 Gianmarco Bellucci1
Gianmarco Bellucci1 Virginia Rinaldi1
Virginia Rinaldi1 Maria Chiara Buscarinu1,2
Maria Chiara Buscarinu1,2 Roberta Reniè1
Roberta Reniè1 Rachele Bigi1Giulia Pellicciari1Emanuele Morena1Carmela Romano1Antonio Marrone1
Rachele Bigi1Giulia Pellicciari1Emanuele Morena1Carmela Romano1Antonio Marrone1 Rosella Mechelli3,4
Rosella Mechelli3,4 Marco Salvetti1,5*
Marco Salvetti1,5* Giovanni Ristori1,2
Giovanni Ristori1,2- 1Centre for Experimental Neurological Therapies (CENTERS), Department of Neurosciences, Mental Health and Sensory Organs, Sapienza University of Rome, Rome, Italy
- 2Neuroimmunology Unit, Istituto di Ricovero e Cura a Carattere Scientifico (IRCCS) Fondazione Santa Lucia, Rome, Italy
- 3Istituto di Ricovero e Cura a Carattere Scientifico (IRCCS) San Raffaele Pisana, Rome, Italy
- 4San Raffaele Roma Open University, Rome, Italy
- 5Istituto di Ricovero e Cura a Carattere Scientifico (IRCCS) Istituto Neurologico Mediterraneo Neuromed, Pozzilli, Italy
Current knowledge on Multiple Sclerosis (MS) etiopathogenesis encompasses complex interactions between the host’s genetic background and several environmental factors that result in dysimmunity against the central nervous system. An old-aged association exists between MS and viral infections, capable of triggering and sustaining neuroinflammation through direct and indirect mechanisms. The novel Coronavirus, SARS-CoV-2, has a remarkable, and still not fully understood, impact on the immune system: the occurrence and severity of both acute COVID-19 and post-infectious chronic illness (long COVID-19) largely depends on the host’s response to the infection, that echoes several aspects of MS pathobiology. Furthermore, other MS-associated viruses, such as the Epstein-Barr Virus (EBV) and Human Endogenous Retroviruses (HERVs), may enhance a mechanistic interplay with the novel Coronavirus, with the potential to interfere in MS natural history. Studies on COVID-19 in people with MS have helped clinicians in adjusting therapeutic strategies during the pandemic; similar efforts are being made for SARS-CoV-2 vaccination campaigns. In this Review, we look over 18 months of SARS-CoV-2 pandemic from the perspective of MS: we dissect neuroinflammatory and demyelinating mechanisms associated with COVID-19, summarize pathophysiological crossroads between MS and SARS-CoV-2 infection, and discuss present evidence on COVID-19 and its vaccination in people with MS.
Introduction
Multiple Sclerosis (MS) is a chronic, immune-mediated disease of the Central Nervous System (CNS) characterized by focal demyelination and neurodegeneration. It is the leading cause of non-traumatic disability in young people, as its onset typically occurs between 20 and 40 years of age. Although the cause(s) of MS remain unknown, a multifactorial model is widely accepted: diverse “environmental” factors may trigger the immune attack and disease progression in genetically susceptible individuals (1–3).
Solid preclinical and clinical data support the role of viral infections in MS etiology. It is plausible that MS neurodegenerative and inflammatory processes start years before the epiphany of radiological and clinical manifestations (4–7). In this context, viral agents are good candidates to uncover the disease, as they are capable to precipitate the dysimmune process via multiple mechanisms: molecular mimicry, bystander activation, epitope spreading, autoreactive immune cell survival and immortalization, regulome modifications (3, 8–10). Thus, the selection and global spread of a novel human-infecting virus represents an experimental challenge to study virus-associated diseases, including MS. The research rush enlightened by SARS-CoV-2 diffusion may help to elucidate the “known unknowns” of virus-host interactions in immune-mediated diseases and accelerate drug tailoring. Furthermore, the novel coronavirus itself—with its pronounced effect on the host’s immune system—may be directly involved in development, or divert the natural history, of dysimmune diseases, depicting a new epidemiological map in the forthcoming years. In this review we will synopsize present knowledge on SARS-COV-2 and neuroinflammation, focusing on the crossroads of SARS-CoV-2 infection and MS physiopathology; we will also reassess current evidence on COVID-19 and SARS-CoV-2 vaccines in people with MS and discuss a potential inference in MS disease course.
SARS-CoV-2 and Neuroinflammation
Human-infecting Coronaviruses include four mild pathogenic strains (229E, NL63, OC43, and HKU1), which cause up to a third of common cold cases, and three highly virulent agents: SARS-CoV-1, MERS, and SARS-CoV-2. The latter pathogen has emerged in China in late 2019 and has quickly become pandemic, due to its extreme infectivity (11). In the acute phase, most of infected patients have no or mild symptoms, but up to 15% cases require hospitalization for a severe interstitial pneumonia, which has caused, to date, more than 4 million deaths.
COVID-19 is a multifaceted and unpredictable syndrome whose outcomes are primarily determined by the host’s immune response. Compared to other respiratory viruses, SARS-CoV-2 elicits a stronger, perduring, auto-aggressive inflammatory response (12), fueled by a massive cytokine release, causing coagulation dysfunction and multiorgan failure (13) in severe cases. The aberrant immune activation is clear-cut in children, who can suffer of a virus-induced multisystem inflammatory syndrome (MIS-C) with prominent autoimmune components (14, 15).
A SARS-CoV-2 neuroinvasive potential has been reported (16) as for other Coronaviruses (17), that have also been isolated in brains and cerebrospinal fluid (CSF) of MS patients with significant frequency (18, 19). Blood-brain barrier pericytes and astrocytes may represent SARS-CoV-2 entry points (20). Neurological symptoms are reported in more than 80% cases during disease history and, opposite to the respiratory distress syndrome, are more frequent in young people (21). Ischemic stroke may be a SARS-CoV-2 specific presentation, especially in people under 50 years of age (22, 23) as a consequence of the hypercoagulation state induced by systemic inflammation. Besides non-specific symptoms (headache, confusional states, dizziness, etc.) that are common complications of viral infections and critical illnesses (24), most of neuro-COVID manifestations share an immune-mediate substrate. In fact, brain lesions associated to COVID-19 reflect both vascular and demyelinating etiologies and are mainly imputable to the intense immune activation with massive neurotoxic cytokines production (in particular, IL1-beta and IL6) (25), rather than a direct infectious cytopathic effect. A prominent example comes from a neuropathological description of vascular and acute disseminated encephalomyelitis (ADEM)-like demyelinating pathology in a COVID-19 patient (26).
Omics studies on brain samples from COVID-19 patients are pinpointing the neuroinflammatory mechanisms underlying neurological involvement. Yang et al. (27) performed single-nucleus transcriptomics and immunohistochemistry of cortex and choroid plexus of eight COVID-19 brains. First, they highlighted that, albeit all major brain cell types resulted affected, glial cells displayed a pro-inflammatory, disease-specific signature. Second, they underscored significant brain barrier inflammation in the choroid plexus, a strategic interface between peripheral blood and cerebrospinal fluid. In COVID-19 brains, choroid barrier cells and glia limitans released chemokines toward brain parenchyma and promoted complement activation, fueling neuroinflammation and neural damages. A similar mechanism was described in MS, where an activated choroid epithelium in response to peripheral inflammation acts as a gateway for brain-homing, pathogenic B and T lymphocytes (28–30). Likely, infiltrating T cells were found in COVID-19 brain parenchyma, in the absence of SARS-CoV-2 RNA transcripts, suggesting an aberrant CNS attack by peripheral immune effectors favored by BBB disruption.
Comparison of transcriptome signatures between COVID-19 brains and chronic CNS diseases (including MS) revealed a significant intersection of dysregulated genes, particularly in glial cells. A notable example comes from RIPK1, whose overactivation in microglia and astrocytes contributes to MS pathology, particularly in the progressive disease (31). Moreover, COVID-19 differentially expressed genes are enriched with GWAS-associated variants of complex neurological traits, suggesting a possible interaction of the novel Coronavirus to the initiation or perpetuation of CNS disorders in individuals who are at risk (27).
Schwabenland et al. (32) found extensive T CD8 cell infiltration, microgliosis, and increased axonal damage in COVID-19 comparable to that seen in long-term MS patients. A disease-specific cluster of CD8+ lymphocytes was present in the perivascular space, causing vascular immunopathology and BBB disruption. Indeed, endothelial cells with viral proteins in the cytoplasm colocalize with innate and adaptive immune effectors, vascular pathology, and axonal damage. The crosstalk between cytotoxic lymphocytes and microglial cells peaks in the formation of microanatomical niches, so-called “microglial nodules,” endowed with a pervasive pro-inflammatory effect that extends throughout the brain.
To decipher the underpinnings of neurological symptoms, Song et al. (33) performed single-cell transcriptomics on immune cells from the CSF and blood of neuro-COVID-19 patients and healthy donors. They found and increased activation of CSF T cells, that strongly interacted with overactive NK and dendritic cells, displaying upregulation of IL-1 and IL-12 pathways and suggesting a coordinated and compartmentalized T-cell based response to CNS antigens. Also, B cells were enriched in COVID-19 CSF compared with controls, and their antibody profile differs from peripheral blood. A subset of intrathecal antibodies—especially anti-Spike—also targets neural antigens. A similar approach reported autoantibodies targeting known antigens (Yo, NMDA receptors) and undetermined epitopes of myelin, endothelium, astrocytes, and neurons, some of which help explain clinical features (e.g., seizures) (34).
SARS-CoV-2 is associated to a growing number of para-infectious and postinfectious neurological conditions that have an immunological substrate (35–38): in particular, Acute Disseminated Encephalomyelitis (ADEM) results extremely frequent (39). Para-infectious ADEM occurrence often correlates with COVID-19 severity, suggesting off-target effects of the hyperinflammatory response involving brain tissue and vasculature (40). Indeed, beyond deep white matter demyelination, these cases often display intraventricular and intraparenchymal hemorrhages (e.g., microbleeds), with features of fulminant necrotizing encephalitis (41–44). Conversely, post-COVID-19 ADEMs appear unrelated to respiratory involvement and do not differ from those triggered by other infections, also in terms of prognosis (45, 46). Interestingly, COVID-19 was shown to trigger an exacerbation of a recent, but remitted, Coxsackie virus–induced ADEM (47). It was also suggested that SARS-CoV-2-associated ADEMs have an atypically frequent involvement of spine—especially spinal gray matter (48).
Literature reviews identified 43 cases of Acute Transverse Myelitis (49) reported until January 2021 in association with COVID-19. As the first reports of neurological complications of SARS-CoV-2 infections did not involve myelitis, such a number of cases (with an estimated incidence of 0.5 per million) resulted somehow unexpected. Of interest, no association has been made between ATM and the other highly virulent beta-coronaviruses, suggesting a special property of SARS-CoV-2 in triggering spine demyelination. Patients’ age ranged from 21 to 73 years, male sex slightly prevailed, and COVID-19 course was prevalently mild. A third of cases developed neurological impairment within 5 days from the onset of COVID-19 symptoms, compatible with para-infectious physiopathology; however, the greatest number of cases represent postinfectious dysimmune complications, with a maximum latency of 6 weeks. Twenty percent of patients had myelitis in the context of an ADEM, which instead involved mostly women. Longitudinally extensive lesions (as in Longitudinally Extended Transverse Myelitis, LETM) involving >3 spinal cord segment appeared typical, occurring in 70% of COVID-19-associated myelitis cases; in a 28-year old woman, demyelination involved the entire spine, from the medulla oblungata to the conus medullaris (50).
More recently, additional and, to some extent, peculiar CNS demyelinating events linked to COVID-19 have been reported. Zoghi et al. (51) described a case of a 21-year-old male that developed encephalomyelitis 2 weeks after mild COVID-19. At admission, he manifested a confusional state with recurrent vomit, paraparesis, and a T8 sensory level. MRI revealed scarcely enhancing FLAIR hyperintensities extending from internal capsules to the cerebral peduncles and pons, marbled-pattern hyperintensities in corpus callosum, and a longitudinally extensive transverse myelitis in the cervical and thoracic spine. CSF analysis showed pleocytosis, hypoglycorrhachia, and hyperprotidorrhachia, without oligoclonal bands (OCBs). Image findings and clinical onset (alike area-postrema syndrome) pointed toward a possible NMOSD, but Aquaporin-4 receptor (AQP4) and myelin oligodendrocyte glycoprotein (MOG) antibodies in the serum and CSF were absent. Treatment with plasma exchange only provided partial recovery. The case was framed as an atypical ADEM attributable to a postinfectious dysimmune state. A more stereotypical acute anti-MOG CNS syndrome following SARS-CoV-2 infection had been previously described by Zhou et al. (52) characterized by bilateral, sequential, severe optic neuritis with contemporary involvement of the cervical and thoracic spine. The finding of anti-AQP4 antibodies in CSF and serum in a case of post-COVID-19 encephalomyeloradiculitis also suggests a nexus between SARS-CoV-2 and neuromyelitis optica spectrum disorder (NMOSDs) pathogenesis (53, 54)
Rodriguez de Antonio et al. (55) reported the case of a 40-year-old woman with asymptomatic SARS-CoV-2 infection that developed, in the late phase of a totally asymptomatic SARS-CoV-2 infection, hypoesthesia and bladder dysfunction. MRI revealed a T5-6 lesion; serological essays found the presence of IgM anti-gangliosides (GD2 and GD3), more commonly seen in peripheral nervous system inflammatory disorder (such as Guillain Barrè Syndrome, GBS). Indeed, IgM anti-GD1b were detected in a post-Covid Acute Motor Axonal Neuropathy (AMAN) variant of GBS (56). However, whether SARS-CoV-2-induced demyelination is endowed with distinct features and biomarkers should be clarified.
COVID-19 vs MS: Immunopathogenetic Crossroads
The above-mentioned immunological pathways dysregulated in COVID-19 largely overlap with MS pathogenetic mechanisms and suggest that SARS-CoV-2 infection could act as an environmental risk factor for disease manifestation in susceptible individuals. This hypothesis is reinforced by clinical and experimental studies that unravel host-pathogen interactions underlying COVID-19 at both virological and genetic levels, intersecting those dealing with the major components of MS etiopathogenesis.
Based on the principle that genetic data inform on multifactorial disease pathogenesis, a system biology study found significant interactions of SARS-CoV-2 with genes associated with autoimmune diseases and comorbidities that predispose to a severe COVID-19 course. The strong association with autoimmunity was peculiar of SARS-CoV-2 with respect to other Coronaviruses and respiratory viruses, in line with the exuberant and autoaggressive immune response that causes COVID-19. Interestingly, MS-associated genes were the most enriched in SARS-CoV-2 host’s interactors, suggesting pathophysiological overlaps that are worth investigating (57). In particular, there are three pivotal crossroads of MS and COVID-19 immunological substrates: the type-1 IFN (IFN-I) response, the TH-17 axis, and the inflammasome pathway.
In MS patients, an impairment in IFN-I response has been widely described at the genetic and transcriptional levels and is confirmed by three decades of successful treatment with IFN-1B formulations (58, 59). Studies on severe COVID-19 patients highlighted a defective IFN-I production, with a consequent delayed and unrestrained T cell response, enhancing both viral spread and cytokine release that predispose to multiorgan failure (60, 61). Notably, the use of beta-interferons decreases the risk of COVID-19 in people with MS (pwMS) (62). Coronaviruses can interfere with IFN-I production and actions through multiple non-structural proteins. It has been demonstrated that SARS-CoV-2 antagonizes IFN-I secretion and signaling more efficiently than SARS-CoV-1 and MERS-CoV (63). These intrinsic viral properties are detrimentally boosted if the host itself displays a blunted IFN system: in severely ill COVID-19 patients, Zhang et al. found an enrichment in genetic variants accounting for an impairment of IFN-I immunity (64), while Bastard et al. reported augmented levels of IFN-I neutralizing antibodies (65). These data may suggest that people at risk for developing MS (just as pwMS not receiving DMTs) may be more susceptible to the virus-induced dysimmune effects; likewise, the repurposing of IFN-beta from MS armamentarium for the fight against COVID-19 has a biological rationale if administered early after the infection, as an exogenous “supplement” to aid viral suppression (35, 66).
Th1 cells producing IFN-gamma and Th17 cells secreting IL-17 are the principal T lymphocyte subsets involved in MS pathogenesis, to the point that most immunomodulating therapies aim to restore immunological balance interfering with their induction and skewing towards the Th2 axis (2). Interest in Th17 cells in MS is growing, since their discovery in 2005. They are induced by IL-6 produced by pathogenic B cells, contribute pleiotropically to CNS inflammation, and are the principal player of the brain autoimmune attack induced by intestinal microbiota alteration in pwMS (36–38). Severe COVID-19 cases exhibit high levels of IFN-γ and IL6 and a skewing towards Th17 cells (67, 68); moreover, IL-17 levels correlate with clinical severity, suggesting the opportunity of therapeutically inhibiting the Th17 axis (69). A sophisticated virus-host interaction analysis discovered that IL-17 receptor A (IL17RA) physically interacts with SARS-CoV-2 Orf8 protein, and highlighted a genetic locus associated with COVID-19 severity whose biological effects relate to IL-17 actions (70). Also, SARS-CoV-2 is capable of infecting enteric cells (71), and microbiota alterations correlate with clinical outcomes (72), so that the contribution of a gut-lung axis including Th17 cells in COVID-19 has been postulated (73). These elements make likely the possibility that Th17 axis could be involved in SARS-CoV-2 effects in pwMS.
Inflammasomes are cytosolic multimeric complexes driving the inflammatory response of the innate immune system. They activate Il-1Beta and IL-18 upon sensing of endogenous (tissue damage) or exogenous (e.g., infections) stressors by pattern-recognition receptors (PRRs), such as NLRP3. In MS, NLRP3-inflammasome significantly contributes to the chronic inflammation led by microglial cells that propels neurodegeneration. IL1B gene is upregulated in blood and brain lesions of primary progressive MS patients (74) due to an NLRP3 overactivity, on which IFN-I may interfere (75). Moreover, gain-of-function variants in NLRP3 and IL1B genes correlate with severity and progression of MS (76). Since the SARS-CoV-1 E, ORF3a, and ORF8b proteins directly activate NLRP3-inflammasome, similar effects are hypothesized in SARS-CoV-2. An increased potency of ORF3a-NLP3 interaction may underly the augmented virulence of the novel coronavirus (77–79). Being the activation of “danger sensors” among the early activators of an immunopathogenetic process, it is plausible that the immune response following SARS-CoV-2 infection could contribute to neuroinflammation in genetically susceptible people, precipitating MS onset or even impacting on progression (80).
SARS-CoV-2 and MS-Associated Viruses: Partners in Crime?
Indirect mechanisms through which SARS-CoV-2 infection may interfere with MS course may be sustained by the interactions with long-studied MS-associated pathogens: the Herpesviridae members Epstein Barr Virus and Human Herpes Virus 6 (HHV-6), as well as the Human Endogenous Retroviruses (HERVs).
A longstanding, quasi-causal evidence supports the Epstein-Barr Virus (EBV) as a necessary environmental interactor (81) involved across the whole spectrum of MS (82). High levels of antibodies against EBV antigens (EBNA1 and VCA) at least triplicate the risk of developing MS; a history of infectious mononucleosis (IM), the clinical syndrome caused by a post-childhood EBV infection, exerts a similar effect (83, 84). The prevalent hypothesis to explain these sero-epidemiological evidences is an augmented immune reactivity reflecting insufficient control of viral infection, due to EBV-specific T lymphocyte exhaustion or scarcity: the quasi-persistent active state in periphery and CNS promotes neuroinflammation and disease progression (85–90).
Lymphopenia and T cell exhaustion, which characterize COVID-19, may impair the control of EBV latency in B cells [also within meningeal follicles (86)], favoring viral reactivation, rising up anti-EBV titers and exacerbating the hyperinflammatory state. Indeed, severe COVID-19 cases display higher prevalence and levels of EBV viremia compared with non-COVID critically ill subjects; the virus’s DNA titers and anti-EBNA1 IgM levels correlate with increased inflammatory markers (C-reactive Protein, IL-6), disease severity, and prognosis (91–93). Paolucci and colleagues screened for opportunistic viral infections 104 moderate to severe COVID-19 cases: EBV was the only detected (95.2% of the ICU patients and in 83.6% Sub-ICU patients). Reactivation correlated with reduction of CD8+ T and NK lymphocyte count, supporting the hypothesis of a defective cell-mediated control (94).
A SARS-CoV-2 -EBV interaction may extend beyond the acute phase and play a role in the pathogenesis of “long-COVID,” or post-COVID-19 syndrome. It manifests with lingering symptoms and/or delayed or long-term complications beyond 4 weeks from the acute phase. Most patients suffer of fatigue or muscle weakness, sleep difficulties, and anxiety or depression, but organ-specific sequelae also play a part (94, 95). Neuropsychiatric involvement appears to be typical and frequent enough that a more specific qualifier of “Post-COVID-19 Neurologic Syndrome” (PCNS) has been coined (96, 97). Gold and colleagues found that two-thirds of COVID-19 long-haulers had EBV reactivation and speculated this might contribute to clinical manifestations.
An integrated, molecular portrait of post-COVID-19 syndrome pathogenesis is still lacking; however, in patients’ and clinicians’ descriptions of this heterogeneous condition, similitudes with Myalgic Encephalomyelitis/Chronic Fatigue syndrome (CFS) stand out (98). This syndrome comprehends extreme physical and cognitive impairment that are typically preceded by an acute infectious disease: EBV and HHV-6 A/B have been historically implicated (99–102). Mechanisms contributing to its pathogenesis include immune exhaustion and dysregulation subsequent to an exuberant acute response to the pathogen (increased cytokines, presence of autoantibodies), a multisystemic hypometabolic state with redox imbalance, intestinal dysbiosis, and diffuse neurological dysfunction (pain hypersensitivity, subtle CNS neuroinflammation with glial cell dysmetabolism, impaired brain connectivity) (103–106). Fatigue concerns more than 75% pwMS during disease course—including the prodromic phase (107)—and is frequently considered as the worse symptom (108, 109). Nonetheless, it is often overlooked by clinicians, also because no drug has proven to be effective in tackling it. A recent randomized clinical trial demonstrated no difference between placebo and three commonly prescribed symptomatic medications (amantadine, modafinil, methylphenidate) in easing MS-related fatigue (110). Beyond clinical similarities—also mirrored by neuroimaging functional studies (111)—there are significant molecular overlaps between CFS and MS, as objectively assessed by immunological studies (112–114). A notable example is the high frequency of circulating CD8+ mucosal-associated invariant T cells (MAIT) in both CFS and MS patients (115), where they bridge intestinal dysbiosis to CNS inflammation (116–118). Given the novelty and the expected high prevalence of post-COVID-19 syndrome, research on CFS and MS-associated fatigue pathomechanisms will hopefully gain momentum, paving the way toward targeted therapies for these conditions.
EBV reactivation could be as a simple byproduct of COVID-19 unbalanced immunity; however, data are emerging that show probable virus-virus synergies at the expense of the host. An in-silico analysis by Vavougios (119) dissected overlapping host gene signatures between SARS-CoV-2 and other co-pathogens. EBV-induced expression pattern was the most significantly enriched, suggesting a mechanistic interplay between the two viruses operating at the host’s transcription level. The interaction nodes shared by the two viruses include Heat-Shock Proteins (HSPA8, HSPA5, HSPD1, etc.), ribonucleoproteins, and post-translational regulators (SYNCRIP, SERBP1, SSBP1).
EBV may also enhance SARS-CoV-2 infectivity. The expression of ACE2, the principal SARS-CoV-2 surface receptor, is regulated by EBV’s ZTA transcription factor and peaks when EBV enters the lytic replicative cycle in epithelial cells. Its reactivation proved to facilitate ACE2-dependent coronavirus infection on pseudovirus assays (120). Neuropilin-1 (NRP1) has been recognized as a coreceptor of SARS-CoV-2, binding the Spike protein after its cleavage by proteases (121, 122). Interestingly, NRP1 also facilitates EBV cell entry, and its interaction with EBV-gB protein activates intracellular kinase pathways resultant in a positive feedback on viral infectivity (123). This transmembrane protein is expressed in various immune cells. Most of studies focused on Tregs, whose action ends in the modulation of Th1 and Th17 subsets (124, 125). Deletion of NRP1 exacerbates Experimental Autoimmune Encephalitis, the most used murine model of MS (124). Moreover, its expression is augmented in endothelial cells of active MS lesions (125) in response to the blood-brain barrier (BBB) disruption.
Alike to EBV, HHV6 is a neurotropic and lymphotropic pathogen that persists lifelong in the host as an episome or through chromosomal integration in glial cells. High titers of anti-HHV6 antibodies are epidemiologically associated to MS development risk, both alone and by interacting with anti-EBNA1 titers (126). Several HHV-6 pathogenetic mechanisms have been pinpointed in MS: enhancing on-site inflammation in MS lesion, mimicry toward myelin antigens, direct provocation of apoptosis of glial cells and consequent epitope spreading, and induction of neuronal death due to glutamate excitotoxicity (10).
Evidence on HHV-6 reactivation during SARS-CoV-2 infection is less solid, and the Herpesvirus is mainly associated to COVID-19 dermatological manifestations (127). Nonetheless, a neurological event associated to SARS-CoV-2/HHV6 interaction was described by Jumah et al. (128): a 61-year-old man experiencing lymphopenia and hypogammaglobulinemia during COVID-19 developed a para-infectious acute myelitis. CSF analysis showed a high HHV-6 viremia, and anti-MOG antibodies were detected in serum. In this case, the two viruses—individually linked to MOG-associated diseases (52, 129)—might have synergized in triggering immune-mediated demyelination. A direct interaction alike to that seen for HHV-6A with EBV in MS is therefore conceivable (130).
HERVs result from the integration of ancient exogenous retroviruses into human genome. Their transcription is finely regulated through epigenetic mechanisms and particularly affected by other viruses’ infection, including EBV and HHV-6. Among HERVs, HERV-W has been linked to MS pathogenesis thus being also named MS-Related Virus (MSRV). Notably, HERVs are also associated to CFS (131). When expressed, HERV-W Env protein acts as a superantigen, activates CNS-oriented T lymphocytes and microglia, sustaining a proinflammatory state and impairing myelin homeostasis (10). These mechanisms translate to MS relapses and progression, to the point that MSRV proteins have been candidate as biomarkers and therapeutic targets in MS (132, 133).
Charvet and colleagues demonstrated that SARS-CoV-2 activates HERV W and HERV-K Env transcription in vitro through the interaction of Spike with ACE2 on T lymphocytes (134). Accordingly, ENV is strongly expressed in blood cells of COVID-19 patients but not in healthy donors, and its level correlates with inflammatory mediators (IL-6, IL-17, and CXCR1), and with markers of T-cell exhaustion (e.g., PD1). Being predictive of a severe prognosis, ENV expression could play a role in immune overactivation and contribute to acute and chronic COVID-19 complications (135). In this context, HERVs could act as endogenous modulators of SARS-CoV-2/host interaction, helping to explain the extreme inter-individual variability in COVID-19 manifestations (136). Along this line, potential interplays of Herpesviruses and HERVs with SARS-CoV-2 might also be conceivably supposed to impact on MS pathogenesis and affect disease course.
COVID-19 and SARS-CoV-2 Vaccines in People With Multiple Sclerosis: What We Know
Relevant to our discussion is the possible role of SARS-CoV-2 infection in unveiling definite MS. To date, four cases have been described in literature, in which, during or post COVID-19, neurological syndromes in young people (three women, one man) appeared as typical onset of MS fulfilling McDonald’s 2017 criteria (Table 1). Such event cascade may be (a) purely casual, i.e., MS onset occurred by chance after SARS-CoV-2 infection; (b) non-specific, i.e., COVID-19 denotes the increase morbidity observed in people that will in a few years develop MS—so-called “MS prodrome” (141); (c) specific, i.e., SARS-CoV-2 infection acts as an environmental risk factor for MS. Of course, given the youth of the novel Coronavirus, evidence is insufficient to take conclusions, and large-scale, case-control studies are awaited to answer.
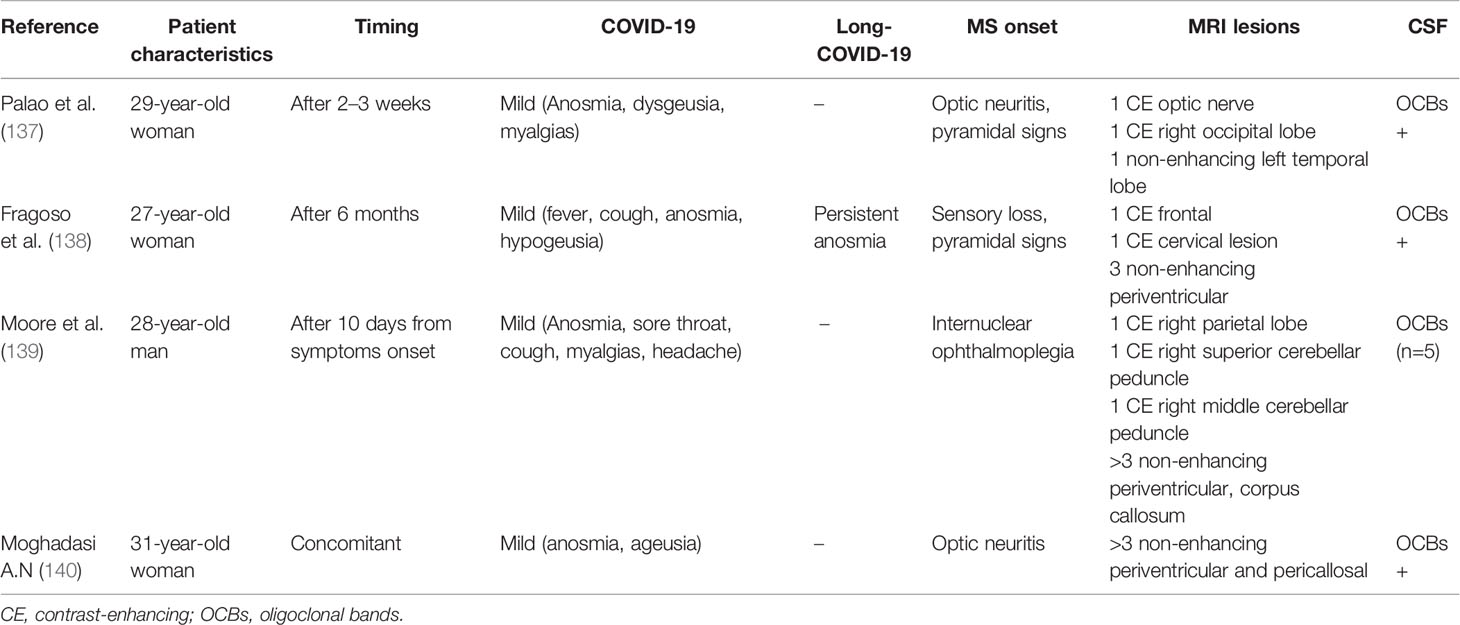
Table 1 Reported cases of MS onset in a temporal relationship with COVID-19 .
The immune activation consequent to a microbial infection often embraces some grade of non-specific systemic inflammation that is known to enhance autoimmunity in people with MS (pwMS) and interfere with the disease course (142–144). Upper respiratory tract infections (URTI), typically caused by adenoviruses, rhinoviruses, flu viruses, and coronaviruses, have been extensively proven to trigger both clinical and radiological MS relapses (145–147). Moreover, Edwards et al. demonstrated that a serological confirmation of URTI corresponded to a threefold augment in MS exacerbation risk (145). Precisely, McDonald’s criteria define relapse as “a monophasic clinical episode with patient-reported symptoms and objective findings typical of multiple sclerosis, reflecting a focal or multifocal inflammatory demyelinating event in the CNS, developing acutely or subacutely, with a duration of at least 24 hours, with or without recovery, and in the absence of fever or infection” (148). Conversely, the term pseudo-relapse refers to recrudescence of previously experienced MS symptoms concomitant to an elevation of body temperature, as during physical activity or infection-related fever. A pseudo-relapse lasts more than the brief, self-limiting exacerbation of a focal deficit typical of the Uhthoff’s phenomenon induced by heat: thus, it is easily distinguished from common day-to-day fluctuations of chronic symptoms, but the discrimination from relapses is more challenging (149).
Emerging data show that SARS-CoV-2 infection can trigger both pseudo-relapses and true relapses. A large community-based study by Garjani et al. (150) showed that the majority of pwMS developing COVID-19 reported clinical worsening of pre-existing neurological symptoms, and 20–35% developed new MS symptoms, mostly motor or sensory in nature, that could last for months following the acute infectious phase (thus being considerable as true relapses). These attacks were referred as worse than those experienced with non-COVID-19 infections. Accordingly, a smaller study from New York University MS Centre reported a 21.1% recrudescence rate (151). DMTs appear to be protective towards SARS-CoV-2-induced relapses. Indeed, pwMS receiving DMTs are less likely to develop new MS symptom during COVID-19 compared to untreated patients (152). On the other hand, a possible “protective” effect of COVID-19 on relapse rate was suggested by Etemadifar and colleagues (153), who conducted a retrospective study on 125 relapsing-remitting pwMS, 56 of whom had laboratory confirmed SARS-CoV-2 infection. Only 7.14% of pwMS in the COVID-19 group experienced a clinical relapse compared with 26.09% of the non-COVID-19 group. The authors hypothesized that COVID-19-associated lymphopenia could have a role in containing the proliferative rate of CNS-targeting T clones. However, no significant difference emerged in a self-comparison assessment of each group: relapse rate of each patient was similar for comparable windows taken before and during the pandemic (i.e., after SARS-CoV-2 infection, for the COVID-19 group). This suggests a possible selection bias: control group included (i) pwMS with an intrinsically higher relapse rate and/or (ii) pwMS receiving mostly first-line, less effective DMTs—which, on the other side, may have reduced their infection risk.
Indeed, since the beginning of the pandemic, research efforts have focused on answering urgent questions: (i) Are pwMS more susceptible to SARS-CoV-2 infection? (ii) What is the clinical course of COVID-19 in pwMS? (iii) What is the impact of DMTs on COVID-19 susceptibility and prognosis? After the first pioneering Italian observational study (62, 154), many others provided converging results that supported clinical practice and reassured both neurologists and patients (155). Besides some discordances among results (mostly imputable to heterogeneity in cohorts’ characteristics and data collection), preliminary findings have been corroborated, thanks to larger samples’ numerosity and pooled analyses (62), allowing to reasonably draw some conclusions. First, pwMS do not appear to have an increased risk of contracting COVID-19. Older age, male sex, and the presence of comorbidities negatively affect the infection’s clinical course in pwMS as in non-MS people. Except for fatigue, which is expectedly more frequent in pwMS, respiratory and systemic symptoms did not differ between pwMS and the general population. Neurological disability and progressive disease course emerged as linked to COVID-19 severity, consistent with a frail status of this subgroup of patients.
In general, DMTs showed an acceptable level of safety with respects to SARS-CoV-2 infection outcomes. Type I IFN formulations showed a protective effect (in line with the role of IFNs in restraining SARS-CoV-2 in the early infectious phase). Instead, a complicated course of COVID-19 occurred more frequently in the following circumstances: pwMS not receiving any DMT, cases under therapy with anti-CD20 agents (ocrelizumab, rituximab), or people who get infected soon after receiving high-dose corticosteroids (62, 155).
Present knowledge reflects the pandemic scenario during the first and second waves of SARS-CoV-2 spread. The emergence of variant of concern, potentially endowed with augmented infectivity and virulence (156), emphasizes the need of continuing clinical and epidemiological monitoring of pwMS. Variant selection and the overall pandemic trend largely depend on ongoing vaccination campaigns, which have started in late 2020, after the speed approval of mRNA-based (Pfizer and Moderna) and adenovirus-vectored formulations (AstraZeneca and Johnson & Johnson’s). Since then, focus has shifted to ascertain safety and efficacy of the anti-COVID-19 vaccines in pwMS.
Trials and real-life experiences with diverse “traditional” vaccine formulations (influenza, tetanus, meningococcus, etc.) showed that active immunization is safe in pwMS. While no specific issues exist for non-live formulations, administration of live-attenuated vaccines (such as VZV) should be programmed before starting DMTs and avoided during immunosuppression owing to the risk of a productive infection (152, 157). A satisfactory level of safety has been promptly confirmed for anti-COVID-19 mRNA-based vaccines: in a study by Achiron et al. (158) on 555 pwMS receiving Pfizer-BioNTech vaccine, frequency and characteristics of vaccine-induced adverse events were almost comparable with non-MS population and more numerous in young people; in a cohort of 239 pwMS surveyed by Lotan and colleagues (159), reactions were even less common. Detailed studies on mRNA-1273 and adenovirus-based vaccines’ safety in pwMS are lacking and awaited to consolidate clinical indications.
In general, most studies have ruled out an increased long-term risk of developing MS (160) as well as a short-term risk of neurological relapse in people with clinically defined MS (161) following vaccinations. However, some caution should be taken when vaccinating pwMS with sustained disease activity and/or a recent clinical or radiological relapse, in which the threshold for inflammatory exacerbation concomitant to the immunization stimulus could be lower (162–165). The same could be hypothesized for people with subclinical CNS inflammation (such as in radiological isolated syndromes, RIS) or genetically at-risk individuals during disease prodrome. Havla et al. (166) described the case of a 28-year-old woman with a familiar history of MS who developed a sensorimotor syndrome 6 days after the first administration of BNT162b2 SARS-CoV-2 vaccine. MRI revealed an active myelitis and non-enhancing, disseminated white matter lesions in the brain; OCBs were present in the CSF, and a diagnosis of MS was made. Early data on cohort BNT162b2 vaccination have not highlighted an increased risk of relapse in the very-short term (158); in another study on the same vaccine, 15.1% of pwMS reported worsening or new neurological symptoms that lasted more than 3 days in 55.6% of cases (159). However, further investigation on the long term is warranted to establish whether anti-COVID-19 vaccines’ safety profile is confirmed in MS.
Concerning vaccine efficacy, studies are ongoing that evaluate serological response as a surrogate of protection against COVID-19 in pwMS. It is known that some DMTs can negatively affect active immunization. Evidence recently reviewed in (167) indicates that fingolimod, natalizumab, alemtuzumab, and the anti-CD20 monoclonal antibodies (ocrelizumab, rituximab) may reduce serological response to “traditional” vaccines (inactivated, conjugate, toxoid). As presumable and in line with studies on rheumatic diseases, anti-CD20 appears to have the greatest impact on antibody production, which is partly related to the levels of circulating B cells at the time of administration (168). Before the pandemic, no studies had been conducted on vaccination efficacy in pwMS receiving the oral B-depleting agent cladribine. Converging results are coming from studies assessing humoral response following natural SARS-CoV-2 infection and vaccinations. Indeed, following COVID-19, lower anti-SARS-CoV-2 IgG titers were found in pwMS receiving fingolimod or anti-CD20, the being latter associated to a completely absent response (169–172). Similar results are coming from observational studies evaluating post-vaccination COVID-19 serology (displayed in Table 2). Fingolimod may attenuate (if not abate) humoral response, especially in pwMS with reduced lymphocyte count at the time of administration; such response is frequently absent in pwMS receiving ocrelizumab or rituximab, and when present, it depends on the time interval between the last drug infusion and vaccination. Other DMTs, including cladribine, did not appear to interfere with post-COVID-19 or post-vaccination serological response. Interestingly, the largest study to date evidenced a significantly higher humoral response to mRNA-1273 vaccine compared to BNT162b2, suggesting its preferential use in pwMS at risk of unsuccessful immunization (179).
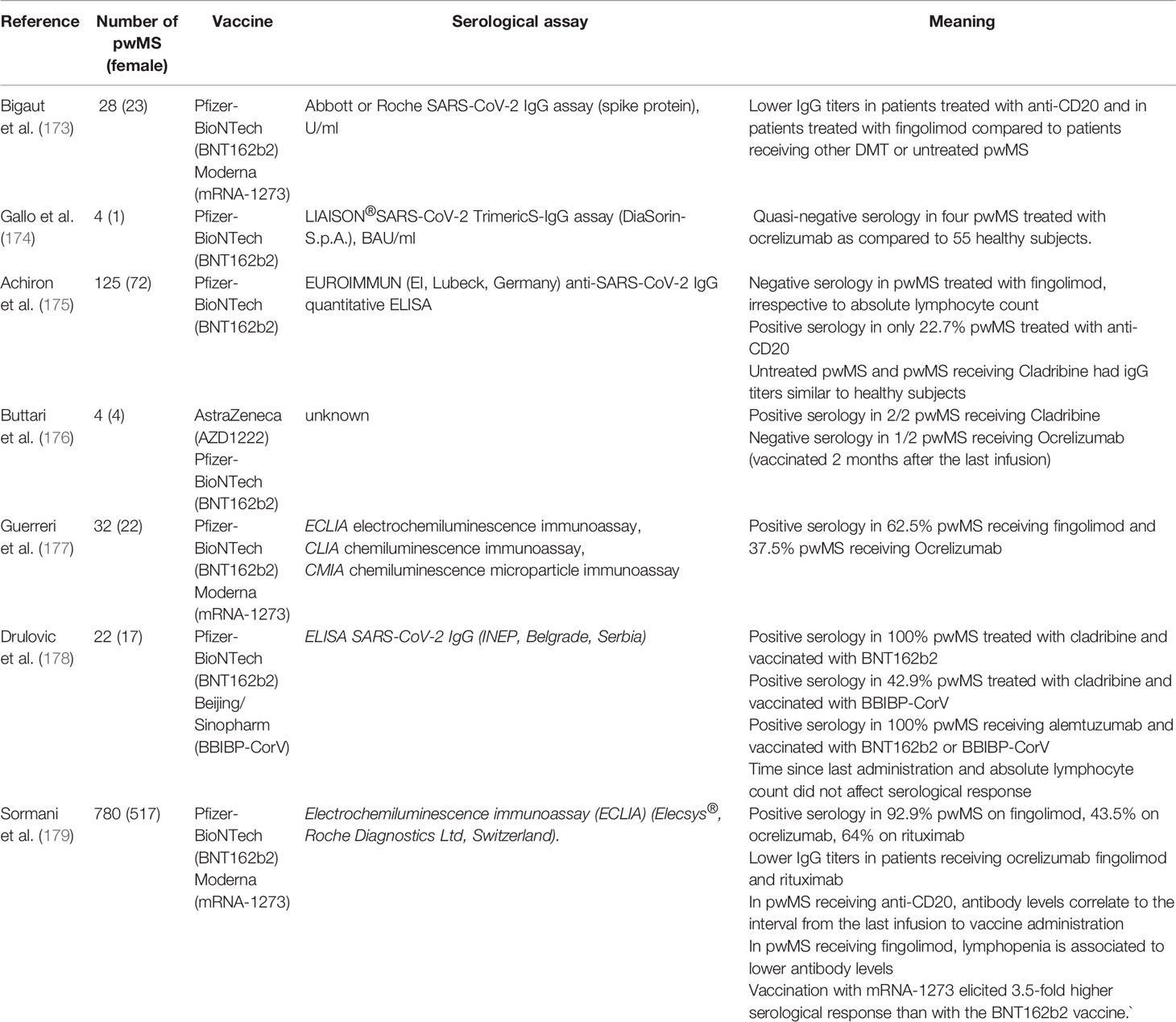
Table 2 Studies on COVID-19 vaccination in people with MS (until July 31st, 2021).
Beyond humoral response, a critical role in adaptive immunity to SARS-CoV-2 is exerted by cytotoxic and helper T cells. As quantitative correlates of serological protection are still unclear, assessing SARS-CoV-2-specific T-cell response in naturally infected or vaccinated subjects is of increasing importance, given that emerging viral variants evading humoral neutralization do not escape cellular responses (180, 181). They appear even more relevant when antibody production is substantially dampened, as in the case of B-cell-depleted pwMS. To this regard, preliminary evidence showed that the majority of pwMS receiving Ocrelizumab who experienced COVID-19 were able to mount an efficient and T-cell response toward SARS-CoV-2 (182). Apostolidis and colleagues (183) performed an extensive immunological study on 20 B-cell depleted pwMS (19 on Ocrelizumab and 1 on Rituximab) following mRNA SARS-CoV-2 vaccination. Besides confirming a significant impairment in antibody and memory B cell responses, they found a robust T cell response, with SARS-CoV-2 specific CD8+ expansion exceeding that of healthy controls, especially in pwMS completely lacking igG production. These findings are in line with some reported cases of favorable course of COVID-19 in pwMS receiving anti-CD20 (184, 185) and reassure about the efficiency of vaccination in immunocompromised patients. Similar immunological investigations are awaited in pwMS receiving other DMTs, in order to get an integrative view on vaccine-induced protection against COVID-19 to optimize immunization strategies.
To sum up, current evidence favors recommending administration of SARS-CoV-2 vaccines to pwMS, as the benefit of protection from COVID-19 largely outweighs the potential risks; timing with respect to DMTs and type of vaccine should be tailored case by case by neurologists, taking into account the clinical history and additional risk factors (186).
Conclusions
Almost 2 years of SARS-CoV-2 pandemic have profoundly impacted on medical world and scientific research, including the field of MS. We believe that the “research rush” triggered by the novel Coronavirus may propel studies on MS; vice versa, deepening our knowledge on MS pathogenesis and its relationship with viral infections could guide the investigations on COVID-19 immunopathogenesis.
Establishing such a virtuous cycle may be relevant to multiple aims. Technological advances rapidly mastered to deal with SARS-CoV-2 could be translated soon to address MS issues: a prominent example comes from the potential application of mRNA-based technology to design tolerizing vaccines that dampen CNS autoimmunity (187). Conversely, some MS DMTs could be repurposed to manage COVID-19-associated immune dysregulation. Clinical trials are already ongoing to evaluate IFN-B, fingolimod, dimethylfumarate; additionally, masitinib (a tyrosine-kinase inhibitor that showed efficacy in phase 2B/3 trials for progressive MS) has been recently highlighted as a potent Coronavirus inhibitor (188).
Furthermore, pinpointing the biological substrates of neuroinflammatory events associated with COVID-19 could help extend knowledge on MS-associated viruses in the etiopathogenesis of both conditions. The possibility that SARS-CoV-2-induced CNS autoimmune syndromes represent a distinct nosological entity should be evaluated through long-term studies (189); likewise, large case-control studies are needed to test a possible association between SARS-CoV-2 infection and MS development. A long follow-up of pwMS that suffered of COVID-19 will evaluate a potential inference in disease natural history, as well as the frequency and manifestation of long-COVID-19 in these patients. To this aim, it will be crucial to perform thorough immunological investigations on biological samples along with epidemiological studies evaluating past SARS-CoV-2 infection or vaccination as clinical variables.
In its transition to endemicity, COVID-19 is expected to become a mild disease of the childhood (190). Will a post-adolescence infection, repeated infection, or vaccine-induced immunity reshape the future epidemiology of MS? While dealing with the unprecedented scenario of a newborn virus pandemic, we might find hints to improve our understanding on this complex disorder.
Author Contributions
GB conceived the idea and wrote the manuscript. GB, VR, MB, RR, RB, GP, EM, CR, AM, and RM performed bibliographic search and contributed to data interpretation. GR and MS reviewed the manuscript. All authors contributed to the article and approved the submitted version.
Funding
MS and GR are supported by CENTERS, a Special Project of, and financed by, FISM—Fondazione Italiana Sclerosi Multipla.
Conflict of Interest
MS received research support and consulting fees from Biogen, Merck, Novartis, Roche, Sanofi, Teva.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Baecher-Allan C, Kaskow BJ, Weiner HL. Multiple Sclerosis: Mechanisms and Immunotherapy. Neuron (2018) 97:742–68. doi: 10.1016/j.neuron.2018.01.021
2. Dendrou CA, Fugger L, Friese MA. Immunopathology of Multiple Sclerosis. Nat Rev Immunol (2015) 15:545–58. doi: 10.1038/nri3871
3. Bordi I, Ricigliano VAG, Umeton R, Ristori G, Grassi F, Crisanti A, et al. Noise in Multiple Sclerosis: Unwanted and Necessary. Ann Clin Trans Neurol (2014) 1:502–11. doi: 10.1002/acn3.72
4. Marrie RA. Mounting Evidence for a Multiple Sclerosis Prodrome. Nat Rev Neurol (2019) 15:689–90. doi: 10.1038/s41582-019-0283-0
5. Giovannoni G. The Neurodegenerative Prodrome in Multiple Sclerosis. Lancet Neurol (2017) 16:413–4. doi: 10.1016/S1474-4422(17)30127-8
6. Wijnands JMA, Zhu F, Kingwell E, Zhao Y, Evans C, Fisk JD, et al. Prodrome in Relapsing-Remitting and Primary Progressive Multiple Sclerosis. Eur J Neurol (2019) 26:1032–6. doi: 10.1111/ene.13925
7. Bjornevik K, Munger KL, Cortese M, Barro C, Healy BC, Niebuhr DW, et al. Serum Neurofilament Light Chain Levels in Patients With Presymptomatic Multiple Sclerosis. JAMA Neurol (2020) 77:58–64. doi: 10.1001/jamaneurol.2019.3238
8. Kakalacheva K, Münz C, Lünemann JD. Viral Triggers of Multiple Sclerosis. Biochim Biophys Acta - Mol Basis Dis (2011) 1812:132–40. doi: 10.1016/j.bbadis.2010.06.012
9. Ricigliano VAG, Handel AE, Sandve GK, Annibali V, Ristori G, Mechelli R, et al. EBNA2 Binds to Genomic Intervals Associated With Multiple Sclerosis and Overlaps With Vitamin D Receptor Occupancy. PloS One (2015) 10:1–11. doi: 10.1371/journal.pone.0119605
10. Mechelli R, Romano C, Reniè R, Manfrè G, Buscarinu MC, Marrone A, et al. Viruses and Neuroinflammation in Multiple Sclerosis. Neuroimmunol Neuroinflamm (2021) 8:1–15. doi: 10.20517/2347-8659.2021.01
11. Hu B. Characteristics of SARS-CoV-2 and COVID-19. Nat Rev Microbiol (2019) 19:141–54. doi: 10.1038/s41579-020-00459-7
12. Subbarao K, Mahanty S. Respiratory Virus Infections: Understanding COVID-19. Immunity (2020) 52:905–9. doi: 10.1016/j.immuni.2020.05.004
13. Giamarellos-Bourboulis EJ, Netea MG, Rovina N, Akinosoglou K, Antoniadou A, Antonakos N, et al. Complex Immune Dysregulation in COVID-19 Patients With Severe Respiratory Failure. Cell Host Microbe (2020), 1–9. doi: 10.1016/j.chom.2020.04.009
14. Consiglio CR, Cotugno N, Sardh F, Pou C, Amodio D, Rodriguez L, et al. The Immunology of Multisystem Inflammatory Syndrome in Children With COVID-19. Cell (2020) 183(4):968–81.e7. doi: 10.1016/j.cell.2020.09.016
15. Gruber CN, Patel RS, Trachtman R, Lepow L, Amanat F, Krammer F, et al. Mapping Systemic Inflammation and Antibody Responses in Multisystem Inflammatory Syndrome in Children (MIS-C). Cell (2020) 1–14. doi: 10.1016/j.cell.2020.09.034
16. Li YC, Bai WZ, Hashikawa T. The Neuroinvasive Potential of SARS-CoV2 may be at Least Partially Responsible for the Respiratory Failure of COVID-19 Patients. J Med Virol (2020), 552–5. doi: 10.1002/jmv.25728
17. Arbour N, Day R, Newcombe J, Talbot PJ. Neuroinvasion by Human Respiratory Coronaviruses. J Virol (2000) 74(19):8913–21. doi: 10.1128/jvi.74.19.8913-8921.2000
18. Burks JS, Devald BL, Jankovsky LD, Gerdes JC. Two Coronaviruses Isolated From Central Nervous System Tissue of Two Multiple Sclerosis Patients. Science (1980) 209(4459):933–4. doi: 10.1126/science.7403860
19. Stewart JN, Mounir S, Talbot PJ. Human Coronavirus Gene Expression in the Brains of Multiple Sclerosis Patients. Virology (1992) 191(1):502–5. doi: 10.1016/0042-6822(92)90220-J
20. Singh M, Bansal V, Feschotte C. A Single-Cell RNA Expression Map of Human Coronavirus Entry Factors. Cell Rep (2020) 32(12):108175. doi: 10.1016/j.celrep.2020.108175
21. Liotta EM, Batra A, Clark JR, Shlobin NA, Hoffman SC, Orban ZS, et al. Frequent Neurologic Manifestations and Encephalopathy-Associated Morbidity in Covid-19 Patients. Ann Clin Trans Neurol (2020) 7(11):2221–30. doi: 10.1002/acn3.51210
22. Fifi JT, Mocco J. COVID-19 Related Stroke in Young Individuals. Lancet Neurol (2020) 19:713–5. doi: 10.1016/S1474-4422(20)30272-6
23. Fridman S, Bullrich MB, Jimenez-Ruiz A, Costantini P, Shah P, Just C, et al. Stroke Risk, Phenotypes, and Death in COVID-19: Systematic Review and Newly Reported Cases. Neurology (2020) 95(24):e3373–85. doi: 10.1212/wnl.0000000000010851
24. Iadecola C, Anrather J, Kamel H. Effects of COVID-19 on the Nervous System. Cell (2020) 183:16–27.e1. doi: 10.1016/j.cell.2020.08.028
25. Bodro M, Compta Y, Llansó L, Esteller D, Doncel-Moriano A, Mesa A, et al. Increased CSF Levels of IL-1β, IL-6, and ACE in SARS-CoV-2-Associated Encephalitis. Neurol Neuroimmunol Neuroinflamm (2020) 7(5):e821. doi: 10.1212/NXI.0000000000000821
26. Reichard RR, Kashani KB, Boire NA, Constantopoulos E, Guo Y, Lucchinetti CF. Neuropathology of COVID-19: A Spectrum of Vascular and Acute Disseminated Encephalomyelitis (ADEM)-Like Pathology. Acta Neuropathol (2020) 140:1–6. doi: 10.1007/s00401-020-02166-2
27. Yang AC, Kern F, Losada PM, Agam MR, Maat CA, Schmartz GP, et al. Dysregulation of Brain and Choroid Plexus Cell Types in Severe COVID-19. Nature (2021), 1–10. doi: 10.1038/s41586-021-03710-0
28. Monaco S, Nicholas R, Reynolds R, Magliozzi R. Intrathecal Inflammation in Progressive Multiple Sclerosis. Int J Mol Sci (2020) 21:1–11. doi: 10.3390/ijms21218217
29. Rodríguez-Lorenzo S, Konings J, van der Pol S, Kamermans A, Amor S, van Horssen J, et al. Inflammation of the Choroid Plexus in Progressive Multiple Sclerosis: Accumulation of Granulocytes and T Cells. Acta Neuropathol Commun (2020) 8:9. doi: 10.1186/s40478-020-0885-1
30. Haas J, Rudolph H, Costa L, Faller S, Libicher S, Würthwein C, et al. The Choroid Plexus Is Permissive for a Preactivated Antigen-Experienced Memory B-Cell Subset in Multiple Sclerosis. Front Immunol (2021) 11:618544. doi: 10.3389/fimmu.2020.618544
31. Zelic M, Pontarelli F, Woodworth L, Zhu C, Mahan A, Ren Y, et al. RIPK1 Activation Mediates Neuroinflammation and Disease Progression in Multiple Sclerosis. Cell Rep (2021) 35(6):109112. doi: 10.1016/j.celrep.2021.109112
32. Schwabenland M, Salié H, Tanevski J, Killmer S, Lago MS, Schlaak AE, et al. Deep Spatial Profiling of Human COVID-19 Brains Reveals Neuroinflammation With Distinct Microanatomical Microglia-T-Cell Interactions. Immunity (2021) 54(7):1594–610. doi: 10.1016/j.immuni.2021.06.002
33. Song E, Bartley CM, Chow RD, Pleasure SJ, Wilson MR, Farhadian Correspondence SF. Divergent and Self-Reactive Immune Responses in the CNS of COVID-19 Patients With Neurological Symptoms. Cell Rep Med (2021) 2(5):100288. doi: 10.1016/j.xcrm.2021.100288
34. Franke C, Ferse C, Kreye J, Reincke SM, Sanchez-Sendin E, Rocco A, et al. High Frequency of Cerebrospinal Fluid Autoantibodies in COVID-19 Patients With Neurological Symptoms. Brain Behav Immun (2021) 93:415–9. doi: 10.1016/J.BBI.2020.12.022
35. Hung IF, Lung K, Tso EY, Liu R, Chung TW, Chu M, et al. Articles Triple Combination of Interferon Beta-1b, Lopinavir – Ritonavir, and Ribavirin in the Treatment of Patients Admitted to Hospital With COVID-19 : An Open-Label, Randomised, Phase 2 Trial. Lancet (2020) 6736:1–10. doi: 10.1016/S0140-6736(20)31042-4
36. Cosorich I, Dalla-Costa G, Sorini C, Ferrarese R, Messina MJ, Dolpady J, et al. High Frequency of Intestinal TH17 Cells Correlates With Microbiota Alterations and Disease Activity in Multiple Sclerosis. Sci Adv (2017) 3(7):e1700492. doi: 10.1126/sciadv.1700492
37. Kadowaki A, Quintana FJ. The Gut–CNS Axis in Multiple Sclerosis. Trends Neurosci (2020) 43(8):622–34. doi: 10.1016/j.tins.2020.06.002
38. Moser T, Akgün K, Proschmann U, Sellner J, Ziemssen T. The Role of TH17 Cells in Multiple Sclerosis: Therapeutic Implications. Autoimmun Rev (2020) 19(10):102647. doi: 10.1016/j.autrev.2020.102647
39. Paterson RW, Brown RL, Benjamin L, Nortley R, Wiethoff S, Bharucha T, et al. The Emerging Spectrum of COVID-19 Neurology: Clinical, Radiological and Laboratory Findings. Brain (2020) 143:3104–20. doi: 10.1093/brain/awaa240
40. Zanin L, Saraceno G, Panciani PP, Renisi G, Signorini L, Migliorati K, et al. SARS-CoV-2 can Induce Brain and Spine Demyelinating Lesions. Acta Neurochirurg (2020) 162:1491–4. doi: 10.1007/s00701-020-04374-x
41. Dixon L, Varley J, Gontsarova A, Mallon D, Tona F, Muir D, et al. COVID-19-Related Acute Necrotizing Encephalopathy With Brain Stem Involvement in a Patient With Aplastic Anemia. Neurol Neuroimmunol Neuroinflamm (2020) 7:789. doi: 10.1212/NXI.0000000000000789
42. Poyiadji N, Shahin G, Noujaim D, Stone M, Patel S, Griffith B. COVID-19–Associated Acute Hemorrhagic Necrotizing Encephalopathy: CT and MRI Features. Radiology (2020) 296:E119–20. doi: 10.1148/RADIOL.2020201187
43. Handa R, Nanda S, Prasad A, Anand R, Zutshi D, Dass SK, et al. Covid-19-Associated Acute Haemorrhagic Leukoencephalomyelitis. Neurol Sci (2020) 41:3023–6. doi: 10.1007/s10072-020-04703-z
44. Varadan B, Shankar A, Rajakumar A, Subramanian S, Sathya AC, Hakeem AR, et al. Acute Hemorrhagic Leukoencephalitis in a COVID-19 Patient—a Case Report With Literature Review. Neuroradiology (2021) 63:1. doi: 10.1007/S00234-021-02667-1
45. Kumar A, Olivera A, Mueller N, Howard J, Lewis A. Delayed SARS-COV-2 Leukoencephalopathy Without Severe Hypoxia. J Neurol Sci (2020) 418:117146. doi: 10.1016/J.JNS.2020.117146
46. Novi G, Rossi T, Pedemonte E, Saitta L, Rolla C, Roccatagliata L, et al. Acute Disseminated Encephalomyelitis After SARS-CoV-2 Infection. Neurol Neuroimmunol Neuroinflamm (2020) 7:797. doi: 10.1212/NXI.0000000000000797
47. Hussein O, Elazim AA, Torbey MT. Covid-19 Systemic Infection Exacerbates Pre-Existing Acute Disseminated Encephalomyelitis (ADEM). J Neuroimmunol (2020) 349:577405. doi: 10.1016/J.JNEUROIM.2020.577405
48. Canavero I, Valentino F, Colombo E, Franciotta D, Ferrandi D, Mussa M, et al. Acute Myelopathies Associated to SARS-CoV-2 Infection: Viral or Immune-Mediated Damage? Travel Med Infect Dis (2021) 40:102000. doi: 10.1016/j.tmaid.2021.102000
49. Román GC, Gracia F, Torres A, Palacios A, Gracia K, Harris D. Acute Transverse Myelitis (ATM):Clinical Review of 43 Patients With COVID-19-Associated ATM and 3 Post-Vaccination ATM Serious Adverse Events With the ChAdOx1 Ncov-19 Vaccine (Azd1222). Front Immunol (2021) 12:653786. doi: 10.3389/fimmu.2021.653786
50. Sarma D, Bilello LA. A Case Report of Acute Transverse Myelitis Following Novel Coronavirus Infection Publication Date License A Case Report of Acute Transverse Myelitis Following Novel Coronavirus Infection. J Clin Pract cases Emergency Med (2020) 4. doi: 10.5811/cpcem.2020.5.47937
51. Zoghi A, Ramezani M, Roozbeh M, Darazam IA. Sahraian MA. A Case of Possible Atypical Demyelinating Event of the Central Nervous System Following COVID-19. Mult Scler Relat Disord (2020) 44:102324. doi: 10.1016/j.msard.2020.102324
52. Zhou S, Jones-Lopez EC, Soneji DJ, Azevedo CJ, Patel VR. Myelin Oligodendrocyte Glycoprotein Antibody-Associated Optic Neuritis and Myelitis in COVID-19. J Neuro-Ophthalmol (2020) 40:398–402. doi: 10.1097/WNO.0000000000001049
53. Corrêa DG, de Souza Lima FC, da Cruz Bezerra D, Coutinho AC, Hygino da Cruz LC. COVID-19 Associated With Encephalomyeloradiculitis and Positive Anti-Aquaporin-4 Antibodies: Cause or Coincidence? Mult Scler J (2021) 27:973–6. doi: 10.1177/1352458520949988
54. Shaw VC, Chander G, Puttanna A. Neuromyelitis Optica Spectrum Disorder Secondary to COVID-19. Br J Hosp Med (2020) 81:1–3. doi: 10.12968/hmed.2020.0401
55. Rodríguez de Antonio LA, González-Suárez I, Fernández-Barriuso I, Rabasa Pérez M. Para-Infectious Anti-GD2/GD3 IgM Myelitis During the Covid-19 Pandemic: Case Report and Literature Review. Mult Scler Relat Disord (2021) 49:102783. doi: 10.1016/j.msard.2021.102783
56. Masuccio FG, Barra M, Claudio G, Claudio S. A Rare Case of Acute Motor Axonal Neuropathy and Myelitis Related to SARS-CoV-2 Infection. J Neurol (2020) 268:2327–30. doi: 10.1007/S00415-020-10219-5
57. Bellucci G, Ballerini C, Mechelli R, Bigi R, Rinaldi V, Reniè R, et al. SARS-CoV-2 Meta-Interactome Suggests Disease-Specific, Autoimmune Pathophysiologies and Therapeutic Targets. F1000Research (2020) 9:992. doi: 10.12688/f1000research.25593.1
58. Annibali V, Mechelli R, Romano S, Buscarinu MC, Fornasiero A, Umeton R, et al. IFN-β and Multiple Sclerosis: From Etiology to Therapy and Back. Cytokine Growth Factor Rev (2015) 26(2):221–8. doi: 10.1016/j.cytogfr.2014.10.010
59. Severa M, Farina C, Salvetti M, Coccia EM. Three Decades of Interferon- β in Multiple Sclerosis: Can We Repurpose This Information for the Management of SARS-CoV2 Infection? Front Immunol (2020) 11:1–4. doi: 10.3389/fimmu.2020.01459
60. Blanco-Melo D, Nilsson-Payant BE, Liu WC, Uhl S, Hoagland D, Møller R, et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell (2020) 181:1036–1045.e9. doi: 10.1016/j.cell.2020.04.026
61. Acharya D, Liu G, Gack MU. Dysregulation of Type I Interferon Responses in COVID-19. Nat Rev Immunol (2020) 20:397–8. doi: 10.1038/s41577-020-0346-x
62. Sormani MP, Salvetti M, Labauge P, Schiavetti I, Zephir H, Carmisciano L, et al. DMTs and Covid-19 Severity in MS: A Pooled Analysis From Italy and France. 1Ann Clin Trans Neurol (2021) 8(8):1738–44. doi: 10.1002/ACN3.51408
63. Xia H, Cao Z, Xie X, Zhang X, Chen JYC, Wang H, et al. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep (2020) 33:108234. doi: 10.1016/j.celrep.2020.108234
64. Zhang Q, Bastard P, Liu Z, Le Pen J, Moncada-Velez M, Chen J, et al. Inborn Errors of Type I IFN Immunity in Patients With Life-Threatening COVID-19. Science (2020) 370(6515). doi: 10.1126/science.abd4570
65. Bastard P, Rosen LB, Zhang Q, Zhang Y, Dorgham K, Béziat V, et al. IgG Autoantibodies Against Type I IFNs in Patients With Severe COVID-19. Science (2020) 370(6515):eabd4585. doi: 10.1126/science.abd4585
66. Lokugamage KG, Hage A, Schindewolf C, Rajsbaum R, Menachery VD. SARS-CoV-2 Is Sensitive to Type I Interferon Pretreatment. J Virol (2020) 94:e01410–20. doi: 10.1128/JVI.01410-20
67. De Biasi S, Meschiari M, Gibellini L, Bellinazzi C, Borella R, Fidanza L, et al. Marked T Cell Activation, Senescence, Exhaustion and Skewing Towards TH17 in Patients With COVID-19 Pneumonia. Nat Commun (2020) 11:3434. doi: 10.1038/s41467-020-17292-4
68. Cao X. COVID-19: Immunopathology and its Implications for Therapy. Nat Rev Immunol (2020) 20:269–70. doi: 10.1038/s41577-020-0308-3
69. Pacha O, Sallman MA, Evans SE. COVID-19: A Case for Inhibiting IL-17? Nat Rev Immunol (2020) 20:345–6. doi: 10.1038/s41577-020-0328-z
70. Gordon DE, Gordon DE, Hiatt J, Bouhaddou M, Rezelj VV, Ulferts S. Comparative Host-Coronavirus Protein Interaction Networks Reveal Pan-Viral Disease Mechanisms. Science (2020) 9403:1–38. doi: 10.1126/science.abe9403
71. Lamers MM, Beumer J, Van Der Vaart J, Knoops K, Puschhof J, Breugem TI, et al. SARS-CoV-2 Productively Infects Human Gut Enterocytes. Science (2020) 369(6499):50–4. doi: 10.1126/science.abc1669
72. Zuo T, Zhang F, Lui GCY, Yeoh YK, Li AYL, Zhan H, et al. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastroenterology (2020) 159(3):944–55.E8. doi: 10.1053/j.gastro.2020.05.048
73. Scaldaferri F, Ianiro G, Privitera G, Lopetuso LR, Vetrone LM, Petito V, et al. The Thrilling Journey of SARS-CoV-2 Into the Intestine: From Pathogenesis to Future Clinical Implications. Inflammatory Bowel Dis (2020) 26(9):1306–14. doi: 10.1093/ibd/izaa181
74. Malhotra S, Costa C, Eixarch H, Keller CW, Amman L, Martínez-Banaclocha H, et al. NLRP3 Inflammasome as Prognostic Factor and Therapeutic Target in Primary Progressive Multiple Sclerosis Patients. Brain (2020) 143(5):1414–30. doi: 10.1093/brain/awaa084
75. Guarda G, Braun M, Staehli F, Tardivel A, Mattmann C, Förster I, et al. Type I Interferon Inhibits Interleukin-1 Production and Inflammasome Activation. Immunity (2011) 34(2):213–23. doi: 10.1016/j.immuni.2011.02.006
76. Soares JL, Oliveira EM, Pontillo A. Variants in NLRP3 and NLRC4 Inflammasome Associate With Susceptibility and Severity of Multiple Sclerosis. Mult Scler Relat Disord (2019) 29:26–34. doi: 10.1016/j.msard.2019.01.023
77. Ong EZ, Chan YFZ, Leong WY, Lee NMY, Kalimuddin S, Haja Mohideen SM, et al. A Dynamic Immune Response Shapes COVID-19 Progression. Cell Host Microbe (2020) 27(6):879–82. doi: 10.1016/j.chom.2020.03.021
78. Freeman TL, Swartz TH. Targeting the NLRP3 Inflammasome in Severe COVID-19. Front Immunol (2020) 11:1518. doi: 10.3389/fimmu.2020.01518
79. van den Berg DF, te Velde AA. Severe COVID-19: NLRP3 Inflammasome Dysregulated. Front Immunol (2020). doi: 10.3389/fimmu.2020.01580
80. Di Stadio A, Romani L, Bernitsas E. Could Sars-Cov2 Affect MS Progression? Mult Scler Relat Disord (2020) 46:102540. doi: 10.1016/j.msard.2020.102540
81. Mechelli R, Umeton R, Policano C, Annibali V, Coarelli G, Ricigliano VAG, et al. A “Candidate-Interactome” Aggregate Analysis of Genome-Wide Association Data in Multiple Sclerosis. PloS One (2013) 8:e63300. doi: 10.1371/journal.pone.0063300
82. Bar-Or A, Pender MP, Khanna R, Steinman L, Hartung HP, Maniar T, et al. Epstein–Barr Virus in Multiple Sclerosis: Theory and Emerging Immunotherapies. Trends Mol Med (2019) 26(3):1–15. doi: 10.1016/j.molmed.2019.11.003
83. Belbasis L, Bellou V, Evangelou E, Ioannidis JPA, Tzoulaki I. Environmental Risk Factors and Multiple Sclerosis: An Umbrella Review of Systematic Reviews and Meta-Analyses. Lancet Neurol (2015) 14(3):263–73. doi: 10.1016/S1474-4422(14)70267-4
84. Ascherio A, Munger KL. Epstein-Barr Virus Infection and Multiple Sclerosis: A Review. J Neuroimmune Pharmacol (2010) 5:271–7. doi: 10.1007/s11481-010-9201-3
85. Angelini DF, Serafini B, Piras E, Severa M, Coccia EM, Rosicarelli B, et al. Increased CD8+ T Cell Response to Epstein-Barr Virus Lytic Antigens in the Active Phase of Multiple Sclerosis. PloS Pathog (2013) 9:e1003220. doi: 10.1371/JOURNAL.PPAT.1003220
86. Serafini B, Rosicarelli B, Magliozzi R, Stigliano E, Aloisi F. Detection of Ectopic B-Cell Follicles With Germinal Centers in the Meninges of Patients With Secondary Progressive Multiple Sclerosis. Brain Pathol (2004) 14:164–74. doi: 10.1111/j.1750-3639.2004.tb00049.x
87. Serafini B, Rosicarelli B, Veroni C, Mazzola GA, Aloisi F. Epstein-Barr Virus-Specific CD8 T Cells Selectively Infiltrate the Multiple Sclerosis Brain and Interact Locally With Virus Infected Cells: Clue for a Virus-Driven Immunopathological Mechanism. J Virol (2019) 93:e00980–19. doi: 10.1128/jvi.00980-19
88. Magliozzi R, Howell O, Vora A, Serafini B, Nicholas R, Puopolo M, et al. Meningeal B-Cell Follicles in Secondary Progressive Multiple Sclerosis Associate With Early Onset of Disease and Severe Cortical Pathology. Brain (2007) 130(4):1089–104. doi: 10.1093/brain/awm038
89. Pender MP, Csurhes PA, Burrows JM, Burrows SR. Defective T-Cell Control of Epstein–Barr Virus Infection in Multiple Sclerosis. Clin Trans Immunol (2017) 6:e126. doi: 10.1038/CTI.2016.87
90. Zivadinov R, Cerza N, Hagemeier J, Carl E, Badgett D, Ramasamy DP, et al. Humoral Response to EBV is Associated With Cortical Atrophy and Lesion Burden in Patients With MS. Neurol - Neuroimmunol Neuroinflamm (2016) 3(1):e190. doi: 10.1212/nxi.0000000000000190
91. Lehner GF, Klein SJ, Zoller H, Peer A, Bellmann R, Joannidis M. Correlation of Interleukin-6 With Epstein–Barr Virus Levels in COVID-19. Crit Care (2020) 24:1–3. doi: 10.1186/S13054-020-03384-6
92. Simonnet A, Engelmann I, Moreau AS, Garcia B, Six S, el Kalioubie A, et al. High Incidence of Epstein–Barr Virus, Cytomegalovirus, and Human-Herpes Virus-6 Reactivations in Critically Ill Patients With COVID-19. Infect Dis Now (2021) 51:296–9. doi: 10.1016/J.IDNOW.2021.01.005
93. Chen T, Song J, Liu H, Zheng H, Chen C. Positive Epstein–Barr Virus Detection in Coronavirus Disease 2019 (COVID-19) Patients. Sci Rep (2021) 11:1–7. doi: 10.1038/s41598-021-90351-y
94. Huang C, Huang L, Wang Y, Li X, Ren L, Gu X, et al. 6-Month Consequences of COVID-19 in Patients Discharged From Hospital: A Cohort Study. Lancet (2021) 397:220–32. doi: 10.1016/S0140-6736(20)32656-8
95. Nalbandian A, Sehgal K, Gupta A, Madhavan MV, McGroder C, Stevens JS, et al. Post-Acute COVID-19 Syndrome. Nat Med (2021) 27:601–6. doi: 10.1038/s41591-021-01283-z
96. Taquet M, Geddes JR, Husain M, Luciano S, Harrison PJ. 6-Month Neurological and Psychiatric Outcomes in 236 379 Survivors of COVID-19: A Retrospective Cohort Study Using Electronic Health Records. Lancet Psychiatry (2021) 8:416–27. doi: 10.1016/S2215-0366(21)00084-5
97. Wijeratne T, Crewther S. Post-COVID 19 Neurological Syndrome (PCNS); a Novel Syndrome With Challenges for the Global Neurology Community. J Neurol Sci (2020) 419:'117179. doi: 10.1016/J.JNS.2020.117179
98. White P. Long COVID: Don’t Consign ME/CFS to History. Nature (2020) 587:197. doi: 10.1038/D41586-020-03136-0
99. Jones JF, Ray CG, Minnich LL, Hicks MJ, Kibler R, Lucas DO. Evidence for Active Epstein-Barr Virus Infection in Patients With Persistent, Unexplained Illnesses: Elevated Anti-Early Antigen Antibodies. Ann Internal Med (1985) 102:1–7. doi: 10.7326/0003-4819-102-1-
100. Straus SE, Tosato G, Armstrong G, Lawley T, Preble OT, Henle W, et al. Persisting Illness and Fatigue in Adults With Evidence of Epstein-Barr Virus Infection. Ann Internal Med (1985) 102:7–16. doi: 10.7326/0003-4819-102-1-7
101. Josephs SF, Henry B, Balachandran N, Strayer D, Peterson D, Komaroff AL. Ablashi D V. HHV-6 Reactivation in Chronic Fatigue Syndrome. Lancet (1991) 337:1346–7. doi: 10.1016/0140-6736(91)93018-5
102. Ablashi DV, Eastman HB, Owen CB, Roman MM, Friedman J, Zabriskie JB, et al. Frequent HHV-6 Reactivation in Multiple Sclerosis (MS) and Chronic Fatigue Syndrome (CFS) Patients. J Clin Virol (2000) 16:179–91. doi: 10.1016/S1386-6532(99)00079-7
103. Komaroff AL, Lipkin WI. Insights From Myalgic Encephalomyelitis/Chronic Fatigue Syndrome May Help Unravel the Pathogenesis of Post-Acute COVID-19 Syndrome. Trends Mol Med (2021) 27(9):895–906. doi: 10.1016/j.molmed.2021.06.002
104. Fukuda K. SSIHMSJDAK. The Chronic Fatigue Syndrome: A Comprehensive Approach to its Definition and Study. International Chronic Fatigue Syndrome Study Group. Ann Intern Med (1994) 121:953–9. doi: 10.7326/0003-4819-121-12-199412150-00009
105. Tomas C, Lodge TA, Potter M, Elson JL, Newton JL, Morten KJ. Assessing Cellular Energy Dysfunction in CFS/ME Using a Commercially Available Laboratory Test. Sci Rep (2019) 9:1–9. doi: 10.1038/s41598-019-47966-z
106. Tanaka M, Ishii A, Watanabe Y. Neural Dysfunction in Chronic Fatigue Syndrome. Adv Neuroimmune Biol (2013) 4(4):291–300. doi: 10.3233/NIB-130065
107. Berger JR, Pocoski J, Preblick R, Boklage S. Fatigue Heralding Multiple Sclerosis. Mult Scler (2013) 19:1526–32. doi: 10.1177/1352458513477924
108. Boerio JLJHAC. Pathophysiology D. And Treatment of Fatigue in Multiple Sclerosis. Rev Neurol (2006) 162:311–20. doi: 10.1016/s0035-3787(06)75017-2
109. Braley TJ, Chervin RD. Fatigue in Multiple Sclerosis: Mechanisms, Evaluation, and Treatment. Sleep (2010) 33:1061. doi: 10.1093/SLEEP/33.8.1061
110. Nourbakhsh B, Revirajan N, Morris B, Cordano C, Creasman J, Manguinao M, et al. Safety and Efficacy of Amantadine, Modafinil, and Methylphenidate for Fatigue in Multiple Sclerosis: A Randomized, Crossover, Double-Blind Trial. Lancet Neurol (2021) 20:38. doi: 10.1016/S1474-4422(20)30354-9
111. Morris G, Berk M, Puri BK. A Comparison of Neuroimaging Abnormalities in Multiple Sclerosis, Major Depression and Chronic Fatigue Syndrome (Myalgic Encephalomyelitis): Is There a Common Cause? Mol Neurobiol (2018) 55:3592–609. doi: 10.1007/s12035-017-0598-z
112. Gaber TA-ZK, Oo WW, Ringrose H. Multiple Sclerosis/Chronic Fatigue Syndrome Overlap: When Two Common Disorders Collide. NeuroRehabilitation (2014) 35:529–34. doi: 10.3233/NRE-141146
113. Morris G, Maes M. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome and Encephalomyelitis Disseminata/Multiple Sclerosis Show Remarkable Levels of Similarity in Phenomenology and Neuroimmune Characteristics. BMC Med (2013) 11:1–23. doi: 10.1186/1741-7015-11-205
114. Natelson BH, Johnson SK, DeLuca J, Sisto S, Ellis SP, Hill N, et al. Reducing Heterogeneity in Chronic Fatigue Syndrome: A Comparison With Depression and Multiple Sclerosis. Clin Infect Dis (1995) 21:1204–10. doi: 10.1093/CLINIDS/21.5.1204
115. Cliff JM, King EC, Lee J-S, Sepúlveda N, Wolf A-S, Kingdon C, et al. Cellular Immune Function in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Front Immunol (2019) 0:796. doi: 10.3389/FIMMU.2019.00796
116. Willing A, Leach OA, Ufer F, Attfield KE, Steinbach K, Kursawe N, et al. CD8+ MAIT Cells Infiltrate Into the CNS and Alterations in Their Blood Frequencies Correlate With IL-18 Serum Levels in Multiple Sclerosis. Eur J Immunol (2014) 44(10):3119–28. doi: 10.1002/eji.201344160
117. Annibali V, Ristori G, Angelini DF, Serafini B, Mechelli R, Cannoni S, et al. Cd161highcd8+T Cells Bear Pathogenetic Potential in Multiple Sclerosis. Brain (2011) 134:542–54. doi: 10.1093/BRAIN/AWQ354
118. Mechelli R, Romano S, Romano C, Morena E, Buscarinu MC, Bigi R, et al. MAIT Cells and Microbiota in Multiple Sclerosis and Other Autoimmune Diseases. Microorganisms (2021) 9:1132. doi: 10.3390/MICROORGANISMS9061132
119. Vavougios GD. A Data-Driven Hypothesis on the Epigenetic Dysregulation of Host Metabolism by SARS Coronaviral Infection: Potential Implications for the SARS-CoV-2 Modus Operandi. Med Hypotheses (2020) 140:109759. doi: 10.1016/j.mehy.2020.109759
120. Verma D, Mel Church T, Swaminathan S. Epstein-Barr Virus Lytic Replication Induces ACE2 Expression and Enhances SARS CoV-2 Pseudotyped Virus Entry in Epithelial Cells. J Virol (2021) 95(13):e00192–21. doi: 10.1128/jvi.00192-21
121. Cantuti-Castelvetri L, Ojha R, Pedro LD, Djannatian M, Franz J, Kuivanen S, et al. Neuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity. Science (2020) 370(6518):856–60. doi: 10.1126/science.abd2985
122. Daly JL, Simonetti B, Klein K, Chen K-E, Williamson MK, Antón-Plágaro C, et al. Neuropilin-1 is a Host Factor for SARS-CoV-2 Infection. Science (2020) 370(6518):861–5. doi: 10.1126/science.abd3072
123. Wang HB, Zhang H, Zhang JP, Li Y, Zhao B, Feng GK, et al. Neuropilin 1 is an Entry Factor That Promotes EBV Infection of Nasopharyngeal Epithelial Cells. Nat Commun (2015) 6:6240. doi: 10.1038/ncomms7240
124. Solomona BD, Muellera C, Chaeb WJ, Alabanzaa LM, Bynoea MS. Neuropilin-1 Attenuates Autoreactivity in Experimental Autoimmune Encephalomyelitis. Proc Natl Acad Sci USA (2011) 108(5):2040–5. doi: 10.1073/pnas.1008721108
125. Wang Y, Cao Y, Mangalam AK, Guo Y, LaFrance-Corey RG, Gamez JD, et al. Neuropilin-1 Modulates Interferon-γ-Stimulated Signaling in Brain Microvascular Endothelial Cells. J Cell Sci (2016) 129(20):3911–21. doi: 10.1242/jcs.190702
126. Illes Z, Gross CC, Geginat J, Bomfim LI, Engdahl E, Gustafsson R, et al. Increased Serological Response Against Human Herpesvirus 6a Is Associated With Risk for Multiple Sclerosis. Front Immunol (2019) 10:2715. doi: 10.3389/fimmu.2019.02715
127. Abadías-Granado I, Navarro-Bielsa A, Morales-Callaghan AM, Roc L, Suso-Estívalez CC, Povar-Echeverría M, et al. COVID-19-Associated Cutaneous Manifestations: Does Human Herpesvirus 6 Play an Aetiological Role? Br J Dermatol (2021) 184:1187–90. doi: 10.1111/BJD.19806
128. Jumah M, Rahman F, Figgie M, Prasad A, Zampino A, Fadhil A, et al. COVID-19, HHV6 and MOG Antibody: A Perfect Storm. J Neuroimmunol (2021) 353:577521. doi: 10.1016/j.jneuroim.2021.577521
129. Vieira JP, Sequeira J, Brito MJ. Postinfectious Anti-Myelin Oligodendrocyte Glycoprotein Antibody Positive Optic Neuritis and Myelitis. J Child Neurol (2017) 996–9. doi: 10.1177/0883073817724927
130. Fierz W. Multiple Sclerosis: An Example of Pathogenic Viral Interaction? Virol J (2017) 14:42. doi: 10.1186/s12985-017-0719-3
131. Rodrigues LS, Nali LH da S, Leal COD, Sabino EC, Lacerda EM, Kingdon CC, et al. HERV-K and HERV-W Transcriptional Activity in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Autoimmun Highlights (2019) 10:1–5. doi: 10.1186/S13317-019-0122-8
132. Hartung H-P, Derfuss T, Cree BA, Sormani MP, Selmaj K, Stutters J, et al. Efficacy and Safety of Temelimab in Multiple Sclerosis: Results of a Randomized Phase 2b and Extension Study. Mult Scler (2021) 135245852110249. doi: 10.1177/13524585211024997
133. Dolei A, Ibba G, Piu C, Serra C. Expression of HERV Genes as Possible Biomarker and Target in Neurodegenerative Diseases. Int J Mol Sci (2019) 20:3706. doi: 10.3390/IJMS20153706
134. Charvet B, Brunel J, Pierquin J, Mathieu C, Perron H. SARS-CoV-2 Induces Transcription of Human Endogenous Retrovirus RNA Followed by Type W Envelope Protein Expression in Human Lymphoid Cells. Res Square (2021). doi: 10.21203/RS.3.RS-301236/V1
135. Balestrieri E, Minutolo A, Petrone V, Fanelli M, Iannetta M, Malagnino V, et al. Evidence of the Pathogenic HERV-W Envelope Expression in T Lymphocytes in Association With the Respiratory Outcome of COVID-19 Patients. EBioMedicine (2021) 66:103341. doi: 10.1016/J.EBIOM.2021.103341
136. El-Shehawi AM, Alotaibi SS, Elseehy MM. Genomic Study of COVID-19 Corona Virus Excludes Its Origin From Recombination or Characterized Biological Sources and Suggests a Role for HERVS in Its Wide Range Symptoms. Cytol Genet (2020) 54:588. doi: 10.3103/S0095452720060031
137. Palao M, Fernández-Díaz E, Gracia-Gil J, Romero-Sánchez C, Díaz-Maroto I, Segura T. Multiple Sclerosis Following SARS-CoV-2 Infection. Mult Scler Relat Disord (2020) 45:102377. doi: 10.1016/j.msard.2020.102377
138. Fragoso YD, Pacheco FAS, Silveira GL, Oliveira RA, Carvalho VM, Martimbianco ALC. COVID-19 in a Temporal Relation to the Onset of Multiple Sclerosis. Mult Scler Relat Disord (2021) 50:102863. doi: 10.1016/j.msard.2021.102863
139. Moore L, Ghannam M, Manousakis G. A First Presentation of Multiple Sclerosis With Concurrent COVID-19 Infection. eNeurologicalSci (2021) 22:100299. doi: 10.1016/j.ensci.2020.100299
140. Naser Moghadasi A. A 31-Year-Old Female Patient With Concurrent Clinical Onset of Multiple Sclerosis and COVID-19: Possible Role of SARS-CoV-2 in the Pathogenesis of Multiple Sclerosis. Autoimmun Rev (2021) 20:102803. doi: 10.1016/j.autrev.2021.102803
141. Makhani N, Tremlett H. The Multiple Sclerosis Prodrome. Nat Rev Neurol (2021) 17:1–7. doi: 10.1038/s41582-021-00519-3
142. Steelman AJ. Infection as an Environmental Trigger of Multiple Sclerosis Disease Exacerbation. Front Immunol (2015) 6:520. doi: 10.3389/fimmu.2015.00520
143. Buljevac D, Flach HZ, Hop WCJ, Hijdra D, Laman JD, Savelkoul HFJ, et al. Prospective Study on the Relationship Between Infections and Multiple Sclerosis Exacerbations. Brain (2002) 125(5):952–60. doi: 10.1093/brain/awf098
144. Marrodan M, Alessandro L, Farez MF, Correale J. The Role of Infections in Multiple Sclerosis. Mult Scler J (2019) 25(7):891–901. doi: 10.1177/1352458518823940
145. Edwards S, Zvartau M, Clarke H, Irving W, Blumhardt LD. Clinical Relapses and Disease Activity on Magnetic Resonance Imaging Associated With Viral Upper Respiratory Tract Infections in Multiple Sclerosis. J Neurol Neurosurg Psychiatry (1998) 64:736–41. doi: 10.1136/jnnp.64.6.736
146. Correale J, Fiol M, Gilmore W. The Risk of Relapses in Multiple Sclerosis During Systemic Infections. Neurology (2006) 67(4):652–9. doi: 10.1212/01.wnl.0000233834.09743.3b
147. Kneider M, Lisovskaja V, Lycke J, Malmeström C, Jakobsen JK, Nerman O, et al. Upper Respiratory Infections and MRI Activity in Relapsing-Remitting MS. Mult Scler (2015) 45(2):83–9. doi: 10.1159/000437371
148. Thompson A, Banwell B, Barkhof F, Carroll W, Coetzee T, Comi G, et al. Diagnosis of Multiple Sclerosis: 2017 Revisions of the McDonald Criteria. Lancet Neurol (2018) 17:162–73. doi: 10.1016/S1474-4422(17)30470-2
149. Thrower BW. Relapse Management in Multiple Sclerosis. Neurologist (2009) 15:1–5. doi: 10.1097/NRL.0B013E31817ACF1A
150. Garjani A, Middleton RM, Hunter R, Tuite- KA, Coles A, Dobson R, et al. COVID-19 Is Associated With New Symptoms of Multiple Sclerosis that Are Prevented by Disease Modifying Therapies. Mult Scler Relat Disord (2021) 52:102939. doi: 10.1016/j.msard.2021.102939
151. Parrotta E, Kister I, Charvet L, Sammarco C, Saha V, Charlson RE, et al. COVID-19 Outcomes in MS: Observational Study of Early Experience From NYU Multiple Sclerosis Comprehensive Care Center. Neurol Neuroimmunol Neuroinflamm (2020) 7(5). doi: 10.1212/NXI.0000000000000835
152. Riva A, Barcella V, Benatti SV, Capobianco M, Capra R, Cinque P, et al. Vaccinations in Patients With Multiple Sclerosis: A Delphi Consensus Statement. Mult Scler J (2021) 27:347–59. doi: 10.1177/1352458520952310
153. Etemadifar M, Sedaghat N, Aghababaee A, Kargaran PK, Maracy MR, Ganjalikhani-Hakemi M, et al. COVID-19 and the Risk of Relapse in Multiple Sclerosis Patients: A Fight With No Bystander Effect? Mult Scler Relat Disord (2021) 51:102915. doi: 10.1016/j.msard.2021.102915
154. Sormani MP. An Italian Programme for COVID-19 Infection in Multiple Sclerosis. Lancet Neurol (2020) 19:481–2. doi: 10.1016/S1474-4422(20)30147-2
155. Barzegar M, Mirmosayyeb O, Gajarzadeh M, Afshari-Safavi A, Nehzat N, Vaheb S, et al. COVID-19 Among Patients With Multiple Sclerosis. Neurol - Neuroimmunol Neuroinflamm (2021) 8:e1001. doi: 10.1212/nxi.0000000000001001
156. Karim SSA. Oliveira T De. New SARS-CoV-2 Variants — Clinical, Public Health, and Vaccine Implications. N Engl J Med (2021) 384:1866–8. doi: 10.1056/NEJMC2100362
157. Otero-Romero S, Ascherio A, Lebrun-Frénay C. Vaccinations in Multiple Sclerosis Patients Receiving Disease-Modifying Drugs. Curr Opin Neurol (2021) 34:322–8. doi: 10.1097/WCO.0000000000000929
158. Achiron A, Dolev M, Menascu S, Zohar D-N, Dreyer-Alster S, Miron S, et al. COVID-19 Vaccination in Patients With Multiple Sclerosis: What We Have Learnt by February 2021. Mult Scler (2021) 27:864–70. doi: 10.1177/13524585211003476
159. Lotan I, Wilf-Yarkoni A, Friedman Y, Stiebel-Kalish H, Steiner I, Hellmann MA. Safety of the BNT162b2 COVID-19 Vaccine in Multiple Sclerosis: Early Experience From a Tertiary MS Center in Israel. Eur J Neurol (2021), ene.15028. doi: 10.1111/ENE.15028
160. Langer-Gould A, Qian L, Tartof SY, Brara SM, Jacobsen SJ, Beaber BE, et al. Vaccines and the Risk of Multiple Sclerosis and Other Central Nervous System Demyelinating Diseases. JAMA Neurol (2014) 71:1506–13. doi: 10.1001/jamaneurol.2014.2633
161. Confavreux C, Suissa S, Saddler P, Bourdès V, Vukusic S. Vaccines in Multiple Sclerosis Study Group. Vaccinations and Risk of Relapses in Multiple Sclerosis. N Engl J Med (2001) 344:319–26. doi: 10.1056/NEJM200102013440501
162. Ristori G, Mechelli R, Salvetti M. Association Between Vaccines and Neuroinflammation: Time, Risks, and Benefits. JAMA Neurol (2015) 72:605–5. doi: 10.1001/JAMANEUROL.2015.71
163. Confavreux C, Suissa S, Saddier P, Bourdès V, Vukusic S. Vaccinations and the Risk of Relapse in Multiple Sclerosis. N Engl J Med (2009) 344:319–26. doi: 10.1056/NEJM200102013440501
164. Buttinelli C, Salvetti M, Ristori G. Vaccinations and Multiple Sclerosis. N Engl J Med (2001) 344:1795–6. doi: 10.1056/NEJM200106073442313.
165. Salvetti M, Pisani A, Bastianello S, Millefiorini E, Buttinelli C, Pozzilli C. Clinical and MRI Assessment of Disease Activity in Patients With Multiple Sclerosis After Influenza Vaccination. J Neurol (1995) 242:143–6. doi: 10.1007/BF00936886
166. Havla J, Schultz Y, Zimmermann H, Hohlfeld R, Danek A, Kümpfel T. First Manifestation of Multiple Sclerosis After Immunization With the Pfizer-BioNTech COVID-19 Vaccine. J Neurol (2021) 1:1. doi: 10.1007/S00415-021-10648-W
167. Woopen C, Schleußner K, Akgün K, Ziemssen T. Approach to SARS-CoV-2 Vaccination in Patients With Multiple Sclerosis. Front Immunol (2021) 0:701752. doi: 10.3389/FIMMU.2021.701752
168. Bar-Or A, Calkwood JC, Chognot C, Evershed J, Fox EJ, Herman A, et al. Effect of Ocrelizumab on Vaccine Responses in Patients With Multiple Sclerosis: The VELOCE Study. Neurology (2020) 95:e1999–2008. doi: 10.1212/WNL.0000000000010380
169. Zabalza A, Cárdenas-Robledo S, Tagliani P, Arrambide G, Otero-Romero S, Carbonell-Mirabent P, et al. COVID-19 in Multiple Sclerosis Patients: Susceptibility, Severity Risk Factors and Serological Response. Eur J Neurol (2020) 28(10):3384–95. doi: 10.1111/ene.14690
170. Sormani MP, Schiavetti I, Landi D, Carmisciano L, de RN, Cordioli C, et al. SARS-CoV-2 Serology After COVID-19 in Multiple Sclerosis: An International Cohort Study. Mult Scler (2021) 135245852110353. doi: 10.1177/13524585211035318
171. Conte WL. Attenuation of Antibody Response to SARS-CoV-2 Infection in Patients With Multiple Sclerosis on Ocrelizumab: A Case-Control Study. Mult Scler Relat Disord (2021) 52:103014. doi: 10.1016/J.MSARD.2021.103014
172. Bigaut K, Kremer L, Fabacher T, Lanotte L, Fleury M-C, Collongues N. Seze J De. Impact of Disease-Modifying Treatments of Multiple Sclerosis on Anti–SARS-CoV-2 Antibodies. Neurol - Neuroimmunol Neuroinflamm (2021) 8:e1055. doi: 10.1212/NXI.0000000000001055
173. Bigaut K, Kremer L, Fleury M, Lanotte L, Collongues N, de Seze J. Impact of Disease-Modifying Treatments on Humoral Response After COVID-19 Vaccination: A Mirror of the Response After SARS-CoV-2 Infection. Rev Neurol (2021) S0035-3787(21):00569–5. doi: 10.1016/j.neurol.2021.05.001
174. Gallo A, Capuano R, Donnarumma G, Bisecco A, Grimaldi E, Conte M, et al. Preliminary Evidence of Blunted Humoral Response to SARS-CoV-2 mRNA Vaccine in Multiple Sclerosis Patients Treated With Ocrelizumab. Neurol Sci (2021) 42:3523–6. doi: 10.1007/s10072-021-05397-7
175. Achiron A, Mandel M, Dreyer-Alster S, Harari G, Magalashvili D, Sonis P, et al. Humoral Immune Response to COVID-19 mRNA Vaccine in Patients With Multiple Sclerosis Treated With High-Efficacy Disease-Modifying Therapies. Ther Adv Neurol Disord (2021) 14. doi: 10.1177/17562864211012835
176. Buttari F, Bruno A, Dolcetti E, Azzolini F, Bellantonio P, Centonze D, et al. COVID-19 Vaccines in Multiple Sclerosis Treated With Cladribine or Ocrelizumab. Mult Scler Relat Disord (2021) 52:102983. doi: 10.1016/j.msard.2021.102983
177. Guerreri S, Lazzarin S, Zanetta C, Nozzolillo A, Filippi M, Moiola L. Serological Response to SARS-CoV-2 Vaccination in Multiple Sclerosis Patients Treated With Fingolimod or Ocrelizumab: An Initial Real-Life Experience. J Neurol (2021) 1:1–5. doi: 10.1007/S00415-021-10663-X
178. Drulovic J, Ivanovic J, Martinovic V, Tamas O, Veselinovic N, Cujic D, et al. Humoral Response to Sars-Cov-2 and Covid-19 Vaccines in Patients With Multiple Sclerosis Treatted With Immune Reconstitution Therapies. Mult Scler Relat Disord (2021) 54:103150. doi: 10.1016/J.MSARD.2021.10315
179. Sormani MP, Inglese M, Schiavetti I, Carmisciano L, Laroni A, Lapucci C, et al. Effect of SARS-CoV-2 mRNA Vaccination in MS Patients Treated With Disease Modifying Therapies. SSRN Electron J (2021). doi: 10.2139/SSRN.3886420
180. Tarke A, Sidney J, Methot N, Yu ED, Zhang Y, Dan JM, et al. Impact of SARS-CoV-2 Variants on the Total CD4+ and CD8+ T cell Reactivity in Infected or Vaccinated Individuals. Cell Rep Med (2021) 2:100355. doi: 10.1016/J.XCRM.2021.100355
181. Geers D, Shamier MC, Bogers S, den Hartog G, Gommers L, Nieuwkoop NN, et al. SARS-CoV-2 Variants of Concern Partially Escape Humoral But Not T Cell Responses in COVID-19 Convalescent Donors and Vaccine Recipients. Sci Immunol (2021) 6:1750. doi: 10.1126/SCIIMMUNOL.ABJ1750
182. Kister I, Krogsgaard M, Mulligan MJ, Patskovsky Y, Voloshyna I, Ferstler N, et al. Preliminary Results of Ongoing, Prospective Study of Antibody and T-Cell Responses to SARS-CoV-2 in Patients With MS on Ocrelizumab or Other Disease-Modifying Therapies, in: American Academy of Neurology 73rd Annual Meeting (2021). Available at: https://www.aan.com/siteassets/home-page/conferences-and-community/annual-meeting/abstracts-and-awards/abstracts/2021-emerging-science-abstracts.pdf (Accessed August 2, 2021).
183. Apostolidis SA, Kakara M, Painter MM, Goel RR, Mathew D, Lenzi K, et al. Altered Cellular and Humoral Immune Responses Following SARS-CoV-2 mRNA Vaccination in Patients With Multiple Sclerosis on Anti-CD20 Therapy. doi: 10.1101/2021.06.23.21259389
184. Suwanwongse K, Shabarek N. Benign Course of COVID-19 in a Multiple Sclerosis Patient Treated With Ocrelizumab. Mult Scler Relat Disord (2020) 42:102201. doi: 10.1016/j.msard.2020.102201
185. Novi G, Mikulska M, Briano F, Toscanini F, Tazza F, Uccelli A, et al. COVID-19 in a MS Patient Treated With Ocrelizumab: Does Immunosuppression Have a Protective Role? Mult Scler Relat Disord (2020) 42:9–10. doi: 10.1016/j.msard.2020.102120
186. Centonze D, Rocca MA, Gasperini C, Kappos L, Hartung H-P, Magyari M, et al. Disease-Modifying Therapies and SARS-CoV-2 Vaccination in Multiple Sclerosis: An Expert Consensus. J Neurol (2021) 10:1. doi: 10.1007/S00415-021-10545-2
187. Krienke C, Kolb L, Diken E, Streuber M, Kirchhoff S, Bukur T, et al. A Noninflammatory mRNA Vaccine for Treatment of Experimental Autoimmune Encephalomyelitis. Science (2021) 371:145–53. doi: 10.1126/SCIENCE.AAY3638
188. Drayman N, DeMarco JK, Jones KA, Azizi S-A, Froggatt HM, Tan K, et al. Masitinib is a Broad Coronavirus 3CL Inhibitor That Blocks Replication of SARS-CoV-2. Science (2021) 373(6557):931–6. doi: 10.1126/SCIENCE.ABG5827
189. Naser Moghadasi A. COVID-19-Related Autoimmune Disorders of Central Nervous System (CRAD-C): Is it a New Entity? Autoimmun Rev (2021) 20:102888. doi: 10.1016/J.AUTREV.2021.102888
Keywords: multiple sclerosis, SARS-CoV-2, COVID-19, virus-host interactions, neuroinflammation, vaccine, virus, EBV—Epstein-Barr Virus
Citation: Bellucci G, Rinaldi V, Buscarinu MC, Reniè R, Bigi R, Pellicciari G, Morena E, Romano C, Marrone A, Mechelli R, Salvetti M and Ristori G (2021) Multiple Sclerosis and SARS-CoV-2: Has the Interplay Started? Front. Immunol. 12:755333. doi: 10.3389/fimmu.2021.755333
Received: 08 August 2021; Accepted: 03 September 2021;
Published: 27 September 2021.
Edited by:
Robert O. Watson, College of Medicine, Texas A&M University, United StatesReviewed by:
Eleonora Tavazzi, Buffalo Neuroimaging Analysis Center, United StatesAngelica Cuapio, Karolinska Institutet, Sweden
Copyright © 2021 Bellucci, Rinaldi, Buscarinu, Reniè, Bigi, Pellicciari, Morena, Romano, Marrone, Mechelli, Salvetti and Ristori. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marco Salvetti, bWFyY28uc2FsdmV0dGlAdW5pcm9tYTEuaXQ=