 Sherif A. El-Kafrawy1,2,3†
Sherif A. El-Kafrawy1,2,3† Aymn T. Abbas1,2,4*†Christopher Oelkrug5†Marwa Tahoon6Sameera Ezzat6,7Alimuddin Zumla1,8,9
Aymn T. Abbas1,2,4*†Christopher Oelkrug5†Marwa Tahoon6Sameera Ezzat6,7Alimuddin Zumla1,8,9 Esam I. Azhar1,2*†
Esam I. Azhar1,2*†- 1Special Infectious Agents Unit-BSL3, King Fahd Medical Research Center, King Abdulaziz University, Jeddah, Saudi Arabia
- 2Department of Medical Laboratory Sciences, Faculty of Applied Medical Sciences, King Abdulaziz University, Jeddah, Saudi Arabia
- 3Department of Clinical Pathology, National Liver Institute, Menoufia University, Shebin El-Kom, Egypt
- 4Biotechnology Research Laboratories, Gastroenterology, Surgery Centre, Mansoura University, Mansoura, Egypt
- 5Oelkrug Enterprises UG (haftungsbeschränkt), Ascheberg, Germany
- 6Epidemiology and Preventive Medicine Department, National Liver Institute, Menoufia University, Shebin El-Kom, Egypt
- 7MARC for Medical Services and Scientific Research, 6th of October City, Giza, Egypt
- 8Department of Infection, Division of Infection and Immunity, Centre for Clinical Microbiology, University College London, London, United Kingdom
- 9National Institute for Health and Care Research (NIHR) Biomedical Research Centre, University College London Hospitals, London, United Kingdom
Antibiotic resistant bacteria are a growing threat to global health security. Whilst the emergence of antimicrobial resistance (AMR) is a natural phenomenon, it is also driven by antibiotic exposure in health care, agriculture, and the environment. Antibiotic pressure and inappropriate use of antibiotics are important factors which drive resistance. Apart from their use to treat bacterial infections in humans, antibiotics also play an important role in animal husbandry. With limited antibiotic options, alternate strategies are required to overcome AMR. Passive immunization through oral, nasal and topical administration of egg yolk-derived IgY antibodies from immunized chickens were recently shown to be effective for treating bacterial infections in animals and humans. Immunization of chickens with specific antigens offers the possibility of creating specific antibodies targeting a wide range of antibiotic-resistant bacteria. In this review, we describe the growing global problem of antimicrobial resistance and highlight the promising potential of the use of egg yolk IgY antibodies for the treatment of bacterial infections, particularly those listed in the World Health Organization priority list.
Introduction
AMR occurs when a microorganism is able to survive in the presence of antibiotics at a concentration that would normally inhibit their growth (1). Antimicrobial resistant (AMR) bacteria are able to survive and thrive through natural selection (2). Multidrug resistance (MDR) refers to bacteria that are resistant to at least three classes of antibiotics. It arises from the presence of resistance-associated genes in the bacterial genome (3). Currently, an estimated 700,000 fatalities are attributed to (AMR) per year. By 2050, AMR could lead to about 10 million deaths per year, as well as a 2% to 3.5% loss in the gross domestic product and global social costs of up to 100 trillion USD (4).
A primary issue with the introduction of a new antibiotic is whether antimicrobial resistance (AMR) to it will eventually emerge (5). Multidrug resistance has been detected among gram-positive pathogens, with penicillin-resistant Streptococcus pneumoniae, methicillin-resistant Staphylococcus aureus, and vancomycin-resistant Enterococcus faecium, being of particular concern (6). Among gram-negative bacteria, resistance to third-generation cephalosporins followed by fluoroquinolones, carbapenems, and currently colistin among Enterobacteriaceae poses a global threat that has resulted in a large increase in mortality and treatment costs and changed the guidelines for the treatment and control of infection (7).
The major driver of resistance evolution is the overuse of antibiotics fostered by factors such as inadequate regulations and misuse, lack of awareness about proper practices and consequent unjustified or unskilled use of antibiotics, the use of antibiotics as growth promotors in poultry and livestock, unrestricted access to antibiotics (8). Antimicrobial resistance is a natural phenomenon that occurs over time and is usually due to genetic changes in an organism. Antimicrobial-resistant organisms are found in humans and all their living environments (animals, plants, water and soil) and can spread from human to human or through zoonotic transmission from animal(or animal products) to humans (9).
Resistance may be caused by one or more of the following mechanisms (2). Enzymatic inactivation of the antimicrobial compound as the case with beta-lactamases (10). Reducing the antimicrobial effect by modifying the metabolic pathways alters bacterial cell walls making antimicrobial agents lose their binding ability to the bacterial target (11). Modifying the antimicrobial targets includes overamplifying the target or altering the permeability of the cell membrane by either decreased influx (porin loss) or increased efflux (efflux pumps) leading to a reduction in the accumulation of antimicrobial agents inside the cell (12). Another mechanism by which bacteria might develop antimicrobial resistance is by acquiring efflux pumps that extrude the antibacterial agent from the cell before reaching the target site (11). However, antimicrobial resistance may be intrinsic or acquired; it can develop through the mutation of existing genes (13, 14), or through the transfer of genes from other species or strains (15, 16).
Strategies employed to overcome AMR include reducing the extensive use of antimicrobials, collecting and analysis of data, avoiding the overuse of antimicrobials in farm animals, and developing novel treatment approaches (17, 18). The development of novel nanoscale antimicrobial agents/nanocomposites has been reported on different microorganisms (18). The limitations, and/or health risks associated with these nano-sized particles need to be taken into consideration (19).
Antimicrobial peptides (AMPs) have been widely tested in the fight against AMR bacterial infections (20, 21). However, overuse of AMPs may result in more resistant forms of bacteria resulting in deadly infections (11). Another potential alternative is bacteriophages; these are bacterial viruses that act as pathogens against bacteria. They are abile to specifically attack and kill only their host bacterial cells (22). Their limitations include the development of antibodies after repeated treatment, rapid inactivation of phages by the spleen, endotoxin contamination of the therapeutic phage preparations from bacterial debris, limited host range, regulation, and bacterial resistance to phages (11). Plant-based therapeutic agents evolved as a therapeutic alternative due to the emergence of AMR infections and the growth of scientific knowledge about herbal medicines as a promising complementary treatment (23).
Antibodies, mostly produced in mammals, provide a useful alternative in the treatment of bacterial infections either directly by targeting bacterial surfaces or indirectly by neutralizing bacterial toxins and the virulence factors that are responsible for infection (24, 25). However, several challenges face the production of IgG antibodies in mammals including the weak immune responses of the antigens used, the pain and distress caused to animals by immunization, blood sample collection, and ultimately sacrifice (26) and the cost of the production, poor shelf life, and the scale-up required for the large-scale production (24, 27).
The search for a more efficient and economical approach for the production of antibodies without the harm caused to the animals has led to a growing interest in egg yolk antibodies (IgY) (28). IgY is an isotype of immunoglobulin found in birds. Large-scale production of antigen-specific IgY can be obtained from eggs laid by immunizing hens with the specific antigens (27). Passive immunization using IgY is a promising alternative approach to combat the emergence of new and current drug-resistant pathogens (29) The yield of IgY antibodies produced in eggs is 18 times superior to the amount produced in rabbits (30) thus reducing the number of animals needed and pain caused by blood collection and sacrifice (31). An extra benefit that IgYs provide is their high content of sialic acid (32) reported to increase the half-life of the drug (33) leading to the increased shelf life of the IgY antibodies. IgY antibodies retain activity through different manufacturing steps and dried IgY batches can keep their biological activity over several years (34, 35).
IgY antibodies are reported as potent preventive and/or therapeutic agents against several viruses such as Influenza A (36, 37), Rotavirus (38), Dengue (39), Zika (40), Ebola (41) and as we reported previously against MERS-CoV (42, 43) and SARS-COV-2 (44). IgYs were also tested for their anti-parasitic activities against Trypanosoma cruzi (45, 46), Cryptosporidium parvum (47), Eimeria (48), and Candida albicans as a fungal infection (49).
According to the WHO, there are 12 bacterial priority pathogens for which novel antibiotics are urgently needed (50). IgY antibodies showed activity against most of these pathogens. The aim of this review is to highlight the potential role that specific IgYs can play in the immunotherapeutic prevention and treatment of these antimicrobial-resistant pathogens.
Safety of the IgY antibodies with the different route of administration
IgYs are safer than IgGs as they do not bind to human Fc receptors or fix mammalian complement components; hence they do not induce dangerous immune responses (51). Hakalehto et al., 2021 reported that IgY antibodies are one of the safest possible therapeutic agents (52). IgY consumed orally is Generally Recognized as Safe (GRAS) by the U.S. Food and Drug Administration (53). Additionally, IgY antibodies have been used orally to treat pulmonary Pseudomonas aeruginosa-infected patients with no negative complications for up to 10 years (54).
Topical applications of the IgY antibodies (55) were reported against S. mutans as a gel or powder in a rat dental caries model and showed inhibition of S. mutans (56, 57). Patients who used toothpaste containing anti-gingivitis IgY showed significant differences in bleeding on probing and gingival index (58, 59). Short-term (three weeks) use of IgY mouth spray resulted in a significant decrease in S. mutans in dental plaque, and low levels of S. mutans were detected for at least 5 weeks after withdrawal of IgY (60).
Nasal delivery is superior in many cases to systemic delivery due to its non-invasive nature, fast onset of action, and low side effects due to targeted delivery (61–63). Anti-P. aeruginosa IgY were reported to inhibit murine pneumonia when administered intranasally (64). The protective effect and safety of intranasally administered anti-SARS-CoV-2 IgY antibodies were confirmed in a mouse model, with no adverse effects observed (65). Other reports showed that the superaficial application of anti-SARS-CoV-2 IgY would not be expected to elicit antibody-dependent enhancement of infection due to its topical application (53).
Systemic administration of IgY has not been clinically evaluated. IgY-based antivenom was given parenterally and showed complete protection in animal models of lethal venomous bites and stings (66–68). Moreover, intraperitoneal administration of polyvalent-specific anti-Zika virus IgY in a mouse model did not induce antibody-dependent enhancement (ADE) and did not display any side effects (40). More in depth studies on the safety and efficacy of IgY systemic delivery are needed before clinical use (69).
IgY antibodies as a candidat to overcome antibiotic resistance
The concept of passive immunization describes the administration of specific antibodies obtained from an immunized donor in a prophylactic or therapeutic setting. In general, passive immunization is a naturally occurring means of transferring immunity from a mother to an offspring, such as through the immunoglobulins contained in breastmilk in mammals and the transfer of IgY antibodies through the egg yolk in chickens (70). The concept of passive immunization has drawn increasing interest in the past several years owing to the increase in antibiotic-resistant pathogens. In general, the IgY technology can be used to develop highly specific antibodies against a vast variety of antigens including bacteria, viruses, and even bacterial enzymes such as beta-lactamase, which is able to inactivate antibiotics. These antibodies are seen as a novel approach to targeting antibiotic-resistant bacteria through passive immunization. The administration of developed and isolated IgYs can provide rapid protection against diseases that are currently unresponsive to antibiotic therapies, including among immunocompromised patients for whom conventional treatment or vaccinations are not effective. The commercial availability of reagents that are specifically designed and developed for use in egg yolk antibody isolation and characterization has increased the amount of research in this area, with the term IgY being internationally recognized in research and industry (71). One of the humane advantages of IgY use is the ability to extract the antibodies from the eggs and not from blood, which makes it more favorable for animal safety and care (72). In the 1990s, several reports investigated the various aspects of IgY technology and applications (73). A particular advantage of IgY-based diagnosis and treatment is that the phylogenetic distance between mammals and birds enables the generation of IgY antibodies against conserved mammalian or pathogen proteins (74). This phylogenetic distance results in no recognition of mammalian Fc receptors and does not trigger the mammalian complement activation in vitro or in vivo (75, 76). IgY provide an added environmental advantage by using nontoxic techniques for purification (77). Precipitation techniques using water dilution and low pH-induced precipitation, as well as polyethylene glycol-, dextran sulfate- and xanthan gum-induced precipitations (78) or NaCl extraction (79) have been employed for IgY purification.
Advantages of avian IgY antibodies over mammalian IgG for passive immunization
IgY antibodies have a large number of advantages over mammalian IgG antibodies such as cost-effectiveness, the short time needed for preparation and production, the wide range of potential pathogen targets, convenience in handling and storage, and the high yield of the target IgYs (53, 80). A hen can be considered a small “factory” for antibody production, as one hen can produce more than 22.5 g of total IgY per year of which 2% to 10% is composed of target-specific antibodies (81). This quantity is the equivalent of the IgG antibody production of 4.3 rabbits over the course of a year; further, this large amount of IgY can be harvested without killing the hen (29). IgY is more resistant to proteolysis than its mammalian IgG counterparts (82), and it has been found to retain 40% of its activity after incubation with trypsin or chymotrypsin for 8 hours (83). Moreover, owing to the phylogenetic distance between birds and mammals, immunizing laying hens twice with the specific antigen is enough to produce a humoral immune response that leads to the transfer of large amounts of specific IgY antibodies to the eggs for several months (53). IgYs are safer than IgGs as they do not bind human Fc receptors or fix mammalian complement components; hence, they do not initiate potentially dangerous immune responses (51). Owing to the lack of a hinge region between the two “arms” of the antibody molecule, the IgY molecular structure is more rigid than that of IgGs and thus somewhat stronger (84, 85).
Advantages of IgY compared with antibiotics
The use of polyclonal IgY against infectious diseases minimizes the risk of developing AMR. Since the antibodies are directed to various antigens of the same microorganism, this lowers the chance of developing resistance to all of these antigens at the same time because they require multiple genes for synthesis (86). Therefore, specific IgY antibodies are promising alternatives for use as antimicrobials in human and veterinary health to combat the emergence of resistant bacteria (28). The use of IgY is environmentally friendly and elicits no undesirable side effects, disease resistance, or toxic residues (87). IgY-based therapy does not cause disruption of the host flora because the treatments target specific disease-causing pathogens (88).
Target and antigen identification
The choice of a specific antigenic target depends on the characteristics of the pathogen and the therapeutic strategy itself. For example, IgY antibodies can target different factors important for the survival of bacteria such as enzymes, toxins, colonization factors, flagella, and mucosal receptors (89, 90). The actual mode of action of passive immunization includes the agglutination of bacteria, inhibition of bacterial adhesion, suppression of virulence factors, toxin neutralization, opsonization and enzyme inactivation (81).
The antigenicity of specific targets that are used to immunize the chickens can be influenced by the immunogen itself, the type of adjuvant used for immunization, route of antigen delivery, frequency of administration, and general avian properties (breed, commensal bacterial footprint, age, egg lying capacity) (91).
Four different target strategies of passive immunization with IgY antibodies can be distinguished and are further displayed in (Figure 1).
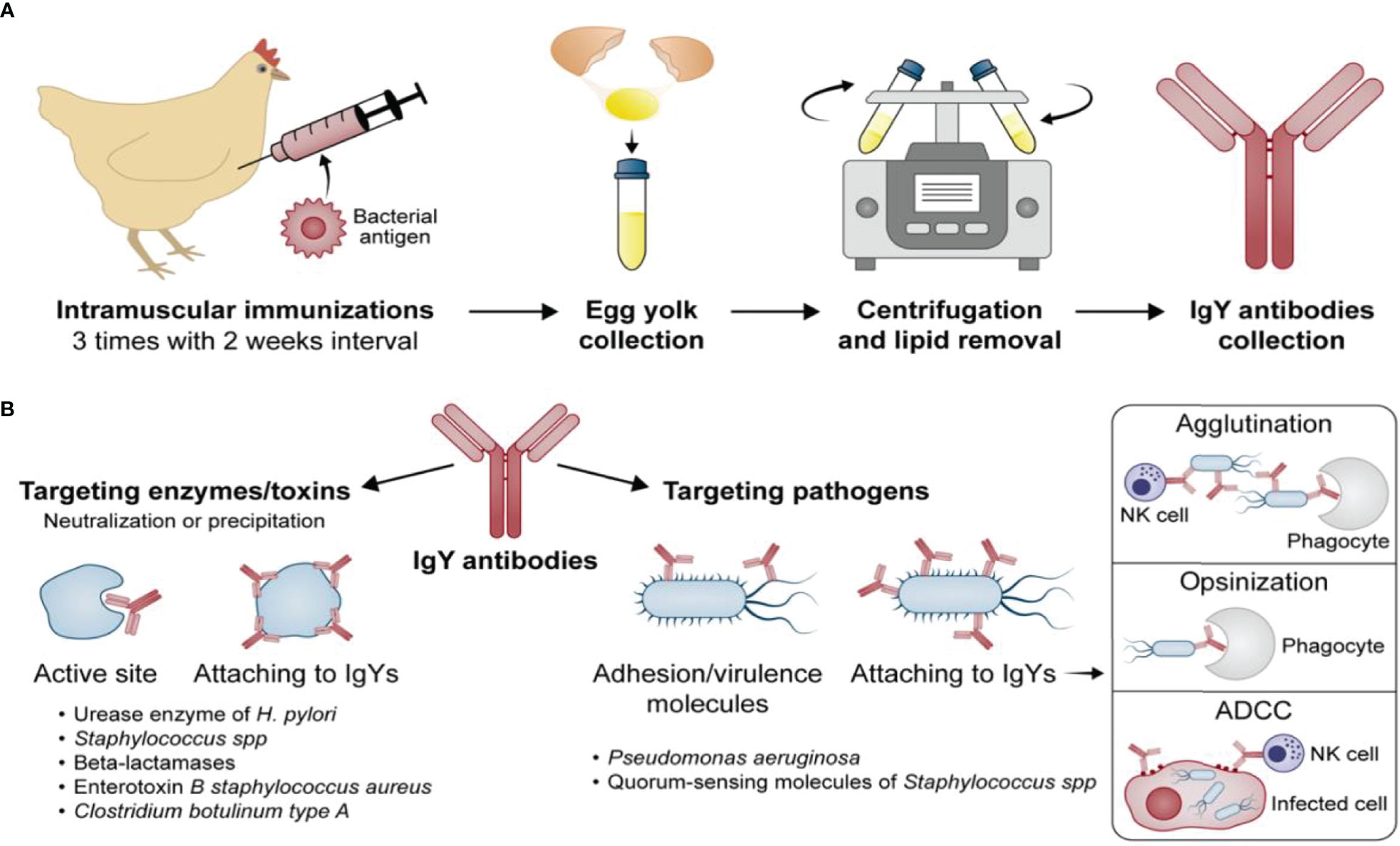
Figure 1 Development of the IgY antibodies (A) and their potential mechanisms of action (B).
Active site/whole enzyme
IgYs have been developed against crucial enzymes such as urease, which is expressed by bacteria such as Helicobacter pylori for survival in harsh conditions (low pH in the gastric mucosa), and beta-lactamases (92), which enable bacteria to degrade antibiotics and survive exposure to them. IgYs inhibit the enzymatic function (urease or beta-lactamase) through blockage of the active site, which prevents the substrate from binding to the enzyme and neutralizes the ability of bacteria to survive adverse conditions.
Research shows that IgYs generated against UreC, one of the subunits of urease, resulted in the prevention and even eradication of antibiotic-resistant H. pylori infections, which cause gastritis and gastric ulcers leading to gastric cancer (93). The addition of IgYs to yogurt as a functional food against H. pylori, for example, makes the usage of IgYs attractive (94).
Further research conducted by LeClaire et al. and Trott et al. (95, 96) also showed promising results regarding the therapeutic usage of IgYs generated against bacterial enzymes/toxins. These researchers reported that IgYs specific to enterotoxin B and botulinum type A neutralize their activity and can therefore prevent and treat infections due to S. aureus and Clostridium spp., respectively.
In addition, Hirai et al. (97) passively immunized mice with three different types of IgYs specific to Vibrio cholerae (anti-01, O139, and anti-cholera toxin B), which effectively prevented cholera infection in the mice.
Adhesion molecules and virulence factors
Several studies have shown that it is possible to generate IgYs that specifically target pathogens directly. These IgYs act by blocking the adhesion/virulence molecules of the bacteria or by coating the whole bacterial cell wall, which suppresses the biological functions of the pathogen itself. This method was adapted by Nilsson et al. (98) who conducted a 12-year study on prophylactic oral IgY treatment against Pseudomonas aeruginosa in 17 patients and reported the prevention of bacterial colonization in most cases. These results clearly indicated the potential of IgYs as a preventative option for respiratory infections. IgY in this treatment is reported to form an antibody barrier preventing P. aeruginosa from entering the lungs through the nose/oropharynx and binding to the epithelial surface of the mucosa. Gargling with IgY at night keeps active concentrations of Anti-Pseudomonas IgY in saliva and oropharyngeal mucosa till the next morning (98).
Furthermore, Dapunt et al. (99) generated IgYs to target quorum-sensing molecules of Staphylococcus spp. associated with implant infections. In particular, the adhesion molecule AtlE (S. epidermidis), which is a member of the autolysin family and mediates the attachment to the implant surface, was identified as a target of interest. Autolysins themselves are a group of enzymes that catalyze the degradation of the bacterial cell wall at specific sites. Besides the quorum-sensing molecules, the bacterial heat shock protein GroEL was also investigated in the context of biofilm formation. GroEL is a highly conserved protein that is homologous to the human heat shock protein 60 (HSP-60) and is essential for protein folding. It was previously shown that bacteria are not able to survive without GroEL, which makes it a perfect candidate as a potential target. Immune cells are able to recognize GroEL, which induces several bactericidal strategies. So far, two scientific reports have mentioned GroEL in relation to IgY and as a potential target. Hermans et al. (100) focused on Campylobacter jejuni in broiler chickens and used whole cell lysate to immunize chickens for the generation of IgY antibodies specific against the pathogen. GroEL was identified via HPLC-MS/MS in the whole cell lysate, and it was predicted that the polyclonal IgYs would also contain an antibody portion against GroEL. Therefore, there is no clear indication of the impact of GroEL-specific IgYs on the growth of C. jejuni. Dapunt et al. (101) have also investigated GroEL-specific IgYs against S. epidermis in biofilm formation. Unfortunately, the impact was only minimal and further research is needed.
Activity of the IgY antibodies against antibiotic-resistant bacteria
Helicobacter pylori
Helicobacter pylori, a gram-negative, spiral-shaped, microaerophilic flagellated human bacterium. It colonizes the stomach causing acute and chronic gastritis developing peptic ulcers in 10% to 20% of cases, gastric adenocarcinoma in 1% to 2%, and gastric lymphoma in less than 1% of cases (102–105). The misuse of antibiotics has increased resistance of H. pylori to CLR and MTZ which has reached 40%–50% and 70%–80% in some areas, respectively (106). As a consequence, antibiotic resistance has reduced the efficacy of standard triple therapy to 70% or lower (107, 108). Passive immunization with IgY directed against specific pathogens offers a potential alternative to antibiotics (109). Several in vitro and in vivo studies were performed to evaluate the efficacy of IgY antibodies against different H. pylori antigens including whole-cell bacteria (93, 110, 111), urease (89, 112–115), vacuolating cytotoxin A (VacA) proteins (116), neutrophil-activating protein (NAP) proteins (117), outer inflammatory protein (OipA) (118), cytotoxin-associated gene A (CagA) (119), and catalase (120). In addition, several H. pylori pathogenesis-related antigens can be used to generate IgY antibodies against different H. pylori targets such as sialic-acid-binding adhesion, blood group antigen binding adhesin, and γ-glutamyl transpeptidase (109). The mechanism of the IgY against H. pylori could be agglutination, help phagocytosis, or neutralization as well as blocking the adherence of the bacteria (109). Reports show that anti-H. pylori IgY antibodies effectively inhibit bacterial growth and adhesion to human gastric epithelial cells in vitro; also decrease H. pylori-induced gastric mucosal injury, improving gastritis, and attenuating gastric urease activity in vivo (109). Previously, we developed specific IgY antibodies from H. pylori cell lysate immunized chickens and evaluated their efficacy in a reliable H. pylori-infected mouse model with marked gastritis that was successfully developed by our research group (121). Results showed a significantly lower degree of infection and gastritis in IgY-treated animals than in untreated animals. A clinical trial was performed on 17 H. pylori asymptomatic volunteers who were orally administered egg powder containing anti-H.pylori urease IgY (Ovalgen, GHEN Corporation Inc., Gifu, Japan) for 4 weeks. The urea breath test (UBT) showed a significant decrease in UBT values although no case showed H. pylori eradication (122). Anti-H. pylori Urease IgY antibodies showed synergistic effects when taken at 3.4 g twice per day combined with lansoprazole (30 mg per day) for 8 weeks in a patient suffering from H. pylori-associated gastritis. The lansoprazole was taken to attenuate acid-induced inactivation of IgY. The treatment showed synergistic effects because IgY was shown to improve drug efficacy by reducing ulcer lesions (122).
Recently, a clinical trial evaluated multivalent IgY antibodies produced in chicken immunized with three recombinant H. pylori antigens urease B, flagellin A and antigen binding adhesion A2 (123). Ther trial included 94 H. pylori-positive volunteers, diagnosed as positive by a 13C UBT value of >4.0%. The recruited subjects were asked to administer a pack containing and 8.9 g of skim milk with 0.1 g of multivalent anti-H. pylori IgY 1 h before each meal for 2 weeks. The clinical symptoms of volunteers were followed up for 6 weeks after cessation of the administration period, followed by a 13C UBT which showed a decrease in UBT value by 56.0% with a total improvement rate of clinical symptoms in volunteers of 87.3%, and H. pylori eradication rate of 30.6% (123). The use of skim milk in this stusy was shown to effectively alleviate the degradation of IgY by pepsin under pH 1.2.
Horie K et al. (124) conducted a clinical trial on 42 volunteers divided into two groups. One group was given regular yogurt and the other was given yogurt mixed with 1.5 g of egg yolk containing about 45 mg of anti-H. pylori urease IgY three times daily. After oral administration of 2 and 4 weeks, the test group showed a significant reduction in the 13C UBT level by 34.19% and 39.3%, respectively with no side effects observed.
Several H. pylori-specific IgY antibodies have been applied for treatment; for example, a Chinese company used IgY antibodies as a raw material to develop chewing pills to treat and prevent H. pylori (125). Future advancements in antibody engineering will increase the application of IgY in passive immunization and therapy against H. pylori infections (125).
Pseudomonas aeruginosa
Pseudomonas aeruginosa is a gram-negative opportunistic bacterium causing chronic respiratory infections in patients with cystic fibrosis, chronic obstructive pulmonary disease, as well as acute infections in immunocompromised patients (126). The excessive use of antibiotics for the treatment accelerates the development of P. aeruginosa-resistant strains, leading to the failure of the antibiotic treatment (127).
A preclinical evaluation of IgY against P. aeruginosa was reported in which the IgY binding to the bacteria was shown to prevent adhesion to the oropharynx and hence prevent bacterial colonization (128).
Anti-Pseudomonas IgY was shown to promote bacterial opsonization and augment the phagocytic activity of human polymorphonuclear neutrophils (PMNs) (129), and it was also found to induce bacterial clearance in an animal model (130). As IgY antibodies do not activate Fc-receptors, the observed IgY-enhanced PMN phagocytosis might not be triggered by the usual receptor-mediated engulfment of opsonized bacteria. Improved phagocytosis might due to recognition of IgY by receptors similar to avian IgY receptors (131) or non-receptor-mediated mechanisms such as alterations in physio-chemical environment of the bacteria, facilitating a more easy and rapid phagocytosis (132). In an animal model, IgY administration to the lung was reported to reduce bacterial burden 100-fold compared with controls, and it was also accompanied by diminished lung inflammation and reduced clinical symptom scores (133). A recent study in mice showed that IgY antibodies provided 100% protection against all strains of P. aeruginosa upon intranasal challenge with 2×107 CFU directly into each nostril mixed with 500 μg of IgYs (133). Moreover, anti-flagellin IgY antibodies conferred protection against P. aeruginosa in a burned wound animal model, and they were found to confer dose-dependent efficacy covering all strain types (133). Sanches et al. (134) described an in vitro experiment showing the synergistic effects of IgY antibodies generated against SPM-1 or VIM-2-producing strains of P. aeruginosa. In that study, chickens were immunized with whole cell lysates of the bacterial strains, and the extracted IgY antibodies were tested in combination with the beta-lactam antibiotics ceftazidime, imipenem, or meropenem. The combination of IgYs and the beta-lactam antibiotics showed increased antimicrobial activity against resistant strains of bacteria. The authors were not able to describe the mode of action within this study.
Urinary tract infections (UTIs) with P. aeruginosa represent a major healthcare problem in disposed patients. P. aeruginosa establishes recalcitrant biofilm infections and can develop antibiotic resistance. In a recent study, P. aeruginosa (PAO1 and PAO3) was mixed with increasing concentrations of specific anti-Pseudomonas IgY (sIgY) or non-specific control IgY (cIgY) in vitro. The study showed a dose-dependent reduction in bacterial growth by the specific IgY at concentrations above 2.5%. In vivo effect of the IgY effect was evaluated in Balb/c mice which showed a reduction in vesical bacterial load by sIgY and cIgY when given the antibodies before infection (135).
A recent study evaluated the intranasal prophylactic effect of the anti-P. aeruginosa IgY antibodies (Pa-IgY) on the colonization of P. aeruginosa in the airways of a porcine model. Pa-IgY was administered through a nebulizer immediately before the administration of P. aeruginosa. A significant reduction was noticed in the Pa-IgY-treated group with the improvement of the physiological parameters (136). IgY against P. aeruginosa is now registered as an orphan drug in the European Medicines Agency (Designation number: EU/3/08/564). Patients were asked to gargle with IgY against P. aeruginosa in order to allow neutralization in the respiratory tract (137, 138). The clinical trial (NCT00633191) continued for about 10 years using a daily mouthwash containing 50 mg of specific IgY. None of the IgY-treated patients in this study became chronically colonized with P. aeruginosa with no side effects (54, 139). The activity of IgY against P. aeruginosa was shown in the saliva and oropharynx after gargle treatment with IgY solution (0.7 mg/ml) for 1 or 2 minutes and it was suggested that specific IgY is able to prevent P. aeruginosa invasion of the lungs (140). Phase III clinical evaluation (Clinicaltrials.gov Identifier: NCT01455675) of the effect of specific IgY antibodies on the recurrence of P. aeruginosa in the sputum of cyctic fibrosis patients asked to gargle with specific IgY solution every night after brushing their teeth. patients were followed up for 24 months or until the next P. aeruginosa infection whichever was first, no side effects were observed. Results showed a good tolerance profile for the immunoglobulin but did not show a clear therapeutic benefit of the anti-P. aeruginosa IgY treatment (141). The authors found that the placebo group reacted with far fewer events than expected, whereas the treated group followed the expected disease outcome. They hypothesized that the non-specific IgY may have unspecific inhibitory effects on protecting against reinfection of P. aeruginosa in cystic fibrosis patients.
Salmonella
The Salmonella species, particularly S. Typhimurium and S. Enteritidis, are human and chicken pathogens (142).
In several studies, IgYs were generated against S. Typhimurium and S. Enteritidis and were found to exhibit significant agglutination (143, 144) and cross-reactivity (145), indicating the potential therapeutic effect of IgY generated against a specific Salmonella serovar infection for treating a broad range of different Salmonella strains.
Effectiveness of Salmonella treatment for Salmonella by specific IgY was shown to be more effective if protected from gastrointestinal degradation (146). The effect of encapsulation on the effect of IgY activity at low pH of the stomach was evaluated in birds against Salmonella Enterica ssp (SI) under in vivo conditions (Table 1). Birds were orally given 1 mL of bacterial suspension and then divided into three groups, group 1 was given Salmonella Immune powdered yolk (SIPY), group 2 was given Salmonella non-immune powdered and the third group was given Salmonella capsulated immune yolk (SCIY). For positive control, Enrofloxacin was added to the drinking water of a fourth group. A significant difference in the reduction of the colonization of SI, evaluated by cecal content, was found between the SCIY group on days 14 and 21 and the SA, SCIY treatments (147).
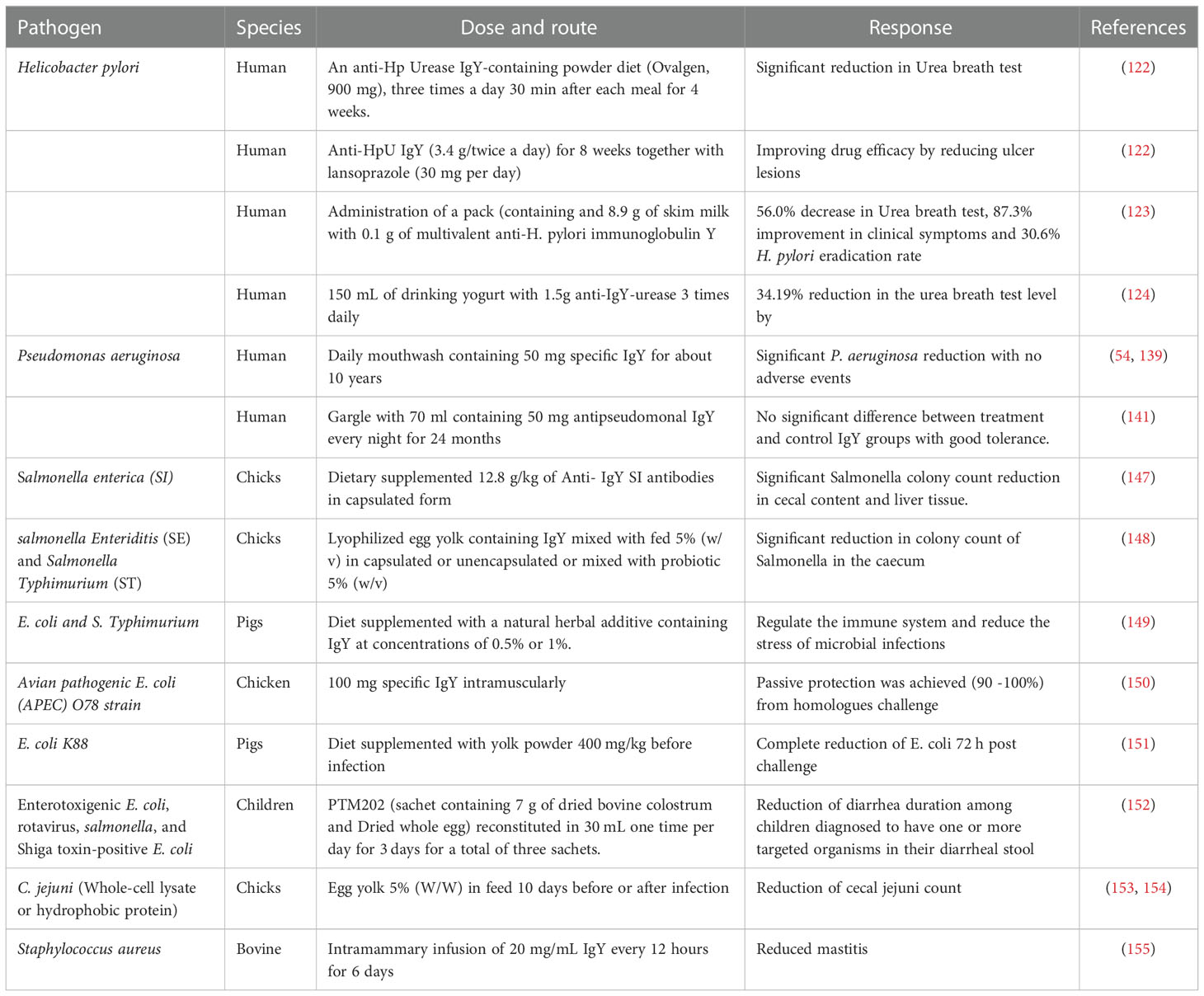
Table 1 Assessment of the efficacy of specific IgY antibodies against AMR for humans and animals.
In another trial, lyophilized egg yolk containing IgY from hens immunized either by Salmonella Enteritidis (SE) or Salmonella Typhimurium (ST) was orally delivered to a day old chicks mixed with their fed (Egg yolk only or encapsulated with liposomes 5% (w/w) or mixed with probiotic 5% (w/v) (Table 1). On day four, all chicks were challenged with (SE) or (ST) by oral inculcation using a blunt needle. One chick from each group was slaughtered on day 7th,14th, 21st and 28th. Salmonella was enumerated in ceacal content using SyBr Green real-time PCR. They found that serotype (SE, ST) specific anti-Salmonella IgY administered orally to the chicks significantly reduced Salmonella count in the caecum and there is no significant difference between the effect of the egg yolk only or encapsulate or egg yolk mixed with probiotics (148). On the other hand, IgY antibodies can resist digestion in the gastrointestinal tract of calves, remaining biologically active (35, 156).
The potential benefit of using IgY anti-Salmonella antibodies was proved when used in combination with probiotics which decreased colonization and fecal shedding in market-aged, young broiler chicks challenged with S. Enteritidis (157). This indicates the potential use of IgY anti-Salmonella antibodies in treating animal infections. In vitro evaluation of anti-Salmonella IgY antibodies in human epithelial Caco2 cells model showed that they prevent adhesion to cells (158).
An interesting study found that specific IgY against Salmonella could modulate the mucosal immune system of infected mice. Anti-S. Typhimurium IgY antibodies were orally administered with 0.4 mL of a solution containing 20 mg/kg once a day for 7 consecutive days after 3 days from infection. Nonspecific IgY or specific IgY has reduced the damage caused by S. Typhimurium challenge, and specific IgY treatment reduced jejunum ulceration, transmural inflammation, and edema significantly better than nonspecific IgY. Specific IgY diminished the effects of S. Typhimurium on the numbers of total T lymphocytes and CD8+ T cells while nonspecific IgY did not have the same effect (159). A commercial product is available in the market that contains IgY antibodies against E. coli and Salmonella (146).
IgY antibodies seem to be protective even if they are nonspecific. A study was performed on pigs infected with E. coli and S. Typhimurium that were fed a diet supplemented with a yolk sac containing 0.5% or 1% of IgY. Results showed oral egg yolk intake has regulated the immune system and reduced the stress due to microbial infections (149).
Escherichia coli
Escherichia coli is a component of the intestinal microbiota with several pathotypes involved in the development of enteric and extraintestinal infections such as sepsis, diarrhea, urinary tract infections, and meningitis (160).
Studies in animal and laboratory settings suggest that IgY targeting animal enteropathogens is effective in the prevention and treatment of diarrheal symptoms (161). Hens may be simultaneously immunized with multiple antigens, resulting in polyvalent IgY antibodies targeting multiple steps of the pathogenesis process (162). Accordingly, the effect of using IgY antibodies as prophylaxis of diarrheal illness caused by enteric pathogens was evaluated in vivo. This IgY has strong inhibitory effects on enterotoxigenic E. coli (ETEC) adherence which is a critical first step in host colonization and subsequent toxin delivery (163). In the veterinary setting, IgY antibodies generated by immunizing hens with selected antigens from E. coli were evaluated for the ability to protect broiler chickens from diseases caused by avian pathogenic E. coli (APEC). Intramuscular IgY (100 mg) injection into broiler chickens followed by challenge with homologous (O78) E. coli through the intra-air sac route 3 days later which resulted in prophylaxis against E. coli-associated respiratory, enteric, and septicemic diseases (150) (Table 1). Another study showed that pigs that administered diets supplemented with 400 mg/kg of IgY targeting E. coli K88 strain before infection were recovered (diarrhea score=0) after 72 h of challenge compare to those treated with non-specific IgY (151) (Table 1). Anti-E. coli O111 IgY antibodies were also found to inhibit the growth of the target pathogens and five other mastitis-causing strains of E. coli (164, 165).
A proprietary mixture of dried bovine colostrum and dried whole egg (PTM202, PanTheryx, Inc., Boulder, CO, USA) was given orally to 301 Guatemalan children with acute non-bloody diarrhea in a randomized, double-blind placebo-controlled trial. The treatment was designed to target enterotoxigenic E. coli, rotavirus, salmonella, and Shiga toxin-positive E. coli (152) (Table 1). The PTM202 treatment given orally as one full reconstituted sachet once a day for 3 days, resulted in the reduction of diarrhea duration among children diagnosed to have one or more targeted organisms in their diarrheal stool at enrollment with no adverse events. The study concluded that this IgY-based treatment represents a potential alternative to treat acute diarrheal disease in low/middle-income communities.
Campylobacter jejuni
Campylobacter species, particularly Campylobacter jejuni, is the most common etiology of human gastroenteritis worldwide (166). C. jejuni is transmitted to humans through poultry products with no effective eradication strategy from poultry production. Whole-cell lysate of C. jejuni was used to immunize chicken and the resulting egg yolk antibodies were fed to 6 days old chicks 5% (wt/wt) (Table 1). The chicks were inoculated orally with C. jejuni strain. Results showed that overall cecal C. jejuni count in chicken treated with C. jejuni IgY was significantly lower than the chicken treated with the nonspecific IgY antibodies. In addition, transmission to contact chicks was completely prevented (153).
In another recent study (154), two novel vaccines, a bacterin of 13 C. jejuni and C. coli strains and a subunit vaccine of six immunodominant Campylobacter antigens, were injected to immunize laying hens producing prolonged high levels of specific IgY in egg yolks. In vivo trial, yolks were orally in broiler feed 5% (wt/wt) for prophylaxis (11 days before infection) resulting in significant reduction in the number of Campylobacter-colonized broilers. In the therapeutic arm of the in vivo trial, administration of the IgY for 3 days mixed with fed 5% (wt/wt) resulted in a significant decrease in C. jejuni counts per infected bird. The hyperimmune yolks showed strong reactivity to a broad spectrum of C. jejuni and C. coli indicating that this passive immunization approach offers possibilities to control Campylobacter colonization in poultry (154).
Acinetobacter baumannii
Acinetobacter baumannii is a gram-negative bacillus that is a common cause of nosocomial infections. It is responsible for hospital-acquired sepsis, ventilator-associated pneumonia, skin and soft tissue infections, wound infections, urinary tract infections, secondary meningitis, and bloodstream infections (167, 168). Nosocomial outbreaks of A. baumannii present a considerable threat to ICU patients and are associated with increased mortality, longer hospital stays, and higher treatment costs (169). The wide use of broad-spectrum antibiotics has caused most A. baumannii strains to develop resistance to multiple antimicrobial agents (170), rendering the bacterial infection difficult to cure (171, 172).
In vitro study indicated that specific IgYs inhibited the growth of pan-drug-resistant A. baumannii (PDR-Ab) in a dose-dependent manner. The antimicrobial efficacy of the two IgYs developed against two A. baumannii strains were comparable to that of cefoperazone/sulbactam. Both IgYs showed significant growth inhibition of PDR-Ab at 20 mg/mL within 24 h (173).
Specific IgYs were reported to enhance bacterial agglutination, causing a CFU reduction rather than directly affecting individual bacteria (173). The binding of IgY to the bacteria was shown to cause cell crenation and structural modification on the cell surface, resulting in reduced bacterial attachment to the mucosa. The same effect was shown for specific IgY against H. pylori attaching to gastric cancer cells (93), and Salmonella attaching to intestinal cells (174, 175).
OmpA and Omp34 are essential virulence factors involved in A. baumannii adhesion to the human lung epithelial cell line. The protective effect of specific anti-acinetobacter IgYs raised against OmpA, Omp34 and inactivation of the whole-cell of A. baumannii was demonstrated (176). The therapeutic activity against the same antigens (OmpA and Omp34 or inactivated whole-cell of A. baumannii) was also conducted in another study, in which BALB/c mice were intranasally administrated 1.18 × l06 to 6 × l08 CFU A. baumannii, after 4 hours, 40 or 100 μg of specific IgY antibodies were intranasally administrated with therapeutic effect in a murine pneumonia model (177). A. baumannii increases its antimicrobial resistance through biofilms formation (178). Intranasal administration of anti- biofilm-associated protein (Bap) IgY antibodies was found to inhibit antibiotic-resistant strains of A. baumannii through the inhibition of biofilm formation (179–181).
Another in vivo study showed that intraperitoneal injection of anti-A. baumannii IgY antibodies in nasally infected BALB/c mice inhibited bacterial growth and protected mice from acute pneumonia induced by A. baumannii, suggesting the potential of these specific IgYs to be used as a new therapeutic alternative to treat PDR-Ab infections in humans (182).
Mycobacterium tuberculosis
Treatment of drug-resistant M. tuberculosis is a major health concern because such cases require second-line antibiotics, which are less effective, more expensive, and more toxic (183). Immunotherapy might provide an alternative for the treatment of drug-resistant TB strains, with promising outcomes and better quality of life for patients (184). In a rat peripheral blood mononuclear cell model, administration of high concentrations of IgY anti-M bacterium tuberculosis (anti-MBTC) increased interleukin (IL)-2 and interferon (IFN) expression (185). Production of these components has a major role in controlling antibody- and cell-mediated immunity, and the study results showed that IgY anti-M. tuberculosis could increase the production of IL-2 and IFN-γ and the proliferation of rat peripheral blood mononuclear cells in a concentration-dependent manner. Hens were immunized with four 80-μg doses of antigen, and anti-MBTC IgY antibodies in eggs were reported to reach a peak concentration at 4 weeks after immunization and to persist for 200 days after immunization. Western blot analysis showed the presence of anti‐MBTC IgY in egg yolks, with molecular weights of approximately 78 kDa (184). The authors concluded that IgY against MBTC may warrant evaluation for use in combination with other immunotherapeutic treatments of tuberculosis.
Staphylococcus aureus
The economic burden of antibiotic-resistant S. aureus (e.g., methicillin-resistant S. aureus) infections affects not only individual patients but also the healthcare systems of different countries owing to the persistence of infection, recurrent infections, a wide spectrum of clinical presentations, and diminished quality of life (186). Passive immunotherapy might provide an alternative for high-risk patients with prolonged hospitalization (187); however, native cross-species antibodies induce violent immune reactions. Additionally, S. aureus immune defenses, such as staphylococcal protein A (SpA) and staphylococcal binding immunoglobulin (Sbi), bind the Fc portion of these antibodies in reverse orientation to avoid complement-mediated killing and phagocytosis. IgY antibodies targeting SpA were tested in vitro and found to completely inhibit the growth of S. aureus at a concentration of 150 μg/mL, and to inhibit biofilm formation by ~45% showing potential use to neutralize these infections (188).
Specific IgY antibodies generated against S. aureus were shown to reduce mastitis during a 6-day intramammary infusion of 100 mg/mL IgY twice a day (155) (Table 1). Specific IgY against encapsulated type 5 (IgY-T5) and type 8 (IgY-T8) and non-encapsulated type 336 (IgY-T336) S. aureus strains (at 5 mg/mL) significantly blocked the internalization of bacteria bovine mammary epithelial cells within 6 h (189). authors suggest that the generated IgY antibodies control mastitis by preventing the uptake rather than by inhibiting the growth of bacteria.
IgY monoclonal antibodies
Successful generation of the monoclonal IgY or IgY fragments in the last few years has increased the functional use of IgY fragments, such as single chain (scFv) (190), chimeric (191), and humanized IgY (192). Monoclonal IgY antibodies combines the benefits of avian IgY antibodies and the features of monoclonal antibodies (193). They have the potential for use as therapeutics in both veterinary and human applications, immunological detection and diagnosis, and for screening and validating biomarkers (193).
Phage display production of IgY monoclonal antibody is more likely to generate a robust immune response against various highly conserved mammalian protein molecules (194). Monoclonal IgY antibodies conjugated with phthalocyanine—a synthetic photosensitizing dye used in near-infrared phototherapy—were recently used against Candida albicans and provided highly effective and specific success in an in vivo skin infection model with no damage to the healthy epithelium (195). In another study, hens were immunized with canine parvovirus VP2 (CPV-VP2) virus-like particles (VLP) and the specific IgY-scFv were generated using the T7 phage display technique (196). Transgenic IgY antibodies containing the bird constant regions and the human variable regions allow the use of the highly specific IgY antibodies against mammalian conserved proteins (197). So far one clinical trial is reported on the parenteral administration of monoclonal IgY product Sym021 (trial ID: NCT03311412) against human programmed cell death protein 1 (PD1) with promising inhibitory binding to the target protein (198).
Marketing of the IgY antibodies for immunotherapy
The vast number of research studies in the past few years has led to an increase in the number of registered products for therapy and diagnosis together with an increase in the number of patents filed and clinical trials registered. For human use several IgY products are registered in the market such as IgY Max (against 26 human-relevant bacteria), Ig-Guard Helico, GastimunHP, and Ovalgen® HP (Helicobacter pylori) (69). For veterinary use, about 56 products are reported at various stages of evaluation, including products in the market such as Ig-Guard Calf, Ig Lock Calves, Globigen® Dia Stop, and IgY DNT (for calf diarrhea); PG-002 (for cow mastitis); Ig-Guard Swine, Ig Lock Pig (for swine diarrhea), Ig-Guard Puppy, ParvoONE®, Ig Lock Canine, GastroMate®, and Guardizen (for pets, especially in canines); Ig-Guard Duck, Ig-Guard Poultry, BIOAb DHV-IgY (for poultry) (69)
The market value of IgY polyclonal antibodies is estimated to be USD 14.2 million by 2027. However, the total antibody market share is only 0.24% (199). About 95% of IgY antibody productions are polyclonal antibodies with future prospects of IgY fragments and monoclonal IgYs (69). Several of the production companies now available provide custom antibody production services (53, 200). As the concept of using IgY antibodies in therapy is a new approach for alternative treatments, the USFDA has enforced strict regulations for the parenteral administration products (193, 201) which mandates more studies on the safety and efficacy of this route of administration. The documentation in these studies is an essential part of the process as well as the utility of standard processes such as good manufacturing practice (GMP) conditions, and immunization of specific- pathogen-free (SPF) birds (69).
Limitations of the IgY antibodies
The susceptibility of IgY to proteolysis is one of the limitations to the oral use of IgY for passive immunotherapy. Although IgY antibodies are resistant to inactivation by the gastric proteolytic enzymes trypsin and chymotrypsin, it is degraded by pepsin (82). To overcome this obstacle, microencapsulation is found to be an effective approach to protect IgY from gastric inactivation (202).
The lack of standardization in the experimental animals (i.e., specific-pathogen-free birds) for the production and extraction and purification procedures of IgY antibodies is one of the major difficulties facing the progress in product licensing so far as well as the consensus on regulation and approval of IgY-based health products (53). More safety studies are needed to evaluate their safety for use as human and veterinary therapeutics. Research is also needed to develop more industrial scale standardized extraction and purification methods to fit the needs of clinical applications (69).
Conclusion and future prospective
MDR pathogens are a growing threat to human health and welfare. The problem requires more research into innovative, and effective approaches including immunotherapies. Economically one of the main advantages of avian immunoglobulins is their cost-effective production, with the benefit of upscaling in the poultry industry, which may allow low-income countries to easily adopt technological capacities in their health systems. Overall, the use and application of IgY antibodies will emerge as an alternative to antibiotics and will help in the design of novel, safe and effective biologicals for the treatment of various MDR pathogens.
Author contributions
SE-K, Conceptualization, Supervision, Writing – original draft, Writing – review & editing; AA, Conceptualization, Supervision, Writing – original draft, Writing – review & editing. CO, Conceptualization, Supervision, Writing – original draft, Writing – review & editing. MT, Writing, original draft, Writing – review & editing. SE Writing, original draft, Writing – review & editing. AZ, Validation, Supervision, Writing – original draft, Writing – review & editing. EA, Conceptualization, Supervision, Writing – original draft, Writing – review & editing. All authors contributed to the article and approved the submitted version.
Funding
The authors extend their appreciation to the Deputyship for Research & Innovation, Ministry of Education in Saudi Arabia for funding this research work through the project number IFPRC-202-141-2020 and King Abdulaziz University, DSR, Jeddah, Saudi Arabia.
Conflict of interest
Author CO was employed by company Oelkrug Enterprises UG.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Sabtu N, Enoch DA, Brown NM. Antibiotic resistance: What, why, where, when and how? Br Med Bull (2015) 116(1):105–13. doi: 10.1093/bmb/ldv041
2. Aslam B, Wang W, Arshad MI, Khurshid M, Muzammil S, Rasool MH, et al. Antibiotic resistance: A rundown of a global crisis. Infection Drug resistance (2018) 11:1645–58. doi: 10.2147/IDR.S173867
3. Merrill K, Hanson SF, Sumner S, Vento T, Veillette J, Webb B. Antimicrobial stewardship: Staff nurse knowledge and attitudes. Am J infection control (2019) 47(10):1219–24. doi: 10.1016/j.ajic.2019.03.022
4. O'Neill J. Tackling drug-resistant infections globally: Final report and recommendations: Review on antimicrobial resistance. (2016), 1–76.
5. Ventola CL. The antibiotic resistance crisis: Part 1: Causes and threats. P t (2015) 40(4):277–83.
6. Munita JM, Bayer AS, Arias CA. Evolving resistance among gram-positive pathogens. Clin Infect Dis (2015) 61 Suppl 2(Suppl 2):S48–57. doi: 10.1093/cid/civ523
7. Shaikh S, Fatima J, Shakil S, Rizvi SM, Kamal MA. Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi J Biol Sci (2015) 22(1):90–101. doi: 10.1016/j.sjbs.2014.08.002
8. Spellberg B, Gilbert DN. The future of antibiotics and resistance: A tribute to a career of leadership by John Bartlett. Clin Infect Dis (2014) 59 Suppl 2(Suppl 2):S71–S5. doi: 10.1093/cid/ciu392
9. WHO. Antimicrobial resistance (2021). Available at: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance.
10. Davies J. Inactivation of antibiotics and the dissemination of resistance genes. Science (1994) 264(5157):375–82. doi: 10.1126/science.8153624
11. Mandal SM, Roy A, Ghosh AK, Hazra TK, Basak A, Franco OL. Challenges and future prospects of antibiotic therapy: From peptides to phages utilization. Front Pharmacol (2014) 5:105. doi: 10.3389/fphar.2014.00105
12. Schmieder R, Edwards R. Insights into antibiotic resistance through metagenomic approaches. Future Microbiol (2012) 7(1):73–89. doi: 10.2217/fmb.11.135
13. Crumplin G, Odell M. Development of resistance to ofloxacin. Drugs (1987) 34(1):1–8. doi: 10.2165/00003495-198700341-00002
14. Martinez J, Baquero F. Mutation frequencies and antibiotic resistance. Antimicrobial Agents chemotherapy (2000) 44(7):1771–7. doi: 10.1128/AAC.44.7.1771-1777.2000
15. Palmer KL, Kos VN, Gilmore MS. Horizontal gene transfer and the genomics of enterococcal antibiotic resistance. Curr Opin Microbiol (2010) 13(5):632–9. doi: 10.1016/j.mib.2010.08.004
16. Hegstad K, Mikalsen T, Coque T, Werner G, Sundsfjord A. Mobile genetic elements and their contribution to the emergence of antimicrobial resistant enterococcus faecalis and enterococcus faecium. Clin Microbiol infection (2010) 16(6):541–54. doi: 10.1111/j.1469-0691.2010.03226.x
17. Laxminarayan R, Duse A, Wattal C, Zaidi AK, Wertheim HF, Sumpradit N, et al. Antibiotic resistance–the need for global solutions. Lancet Infect Dis (2013) 13(12):1057–98. doi: 10.1016/S1473-3099(13)70318-9
18. Akhtar M, Swamy MK, Umar A, Al Sahli AA. Biosynthesis and characterization of silver nanoparticles from methanol leaf extract of cassia didymobotyra and assessment of their antioxidant and antibacterial activities. J Nanoscience Nanotechnology (2015) 15(12):9818–23. doi: 10.1166/jnn.2015.10966
19. Rudramurthy GR, Swamy MK, Sinniah UR, Ghasemzadeh A. Nanoparticles: Alternatives against drug-resistant pathogenic microbes. Molecules (2016) 21(7):836. doi: 10.3390/molecules21070836
20. Hancock RE, Lehrer R. Cationic peptides: A new source of antibiotics. Trends Biotechnol (1998) 16(2):82–8. doi: 10.1016/S0167-7799(97)01156-6
21. Ganz T. Defensins: Antimicrobial peptides of innate immunity. Nat Rev Immunol (2003) 3(9):710–20. doi: 10.1038/nri1180
22. Sulakvelidze A, Alavidze Z, Morris Jg Jr. Bacteriophage therapy antimicrobe agents chemother American Society for Microbiology (ASM) (2001) 45(3):649–59.
23. Sasidharan S, Chen Y, Saravanan D, Sundram K, Latha LY. Extraction, isolation and characterization of bioactive compounds from plants’ extracts. Afr J traditional complementary Altern Medicines (2011) 8(1):5-9. doi: 10.4314/ajtcam.v8i1.60483
24. Ghosh C, Sarkar P, Issa R, Haldar J. Alternatives to conventional antibiotics in the era of antimicrobial resistance. Trends Microbiol (2019) 27(4):323–38. doi: 10.1016/j.tim.2018.12.010
25. Michael A, Meenatchisundaram S, Parameswari G, Subbraj T, Selvakumaran R, Ramalingam S. Chicken egg yolk antibodies (Igy) as an alternative to mammalian antibodies. Indian J Sci Technol (2010) 3(4):468–74. doi: 10.17485/ijst/2010/v3i4.24
27. Seixas AM, Sousa SA, Leitão JH. Antibody-based immunotherapies as a tool for tackling multidrug-resistant bacterial infections. Vaccines (2022) 10(11):1789. doi: 10.3390/vaccines10111789
28. Pereira E, Van Tilburg M, Florean E, Guedes M. Egg yolk antibodies (Igy) and their applications in human and veterinary health: A review. Int Immunopharmacol (2019) 73:293–303. doi: 10.1016/j.intimp.2019.05.015
29. Kovacs-Nolan J, Mine Y. Egg yolk antibodies for passive immunity. Annu Rev Food Sci Technol (2012) 3:163–82. doi: 10.1146/annurev-food-022811-101137
30. Schade R, Hlinak A. Egg yolk antibodies, state of the art and future prospects. ALTEX-Alternatives to Anim experimentation (1996) 13(Supp1):5–9.
31. Akita EM, Nakai S. Immunoglobulins from egg yolk: Isolation and purification. J Food Sci (1992) 57(3):629–34. doi: 10.1111/j.1365-2621.1992.tb08058.x
32. Gilgunn S, Millán Martín S, Wormald MR, Zapatero-Rodríguez J, Conroy PJ, O’Kennedy RJ, et al. Comprehensive n-glycan profiling of avian immunoglobulin y. PloS One (2016) 11(7):e0159859. doi: 10.1371/journal.pone.0159859
33. Liu L. Antibody glycosylation and its impact on the pharmacokinetics and pharmacodynamics of monoclonal antibodies and fc-fusion proteins. J Pharm Sci (2015) 104(6):1866–84. doi: 10.1002/jps.24444
34. Larsson A, Bålöw R-M, Lindahl TL, Forsberg P-O. Chicken antibodies: Taking advantage of evolution–a review. Poultry Sci (1993) 72(10):1807–12. doi: 10.3382/ps.0721807
35. Vega C, Bok M, Saif L, Fernandez F, Parreño V. Egg yolk igy antibodies: A therapeutic intervention against group a rotavirus in calves. Res veterinary Sci (2015) 103:1–10. doi: 10.1016/j.rvsc.2015.09.005
36. Nguyen HH, Tumpey TM, Park H-J, Han G-Y, Lee J, Byun Y-H, et al. Avian antibodies to combat potential H5n1 pandemic and seasonal influenza. INFLUENZA AND OTHER Respir VIRUSES (2011) 5:233–6.
37. Wallach MG, Webby RJ, Islam F, Walkden-Brown S, Emmoth E, Feinstein R, et al. Cross-protection of chicken immunoglobulin y antibodies against H5n1 and H1n1 viruses passively administered in mice. Clin Vaccine Immunol (2011) 18(7):1083–90. doi: 10.1128/CVI.05075-11
38. Thu HM, Myat TW, Win MM, Thant KZ, Rahman S, Umeda K, et al. Chicken egg yolk antibodies (Igy) for prophylaxis and treatment of rotavirus diarrhea in human and animal neonates: A concise review. Food Sci Anim Resour (2017) 37(1):1–9. doi: 10.5851/kosfa.2017.37.1.1
39. Fink AL, Williams KL, Harris E, Alvine TD, Henderson T, Schiltz J, et al. Dengue virus specific igy provides protection following lethal dengue virus challenge and is neutralizing in the absence of inducing antibody dependent enhancement. PloS Negl Trop Dis (2017) 11(7):e0005721. doi: 10.1371/journal.pntd.0005721
40. O’Donnell KL, Meberg B, Schiltz J, Nilles ML, Bradley DS. Zika virus-specific igy results are therapeutic following a lethal zika virus challenge without inducing antibody-dependent enhancement. Viruses (2019) 11(3):301. doi: 10.3390/v11030301
41. Zhang Y, Wei Y, Li Y, Wang X, Liu Y, Tian D, et al. Igy antibodies against Ebola virus possess post-exposure protection and excellent thermostability. bioRxiv (2020). doi: 10.1101/2020.05.21.108159
42. Abbas AT, El-Kafrawy SA, Sohrab SS, Tabll AA, Hassan AM, Iwata-Yoshikawa N, et al. Vaccines (2020) 8(4):634. doi: 10.3390/vaccines8040634
43. El-Kafrawy SA, Abbas AT, Sohrab SS, Tabll AA, Hassan AM, Iwata-Yoshikawa N, et al. Immunotherapeutic efficacy of igy antibodies targeting the full-length spike protein in an animal model of middle East respiratory syndrome coronavirus infection. Pharm (Basel) (2021) 14(6):1–17. doi: 10.3390/ph14060511
44. El-Kafrawy SA, Odle A, Abbas AT, Hassan AM, Abdel-Dayem UA, Qureshi AK, et al. Sars-Cov-2-Specific immunoglobulin y antibodies are protective in infected mice. PloS Pathog (2022) 18(9):e1010782. doi: 10.1371/journal.ppat.1010782
45. Grando TH, Baldissera MD, de Sá MF, do Carmo GM, Porto BCZ, Aguirre GS, et al. Avian antibodies (Igy) against trypanosoma cruzi: Purification and characterization studies. J Immunol Methods (2017) 449:56–61. doi: 10.1016/j.jim.2017.07.002
46. Grando TH, Baldissera MD, Do Carmo G, Oliveira CB, Santi ET, Doleski PH, et al. Ecto-enzymes activities in splenic lymphocytes of mice experimentally infected by trypanosoma cruzi and treated with specific avian immunoglobulins: An attempt to improve the immune response. Mol Cell Biochem (2018) 448(1):9–15. doi: 10.1007/s11010-018-3308-x
47. Kobayashi C, Yokoyama H, Van Nguyen S, Kodama Y, Kimata T, Izeki M. Effect of egg yolk antibody on experimental cryptosporidium parvum infection in scid mice. Vaccine (2004) 23(2):232–5. doi: 10.1016/j.vaccine.2004.05.034
48. Wallach M, Smith NC, Petracca M, Miller CM, Eckert J, Braun R. Eimeria maxima gametocyte antigens: Potential use in a subunit maternal vaccine against coccidiosis in chickens. Vaccine (1995) 13(4):347–54. doi: 10.1016/0264-410X(95)98255-9
49. Ibrahim E-SM, Rahman AS, Isoda R, Umeda K, Van Sa N, Kodama Y. In vitro and in vivo effectiveness of egg yolk antibody against candida albicans (Anti-Ca igy). Vaccine (2008) 26(17):2073–80. doi: 10.1016/j.vaccine.2008.02.046
50. Group WPPLW. Discovery, research, and development of new antibiotics: The who priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis (2018) 18(3):318–27. doi: 10.1016/S1473-3099(17)30753-3
51. Carlander D, Stålberg J, Larsson A. Chicken antibodies: A clinical chemistry perspective. Upsala J Med Sci (1999) 104(3):179–89. doi: 10.3109/03009739909178961
52. Hakalehto E. Chicken igy antibodies provide mucosal barrier against sars-Cov-2 virus and other pathogens. Israel Med Assoc Journal: IMAJ (2021) 23(4):208–11.
53. Lee L, Samardzic K, Wallach M, Frumkin LR, Mochly-Rosen D. Immunoglobulin y for potential diagnostic and therapeutic applications in infectious diseases. Front Immunol (2021) 12:696003. doi: 10.3389/fimmu.2021.696003
54. Nilsson E, Kollberg H, Johannesson M, Wejåker P-E, Carlander D, Larsson A. More than 10 years' continuous oral treatment with specific immunoglobulin y for the prevention of pseudomonas aeruginosa infections: A case report. J medicinal Food (2007) 10(2):375–8. doi: 10.1089/jmf.2006.214
55. Begley TH. Methods and approaches used by fda to evaluate the safety of food packaging materials. Food Additives Contaminants (1997) 14(6-7):545–53. doi: 10.1080/02652039709374566
56. Bachtiar EW, Afdhal A, Meidyawati R, Soejoedono RD, Poerwaningsih E. Effect of topical anti-streptococcus mutans igy gel on quantity of s. mutans on rats’ tooth surface. Acta Microbiologica Immunologica Hungarica (2016) 63(2):159–69. doi: 10.1556/030.63.2016.2.2
57. Krüger C, Pearson S, Kodama Y, Smith AV, Bowen W, Hammarström L. The effects of egg-derived antibodies to glucosyltransferases on dental caries in rats. Caries Res (2004) 38(1):9–14. doi: 10.1159/000073914
58. Xi-ping F, Dan-ying T, Jian-fen C. Clinical study on the effect of anti-gingivitis igy toothpaste in control of gingivitis and dental plaque. Shanghai J Stomatology (2016) 25(4):453.
59. Zhang S, Xing P, Guo G, Liu H, Lin D, Dong C, et al. Development of microbeads of chicken yolk antibodies against clostridium difficile toxin a for colonic-specific delivery. Drug Delivery (2016) 23(6):1940–7. doi: 10.3109/10717544.2015.1022836
60. Zhou Z, Zhou R, Tang Z. Effects of topical application of immunoglobulin yolk on mutans streptococci in dental plaque. Hua xi kou qiang yi xue za zhi= Huaxi kouqiang yixue zazhi= West China J Stomatology (2003) 21(4):295–7.
61. Keller L-A, Merkel O, Popp A. Intranasal drug delivery: Opportunities and toxicologic challenges during drug development. Drug Delivery Trans Res (2022) 12(4):735–57. doi: 10.1007/s13346-020-00891-5
62. Hemmingsson P, Hammarström L. Nasal administration of immunoglobulin as effective prophylaxis against infections in elite cross-country skiers. Scandinavian J Infect Dis (1993) 25(6):783–5. doi: 10.3109/00365549309008580
63. Heikkinen T, Ruohola A, Ruuskanen O, Waris M, Uhari M, Hammarström L. Intranasally administered immunoglobulin for the prevention of rhinitis in children. Pediatr Infect Dis J (1998) 17(5):367–72. doi: 10.1097/00006454-199805000-00004
64. Thomsen K, Christophersen L, Lerche CJ, Holmgaard DB, Calum H, Høiby N, et al. Azithromycin potentiates avian igy effect against pseudomonas aeruginosa in a murine pulmonary infection model. Int J antimicrobial Agents (2021) 57(1):106213. doi: 10.1016/j.ijantimicag.2020.106213
65. Fan W, Sun S, Zhang N, Zhang Y, Jiao P, Wang J, et al. Nasal delivery of thermostable and broadly neutralizing antibodies protects mice against sars-Cov-2 infection. Signal transduction targeted Ther (2022) 7(1):1–4. doi: 10.1038/s41392-022-00911-5
66. da Rocha DG, Fernandez JH, de Almeida CMC, da Silva CL, Magnoli FC, da Silva OÉ, et al. Development of igy antibodies against anti-snake toxins endowed with highly lethal neutralizing activity. Eur J Pharm Sci (2017) 106:404–12. doi: 10.1016/j.ejps.2017.05.069
67. Sifi A, Adi-Bessalem S, Laraba-Djebari F. Development of a new approach of immunotherapy against scorpion envenoming: Avian igys an alternative to equine iggs. Int Immunopharmacol (2018) 61:256–65. doi: 10.1016/j.intimp.2018.06.013
68. Leiva CL, Geoghegan P, Lammer M, Cangelosi A, Mariconda V, Celi AB, et al. In vivo neutralization of bee venom lethality by igy antibodies. Mol Immunol (2021) 135:183–90. doi: 10.1016/j.molimm.2021.04.015
69. Yakhkeshi S, Wu R, Chelliappan B, Zhang X. Trends in industrialization and commercialization of igy technology. Front Immunol (2022) 13. doi: 10.3389/fimmu.2022.991931
70. Casadevall A, Scharff MD. Serum therapy revisited: Animal models of infection and development of passive antibody therapy. Antimicrobial Agents chemotherapy (1994) 38(8):1695–702. doi: 10.1128/AAC.38.8.1695
71. Schade R, Hlinak A. Egg yolk antibodies, state of the art and future prospects. Altex (1996) 13(5):5–9.
72. Gruber FJ, Joeckel R. Allgemeine grundlagen. In: Gruber FJ, Joeckel R, editors. Formelsammlung für das vermessungswesen. Wiesbaden: Vieweg+Teubner Verlag (2004). p. 1–6.
73. Schade R, Calzado EG, Sarmiento R, Chacana PA, Porankiewicz-Asplund J, Terzolo HR. Chicken egg yolk antibodies (Igy-technology): A review of progress in production and use in research and human and veterinary medicine. Altern Lab Anim (2005) 33(2):129–54. doi: 10.1177/026119290503300208
74. Nilsson E, Larsson A. Chicken anti-protein l for the detection of small amounts of protein l in the presence of igg. Hybridoma (2005) 24(2):112–4. doi: 10.1089/hyb.2005.24.112
75. Carlander D, Larsson A. Avian antibodies can eliminate interference due to complement activation in Elisa. Upsala J Med Sci (2009) 106(3):189–95. doi: 10.3109/2000-1967-145
76. Sesarman A, Mihai S, Chiriac MT, Olaru F, Sitaru AG, Thurman JM, et al. Binding of avian igy to type vii collagen does not activate complement and leucocytes and fails to induce subepidermal blistering in mice. Br J Dermatol (2008) 158(3):463–71. doi: 10.1111/j.1365-2133.2007.08388.x
77. Yokoyama H, Peralta R, Sendo S, Ikemori Y, Kodama Y. Detection of passage and absorption of chicken egg yolk immunoglobulins in the gastrointestinal tract of pigs by use of enzyme-linked immunosorbent assay and fluorescent antibody testing. Am J veterinary Res (1993) 54(6):867–72.
78. Akita E, Nakai S. Production and purification of fab′ fragments from chicken egg yolk immunoglobulin y (Igy). J Immunol Methods (1993) 162(2):155–64. doi: 10.1016/0022-1759(93)90380-P
79. Kubickova B, Majerova B, Hadrabova J, Noskova L, Stiborova M, Hodek P. Effect of chicken antibodies on inflammation in human lung epithelial cell lines. Neuro Endocrinol Lett (2014) 35(Suppl 2):99–104.
80. Carlander D, Kollberg H, Wejåker P-E, Larsson A. Peroral immunotheraphy with yolk antibodies for the prevention and treatment of enteric infections. Immunologic Res (2000) 21(1):1–6. doi: 10.1385/IR:21:1:1
81. Xu Y, Li X, Jin L, Zhen Y, Lu Y, Li S, et al. Application of chicken egg yolk immunoglobulins in the control of terrestrial and aquatic animal diseases: A review. Biotechnol Adv (2011) 29(6):860–8. doi: 10.1016/j.biotechadv.2011.07.003
82. Hatta H, Tsuda K, Akachi S, Kim M, Yamamoto T, Ebina T. Oral passive immunization effect of anti-human rotavirus igy and its behavior against proteolytic enzymes. Bioscience biotechnology Biochem (1993) 57(7):1077–81. doi: 10.1271/bbb.57.1077
83. Davalos-Pantoja L, Ortega-Vinuesa J, Bastos-Gonzalez D, Hidalgo-Alvarez R. A comparative study between the adsorption of igy and igg on latex particles. J Biomaterials Science Polymer Edition (2000) 11(6):657–73. doi: 10.1163/156856200743931
84. Cray C, Villar D. Cross-reactivity of anti-chicken igy antibody with immunoglobulins of exotic avian species. Veterinary Clin Pathol (2008) 37(3):328–31. doi: 10.1111/j.1939-165X.2008.00055.x
85. Larsson A, Sjöquist J. Chicken igy: Utilizing the evolutionary difference. Comp immunology Microbiol Infect Dis (1990) 13(4):199–201. doi: 10.1016/0147-9571(90)90088-B
86. Rahman S, Van Nguyen S, Icatlo FC Jr., Umeda K, Kodama Y. Oral passive igy-based immunotherapeutics: A novel solution for prevention and treatment of alimentary tract diseases. Hum Vaccines immunotherapeutics (2013) 9(5):1039–48. doi: 10.4161/hv.23383
88. Casadevall A, Dadachova E, Pirofski L-a. Passive antibody therapy for infectious diseases. Nat Rev Microbiol (2004) 2(9):695–703. doi: 10.1038/nrmicro974
89. Malekshahi ZV, Gargari SLM, Rasooli I, Ebrahimizadeh W. Treatment of helicobacter pylori infection in mice with oral administration of egg yolk-driven anti-urec immunoglobulin. Microbial pathogenesis (2011) 51(5):366–72. doi: 10.1016/j.micpath.2011.06.002
90. Jin L, Baidoo SK, Marquardt RR, Frohlich AA. In vitro inhibition of adhesion of enterotoxigenic escherichia coli K88 to piglet intestinal mucus by egg-yolk antibodies. FEMS Immunol Med Microbiol (1998) 21(4):313–21. doi: 10.1111/j.1574-695X.1998.tb01179.x
91. Schade R, Calzado EG, Sarmiento R, Chacana PA, Porankiewicz-Asplund J, Terzolo HR. Chicken egg yolk antibodies (Igy-technology): A review of progress in production and use in research and human and veterinary medicine. Alternatives to Lab Anim (2005) 33(2):129–54. doi: 10.1177/026119290503300208
92. Jonsson AK, Larsson A, Tängdén T, Melhus Å, Lannergård A. A trial with igy chicken antibodies to eradicate faecal carriage of klebsiella pneumoniae and escherichia coli producing extended-spectrum beta-lactamases. Infection Ecol Epidemiol (2015) 5:28224. doi: 10.3402/iee.v5.28224
93. Shin J-H, Yang M, Nam SW, Kim JT, Myung NH, Bang W-G, et al. Use of egg yolk-derived immunoglobulin as an alternative to antibiotic treatment for control of helicobacter pylori infection. Clin Vaccine Immunol (2002) 9(5):1061–6. doi: 10.1128/CDLI.9.5.1061-1066.2002
94. Hatta H, Horimoto Y. Applications of egg yolk antibody (Igy) in diagnosis reagents and in prevention of diseases. Eggs as Funct Foods Nutraceuticals Hum Health (2019), 305–28. doi: 10.1039/9781788013833-00305
95. LeClaire RD, Hunt RE, Bavari S. Protection against bacterial superantigen staphylococcal enterotoxin b by passive vaccination. Infection Immun (2002) 70(5):2278–81. doi: 10.1128/IAI.70.5.2278-2281.2002
96. Trott D, Yang M, Gonzalez J, Larson A, Tepp W, Johnson E, et al. Egg yolk antibodies for detection and neutralization of clostridium botulinum type a neurotoxin. J Food Prot (2009) 72(5):1005–11. doi: 10.4315/0362-028X-72.5.1005
97. Hirai K, Arimitsu H, Umeda K, Yokota K, Shen L, Ayada K, et al. Passive oral immunization by egg yolk immunoglobulin (Igy) to vibrio cholerae effectively prevents cholera. Acta Med Okayama (2010) 64(3):163–70. doi: 10.18926/AMO/40008
98. Nilsson E, Larsson A, Olesen HV, Wejåker PE, Kollberg H. Good effect of igy against pseudomonas aeruginosa infections in cystic fibrosis patients. Pediatr pulmonology (2008) 43(9):892–9. doi: 10.1002/ppul.20875
99. Dapunt U, Prior B, Oelkrug C, Kretzer JP. Igy targeting bacterial quorum-sensing molecules in implant-associated infections. Molecules (2020) 25(17):4027. doi: 10.3390/molecules25174027
100. Hermans D, Van Steendam K, Verbrugghe E, Verlinden M, Martel A, Seliwiorstow T, et al. Passive immunization to reduce campylobacter jejuni colonization and transmission in broiler chickens. Veterinary Res (2014) 45(1):27. doi: 10.1186/1297-9716-45-27
101. Dapunt U, Prior B, Oelkrug C, Kretzer JP. Igy targeting bacterial quorum-sensing molecules in implant-associated infections. Molecules (2020) 25(17):1–12. doi: 10.3390/molecules25174027
102. Bauer B, Meyer TF. The human gastric pathogen helicobacter pylori and its association with gastric cancer and ulcer disease. Ulcers (2011) 2011:1-23. doi: 10.1155/2011/340157
103. Blaser MJ, Atherton JC. Helicobacter pylori persistence: Biology and disease. J Clin Invest (2004) 113(3):321–33. doi: 10.1172/JCI20925
104. Vakil N, Megraud F. Eradication therapy for helicobacter pylori. Gastroenterology (2007) 133(3):985–1001. doi: 10.1053/j.gastro.2007.07.008
105. Fukase K, Kato M, Kikuchi S, Inoue K, Uemura N, Okamoto S, et al. Effect of eradication of helicobacter pylori on incidence of metachronous gastric carcinoma after endoscopic resection of early gastric cancer: An open-label, randomised controlled trial. Lancet (2008) 372(9636):392–7. doi: 10.1016/S0140-6736(08)61159-9
106. Thung I, Aramin H, Vavinskaya V, Gupta S, Park J, Crowe S, et al. The global emergence of helicobacter pylori antibiotic resistance. Alimentary Pharmacol Ther (2016) 43(4):514–33. doi: 10.1111/apt.13497
107. Liu WZ, Xie Y, Lu H, Cheng H, Zeng ZR, Zhou LY, et al. Fifth Chinese national consensus report on the management of helicobacter pylori infection. Helicobacter (2018) 23(2):e12475. doi: 10.1111/hel.12475
108. De Francesco V, Zullo A, Fiorini G, Saracino IM, Pavoni M, Vaira D. Role of mic levels of resistance to clarithromycin and metronidazole in helicobacter pylori eradication. J Antimicrobial Chemotherapy (2019) 74(3):772–4. doi: 10.1093/jac/dky469
109. Zhang L, Xiao Y, Ji L, Lin M, Zou Y, Zhao J, et al. Potential therapeutic effects of egg yolk antibody (Igy) in helicobacter pylori infections─ a review. J Agric Food Chem (2021) 69(46)):13691–9. doi: 10.1021/acs.jafc.1c05398
110. Yang Y-H, Park D, Yang G, Lee SH, Bae DK, Kyung J, et al. Anti-helicobacter pylori effects of igy from egg York of immunized hens. Lab Anim Res (2012) 28(1):55–60. doi: 10.5625/lar.2012.28.1.55
111. Wang B, Yang J, Cao S, Wang H, Pan X, Zhu J, et al. Preparation of specific anti-helicobacter pylori yolk antibodies and their antibacterial effects. Int J Clin Exp Pathol (2014) 7(10):6430.
112. Shin J-H, Nam S-W, Kim J-T, Yoon J-B, Bang W-G, Roe I-H. Identification of immunodominant helicobacter pylori proteins with reactivity to h. pylori-specific egg-yolk immunoglobulin. J Med Microbiol (2003) 52(3):217–22. doi: 10.1099/jmm.0.04978-0
113. Shin J-H, Roe I-H, Kim H-G. Production of anti-helicobacter pylori urease-specific immunoglobulin in egg yolk using an antigenic epitope of h. pylori urease. J Med Microbiol (2004) 53(1):31–4. doi: 10.1099/jmm.0.05327-0
114. Nomura S, Suzuki H, Masaoka T, Kurabayashi K, Ishii H, Kitajima M, et al. Effect of dietary anti-urease immunoglobulin y on helicobacter pylori infection in Mongolian gerbils. Helicobacter (2005) 10(1):43–52. doi: 10.1111/j.1523-5378.2005.00290.x
115. Aiba Y, Umeda K, Rahman S, Nguyen SV, Komatsu Y. Synergistic effect of anti-helicobacter pylori urease immunoglobulin y from egg yolk of immunized hens and lactobacillus johnsonii no. 1088 to inhibit the growth of helicobacter pylori in vitro and in vivo. Vaccine (2019) 37(23):3106–12. doi: 10.1016/j.vaccine.2019.04.045
116. Hong KS, Ki M-R, Ullah HA, Lee E-J, Kim YD, Chung M-J, et al. Preventive effect of anti-vaca egg yolk immunoglobulin (Igy) on helicobacter pylori-infected mice. Vaccine (2018) 36(3):371–80. doi: 10.1016/j.vaccine.2017.11.082
117. Borhani K, Mobarez AM, Khabiri AR, Behmanesh M, Khoramabadi N. Inhibitory effects of rhp-nap igy against helicobacter pylori attachment to ags cell line. Microbial pathogenesis (2016) 97:231–5. doi: 10.1016/j.micpath.2016.06.004
118. Borhani K, Mobarez AM, Khabiri AR, Behmanesh M, Khoramabadi N. Production of specific igy helicobacter pylori recombinant oipa protein and assessment of its inhibitory effects towards attachment of h. pylori to ags cell line. Clin Exp Vaccine Res (2015) 4(2):177–83. doi: 10.7774/cevr.2015.4.2.177
119. Malfertheiner P, Selgrad M, Wex T, Romi B, Borgogni E, Spensieri F, et al. Efficacy, immunogenicity, and safety of a parenteral vaccine against helicobacter pylori in healthy volunteers challenged with a cag-positive strain: A randomised, placebo-controlled phase 1/2 study. Lancet Gastroenterol Hepatol (2018) 3(10):698–707. doi: 10.1016/S2468-1253(18)30125-0
120. Miyashita M, Joh T, Watanabe K, Todoroki I, Seno K, Ohara H, et al. Immune responses in mice to intranasal and intracutaneous administration of a DNA vaccine encoding helicobacter pylori-catalase. Vaccine (2002) 20(17-18):2336–42. doi: 10.1016/S0264-410X(02)00104-4
121. Attallah AM, Abbas AT, Ismail H, Abdel-Raouf M, El-Dosoky I. Efficacy of passive immunization with igy antibodies to a 58-kda h. pylori antigen on severe gastritis in Balb/C mouse model. J Immunoassay Immunochemistry (2009) 30(4):359–77. doi: 10.1080/15321810903187922
122. Suzuki H, Nomura S, Masaoka T, Goshima H, Kamata N, Kodama Y, et al. Effect of dietary anti-helicobacter pylori-urease immunoglobulin y on helicobacter pylori infection. Alimentary Pharmacol Ther (2004) 20:185–92. doi: 10.1111/j.1365-2036.2004.02027.x
123. Guo R, Wu S, Guan L, Xie Y, Yang X, Li Z, et al. Dietary multivalent anti-helicobacter pylori immunoglobulin y significantly increase the h. pylori eradication and improve the clinical symptoms in patients. Helicobacter (2021) 26(5):e12843. doi: 10.1111/hel.12843
124. Horie K, Horie N, Abdou A, Yang J-O, Yun S-S, Chun H-N, et al. Suppressive effect of functional drinking yogurt containing specific egg yolk immunoglobulin on helicobacter pylori in humans. J dairy Sci (2004) 87(12):4073–9. doi: 10.3168/jds.S0022-0302(04)73549-3
125. Zhang L, Xiao Y, Ji L, Lin M, Zou Y, Zhao J, et al. Potential therapeutic effects of egg yolk antibody (Igy) in helicobacter pylori infections─ A review. J Agric Food Chem (2021) 69(46):13691–9. doi: 10.1021/acs.jafc.1c05398
126. Sainz-Mejías M, Jurado-Martín I, McClean S. Understanding pseudomonas aeruginosa–host interactions: The ongoing quest for an efficacious vaccine. Cells (2020) 9(12):2617. doi: 10.3390/cells9122617
127. Hirsch EB, Tam VH. Impact of multidrug-resistant pseudomonas aeruginosa infection on patient outcomes. Expert Rev pharmacoeconomics outcomes Res (2010) 10(4):441–51. doi: 10.1586/erp.10.49
128. Nilsson E, Larsson A, Olesen HV, Wejaker PE, Kollberg H. Good effect of igy against pseudomonas aeruginosa infections in cystic fibrosis patients. Pediatr pulmonology (2008) 43(9):892–9. doi: 10.1002/ppul.20875
129. Thomsen K, Christophersen L, Bjarnsholt T, Jensen PO, Moser C, Hoiby N. Anti-pseudomonas aeruginosa igy antibodies induce specific bacterial aggregation and internalization in human polymorphonuclear neutrophils. Infection Immun (2015) 83(7):2686–93. doi: 10.1128/iai.02970-14
130. Thomsen K, Christophersen L, Bjarnsholt T, Jensen PO, Moser C, Hoiby N. Anti-pseudomonas aeruginosa igy antibodies augment bacterial clearance in a murine pneumonia model. J cystic fibrosis (2016) 15(2):171–8. doi: 10.1016/j.jcf.2015.08.002
131. Schreiner B, Viertlboeck BC, Göbel TW. A striking example of convergent evolution observed for the ggfcr: Igy interaction closely resembling that of mammalian fcr: Igg. Dev Comp Immunol (2012) 36(3):566–71. doi: 10.1016/j.dci.2011.09.013
132. Thomsen K, Christophersen L, Jensen PØ, Bjarnsholt T, Moser C, Høiby N. Anti-pseudomonas aeruginosa igy antibodies promote bacterial opsonization and augment the phagocytic activity of polymorphonuclear neutrophils. Hum Vaccines immunotherapeutics (2016) 12(7):1690–9. doi: 10.1080/21645515.2016.1145848
133. Ahmadi TS, Gargari SLM, Talei D. Anti-flagellin igy antibodies protect against pseudomonas aeruginosa infection in both acute pneumonia and burn wound murine models in a non-Type-Specific mode. Mol Immunol (2021) 136:118–27. doi: 10.1016/j.molimm.2021.06.002
134. Sanches RF, Dos Santos Ferraro ACN, Marroni FEC, Venancio EJ. Synergistic activity between beta-lactams and igy antibodies against pseudomonas aeruginosa in vitro. Mol Immunol (2022) 148:1–5. doi: 10.1016/j.molimm.2022.05.010
135. Schwartz FA, Christophersen L, Thomsen K, Baekdal S, Pals Bendixen M, Jørgensen M, et al. Chicken igy reduces the risk of pseudomonas aeruginosa urinary tract infections in a murine model. Front Microbiol (2022), 3582. doi: 10.3389/fmicb.2022.988386
136. Otterbeck A, Hanslin K, Lantz EL, Larsson A, Stålberg J, Lipcsey M. Inhalation of specific anti-pseudomonas aeruginosa igy antibodies transiently decreases p. aeruginosa colonization of the airway in mechanically ventilated piglets. Intensive Care Med Exp (2019) 7(1):1–10. doi: 10.1186/s40635-019-0246-1
137. Leiva CL, Gallardo MJ, Casanova N, Terzolo H, Chacana P. Igy-technology (Egg yolk antibodies) in human medicine: A review of patents and clinical trials. Int Immunopharmacol (2020) 81:106269. doi: 10.1016/j.intimp.2020.106269
138. Schuller Y, Biegstraaten M, Hollak CE, Klümpen H-J, Gispen-de Wied CC, Stoyanova-Beninska V. Oncologic orphan drugs approved in the eu–do clinical trial data correspond with real-world effectiveness? Orphanet J rare Dis (2018) 13(1):1–11. doi: 10.1186/s13023-018-0900-9
139. Kollberg H, Carlander D, Olesen H, Wejåker PE, Johannesson M, Larsson A. Oral administration of specific yolk antibodies (Igy) may prevent pseudomonas aeruginosa infections in patients with cystic fibrosis: A phase I feasibility study. Pediatr pulmonology (2003) 35(6):433–40. doi: 10.1002/ppul.10290
140. Carlander D, Kollberg H, Larsson A. Retention of specific yolk igy in the human: Oral cavity. BioDrugs (2002) 16(6):433–7. doi: 10.2165/00063030-200216060-00004
142. Broom LJ, Kogut MH. Deciphering desirable immune responses from disease models with resistant and susceptible chickens. Poultry Sci (2019) 98(4):1634–42. doi: 10.3382/ps/pey535
143. Mine Y. Separation of salmonella enteritidis from experimentally contaminated liquid eggs using a hen igy immobilized immunomagnetic separation system. J Agric Food Chem (1997) 45(10):3723–7. doi: 10.1021/jf9701998
144. Terzolo H, Sandoval V, Caffer M, Terragno R, Alcain A. Agglutination of hen egg-yolk immunoglobulins (Igy) against salmonella enterica, serovar enteritidis. Rev Argent Microbiologia (1998) 30(2):84–92.
145. Lee E, Sunwoo H, Menninen K, Sim J. In vitro studies of chicken egg yolk antibody (Igy) against salmonella enteritidis and salmonella typhimurium. Poultry Sci (2002) 81(5):632–41. doi: 10.1093/ps/81.5.632
146. Chelliappan B, Michael A, Vega CG, Zhong F, Zhang X, Morgan PM. Applications of igy in veterinary medicine. In: Zhang X-Y, Vieira-Pires RS, Morgan PM, Schade R, editors. Igy-technology: Production and application of egg yolk antibodies: Basic knowledge for a successful practice. Cham: Springer International Publishing (2021). p. 205–35.
147. Isfahani NH, Rahimi S, Rasaee MJ, Torshizi MAK, Salehi TZ, Grimes JL. The effect of capsulated and noncapsulated Egg-Yolk–specific antibody to reduce colonization in the intestine of salmonella enterica ssp. enterica serovar infantis–challenged broiler chickens. Poultry Sci (2020) 99(3):1387–94. doi: 10.1016/j.psj.2019.11.019
148. Karabasanavar N, Chakkodabail Benakabhat M, Agalagandi Gopalakrishna S, Bagalkote PS, Basayya Hiremath J, Shivanagowda Patil G, et al. Polyclonal hen egg yolk antibodies could confer passive protection against salmonella serotypes in broiler chicks. J Food Saf (2022) 42(4):e12987. doi: 10.1111/jfs.12987
149. Park H-S, Park K-I, Nagappan A, Lee D-H, Kang S-R, Kim J-A, et al. Proteomic analysis of effects on natural herb additive containing immunoglobulin yolksac (Igy) in pigs. Am J Chin Med (2011) 39(03):477–88. doi: 10.1142/S0192415X1100897X
150. Kariyawasam S, Wilkie B, Gyles C. Resistance of broiler chickens to escherichia coli respiratory tract infection induced by passively transferred egg-yolk antibodies. Veterinary Microbiol (2004) 98(3-4):273–84. doi: 10.1016/j.vetmic.2003.10.022
151. Wang Z, Li J, Li J, Li Y, Wang L, Wang Q, et al. Protective effect of chicken egg yolk immunoglobulins (Igy) against enterotoxigenic escherichia coli K88 adhesion in weaned piglets. BMC veterinary Res (2019) 15(1):1–12. doi: 10.1186/s12917-019-1958-x
152. Gaensbauer JT, Melgar MA, Calvimontes DM, Lamb MM, Asturias EJ, Contreras-Roldan IL, et al. Efficacy of a bovine colostrum and egg-based intervention in acute childhood diarrhoea in Guatemala: A randomised, double-blind, placebo-controlled trial. BMJ Global Health (2017) 2(4):e000452. doi: 10.1136/bmjgh-2017-000452
153. Hermans D, Van Steendam K, Verbrugghe E, Verlinden M, Martel A, Seliwiorstow T, et al. Passive immunization to reduce campylobacter jejuni colonization and transmission in broiler chickens. Veterinary Res (2014) 45(1):1–12. doi: 10.1186/1297-9716-45-27
154. Vandeputte J, Martel A, Canessa S, Van Rysselberghe N, De Zutter L, Heyndrickx M, et al. Reducing campylobacter jejuni colonization in broiler chickens by in-feed supplementation with hyperimmune egg yolk antibodies. Sci Rep (2019) 9(1):1–10. doi: 10.1038/s41598-019-45380-z
155. Zhen Y-H, Jin L-J, Li X-Y, Guo J, Li Z, Zhang B-J, et al. Efficacy of specific egg yolk immunoglobulin (Igy) to bovine mastitis caused by staphylococcus aureus. Veterinary Microbiol (2009) 133(4):317–22. doi: 10.1016/j.vetmic.2008.07.016
156. Vega C, Bok M, Chacana P, Saif L, Fernandez F, Parreño V. Egg yolk igy: Protection against rotavirus induced diarrhea and modulatory effect on the systemic and mucosal antibody responses in newborn calves. Veterinary Immunol immunopathology (2011) 142(3-4):156–69. doi: 10.1016/j.vetimm.2011.05.003
157. Rahimi S, Shiraz ZM, Salehi TZ, Torshizi MK, Grimes JL. Prevention of salmonella infection in poultry by specific egg-derived antibody. Int J Poult Sci (2007) 6(4):230–5. doi: 10.3923/ijps.2007.230.235
158. Chalghoumi R, Thewis A, Beckers Y, Marcq C, Portetelle D, Schneider Y-J. Adhesion and growth inhibitory effect of chicken egg yolk antibody (Igy) on salmonella enterica serovars enteritidis and typhimurium in vitro. Foodborne Pathog Dis (2009) 6(5):593–604. doi: 10.1089/fpd.2008.0258
159. Li X, Yao Y, Wang X, Zhen Y, Thacker PA, Wang L, et al. Chicken egg yolk antibodies (Igy) modulate the intestinal mucosal immune response in a mouse model of salmonella typhimurium infection. Int Immunopharmacol (2016) 36:305–14. doi: 10.1016/j.intimp.2016.04.036
160. Kaper J, Nataro J, Mobley H. Nature reviews. microbiology. Nat Rev Microbiol (2004) 2(2):123–40. doi: 10.1038/nrmicro818
161. Diraviyam T, Zhao B, Wang Y, Schade R, Michael A, Zhang X. Effect of chicken egg yolk antibodies (Igy) against diarrhea in domesticated animals: A systematic review and meta-analysis. PloS One (2014) 9(5):e97716. doi: 10.1371/journal.pone.0097716
162. Aizenshtein E, Yosipovich R, Kvint M, Shadmon R, Krispel S, Shuster E, et al. Practical aspects in the use of passive immunization as an alternative to attenuated viral vaccines. Vaccine (2016) 34(22):2513–8. doi: 10.1016/j.vaccine.2016.03.051
163. Brumfield K, Seo H, Idegwu N, Artman C, Gonyar L, Nataro J, et al. Feasibility of avian antibodies as prophylaxis against enterotoxigenic escherichia coli colonization. Front Immunol (2022) 13. doi: 10.3389/fimmu.2022.1011200
164. Zhen Y-H, Jin L-J, Guo J, Li X-Y, Lu Y-N, Chen J, et al. Characterization of specific egg yolk immunoglobulin (Igy) against mastitis-causing escherichia coli. Veterinary Microbiol (2008) 130(1-2):126–33. doi: 10.1016/j.vetmic.2007.12.014
165. Michael A, Subbraj T, Diraviam T, Shanmugam V. Isolation, purification and neutralizing potential of chicken egg yolk immunoglobulin (Igy) against mastitis causing escherichia coli in dairy cows in coimbatore district. Int J Drug Dev Res (2011) 3(2):147–53.
166. Endtz HP. Campylobacter infections. In: Hunter's tropical medicine and emerging infectious diseases. Elsevier (2020). p. 507–11.
167. Li S, Li H, Qi T, Yan X, Wang B, Guan J, et al. Comparative transcriptomics analyses of the different growth states of multidrug-resistant acinetobacter baumannii. Biomedicine Pharmacotherapy (2017) 85:564–74. doi: 10.1016/j.biopha.2016.11.065
168. Endimiani A, Hujer KM, Hujer AM, Bertschy I, Rossano A, Koch C, et al. Acinetobacter baumannii isolates from pets and horses in Switzerland: Molecular characterization and clinical data. J antimicrobial chemotherapy (2011) 66(10):2248–54. doi: 10.1093/jac/dkr289
169. Luo G, Spellberg B, Gebremariam T, Bolaris M, Lee H, Fu Y, et al. Diabetic murine models for acinetobacter baumannii infection. J antimicrobial chemotherapy (2012) 67(6):1439–45. doi: 10.1093/jac/dks050
170. Shin JA, Chang YS, Kim HJ, Kim SK, Chang J, Ahn CM, et al. Clinical outcomes of tigecycline in the treatment of multidrug-resistant acinetobacter baumannii infection. Yonsei Med J (2012) 53(5):974–84. doi: 10.3349/ymj.2012.53.5.974
171. Munoz-Price LS, Weinstein RA. Acinetobacter infection. New Engl J Med (2008) 358(12):1271–81. doi: 10.1056/NEJMra070741
172. Sopirala MM, Mangino JE, Gebreyes WA, Biller B, Bannerman T, Balada-Llasat J-M, et al. Synergy testing by etest, microdilution checkerboard, and time-kill methods for pan-Drug-Resistant acinetobacter baumannii. Antimicrobial Agents chemotherapy (2010) 54(11):4678–83. doi: 10.1128/AAC.00497-10
173. Tsubokura K, Berndtson E, Bogstedt A, Kaijser B, Kim M, Ozeki M, et al. Oral administration of antibodies as prophylaxis and therapy in campylobacter jejuni-infected chickens. Clin Exp Immunol (1997) 108(3):451–5. doi: 10.1046/j.1365-2249.1997.3901288.x
174. Sugita-Konishi Y, Shibata K, Yun SS, Hara-Kudo Y, Yamaguchi K, Kumagai S. Immune functions of immunoglobulin y isolated from egg yolk of hens immunized with various infectious bacteria. Bioscience biotechnology Biochem (1996) 60(5):886–8. doi: 10.1271/bbb.60.886
175. Sugita-Konishi Y, Ogawa M, Arai S, Kumagai S, Igimi S, Shimizu M. Blockade of salmonella enteritidis passage across the basolateral barriers of human intestinal epithelial cells by specific antibody. Microbiol Immunol (2000) 44(6):473–9. doi: 10.1111/j.1348-0421.2000.tb02522.x
176. Jahangiri A, Owlia P, Rasooli I, Salimian J, Derakhshanifar E, Naghipour Erami A, et al. Specific egg yolk antibodies (Igy) confer protection against acinetobacter baumannii in a murine pneumonia model. J Appl Microbiol (2019) 126(2):624–32. doi: 10.1111/jam.14135
177. Jahangiri A, Owlia P, Rasooli I, Salimian J, Derakhshanifar E, Aghajani Z, et al. Specific egg yolk immunoglobulin as a promising non-antibiotic biotherapeutic product against acinetobacter baumannii pneumonia infection. Sci Rep (2021) 11(1):1–11. doi: 10.1038/s41598-021-81356-8
178. Rao RS, Karthika RU, Singh S, Shashikala P, Kanungo R, Jayachandran S, et al. Correlation between biofilm production and multiple drug resistance in imipenem resistant clinical isolates of acinetobacter baumannii. Indian J Med Microbiol (2008) 26(4):333–7. doi: 10.1016/S0255-0857(21)01809-0
179. Goh HS, Beatson SA, Totsika M, Moriel DG, Phan M-D, Szubert J, et al. Molecular analysis of the acinetobacter baumannii biofilm-associated protein. Appl Environ Microbiol (2013) 79(21):6535–43. doi: 10.1128/AEM.01402-13
180. Thummeepak R, Kongthai P, Leungtongkam U, Sitthisak S. Distribution of virulence genes involved in biofilm formation in multi-drug resistant acinetobacter baumannii clinical isolates. Int Microbiol (2016) 19(2):121–9. doi: 10.2436/20.1501.01.270
181. Ranjbar A, Rasooli I, Jahangiri A, Ramezanalizadeh F. Specific egg yolk antibody raised to biofilm associated protein (Bap) is protective against murine pneumonia caused by acinetobacter baumannii. Sci Rep (2022) 12(1):1–9. doi: 10.1038/s41598-022-16894-w
182. Shi H, Zhu J, Zou B, Shi L, Du L, Long Y, et al. Effects of specific egg yolk immunoglobulin on pan-Drug-Resistant acinetobacter baumannii. Biomedicine Pharmacotherapy (2017) 95:1734–42. doi: 10.1016/j.biopha.2017.09.112
183. Gygli SM, Borrell S, Trauner A, Gagneux S. Antimicrobial resistance in mycobacterium tuberculosis: Mechanistic and evolutionary perspectives. FEMS Microbiol Rev (2017) 41(3):354–73. doi: 10.1093/femsre/fux011
184. Sudjarwo SA, Eraiko K, Sudjarwo GW, Koerniasari E. The potency of chicken egg yolk immunoglobulin (Igy) specific as immunotherapy to mycobacterium tuberculosis infection. J advanced Pharm Technol Res (2017) 8(3):91–6. doi: 10.4103/japtr.JAPTR_167_16
185. Sudjarwo SA, Eraiko K, Sudjarwo GW, Koerniasari. The activity of immunoglobulin y anti-mycobacterium tuberculosis on proliferation and cytokine expression of rat peripheral blood mononuclear cells. Pharmacognosy Res (2017) 9(Suppl 1):S5. doi: 10.4103/pr.pr_66_17
186. Reddy PN, Srirama K, Dirisala VR. An update on clinical burden, diagnostic tools, and therapeutic options of staphylococcus aureus. Infect Diseases: Res Treat (2017) 10:1179916117703999. doi: 10.1177/1179916117703999
187. Nomura Y, Yoshinaga M, Masuda K, Takei S, Miyata K. Maternal antibody against toxic shock syndrome toxin-1 may protect infants younger than 6 months of age from developing Kawasaki syndrome. J Infect Dis (2002) 185(11):1677–80. doi: 10.1086/340513
188. Kota RK, Reddy PN, Sreerama K. Application of igy antibodies against staphylococcal protein a (Spa) of staphylococcus aureus for detection and prophylactic functions. Appl Microbiol Biotechnol (2020) 104(21):9387–98. doi: 10.1007/s00253-020-10912-5
189. Wang L-H, Li X-Y, Jin L-J, You J-S, Zhou Y, Li S-Y, et al. Characterization of chicken egg yolk immunoglobulins (Igys) specific for the most prevalent capsular serotypes of mastitis-causing staphylococcus aureus. Veterinary Microbiol (2011) 149(3-4):415–21. doi: 10.1016/j.vetmic.2010.11.029
190. Tian Z, Zhang X. Progress on research of chicken igy antibody-fcry receptor combination and transfer. J Receptors Signal Transduction (2012) 32(5):231–7. doi: 10.3109/10799893.2012.703207
191. Nishibori N, Horiuchi H, Furusawa S, Matsuda H. Humanization of chicken monoclonal antibody using phage-display system. Mol Immunol (2006) 43(6):634–42. doi: 10.1016/j.molimm.2005.04.002
192. Andris-Widhopf J, Rader C, Steinberger P, Fuller R, Barbas Iii CF. Methods for the generation of chicken monoclonal antibody fragments by phage display. J Immunol Methods (2000) 242(1-2):159–81. doi: 10.1016/S0022-1759(00)00221-0
193. Zhang X, Chen H, Tian Z, Chen S, Schade R. Chicken monoclonal igy antibody: A novel antibody development strategy. Avian Biol Res (2010) 3(3):97–106. doi: 10.3184/175815510X12823014530963
194. Bossy-Wetzel E, Schwarzenbacher R, Lipton SA. Molecular pathways to neurodegeneration. Nat Med (2004) 10(7):S2–9. doi: 10.1038/nm1067
195. Yasui H, Takahashi K, Taki S, Shimizu M, Koike C, Umeda K, et al. Near infrared photo-antimicrobial targeting therapy for candida albicans. Advanced Ther (2021) 4(2):2000221. doi: 10.1002/adtp.202000221
196. Ge S, Xu L, Li B, Zhong F, Liu X, Zhang X. Canine parvovirus is diagnosed and neutralized by chicken igy-scfv generated against the virus capsid protein. Veterinary Res (2020) 51(1):1–11. doi: 10.1186/s13567-020-00832-7
197. Ching KH, Collarini EJ, Abdiche YN, Bedinger D, Pedersen D, Izquierdo S, et al. Chickens with humanized immunoglobulin genes generate antibodies with high affinity and broad epitope coverage to conserved targets. mAbs (2018) 10(1):71–80. doi: 10.1080/19420862.2017.1386825
198. Gjetting T, Gad M, Fröhlich C, Lindsted T, Melander MC, Bhatia VK, et al. Sym021, a promising anti-Pd1 clinical candidate antibody derived from a new chicken antibody discovery platform. mAbs (2019). doi: 10.1080/19420862.2019.1596514
199. Vieira-Pires RS, Ahn Chul H, Bok M, Caulfield CD, Chacana P, Elahi F, Larsson AO, et al. “IgY Industries and Markets.” IgY-Technology: Production and Application of Egg Yolk Antibodies (2021) (Cham: Springer), 279-308. doi: 10.1007/978-3-030-72688-1_17
200. Wu R, Yakhkeshi S, Zhang X. Scientometric analysis and perspective of igy technology study. Poultry Sci (2022) 101(4):101713. doi: 10.1016/j.psj.2022.101713
201. Lee W, Atif AS, Tan SC, Leow CH. Insights into the chicken igy with emphasis on the generation and applications of chicken recombinant monoclonal antibodies. J Immunol Methods (2017) 447:71–85. doi: 10.1016/j.jim.2017.05.001
Keywords: IgY antibodies, Bacteria, AMR, antibiotics, passive immunization
Citation: El-Kafrawy SA, Abbas AT, Oelkrug C, Tahoon M, Ezzat S, Zumla A and Azhar EI (2023) IgY antibodies: The promising potential to overcome antibiotic resistance. Front. Immunol. 14:1065353. doi: 10.3389/fimmu.2023.1065353
Received: 09 October 2022; Accepted: 05 January 2023;
Published: 20 January 2023.
Edited by:
Pablo Chacana, Instituto Nacional de Tecnología Agropecuaria, ArgentinaReviewed by:
Saeed Yakhkeshi, Royan institute for Stem Cell Biology and Technology (RI-SCBT), IranCARLA BUSTOS, National Scientific and Technical Research Council (CONICET), Argentina
Copyright © 2023 El-Kafrawy, Abbas, Oelkrug, Tahoon, Ezzat, Zumla and Azhar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aymn T. Abbas, YXRhYmRhbGhhZGlAa2F1LmVkdS5zYQ==; Esam I. Azhar, ZWF6aGFyQGthdS5lZHUuc2E=
†These authors have contributed equally to this work