 Shawn Abeynaike1,2
Shawn Abeynaike1,2 Silke Paust1,2*
Silke Paust1,2*- 1Department of Immunology and Microbiology, The Scripps Research Institute, La Jolla, CA, United States
- 2The Skaggs Graduate Program in Chemical and Biological Sciences, The Scripps Research Institute, La Jolla, CA, United States
With the discovery of antiretroviral therapy, HIV-1 infection has transitioned into a manageable but chronic illness, which requires lifelong treatment. Nevertheless, complete eradication of the virus has still eluded us. This is partly due to the virus’s ability to remain in a dormant state in tissue reservoirs, ‘hidden’ from the host’s immune system. Also, the high mutation rate of HIV-1 results in escape mutations in response to many therapeutics. Regardless, the development of novel cures for HIV-1 continues to move forward with a range of approaches from immunotherapy to gene editing. However, to evaluate in vivo pathogenesis and the efficacy and safety of therapeutic approaches, a suitable animal model is necessary. To this end, the humanized mouse was developed by McCune in 1988 and has continued to be improved on over the past 30 years. Here, we review the variety of humanized mouse models that have been utilized through the years and describe their specific contribution in translating HIV-1 cure strategies to the clinic.
Introduction
Human immunodeficiency virus-1 (HIV-1) was first discovered in 1983 by the laboratory of Luc Montagniers at the Pasteur Institute, by culturing T cells isolated from the lymph nodes of a patient with early symptoms of AIDS (1). Since its discovery, the battle to control and eradicate HIV-1 has been long and tumultuous. Close to four decades have passed. While great strides have been made in managing HIV and preventing the onset of AIDS in patients via daily treatment with combination antiretroviral therapy (ART) (2), a successful vaccine or curative treatment has yet to be developed. The key to research on HIV cure therapy is using a suitable animal model, as a comprehensive analysis of the human immune system is limited due to ethical and practical restrictions. Humans and chimpanzees are the natural hosts for HIV-1 replication. However, due to ethical and practical reasons are not amenable to most methods of experimentation. Mice reconstituted with human immune systems and non-human primates are the two animal models that have received the most attention in investigating HIV-1 pathogenesis. Non-human primate models provide many advantages, such as being a natural host for the closely related Simian Immunodeficiency Virus (SIV) and the chimeric SHIV virus and having similar anatomical and physiological features to humans. The complete range of advantages and disadvantages of these models have been reviewed elsewhere (3–5). In contrast, humanized mice contain human CD4+ T cells, which are permissible to HIV-1 infection and simultaneously allow in-depth analysis of the human immune response to HIV-1 pathogenesis in vivo. Humanized mice have provided important insights into preventative approaches to HIV-1 infection (6–8). These approaches have been reviewed in detail elsewhere (9, 10). Here we review the history and development of humanized mouse models (Tables 1 and 2) and describe their applications in a wide range of novel approaches for HIV-1 eradication (Table 3).
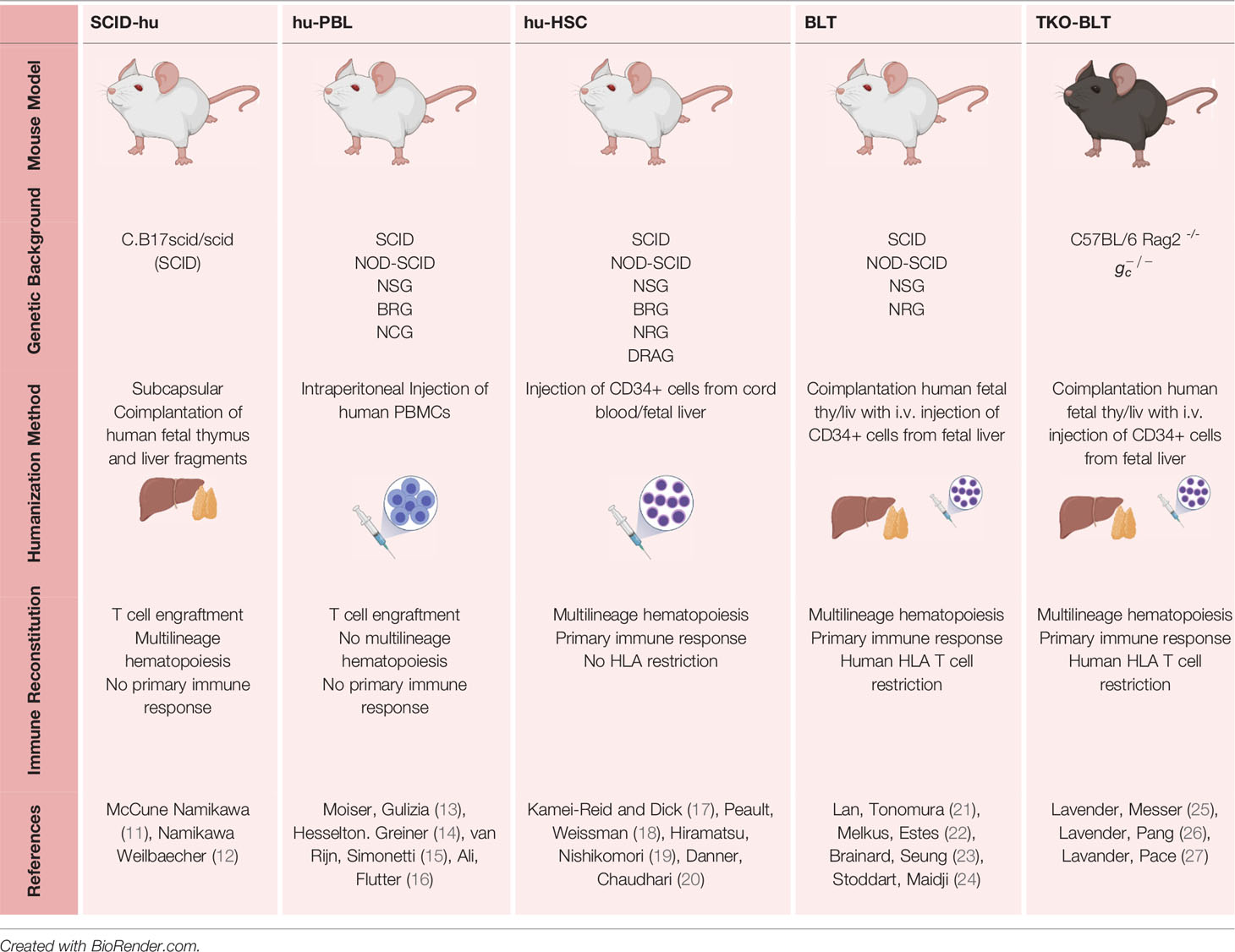
Table 1 Summary of Humanized mouse models and their tissue-based chimerism.
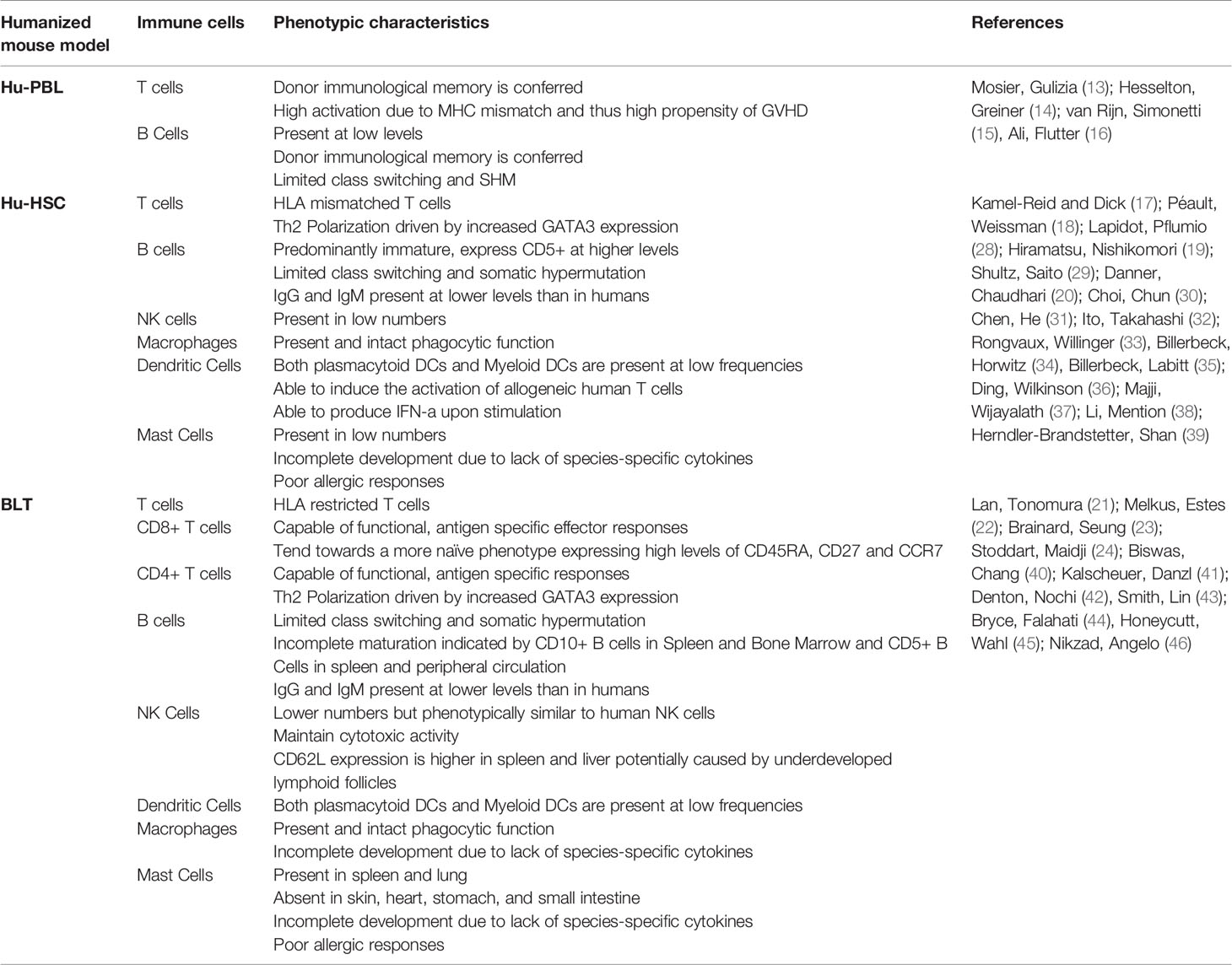
Table 2 Summary of known immunophenotypic characteristics of humanized mice.
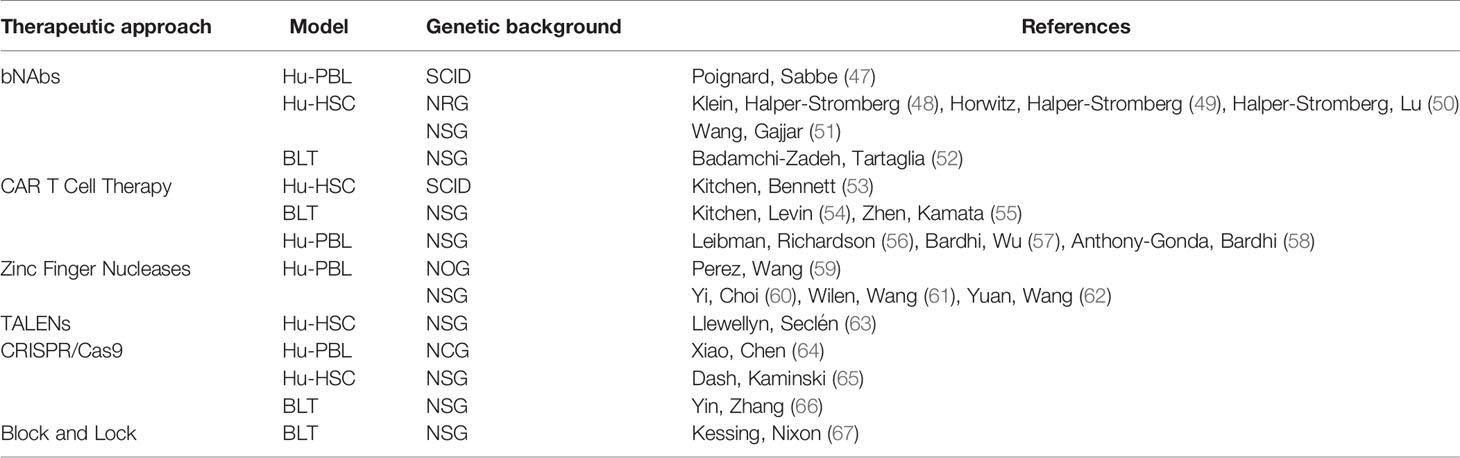
Table 3 Summary of therapeutic approaches and humanized mouse models utilized in their testing.
Humanized Mouse Models
SCID-hu Mouse
Severe combined immunodeficiency (SCID) is a debilitating disease characterized by T and B lymphocyte differentiation impairment. Those affected show high susceptibility to recurring infections from viruses, bacteria, and fungi and leads to death within the first two years of life unless treated by stem cell transplant (68). Bosma, Custer (69) observed mice of ty -65the C.B-17 strain that had impaired lymphopoiesis, caused by an autosomal recessive mutation (scid). These mice became the first mouse model of SCID, with mice homozygous for this recessive gene showing hypogammaglobulinemia and deficiency in functional T and B lymphocytes (69).
In 1988, McCune reasoned that if human hematopoietic stem cells (HSCs) and human thymus were introduced together into a mouse unable to reject them, T cell development and maturation could proceed in a fashion mimicking human physiology (11). McCune implanted C.B17 scid/scid mice (SCID mice) with human fetal thymus and injected them with human fetal liver cells, either intravenously or intrathymically (11). Mice were sub-lethally irradiated to ensure full reconstitution, as was seen previously in SCID mice implanted with long-term bone marrow cultures (70). These so-called SCID-hu mice showed human T cells in peripheral circulation at 6-7 weeks post-transplantation, but this population was no longer seen by 10-12 weeks (11). They also identified that the intravenously injected fetal liver cells could home to the implanted fetal thymus, and after 10 weeks, can be found in the peripheral circulation. If the thymus is engrafted in SCID mice alone, it eventually recedes over time. Therefore, a source of human progenitor cells needs to be provided (11, 71). Human fetal liver, in particular, is used as it is the primary site of hematopoiesis in humans between 8 to 24 weeks of gestation. The human fetal liver also contains progenitors for lymphoid, myeloid, and erythroid lineages (72) but limited numbers of mature CD3+ T cells (73). This reduces the risk of graft-versus-host disease (12) and allows human immune cells to engraft more efficiently than an adult or post-natal tissue (74).
Shortly after, Namikawa, Weilbaecher (12) implanted fetal thymus and liver concurrently. The resulting Thy/Liv mice show prolonged reconstitution with human immune cells with minimal graft-versus-host disease. These mice showed increased longevity in lymphopoiesis, even up to 15 months post-transplantation.
The SCID-hu mouse provided us with the first mouse model that could be used to study HIV-1 pathogenesis (75, 76). The first infection of a humanized mouse model (SCID-hu) with HIV-1JRCSF was in 1988, shortly after McCune had developed the model (76). McCune, Namikawa (77) utilized this model to confirm, for the first time, the efficacy of the nucleoside analog azidothymidine (AZT) as an ART for HIV-1.
While providing a great stride forward in using humanized mice in HIV cure research, SCID-hu mice had some limitations. While the rapid generation of human thymocytes and naïve T cells was possible, mature T cells are primarily restricted to the implanted thy/liver organoid. Furthermore, these mice do not produce functional immune responses that recapitulate the human immune response.
Peripheral Blood Leukocyte (Hu-PBL) Mouse
Mosier and colleagues conducted the first iteration of humanizing SCID mice by transferring human PBMCs in 1988 (13). These mice were susceptible to HIV-1 infection, with 50% of them presenting with detectable viral RNA 16 weeks post-infection (78). Infection of hu-PBL-SCID mice as early as 2 hours post reconstitution has led to productive HIV-1 infection and a dramatic decrease in CD4+ T cell numbers (79).
The establishment of non-obese diabetic (NOD) mice further progressed the field of humanized mice (80, 81). These NOD mice showed defects in the innate immune system, resulting in better engraftment when crossed with SCID mice. Further development came from crossing these so-called NOD-SCID mice with those who had the IL-2Rγ-chain null mutation (82) or truncation of the intracellular signaling domain (83). This common cytokine receptor gamma chain is required for the signaling of IL-2, IL-4. IL-7, IL-9 IL-15, and IL-21 (84–90) and its deletion completely abrogates murine Natural Killer (NK) cell development and function (82) which has shown to negatively impact engraftment of human lymphoid cells in mice (91). Modern versions of hu-PBL mice utilize NOD/SCID/IL2rγ-null (NSG) or BALB/c-Rag2null IL-2Rγnull (BRG) backgrounds as they provide better reconstitution (14–16). These mice show partial functionality (13, 78, 92, 93) but lack de novo multilineage hematopoiesis and therefore are absent of a primary immune response. They are also amenable to HIV-1 infection (14, 94, 95) and are responsive to HAART (95). This model is particularly suited to studying GVHD (16, 96, 97), simultaneously creating a significant limitation in this model’s utility for long-term HIV-1 studies.
Hematopoietic Stem Cell (HSC) Mouse
Following the early success of reconstituting human immune systems in SCID mice, Kamel-Reid and Dick (17) intravenously infused hematopoietic stem cells from human bone marrow into the SCID mouse as well as the bg/nu/xid mouse, which are deficient in natural killer responses. They found bg/nu/xid mice show higher levels of human progenitors and, in addition to T and B cells, found that macrophage progenitors can be isolated and cultured in vitro (17). It was subsequently identified that by providing these HSC humanized mice with erythropoietin, human mast cell growth factor and human IL-3 stimulated the immature cells from human bone marrow to differentiate into cells of the erythroid, myeloid, and lymphoid lineages (28).
The cell surface sialomucin-like adhesion molecule CD34 is widely accepted as a marker for human HSCs (98, 99), having showing both short-term (98) and long-term (100) colony-forming potential in vitro and allowing the differentiation of both myeloid and lymphoid cell lineages in NOD/SCID mice (101). Péault, Weissman (18) isolated CD34+ cells from human bone marrow and human fetal liver and was able to reconstitute human fetal thymus in vitro and then implanted them into SCID mice. The third source of HSCs is umbilical cord blood (UCB). High levels of HSCs can be isolated from fetal liver, bone marrow, and UCB, although they produce varying levels of lymphoid, myeloid, and erythroid progeny (102). Hao, Shah (103) showed that CD34+CD38- cells isolated from UCB have a higher cloning efficiency, proliferate more rapidly in response to cytokine stimulation, and generate more progeny than those derived from human bone marrow. Furthermore, as few as 500 CD34+CD38- cells separated from UCB were able to reconstitute NOD/SCID mice (104).
Several mouse backgrounds have been utilized for reconstitution with HSCs, including NOD/SCID (104), NOD/Shi-Scid mice (105). NOD/Shi-scid/IL2Rγnull (NOG) (106) and NSG (19, 107). McDermott et al. performed a comparative analysis of each mouse background (108). They used the current optimized methods for each mouse model, which included interfemoral injection of HSCs to all mice and anti-CD122 (IL-2Rβ) treatment for NOD/Lt-scid and NOD/Shi-scid, to inhibit NK cell activity (109). They identified that NSG and NOG mice had superior engraftment in the thymus and spleen compared to NOD/Lt-scid and NOD/Shi-scid mice. Also, NSG mice showed 1.5-fold greater engraftment in the bone marrow over NOG, NOD/Lt-scid, and NOD/Shi-scid mice (108). Finally, at limiting doses of HSCs, female NSGs showed improved engraftment compared to males (108).
Successful infection with HIV-1 through multiple routes have been demonstrated in CD34+ reconstituted NOG (110–114), BRG (115–117), NRG (118) and NSG mice (65, 119–124). They are also responsive to HAART (65, 118, 119) and have significant potential to contribute to our understanding of HIV-1 latency and therapeutic studies.
DRAG Mouse
Impaired T cell development and consequent lack of antibody class switching in HSC humanized mice is mainly attributed to the lack of donor matched HLA molecules in the mouse thymus. Lack of a thymic environment hampers the negative selection of self-reactive T cells by which autoimmunity is prevented. Danner, Chaudhari (20) hypothesized that by expressing human HLA class II molecules in a transgenic mouse model, they could rescue the development of CD4+ T cells and consequently B cell development and antibody class switching. They generated NOD.Rag1KO.IL2RccKO mice expressing HLA-DR4 (0401), abbreviated DRAG, and at 4-6 weeks old intravenously injected them with CD34+ HSCs isolated from HLA-DR*0401 positive UCB (20). When compared against HLA mismatched recipients, higher levels of reconstitution of CD4 + T cells were seen in DRAG mice comparable to human blood levels. However, levels of CD8+ T cells did not show as drastic an increase. These mice showed the presence of dendritic cells (2.9% in spleen) and NK cells (0.05% in spleen), although their development appeared to be unaffected by HLA-DR4 expression. T cells isolated from DRAG mice showed vigorous responses, similar to that of PBMCs from healthy volunteers upon stimulation with either CD3/CD28 or PMA/ionomycin. B cell reconstitution was seen at similar levels to control mice, although IgM levels were significantly higher in DRAG mice. Interestingly, DRAG mice showed substantial IgG reconstitution, confirming that the mechanism of immunoglobulin class switching is rescued in DRAG mice, a feature lacking in most other humanized mouse models. Further, DRAG mice showed reconstituted plasma levels of all human IgG subclasses, with IgG2 being the most prevalent.
Studies have shown that DRAG mice are susceptible to HIV-1 infection, similar to other HSC reconstituted mice (125, 126). The replication-competent virus was isolated from plasma, lymph nodes, bone marrow, spleen, gut, brain, and female reproductive tissue upon a single intravaginal challenge of purified primary HIV-1 (125). High reconstitution in the gut and female reproductive tract, particularly with CXCR5+PD-1++ Follicular T helper cells, which are highly permissive to HIV-1 infection, are potentially a significant contributor to the above (126). Also, plasma viral loads were stable as far as 18 weeks post-infection, making the DRAG mouse a suitable model system, particularly for long-term vaccination studies.
Bone Marrow-Liver-Thymus (BLT) Mouse
While implantation of Thy/Liv into SCID or NOD/SCID mice results in thymopoiesis and functional T cells, strong in vivo immune responses are not observed. In 2006 (21), Lan et al. combined the implantation of fetal Thy/Liv with the simultaneous transplantation of CD34+ fetal liver cells in NOD/SCID mice (21). In the same year, Melkus, Estes (22) performed a similar transplant on the NOD-SCID background, coining the term ‘BLT’ to describe these bone marrow-liver-thymus humanized mice (22). A significant advantage of the BLT model is that it allows the development of MHC-restricted T cells due to the presence of an autologous human thymic environment (22). This iteration of the humanized mouse showed stronger in vivo immune responses and repopulates with multiple lineages of immune cells, including T cells, B cells, NK cells, Dendritic Cells, Neutrophils, and Monocytes distributed through multiple organs, including bone marrow, lymph nodes, spleen, thymus, liver, lung, digestive and reproductive tracts (21–23, 42, 43, 46, 127, 128).
Soon after its development, Sun, Denton (128) identified that BLTs contain CD4+ T cells throughout the gut-associated lymphoid tissue (GALT), including the colon and rectum, and thus hypothesized that they would be ideal for modeling intrarectal transmission, a predominant form of HIV-1 transmission. BLT mice were inoculated intrarectally with a single dose of cell-free HIV-1 (LAI CXCR4 strain), and six out of seven infected BLTs were found positive for viral RNA and the presence of p24 antigen (128). Similarly, Denton, Estes (127) showed that BLTs were susceptible to vaginal transmission of HIV-1 (CCR5-tropic JR-CSF). Infection was prevented in BLTs pre-treated with a daily dose of the ARTs emtricitabine and tenofovir diosoproxil fumarate (FTC/TDF), making the BLT model additionally suited for preclinical testing of pre-exposure prophylactics (PrEP). Subsequent studies of topically administered 1% tenofovir in BLT mice also showed partial protection against vaginal transmission of HIV-1 (129).
Brainard, Seung (23) infected BLT mice made with mice of both NOD/SCID and NSG backgrounds, implanting tissue under both right and left kidney capsules. Stoddart, Maidji (24) also demonstrated that NSG reconstitution was highly superior to NOD/SCID reconstitution (24). However, Denton, Nochi (42) showed higher levels of intraepithelial lymphocytes in the small and large intestines of NOD/SCID BLTs.
The functional human cellular immune response observed in BLTs makes it a particularly valuable model for studying HIV specific immunity. HIV infection in BLT humanized mice is associated with both CD8+ T cell activation (23, 130, 131) as well as HIV-specific IgM and IgG (23, 40). HIV-1 specific CD8+ T cell responses show strong similarities to human cellular immunity and result in rapid viral escape in BLT mice (131). Furthermore, mice created with tissue from elite controllers that express the HLA-B*57 allele exhibited enhanced Gag mediated control of viremia (131).
While substantial reconstitution with B cells is observed in BLT mice, they are considered primarily immature, and antibody class-switching is thought to be defective. Interestingly, it has been speculated that the humoral immunity observed in BLT mice in response to viral infection is driven by extra-follicular or ‘innate-like’ B cells rather than conventional post-germinal B cells (40, 132, 133). This inadequate antibody response has led to the proposal of the BLT mouse as a model for hypogammaglobulinemia (134).
Crucial to HIV-1 therapeutic studies is the establishment of latency in the BLT mouse model. Denton, Olesen (135) used a commonly used combination ART regimen, a combination of tenofovir, emtricitabine, and raltegravir, to suppress viral replication in BLT humanized mice, and thereafter isolated CD4+ T cells and cultured them ex vivo. Stimulation of these CD4+ T cells with Phytohemagglutinin and IL-2 led to a rebound in viral load (135).
Triple Knockout (TKO)-BLT
While modern BLT and humanized mouse models primarily use NOD/SCID backgrounds, there is the potential benefit of developing a BLT model on the C57BL/6 background due to the wide availability of transgenes and genes inactivation as well as its relative resistance to radiation. A significant barrier to this is the expression of a different form of the signal recognition protein α (SIRPα) receptor by C57BL/6 mice, which does not recognize the human ligand CD47 (integrin associated protein) unlike in NOD mice (136, 137). Recognition of CD47 on transplanted human cells by SIRPα on mouse macrophages leads to the transmission of inhibitory signals preventing their phagocytosis (138, 139). The Hasenkrug lab demonstrated that creating C57BL/6 Rag2-/-γc-/- (TKO) facilitated a CD47-negative environment, which leads to tolerizing of cells without the need for the SIRPα-CD47 interaction, thereby allowing successful long-term human immune systems in TKO-BLT mice (25, 26).
The TKO-BLT mice showed no clinical signs of graft-versus-host disease (GVHD) up to 29 weeks post-transplantation compared to NSG-BLT mice, of which a third had died from lethal GVHD at the same timepoint (26). Furthermore, TKO-BLT mice developed human GALT, including CD4+ T cells, supporting mucosal HIV infection through intraperitoneal and intrarectal routes (26). HIV-1 infected TKO-BLTs showed HIV gp120JR-CSF specific IgG antibodies demonstrating both antibody responses and T-cell dependent class switching (26). Furthermore, cells from the spleens of HIV infected TKO-BLTs were assayed by IFN-γ ELISPOT and were shown to be responsive to a broad range of peptide pools spanning the HIV-1JR-CSF proteome (26). Similar to NSG-BLT mice, the TKO-BLT can also recapitulate HIV-1 latency upon treatment with ART up to 18 weeks, with viral rebound seen upon removal of therapy (27). GVHD is a limitation in the BLT humanized model (140). Resistance to GVHD makes the TKO-BLT particularly suited to long-term studies required to study HIV-1 latency and measure the lasting efficacy of current curative approaches.
Humanized Mice in HIV-1 Cure Research
The primary barrier to eradicating HIV-1 is its integration into the host genome and continued persistence in a non-replicating or ‘latent’ state, even under ART (141–143). Furthermore, chronic infection with HIV-1 leads to significant disruptions in the host’s immune response (144). Consequently, latently infected cells are difficult to find, and the host immune response is inefficient in killing these cells. Therefore, innovative strategies are required to find, measure, and ultimately eradicate the latent HIV-1 reservoir.
Two definitions exist for a cure for HIV. The complete eradication of the virus from all cells in the body is defined as a sterilizing cure. In contrast, a functional cure would be a treatment that would lead to HIV+ people being able to halt ARV treatment while maintaining long-term viral suppression and preventing transmission of the virus. The humanized mouse models described in this review have played a vital role in pre-clinical testing of many approaches to an HIV-1 cure, with some directly leading to clinical trials.
Latency Reversal
One major strategy in the fight for a sterilizing cure is the “shock” and “kill” approach. This strategy involves utilizing latency reversal agents to ‘shock’ the latently infected cells into resuming replication of the virus, allowing the immune system to recognize and kill infected cells or for virus-induced cytolysis to occur. Several classes of LRAs have been identified which include histone deacetylase inhibitors (HDACis) (145–147), histone methyltransferase (HMT) inhibitors (148), DNA methyltransferase inhibitors, Protein Kinase C (PKC) activators (149), bromodomain inhibitors (150), Disulfiram (a drug used to treat chronic alcoholism) (151), agonists to Toll-like receptor 7 (152) and cytokines such as IL-15 (153).
One of the most well-studied classes of LRAs, HDACi’s work by inhibiting the enzymes that remove acetyl groups from histones, leaving them in an open state, and increasing the ability for transcription to resume. While HDAC inhibitors such as Panobinostat successfully increase levels of histone acetylation in vivo (154) and reactivation of latently infected cells ex vivo (149, 155), reactivation of HIV-1 replication was not seen in ART-treated HIV-1 infected BLT mice (154).
Other classes of LRAs impact cellular factors such as NF-κB and pTEFB by either increasing their expression or activating them. Protein kinase C (PKC) agonists are one such class that activates the NF-κB pathway leading to reactivation of latent HIV-1 (156, 157). Prostratin, a PKC agonist, was one of the earliest LRAs of this class identified with potency in reactivating latent HIV-1 (158). Additionally, IDB and Bryostatin 1 can effectively reactivate viruses in cells isolated from HIV-infected patients (149, 159). Marsden, Loy (160) identified a synthetic analog of the PKC modulator bryostatin 1, SUW133, which when intraperitoneally injected into ART-treated HIV-1 infected BLT mice, reactivated HIV-1 more potently, was more tolerable than the natural analog, and led to infected cells’ death.
Bobardt, Kuo (161) showed that an inhibitor apoptosis protein antagonist (IAPa) reactivates latent HIV-1 by degrading the ubiquitin ligase BIRC2, a repressor of the non-canonical NF-κB pathway. Substantial reactivation was seen ex vivo in latently infected CD4+ T cells isolated from ART-treated HIV-1 infected BLT mice at much greater levels than the HDACis panobinostat and vorinostat as well as another IAPa LCL-161 (161). Similar results were seen in vivo with AZD5582, which also works as a SMAC mimetic and activates the non-canonical NF-kB pathway (162). Upon a single dose of 3mgkg-1 of AZD5582 HIV-1 RNA was detected in 3 out of 6 and 3 out of 4 ART-treated HIV-1 infected BLT mice, with no signs of general toxicity (162).
Specific cytokines have dual potential to both reactivate HIV-1 replication and simultaneously activate host immune cells such as NK cells to kill infected cells (153, 163, 164). Recently, the IL-15 ‘super-agonist’ N-803 tested in the BLT humanized model showed reactivation of the viral reservoir only upon depletion of CD8+ T cells, showing further nuance in the interactions between LRAs, viral reactivation, and immune cells (165).
Interestingly, Llewellyn, Alvarez-Carbonell (166) found that HSC mice (NSG fetal liver CD34+) reconstitute with human microglial cells in the brain when endogenous microglia are depleted using the chemotherapeutic agent busulfan at much greater levels than mice pretreated with irradiation. These human-derived microglial cells could be infected with HIV-1 and, upon exposure to the MAO inhibitor phenelzine, they could reactivate latent virus ex vivo (166). These results further emphasize the potential for humanized mouse models in studying latency reversal in multiple tissue reservoirs of HIV-1.
Unfortunately, while many LRAs have successfully reactivated HIV, the subsequent killing by viral cytopathic effects, killing by host immune cells, or intensifying ART has proven ineffective in the clinic (167). This is partly due to dysfunctional CTLs (168) and NK cells (169–171) and due to the presence of escape mutant viruses in the latent reservoir (172). Therefore, to enhance the ‘kill’ arm, multiple immunotherapies, gene therapies, and pharmacological interventions have been explored.
Broadly Neutralizing Antibodies (bNAbs)
The HIV-1 envelope protein (env) is a trimeric glycoprotein of gp41 and gp120 heterodimers and is on the virus’s surface and consists of multiple sites that can be targeted by host antibodies. The majority of neutralizing antibodies that develop in humans are strain-specific, but a small subset of antibodies can bind to and neutralize a broad range of HIV-1 strains (173–176). Initial attempts to utilized these broadly neutralizing antibodies (bNAbs) in HIV-1 therapy proved unsuccessful in both humanize mice (47) as well as in the clinic (177, 178). However, with the advent of single-cell antibody cloning methods, highly potent bNAbs have been identified (179). Over the past decade, humanized mice have proved instrumental in developing therapies utilizing bNAbs for viral suppression and eradicating latently infected cells.
Klein, Halper-Stromberg (48) showed that treatment with a combination of bNAbs (45-46G54W, PG16, PGT128, 10-1074, and 3BC176) that target different epitopes could suppress viral replication in HIV-1 infected hu-HSC mice. These humanized mice were then infected with HIV-1YU2, and treated with a penta-mix treatment therapy reducing viral loads to undetectable levels for up to 60 days post removal of therapy compared to only 10 days when treated with ART. Shortly after, Horwitz, Halper-Stromberg (49) showed in the same mouse model that lowering the viral load by ART followed by treatment with bNAb monotherapy (3BNC117, PG16, or 10-1074) can similarly lead to control of viremia until antibody titers dropped to low or undetectable levels. Also, a single injection of adeno-associated virus directing expression of a single bNAb suppressed viral replication after the termination of ART (49). Following these studies, Halper-Stromberg, Lu (50) hypothesized that these capabilities of bNAbs could be harnessed to amplify the “kill” arm with bNAb therapy after reactivation of the latent reservoir. They used three LRAs; the HDACi Vorinostat, the BET protein inhibitor I-BET151, and CTLA, a T cell inhibitory pathway blocker given individually or in combination (50). They discovered that combination treatment with the three LRAs with the trimix bNAb treatment (3BNC117, PG16, or 10-1074) in humanized mice reduced the viral reservoir, evidenced by reduced viral rebound at significantly higher levels than with single LRAs (50). They further showed that bNAbs exert this effect primarily through Fc mediated functions by comparing their bNAbs to those with mutated Fc regions (50).
These results provided the preclinical evidence that combination bNAb therapy is a critical tool that can be utilized in HIV-1 eradication and has since led to the translation of these ideas to the clinic (180). In an open-label clinical trial, Caskey, Klein (181) showed that monotherapy with a single 30mgkg-1 infusion of 3BNC117, the antibody targeting the CD4 binding site, was able to suppress viral load in HIV-1 infected individuals for up to 28 days. Also, in the following phase II trial, the same therapy suppressed HIV-1 viral load in infected individuals on average for 10 weeks, upon four doses given two weeks apart (182). Further investigation by adoptive transfer of patient T cells into NRG mice identified that these effects are not limited to preventing new infection of cells, but also aids in the clearance of infected cells by Fcγ receptor-mediated engagement (183). Shortly after, Caskey, Schoofs (184) tested 10-1074, which targets a glycan on the V3 loop of the HIV-1 envelope spike, as monotherapy in HIV-1 infected patients. Similarly, at a dose of 30mgkg-1, 11 out of 13 HIV-1 positive individuals showed suppression of viral loads. The two that failed to respond carried mutations leading to single-amino-acid changes, making them 10-1074 resistant before treatment (184). Unfortunately, while monotherapy with bNAbs was proven to be safe and efficacious, in both cases, it led to viral escape (181, 183, 184).
In contrast to monotherapy, but similar to the results in humanized mice, combination therapy with 3BNC117 and 10-1074 at a dose of 30mgkg- was found to be both safe and more effective at viral suppression in humans (185–187). When administered upon treatment interruption three times at three weeks apart, 11 HIV-1 infected individuals resulted in viral suppression for a median of 21 weeks post final dose of antibody therapy (186). Furthermore, none of the rebounding viruses showed resistance to both therapeutic bNAbs. Similar to data generated in humanized mice, these data demonstrate the enhanced utility of combination bNAb therapy to suppress viral replication (186). Moreover, combination bNAb therapy during treatment interruption led to increases in antigen-specific CD8+ T cells that expressed IFN-γ. TNF-α, MIP1-β, and CD107a in all individuals at 6-7 weeks while plasma bNAb levels were at their peak (188). While latency reversal agents combined with bNAB therapy have yet to be tested in the clinic, the evidence provided so far shows a clear line from preclinical experiments in humanized mice to the successful translation of these therapies into the clinic.
Chimeric Antigen Receptor (CAR) T Cell Immunotherapy
Since the advent of CAR T cell therapies and their successful use in treating B cell malignancies (189–191), several approaches have been taken to utilize this technology to target HIV-1 infected cells.
Kitchen, Bennett (53) transduced HSCs isolated from human fetal liver with an HLA-A*0201 restricted T cell receptor (TCR) targeted towards the HIV-1 gag SL9 epitope and injected them into human thymic implants in SCID-hu mice, providing them with an optimal environment for development. This led to the development of anti-HIV TCR+ CD8+ T cells, which produced Interferon-γ (IFN- γ) and lyse target cells in response to SL9 peptide stimulation ex vivo when biopsied from the thymus after seven weeks (53). In a subsequent study Kitchen, Levin (54) transduced HSCs with the same vector. They tested the ability of these anti-HIV TCR+ CTLs to suppress viral replication in a modified NSG-BLT model, which received a fetal thymus/liver implant along with the transduced HSCs. They found a significant decrease in plasma viremia at two weeks and six weeks and a higher percentage of CD4+ cells in mice transduced with the HIV-1 specific TCR when compared against a non-specific TCR control (54). Similar results were seen upon analyses of other organs of infected mice, with lower HIV-1 DNA at six weeks in the spleen, bone marrow, and human thymus (54). Importantly, their analysis of the viral RNA in the blood showed that within this time, no viral escape had occurred in response to the selective pressure of the SL9 specific TCR (54).
While modification with molecularly cloned anti-HIV TCRs showed to be effective, its application is restricted by HLA type, and several effective HIV CTLs use uncommon HLA alleles (192). A second alternative approach is a CD4ζ CAR, a chimeric molecule consisting of the extracellular and transmembrane domains of the human CD4 molecule fused to the signaling molecule of the CD3 ζ chain. Zhen, Kamata (55) transduced human HSPCs with an HIV-specific CD4ζ CAR and two antiviral genes (Triple CAR vector) and transferred them into NSGs transplanted with fetal thymus and liver (55). The antiviral genes included a small hairpin (sh) RNA molecule specific to human CCR5 and shRNA targeting specific HIV-1 long-terminal repeat (LTR) sequences to prevent the newly CD4 CAR-expressing cells from being infected by HIV-1, an obstacle previously observed with this approach (55). The Triple CAR construct was expressed on T cells, NK cells, B cells, and myeloid cells (55). They found these cells are resistant to HIV infection themselves, suppressed HIV replication in vivo, and isolated CD4ζ CAR-expressing cells could produce IFN- γ and tumor necrosis factor (TNF)-α when cultured with virally infected cells (55).
While CD4ζ CARs showed promise in vitro and in preclinical studies, clinical trials showed that while they survive for at least 11 years post-transfusion, they could not suppress the viral reservoir in a sustained manner (193). Therefore, Leibman, Richardson (56) decided to identify several components of the CD4ζ CAR construct to be optimized to improve their antiviral function. First, they switched from a murine retroviral vector (MMLV) to a lentiviral vector, as MMLV targets promoter regions while lentiviral vectors preferentially integrate into open reading frames (56). Next, they switched from the PGK promoter to the EF1α promotor, which induces higher expression better sustained as T cells reach a resting state (56). Thirdly, they swapped the CD4 transmembrane domain to a CD8a transmembrane domain to promote CAR dimerization (56). Finally, they assessed the inclusion of different co-stimulatory receptors such as 4-1BB and CD28 (56). While each modification individually produced significant improvements to target cell killing in vitro, in combination, they resulted in a 50-fold increase in potency over the original construct (56). To test their optimized CD4ζ CARs in vivo, they utilized a modified hu-PBL mouse as an HIV-1 treatment model. Briefly, they transferred CD8+ T cell-depleted PBMCs into NSGs, transfused them with HIV-1 infected CD4+ T cells, treated with 200mg/kg TDF for four days, and after removal of ART, transfused the mice with their CD4ζ CAR transduced CD8+ T cells (56). They found that mice infused with the optimized CD4ζ CARs T cells controlled HIV-1 replication better and expanded to greater levels in vivo than their first-generation counterparts (56).
In addition to CD4ζ, CARs with a single-chain variable fragment (scFv) derived from bNabs have been utilized. However recent clinical trials using bNAb monotherapy led to viral rebound upon interruption of ART (181, 184, 194). Bardhi, Wu (57) showed that a hexavalent fusion protein which consisted of m36.4, the scFv heavy chain only domain which targets the gp120 co-receptor binding site and mD1.22 an engineered mutant of the CD4 extracellular domain when intravenously injected into a modified hu-PBL model induced NK cell-mediated killing of HIV-1 infected cells. The model they used involved injecting human PBMCs intrasplenically (hu-spl-PBMC-NSG) into NSG mice followed by intrasplenic inoculation with HIV-1 (57, 58, 164). Subsequently, Anthony-Gonda, Bardhi (58) utilized m36.4, mD1.22, and a fusion inhibitor peptide C46, individually or in combination, to develop novel multi-specific CAR constructs. Using their hu-spl-PBMC-NSG model, they discovered that their duoCAR construct, by which m136.4 and mD1.22 were expressed as individual CARs on the same cell, was able to suppress HIV-1 infection for up to 30 days while mitigating CD4+ T cell depletion, compared to their monoCAR or untransduced counterparts (58). Altogether, these findings clearly show the utility of various humanized mouse models in elucidating CAR-based immunotherapy in vivo efficacy in the clearance of HIV-1 infected cells.
Gene Editing With Designer Nucleases
The first patient cured of HIV to date was treated for myeloid leukemia with total body irradiation and two allogeneic hematopoietic stem-cell transplants (HSCT) from donors with the CCR5Δ32/Δ32 mutations (195). While the second patient exhibiting HIV-1 remission reported in 2019 received no radiation and only one HSCT transplant (196). These treatment methods are not feasible for the wide-scale treatment of otherwise healthy HIV-positive people. However, these successes have inspired the gene-editing field to target the CCR5 gene to induce resistance to HIV-1 infection (at least through CCR5 tropic HIV-1).
To date, three major types of nucleases are used for genome editing, namely, zinc finger nucleases (ZFN), transcription activator-like effector nucleases (TALEN), and the Cas9 endonuclease with clustered regularly interspaced short palindromic repeats (CRISPR). All three utilize a similar overall mechanism of binding a target DNA sequence, creating a double-stranded break allowing the DNA to undergo repair through the error-prone non-homologous end joining (NHEJ), resulting in small insertions or deletions disrupting gene expression [reviewed in (197)].
ZFN consist of pairs of zinc finger DNA binding domains that recognize three base pairs of DNA and are covalently linked to complementary halves of a FokI restriction endonuclease. ZFNs have been utilized to confer resistance to HIV-1 infection in resting CD4+ T cells through the disruption of the CCR5 gene in humanized mouse models (59, 60). Similarly, Holt, Wang (123) showed human HSCs in which CCR5 was disrupted by ZFN, were able to engraft in NSG mice successfully and led to the development of CCR5-tropic HIV-1 resistant cells. Also, ZFNs targeting the CXCR4 co-receptor on CD4+ T cells have shown success in protecting from HIV-1 infection in humanized mice, although resistance was eventually lost upon selection for CCR5-tropic mutants (61, 62). Following these promising results in humanized mice, the first phase 1 human clinical trial with ZFN-CCR5 mutated CD4+ T cells was conducted, showing that these genetically modified CD4+ T cells were more resistant to HIV-1 infection and led to decrease viral loads during ART interruption (198).
The gene-editing field was revolutionized by the discovery of CRISPR/Cas9 in prokaryotes (199, 200) and its role in bacterial and archeal adaptive immunity against invading viruses (201). Unlike ZFNs, CRISPR/Cas9 relies on short sequences of guide RNA (gRNA) to target the Cas9 nuclease to the complementary DNA sequence to be edited. This feature of CRISPR/Cas9 makes it more sequence-specific and easier to design and produce, leading to its first use in gene editing by Jinek, Chylinski (202), a landmark discovery leading to a Nobel prize award in chemistry in 2020. Not long after, CRISPR/Cas9 was used to target the HIV-1 LTR in human cell lines resulting in a significant decrease in HIV-1 expression upon stimulation (203). Recently, CRISPR with Staphylococcus aureus Cas9 (SaCas9) delivered by a lentiviral vector improved primary CD4+ T cell resistance to HIV-1 infection. Also, transplanting these cells into NCG humanized mice resulted in enhanced survival upon challenge with CCR5 tropic HIV-1YU2 (64).
In addition to targeting host factors, another gene-editing approach that has picked up steam is to directly target the integrated HIV-1 provirus for excision (203–205). This approach has been used to remove the entire HIV-1 genome between the 5’ and 3 LTRs from latently infected human CD4+ T cells (206). Humanized mice have shown to be invaluable in testing these approaches in a model that accurately recapitulates HIV latency, as is seen in humans. Yin, Zhang (66) discovered that in vivo excision of the HIV-1 provirus with a SaCas9 with multiplex single-guide RNAs (sgRNA) was successful in a small cohort of BLT mice. A TALEN targeting the TATA-box of the HIV-1 LTR has also been used successfully to clear HIV-1 ex vivo on splenocytes derived from humanized mice (63).
Recently the Gendelmann lab combined long-acting slow-effective release (LASER) ART treatment with consequent CRISPR/Cas9 treatment to excise HIV DNA (65). They showed in 3 donor cohorts of hu-HSC mice, over a third 9/23 of the mice showed no rebound of the infectious virus following combination treatment than LASER ART or CRISPR/Cas9 treatment alone (65). Multiple organs, including the spleen, bone marrow, gut, brain, liver, kidney, and lung of these mice were assessed and showed no viral rebound (65). This data provides a solid foundation for the efficacy of these gene-editing approaches in vivo and can pave the way for human clinical trials.
Block and Lock
While the therapeutic approaches discussed so far have focused on the complete eradication of HIV-1 reservoirs, the Valente lab took an alternate approach to lock HIV-1 in its latent state by targeting the HIV-1 Tat protein (67, 207). Tat is transcribed early in the HIV-1 lifecycle and recruits the necessary transcription factors to enhance viral transcription and stabilize elongation (208). Specifically, Tat binds the 5’ terminal region of the transactivation response element (TAR) (208) on HIV mRNA and recruits the positive transcription elongation factor B (PTEF-b) (209). PTEFb is composed of cyclin T1 and cyclin-dependent kinase 9 (CDK9), promoting transcription elongation from the viral promoter (210). Also, Tat has shown to recruit chromatin remodeling factors such as SWI/SNF (211, 212) and histone acetyltransferases (213–215) allowing chromatin to remain in an open state and thus allowing easier access to transcription factors such as nuclear factor-kB (NFkB).
Targeting viral factors such as Tat is a major focus of antiretroviral therapy research, mainly due to the absence of a host cellular homolog, resulting in less toxicity. The Valente Lab discovered that didehydro-cortistatin A (dCA), a synthetic analog of the natural product cortistatin A, potently inhibits Tat-dependent transcription activity in both acutely and chronically infected cells (216). dCA was also found to act additively with conventional HAART, inhibiting spontaneous viral particle release from CD4+ T cells in virally suppressed subjects (216). Soon after that, they showed that dCA effectively inhibits viral reactivation by a PKC agonist or by antigenic stimulation of primary latently infected cells isolated from ART-treated individuals (207). Furthermore, they demonstrated that dCA, unlike conventional ART, can reduce the base level cell-associated HIV-1 RNA production by decreasing RNA Polymerase II recruitment to the viral promoter (207). Primary CD4+ T cell cultures alone cannot fully capture the characteristics of latently reservoirs, as these use clonal HIV strains and specific cytokine cocktails to prolong lifespan. These conditions may transform these cells and alter cell-subset representation. Therefore, in a subsequent study, Kessing, Nixon (67) tested the ability of dCA to suppress viral reactivation in a BLT mouse model. Humanized mice were infected with HIV-1JRCSF and treated with ART for three weeks, at which point dCA was combined with ART for the fourth week (67). Strikingly, upon treatment interruption, viral rebound was delayed up to 19 days in mice treated with dCA compared to the controls, which showed rebound between 3-7 days upon treatment interruption (67). Unfortunately, certain viruses develop resistance to dCA through a combination of mutations in Nef and Vpr that increase NF-κB activity and lead to a higher Tat-independent basal transcription (217). The discovery of additional transcriptional inhibitors that promote deep latency through different mechanisms, used in combination with dCA could be the way forward for the block and lock strategy. In vivo analysis in humanized mouse models will be critical in their translation to the clinic.
Limitations of Humanized Mice
Despite the fact that they are an advantageous small animal model for HIV-1 cure research, humanized mice have their limitations. Hu-PBL mice and, to a lesser extent, hu-HSC and BLT mice occasionally develop graft-versus-host disease. GvHD is characterized by lymphocytic infiltration, progressive inflammation, and sclerosis in multiple organs, eventually leading to death (140). Moreover, the associated excessive inflammation can confound results from long-term HIV-1 studies. The use of NSG mice over NOD/SCID mice for the construction of BLT humanized mice was found to delay the onset of GVHD, although it did not decrease its incidence (140). Furthermore, even with many advances over the years, here remain deficiencies in the immune response of humanized mice compared to a complete human immune system. First, they lack robust humoral immune responses due to limited class switch recombination and mutation rates, limiting the study of B cell responses, particularly in vaccination studies (23, 134). Also, hu-PBL, and hu-HSC mice lack human HLA restriction of T cells, leading to deficiencies in the development and functionality of T cells, a significant advantage of the BLT model.
On the other hand, the construction of BLT mice requires significant technical expertise to produce, and access to fetal tissue for scientific research continues to become scarce. Moreover, the murine environment lacks human cytokines and poor cross-reactivity with murine cytokines leads to incomplete development of some human myeloid and lymphoid cell subsets. Several solutions for this include treatment with exogenous human cytokines. For instance, periodic intraperitoneal injection of human IL-15/IL-15Rα is used to support the proliferation and survival of NK cells (46, 218). Other strategies involve knock-in and transgenic human cytokine insertions into immunodeficient mice. For example, human IL-2 and IL-15 transgenics (219), IL-15 and SIRPα knock-in (39), and IL-7 and IL-15 knock-in mice (220) promote NK cell development in humanized mice. Also, mice that comprise multiples gene modifications to produce the cytokines macrophage-colony stimulating factor (M-CSF), IL-3, granulocyte-colony stimulating factor (GM-CSF), and thrombopoietin have been established to support monocytes and macrophage development (33). While these modified humanized mouse models show potential in producing a more comprehensive innate immune system, their utility in HIV-1 persistence and therapeutic studies has yet to be evaluated.
Another limitation to long term persistence studies is the limited lifespan of mice (221), making studies lasting several years impractical. In these cases, an NHP model would be more suitable. Even though small animals are easier to house and maintain, they bring the additional constraint of having lower blood volumes and cell numbers for analyses, making ex vivo assays technically challenging to perform. Furthermore, the murine basal metabolic rate is different from that of humans and needs to be considered, particularly when dealing with pharmacological approaches (222). Lastly, the murine gut microbiome differs significantly from that of humans. This important topic has been reviewed in detail elsewhere (223, 224). The microbiome has several implications in HIV-1 infection, pathogenesis, and the host immune response [reviewed in (225–228)], and the microbiomes of inbred mice in a controlled environment aren’t capable of fully recapitulating these effects. To skew the microbiome to resemble that seen in humans, Daharsh, Zhang (229) treated BLT mice with broad-spectrum antibiotics to deplete the murine gut microbiome, followed by fecal transplants from healthy human donor samples via oral gavage. While this approach shifted the murine microbiome towards the corresponding human donors, they didn’t wholly resemble a human gut microbiome. Future developments may improve this premise, leading to a more comprehensive mouse model mimicking both a human immune system and the human gut microbiome.
Conclusion
Regardless of its limitations, the humanized mouse is currently the only small animal model for HIV-1 and has proved to be an incredible resource in many facets of HIV-1 research in the past four decades. Large cohorts of mice can be generated from tissues derived from a single human donor, resulting in animals with genetically identical human immune cells. This approach allows us to control genetic variables and perform specific experiments such as adoptive transfers of lymphocytes, which would not be possible in most NHPs. Furthermore, humanized mice do not require a surrogate virus for infection and are susceptible to multiple routes of HIV-1 transmission. Moreover, tissue reservoirs harboring latent viruses can be analyzed, allowing a more straightforward comparison of the systemic effects of administered therapeutics, which is exceedingly essential for HIV-1 eradication efforts. Humanized mice have been critical in HIV-1 research, and we expect they will continue to play a significant role in ultimately translating a cure from bench to bedside.
Author Contributions
SA and SP conceptualized the content of and wrote the article. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by NIH RO1 AI116282 (SP), and unrestricted funds from The Scripps Research Institute, La Jolla, CA (SP).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Barre-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Sci (N Y NY) (1983) 220(4599):868–71. doi: 10.1126/science.6189183
2. Arts EJ, Hazuda DJ. HIV-1 antiretroviral drug therapy. Cold Spring Harb Perspect Med (2012) 2(4):a007161–a. doi: 10.1101/cshperspect.a007161
3. Apetrei C, Pandrea I, Mellors JW. Nonhuman primate models for HIV cure research. PloS Pathog (2012) 8(8):e1002892–e. doi: 10.1371/journal.ppat.1002892
4. Evans DT, Silvestri G. Nonhuman primate models in AIDS research. Curr Opin HIV AIDS (2013) 8(4):255–61. doi: 10.1097/COH.0b013e328361cee8
5. Van Rompay KKA, Tackling HIV. and AIDS: contributions by non-human primate models. Lab Anim (2017) 46(6):259–70. doi: 10.1038/laban.1279
6. Sun M, Li Y, Yuan Z, Lu W, Kang G, Fan W, et al. VRC01 antibody protects against vaginal and rectal transmission of human immunodeficiency virus 1 in hu-BLT mice. Arch Virol (2016) 161(9):2449–55. doi: 10.1007/s00705-016-2942-4
7. Pardi N, Secreto AJ, Shan X, Debonera F, Glover J, Yi Y, et al. Administration of nucleoside-modified mRNA encoding broadly neutralizing antibody protects humanized mice from HIV-1 challenge. Nat Commun (2017) 8:14630. doi: 10.1038/ncomms14630
8. Baum MM, Ramirez CM, Moss JA, Gunawardana M, Bobardt M, Gallay PA. Highly synergistic drug combination prevents vaginal HIV infection in humanized mice. Sci Rep (2020) 10(1):12995. doi: 10.1038/s41598-020-69937-5
9. Denton PW, Garcia JV. Mucosal HIV-1 transmission and prevention strategies in BLT humanized mice. Trends Microbiol (2012) 20(6):268–74. doi: 10.1016/j.tim.2012.03.007
10. Tager AM, Pensiero M, Allen TM. Recent advances in humanized mice: accelerating the development of an HIV vaccine. J Infect Dis (2013) 208Suppl 2(Suppl 2):S121–S4. doi: 10.1093/infdis/jit451
11. McCune JM, Namikawa R, Kaneshima H, Shultz LD, Lieberman M, Weissman IL. The SCID-hu mouse: murine model for the analysis of human hematolymphoid differentiation and function. Sci (N Y NY) (1988) 241(4873):1632–9. doi: 10.1126/science.2971269
12. Namikawa R, Weilbaecher KN, Kaneshima H, Yee EJ, McCune JM. Long-term human hematopoiesis in the SCID-hu mouse. J Exp Med (1990) 172(4):1055–63. doi: 10.1084/jem.172.4.1055
13. Mosier DE, Gulizia RJ, Baird SM, Wilson DB. Transfer of a functional human immune system to mice with severe combined immunodeficiency. Nature (1988) 335(6187):256–9. doi: 10.1038/335256a0
14. Hesselton RM, Greiner DL, Mordes JP, Rajan TV, Sullivan JL, Shultz LD. High levels of human peripheral blood mononuclear cell engraftment and enhanced susceptibility to human immunodeficiency virus type 1 infection in NOD/LtSz-scid/scid mice. J Infect Dis (1995) 172(4):974–82. doi: 10.1093/infdis/172.4.974
15. van Rijn RS, Simonetti ER, Hagenbeek A, Hogenes MCH, de Weger RA, Canninga-van Dijk MR, et al. A new xenograft model for graft-versus-host disease by intravenous transfer of human peripheral blood mononuclear cells in RAG2-/- γc-/- double-mutant mice. Blood (2003) 102(7):2522–31. doi: 10.1182/blood-2002-10-3241
16. Ali N, Flutter B, Sanchez Rodriguez R, Sharif-Paghaleh E, Barber LD, Lombardi G, et al. Xenogeneic graft-versus-host-disease in NOD-scid IL-2Rγnull mice display a T-effector memory phenotype. PloS One (2012) 7(8):e44219. doi: 10.1371/journal.pone.0044219
17. Kamel-Reid S, Dick J. Engraftment of immune-deficient mice with human hematopoietic stem cells. Sci (N Y NY) (1988) 242(4886):1706–9. doi: 10.1126/science.2904703
18. Péault B, Weissman IL, Baum C, McCune JM, Tsukamoto A. Lymphoid reconstitution of the human fetal thymus in SCID mice with CD34+ precursor cells. J Exp Med (1991) 174(5):1283–6. doi: 10.1084/jem.174.5.1283
19. Hiramatsu H, Nishikomori R, Heike T, Ito M, Kobayashi K, Katamura K, et al. Complete reconstitution of human lymphocytes from cord blood CD34+ cells using the NOD/SCID/gammacnull mice model. Blood (2003) 102(3):873–80. doi: 10.1182/blood-2002-09-2755
20. Danner R, Chaudhari SN, Rosenberger J, Surls J, Richie TL, Brumeanu TD, et al. Expression of HLA class II molecules in humanized NOD.Rag1KO.IL2RgcKO mice is critical for development and function of human T and B cells. PloS One (2011) 6(5):e19826. doi: 10.1371/journal.pone.0019826
21. Lan P, Tonomura N, Shimizu A, Wang S, Yang Y-G. Reconstitution of a functional human immune system in immunodeficient mice through combined human fetal thymus/liver and CD34+ cell transplantation. Blood (2006) 108(2):487–92. doi: 10.1182/blood-2005-11-4388
22. Melkus MW, Estes JD, Padgett-Thomas A, Gatlin J, Denton PW, Othieno FA, et al. Humanized mice mount specific adaptive and innate immune responses to EBV and TSST-1. Nat Med (2006) 12(11):1316–22. doi: 10.1038/nm1431
23. Brainard DM, Seung E, Frahm N, Cariappa A, Bailey CC, Hart WK, et al. Induction of robust cellular and humoral virus-specific adaptive immune responses in human immunodeficiency virus-infected humanized BLT mice. J Virol (2009) 83(14):7305–21. doi: 10.1128/JVI.02207-08
24. Stoddart CA, Maidji E, Galkina SA, Kosikova G, Rivera JM, Moreno ME, et al. Superior human leukocyte reconstitution and susceptibility to vaginal HIV transmission in humanized NOD-scid IL-2Rγ(-/-) (NSG) BLT mice. Virology (2011) 417(1):154–60. doi: 10.1016/j.virol.2011.05.013
25. Lavender KJ, Messer RJ, Race B, Hasenkrug KJ. Production of bone marrow, liver, thymus (BLT) humanized mice on the C57BL/6 Rag2(-/-)γc(-/-)CD47(-/-) background. J Immunol Methods (2014) 407:127–34. doi: 10.1016/j.jim.2014.04.008
26. Lavender KJ, Pang WW, Messer RJ, Duley AK, Race B, Phillips K, et al. BLT-humanized C57BL/6 Rag2-/-γc-/-CD47-/- mice are resistant to GVHD and develop B- and T-cell immunity to HIV infection. Blood (2013) 122(25):4013–20. doi: 10.1182/blood-2013-06-506949
27. Lavender KJ, Pace C, Sutter K, Messer RJ, Pouncey DL, Cummins NW, et al. An advanced BLT-humanized mouse model for extended HIV-1 cure studies. AIDS (2018) 32(1):1–10. doi: 10.1097/QAD.0000000000001674
28. Lapidot T, Pflumio F, Doedens M, Murdoch B, Williams DE, Dick JE. Cytokine stimulation of multilineage hematopoiesis from immature human cells engrafted in SCID mice. Sci (N Y NY) (1992) 255(5048):1137–41. doi: 10.1126/science.1372131
29. Shultz LD, Saito Y, Najima Y, Tanaka S, Ochi T, Tomizawa M, et al. Generation of functional human T-cell subsets with HLA-restricted immune responses in HLA class I expressing NOD/SCID/IL2r gamma(null) humanized mice. Proc Natl Acad Sci USA (2010) 107(29):13022–7. doi: 10.1073/pnas.1000475107
30. Choi B, Chun E, Kim M, Kim ST, Yoon K, Lee KY, et al. Human B cell development and antibody production in humanized NOD/SCID/IL-2Rγ(null) (NSG) mice conditioned by busulfan. J Clin Immunol (2011) 31(2):253–64. doi: 10.1007/s10875-010-9478-2
31. Chen Q, He F, Kwang J, Chan JKY, Chen J. GM-CSF and IL-4 Stimulate Antibody Responses in Humanized Mice by Promoting T, B, and Dendritic Cell Maturation. J Immunol (2012) 189(11):5223–9. doi: 10.4049/jimmunol.1201789
32. Ito R, Takahashi T, Katano I, Kawai K, Kamisako T, Ogura T, et al. Establishment of a Human Allergy Model Using Human IL-3/GM-CSF–Transgenic NOG Mice. J Immunol (2013) 191(6):2890–9. doi: 10.4049/jimmunol.1203543
33. Rongvaux A, Willinger T, Martinek J, Strowig T, Gearty SV, Teichmann LL, et al. Development and function of human innate immune cells in a humanized mouse model. Nat Biotechnol (2014) 32(4):364–72. doi: 10.1038/nbt.2858
34. Billerbeck E, Horwitz JA, Labitt RN, Donovan BM, Vega K, Budell WC, et al. Characterization of human antiviral adaptive immune responses during hepatotropic virus infection in HLA-transgenic human immune system mice. J Immunol (Baltimore Md 1950) (2013) 191(4):1753–64. doi: 10.4049/jimmunol.1201518
35. Billerbeck E, Labitt RN, Vega K, Frias-Staheli N, Dorner M, Xiao JW, et al. Insufficient interleukin-12 signalling favours differentiation of human CD4+ and CD8+ T cells into GATA-3+ and GATA-3+ T-bet+ subsets in humanized mice. Immunology (2014) 143(2):202–18. doi: 10.1111/imm.12304
36. Ding Y, Wilkinson A, Idris A, Fancke B, O’Keeffe M, Khalil D, et al. FLT3-Ligand Treatment of Humanized Mice Results in the Generation of Large Numbers of CD141+ and CD1c+ Dendritic Cells In Vivo. J Immunol (2014) 192(4):1982–9. doi: 10.4049/jimmunol.1302391
37. Majji S, Wijayalath W, Shashikumar S, Pow-Sang L, Villasante E, Brumeanu TD, et al. Differential effect of HLA class-I versus class-II transgenes on human T and B cell reconstitution and function in NRG mice. Sci Rep (2016) 6:28093. doi: 10.1038/srep28093
38. Li Y, Mention JJ, Court N, Masse-Ranson G, Toubert A, Spits H, et al. A novel Flt3-deficient HIS mouse model with selective enhancement of human DC development. Eur J Immunol (2016) 46(5):1291–9. doi: 10.1002/eji.201546132
39. Herndler-Brandstetter D, Shan L, Yao Y, Stecher C, Plajer V, Lietzenmayer M, et al. Humanized mouse model supports development, function, and tissue residency of human natural killer cells. Proc Natl Acad Sci (2017) 114(45):E9626–E34. doi: 10.1073/pnas.1705301114
40. Biswas S, Chang H, Sarkis PT, Fikrig E, Zhu Q, Marasco WA. Humoral immune responses in humanized BLT mice immunized with West Nile virus and HIV-1 envelope proteins are largely mediated via human CD5+ B cells. Immunology (2011) 134(4):419–33. doi: 10.1111/j.1365-2567.2011.03501.x
41. Kalscheuer H, Danzl N, Onoe T, Faust T, Winchester R, Goland R, et al. A model for personalized in vivo analysis of human immune responsiveness. Sci Transl Med (2012) 4(125):125ra30. doi: 10.1126/scitranslmed.3003481
42. Denton PW, Nochi T, Lim A, Krisko JF, Martinez-Torres F, Choudhary SK, et al. IL-2 receptor γ-chain molecule is critical for intestinal T-cell reconstitution in humanized mice. Mucosal Immunol (2012) 5(5):555–66. doi: 10.1038/mi.2012.31
43. Smith DJ, Lin LJ, Moon H, Pham AT, Wang X, Liu S, et al. Propagating Humanized BLT Mice for the Study of Human Immunology and Immunotherapy. Stem Cells Dev (2016) 25(24):1863–73. doi: 10.1089/scd.2016.0193
44. Bryce PJ, Falahati R, Kenney LL, Leung J, Bebbington C, Tomasevic N, et al. Humanized mouse model of mast cell-mediated passive cutaneous anaphylaxis and passive systemic anaphylaxis. J Allergy Clin Immunol (2016) 138(3):769–79. doi: 10.1016/j.jaci.2016.01.049
45. Honeycutt JB, Wahl A, Baker C, Spagnuolo RA, Foster J, Zakharova O, et al. Macrophages sustain HIV replication in vivo independently of T cells. J Clin Invest (2016) 126(4):1353–66. doi: 10.1172/JCI84456
46. Nikzad R, Angelo LS, Aviles-Padilla K, Le DT, Singh VK, Bimler L, et al. Human natural killer cells mediate adaptive immunity to viral antigens. Sci Immunol (2019) 4(35):eaat8116. doi: 10.1126/sciimmunol.aat8116
47. Poignard P, Sabbe R, Picchio GR, Wang M, Gulizia RJ, Katinger H, et al. Neutralizing antibodies have limited effects on the control of established HIV-1 infection in vivo. Immunity (1999) 10(4):431–8. doi: 10.1016/S1074-7613(00)80043-6
48. Klein F, Halper-Stromberg A, Horwitz JA, Gruell H, Scheid JF, Bournazos S, et al. HIV therapy by a combination of broadly neutralizing antibodies in humanized mice. Nature (2012) 492(7427):118–22. doi: 10.1038/nature11604
49. Horwitz JA, Halper-Stromberg A, Mouquet H, Gitlin AD, Tretiakova A, Eisenreich TR, et al. HIV-1 suppression and durable control by combining single broadly neutralizing antibodies and antiretroviral drugs in humanized mice. Proc Natl Acad Sci (2013) 110(41):16538–43. doi: 10.1073/pnas.1315295110
50. Halper-Stromberg A, Lu C-L, Klein F, Horwitz Joshua A, Bournazos S, Nogueira L, et al. Broadly Neutralizing Antibodies and Viral Inducers Decrease Rebound from HIV-1 Latent Reservoirs in Humanized Mice. Cell (2014) 158(5):989–99. doi: 10.1016/j.cell.2014.07.043
51. Wang P, Gajjar MR, Yu J, Padte NN, Gettie A, Blanchard JL, et al. Quantifying the contribution of Fc-mediated effector functions to the antiviral activity of anti-HIV-1 IgG1 antibodies in vivo. Proc Natl Acad Sci U S A (2020) 117(30):18002–9. doi: 10.1073/pnas.2008190117
52. Badamchi-Zadeh A, Tartaglia LJ, Abbink P, Bricault CA, Liu P-T, Boyd M, et al. Therapeutic Efficacy of Vectored PGT121 Gene Delivery in HIV-1-Infected Humanized Mice. J Virol (2018) 92(7):e01925–17. doi: 10.1128/JVI.01925-17
53. Kitchen SG, Bennett M, Galić Z, Kim J, Xu Q, Young A, et al. Engineering antigen-specific T cells from genetically modified human hematopoietic stem cells in immunodeficient mice. PloS One (2009) 4(12):e8208. doi: 10.1371/journal.pone.0008208
54. Kitchen SG, Levin BR, Bristol G, Rezek V, Kim S, Aguilera-Sandoval C, et al. In vivo suppression of HIV by antigen specific T cells derived from engineered hematopoietic stem cells. PloS Pathog (2012) 8(4):e1002649. doi: 10.1371/journal.ppat.1002649
55. Zhen A, Kamata M, Rezek V, Rick J, Levin B, Kasparian S, et al. HIV-specific Immunity Derived From Chimeric Antigen Receptor-engineered Stem Cells. Mol Ther J Am Soc Gene Ther (2015) 23(8):1358–67. doi: 10.1038/mt.2015.102
56. Leibman RS, Richardson MW, Ellebrecht CT, Maldini CR, Glover JA, Secreto AJ, et al. Supraphysiologic control over HIV-1 replication mediated by CD8 T cells expressing a re-engineered CD4-based chimeric antigen receptor. PloS Pathog (2017) 13(10):e1006613. doi: 10.1371/journal.ppat.1006613
57. Bardhi A, Wu Y, Chen W, Li W, Zhu Z, Zheng JH, et al. Potent In Vivo NK Cell-Mediated Elimination of HIV-1-Infected Cells Mobilized by a gp120-Bispecific and Hexavalent Broadly Neutralizing Fusion Protein. J Virol (2017) 91(20):e00937–17. doi: 10.1128/JVI.00937-17
58. Anthony-Gonda K, Bardhi A, Ray A, Flerin N, Li M, Chen W, et al. Multispecific anti-HIV duoCAR-T cells display broad in vitro antiviral activity and potent in vivo elimination of HIV-infected cells in a humanized mouse model. Sci Trans Med (2019) 11(504):eaav5685. doi: 10.1126/scitranslmed.aav5685
59. Perez EE, Wang J, Miller JC, Jouvenot Y, Kim KA, Liu O, et al. Establishment of HIV-1 resistance in CD4+ T cells by genome editing using zinc-finger nucleases. Nat Biotechnol (2008) 26(7):808–16. doi: 10.1038/nbt1410
60. Yi G, Choi JG, Bharaj P, Abraham S, Dang Y, Kafri T, et al. CCR5 Gene Editing of Resting CD4(+) T Cells by Transient ZFN Expression From HIV Envelope Pseudotyped Nonintegrating Lentivirus Confers HIV-1 Resistance in Humanized Mice. Mol Ther Nucleic Acids (2014) 3:e198. doi: 10.1038/mtna.2014.52
61. Wilen CB, Wang J, Tilton JC, Miller JC, Kim KA, Rebar EJ, et al. Engineering HIV-Resistant Human CD4+ T Cells with CXCR4-Specific Zinc-Finger Nucleases. PloS Pathog (2011) 7(4):e1002020. doi: 10.1371/journal.ppat.1002020
62. Yuan J, Wang J, Crain K, Fearns C, Kim KA, Hua KL, et al. Zinc-finger Nuclease Editing of Human cxcr4 Promotes HIV-1 CD4+ T Cell Resistance and Enrichment. Mol Ther (2012) 20(4):849–59. doi: 10.1038/mt.2011.310
63. Llewellyn GN, Seclén E, Wietgrefe S, Liu S, Chateau M, Pei H, et al. Humanized Mouse Model of HIV-1 Latency with Enrichment of Latent Virus in PD-1(+) and TIGIT(+) CD4 T Cells. J Virol (2019) 93(10):e02086–18. doi: 10.1128/JVI.02086-18
64. Xiao Q, Chen S, Wang Q, Liu Z, Liu S, Deng H, et al. CCR5 editing by Staphylococcus aureus Cas9 in human primary CD4+ T cells and hematopoietic stem/progenitor cells promotes HIV-1 resistance and CD4+ T cell enrichment in humanized mice. Retrovirology (2019) 16(1):15. doi: 10.1186/s12977-019-0477-y
65. Dash PK, Kaminski R, Bella R, Su H, Mathews S, Ahooyi TM, et al. and CRISPR Treatments Eliminate HIV-1 in a Subset of Infected Humanized Mice. Nat Commun (2019) 10(1):2753. doi: 10.1038/s41467-019-10366-y
66. Yin C, Zhang T, Qu X, Zhang Y, Putatunda R, Xiao X, et al. In Vivo Excision of HIV-1 Provirus by saCas9 and Multiplex Single-Guide RNAs in Animal Models. Mol Ther J Am Soc Gene Ther (2017) 25(5):1168–86. doi: 10.1016/j.ymthe.2017.03.012
67. Kessing CF, Nixon CC, Li C, Tsai P, Takata H, Mousseau G, et al. In Vivo Suppression of HIV Rebound by Didehydro-Cortistatin A, a “Block-and-Lock” Strategy for HIV-1 Treatment. Cell Rep (2017) 21(3):600–11. doi: 10.1016/j.celrep.2017.09.080
68. Gatti RA, Meuwissen HJ, Allen HD, Hong R, Good RA. Immunological reconstitution of sex-linked lymphopenic immunological deficiency. Lancet (London England) (1968) 2(7583):1366–9. doi: 10.1016/S0140-6736(68)92673-1
69. Bosma GC, Custer RP, Bosma MJ. A severe combined immunodeficiency mutation in the mouse. Nature (1983) 301(5900):527–30. doi: 10.1038/301527a0
70. Fulop GM, Phillips RA. Full reconstitution of the immune deficiency in scid mice with normal stem cells requires low-dose irradiation of the recipients. J Immunol (Baltimore Md 1950) (1986) 136(12):4438–43.
71. McCune J, Kaneshima H, Krowka J, Namikawa R, Outzen H, Peault B, et al. The SCID-hu mouse: a small animal model for HIV infection and pathogenesis. Annu Rev Immunol (1991) 9:399–429. doi: 10.1146/annurev.iy.09.040191.002151
72. Moore MAS, Owen JJT. STEM-CELL MIGRATION IN DEVELOPING MYELOID AND LYMPHOID SYSTEMS. Lancet (1967) 290(7517):658–9. doi: 10.1016/S0140-6736(67)90693-9
73. Namikawa R, Mizuno T, Matsuoka H, Fukami H, Ueda R, Itoh G, et al. Ontogenic development of T and B cells and non-lymphoid cells in the white pulp of human spleen. Immunology (1986) 57(1):61–9.
74. Barry TS, Jones DM, Richter CB, Haynes BF. Successful engraftment of human postnatal thymus in severe combined immune deficient (SCID) mice: differential engraftment of thymic components with irradiation versus anti-asialo GM-1 immunosuppressive regimens. J Exp Med (1991) 173(1):167–80. doi: 10.1084/jem.173.1.167
75. Aldrovandi GM, Feuer G, Gao L, Jamieson B, Kristeva M, Chen IS, et al. The SCID-hu mouse as a model for HIV-1 infection. Nature (1993) 363(6431):732–6. doi: 10.1038/363732a0
76. Namikawa R, Kaneshima H, Lieberman M, Weissman IL, McCune JM. Infection of the SCID-hu mouse by HIV-1. Sci (N Y NY) (1988) 242(4886):1684–6. doi: 10.1126/science.3201256
77. McCune J, Namikawa R, Shih C, Rabin L, Kaneshima H. Suppression of HIV infection in AZT-treated SCID-hu mice. Sci (N Y NY) (1990) 247(4942):564–6. doi: 10.1126/science.2300816
78. Torbett BE, Picchio G, Mosier DE. hu-PBL-SCID Mice: A Model for Human Immune Function, AIDS, and Lymphomagenesis. Immunol Rev (1991) 124(1):139–64. doi: 10.1111/j.1600-065X.1991.tb00620.x
79. Rizza P, Santini SM, Logozzi MA, Lapenta C, Sestili P, Gherardi G, et al. T-cell dysfunctions in hu-PBL-SCID mice infected with human immunodeficiency virus (HIV) shortly after reconstitution: in vivo effects of HIV on highly activated human immune cells.s. J Virol (1996) 70(11):7958–64. doi: 10.1128/JVI.70.11.7958-7964.1996
80. Makino S, Kunimoto K, Muraoka Y, Mizushima Y, Katagiri K, Tochino Y. Breeding of a non-obese, diabetic strain of mice. Jikken Dobutsu (1980) 29(1):1–13. doi: 10.1538/expanim1978.29.1_1
81. Leiter EH, Serreze DV, Prochazka M. The genetics and epidemiology of diabetes in NOD mice. Immunol Today (1990) 11(5):147–9. doi: 10.1016/0167-5699(90)90057-G
82. Cao X, Shores EW, Hu-Li J, Anver MR, Kelsall BL, Russell SM, et al. Defective lymphoid development in mice lacking expression of the common cytokine receptor gamma chain. Immunity (1995) 2(3):223–38. doi: 10.1016/1074-7613(95)90047-0
83. Ohbo K, Suda T, Hashiyama M, Mantani A, Ikebe M, Miyakawa K, et al. Modulation of hematopoiesis in mice with a truncated mutant of the interleukin-2 receptor gamma chain. Blood (1996) 87(3):956–67. doi: 10.1182/blood.V87.3.956.bloodjournal873956
84. Russell S, Keegan A, Harada N, Nakamura Y, Noguchi M, Leland P, et al. Interleukin-2 receptor gamma chain: a functional component of the interleukin-4 receptor. Sci (N Y NY) (1993) 262(5141):1880–3. doi: 10.1126/science.8266078
85. Kondo M, Takeshita T, Ishii N, Nakamura M, Watanabe S, K-i A, et al. Sharing of the Interleukin-2 (IL-2) receptor ?? chain between receptors for IL-2 and IL-4. Sci (N Y NY) (1994) 262:1874–7. doi: 10.1126/science.8266076
86. Noguchi M, Nakamura Y, Russell S, Ziegler S, Tsang M, Cao X, et al. Interleukin-2 receptor gamma chain: a functional component of the interleukin-7 receptor. Sci (N Y NY) (1993) 262(5141):1877–80. doi: 10.1126/science.8266077
87. Kondo M, Takeshita T, Higuchi M, Nakamura M, Sudo T, Nishikawa S, et al. Functional participation of the IL-2 receptor gamma chain in IL-7 receptor complexes. Sci (N Y NY) (1994) 263(5152):1453–4. doi: 10.1126/science.8128231
88. Kimura Y, Takeshita T, Kondo M, Ishii N, Nakamura M, Van Snick J, et al. Sharing of the IL-2 receptor gamma chain with the functional IL-9 receptor complex. Int Immunol (1995) 7(1):115–20. doi: 10.1093/intimm/7.1.115
89. Giri JG, Ahdieh M, Eisenman J, Shanebeck K, Grabstein K, Kumaki S, et al. Utilization of the beta and gamma chains of the IL-2 receptor by the novel cytokine IL-15. EMBO J (1994) 13(12):2822–30. doi: 10.1002/j.1460-2075.1994.tb06576.x
90. Habib T, Senadheera S, Weinberg K, Kaushansky K. The common gamma chain (gamma c) is a required signaling component of the IL-21 receptor and supports IL-21-induced cell proliferation via JAK3. Biochemistry (2002) 41(27):8725–31. doi: 10.1021/bi0202023
91. Christianson SW, Greiner DL, Schweitzer IB, Gott B, Beamer GL, Schweitzer PA, et al. Role of natural killer cells on engraftment of human lymphoid cells and on metastasis of human T-lymphoblastoid leukemia cells in C57BL/6J-scid mice and in C57BL/6J-scid bg mice. Cell Immunol (1996) 171(2):186–99. doi: 10.1006/cimm.1996.0193
92. Ifversen P, Borrebaeck CA. SCID-hu-PBL: a model for making human antibodies? Semin Immunol (1996) 8(4):243–8. doi: 10.1006/smim.1996.0030
93. Murphy WJ, Taub DD, Longo DL. The huPBL-SCID mouse as a means to examine human immune function in vivo. Semin Immunol (1996) 8(4):233–41. doi: 10.1006/smim.1996.0029
94. Koyanagi Y, Tanaka Y, Kira J, Ito M, Hioki K, Misawa N, et al. Primary human immunodeficiency virus type 1 viremia and central nervous system invasion in a novel hu-PBL-immunodeficient mouse strain. J Virol (1997) 71(3):2417–24. doi: 10.1128/JVI.71.3.2417-2424.1997
95. Kim KC, Choi BS, Kim KC, Park KH, Lee HJ, Cho YK, et al. A Simple Mouse Model for the Study of Human Immunodeficiency Virus. AIDS Res Hum Retroviruses (2016) 32(2):194–202. doi: 10.1089/aid.2015.0211
96. Mutis T, van Rijn RS, Simonetti ER, Aarts-Riemens T, Emmelot ME, van Bloois L, et al. Human Regulatory T Cells Control Xenogeneic Graft-versus-Host Disease Induced by Autologous T Cells in RAG2–/–γc–/– Immunodeficient Mice. Clin Cancer Res (2006) 12(18):5520–5. doi: 10.1158/1078-0432.CCR-06-0035
97. Gregoire-Gauthier J, Durrieu L, Duval A, Fontaine F, Dieng MM, Bourgey M, et al. Use of immunoglobulins in the prevention of GvHD in a xenogeneic NOD/SCID/γc– mouse model. Bone Marrow Transplant (2012) 47(3):439–50. doi: 10.1038/bmt.2011.93
98. Civin CI, Strauss LC, Brovall C, Fackler MJ, Schwartz JF, Shaper JH. Antigenic analysis of hematopoiesis. III. A hematopoietic progenitor cell surface antigen defined by a monoclonal antibody raised against KG-1a cells. J Immunol (Baltimore Md 1950) (1984) 133(1):157–65.
99. Andrews RG, Singer JW, Bernstein ID. Precursors of colony-forming cells in humans can be distinguished from colony-forming cells by expression of the CD33 and CD34 antigens and light scatter properties. J Exp Med (1989) 169(5):1721–31. doi: 10.1084/jem.169.5.1721
100. Sutherland HJ, Eaves CJ, Eaves AC, Dragowska W, Lansdorp PM. Characterization and partial purification of human marrow cells capable of initiating long-term hematopoiesis in vitro. Blood (1989) 74(5):1563–70. doi: 10.1182/blood.V74.5.1563.1563
101. Bhatia M, Wang JC, Kapp U, Bonnet D, Dick JE. Purification of primitive human hematopoietic cells capable of repopulating immune-deficient mice. Proc Natl Acad Sci U S A (1997) 94(10):5320–5. doi: 10.1073/pnas.94.10.5320
102. Holyoake TL, Nicolini FE, Eaves CJ. Functional differences between transplantable human hematopoietic stem cells from fetal liver, cord blood, and adult marrow. Exp Hematol (1999) 27(9):1418–27. doi: 10.1016/S0301-472X(99)00078-8
103. Hao Q, Shah A, Thiemann F, Smogorzewska E, Crooks G. A functional comparison of CD34 + CD38- cells in cord blood and bone marrow. Blood (1995) 86(10):3745–53. doi: 10.1182/blood.V86.10.3745.bloodjournal86103745
104. Larochelle A, Vormoor J, Hanenberg H, Wang JCY, Bhatia M, Lapidot T, et al. Identification of primitive human hematopoietic cells capable of repopulating NOD/SCID mouse bone marrow: Implications for gene therapy. Nat Med (1996) 2(12):1329–37. doi: 10.1038/nm1296-1329
105. Ueda T, Yoshino H, Kobayashi K, Kawahata M, Ebihara Y, Ito M, et al. Hematopoietic repopulating ability of cord blood CD34(+) cells in NOD/Shi-scid mice. Stem Cells (2000) 18(3):204–13. doi: 10.1634/stemcells.18-3-204
106. Yahata T, Ando K, Nakamura Y, Ueyama Y, Shimamura K, Tamaoki N, et al. Functional human T lymphocyte development from cord blood CD34+ cells in nonobese diabetic/Shi-scid, IL-2 receptor gamma null mice. J Immunol (Baltimore Md 1950) (2002) 169(1):204–9. doi: 10.4049/jimmunol.169.1.204
107. Shultz LD, Lyons BL, Burzenski LM, Gott B, Chen X, Chaleff S, et al. Human Lymphoid and Myeloid Cell Development in NOD/LtSz-scid IL2Rγnull Mice Engrafted with Mobilized Human Hemopoietic Stem Cells. J Immunol (2005) 174(10):6477–89. doi: 10.4049/jimmunol.174.10.6477
108. McDermott SP, Eppert K, Lechman ER, Doedens M, Dick JE. Comparison of human cord blood engraftment between immunocompromised mouse strains. Blood (2010) 116(2):193–200. doi: 10.1182/blood-2010-02-271841
109. Tanaka T, Kitamura F, Nagasaka Y, Kuida K, Suwa H, Miyasaka M. Selective long-term elimination of natural killer cells in vivo by an anti-interleukin 2 receptor beta chain monoclonal antibody in mice. J Exp Med (1993) 178(3):1103–7. doi: 10.1084/jem.178.3.1103
110. Watanabe S, Terashima K, Ohta S, Horibata S, Yajima M, Shiozawa Y, et al. Hematopoietic stem cell–engrafted NOD/SCID/IL2Rγnull mice develop human lymphoid systems and induce long-lasting HIV-1 infection with specific humoral immune responses. Blood (2006) 109(1):212–8. doi: 10.1182/blood-2006-04-017681
111. Watanabe S, Ohta S, Yajima M, Terashima K, Ito M, Mugishima H, et al. Humanized NOD/SCID/IL2Rγnull Mice Transplanted with Hematopoietic Stem Cells under Nonmyeloablative Conditions Show Prolonged Life Spans and Allow Detailed Analysis of Human Immunodeficiency Virus Type 1 Pathogenesis. J Virol (2007) 81(23):13259–64. doi: 10.1128/JVI.01353-07
112. Nie C, Sato K, Misawa N, Kitayama H, Fujino H, Hiramatsu H, et al. Selective infection of CD4+ effector memory T lymphocytes leads to preferential depletion of memory T lymphocytes in R5 HIV-1-infected humanized NOD/SCID/IL-2Rgammanull mice. Virology (2009) 394(1):64–72. doi: 10.1016/j.virol.2009.08.011
113. Sato K, Izumi T, Misawa N, Kobayashi T, Yamashita Y, Ohmichi M, et al. Remarkable Lethal G-to-A Mutations in vif-Proficient HIV-1 Provirus by Individual APOBEC3 Proteins in Humanized Mice. J Virol (2010) 84(18):9546–56. doi: 10.1128/JVI.00823-10
114. Sato K, Nie C, Misawa N, Tanaka Y, Ito M, Koyanagi Y. Dynamics of memory and naïve CD8+ T lymphocytes in humanized NOD/SCID/IL-2Rgammanull mice infected with CCR5-tropic HIV-1. Vaccine (2010) 28 Suppl 2:B32–7. doi: 10.1016/j.vaccine.2009.10.154
115. Berges BK, Akkina SR, Folkvord JM, Connick E, Akkina R. Mucosal transmission of R5 and X4 tropic HIV-1 via vaginal and rectal routes in humanized Rag2-/- gammac -/- (RAG-hu) mice. Virology (2008) 373(2):342–51. doi: 10.1016/j.virol.2007.11.020
116. Akkina R, Berges BK, Palmer BE, Remling L, Neff CP, Kuruvilla J, et al. Humanized Rag1-/- γc-/- mice support multilineage hematopoiesis and are susceptible to HIV-1 infection via systemic and vaginal routes. PloS One (2011) 6(6):e20169. doi: 10.1371/journal.pone.0020169
117. Van Duyne R, Cardenas J, Easley R, Wu W, Kehn-Hall K, Klase Z, et al. Effect of transcription peptide inhibitors on HIV-1 replication. Virology (2008) 376(2):308–22. doi: 10.1016/j.virol.2008.02.036
118. Cheng L, Ma J, Li G, Su L. Humanized Mice Engrafted With Human HSC Only or HSC and Thymus Support Comparable HIV-1 Replication, Immunopathology, and Responses to ART and Immune Therapy. Front Immunol (2018) 9:817–. doi: 10.3389/fimmu.2018.00817
119. Satheesan S, Li H, Burnett JC, Takahashi M, Li S, Wu SX, et al. HIV Replication and Latency in a Humanized NSG Mouse Model during Suppressive Oral Combinational Antiretroviral Therapy. J Virol (2018) 92(7):e02118–17. doi: 10.1128/JVI.02118-17
120. Kumar P, Ban HS, Kim SS, Wu H, Pearson T, Greiner DL, et al. T cell-specific siRNA delivery suppresses HIV-1 infection in humanized mice. Cell (2008) 134(4):577–86. doi: 10.1016/j.cell.2008.06.034
121. Gorantla S, Makarov E, Finke-Dwyer J, Gebhart CL, Domm W, Dewhurst S, et al. CD8+ Cell Depletion Accelerates HIV-1 Immunopathology in Humanized Mice. J Immunol (2010) 184(12):7082–91. doi: 10.4049/jimmunol.1000438
122. Gorantla S, Makarov E, Finke-Dwyer J, Castanedo A, Holguin A, Gebhart CL, et al. Links between progressive HIV-1 infection of humanized mice and viral neuropathogenesis. Am J Pathol (2010) 177(6):2938–49. doi: 10.2353/ajpath.2010.100536
123. Holt N, Wang J, Kim K, Friedman G, Wang X, Taupin V, et al. Human hematopoietic stem/progenitor cells modified by zinc-finger nucleases targeted to CCR5 control HIV-1 in vivo. Nat Biotechnol (2010) 28(8):839–47. doi: 10.1038/nbt.1663
124. Dash PK, Gorantla S, Gendelman HE, Knibbe J, Casale GP, Makarov E, et al. Loss of neuronal integrity during progressive HIV-1 infection of humanized mice. J Neurosci (2011) 31(9):3148–57. doi: 10.1523/JNEUROSCI.5473-10.2011
125. Kim J, Peachman KK, Jobe O, Morrison EB, Allam A, Jagodzinski L, et al. Tracking Human Immunodeficiency Virus-1 Infection in the Humanized DRAG Mouse Model. Front Immunol (2017) 8:1405–. doi: 10.3389/fimmu.2017.01405
126. Allam A, Majji S, Peachman K, Jagodzinski L, Kim J, Ratto-Kim S, et al. TFH cells accumulate in mucosal tissues of humanized-DRAG mice and are highly permissive to HIV-1. Sci Rep (2015) 5(1):10443. doi: 10.1038/srep10443
127. Denton PW, Estes JD, Sun Z, Othieno FA, Wei BL, Wege AK, et al. Antiretroviral pre-exposure prophylaxis prevents vaginal transmission of HIV-1 in humanized BLT mice. PloS Med (2008) 5(1):e16. doi: 10.1371/journal.pmed.0050016
128. Sun Z, Denton PW, Estes JD, Othieno FA, Wei BL, Wege AK, et al. Intrarectal transmission, systemic infection, and CD4+ T cell depletion in humanized mice infected with HIV-1. J Exp Med (2007) 204(4):705–14. doi: 10.1084/jem.20062411
129. Denton PW, Othieno F, Martinez-Torres F, Zou W, Krisko JF, Fleming E, et al. One percent tenofovir applied topically to humanized BLT mice and used according to the CAPRISA 004 experimental design demonstrates partial protection from vaginal HIV infection, validating the BLT model for evaluation of new microbicide candidates. J Virol (2011) 85(15):7582–93. doi: 10.1128/JVI.00537-11
130. Long BR, Stoddart CA. Alpha Interferon and HIV Infection Cause Activation of Human T Cells in NSG-BLT Mice. J Virol (2012) 86(6):3327–36. doi: 10.1128/JVI.06676-11
131. Dudek TE, No DC, Seung E, Vrbanac VD, Fadda L, Bhoumik P, et al. Rapid Evolution of HIV-1 to Functional CD8+ T Cell Responses in Humanized BLT Mice. Sci Trans Med (2012) 4(143):143ra98–ra98. doi: 10.1126/scitranslmed.3003984
132. Chang H, Biswas S, Tallarico AS, Sarkis PTN, Geng S, Panditrao MM, et al. Human B-cell ontogeny in humanized NOD/SCID γc(null) mice generates a diverse yet auto/poly- and HIV-1-reactive antibody repertoire. Genes Immun (2012) 13(5):399–410. doi: 10.1038/gene.2012.16
133. Seung E, Tager AM. Humoral immunity in humanized mice: a work in progress. J Infect Dis (2013) 208Suppl 2(Suppl 2):S155–S9. doi: 10.1093/infdis/jit448
134. Martinez-Torres F, Nochi T, Wahl A, Garcia JV, Denton PW. Hypogammaglobulinemia in BLT Humanized Mice – An Animal Model of Primary Antibody Deficiency. PloS One (2014) 9(10):e108663. doi: 10.1371/journal.pone.0108663
135. Denton PW, Olesen R, Choudhary SK, Archin NM, Wahl A, Swanson MD, et al. Generation of HIV Latency in Humanized BLT Mice. J Virol (2012) 86(1):630–4. doi: 10.1128/JVI.06120-11
136. Takenaka K, Prasolava TK, Wang JCY, Mortin-Toth SM, Khalouei S, Gan OI, et al. Polymorphism in Sirpa modulates engraftment of human hematopoietic stem cells. Nat Immunol (2007) 8(12):1313–23. doi: 10.1038/ni1527
137. Yamauchi T, Takenaka K, Urata S, Shima T, Kikushige Y, Tokuyama T, et al. Polymorphic Sirpa is the genetic determinant for NOD-based mouse lines to achieve efficient human cell engraftment. Blood (2013) 121(8):1316–25. doi: 10.1182/blood-2012-06-440354
138. Oldenborg P-A, Zheleznyak A, Fang Y-F, Lagenaur CF, Gresham HD, Lindberg FP. Role of CD47 as a Marker of Self on Red Blood Cells. Sci (N Y NY) (2000) 288(5473):2051–4. doi: 10.1126/science.288.5473.2051
139. Blazar BR, Lindberg FP, Ingulli E, Panoskaltsis-Mortari A, Oldenborg PA, Iizuka K, et al. CD47 (integrin-associated protein) engagement of dendritic cell and macrophage counterreceptors is required to prevent the clearance of donor lymphohematopoietic cells. J Exp Med (2001) 194(4):541–9. doi: 10.1084/jem.194.4.541
140. Greenblatt MB, Vrbanac V, Tivey T, Tsang K, Tager AM, Aliprantis AO. Graft versus host disease in the bone marrow, liver and thymus humanized mouse model. PloS One (2012) 7(9):e44664. doi: 10.1371/journal.pone.0044664
141. Barton K, Winckelmann A, Palmer S. HIV-1 Reservoirs During Suppressive Therapy. Trends Microbiol (2016) 24(5):345–55. doi: 10.1016/j.tim.2016.01.006
142. Siliciano RF. The Challenge of Viral Reservoirs in HIV-1 Infection. Annu Rev Med (2002) 53(1):557–93. doi: 10.1146/annurev.med.53.082901.104024
143. Murray AJ, Kwon KJ, Farber DL, Siliciano RF. The Latent Reservoir for HIV-1: How Immunologic Memory and Clonal Expansion Contribute to HIV-1 Persistence. J Immunol (2016) 197(2):407–17. doi: 10.4049/jimmunol.1600343
144. Migueles SA, Connors M. Success and failure of the cellular immune response against HIV-1. Nat Immunol (2015) 16(6):563–70. doi: 10.1038/ni.3161
145. Archin NM, Liberty AL, Kashuba AD, Choudhary SK, Kuruc JD, Crooks AM, et al. Administration of vorinostat disrupts HIV-1 latency in patients on antiretroviral therapy. Nature (2012) 487(7408):482–5. doi: 10.1038/nature11286
146. Rasmussen TA, Tolstrup M, Brinkmann CR, Olesen R, Erikstrup C, Solomon A, et al. Panobinostat, a histone deacetylase inhibitor, for latent-virus reactivation in HIV-infected patients on suppressive antiretroviral therapy: a phase 1/2, single group, clinical trial. Lancet HIV (2014) 1(1):e13–21. doi: 10.1016/S2352-3018(14)70014-1
147. Søgaard OS, Graversen ME, Leth S, Olesen R, Brinkmann CR, Nissen SK, et al. The Depsipeptide Romidepsin Reverses HIV-1 Latency In Vivo. PloS Pathog (2015) 11(9):e1005142. doi: 10.1371/journal.ppat.1005142
148. Bouchat S, Gatot JS, Kabeya K, Cardona C, Colin L, Herbein G, et al. Histone methyltransferase inhibitors induce HIV-1 recovery in resting CD4(+) T cells from HIV-1-infected HAART-treated patients. AIDS (2012) 26(12):1473–82. doi: 10.1097/QAD.0b013e32835535f5
149. Spivak AM, Bosque A, Balch AH, Smyth D, Martins L, Planelles V. Ex Vivo Bioactivity and HIV-1 Latency Reversal by Ingenol Dibenzoate and Panobinostat in Resting CD4+ T Cells from Aviremic Patients. Antimicrobial Agents Chemother (2015) 59(10):5984–91. doi: 10.1128/AAC.01077-15
150. Banerjee C, Archin N, Michaels D, Belkina AC, Denis GV, Bradner J, et al. BET bromodomain inhibition as a novel strategy for reactivation of HIV-1. J Leukoc Biol (2012) 92(6):1147–54. doi: 10.1189/jlb.0312165
151. Doyon G, Zerbato J, Mellors JW, Sluis-Cremer N. Disulfiram reactivates latent HIV-1 expression through depletion of the phosphatase and tensin homolog. AIDS (2013) 27(2):F7–f11. doi: 10.1097/QAD.0b013e3283570620
152. Lim S-Y, Osuna CE, Hraber PT, Hesselgesser J, Gerold JM, Barnes TL, et al. TLR7 agonists induce transient viremia and reduce the viral reservoir in SIV-infected rhesus macaques on antiretroviral therapy. Sci Trans Med (2018) 10(439):eaao4521. doi: 10.1126/scitranslmed.aao4521
153. Watson DC, Moysi E, Valentin A, Bergamaschi C, Devasundaram S, Fortis SP, et al. Treatment with native heterodimeric IL-15 increases cytotoxic lymphocytes and reduces SHIV RNA in lymph nodes. PloS Pathog (2018) 14(2):e1006902. doi: 10.1371/journal.ppat.1006902
154. Tsai P, Wu G, Baker CE, Thayer WO, Spagnuolo RA, Sanchez R, et al. In vivo analysis of the effect of panobinostat on cell-associated HIV RNA and DNA levels and latent HIV infection. Retrovirology (2016) 13(1):36. doi: 10.1186/s12977-016-0268-7
155. Laird GM, Bullen CK, Rosenbloom DI, Martin AR, Hill AL, Durand CM, et al. Ex vivo analysis identifies effective HIV-1 latency-reversing drug combinations. J Clin Invest (2015) 125(5):1901–12. doi: 10.1172/JCI80142
156. Jiang G, Dandekar S. Targeting NF-κB signaling with protein kinase C agonists as an emerging strategy for combating HIV latency. AIDS Res Hum Retroviruses (2015) 31(1):4–12. doi: 10.1089/aid.2014.0199
157. Rice AP. Cyclin-dependent kinases as therapeutic targets for HIV-1 infection. Expert Opin Ther Targets (2016) 20(12):1453–61. doi: 10.1080/14728222.2016.1254619
158. Wender PA, Kee J-M, Warrington JM. Practical Synthesis of Prostratin, DPP, and Their Analogs, Adjuvant Leads Against Latent HIV. Sci (N Y NY) (2008) 320(5876):649–52. doi: 10.1126/science.1154690
159. Gutiérrez C, Serrano-Villar S, Madrid-Elena N, Pérez-Elías MJ, Martín ME, Barbas C, et al. Bryostatin-1 for latent virus reactivation in HIV-infected patients on antiretroviral therapy. AIDS (2016) 30(9):1385–92. doi: 10.1097/QAD.0000000000001064
160. Marsden MD, Loy BA, Wu X, Ramirez CM, Schrier AJ, Murray D, et al. In vivo activation of latent HIV with a synthetic bryostatin analog effects both latent cell “kick” and “kill” in strategy for virus eradication. PloS Pathog (2017) 13(9):e1006575. doi: 10.1371/journal.ppat.1006575
161. Bobardt M, Kuo J, Chatterji U, Chanda S, Little SJ, Wiedemann N, et al. The inhibitor apoptosis protein antagonist Debio 1143 Is an attractive HIV-1 latency reversal candidate. PloS One (2019) 14(2):e0211746–e. doi: 10.1371/journal.pone.0211746
162. Nixon CC, Mavigner M, Sampey GC, Brooks AD, Spagnuolo RA, Irlbeck DM, et al. and SIV latency reversal via non-canonical NF-κB signalling in vivo. Nature (2020) 578(7793):160–5. doi: 10.1038/s41586-020-1951-3
163. Garrido C, Abad-Fernandez M, Tuyishime M, Pollara JJ, Ferrari G, Soriano-Sarabia N, et al. Interleukin-15-Stimulated Natural Killer Cells Clear HIV-1-Infected Cells following Latency Reversal Ex Vivo. J Virol (2018) 92(12):e00235–18. doi: 10.1128/JVI.00235-18
164. Seay K, Church C, Zheng JH, Deneroff K, Ochsenbauer C, Kappes JC, et al. In Vivo Activation of Human NK Cells by Treatment with an Interleukin-15 Superagonist Potently Inhibits Acute In Vivo HIV-1 Infection in Humanized Mice. J Virol (2015) 89(12):6264–74. doi: 10.1128/JVI.00563-15
165. McBrien JB, Mavigner M, Franchitti L, Smith SA, White E, Tharp GK, et al. Robust and persistent reactivation of SIV and HIV by N-803 and depletion of CD8+ cells. Nature (2020) 578(7793):154–9. doi: 10.1038/s41586-020-1946-0
166. Llewellyn GN, Alvarez-Carbonell D, Chateau M, Karn J, Cannon PM. HIV-1 infection of microglial cells in a reconstituted humanized mouse model and identification of compounds that selectively reverse HIV latency. J Neurovirol (2018) 24(2):192–203. doi: 10.1007/s13365-017-0604-2
167. Rasmussen TA, Lewin SR. Shocking HIV out of hiding: where are we with clinical trials of latency reversing agents? Curr Opin HIV AIDS (2016) 11(4):394–401. doi: 10.1097/COH.0000000000000279
168. Shan L, Deng K, Shroff NS, Durand CM, Rabi SA, Yang H-C, et al. Stimulation of HIV-1-specific cytolytic T lymphocytes facilitates elimination of latent viral reservoir after virus reactivation. Immunity (2012) 36(3):491–501. doi: 10.1016/j.immuni.2012.01.014
169. Mikulak J, Oriolo F, Zaghi E, Di Vito C, Mavilio D. Natural killer cells in HIV-1 infection and therapy. AIDS (2017) 31(17):2317–30. doi: 10.1097/QAD.0000000000001645
170. Nabatanzi R, Bayigga L, Cose S, Rowland-Jones S, Canderan G, Joloba M, et al. Aberrant natural killer (NK) cell activation and dysfunction among ART-treated HIV-infected adults in an African cohort. Clin Immunol (2019) 201:55–60. doi: 10.1016/j.clim.2019.02.010
171. Costanzo MC, Kim D, Creegan M, Lal KG, Ake JA, Currier JR, et al. Transcriptomic signatures of NK cells suggest impaired responsiveness in HIV-1 infection and increased activity post-vaccination. Nat Commun (2018) 9(1):1212. doi: 10.1038/s41467-018-03618-w
172. Deng K, Pertea M, Rongvaux A, Wang L, Durand CM, Ghiaur G, et al. Broad CTL response is required to clear latent HIV-1 due to dominance of escape mutations. Nature (2015) 517(7534):381–5. doi: 10.1038/nature14053
173. Gorny MK, Conley AJ, Karwowska S, Buchbinder A, Xu JY, Emini EA, et al. Neutralization of diverse human immunodeficiency virus type 1 variants by an anti-V3 human monoclonal antibody. J Virol (1992) 66(12):7538–42. doi: 10.1128/JVI.66.12.7538-7542.1992
174. Burton DR, Pyati J, Koduri R, Sharp SJ, Thornton GB, Parren PW, et al. Efficient neutralization of primary isolates of HIV-1 by a recombinant human monoclonal antibody. Sci (N Y NY) (1994) 266(5187):1024–7. doi: 10.1126/science.7973652
175. Trkola A, Purtscher M, Muster T, Ballaun C, Buchacher A, Sullivan N, et al. Human monoclonal antibody 2G12 defines a distinctive neutralization epitope on the gp120 glycoprotein of human immunodeficiency virus type 1. J Virol (1996) 70(2):1100–8. doi: 10.1128/JVI.70.2.1100-1108.1996
176. Zwick MB, Labrijn AF, Wang M, Spenlehauer C, Saphire EO, Binley JM, et al. Broadly neutralizing antibodies targeted to the membrane-proximal external region of human immunodeficiency virus type 1 glycoprotein gp41. J Virol (2001) 75(22):10892–905. doi: 10.1128/JVI.75.22.10892-10905.2001
177. Trkola A, Kuster H, Rusert P, Joos B, Fischer M, Leemann C, et al. Delay of HIV-1 rebound after cessation of antiretroviral therapy through passive transfer of human neutralizing antibodies. Nat Med (2005) 11(6):615–22. doi: 10.1038/nm1244
178. Mehandru S, Vcelar B, Wrin T, Stiegler G, Joos B, Mohri H, et al. Adjunctive Passive Immunotherapy in Human Immunodeficiency Virus Type 1-Infected Individuals Treated with Antiviral Therapy during Acute and Early Infection. J Virol (2007) 81(20):11016–31. doi: 10.1128/JVI.01340-07
179. Walker LM, Burton DR. Passive immunotherapy of viral infections: ‘super-antibodies’ enter the fray. Nat Rev Immunol (2018) 18(5):297–308. doi: 10.1038/nri.2017.148
180. Caskey M, Klein F, Nussenzweig MC. Broadly neutralizing anti-HIV-1 monoclonal antibodies in the clinic. Nat Med (2019) 25(4):547–53. doi: 10.1038/s41591-019-0412-8
181. Caskey M, Klein F, Lorenzi JC, Seaman MS, West AP Jr., Buckley N, et al. Viraemia suppressed in HIV-1-infected humans by broadly neutralizing antibody 3BNC117. Nature (2015) 522(7557):487–91. doi: 10.1038/nature14411
182. Scheid JF, Horwitz JA, Bar-On Y, Kreider EF, Lu C-L, Lorenzi JCC, et al. HIV-1 antibody 3BNC117 suppresses viral rebound in humans during treatment interruption. Nature (2016) 535(7613):556–60. doi: 10.1038/nature18929
183. Lu C-L, Murakowski DK, Bournazos S, Schoofs T, Sarkar D, Halper-Stromberg A, et al. Enhanced clearance of HIV-1–infected cells by broadly neutralizing antibodies against HIV-1 in vivo. Sci (N Y NY) (2016) 352(6288):1001–4. doi: 10.1126/science.aaf1279
184. Caskey M, Schoofs T, Gruell H, Settler A, Karagounis T, Kreider EF, et al. Antibody 10-1074 suppresses viremia in HIV-1-infected individuals. Nat Med (2017) 23(2):185–91. doi: 10.1038/nm.4268
185. Bar-On Y, Gruell H, Schoofs T, Pai JA, Nogueira L, Butler AL, et al. Safety and antiviral activity of combination HIV-1 broadly neutralizing antibodies in viremic individuals. Nat Med (2018) 24(11):1701–7. doi: 10.1038/s41591-018-0186-4
186. Mendoza P, Gruell H, Nogueira L, Pai JA, Butler AL, Millard K, et al. Combination therapy with anti-HIV-1 antibodies maintains viral suppression. Nature (2018) 561(7724):479–84. doi: 10.1038/s41586-018-0531-2
187. Cohen YZ, Butler AL, Millard K, Witmer-Pack M, Levin R, Unson-O’Brien C, et al. Safety, pharmacokinetics, and immunogenicity of the combination of the broadly neutralizing anti-HIV-1 antibodies 3BNC117 and 10-1074 in healthy adults: A randomized, phase 1 study. PloS One (2019) 14(8):e0219142. doi: 10.1371/journal.pone.0219142
188. Niessl J, Baxter AE, Mendoza P, Jankovic M, Cohen YZ, Butler AL, et al. Combination anti-HIV-1 antibody therapy is associated with increased virus-specific T cell immunity. Nat Med (2020) 26(2):222–7. doi: 10.1038/s41591-019-0747-1
189. Park JH, Geyer MB, Brentjens RJ. CD19-targeted CAR T-cell therapeutics for hematologic malignancies: interpreting clinical outcomes to date. Blood (2016) 127(26):3312–20. doi: 10.1182/blood-2016-02-629063
190. Zhang WY, Wang Y, Guo YL, Dai HR, Yang QM, Zhang YJ, et al. Treatment of CD20-directed Chimeric Antigen Receptor-modified T cells in patients with relapsed or refractory B-cell non-Hodgkin lymphoma: an early phase IIa trial report. Signal Transduct Targ Ther (2016) 1:16002. doi: 10.1038/sigtrans.2016.2
191. Fry TJ, Shah NN, Orentas RJ, Stetler-Stevenson M, Yuan CM, Ramakrishna S, et al. CD22-targeted CAR T cells induce remission in B-ALL that is naive or resistant to CD19-targeted CAR immunotherapy. Nat Med (2018) 24(1):20–8. doi: 10.1038/nm.4441
192. Zhen A, Kitchen S. Stem-cell-based gene therapy for HIV infection. Viruses (2013) 6(1):1–12. doi: 10.3390/v6010001
193. Scholler J, Brady TL, Binder-Scholl G, Hwang W-T, Plesa G, Hege KM, et al. Decade-Long Safety and Function of Retroviral-Modified Chimeric Antigen Receptor T Cells. Sci Trans Med (2012) 4(132):132ra53–ra53. doi: 10.1126/scitranslmed.3003761
194. Bar KJ, Sneller MC, Harrison LJ, Justement JS, Overton ET, Petrone ME, et al. Effect of HIV Antibody VRC01 on Viral Rebound after Treatment Interruption. New Engl J Med (2016) 375(21):2037–50. doi: 10.1056/NEJMoa1608243
195. Hutter G, Nowak D, Mossner M, Ganepola S, Mussig A, Allers K, et al. Long-term control of HIV by CCR5 Delta32/Delta32 stem-cell transplantation. New Engl J Med (2009) 360(7):692–8. doi: 10.1056/NEJMoa0802905
196. Gupta RK, Abdul-Jawad S, McCoy LE, Mok HP, Peppa D, Salgado M, et al. HIV-1 remission following CCR5Delta32/Delta32 haematopoietic stem-cell transplantation. Nature (2019) 568(7751):244–8. doi: 10.1038/s41586-019-1027-4
197. Drake MJ, Bates P. Application of gene-editing technologies to HIV-1. Curr Opin HIV AIDS (2015) 10(2):123–7. doi: 10.1097/COH.0000000000000139
198. Tebas P, Stein D, Tang WW, Frank I, Wang SQ, Lee G, et al. Gene Editing of CCR5 in Autologous CD4 T Cells of Persons Infected with HIV. New Engl J Med (2014) 370(10):901–10. doi: 10.1056/NEJMoa1300662
199. Ishino Y, Shinagawa H, Makino K, Amemura M, Nakata A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J Bacteriol (1987) 169(12):5429–33. doi: 10.1128/JB.169.12.5429-5433.1987
200. Jansen R, Embden JD, Gaastra W, Schouls LM. Identification of genes that are associated with DNA repeats in prokaryotes. Mol Microbiol (2002) 43(6):1565–75. doi: 10.1046/j.1365-2958.2002.02839.x
201. Barrangou R, Fremaux C, Deveau H, Richards M, Boyaval P, Moineau S, et al. CRISPR provides acquired resistance against viruses in prokaryotes. Sci (N Y NY) (2007) 315(5819):1709–12. doi: 10.1126/science.1138140
202. Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Sci (N Y NY) (2012) 337(6096):816–21. doi: 10.1126/science.1225829
203. Ebina H, Misawa N, Kanemura Y, Koyanagi Y. Harnessing the CRISPR/Cas9 system to disrupt latent HIV-1 provirus. Sci Rep (2013) 3(1):2510. doi: 10.1038/srep02510
204. Qu X, Wang P, Ding D, Li L, Wang H, Ma L, et al. Zinc-finger-nucleases mediate specific and efficient excision of HIV-1 proviral DNA from infected and latently infected human T cells. Nucleic Acids Res (2013) 41(16):7771–82. doi: 10.1093/nar/gkt571
205. Hu W, Kaminski R, Yang F, Zhang Y, Cosentino L, Li F, et al. RNA-directed gene editing specifically eradicates latent and prevents new HIV-1 infection. Proc Natl Acad Sci U States America (2014) 111(31):11461–6. doi: 10.1073/pnas.1405186111
206. Adams Waldorf KM, Stencel-Baerenwald JE, Kapur RP, Studholme C, Boldenow E, Vornhagen J, et al. Fetal brain lesions after subcutaneous inoculation of Zika virus in a pregnant nonhuman primate. Nat Med (2016) 22(11):1256–9. doi: 10.1038/nm.4193
207. Mousseau G, Kessing CF, Fromentin R, Trautmann L, Chomont N, Valente ST. The Tat Inhibitor Didehydro-Cortistatin A Prevents HIV-1 Reactivation from Latency. mBio (2015) 6(4):e00465. doi: 10.1128/mBio.00465-15
208. Laspia MF, Rice AP, Mathews MB. HIV-1 Tat protein increases transcriptional initiation and stabilizes elongation. Cell (1989) 59(2):283–92. doi: 10.1016/0092-8674(89)90290-0
209. Mancebo HS, Lee G, Flygare J, Tomassini J, Luu P, Zhu Y, et al. P-TEFb kinase is required for HIV Tat transcriptional activation in vivo and in vitro. Genes Dev (1997) 11(20):2633–44. doi: 10.1101/gad.11.20.2633
210. Wei P, Garber ME, Fang S-M, Fischer WH, Jones KA. A Novel CDK9-Associated C-Type Cyclin Interacts Directly with HIV-1 Tat and Mediates Its High-Affinity, Loop-Specific Binding to TAR RNA. Cell (1998) 92(4):451–62. doi: 10.1016/S0092-8674(00)80939-3
211. Mahmoudi T, Parra M, Vries RG, Kauder SE, Verrijzer CP, Ott M, et al. The SWI/SNF chromatin-remodeling complex is a cofactor for Tat transactivation of the HIV promoter. J Biol Chem (2006) 281(29):19960–8. doi: 10.1074/jbc.M603336200
212. Tréand C, du Chéné I, Brès V, Kiernan R, Benarous R, Benkirane M, et al. Requirement for SWI/SNF chromatin-remodeling complex in Tat-mediated activation of the HIV-1 promoter. EMBO J (2006) 25(8):1690–9. doi: 10.1038/sj.emboj.7601074
213. Benkirane M, Chun RF, Xiao H, Ogryzko VV, Howard BH, Nakatani Y, et al. Activation of integrated provirus requires histone acetyltransferase. p300 and P/CAF are coactivators for HIV-1 Tat. J Biol Chem (1998) 273(38):24898–905. doi: 10.1074/jbc.273.38.24898
214. Col E, Caron C, Seigneurin-Berny D, Gracia J, Favier A, Khochbin S. The histone acetyltransferase, hGCN5, interacts with and acetylates the HIV transactivator, Tat. J Biol Chem (2001) 276(30):28179–84. doi: 10.1074/jbc.M101385200
215. Marzio G, Tyagi M, Gutierrez MI, Giacca M. HIV-1 Tat transactivator recruits p300 and CREB-binding protein histone acetyltransferases to the viral promoter. Proc Natl Acad Sci (1998) 95(23):13519–24. doi: 10.1073/pnas.95.23.13519
216. Mousseau G, Clementz MA, Bakeman WN, Nagarsheth N, Cameron M, Shi J, et al. An analog of the natural steroidal alkaloid cortistatin A potently suppresses Tat-dependent HIV transcription. Cell Host Microbe (2012) 12(1):97–108. doi: 10.1016/j.chom.2012.05.016
217. Mousseau G, Aneja R, Clementz MA, Mediouni S, Lima NS, Haregot A, et al. Resistance to the Tat Inhibitor Didehydro-Cortistatin A Is Mediated by Heightened Basal HIV-1 Transcription. mBio (2019) 10(4):e01750–18. doi: 10.1128/mBio.01750-18
218. Huntington ND, Legrand N, Alves NL, Jaron B, Weijer K, Plet A, et al. IL-15 trans-presentation promotes human NK cell development and differentiation in vivo. J Exp Med (2009) 206(1):25–34. doi: 10.1084/jem.20082013
219. Katano I, Nishime C, Ito R, Kamisako T, Mizusawa T, Ka Y, et al. Long-term maintenance of peripheral blood derived human NK cells in a novel human IL-15- transgenic NOG mouse. Sci Rep (2017) 7(1):17230. doi: 10.1038/s41598-017-17442-7
220. Matsuda M, Ono R, Iyoda T, Endo T, Iwasaki M, Tomizawa-Murasawa M, et al. Human NK cell development in hIL-7 and hIL-15 knockin NOD/SCID/IL2rgKO mice. Life Sci Alliance (2019) 2(2):e201800195. doi: 10.26508/lsa.201800195
221. Demetrius L. Of mice and men. When it comes to studying ageing and the means to slow it down, mice are not just small humans. EMBO Rep (2005) 6 Spec No(Suppl 1):S39–44. doi: 10.1038/sj.embor.7400422
222. Perlman RL. Mouse models of human disease: An evolutionary perspective. Evol Med Public Health (2016) 2016(1):170–6. doi: 10.1093/emph/eow014
223. Nguyen TL, Vieira-Silva S, Liston A, Raes J. How informative is the mouse for human gut microbiota research? Dis Model Mech (2015) 8(1):1–16. doi: 10.1242/dmm.017400
224. Hugenholtz F, de Vos WM. Mouse models for human intestinal microbiota research: a critical evaluation. Cell Mol Life Sci (2018) 75(1):149–60. doi: 10.1007/s00018-017-2693-8
225. Saxena D, Li Y, Yang L, Pei Z, Poles M, Abrams WR, et al. Human microbiome and HIV/AIDS. Curr HIV/AIDS Rep (2012) 9(1):44–51. doi: 10.1007/s11904-011-0103-7
226. Zilberman-Schapira G, Zmora N, Itav S, Bashiardes S, Elinav H, Elinav E. The gut microbiome in human immunodeficiency virus infection. BMC Med (2016) 14(1):83. doi: 10.1186/s12916-016-0625-3
227. Liu J, Williams B, Frank D, Dillon SM, Wilson CC, Landay AL. Inside Out: HIV, the Gut Microbiome, and the Mucosal Immune System. J Immunol (Baltimore Md 1950) (2017) 198(2):605–14. doi: 10.4049/jimmunol.1601355
228. Crakes KR, Jiang G. Gut Microbiome Alterations During HIV/SIV Infection: Implications for HIV Cure. Front Microbiol (2019) 10:1104–. doi: 10.3389/fmicb.2019.01104
Keywords: humanized mice, BLT, DRAG, HIV-1 infection, viral latency, latency reversal, immunotherapy, gene therapy
Citation: Abeynaike S and Paust S (2021) Humanized Mice for the Evaluation of Novel HIV-1 Therapies. Front. Immunol. 12:636775. doi: 10.3389/fimmu.2021.636775
Received: 02 December 2020; Accepted: 01 March 2021;
Published: 01 April 2021.
Edited by:
Qingfeng Chen, Institute of Molecular and Cell Biology (A*STAR), SingaporeReviewed by:
Mangai Asokan, Independent Researcher, Bengaluru, IndiaNamal P. M. Liyanage, The Ohio State University, United States
Copyright © 2021 Abeynaike and Paust. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Silke Paust, cGF1c3RAc2NyaXBwcy5lZHU=