranodal reinfusion of cDC1s from patients after isolation them from PBMCs, maturation, and antigen-loading could be an ex vivo approach. In vivo, intratumoral XCL1 injection could recruit cDC1s into the tumor. Injected or attracted cross-presenting cDC1s in the tumor enhance CD8+ T cells activation. Monoclonal antibodies initiate antibody-dependent cellular cytotoxicity (ADCC) by NK cells leading to increased cross-presentation of tumor-cell derived antigens by cDC1s.
Besides targeting XCL1 to increase CD8+ T cell and NK cell numbers within the TME, direct administration of cDC1s into the TME could also be an approach. In mice, the application of cDC1s loaded with tumor-cell derived antigens enhanced tumor T cell infiltration and reduced tumor cell growth (128). DC therapy in the clinic shifts the application of monocyte-derived DC towards the application of blood-derived cDC2s and the combination of cDC2s and pDCs (129). Even though cDC1s are associated with improved survival in different types of cancer, the application of cDC1s is not yet assessed due to their low frequency in human blood (14, 93, 104, 129). Proposed alternatives to overcome the low abundance of cDC1s are the ex vivo generation of cDC1s from progenitor cells or expanding cDC1 progenitors in vivo (94, 130, 131). In vitro stimulation of human hematopoietic progenitors with Notch signaling and FLT3L induces cDC differentiation and yields phenotypical cDC1s with cross-presenting abilities (132). In mice, injection of FLT3L and intratumoral TLR3 stimulus poly IC, leads to expansion of cDC1 progenitor cells in the bone marrow and promotes cDC1s accumulation within the TME (131).
Monoclonal Antibody Treatment
Checkpoint blockade therapies targeting CTLA-4 and PD-1/PD-L1 are successfully used as cancer immunotherapy (133). PD-1 and CTLA-4 expression on NK cells is upregulated in several types of cancer and is associated with reduced cytotoxicity and cytokine secretion (66, 82, 134, 135). Especially in low MHC-I expressing tumors, the effect of mAbs blocking PD-1/PDL-1 might partly be facilitated by NK cells (136). Antibodies targeting PD-1 or CD40 stimulate IFN-γ secretion, which drives IL-12 production by cDC1s that licenses cytotoxic T cell responses in both mice and cancer patients (137). Mice lacking cDC1s display no specific T cell response upon mAbs targeting of PD-1, indicating that the effectiveness of anti-PD1 mAbs depends on the presence of cDC1s within the tumor (94, 131, 138). Moreover, the expression of PD-L1 on DC seems to influence the efficacy of anti-PD-L1 mAb treatment as mice lacking PD-L1 expression on DCs show no response to anti-PD-L1 mAb treatment (100). Based on pre-clinical data, it is proposed that therapies recruiting and activating NK cells within the TME could increase cDC1s within the tumor, benefitting responsiveness to checkpoint inhibitors (94). In line with that, enhanced NK cell activity in mice resulted in increased anti-PD-1 and anti-CTLA4 responses and a better control of tumor growth (139). In melanoma patients, anti-PD-1 treatment response correlates with the presence of cDC1s and NK cells within the TME (94).
Other mAbs target tumor-associated antigens like Her2/neu which are overexpressed on malignant cells compared to healthy cells (140). Monoclonal Ab administration induces NK cell-mediated ADCC (47). NK cells activated via ADCC release soluble factors like cytotoxic granules and cytokines (IFN-γ and TNF-α). The uptake of cytotoxic granules initiates tumor cell apoptosis causing the release of tumor-cell derived antigens, which can be taken up and presented by DCs (47, 141). Simultaneously, IFN-γ and TNF-α secreted by NK cells activate cDC1s to cross-present antigens leading to an antigen-specific CD8+ T cell activation (47, 142). Moreover, mAb-activated NK cells can enhance NK cell-cDC1 cross-talk by activating cDC1s to secrete IL-12 which amplifies NK cell activation (141).
In some tumor types, inhibitory receptors such as KIRs and NKG2A are upregulated on NK cells (3, 143, 144). Therefore, mAb against these inhibitory receptors could be targets for blocking immune inhibition and thereby increase the cytotoxic potential of NK cells and anti-tumor immunity (145). mAbs targeting KIRs, NKGA2, and other inhibitory receptors are currently investigated in clinical trials and are until now considered safe with limited side effects (3, 145).
Conclusion
Both cDC1s and NK cells are important for a proper anti-tumor immune response. cDC1s due to their specialization in cross-presenting tumor antigens and initiating an antigen-specific T cell response. NK cells because of cytotoxic and immunomodulatory functions. Even though both cell types can be affected by immunosuppressive factors in the TME, the NK-cDC1 cross-talk within this microenvironment establishes the cooperative nature of protective immunity against tumors. cDC1s, as producer of IL-12, CXCR9, CXCR10, and CCL5, attract NK cells to the TME, initiate cytokine production and boost NK cell cytotoxicity. At the same time, NK cells enhance cDC1 accumulation in the TME via XCL1 and CCL4, CCL5 secretion, and favor differentiation of cDC precursors to cDC1s. NK cells further stimulate DC maturation via cytokine secretion and CD40 ligation. Moreover, NK cell-mediated lysis of target cell releases cell debris including tumor-antigens, processed by cDC1s and cross-presented to CD8+ T cells. Thus, NK cell-cDC1 cross-talk is a promising target for immunotherapy to improve the clinical outcome of cancer patients.
Author Contributions
JB wrote the manuscript and designed the figures. TZ and RS helped with the literature and wrote the manuscript. GS, IV, and GF-G designed and reviewed the manuscript. All authors contributed to the article and approved the submitted version.
Funding
NHealth Holland/SGF grant DC4Balance (LSHM18056-SGF) and EU grant PROCROP (grant nr. 635122).
Acknowledgement
Images were self-made using the server medical art.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Paul S, Lal G. The Molecular Mechanism of Natural Killer Cells Function and Its Importance in Cancer Immunotherapy. Front Immunol (2017) 8:1124–4. doi: 10.3389/fimmu.2017.01124
2. Long EO, Kim HS, Liu D, Peterson ME, Rajagopalan S. Controlling Natural Killer Cell Responses: Integration of Signals for Activation and Inhibition. Annu Rev Immunol (2013) 31(1):227–58. doi: 10.1146/annurev-immunol-020711-075005
3. Pende D, Falco M, Vitale M, Cantoni C, Vitale C, Munari E, et al. Killer Ig-Like Receptors (KIRs): Their Role in NK Cell Modulation and Developments Leading to Their Clinical Exploitation. Front Immunol (2019) 10(1179). doi: 10.3389/fimmu.2019.01179
4. Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature (1998) 392:245. doi: 10.1038/32588
5. Collin M, McGovern N, Haniffa M. Human dendritic cell subsets. Immunology (2013) 140(1):22–30. doi: 10.1111/imm.12117
6. Cancel J-C, Crozat K, Dalod M, Mattiuz R. Are Conventional Type 1 Dendritic Cells Critical for Protective Antitumor Immunity and How? Front Immunol (2019) 10(9). doi: 10.3389/fimmu.2019.00009
7. Guilliams M, Ginhoux F, Jakubzick C, Naik SH, Onai BU, Schraml BU, et al. Dendritic cells, monocytes and macrophages: a unified nomenclature based on ontogeny. Nat Rev Immunol (2014) 14:571. doi: 10.1038/nri3712
8. Schlitzer A, Ginhoux F. Organization of the mouse and human DC network. Curr Opin Immunol (2014) 26:90–9. doi: 10.1016/j.coi.2013.11.002
9. Robbins SH, Walzer T, Dembélé D, Thibault C, Defays A, Bessou G, et al. Novel insights into the relationships between dendritic cell subsets in human and mouse revealed by genome-wide expression profiling. Genome Biol (2008) 9(1):R17–7. doi: 10.1186/gb-2008-9-1-r17
10. van der Aa E, van Montfoort N, Woltman AM. BDCA3+CLEC9A+ human dendritic cell function and development. Semin Cell Dev Biol (2015) 41:39–48. doi: 10.1016/j.semcdb.2014.05.016
11. Jongbloed SL, Kassianos AJ, McDonald KJ, Clark GJ, Ju X, Angel CE, et al. Human CD141+ (BDCA-3)+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J Exp Med (2010) 207(6):1247–60. doi: 10.1084/jem.20092140
12. Bachem A, Güttler S, Hartung E, Ebstein F, Schaefer M, Tannert A, et al. Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells. J Exp Med (2010) 207(6):1273–81. doi: 10.1084/jem.20100348
13. Crozat K, Guiton R, Contreras V, Feuillet V, Dutertre C-A, Ventre E, et al. The XC chemokine receptor 1 is a conserved selective marker of mammalian cells homologous to mouse CD8alpha+ dendritic cells. J Exp Med (2010) 207(6):1283–92. doi: 10.1084/jem.20100223
14. Böttcher JP, Bonavita E, Chakravarty P, Blees H, Cabeza-Cabrerizo M, Sammicheli S, et al. NK Cells Stimulate Recruitment of cDC1 into the Tumor Microenvironment Promoting Cancer Immune Control. Cell (2018) 172(5):1022–1037.e14. doi: 10.1016/j.cell.2018.01.004
15. Villadangos JA, Shortman K. Found in translation: the human equivalent of mouse CD8+ dendritic cells. J Exp Med (2010) 207(6):1131–4. doi: 10.1084/jem.20100985
16. Guilliams M, Dutertre C-A, Scott CL, McGovern N, Sichien D, Chakarov S, et al. Unsupervised High-Dimensional Analysis Aligns Dendritic Cells across Tissues and Species. Immunity (2016) 45(3):669–84. doi: 10.1016/j.immuni.2016.08.015
17. Chu C-C, Ali N, Karagiannis P, Di Meglio P, Skowera A, Napolitano L, et al. Resident CD141 (BDCA3)+ dendritic cells in human skin produce IL-10 and induce regulatory T cells that suppress skin inflammation. J Exp Med (2012) 209(5):935–45. doi: 10.1084/jem.20112583
18. Villani A-C, Satija R, Reynolds G, Sarkizova S, Shekhar K, Fletcher J, et al. Single-cell RNA-seq reveals new types of human blood dendritic cells, monocytes, and progenitors. Science (2017) 356(6335):eaah4573. doi: 10.1126/science.aah4573
19. Schreibelt G, Klinkenberg LJJ, Cruz LJ, Tacken PJ, Tel J, Kreutz M, et al. The C-type lectin receptor CLEC9A mediates antigen uptake and (cross-)presentation by human blood BDCA3+ myeloid dendritic cells. Blood (2012) 119(10):2284–92. doi: 10.1182/blood-2011-08-373944
20. Huysamen C, Willment JA, Dennehy KM, Brown GD. CLEC9A Is a Novel Activation C-type Lectin-like Receptor Expressed on BDCA3+ Dendritic Cells and a Subset of Monocytes. J Biol Chem (2008) 283(24):16693–701. doi: 10.1074/jbc.M709923200
21. Thul PJ, Åkesson L, Wiking M, Mahdessian D, Geladaki A, Ait Blal H, et al. A subcellular map of the human proteome. Science (2017) 356(6340):eaal3321. doi: 10.1126/science.aal3321
22. Sancho D, Joffre OP, Keller AM, Rogers NC, Martínez D, Hernanz-Falcön P, et al. Identification of a dendritic cell receptor that couples sensing of necrosis to immunity. Nature (2009) 458:899. doi: 10.1038/nature07750
23. Dorner BG, Dorner MB, Zhou X, Opitz C, Mora A, Güttler S, et al. Selective Expression of the Chemokine Receptor XCR1 on Cross-presenting Dendritic Cells Determines Cooperation with CD8+ T Cells. Immunity (2009) 31(5):823–33. doi: 10.1016/j.immuni.2009.08.027
24. Maier B, Leader AM, Chen ST, Tung N, Chang C, LeBerichel J, et al. A conserved dendritic-cell regulatory program limits antitumour immunity. Nature (2020) 580(7802):257–62. doi: 10.1038/s41586-020-2326-5
25. Huang A, Golumbek P, Ahmadzadeh M, Jaffee E, Pardoll D, Levitsky H. Role of bone marrow-derived cells in presenting MHC class I-restricted tumor antigens. Science (1994) 264(5161):961–5. doi: 10.1126/science.7513904
26. Hildner K, Edelson BT, Purtha WE, Diamond M, Matsushita H, Kohyama M, et al. Batf3 deficiency reveals a critical role for CD8alpha+ dendritic cells in cytotoxic T cell immunity. Science (New York NY) (2008) 322(5904):1097–100. doi: 10.1126/science.1164206
27. Poulin LF, Reyal Y, Uronen-Hansson H, Schraml BU, Sancho D, Murphy KM, et al. DNGR-1 is a specific and universal marker of mouse and human Batf3-dependent dendritic cells in lymphoid and nonlymphoid tissues. Blood (2012) 119(25):6052–62. doi: 10.1182/blood-2012-01-406967
28. Hammerich L, Marron TU, Upadhyay R, Svensson-Arvelund J, Dhainaut M, Hussein S, et al. Systemic clinical tumor regressions and potentiation of PD1 blockade with in situ vaccination. Nat Med (2019) 25(5):814–24. doi: 10.1038/s41591-019-0410-x
29. Wylie B, Seppanen E, Xiao K, Zemek R, Zanker D, Prato S, et al. Cross-presentation of cutaneous melanoma antigen by migratory XCR1(+)CD103(-) and XCR1(+)CD103(+) dendritic cells. Oncoimmunology (2015) 4(8):e1019198–e1019198. doi: 10.1080/2162402X.2015.1019198
30. Tel J, Schreibelt G, Sittig SP, Mathan TSM, Buschow SI, Cruz LJ, et al. Human plasmacytoid dendritic cells efficiently cross-present exogenous Ags to CD8+ T cells despite lower Ag uptake than myeloid dendritic cell subsets. Blood (2013) 121(3):459–67. doi: 10.1182/blood-2012-06-435644
31. Nizzoli G, Krietsch J, Weick A, Steinfelder S, Facciotti F, Gruarin P, et al. Human CD1c+ dendritic cells secrete high levels of IL-12 and potently prime cytotoxic T-cell responses. Blood (2013) 122(6):932–42. doi: 10.1182/blood-2013-04-495424
32. Tullett KM, Lahoud MH, Radford KJ. Harnessing Human Cross-Presenting CLEC9A(+)XCR1(+) Dendritic Cells for Immunotherapy. Front Immunol (2014) 5:239–9. doi: 10.3389/fimmu.2014.00239
33. Cohn L, Chatterjee B, Esselborn F, Smed-Sörensen A, Nakamura N, Chalouni C, et al. Antigen delivery to early endosomes eliminates the superiority of human blood BDCA3+ dendritic cells at cross presentation. J Exp Med (2013) 210(5):1049–63. doi: 10.1084/jem.20121251
34. Chiang M-C, Tullett KM, Lee YS, Idris A, Ding Y, McDonald KJ, et al. Differential uptake and cross-presentation of soluble and necrotic cell antigen by human DC subsets. Eur J Immunol (2016) 46(2):329–39. doi: 10.1002/eji.201546023
35. Joffre OP, Segura E, Savina A, Amigorena S. Cross-presentation by dendritic cells. Nat Rev Immunol (2012) 12:557. doi: 10.1038/nri3254
36. Fu C, Jiang A. Dendritic Cells and CD8 T Cell Immunity in Tumor Microenvironment. Front Immunol (2018) 9:3059–9. doi: 10.3389/fimmu.2018.03059
37. Sánchez-Arráez A, Teijeira A, Sánchez-Paulete AR, Garasa S, Melero I, Sancho D, et al. Antigen cross-presentation and T-cell cross-priming in cancer immunology and immunotherapy. Ann Oncol (2017) 28(suppl_12):xii44–55. doi: 10.1093/annonc/mdx237
38. Freud AG, Mundy-Bosse BL, Yu J, Caligiuri MA. The Broad Spectrum of Human Natural Killer Cell Diversity. Immunity (2017) 47(5):820–33. doi: 10.1016/j.immuni.2017.10.008
39. Smith SL, Kennedy PR, Stacey KB, Worboys JD, Yarwood A, Seo S, et al. Diversity of peripheral blood human NK cells identified by single-cell RNA sequencing. Blood Adv (2020) 4(7):1388–406. doi: 10.1182/bloodadvances.2019000699
40. Garrido F, Ruiz-Cabello F, Aptsiauri N. Rejection versus escape: the tumor MHC dilemma. Cancer Immunol Immunother (2017) 66(2):259–71. doi: 10.1007/s00262-016-1947-x
41. Shimasaki N, Jain A, Campana D. NK cells for cancer immunotherapy. Nat Rev Drug Discov (2020) 19(3):200–18. doi: 10.1038/s41573-019-0052-1
42. Mukherjee S, Jensen H, Stewart W, Stewart D, Ray WC, Chen S-Y, et al. In silico modeling identifies CD45 as a regulator of IL-2 synergy in the NKG2D-mediated activation of immature human NK cells. Sci Signal (2017) 10(485):eaai9062. doi: 10.1126/scisignal.aai9062
43. Miller JS, Morishima C, McNeel DG, Patel MR, Kohrt HEK, Thompson JA, et al. A First-in-Human Phase I Study of Subcutaneous Outpatient Recombinant Human IL15 (rhIL15) in Adults with Advanced Solid Tumors. Clin Cancer Res (2018) 24(7):1525–35. doi: 10.1158/1078-0432.CCR-17-2451
44. Sakellariou C, Elhage O, Papaevangelou E, Giustarini G, Esteves AM, Smolarek D, et al. Prostate cancer cells enhance interleukin-15-mediated expansion of NK cells. BJU Int (2020) 125(1):89–102. doi: 10.1111/bju.14893
45. Gwalani LA, Orange JS. Single Degranulations in NK Cells Can Mediate Target Cell Killing. J Immunol (Baltimore Md 1950) (2018) 200(9):3231–43. doi: 10.4049/jimmunol.1701500
46. Krzewski K, Coligan JE. Human NK cell lytic granules and regulation of their exocytosis. Front Immunol (2012) 3:335–5. doi: 10.3389/fimmu.2012.00335
47. Wang W, Erbe AK, Hank JA, Morris ZS, Sondel PM. NK Cell-Mediated Antibody-Dependent Cellular Cytotoxicity in Cancer Immunotherapy. Front Immunol (2015) 6:368–8. doi: 10.3389/fimmu.2015.00368
48. Mocikat R, Braumüller H, Gumy A, Egeter O, Ziegler H, Reusch U, et al. Natural Killer Cells Activated by MHC Class ILow Targets Prime Dendritic Cells to Induce Protective CD8 T Cell Responses. Immunity (2003) 19(4):561–9. doi: 10.1016/S1074-7613(03)00264-4
49. Hanahan D, Weinberg RA. Hallmarks of Cancer: The Next Generation. Cell (2011) 144(5):646–74. doi: 10.1016/j.cell.2011.02.013
50. Vinay DS, Ryan EP, Pawelec G, Talib WH, Stagg J, Elkord E, et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin Cancer Biol (2015) 35:S185–98. doi: 10.1016/j.semcancer.2015.03.004
51. Seliger B. The link between MHC class I abnormalities of tumors, oncogenes, tumor suppressor genes, and transcription factors. J Immunotoxicol (2014) 11(4):308–10. doi: 10.3109/1547691X.2013.875084
52. Gabrilovich DI, Chen HL, Girgis KR, Cunningham HT, Meny GM, Nadaf S, et al. Production of vascular endothelial growth factor by human tumors inhibits the functional maturation of dendritic cells. Nat Med (1996) 2(10):1096–103. doi: 10.1038/nm1096-1096
53. Wu AA, Drake V, Huang HS, Chiu S, Zheng L. Reprogramming the tumor microenvironment: tumor-induced immunosuppressive factors paralyze T cells. Oncoimmunology (2015) 4(7):e1016700–e1016700. doi: 10.1080/2162402X.2015.1016700
54. Gerlini G, Tun-Kyi A, Dudli C, Burg G, Pimpinelli N, Nestle FO, et al. Metastatic melanoma secreted IL-10 down-regulates CD1 molecules on dendritic cells in metastatic tumor lesions. Am J Pathol (2004) 165(6):1853–63. doi: 10.1016/S0002-9440(10)63238-5
55. van der Woude LL, Gorris MAJ, Halilovic A, Figdor CG, de Vries IJM. Migrating into the Tumor: a Roadmap for T Cells. Trends Cancer (2017) 3(11):797–808. doi: 10.1016/j.trecan.2017.09.006
56. Zhang L, Conejo-Garcia JR, Katsaros D, Gimotty PA, Massobrio M, Regnani G, et al. Intratumoral T Cells, Recurrence, and Survival in Epithelial Ovarian Cancer. N Engl J Med (2003) 348(3):203–13. doi: 10.1056/NEJMoa020177
57. Naito Y, Saito K, Shiiba K, Ohuchi A, Saigenji K, Nagura H, et al. CD8+ T Cells Infiltrated within Cancer Cell Nests as a Prognostic Factor in Human Colorectal Cancer. Cancer Res (1998) 58(16):3491–4.
58. O’Donnell JS, Teng MWL, Smyth MJ. Cancer immunoediting and resistance to T cell-based immunotherapy. Nat Rev Clin Oncol (2019) 16(3):151–67. doi: 10.1038/s41571-018-0142-8
59. Tumeh PC, Harview CL, Yearley JH, Shintaku IP, Taylor EJM, Robert L, et al. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature (2014) 515(7528):568–71. doi: 10.1038/nature13954
60. Görgün GT, Whitehill G, Anderson JL, Hideshima T, Maguire C, Laubach J, et al. Tumor-promoting immune-suppressive myeloid-derived suppressor cells in the multiple myeloma microenvironment in humans. Blood (2013) 121(15):2975–87. doi: 10.1182/blood-2012-08-448548
61. Darrasse-Jèze G, Podsypanina K. How numbers, nature, and immune status of foxp3(+) regulatory T-cells shape the early immunological events in tumor development. Front Immunol (2013) 4:292–2. doi: 10.3389/fimmu.2013.00292
62. Eckl J, Buchner A, Prinz PU, Riesenberg R, Siegert SI, Kammerer R, et al. Transcript signature predicts tissue NK cell content and defines renal cell carcinoma subgroups independent of TNM staging. J Mol Med (2012) 90(1):55–66. doi: 10.1007/s00109-011-0806-7
63. Melaiu O, Lucarini V, Cifaldi L, Fruci D. Influence of the Tumor Microenvironment on NK Cell Function in Solid Tumors. Front Immunol (2020) 10(3038). doi: 10.3389/fimmu.2019.03038
64. Coca S, Perez-Piqueras J, Martinez D, Colmenarejo A, Saez MA, Vallejo C, et al. The prognostic significance of intratumoral natural killer cells in patients with colorectal carcinoma. Cancer (1997) 79(12):2320–8. doi: 10.1002/(SICI)1097-0142(19970615)79:12<2320::AID-CNCR5>3.0.CO;2-P
65. Carrega P, Morandi B, Costa R, Frumento G, Forte G, Altavilla G, et al. Natural killer cells infiltrating human nonsmall-cell lung cancer are enriched in CD56brightCD16– cells and display an impaired capability to kill tumor cells. Cancer (2008) 112(4):863–75. doi: 10.1002/cncr.23239
66. Russick J, Joubert P-E, Gillard-Bocquet M, Torset C, Meylan M, Petitprez F, et al. Natural killer cells in the human lung tumor microenvironment display immune inhibitory functions. J Immunother Cancer (2020) 8(2):e001054. doi: 10.1136/jitc-2020-001054
67. Pasero C, Gravis G, Granjeaud S, Guerin M, Thomassin-Piana J, Rocchi P, et al. Highly effective NK cells are associated with good prognosis in patients with metastatic prostate cancer. Oncotarget (2015) 6(16):14360–73. doi: 10.18632/oncotarget.3965
68. Pasero C, Gravis G, Guerin M, Granjeaud S, Thomassin-Piana J, Rocchi P, et al. Inherent and Tumor-Driven Immune Tolerance in the Prostate Microenvironment Impairs Natural Killer Cell Antitumor Activity. Cancer Res (2016) 76(8):2153–65. doi: 10.1158/0008-5472.CAN-15-1965
69. de Andrade LF, Lu Y, Luoma A, Ito Y, Pan D, Pyrdol JW, et al. Discovery of specialized NK cell populations infiltrating human melanoma metastases. JCI Insight (2019) 4(23):e133103. doi: 10.1172/jci.insight.133103
70. Chiossone L, Dumas P-Y, Vienne M, Vivier E. Natural killer cells and other innate lymphoid cells in cancer. Nat Rev Immunol (2018) 18(11):671–88. doi: 10.1038/s41577-018-0061-z
71. Peng L-S, Zhang J-Y, Teng Y-S, Zhao Y-L, Wang T-T, Mao F-Y, et al. Tumor-Associated Monocytes/Macrophages Impair NK-Cell Function via TGFβ1 in Human Gastric Cancer. Cancer Immunol Res (2017) 5(3):248–56. doi: 10.1158/2326-6066.CIR-16-0152
72. Ghiringhelli F, Ménard C, Terme M, Flament C, Taieb J, Chaput N, et al. CD4+CD25+ regulatory T cells inhibit natural killer cell functions in a transforming growth factor-beta-dependent manner. J Exp Med (2005) 202(8):1075–85. doi: 10.1084/jem.20051511
73. Li H, Han Y, Guo Q, Zhang M, Cao X. Cancer-Expanded Myeloid-Derived Suppressor Cells Induce Anergy of NK Cells through Membrane-Bound TGF-β1. J Immunol (2009) 182(1):240–9. doi: 10.4049/jimmunol.182.1.240
74. Batlle E, Massagué J. Transforming Growth Factor-β Signaling in Immunity and Cancer. Immunity (2019) 50(4):924–40. doi: 10.1016/j.immuni.2019.03.024
75. Trotta R, Dal Col J, Yu J, Ciarlariello D, Thomas B, Zhang X, et al. TGF-beta utilizes SMAD3 to inhibit CD16-mediated IFN-gamma production and antibody-dependent cellular cytotoxicity in human NK cells. J Immunol (Baltimore Md 1950) (2008) 181(6):3784–92. doi: 10.4049/jimmunol.181.6.3784
76. Castriconi R, Cantoni C, Della Chiesa M, Vitale M, Marcenaro E, Conte R, et al. Transforming growth factor β1 inhibits expression of NKp30 and NKG2D receptors: Consequences for the NK-mediated killing of dendritic cells. Proc Natl Acad Sci (2003) 100(7):4120–5. doi: 10.1073/pnas.0730640100
77. Holt D, Ma X, Kundu N, Fulton A. Prostaglandin E(2) (PGE (2)) suppresses natural killer cell function primarily through the PGE(2) receptor EP4. Cancer Immunol Immunother (2011) 60(11):1577–86. doi: 10.1007/s00262-011-1064-9
78. Park A, Lee Y, Kim MS, Kang YJ, Park YJ, Jung H, et al. Prostaglandin E2 Secreted by Thyroid Cancer Cells Contributes to Immune Escape Through the Suppression of Natural Killer (NK) Cell Cytotoxicity and NK Cell Differentiation. Front Immunol (2018) 9:1859–9. doi: 10.3389/fimmu.2018.01859
79. Concha-Benavente F, Kansy B, Moskovitz J, Moy J, Chandran U, Ferris RL. PD-L1 Mediates Dysfunction in Activated PD-1(+) NK Cells in Head and Neck Cancer Patients. Cancer Immunol Res (2018) 6(12):1548–60. doi: 10.1158/2326-6066.CIR-18-0062
80. Alvarez M, Simonetta F, Baker J, Pierini A, Wenokur AS, Morrison AR, et al. Regulation of murine NK cell exhaustion through the activation of the DNA damage repair pathway. JCI Insight (2019) 5(14):e127729. doi: 10.1172/jci.insight.127729
81. Parodi M, Raggi F, Cangelosi D, Manzini C, Balsamo M, Blengio F, et al. Hypoxia Modifies the Transcriptome of Human NK Cells, Modulates Their Immunoregulatory Profile, and Influences NK Cell Subset Migration. Front Immunol (2018) 9:2358–8. doi: 10.3389/fimmu.2018.02358
82. Ben-Shmuel A, Biber G, Barda-Saad M. Unleashing Natural Killer Cells in the Tumor Microenvironment–The Next Generation of Immunotherapy? Front Immunol (2020) 11(275). doi: 10.3389/fimmu.2020.00275
84. Perrot I, Blanchard D, Freymond N, Isaac S, Guibert B, Pachéco I, et al. Dendritic Cells Infiltrating Human Non-Small Cell Lung Cancer Are Blocked at Immature Stage. J Immunol (2007) 178(5):2763–9. doi: 10.4049/jimmunol.178.5.2763
85. Hartmann E, Wollenberg B, Rothenfusser S, Wagner M, Wellisch D, Mack B, et al. Identification and Functional Analysis of Tumor-Infiltrating Plasmacytoid Dendritic Cells in Head and Neck Cancer. Cancer Res (2003) 63(19):6478–87.
86. Krempski J, Karyampudi L, Behrens MD, Erskine CL, Hartmann L, Dong H, et al. Tumor-infiltrating programmed death receptor-1+ dendritic cells mediate immune suppression in ovarian cancer. J Immunol (Baltimore Md 1950) (2011) 186(12):6905–13. doi: 10.4049/jimmunol.1100274
87. Harimoto H, Shimizu M, Nakagawa Y, Nakatsuka K, Wakabayashi A, Sakamoto C, et al. Inactivation of tumor-specific CD8⁺ CTLs by tumor-infiltrating tolerogenic dendritic cells. Immunol Cell Biol (2013) 91(9):545–55. doi: 10.1038/icb.2013.38
88. Tran Janco JM, Lamichhane P, Karyampudi L, Knutson KL. Tumor-infiltrating dendritic cells in cancer pathogenesis. J Immunol (Baltimore Md 1950) (2015) 194(7):2985–91. doi: 10.4049/jimmunol.1403134
89. Faget J, Sisirak V, Blay JY, Caux C, Bendriss-Vermare N, Ménétrier-Caux C. ICOS is associated with poor prognosis in breast cancer as it promotes the amplification of immunosuppressive CD4(+) T cells by plasmacytoid dendritic cells. Oncoimmunology (2013) 2(3):e23185–5. doi: 10.4161/onci.23185
90. Labidi-Galy SI, Treilleux I, Goddard-Leon S, Combes J-D, Blay J-Y, Ray-Coquard I, et al. Plasmacytoid dendritic cells infiltrating ovarian cancer are associated with poor prognosis. Oncoimmunology (2012) 1(3):380–2. doi: 10.4161/onci.18801
91. Böttcher JP, Reis E Sousa C. The Role of Type 1 Conventional Dendritic Cells in Cancer Immunity. Trends Cancer (2018) 4(11):784–92. doi: 10.1016/j.trecan.2018.09.001
92. Spranger S, Bao R, Gajewski TF. Melanoma-intrinsic β-catenin signalling prevents anti-tumour immunity. Nature (2015) 523:231. doi: 10.1038/nature14404
93. Broz ML, Binnewies M, Boldajipour B, Nelson AE, Pollack JL, Erle DJ, et al. Dissecting the tumor myeloid compartment reveals rare activating antigen-presenting cells critical for T cell immunity. Cancer Cell (2014) 26(5):638–52. doi: 10.1016/j.ccell.2014.09.007
94. Barry KC, Hsu J, Broz ML, Cueto FJ, Binnewies M, Combes AJ, et al. A natural killer-dendritic cell axis defines checkpoint therapy-responsive tumor microenvironments. Nat Med (2018) 24(8):1178–91. doi: 10.1038/s41591-018-0085-8
95. Spranger S, Dai D, Horton B, Gajewski TF. Tumor-Residing Batf3 Dendritic Cells Are Required for Effector T Cell Trafficking and Adoptive T Cell Therapy. Cancer Cell (2017) 31(5):711–23.e4. doi: 10.1016/j.ccell.2017.04.003
96. Dikshit A, Anderson CM, Zhang B, Ma X-J. Conventional type 1 dendritic cells and natural killer cells demonstrate strong correlation to cytotoxic T lymphocyte infiltration in cervical cancer tumors. J Immunol (2020) 204(1 Supplement):243.11–1. doi: 10.1158/1538-7445.AM2020-3306
97. Martínez-López M, Iborra S, Conde-Garrosa R, Sancho D. Batf3-dependent CD103+ dendritic cells are major producers of IL-12 that drive local Th1 immunity against Leishmania major infection in mice. Eur J Immunol (2015) 45(1):119–29. doi: 10.1002/eji.201444651
98. Mittal D, Vijayan D, Putz EM, Aguilera AR, Markey KA, Straube J, et al. Interleukin-12 from CD103+ Batf3-Dependent Dendritic Cells Required for NK-Cell Suppression of Metastasis. Cancer Immunol Res (2017) 5(12):1098–108. doi: 10.1158/2326-6066.CIR-17-0341
99. Ruffell B, Chang-Strachan D, Chan V, Rosenbusch A, Ho CMT, Pryer N, et al. Macrophage IL-10 blocks CD8+ T cell-dependent responses to chemotherapy by suppressing IL-12 expression in intratumoral dendritic cells. Cancer Cell (2014) 26(5):623–37. doi: 10.1016/j.ccell.2014.09.006
100. Peng Q, Qiu X, Zhang Z, Zhang S, Zhang Y, Liang Y, et al. PD-L1 on dendritic cells attenuates T cell activation and regulates response to immune checkpoint blockade. Nat Commun (2020) 11(1):4835. doi: 10.1038/s41467-020-18570-x
101. Zelenay S, van der Veen AG, Böttcher JP, Snelgrove KJ, Rogers N, Acton SE, et al. Cyclooxygenase-Dependent Tumor Growth through Evasion of Immunity. Cell (2015) 162(6):1257–70. doi: 10.1016/j.cell.2015.08.015
102. Seyfizadeh N, Muthuswamy R, Mitchell DA, Nierkens S, Seyfizadeh N. Migration of dendritic cells to the lymph nodes and its enhancement to drive anti-tumor responses. Crit Rev Oncol Hematol (2016) 107:100–10. doi: 10.1016/j.critrevonc.2016.09.002
103. Haessler U, Pisano M, Wu M, Swartz MA. Dendritic cell chemotaxis in 3D under defined chemokine gradients reveals differential response to ligands CCL21 and CCL19. Proc Natl Acad Sci U S A (2011) 108(14):5614–9. doi: 10.1073/pnas.1014920108
104. Roberts EW, Broz ML, Binnewies M, Headley MB, Nelson AE, Wolf DM, et al. Critical Role for CD103+/CD141+ Dendritic Cells Bearing CCR7 for Tumor Antigen Trafficking and Priming of T Cell Immunity in Melanoma. Cancer Cell (2016) 30(2):324–36. doi: 10.1016/j.ccell.2016.06.003
105. Lanitis E, Dangaj D, Irving M, Coukos G, et al. Mechanisms regulating T-cell infiltration and activity in solid tumors. Ann Oncol (2017) 28(suppl_12):xii18–32. doi: 10.1093/annonc/mdx238
106. Harlin H, Meng Y, Peterson AC, Zha Y, Tretiakova M, Slingluff C, et al. Chemokine expression in melanoma metastases associated with CD8+ T-cell recruitment. Cancer Res (2009) 69(7):3077–85. doi: 10.1158/0008-5472.CAN-08-2281
107. Susek KH, Karvouni M, Alici E, Lundqvist A. The Role of CXC Chemokine Receptors 1–4 on Immune Cells in the Tumor Microenvironment. Front Immunol (2018) 9(2159). doi: 10.3389/fimmu.2018.02159
108. Mikucki ME, Fisher DT, Matsuzaki J, Skitzki JJ, Gaulin NB, Muhitch JB, et al. Non-redundant requirement for CXCR3 signalling during tumoricidal T-cell trafficking across tumour vascular checkpoints. Nat Commun (2015) 6:7458–8. doi: 10.1038/ncomms8458
109. Tokunaga R, Zhang W, Naseem M, Puccini A, Berger MD, Soni S, et al. CXCL9, CXCL10, CXCL11/CXCR3 axis for immune activation - A target for novel cancer therapy. Cancer Treat Rev (2018) 63:40–7. doi: 10.1016/j.ctrv.2017.11.007
110. Castriconi R, Carrega P, Dondero A, Bellora F, Casu B, Regis S, et al. Molecular Mechanisms Directing Migration and Retention of Natural Killer Cells in Human Tissues. Front Immunol (2018) 9:2324–4. doi: 10.3389/fimmu.2018.02324
111. Kremer V, Ligtenberg MA, Zendehdel R, Seitz C, Duivenvoorden A, Wennerberg E, et al. Genetic engineering of human NK cells to express CXCR2 improves migration to renal cell carcinoma. J Immunother Cancer (2017) 5(1):73. doi: 10.1186/s40425-017-0275-9
112. Carrega P, Bonaccorsi I, Di Carlo E, Morandi B, Paul P, Rizzello V, et al. CD56brightPerforinlow Noncytotoxic Human NK Cells Are Abundant in Both Healthy and Neoplastic Solid Tissues and Recirculate to Secondary Lymphoid Organs via Afferent Lymph. J Immunol (2014) 192(8):3805–15. doi: 10.4049/jimmunol.1301889
113. Castriconi R, Dondero A, Bellora F, Moretta L, Castellano A, Locatelli F, et al. Neuroblastoma-Derived TGF-β1 Modulates the Chemokine Receptor Repertoire of Human Resting NK Cells. J Immunol (2013) 190(10):5321–8. doi: 10.4049/jimmunol.1202693
114. Wong JL, Berk E, Edwards RP, Kalinski P. IL-18–Primed Helper NK Cells Collaborate with Dendritic Cells to Promote Recruitment of Effector CD8+ T Cells to the Tumor Microenvironment. Cancer Res (2013) 73(15):4653–62. doi: 10.1158/0008-5472.CAN-12-4366
115. Fehniger TA, Shah MH, Turner MJ, VanDeusen JB, Whitman SP, Cooper MA, et al. Differential Cytokine and Chemokine Gene Expression by Human NK Cells Following Activation with IL-18 or IL-15 in Combination with IL-12: Implications for the Innate Immune Response. J Immunol (1999) 162(8):4511–20.
116. Gerosa F, Baldani-Guerra B, Nisii C, Marchesini V, Carra G, Trinchieri G. Reciprocal Activating Interaction between Natural Killer Cells and Dendritic Cells. J Exp Med (2002) 195(3):327–33. doi: 10.1084/jem.20010938
117. Ma DY, Clark EA. The role of CD40 and CD154/CD40L in dendritic cells. Semin Immunol (2009) 21(5):265–72. doi: 10.1016/j.smim.2009.05.010
118. Ferlazzo G, Pack M, Thomas D, Paludan C, Schmid D, Strowig T, et al. Distinct roles of IL-12 and IL-15 in human natural killer cell activation by dendritic cells from secondary lymphoid organs. Proc Natl Acad Sci U S A (2004) 101(47):16606. doi: 10.1073/pnas.0407522101
119. Cooper MA, Bush JE, Fehniger TA, VanDeusen JB, Waite RE, Liu Y, et al. In vivo evidence for a dependence on interleukin 15 for survival of natural killer cells. Blood (2002) 100(10):3633–8. doi: 10.1182/blood-2001-12-0293
120. Jyothi MD, Khar A. Regulation of CD40L expression on natural killer cells by interleukin-12 and interferon γ: its role in the elicitation of an effective antitumor immune response. Cancer Immunol Immunother (2000) 49(10):563–72. doi: 10.1007/s002620000151
121. Pulendran B, Smith JL, Caspary G, Brasel K, Pettit D, Maraskovsky E, et al. Distinct dendritic cell subsets differentially regulate the class of immune response in vivo. Proc Natl Acad Sci U S A (1999) 96(3):1036–41. doi: 10.1073/pnas.96.3.1036
122. Kroczek R, Henn V. The Role of XCR1 and its Ligand XCL1 in Antigen Cross-Presentation by Murine and Human Dendritic Cells. Front Immunol (2012) 3(14). doi: 10.3389/fimmu.2012.00014
123. Hartung E, Becker M, Bachem A, Reeg N, Jäkel E, Hutloff A, et al. Induction of Potent CD8 T Cell Cytotoxicity by Specific Targeting of Antigen to Cross-Presenting Dendritic Cells In Vivo via Murine or Human XCR1. J Immunol (2015) 194(3):1069–79. doi: 10.4049/jimmunol.1401903
124. Fossum E, Grødeland G, Terhorst D, Tveita AA, Vikse E, Mjaaland S, et al. Vaccine molecules targeting Xcr1 on cross-presenting DCs induce protective CD8+ T-cell responses against influenza virus. Eur J Immunol (2015) 45(2):624–35. doi: 10.1002/eji.201445080
125. Chen K, Wu Z, Zhao H, Wang Y, Ge Y, Wang D, et al. XCL1/Glypican-3 Fusion Gene Immunization Generates Potent Antitumor Cellular Immunity and Enhances Anti–PD-1 Efficacy. Cancer Immunol Res (2020) 8(1):81–93. doi: 10.1158/2326-6066.CIR-19-0210
126. Matsuo K, Kitahata K, Kawabata F, Kamei M, Hara Y, Takamura S, et al. A Highly Active Form of XCL1/Lymphotactin Functions as an Effective Adjuvant to Recruit Cross-Presenting Dendritic Cells for Induction of Effector and Memory CD8+ T Cells. Front Immunol (2018) 9(2775). doi: 10.3389/fimmu.2018.02775
127. Kelner G, Kennedy J, Bacon K, Kleyensteuber S, Largaespada D, Jenkins N, et al. Lymphotactin: a cytokine that represents a new class of chemokine. Science (1994) 266(5189):1395–9. doi: 10.1126/science.7973732
128. Wculek SK, Amores-Iniesta J, Conde-Garrosa R, Khouili SC, Melero I, Sancho D. Effective cancer immunotherapy by natural mouse conventional type-1 dendritic cells bearing dead tumor antigen. J Immunother Cancer (2019) 7(1):100–0. doi: 10.1186/s40425-019-0565-5
129. Bol KF, Schreibelt G, Rabold K, Wculek SK, Schwarze JK, Dzionek A, et al. The clinical application of cancer immunotherapy based on naturally circulating dendritic cells. J Immunother Cancer (2019) 7(1):109. doi: 10.1186/s40425-019-0580-6
130. Balan S, Arnold-Schrauf C, Abbas A, Couespel N, Savoret J, Imperatore F, et al. Large-Scale Human Dendritic Cell Differentiation Revealing Notch-Dependent Lineage Bifurcation and Heterogeneity. Cell Rep (2018) 24(7):1902–1915.e6. doi: 10.1016/j.celrep.2018.07.033
131. Salmon H, Idoyaga J, Rahman A, Leboeuf R, Remark R, Jordan S, et al. Expansion and Activation of CD103(+) Dendritic Cell Progenitors at the Tumor Site Enhances Tumor Responses to Therapeutic PD-L1 and BRAF Inhibition. Immunity (2016) 44(4):924–38. doi: 10.1016/j.immuni.2016.03.012
132. Kirkling ME, Cytlak U, Lau CM, Lewis KL, Resteu A, Khodadadi-Jamayran A, et al. Notch Signaling Facilitates In Vitro Generation of Cross-Presenting Classical Dendritic Cells. Cell Rep (2018) 23(12):3658–3672.e6. doi: 10.1016/j.celrep.2018.05.068
133. Topalian SL, Drake CG, Pardoll DM. Immune checkpoint blockade: a common denominator approach to cancer therapy. Cancer Cell (2015) 27(4):450–61. doi: 10.1016/j.ccell.2015.03.001
134. Mariotti FR, Petrini S, Ingegnere T, Tumino N, Besi F, Scordamaglia F, et al. PD-1 in human NK cells: evidence of cytoplasmic mRNA and protein expression. OncoImmunology (2019) 8(3):1557030. doi: 10.1080/2162402X.2018.1557030
135. Pesce S, Greppi M, Tabellini G, Rampinelli F, Parolini S, Olive D, et al. Identification of a subset of human natural killer cells expressing high levels of programmed death 1: A phenotypic and functional characterization. J Allergy Clin Immunol (2017) 139(1):335–346.e3. doi: 10.1016/j.jaci.2016.04.025
136. Hsu J, Hodgins JJ, Marathe M, Nicolai CJ, Bourgeois-Daigneault M-C, Trevino TN, et al. Contribution of NK cells to immunotherapy mediated by PD-1/PD-L1 blockade. J Clin Invest (2018) 128(10):4654–68. doi: 10.1172/JCI99317
137. Garris CS, Arlauckas SP, Kohler CS, Trefny MP, Garren S, Piot C, et al. Successful Anti-PD-1 Cancer Immunotherapy Requires T Cell-Dendritic Cell Crosstalk Involving the Cytokines IFN-γ and IL-12. Immunity (2018) 49(6):1148–1161.e7. doi: 10.1016/j.immuni.2018.09.024
138. Sánchez-Paulete AR, Cueto FJ, Martínez-López M, Labiano S, Morales-Kastresana A, Rodríguez-Ruiz ME, et al. Cancer Immunotherapy with Immunomodulatory Anti-CD137 and Anti-PD-1 Monoclonal Antibodies Requires BATF3-Dependent Dendritic Cells. Cancer Discov (2016) 6(1):71–9. doi: 10.1158/2159-8290.CD-15-0510
139. Blake SJ, Stannard K, Liu J, Allen S, Yong MC, Mittal D, et al. Suppression of Metastases Using a New Lymphocyte Checkpoint Target for Cancer Immunotherapy. Cancer Discov (2016) 6(4):446–59. doi: 10.1158/2159-8290.CD-15-0944
140. Sun Z, Fu Y-X, Peng H. Targeting tumor cells with antibodies enhances anti-tumor immunity. Biophys Rep (2018) 4(5):243–53. doi: 10.1007/s41048-018-0070-2
141. Lee SC, Srivastava RM, López-Albaitero A, Ferrone S, Ferris RL, et al. Natural killer (NK): dendritic cell (DC) cross talk induced by therapeutic monoclonal antibody triggers tumor antigen-specific T cell immunity. Immunol Res (2011) 50(2-3):248–54. doi: 10.1007/s12026-011-8231-0
142. Deauvieau F, Ollion V, Doffin A-C, Achard C, Fonteneau J-F, Verronese E, et al. Human natural killer cells promote cross-presentation of tumor cell-derived antigens by dendritic cells. Int J Cancer (2015) 136(5):1085–94. doi: 10.1002/ijc.29087
143. Monaco EL, Tremante E, Cerboni C, Melucci E, Sibilio L, Zingoni A, et al. Human Leukocyte Antigen E Contributes to Protect Tumor Cells from Lysis by Natural Killer Cells. Neoplasia (2011) 13(9):822–IN14. doi: 10.1593/neo.101684
144. Sun C, Xu J, Huang Q, Huang M, Wen H, Zhang C, et al. High NKG2A expression contributes to NK cell exhaustion and predicts a poor prognosis of patients with liver cancer. OncoImmunology (2017) 6(1):e1264562. doi: 10.1080/2162402X.2016.1264562
Keywords: natural killer cells, conventional type 1 DCs, cross-talk, tumor microenvironment, immunotherapy
Citation: Bödder J, Zahan T, van Slooten R, Schreibelt G, de Vries IJM and Flórez-Grau G (2021) Harnessing the cDC1-NK Cross-Talk in the Tumor Microenvironment to Battle Cancer. Front. Immunol. 11:631713. doi: 10.3389/fimmu.2020.631713
Received: 20 November 2020; Accepted: 29 December 2020;
Published: 19 February 2021.
Edited by:
Erik Wennerberg, Weill Cornell Medicine, United StatesReviewed by:
Alessandro Poggi, San Martino Hospital (IRCCS), ItalyFabian Benencia, Ohio University, United States
Copyright © 2021 Bödder, Zahan, van Slooten, Schreibelt, de Vries and Flórez-Grau. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: I. Jolanda M. de Vries, Sm9sYW5kYS5kZXZyaWVzQHJhZGJvdWR1bWMubmw=