 Matheus Braga1
Matheus Braga1 Fernanda Formaggi Lara-Armi1
Fernanda Formaggi Lara-Armi1 Janisleya Silva Ferreira Neves1Marco Antônio Rocha-Loures2Mariana de Souza Terron-Monich3Larissa Danielle Bahls-Pinto3
Janisleya Silva Ferreira Neves1Marco Antônio Rocha-Loures2Mariana de Souza Terron-Monich3Larissa Danielle Bahls-Pinto3 Quirino Alves de Lima Neto1,3Joana Maira Valentini Zacarias1,3
Quirino Alves de Lima Neto1,3Joana Maira Valentini Zacarias1,3 Ana Maria Sell1,3
Ana Maria Sell1,3 Jeane Eliete Laguila Visentainer1,3*
Jeane Eliete Laguila Visentainer1,3*- 1Post Graduation Program in Biosciences and Physiopathology, Department of Clinical Analysis and Biomedicine, State University of Maringá, Paraná, Brazil
- 2Rheumatology Division, Department of Medicine, State University of Maringá, Paraná, Brazil
- 3Immunogenetics Laboratory, Department of Basic and Health Science, State University of Maringá, Paraná, Brazil
Ankylosing spondylitis (AS) is a chronic autoimmune inflammatory disease that mainly affects the axial and sacroiliac joints. Single-nucleotide polymorphisms (SNPs) in genes encoding cytokines have been associated with AS, which can interfere with the production of these cytokines and contribute to the development of AS. In order to contribute to a better understanding of the pathology of AS, our objective was to investigate a possible association of the IL10 −1082 A>G SNP (rs1800896) with AS and to evaluate the serum levels of TNF-α, IL-10, IL-17A, and IL-17F in AS patients and controls comparing them with their respective genotypes (TNF rs1800629, IL10 rs1800896, IL17A rs2275913, and IL17F rs763780). Patients and controls were selected from the Maringá University Hospital and the Maringá Rheumatism Clinic, in Paraná State, Southern Brazil, and they were diagnosed by the ASAS Criteria. In total, 149 patients and 169 controls were genotyped for the IL10 −1082 A>G polymorphism using a polymerase chain reaction with sequence specific primers (PCR-SSP); the measurement of TNF-α serum levels was performed through the immunofluorimetric test and IL-10, IL-17A, and IL-17F using an ELISA test. There was a high frequency of the IL10 −1082 G allele in AS patients compared with controls with an odds ratio of 1.83 and 95% confidence interval of 1.32 to 2.54, and a significant difference in the genotype frequencies of the IL10 −1082 A/G+G/G between patients and healthy controls, with an odds ratio of 3.01 and 95% confidence interval of 1.75 to 5.17. In addition, increased serum levels of IL-10 were observed in AS patients: 2.38 (IQR, 0.91) pg/ml compared with controls 1.72 (IQR 0.93) pg/ml (P = 0.01). Our results also showed an association between IL17F rs763780 C/T+T/T genotypes and increased serum levels of IL-17F in patients with AS and also in controls. We can conclude that patients with the A/G and G/G genotypes for −1082 A>G (rs1800896) in the IL10 gene are three times more likely to develop AS, that the serum level of IL-10 was higher in AS patients and that the IL17F rs763780 polymorphism can affect the levels of IL-17F in the serum of patients and controls in the same way.
Introduction
Ankylosing spondylitis (AS) is a chronic autoimmune inflammatory disease that mainly affects the axial and sacroiliac joints, causing severe pain. In more advanced cases, this inflammation can lead to fibrosis and calcification, spinal fusion, and consequently, a loss of flexibility (1, 2). Although the major histocompatibility complex (MHC) class I allele HLA-B*27 is strongly associated with AS, genome-wide association studies have revealed a number of other risk factors for the disease, including several innate immune-related pathways and cytokines (3).
IL-10 is an immunomodulatory cytokine encoded by the IL10 gene on chromosome 1q31–32, containing five exons separated by four introns (4). Polymorphisms located in the 5′-flanking region of the IL10 gene, at positions −1082 A>G, −819 T>C, and −592 A>C, are known to be involved in regulating the production of IL-10, with the first being the best characterised of them (5).
Three single nucleotide polymorphisms (SNPs) in genes that encode inflammatory cytokines have been associated with AS, in a previous study by our research group (TNF rs1800629, IL17A rs2275913, and IL17F rs763780) (6). These polymorphisms can interfere with the level of production of these cytokines and contribute to the development of AS (7–10).
Inflammatory cytokines, such as tumour necrosis factor alpha (TNF-α) and interleukin 17 (IL-17), are known to be involved in AS (11, 12). TNF-α is a key component of the immune system and a potent pro-inflammatory cytokine that is highly produced after infection or tissue damage (13) and is present in high concentrations in patients with AS (12). IL-17 is a pro-inflammatory cytokine that contributes to the pathogenesis of several inflammatory diseases. The IL-17 family consists of six structurally related cytokines (IL-17A, IL-17B, IL-17C, IL-17D, IL-17E, and IL-17F) (14). In rheumatological diseases, this interleukin causes cartilage damage in an experimental animal model (15).
The pathogenesis of AS has not been fully elucidated, and some cytokines may play a key role in the disease. In order to elucidate the pathology of AS and identify new genetic markers, we investigated a possible association of the IL10 SNP −1082 A>G (rs1800896) with AS, and evaluated the serum levels of TNF-α, IL-10, IL-17A, and IL-17F in AS patients and controls compared with their respective genotypes (TNF rs1800629, IL17A rs2275913, IL17F rs763780, and IL10 rs1800896).
Methodology
Study Population
Unrelated patients were diagnosed by the ASAS Criteria 2009 (16) for axial spondyloarthritis (SpA) and the ASAS Criteria 2011 (17) for peripheral SpA. Patients were selected from the Maringá University Hospital-Maringá State University and the Maringá Rheumatism Clinic, in Paraná State, Southern Brazil. All patients had magnetic resonance imaging of the sacroiliac joints and were evaluated for the presence of HLA-B*27. In addition, individuals were selected to form the control group, following the criteria for inclusion: no autoimmune and/or rheumatic diseases, unrelated to the patient group, and belonging to the same ethnic group as the patients, and from the same region. This research was approved by the Standing Committee of Ethics of the State University of Maringá (CAAE 687.222/2014), and all participants signed an informed consent form.
Sample Collection and DNA Extraction
Peripheral blood was collected from case and control groups using tubes without anticoagulants for the determination of serum levels of cytokines and using EDTA-anticoagulant for DNA extraction. Genomic DNA was extracted from blood samples using the extraction kit Biopur® (BIOMETRIX Diagnostic, Curitiba, Paraná, Brazil), according to the manufacturer’s recommendations.
Determination of IL10 Gene Polymorphism
One hundred and forty-nine patients and 169 controls were genotyped for the IL10 −1082 A>G polymorphism using a polymerase chain reaction with sequence specific primers (PCR-SSP), and a methodology adapted from (18). The primer sequences used are shown in Table 1.
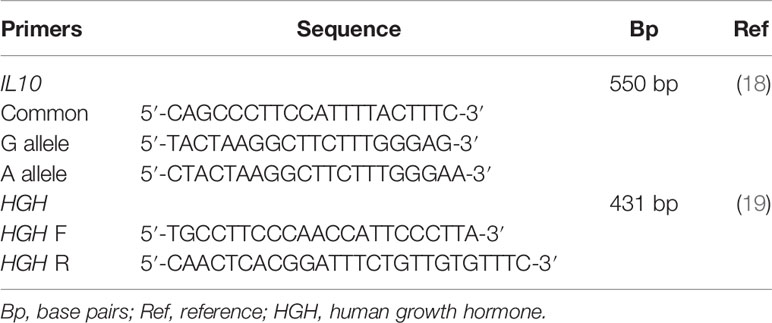
Table 1 Primer sequences.
The protocol was adapted to a final volume of 10 µL of the reaction mixture containing 50 ng of DNA template, 1.8 ng/μM of each IL10 primer, 0.75 ng/µM of each Human growth hormone (HGH) for internal control, 0.19 mM of dNTP mixture, 1.5 mM of MgCl2 for allele G reaction and 2.0 mM for allele A reaction, 0.47× Reaction Buffer, and 0.55 U of Taq DNA polymerase.
PCR conditions for amplification included an initial melting step of 5 min at 94°C, followed by 32 cycles of 40 s at 95°C, 50 s at 65°C, 40 s at 72°C, and a final elongation at 72°C for 7 min. After amplification, PCR products were visualised in a 2% agarose gel stained with SYBR™ Safe DNA Gel Stain dye (Invitrogen, Carlsbad, CA, USA) using the Quantum ultraviolet gel documentation system.
For methodology validation, samples previously genotyped in our laboratory using a commercial kit (One Lambda Cytokine Genotyping Primer Pack, One Lambda) were used as positive controls for each of the three genotypes obtained (A/A, G/A, and G/G) (Figure 1).
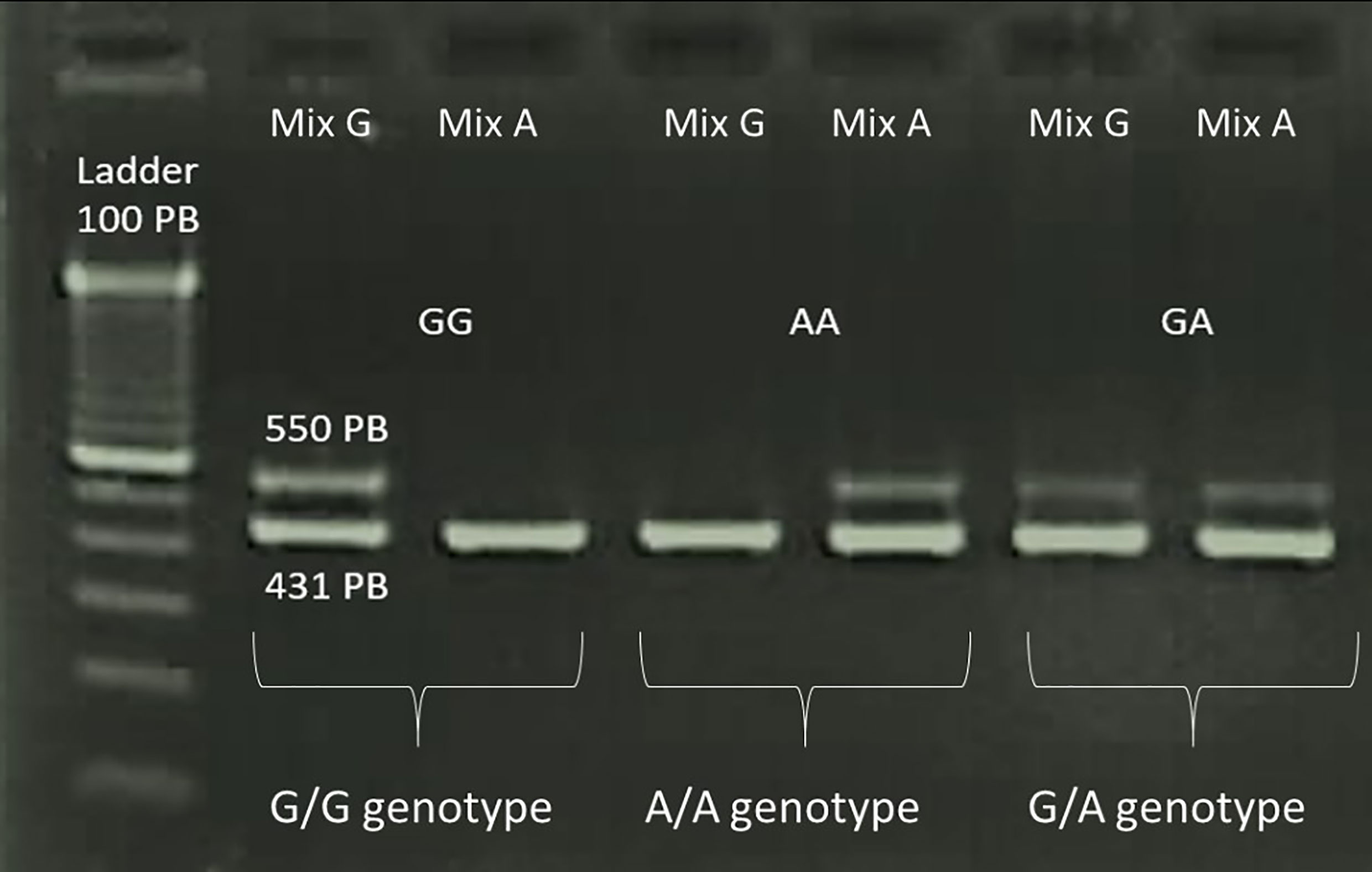
Figure 1 Amplified products of the PCR-SSP assay for analysing the −1082 A>G polymorphism of IL10 visualised in 2% agarose gel. Bands of 550 bp correspond to the IL10 fragment and bands of 431 bp correspond to the HGH fragment, which was used as an internal control for the reaction.
Determination of Serum Levels of Cytokines
The samples selected for cytokine measurement were randomly chosen, paired by age, gender, and genotype. For IL-10, 28 AS patients and 20 controls genotyped for IL10 rs1800896 were selected, with 12 being A/A, and 16 being A/G+G/G for AS and 10 being A/A, and 10 being A/G+A/A for controls. TNF, IL17A, and IL17F gene polymorphisms were previously determined by our group for the same samples and the polymorphisms analysed were TNF rs1800629, IL17A rs2275913, and IL17F rs763780 (6). 35 patients were selected for the analysis of TNF-α levels, with 17 being G/G, and 20 being G/A+A/A for AS, and 15 controls were also included, of which 8 were G/G and 7 were G/A+A/A. For the IL-17A analysis, 45 AS patients, including 21 G/G and 24 G/A+A/A, and 15 controls, including 5 G/G and 10 G/A+A/A, were selected. For the IL-17F analysis, 40 patients were selected, with 24 being T/T, and 16 being C/T+C/C; for the 14 controls, there were seven T/T and seven C/T+C/C. The characteristics of AS patients and controls selected for measurement of cytokines are shown in Table 2. There were no significant differences between the patients with AS and controls for gender and age distribution for any group of cytokines (P > 0.05).
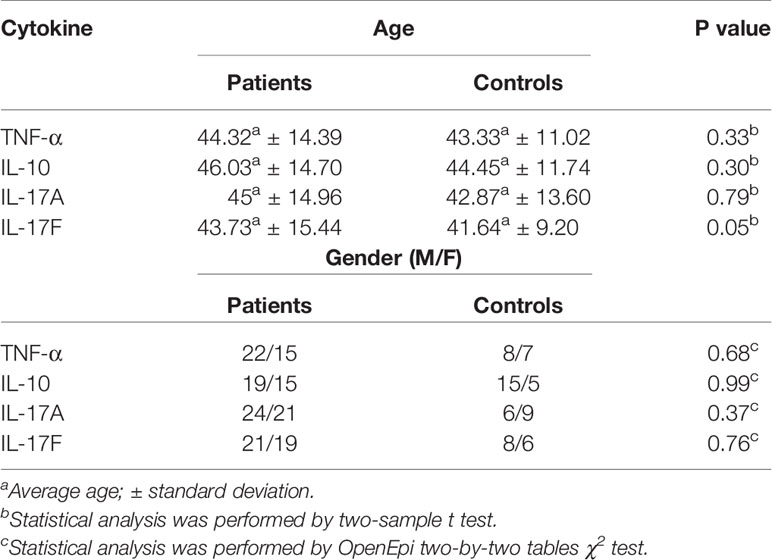
Table 2 Characteristics of AS patients and controls selected for cytokines measurement.
The determination of serum levels of TNF-α was performed using the Kit Luminex Technology with the ProcartaPlex™ Immunoassay (Invitrogen, ThermoFisher Scientific, Inc., Burlington, Ontario, Canada). The determination of IL-10 serum levels was performed using the human IL-10 uncoated ELISA kit (Affymetrix eBioscience, North America, USA). The determination of serum levels of IL-17A was performed using the Human IL-17A platinum ELISA Kit (Affymetrix eBioscience, North America, USA), while the determination of serum levels of IL-17F cytokine was performed using the Human IL-17F ELISA Kit (Affymetrix eBioscience, North America, USA). The minimum detection limit for each kit was 0.5 picograms per ml (pg/ml) for TNF-α, 1.6 pg/ml for IL-10, 0.5 pg/ml for IL-17A, and 3.3 pg/ml for IL-17F.
Statistical Analysis
The QUANTO 1.2.4 software (http://biostats.usc.edu/software) was used to calculate the sample size using the following values: prevalence of the less frequent allele 0.30%, population risk 0.01%, power 0.80, and OR >3.0. For genetic statistical analyses and to estimate the distribution of genotype frequencies according to the Hardy-Weinberg equilibrium (HWE), we applied the software SNPStats (available at: https://www.snpstats.net/start.htm) (20), which was also used for detecting the odds ratio balance, with a 95% confidence interval (CI) after adjustment for age and HLA*B27 marker. The Akaike information criterion (AIC) was used to choose the inheritance model that best fits the data. The OpenEpi program was used to calculate the allele association using the two-by-two tables test (https://www.openepi.com/Menu/OE_Menu.htm) (21).
To compare the serum levels of cytokines with their respective genotypes, the BioEstat 5.0 program was used. The Shapiro-Wilk test was used to calculate the normality of the samples and the Mann-Whitney test to compare the samples. All values of serum concentration cytokines were expressed as median and Interquartile range (IQR); differences were considered statistically significant if P<0.05.
Results
The characteristics and clinical data of individuals genotyped for the IL10 −1082 A>G polymorphism are shown in Table 3. There were no significant differences between AS patients and controls for gender distribution (P>0.05). HLA-B*27 was previously analysed using PCR-SSP according to the method published by our research group (22) and it was more frequent in patients with AS than in controls (P<0.0001).
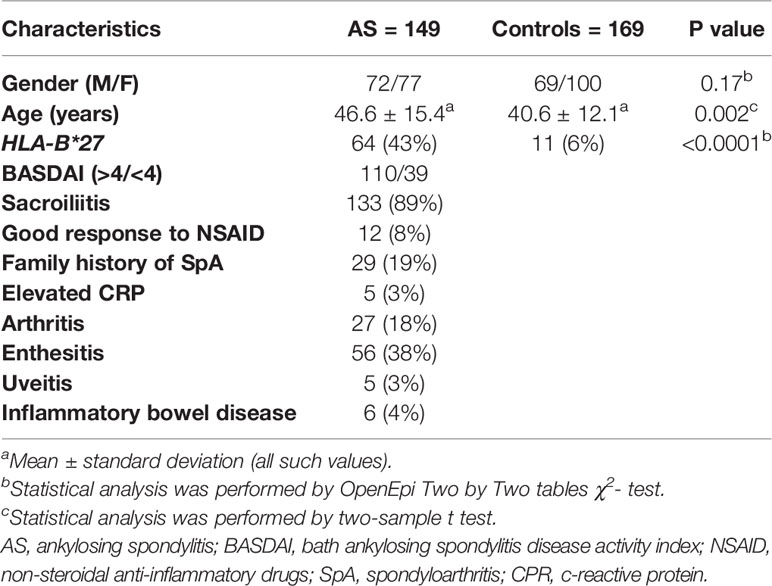
Table 3 Characteristics of AS patients and healthy controls.
Determination of the IL10 gene polymorphism by PCR-SSP demonstrated an increased frequency of the −1082 G allele in AS patients compared to controls (Table 4). In addition, there was significant difference in the genotype frequency of the IL10 −1082 A>G polymorphism between patients and controls. To estimate the association between the genotype and AS occurrence, the disease odds ratio (OR) and 95% confidence interval (CI) were assessed and adjusted for age and HLA*B27 marker. The results suggest that the −1082 A/G and G/G genotypes were associated with disease in a dominant model with an increased risk for AS (OR, 3.01; 95% CI, 1.75–5.17). The IL10 genotype frequency distributions were in Hardy-Weinberg equilibrium in patients and controls (P>0.05).
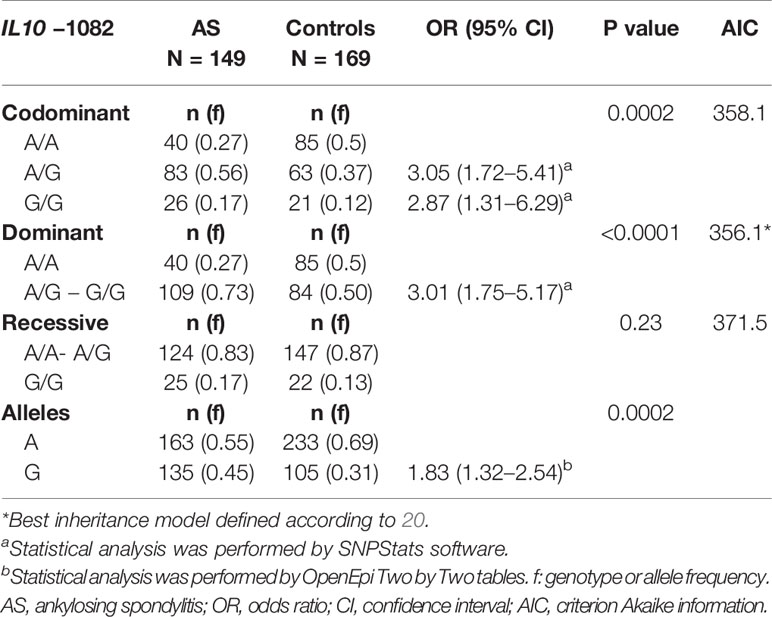
Table 4 Allele and genotype frequency distribution for IL10 −1082 A>G and the association with the risk for ankylosing spondylitis, regardless of HLA-B*27 marker and age in different genetic models.
Results of the analysis of the association between IL10 −1082 A>G (rs1800896) polymorphism with different clinical data of AS are shown in Table 5. There was no statistical difference between IL10 genotypes or alleles in relation to clinical data of AS. The smaller number of patients in each subgroup could be related to these results.
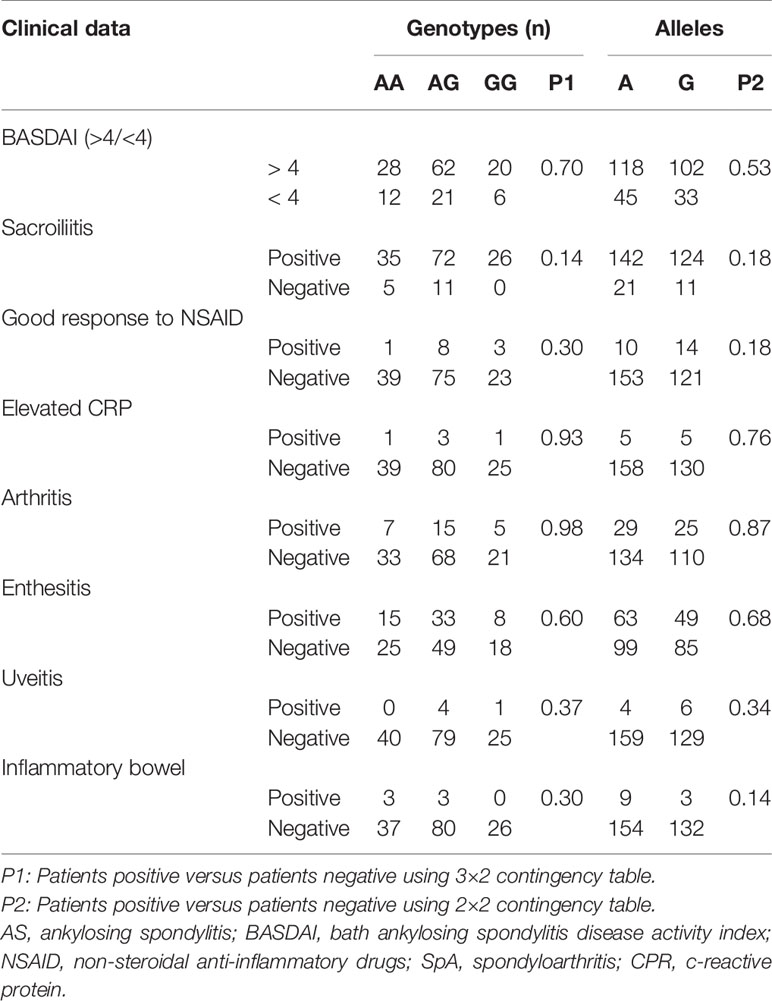
Table 5 Analysis of IL10 gene polymorphisms (rs1800896) in AS patients separated according to clinical data.
The analyses of serum cytokine levels in AS patients and controls are shown in Figure 2. Higher serum levels of IL-10 were observed in patients, as the levels of IL-10 was 2.38 (IQR 0.91) pg/mL in patients and 1.72 (IQR, 0.93) pg/ml in controls (P=0.01) (Figure 2B). There were no statistically significant differences in the serum levels of the other cytokines when patients were compared to controls. The serum levels of the cytokines in patients were as follows: TNF-α 2.14 (IQR, 3.65) pg/ml, IL-17A 22.12 (IQR, 51.97) pg/ml, and IL-17F 100.13 (IQR, 1273.65) pg/ml. For controls, the serum levels were as follows: TNF-α 1.39 (IQR, 3.14) pg/ml; IL-17A 33.50 (IQR, 339) pg/ml, and IL-17F 251.04 (IQR, 765.45) pg/ml.

Figure 2 Comparison between patients and controls for cytokine levels. (A) Comparison of TNF-α levels between patients and controls; (B) Comparison of IL-10 levels between patients and controls; (C) Comparison of IL-17A levels between patients and controls; (D) Comparison of IL-17F levels between patients and controls. AS, ankylosing spondylitis; ns, not significant.
The analyses of TNF-α, IL-10, IL-17A, and IL-17F serum cytokine levels with genotypes in AS patients and controls are shown in Figure 3. Higher serum levels of IL-17F were observed in patients with the IL17F rs763780 T/C+C/C genotypes [796.04 (IQR, 1496.23) pg/ml compared to patients with the T/T genotype 43.77 (IQR, 384.1) pg/ml] (p=0.03) (Figure 3H). The same association was found for controls: T/C+C/C 570.10 (IQR, 1325.23) pg/ml compared with T/T 25.36 (IQR, 142.73) pg/ml (P=0.02) (Figure 3I).
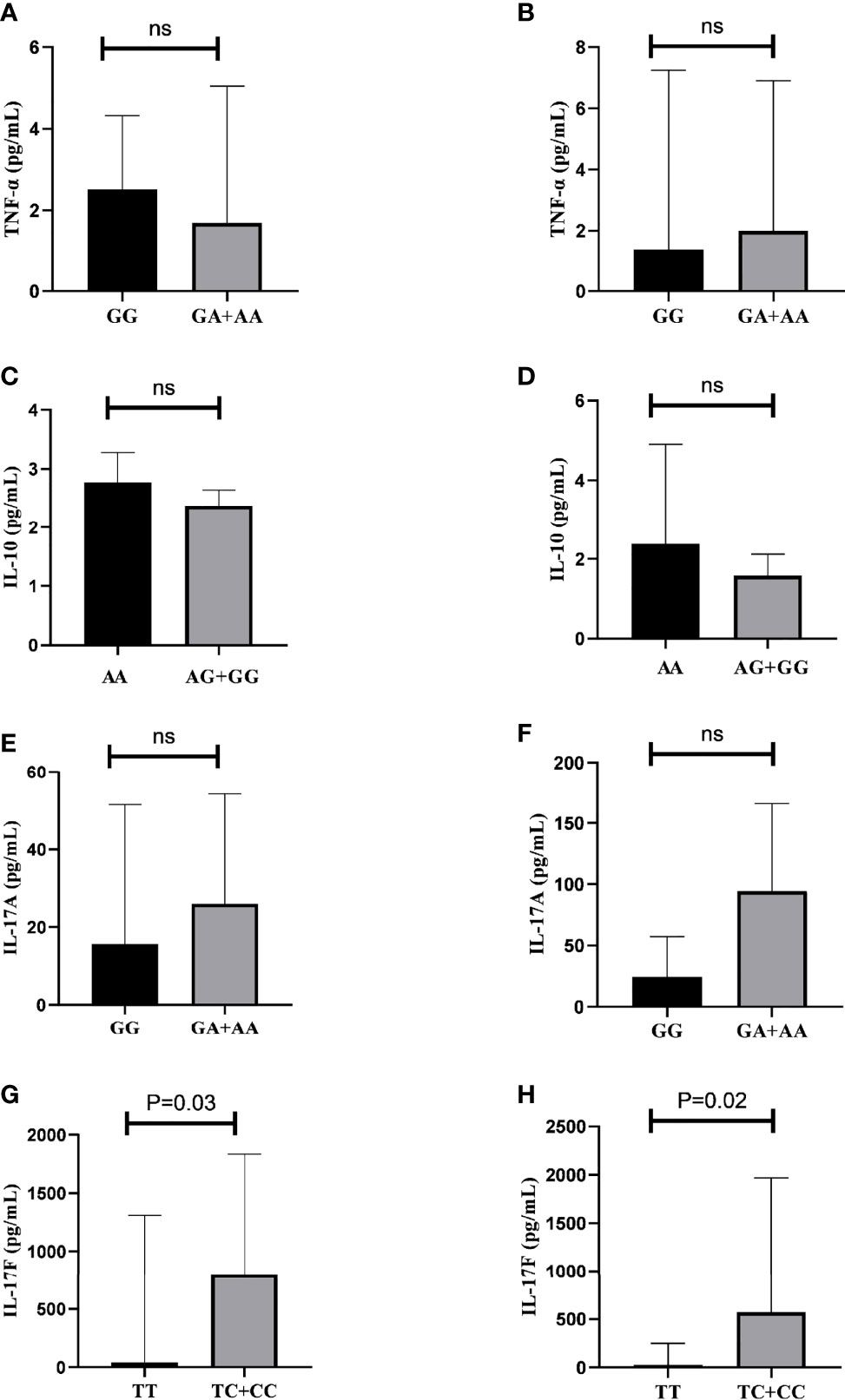
Figure 3 Relation between serum cytokine levels and genotypes in AS patients and controls. (A) Comparison of TNF-α levels in patients with TNF rs1800629 G/G vs. G/A+AA genotypes. (B) Comparison of TNF-α levels in controls with TNF rs1800629 G/G vs. G/A+A/A genotypes. (C) Comparison of IL-10 levels in patients with IL10 rs1800896 A/A vs. A/G+G/G genotypes. (D) Comparison of IL-10 levels in controls with IL10 rs1800896 A/A vs. A/G+G/G genotypes. (E) Comparison of IL-17A levels in patients with IL17A rs2275913 G/A+A/A vs. G/G genotypes. (F) Comparison of IL-17A levels in controls with IL17A rs2275913 G/A+A/A vs. G/G genotypes. (G) Comparison of IL-17F levels in patients with IL17F rs763780 T/C+C/C vs. T/T genotypes. (H) Comparison of IL-17F levels in controls with IL17F rs763780 T/C+C/C vs. T/T genotypes; ns, not significant.
No statistically significant difference was found for the other cytokines. TNF-α serum level in AS patients carrying TNF rs1800629 G/A+A/A was 1.70 pg/ml (IQR, 4.71) and 2.51 (IQR, 2.21) pg/ml for those carrying G/G (Figure 3A). For controls, it was 2.00 (IQR, 4.73) pg/ml for those carrying G/A+A/A and 1.39 (IQR, 1.37) pg/ml for G/G (Figure 3B). Serum levels of IL-10 in AS patients carrying IL10 rs1800896 A/G +A/A were 2.38 (IQR, 0.45) pg/ml and 2.77 (IQR 0.91) pg/mL in those carrying A/A (Figure 3C); for controls carrying A/G +A/A, it was 1.58 (IQR, 0.68) pg/ml and for A/A 2.12 (IQR 1.19) pg/ml (Figure 3D). Serum levels of IL-17A were 26.00 (IQR, 51.6) pg/ml in AS patients carrying the IL17A rs2275913 G/A+A/A genotypes and 15.63 (IQR, 49.6) pg/ml for those carrying the G/G genotype (Figure 3E); for controls, it was 94.63 (IQR, 129.23) pg/ml for individuals carrying G/A+A/A and 24.37 (IQR, 30.43) pg/ml for those carrying G/G (Figure 3F).
Discussion
In this study, we found that the IL10 −1082 G allele and the A/G+G/G genotypes (in a dominant inheritance model) were associated with the risk of AS. It is worth mentioning that our analyses of the IL10 -1082 polymorphism were adjusted for age and presence of HLA-B*27, and that we found this association to be independent of these factors, which makes the evidence of an association between the IL10 gene and AS more powerful.
Despite the population difference, another study conducted with a Chinese population also found an association with the same polymorphism and AS (23). The Paraná population mainly consists of people of European origin (80.6%), with a small contribution from African (12.5%), and Indigenous (7.0%) populations (24, 25). Hence, patients and healthy controls in our study were classified as mixed ethnic groups, according to phenotypic characteristics.
Our study also showed higher serum levels of IL-10 in AS patients compared to controls, similarly to the Chinese study (23). Increased levels of IL-10 and IL-10–positive T cells were detected in synovial fluid from AS patients in other studies (26, 27). Although these results seem controversial because of IL-10 being an anti-inflammatory cytokine, a way to better understand the role of IL-10 in the pathogenesis of disease can be through the M2 macrophages. Some studies have shown an increase in M2 CD163+ in synovial biopsies and inflamed colon of the patients with AS, and CD163 synovial over-expression has been shown to be correlated with disease (28, 29); in this context, IL-10 can alter the phenotype of macrophages from M1 to M2 (30). Furthermore, it has been proposed that the M2 macrophages express less IFN-γ, leading to an increased Th17 response since IFN-γ can suppress IL-17 expression (31), and it has been reported that the IL-17 is one of the main cytokines involved in the pathogenesis of AS (28, 29).
In this study, we found no statistical differences between the levels of TNF-α, IL-17A, and IL-17F among patients and controls, despite some studies having shown an increase in inflammatory cytokines, such as IL-17 and TNF-α in AS (32, 33). We must take into account that our patients were being treated with non-steroidal anti-inflammatory drugs (NSAIDs), and anti-TNF therapy for longer than 6 months.
In order to better understand the influence of SNPs associated with AS, IL10 rs1800896 in the present study, as well as TNF rs1800629, IL17A rs2275913, IL17F rs763780 that were associated with the risk of developing AS in a previous study by our group (6), we compared the serum levels of the cytokines between the risk genotypes and non-risk genotypes in patients and controls.
With regard to TNF-α, we found no differences in the serum levels of this cytokine between the TNF rs1800629 genotype G/G compared to the risk genotypes G/A+A/A. The G/A+A/A genotypes have also been reported to be major producers of TNF-α in congenital heart disease (34). The A allele was found to be the higher producer of TNF-α in an in vitro study (35). Although we did not observe this association, we should also consider the use of anti-TNF in the treatment.
Many studies show that the IL10 −1082 genotypes A/G+G/G were the largest producers of IL-10 (23, 36–38). Despite the association of A/G+G/G genotypes with AS and increased levels of IL-10 in patients, we did not find any association between these genotypes and the levels of IL-10, suggesting that other SNPs could influence the production of the cytokine.
We also found no differences in the serum levels of IL-17A cytokine between G/G and G/A+A/A genotypes. The genotypes G/A and A/A have been reported to be associated with the increased production of IL-17A in a recent study on coronary disease (39). However, this relationship is controversial, as other studies have shown an increase in IL-17A in patients with the A/A genotype (40), a lack of association with this SNP and production (41), and high levels in individuals carrying the genotypes GG+GA (42). These differences could suggest that the polymorphism (rs2275913) does not influence IL-17A levels.
The assessment of the association between IL-17F levels and the IL17F T>C (rs763780) polymorphism showed an association between increased IL-17F production and the T/C+C/C genotypes in patients and controls. These results suggest that variations in the sequence of the specific nucleotides of IL17F can have a direct effect on the production of the encoded protein, IL-17F. Another study with psoriasis also reported a higher level of IL-17F in patients with the T/C+C/C genotypes (43).
However, we must take into account some limitations of our study. It is a cross-sectional study, although the association between the polymorphism (rs763780) and IL-17F levels has been confirmed. It was not possible to establish a causal relationship, requiring experimental studies and analysis of cytokines in tissue and synovial fluid samples. Another weakness of our study was the impossibility of using patients who had not yet used any type of medication.
We can conclude that the IL10 −1082 G allele and the A/G+G/G genotypes (in a dominant inheritance model) were risk factors for AS, regardless of HLA-B*27 and age, and that serum IL-10 levels were increased in patients with AS compared to controls.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics Statement
This research was approved by the standing committee of ethics of the State University of Maringá (CAAE 687.222/2014). The patients/participants provided their written informed consent to participate in this study.
Author Contributions
MB executed the research protocol, analyzed the data, and wrote the manuscript. MT-M, LB-P and JZ contributed to the writing and critical analysis of the manuscript. JN and MR-L contributed to the collection of data, sample and classification of patients. FL-A contributed to the implementation of the protocols. QN supervised the analyses and the execution of this research. AS and JV developed the original idea and provided intellectual input and critical analysis of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Laboratory of Immunogenetics (Proc. n.1589/2017-CSD-UEM).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors wish to thank the patients and volunteers for their valuable participation in the study, the Post-Graduation Program in Biosciences and Physiopathology, UEM, CAPES, Fundação Araucária-PR, and CNPq.
References
1. Pedersen SJ, Maksymowych WP. The Pathogenesis of Ankylosing Spondylitis: An Update. Curr Rheumatol Rep (2019) 21:1–10. doi: 10.1007/s11926-019-0856-3
2. Zhu W, He X, Cheng K, Zhang L, Chen D, Wang X, et al. Ankylosing Spondylitis: Etiology, Pathogenesis, and Treatments. Bone Res (2019) 7:1–16. doi: 10.1038/s41413-019-0057-8
3. Vanaki N, Aslani S, Jamshidi A, Mahmoudi M. Role of Innate Immune System in the Pathogenesis of Ankylosing Spondylitis. BioMed Pharmacother (2018) 105:130–43. doi: 10.1016/j.biopha.2018.05.097
4. Moore KW, de Waal Malefyt R, Coffman RL, O’Garra A. Interleukin-10 and the interleukin-10 Receptor. Annu Rev Immunol (2001) 19:683–765. doi: 10.1146/annurev.immunol.19.1.683
5. Turner D, Williams DM, Sankaran D, Lazarus M, Sinnott PJ, Hutchinson IV. An Investigation of Polymorphism in the Interleukin-10 Gene Promoter. Eur J immunogenetics (1997) 24:1–8. doi: 10.1111/j.1365-2370.1997.tb00001.x
6. Rocha Loures MA, Macedo LC, Reis DM, Oliveira CF, Meneguetti JL, Martines GF, et al. Influence of TNF and IL17 Gene Polymorphisms on the Spondyloarthritis Immunopathogenesis, Regardless of HLA-B27, in a Brazilian Population. Mediators Inflammation (2018) 2018:1395823. doi: 10.1155/2018/1395823
7. Sode J, Bank S, Vogel U, Andersen PS, Sørensen SB, Bojesen AB, et al. Genetically Determined High Activities of the TNF-Alpha, IL23/IL17, and NFkB Pathways Were Associated With Increased Risk of Ankylosing Spondylitis. BMC Med Genet (2018) 19:1–16. doi: 10.1186/s12881-018-0680-z
8. Vargas-Alarcón G, Casasola-Vargas J, Rodríguez-Pérez JM, Huerta-Sil G, Pérez-Hernández N, Londoño J, et al. Tumor Necrosis Factor-α Promoter Polymorphisms in Mexican Patients With Spondyloarthritis. Hum Immunol (2006) 67:826–32. doi: 10.1016/j.humimm.2006.07.009
9. Romero-Sánchez C, Londoño J, Delgado G, Jaimes DA, De Avila J, Mora A, et al. Association of Tumor Necrosis Factor Alpha-308 Promoter Polymorphism With Spondyloarthritides Patients in Colombia. Rheumatol Int (2012) 32:2195–7. doi: 10.1007/s00296-011-1883-1
10. Yang B, Xu Y, Liu X, Huang Z, Wang L. Il-23R and IL-17A Polymorphisms Correlate With Susceptibility of Ankylosing Spondylitis in a Southwest Chinese Population. Oncotarget (2017) 8:70310. doi: 10.18632/oncotarget.20319
11. Shen H, Goodall JC, Hill Gaston JS. Frequency and Phenotype of Peripheral Blood Th17 Cells in Ankylosing Spondylitis and Rheumatoid Arthritis. Arthritis Rheumatism: Off J Am Coll Rheumatol (2009) 60:1647–56. doi: 10.1002/art.24568
12. Chou C-T, Huo A-P, Chang H-N, Tsai C-Y, Chen W-S, Wang H-P. Cytokine Production From Peripheral Blood Mononuclear Cells in Patients With Ankylosing Spondylitis and Their First-Degree Relatives. Arch Med Res (2007) 38:190–5. doi: 10.1016/j.arcmed.2006.09.008
13. Van Hauwermeiren F, Vandenbroucke RE, Libert C. Treatment of TNF Mediated Diseases by Selective Inhibition of Soluble TNF or TNFR1. Cytokine Growth factor Rev (2011) 22:311–9. doi: 10.1016/j.cytogfr.2011.09.004
14. Kawaguchi M, Adachi M, Oda N, Kokubu F, Huang S-K. IL-17 Cytokine Family. J Allergy Clin Immunol (2004) 114:1265–73. doi: 10.1016/j.jaci.2004.10.019
15. Koenders MI, Marijnissen RJ, Devesa I, Lubberts E, Joosten LAB, Roth J, et al. Tumor Necrosis Factor–Interleukin-17 Interplay Induces S100A8, Interleukin-1β, and Matrix Metalloproteinases, and Drives Irreversible Cartilage Destruction in Murine Arthritis: Rationale for Combination Treatment During Arthritis. Arthritis Rheumatism (2011) 63:2329–39. doi: 10.1002/art.30418
16. Rudwaleit M, van der Heijde D, Landewé R, Listing J, Akkoc N, Brandt J, et al. The Development of Assessment of SpondyloArthritis International Society Classification Criteria for Axial Spondyloarthritis (Part II): Validation and Final Selection. Ann Rheumatic Diseases (2009) 68:777–83. doi: 10.1136/ard.2009.108233corr1
17. Rudwaleit MV, van der Heijde D, Landewé R, Akkoc N, Brandt J, Chou CT, et al. The Assessment of SpondyloArthritis International Society Classification Criteria for Peripheral Spondyloarthritis and for Spondyloarthritis in General. Ann Rheumatic Diseases (2011) 70:25–31. doi: 10.1136/ard.2010.133645
18. Abdolrahim-Zadeh H, Hakkakian N, Asadollahi R, Gharesifard B, Sarvari J, Kamali-Sarvestani E, et al. Interleukin-10 Promoter Polymorphisms and Breast Cancer Risk in Iranian Women. Iranian J Immunol (2005) 2:158–65.
19. Silvestre APA, Zacarias JMV, Guelsin GAS, Visentainer JEL, Sell AM. Genetic Polymorphisms of Human Platelet Antigens in Euro-African and Japanese Descendants From Parana, Southern Brazil. Platelets (2017) 28:607–10. doi: 10.1080/09537104.2016.1257785
20. Solé X, Guinó E, Valls J, Iniesta R, Moreno V. Snpstats: A Web Tool for the Analysis of Association Studies. Bioinformatics (2006) 22:1928–9. doi: 10.1093/bioinformatics/btl268
21. Dean AG, Sullivan KM, Soe MM. Openepi: Open Source Epidemiologic Statistics for Public Health, Version. (2014). Available at: www.OpenEpi.com, updated 2013/04/06. (Accessed November 22, 2020).
22. Lara-Armi FF, Visentainer JEL, Alves HV, Rocha-Loures MA, Neves JSF, Colli CM, et al. Optimization of HLA-B* 27 ALLELE Genotyping by PCR-SSP. Clinics (2020) 75:e1840. doi: 10.6061/clinics/2020/e1840
23. Lv C, Wang Y, Wang J, Zhang H, Xu H, Zhang D. Association of Interleukin-10 Gene Polymorphisms With Ankylosing Spondylitis. Clin Invest Med (2011) 34:370–6. doi: 10.25011/cim.v34i6.15898
24. Probst CM, Bompeixe EP, Pereira NF, De O Dalalio MM, Visentainer JEL, Tsuneto LT, et al. HLA Polymorphism and Evaluation of European, African, and Amerindian Contribution to the White and Mulatto Populations From Paraná, Brazil. Hum Biol (2000) 72:597.
25. Reis PG, Ambrosio-Albuquerque EP, Fabreti-Oliveira RA, Moliterno RA, de Souza VH, Sell AM, et al. Hla-A,-B,-DRB1,-DQA1, and-DQB1 Profile in a Population From Southern Brazil. Hla (2018) 92:298–303. doi: 10.1111/tan.13368
26. Keller C, Webb A, Davis J. Cytokines in the Seronegative Spondyloarthropathies and Their Modification by TNF Blockade: A Brief Report and Literature Review. Ann Rheumatic Diseases (2003) 62:1128–32. doi: 10.1136/ard.2003.011023
27. Baeten D, Van Damme N, Van den Bosch F, Kruithof E, De Vos M, Mielants H, et al. Impaired Th1 Cytokine Production in Spondyloarthropathy Is Restored by Anti-TNFα. Ann Rheumatic Diseases (2001) 60:750–5. doi: 10.1136/ard.60.8.750
28. Baeten D, Kruithof E, De Rycke L, Boots AM, Mielants H, Veys EM, et al. Infiltration of the Synovial Membrane With Macrophage Subsets and Polymorphonuclear Cells Reflects Global Disease Activity in Spondyloarthropathy. Arthritis Res Ther (2005) 7:R359. doi: 10.1186/ar1501
29. Ciccia F, Alessandro R, Rizzo A, Accardo-Palumbo A, Raimondo S, Raiata F, et al. Macrophage Phenotype in the Subclinical Gut Inflammation of Patients With Ankylosing Spondylitis. Rheumatology (2013) 53:104–13. doi: 10.1093/rheumatology/ket323
30. Deng B, Wehling-Henricks M, Villalta SA, Wang Y, Tidball JG. Il-10 Triggers Changes in Macrophage Phenotype That Promote Muscle Growth and Regeneration. J Immunol (2012) 189:3669–80. doi: 10.4049/jimmunol.1103180
31. Chu CQ, Swart D, Alcorn D, Tocker J, Elkon KB. Interferon-γ Regulates Susceptibility to Collagen-Induced Arthritis Through Suppression of Interleukin-17. Arthritis Rheumatism (2007) 56:1145–51. doi: 10.1002/art.22453
32. Mei Y, Pan F, Gao J, Ge R, Duan Z, Zeng Z, et al. Increased Serum Il-17 and IL-23 in the Patient With Ankylosing Spondylitis. Clin Rheumatol (2011) 30:269–73. doi: 10.1007/s10067-010-1647-4
33. Wang J, Zhao Q, Wang G, Yang C, Xu Y, Li Y, et al. Circulating Levels of Th1 and Th2 Chemokines in Patients With Ankylosing Spondylitis. Cytokine (2016) 81:10–4. doi: 10.1016/j.cyto.2016.01.012
34. Pan J, Hu J, Qi X, Xu L. Association Study of a Functional Variant of TNF-α Gene and Serum TNF-α Level With the Susceptibility of Congenital Heart Disease in a Chinese Population. Postgraduate Med J (2019) 95:547–51. doi: 10.1136/postgradmedj-2019-136621
35. Kroeger KM, Steer JH, Joyce DA, Abraham LJ. Effects of Stimulus and Cell Type on the Expression of the– 308 Tumour Necrosis Factor Promoter Polymorphism. Cytokine (2000) 12:110–9. doi: 10.1006/cyto.1999.0529
36. Miteva LD, Stanilov NS, Deliysky TS, Stanilova SA. Significance of -1082A/G Polymorphism of IL10 Gene for Progression of Colorectal Cancer and IL-10 Expression. Tumour Biol (2014) 35:12655–64. doi: 10.1007/s13277-014-2589-2
37. Fathy MM, Elsaadany HF, Ali YF, Farghaly MAA, Hamed ME, Ibrahim HE, et al. Association of IL-10 Gene Polymorphisms and Susceptibility to Juvenile Idiopathic Arthritis in Egyptian Children and Adolescents: A Case-Control Study. Ital J pediatrics (2017) 43:9. doi: 10.1186/s13052-017-0328-1
38. Azab SF, Abdalhady MA, Elsaadany HF, Elkomi MA, Elhindawy EM, Sarhan DT, et al. Interleukin-10-1082 G/A Gene Polymorphisms in Egyptian Children With CAP: A Case–Control Study. Medicine (2016) 95:e4013. doi: 10.1097/MD.0000000000004013
39. Ghaznavi H, Soltanpour MS. Association Study Between Rs2275913 Genetic Polymorphism and Serum Levels of IL-17A With Risk of Coronary Artery Disease. Mol Biol Res Commun (2020) 9:35–40. doi: 10.22099/mbrc.2020.35442.1463
40. Pasha HF, Tantawy EA, Youssef MA. Osteopontin and Interleukin-17A Genes Polymorphisms in Egyptian Systemic Lupus Erythematosus Patients: A Relation to Disease Activity and Severity. Gene (2019) 702:107–13. doi: 10.1016/j.gene.2019.02.100
41. Ocejo-Vinyals JG, de Mateo EP, Hoz MÁ, Arroyo JL, Agüero R, Ausín F, et al. The IL-17 G-152a Single Nucleotide Polymorphism Is Associated With Pulmonary Tuberculosis in Northern Spain. Cytokine (2013) 64:58–61. doi: 10.1016/j.cyto.2013.05.022
42. Huang H-T, Lu Y-L, Wang R, Qin H-M, Wang C-F, Wang J-L, et al. The Association of IL-17A Polymorphisms With Il-17a Serum Levels and Risk of Ischemic Stroke. Oncotarget (2017) 8:103499. doi: 10.18632/oncotarget.21498
Keywords: autoimmune disease, spondyloarthritis, rheumatological diseases, interleukin-10, interleukin-17, tumour necrosis factor alpha, single nucleotide polymorphisms, genetic polymorphism
Citation: Braga M, Lara-Armi FF, Neves JSF, Rocha-Loures MA, Terron-Monich MS, Bahls-Pinto LD, de Lima Neto QA, Zacarias JMV, Sell AM and Visentainer JEL (2021) Influence of IL10 (rs1800896) Polymorphism and TNF-α, IL-10, IL-17A, and IL-17F Serum Levels in Ankylosing Spondylitis. Front. Immunol. 12:653611. doi: 10.3389/fimmu.2021.653611
Received: 14 January 2021; Accepted: 09 June 2021;
Published: 05 July 2021.
Edited by:
Katarzyna Bogunia-Kubik, Hirszfeld Institute of Immunology and Experimental Therapy (PAN), PolandReviewed by:
Agnieszka Paradowska-Gorycka, National Institute of Geriatrics, Rheumatology, and Rehabilitation, PolandRoberto Padoan, University of Padua, Italy
Copyright © 2021 Braga, Lara-Armi, Neves, Rocha-Loures, Terron-Monich, Bahls-Pinto, de Lima Neto, Zacarias, Sell and Visentainer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jeane Eliete Laguila Visentainer, amVsdmlzZW50YWluZXJAdWVtLmJy