 James R. G. Adams
James R. G. Adams Jai Mehat3
Jai Mehat3 Roberto La Ragione
Roberto La Ragione Shahriar Behboudi
Shahriar Behboudi- 1School of Veterinary Medicine, Faculty of Health and Medical Sciences, University of Surrey, Guildford, United Kingdom
- 2Avian Immunology, The Pirbright Institute, Woking, United Kingdom
- 3School of Biosciences, Faculty of Health and Medical Sciences, University of Surrey, Guildford, United Kingdom
The widespread use of antibiotics in the poultry industry has led to the emergence of antibiotic-resistant bacteria, which pose a significant health risk to humans and animals. These public health concerns, which have led to legislation limiting antibiotic use in animals, drive the need to find alternative strategies for controlling and treating bacterial infections. Modulation of the avian innate immune system using immunostimulatory compounds provides a promising solution to enhance poultry immune responses to a broad range of bacterial infections without the risk of generating antibiotic resistance. An array of immunomodulatory compounds have been investigated for their impact on poultry performance and immune responses. However, further research is required to identify compounds capable of controlling bacterial infections without detrimentally affecting bird performance. It is also crucial to determine the safety and effectiveness of these compounds in conjunction with poultry vaccines. This review provides an overview of the various immune modulators known to enhance innate immunity against avian bacterial pathogens in chickens, and describes the mechanisms involved.
1 Introduction
By 2050, the world’s population is predicted to increase to 9.8 billion people, threatening food security and causing shifts in consumer purchasing patterns (1, 2). Animal-derived meat products are central to the current food supply, accounting for 40% of the agricultural production value and 13% of the calories consumed globally. Furthermore, based on consumer patterns meat consumption is predicted to increase by between 62% to 144% by 2050, with consumption of poultry alone predicted to double (2, 3). To accommodate this increased demand, the poultry industry must expand beyond its current size or innovate its current production methods. However, accommodating this increased demand incurs greater costs, difficulty in maintaining production yields, meat quality issues, and animal welfare concerns (4, 5). Infectious diseases such as mycoplasmosis and laryngotracheitis are significant contributors to these factors, with increased production intensity resulting in the emergence of diseases and antimicrobial resistance (AMR). Jones et al. (2019) observed that diseases associated with production, namely respiratory diseases/conditions (ascites and infectious bronchitis), enteric diseases (coccidiosis and clostridiosis), locomotory diseases (tibial dyschondroplasia, foot pad dermatitis, and keel bone damage), colibacillosis affecting reproduction, and injurious feather pecking, can increase bird mortality by up to 336% in broiler flocks and 71.5% in layer flocks (6). These conditions were also observed to reduce productivity, with broiler body weight reduced by as much as 17.7%, and egg numbers and egg weight reduced by 32.9% and 8.7%, respectively (6). The incidence of diseases within poultry flocks can stem from a range of sources, including viruses, parasites, and bacteria, with the latter being the focus of this review.
To ensure productivity and maintain animal welfare, mitigation of the emergence and spread of bacterial diseases is essential. Current management strategies are focused on biosecurity, antibiotics, and vaccines. Biosecurity broadly refers to the measures taken to prevent the introduction or minimize the spread of infectious diseases (7). The elimination of pathogenic bacteria such as Salmonella within flocks can pose a significant challenge because of their ubiquitous nature. However, the application of sanitation technologies, combined with effective surveillance, prevention, and management strategies can result in the reduction or eradication of epidemiologically important human pathogens, such as Salmonella Typhi, in higher income countries (8). In poultry, sanitation strategies typically aim to reduce contamination of feed, water, and hatching eggs, as these are frequently seen as common points of disease transmission (9). These strategies include improving the cleanliness of the hatching area and sanitization of eggs using disinfectants or UV radiation (9, 10). Improvements in feed and water supply by pasteurization and chlorination allow the number of pathogenic strains within the gastrointestinal tract and feces to be reduced in a non-invasive manner, limiting the spread of bacterial diseases such as colibacillosis (11, 12). Improvement in air and litter quality have also been demonstrated to reduce the risk of outbreaks, highlighting the importance of environmental management (13).
Antibiotics have historically been applied on a flock-wide scale to prevent and treat bacterial diseases, as well as to promote bird growth and egg production (14). Tetracyclines, aminoglycosides, and fluoroquinolones are among the antibiotic classes approved for use by regulatory authorities in the UK, Brazil, China, and Europe, and where permitted can be employed prophylactically (14, 15). Glycolipids, macrolides, and glycopeptides are used as growth-promoting antibiotics (16) but have been prohibited for use in agriculture in the EU since 2006, with recent legislation further restricting their use (17, 18). Similar legislative efforts and antibiotic stewardship campaigns have been seen globally, including in the US, Japan, Denmark, China, and India (19, 20). However, the use of agricultural antibiotics remains prevalent in low- and middle-income countries because of their reliance on food and animals as exports, as well as food insecurity (20). The historic, current, and misuse of antibiotics have led to the selection and promotion of antimicrobial resistance (AMR) within bacterial populations. The presence of antimicrobials within an environment following misuse results in selection pressure, eliminating susceptible organisms and allowing those with acquired or intrinsic resistance to survival and multiplication (21). The selection of AMR, particularly within populations associated with infectious diseases, is regarded as one of the most pressing issues facing global public and animal health (14, 22, 23).
In addition to antibiotics and improved sanitation, several types of vaccines are employed to protect against viral, parasitic, and bacterial pathogens, including live attenuated, inactivated/killed, nucleic acid-based, and subunit vaccines (24, 25). Autogenous vaccination, generated by the isolation and inactivation of bacteria from diseased animals for immunization of flocks, has also been frequent use, but its use is now restricted within the EU to emergency cases when licensed vaccines are available, thus their usage may change in the future (26). Vaccination also suffers from a lack of long-term efficiency due to genomic diversity and drift within pathogens, leading to reduced protection (25, 27). Furthermore, issues have also been identified with the administration of vaccines, with some requiring multiple doses and uneven application resulting in reduced efficacy (28). Administration-associated issues can be mitigated through the application of vaccines through spray, orally, or in ovo (29), but the specificity of vaccination still demands the application of multiple vaccines, each targeting an individual pathogen or pathogen group, driving up costs, and increasing the labor required for production (24, 27).
This growing desire for non-antibiotic treatments has led to an increased interest in alternative control strategies. Bacteriophage therapy, the use of viruses to control and treat bacterial pathogens, has seen greater focus, with over a dozen bacteriophage products for use in poultry coming to market, with many receiving “generally recognized as safe” certification by the US Food and Drug Administration (30). Furthermore, a range of non-antibiotic methods have also been employed to manage infectious diseases, such as pre- and probiotics (31), plant-derived phytochemicals (32), and organic or short-chain fatty acids (33), many of which have been observed to directly inhibit the growth of pathogenic bacteria. However, bacterial diseases remain a key issue in poultry production and may cripple poultry production in the future. Therefore, novel management strategies to supplement or replace existing methods are required. Notably, the ability to generate sustained heterologous protection through minimal application, while avoiding AMR selection is strongly desired. A proposed alternative to the use of antibiotics and traditional vaccination is to target the innate immune system using immunostimulatory compounds.
2 Overview of the avian immune system
2.1 Innate immune system
Like most vertebrates, the avian immune system consists of a rapid non-specific innate immune system and a highly specialized adaptive immune system (34). The avian innate immune system consists of a broad range of physiological barriers as well as effector cells that generate rapid broad-spectrum responses encoded by germline genes. Several functionally distinct effector cells are present within the avian innate immune system, including natural killer (NK)-cells, heterophils (the avian homologue of neutrophils), macrophages, and innate-like T cells (e.g., γ-δ T cells), which were explored in detail here (35).
Avian monocytes are the major phagocytic component of the blood, which originate from bone marrow stem cells (BMSCs) and can differentiate into monoblasts, pro-monocytes, and monocytes. Avian monocytes can further differentiate into macrophages which populate and reside in different tissues (36). In contrast to mammals, the avian respiratory system lacks alveoli and a resident respiratory macrophage population, with lung lavages of healthy birds rarely being able to recover immune cells (36, 37). In birds, highly phagocytic free avian respiratory macrophages (FARMs) with effective antimicrobial activity appear to be key factors in pulmonary defense (38). FARM rapidly translocates to the lung following challenge, with lung macrophage population seen to increase dramatically following avirulent Pasteurella multocida intratracheal immunization (39), as well as E. coli and Salmonella challenge (36, 39). Induction of macrophage recruitment has been shown to provide non-specific protection, with intratracheal dosing of chickens with P. multocida Chloral vaccination strain protecting against virulent E. coli air-sac challenge 7 h later (40). This protection is attributed to the increased recruitment of macrophages to the site of infection or induction of innate immune memory.
The phagocytic function of avian macrophages is well characterized, and internalization has been demonstrated to increase when targets have been opsonized, suggesting a key role of receptors in this process, with Fc-υ and complement receptors for IgY and C3 opsonization, respectively, suggested to be significant (36). In addition, macrophages are activated by pattern recognition receptors (PRRs) that recognize pathogen-associated molecular patterns (PAMPs), including lipopolysaccharides (LPSs), flagellin, and foreign nucleic acids, triggering a physiological or signaling response (35, 41).
Toll-like receptors (TLRs) are the most studied class of PRR and, upon stimulation, activate the NF-κB and type I interferon (IFN) pathways (42). These release signaling molecules, including inflammatory cytokines and chemokines, as well as type I IFNs, leading to the recruitment of other immune cells (35). In total, 13 TLRs have been identified in mammalian species, of which orthologues of six have been found in chickens, alongside four additional TLRs. Chicken TLRs (chTLRs) 3, 4, 5, and 7, as well as duplicated chTLRs 2A and 2B, are direct orthologs of mammalian TLRs of the same name. Furthermore, the chicken repertoire also includes chTLR 21, an ortholog of TLR 21 in fish and amphibians, and chTLR 1LA, 1LB, and 15, which have only been reported in avian species (43). Several TLRs are associated with the response to bacteria, with disruption of chTLRs 1, 2, 4, 5, 7, 15, and 21 each of which associated with susceptibility to E. coli and Salmonella infection (43, 44).
2.2 Adaptive immune system
In contrast to the low specificity of the innate immune system, the avian adaptive immune system is highly targeted and associated with the generation of immunological memory (34). The avian adaptive immune system is generally divided into two different responses: cell-mediated responses aimed at the clearance of intracellular pathogens and humoral immune responses targeting extracellular pathogens (35).
Cell-mediated responses are primarily actioned by T cells that develop within avian species in a manner strikingly like that seen in mammals. The thymus is initially colonized by mesodermal hemotopoietic cells during embryonic development, with these progenitor T cells undergoing differentiation and T-cell receptor (TCR) gene rearrangement; prior to migration to the peripheries (45). Antigen recognition is facilitated by TCRαβ, which is a heterodimeric surface receptor composed of two immunoglobulin superfamily domains. These domains combine to form TCRαβ heterodimers. In chickens, the two T-cell lineages can be distinguished based on the types of chains they possess, which include αVβ1 and αVβ2 chains. T cells expressing αVβ1 chains from infected or vaccinated chickens, but not naïve birds, recognize peptides from avian viruses in association with classical MHC molecules (46). Thus, T cells expressing αVβ1 chains are part of adaptive immunity. However, it remains unclear whether T cells expressing αVβ2 chains are also part of adaptive immunity, as no specific antigens recognized by this TCR have yet been identified (46). Moreover, the CD4 and CD8 co-receptors, which bind to major histocompatibility (MHC) I and MHC II, respectively, have also been identified in chickens and allow for further classification. However, unlike in mammals, a clear distinction of cell populations into cytotoxic and helper cells has not been demonstrated, but evidence suggests that both conventional CD4+ and CD8+ cells and innate-like T cells, such as γδ T cells, are involved in the control of pathogens via cytotoxicity or production of cytokines (46–52).
Like mammals, humoral responses in chickens are mediated by B cells. However, the origin of B cells is unique to birds, as they develop in the bursa of Fabricius. Akin to the development of T cells, the bursa is colonized by lymphoid precursors during embryonic development, before migration to the periphery. B cells generate antibodies specific to the encountered pathogenic threats, which requires the development of a diverse antibody repertoire. In mammals, this typically occurs through immunoglobulin (Ig) gene rearrangement; however, while this still occurs in birds, the primary way in which diversity is generated is through somatic gene conversion (53). While Ig gene rearrangement occurs continuously within the bone marrow of mammals, somatic gene conversion occurs entirely during the period of embryonic development in avian species (54). The activation of B cells within the periphery of chickens is identical to that reported in mammals, with the binding and processing of antigens by the B cell receptor followed by activation by helper T cells inducing CD40 activation. Chicken B cells can then differentiate into plasma cells, allowing the secretion of Ig that is capable of binding to and opsonizing extracellular pathogens (53). This allows the generation of long-term protective responses following vaccination, with increased Ig production following a secondary challenge (55).
2.3 Innate immune memory
The innate immune system evolved more than 600 million years ago in early eukaryotic cells to preserve cellular integrity and resist exogenous assault (56). The adaptive immune system is comparatively new, developing in the first jawed vertebrates approximately 450 million years ago (57). However, the adaptive immune system or ortholog has not evolved in invertebrates and plants, yet many of these organisms can survive for decades despite numerous encounters with rapidly evolving bacterial threats. Organisms lacking an adaptive immune system have also been observed to respond more effectively to secondary pathogenic exposure, with green bean plants and mealworm beetles demonstrated to resist reinfection following immunization with heat-attenuated bacterial pathogens (58–60). Furthermore, these immune memory responses in invertebrates and plants contrast with those typically seen within the vertebrate adaptive immune system, protecting against both homologous and heterologous secondary challenges, rather than being highly specific (61). These observations have led to a closer examination and eventual revaluation of the role of memory within the innate immune system of vertebrates.
Early evidence suggests the existence of non-specific innate immune memory in vertebrates, centered on historic trends of significantly reduced heterologous infections following the introduction of smallpox and Bacillus Calmette-Guerin (BCG) vaccines in human populations (62). As non-specific protection is atypical for the immunological memory of the adaptive immune system, innate immune memory has been suggested as the probable cause. This culminated in the identification of four distinct memory profiles within innate immune cells, particularly myeloid cells, including monocytes, granulocytes, dendritic cells, and macrophages (63). These four phenotypes were trained immunity, priming, tolerance, and differentiation. Trained immunity (Figure 1A) is defined as a functional adaptive program within innate immune cells brought on by a change in the activation state following exposure to a primary stimulus resulting in epigenetic modifications that persist despite a return to basal activation levels after removal of the stimulus. Epigenetic modifications result in enhanced responses following reactivation via homologous or heterologous stimuli with increased cell functionality and/or gene expression (66). Meanwhile, priming (Figure 1B) showed a similar change in the functional state within immune cells following stimulation, but this did not result in a return to baseline prior to secondary activation. Ultimately, this results in an augmented response to the subsequent challenge, which can be cumulative or synergistic (66). Tolerance (Figure 1C), on the other hand, is characterized by a reduction in the responsiveness of immune cells to a secondary challenge following a change in functional state by primary stimulation. This is typified by reduced metabolic activity and silencing of genes associated with inflammation (67). The final memory profile associated with innate cells is differentiation (Figure 1D), a long-term change in a cell’s functional state when a naïve immune cell becomes mature (66). Targeting this underutilized functionality of the innate immune system may lead to therapies with the potential to protect the host from a range of pathogens (62). This potential broad-spectrum protection is of great interest to the poultry industry as a management strategy for infectious disease in the post-antibiotic era.
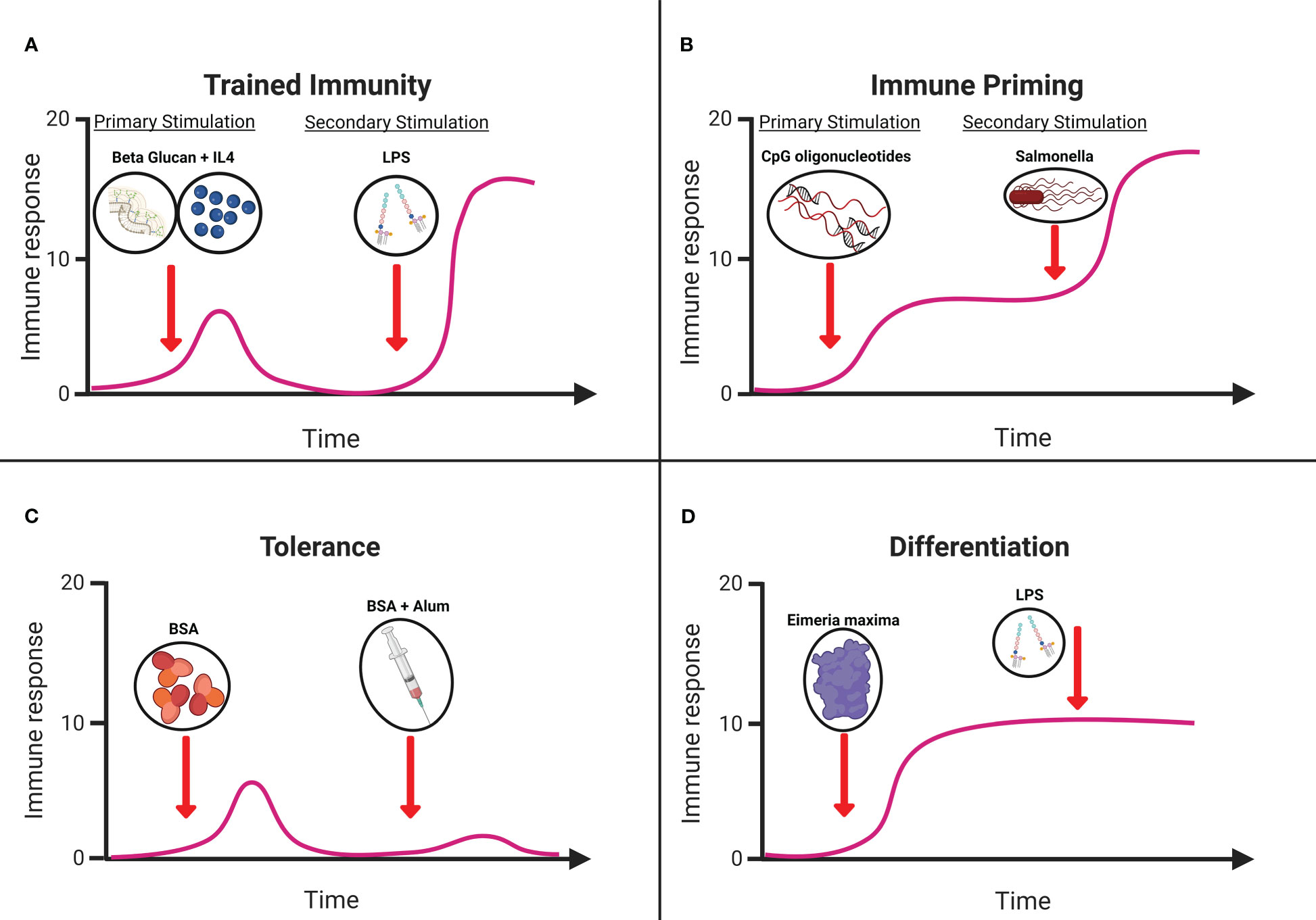
Figure 1 Graphical representation of (A) trained immunity. Treatment of primary chicken monocytes with Beta glucan microparticles and IL-4 significantly increased immune responsiveness to LPS (64). (B) Immune priming. Immunization with CpG oligonucleotides protecting chickens against Salmonella in vivo (65). (C) Tolerance. Dietary supplementation of bovine serum albumin (BSA) reduces immune responsiveness. (D) Differentiation. In vitro stimulation splenic dendritic cells with Eimeria maxima antigens resulting in morphological and functional changes. Schematic adapted from (66) created with Biorender.com.
3 Potential innate immune modulators
The advances we have made in our understanding of how the innate immune system works and its ability to improve through ‘training’ opens a clear opportunity to use it as a means of improving performance in animal husbandry. The ability to generate effective protection against a wide range of pathogens through the application of non-antibiotic agents holds great promise. Here, we present compounds that are potentially suitable for use in the poultry industry Table 1.

Table 1 Summary of immune modulators and their effects.
3.1 Prebiotics and probiotics
Prebiotics are indigestible carbohydrates that selectively promote the growth or activity of one or more bacterial species within the GI tract of the host to improve host health (130). Fructooligosaccharides (FOS), galactooligosaccharides (GOS), and mannanoligoasaccharides (MOS) are the most frequently utilized carbohydrates because of their inability to be broken down by vertebrate GI but can be metabolized by members of the microbiota (131). Prebiotics represent a viable alternative to growth-promoting antibiotics to improve growth, likely because of enhanced development of the avian innate intestinal barrier, resulting in improved nutrient uptake as well as innate responses. The innate intestinal barrier is key in host defense against infectious diseases, containing a range of cell types that directly contribute to host defense, such as enterocytes and Paneth cells which secrete antimicrobial peptides and prevent colonization of the intestinal surface by pathogenic bacteria. Moreover, the lamina propria within the epithelium contains antigen-presenting dendritic cells that facilitate crosstalk between innate and adaptive immune systems (132). MOS administration has been observed to modulate the innate immune system within the avian gut, increasing the goblet cell number, size, and density (68–70). Furthermore, prebiotic administration has also been shown to modulate innate effector cell responses as inclusion of MOS in broiler diets increased TLR2b, TLR4, IL-12p35, and IFN-γ expression within the GI tract (71). β-1,4-Mannobiose (MNB) supplementation of broiler diets also results in an increase in antigen presentation, host defense, and interferon-related genes (72). The in ovo delivery of inulin also increased the heterophil/leucocyte ratio and phagocytic ability at days 21 and 35 post-hatch. Additionally, the same study observed that in ovo administration of a GOS-based commercial prebiotic also increased leukocyte oxidative potential (73). Collectively, this suggests that prebiotics can effectively enhance the innate immune response within the avian GI tract, likely improving host responses to bacterial challenge, which, combined with the ability of prebiotics, inhibits pathogenic bacterial colonization of the GI tract through direct binding to pathogens or competitive exclusion (133).
Probiotics are live microorganisms that can confer health benefits to the host if administered in adequate amounts (134). The administration of probiotics can prevent infection through several mechanisms, including maintaining intestinal microflora, altering metabolism by enzymatic modulation, improving feed conversion, and stimulating the immune system (135). Several studies have identified an association between the microbiota and burden of pathogenic bacteria, with an increased Campylobacter load in the ceca of broiler chickens correlating with increased Enterobacteriaceae and reduced Lactobacillus abundance, respectively (136). This highlights the importance of microbiota composition and its effects on the burden of bacterial diseases in poultry. Therefore, the application of pre- and probiotics, aims to modulate microbial communities to improve performance and prevent disease.
One way in which probiotics may contribute to the modulation of the innate immune system is through the enhancement of epithelial barrier function, with oral gavage of L. reuteri in young chicks, resulting in enhanced intestinal cell proliferation and promotion of differentiation into mucin-producing goblet cells, likely due to the activation of Wnt/β-catenin and Mucin 2 expression, respectively. These effects, along with enhanced lysozyme expression following L. reuteri administration, suggest an improved innate barrier function in the intestinal tract following probiotic treatment (74). Administration of E. faecium to broiler chicks has also been shown to induce intestinal cell proliferation and reduce the number of apoptotic cells (76). However, this effect may vary depending on the phenotypic profile of the microbe, and a later study observed that oral administration of L. reuteri enhanced the villus height: crypt depth ratio but did not increase goblet cell numbers or mucin 2 expression (75). Pre-stimulation of the HD11 chicken macrophage-like cell line with the yeast probiotic Saccharomyces boulardii has also been observed to significantly increase phagocytosis and bactericidal activity against C. perfringens. Notably, pre-stimulation downregulates C. perfringens-induced pro-inflammatory responses, reducing the expression of IL-6, IL-10, TNF-α, and inducible nitric oxide synthase (iNOS) (77). Together, these results suggest an enhanced innate antimicrobial response, and a simultaneous reduction in inflammatory responses, which may be detrimental to bird performance and health. Dietary supplementation with B. subtilis has also resulted in immunomodulation in broiler chickens, significantly increasing pro-inflammatory cytokine production but greatly reducing antibody levels specific to Eimeria spp. and C. perfringens following challenge (78). These changes may suggest a shift in the immune response away from adaptation to the innate immune response following B. subtilis supplementation.
The role of gut microbiota in health and disease has been further elucidated in recent years, with dysbiosis and disruption of the gut microbiota linked to increased disease risk. The importance of microbial crosstalk within the gut–lung axis in mammals has also been observed, with disruption resulting in increased susceptibility to respiratory infections (137). Furthermore, the application of probiotics to modulate the gut microbiota has been shown to provide protection against respiratory diseases in humans, blocking the COVID-19 virus from entering and proliferating within host cells, as well as suppressing inflammatory responses (138). Further research is necessary to confirm the presence of the same axis within poultry, but evidence suggests that manipulation of the bird microbiota also has the capacity to directly modulate the innate immune system. This was clearly demonstrated with the inoculation of newly hatched broiler chickens with a microbial cocktail derived from adult birds, resulting in higher numbers of IL-2Rα+ and activated NK cells within the intestine at days 3 and 35, respectively. Increased NK cell activation was also observed in the blood and spleen of birds inoculated with adult-derived microbiota (79). This suggests that the composition of the microbiota has a significant impact on immunity in young birds and could be a valuable target for probiotic applications in the future.
Probiotics have historically been associated with improvements in gut health, along with recent developments in elucidating the role of the gut microbiota in the prevention of diseases within other organ systems, such as the respiratory system (137). Moreover, the microbiota has also been implicated in the development of immunity, particularly its role in the host response to vaccination in humans. It has been observed that microbiota composition in early life is associated with responsiveness to immunization (139, 140). This may be the case in poultry, with antibiotic depletion of the microbiota resulting in impaired IgM and IgY antibodies, and IFN-γ responses to H9N2 vaccination, with proper immune response restored following fecal microbial transplantation (141). However, further research is required to determine the capacity of the poultry microbiota to influence immunity to bacterial vaccination.
3.2 Beta-glucans
Beta-glucans are heterologous glucose-based polysaccharides produced by many fungi, bacteria, and plants. The pharmacological potential of the β- (1–3)-linked β-D-glucopyranosyl backbone and varying β- (1–6)-linked side chains have been extensively studied (142). Beta-glucans from a variety of sources have been tested for their efficacy in improving avian health and response to infection. Several studies have highlighted the benefits of dietary β-glucan supplementation in the enhancement of growth and nutrient digestibility in farmed animals, including pigs (143), cattle (144), and fish (145). In poultry, dietary supplementation with S. cerevisiae beta-glucan polysaccharide has been demonstrated to result in significant improvement in chick weight gain and improved FCR (80, 146–150). Beta-glucan supplementation also increases goblet cell numbers within the jejunum following Salmonella Typhimurium infection (82). This suggests that β-glucan may contribute to the maintenance of an effective gut barrier, which is critical for health and disease (151). Beta-glucan use has also been demonstrated to help prevent bacterial infection, with the inclusion of purified beta-glucan in the feed of white-leghorn chickens significantly reducing invasion and colonization of visceral organs by Salmonella Enteritidis in young chickens compared to a diet without the beta-glucan ration (83). Supplementation of the turkey diets with commercial β-glucan feed additives also resulted in reduced E. coli colonization in the air sacs and livers after challenge at one week of age with and without the birds undergoing transport stress (84). Continuous Beta-glucan supplementation of the turkey diets at 1 during the 16-week rearing period also significantly reduced the isolation of Salmonella and Campylobacter from the caeca of birds exposed to transport stress (81). This improved responsiveness to bacterial challenge is suggested to be a result of the ability of beta-glucan to bind to and interact with white blood cells such as macrophages via the complement receptor 3 and dectin-1 receptors, modulating immune responses (152). Activation of dectin-1 receptors has been demonstrated to amplify toll-like receptor (TLR) responses, including the production of cytokines and reactive oxygen species (ROS) and promoting phagocytosis (153, 154). Beta-glucan microparticles have been shown to induce the development of a trained immunity phenotype in primary chicken monocytes when co-administered with interleukin-4 (IL-4). After secondary stimulation with LPS, trained broiler and layer BMDMs showed significantly increased NO production and surface expression of colony-stimulating factor 1 receptor (CSF1R), CD40, and major histocompatibility complex class II (MHC-II) (64, 155). This increased NO response after secondary stimulation aligns with the trained immune phenotype observed in mammalian models (156). This highlights the potential of β-glucan to act as a modulator of innate immunity in poultry. In vivo, dietary supplementation with β-glucan in one-day-old chicks has also resulted in altered cytokine profiles in peripheral blood serum and increased expression of the pro-inflammatory factors interleukin (IL)-1, IL-2, interferon (IFN)-γ, and tumor necrosis factor-alpha (TNF-α) (80), factors associated with inflammatory responses to infections. The direct effect of dietary supplementation on the functionality of innate immune effector cells was investigated by isolating heterophils. This showed that heterophiles isolated from β-glucan-fed chickens had significantly increased phagocytosis, antibacterial activity, and oxidative burst capacity (83). In addition to heterophils, beta-glucan supplementation of one-day-old broiler chicks resulted in increased macrophage phagocytic activity and proportion of CD4 and CD8 positive lymphocytes (85). Modulation of innate effector cell functionality was also observed in turkey poults following dietary supplementation with commercial beta-glucan supplement. Isolation and examination of peripheral blood leukocytes at three weeks of age revealed an increased level of heterophils in the peripheral blood, with increased oxidative burst potential (84). Furthermore, dietary supplementation with 1% sophy β-glucan in Peking ducks significantly increased the proliferation of peripheral blood mononuclear cells in a dose-dependent manner (86), suggesting increased cellular immunity.
3.3 Pathogen associated molecular patterns
Bacterial, viral, fungal, and parasitic pathogens contain various components that are recognized by and stimulate immune cells. Vaccination with specific antigens has proven to be effective as a means of generating protection. However, in recent years, there has been increasing research into how stimulation of the innate immune system by PAMPs elicits a protective immune response. Single-stranded DNA PAMP Cytosine-phosphorothioate-guanine oligodeoxynucleotides (CpG ODN) have been identified as potent immunostimulatory molecules, with potential utility in cancer immunotherapy (157, 158), vaccine adjuvants (159), and mucosal vaccination (160). In poultry, immunostimulation with synthetic CpG ODN has been demonstrated to confer protection against bacterial infections. Subcutaneous or intramuscular administration of 10 ug or 50 µg to 22-day old broiler chicks significantly improved the survival of birds after E. coli challenge at 25 days post-hatching (87). Furthermore, intramuscular administration of 50 µg CpG-OGN to neonatal broiler chicks also significantly improved survival following virulent E. coli and Salmonella Typhimurium challenge (65, 88, 89). Treatment also resulted in a dramatic reduction in the frequency with which bacteria were isolated in CpG OGN-treated birds compared to the untreated group (65, 87–89), as well as a significant reduction in clinicopathological presentations following infection (65, 87, 89). Similar findings have also been reported in newly hatched birds treated with intraperitoneal administration of CpG ODN. Here significantly reduced colonization of visceral organs was observed in treated chicks following Salmonella enteritidis challenge (90). Needle-free intrapulmonary administration of 4 mg CpG-ODN per chamber between 6 h and five days before E. coli challenge was also demonstrated to reduce clinical presentations, frequency of bacterial isolation, and mortality without affecting the growth of birds (91). The efficacy of protection induced by CpG ODN was enhanced by formulation with carbon nanotubes and lipid surfactant delivery systems, which further reduced mortality and clinical presentations of colibacillosis disease (92). However, owing to the scale of commercial poultry farming, individually administered immunization is of little use. This has led to investigations into alternative routes, such as in ovo delivery, which can be achieved on a larger scale. Indeed, in ovo immunization with 50 µg of CpG ODN on day 18 of incubation significantly improved protection against E. coli or Salmonella typhimurium infection two days post hatch. This also reduces frequency of bacterial isolation from the air sacs of birds (88, 89). The protection rate against post-hatch E. coli challenge was further improved by in ovo immunization with the formulation of CpG OGNs with carbon nanotubes or lipid surfactants, without causing any adverse effects (92). Furthermore, in ovo administration of CpG ODNs significantly improved protection against an experimental model of colibacillosis yolk sac infection, which is a significant cause of early mortality in chicks (93). The mechanism involved in the protective effect of CpG ODN administration may be its ability to modulate the innate immune system responsiveness, which is characterized by significantly enhanced degranulation and oxidative burst responses in heterophils (90). Moreover, CpG ODN-induced immune responses have also resulted in a shift in metabolic profiles in innate immune cells, greatly upregulating glycolysis and fatty acid synthesis while downregulating ketogenesis and fatty acid β-oxidation (65). These metabolic shifts are strikingly like those observed in metabolic remodeling after the induction of trained immunity. Under these conditions, trained immune cells display significantly enhanced glycolysis, increased fatty acid metabolism, and oxidative phosphorylation pathways, leading to the promotion of epigenetic changes involved in proinflammatory responses (161). In fact, protection against E. coli meningitis in a murine model by intraperitoneal administration of CpG ODN has been cited as evidence for trained immunity in mammals (63, 162), suggesting a likely role for the innate program within the protection observed within poultry.
The use of PAMPs as immunostimulants for the induction of protective immunity in poultry has also been investigated. Lipopolysaccharides (LPS) form an important part of the outer membrane of gram-negative bacteria and are recognized by TLR4, resulting in a potent inflammatory response and the induction of cytokine expression (163). LPS from the plant symbiotic bacterium, Pantoea agglomerans, commonly found in edible plants (164), has been tested to prime the innate immune system against infection. Previously, its use in invertebrates was shown to improved phagocytic responses in shrimp (165) and carp (166). Oral administration of 10 ug/kg body weight/day LPS to broiler chicks significantly reduced their mortality within the first 10 weeks of age. Furthermore, analysis of the functional abilities of peripheral blood monocytes from birds administered LPS showed an increase in their phagocytosis, as well as NO and IL-1β production after stimulation with heat-killed E. coli (94). P. agglomerans LPS also inhibited dexamethasone-induced immunosuppression, cell death in thymic and bursal lymphocytes, and significantly increased the relative weight of both organs. This treatment also increased antibody production after S. enteritidis vaccination compared with that in control birds (95).
3.4 Microbial metabolites
The microbiota has been identified as inextricably linked to the development and function of the immune system, with dysbiosis associated with improper immunological responses in mammals (167) and chickens (168). Microbiota has been shown to communicate with the immune system, particularly at mucosal surfaces, through the production of a variety of molecules, often referred to as metabolites (169). These metabolites include short-chain fatty acids (SCFAs), secreted proteins or peptides, organic acids, biosurfactants, flavonoids, and vitamins (170).
Broadly defined as exogenously or endogenously generated intermediates or products of metabolism, metabolites can modulate the vertebrate immune system (169). Bile acids, known as secondary bile acids (SBAs), are host-derived molecules that undergo microbial transformation by members of the gut microbiota and gain novel functions (171). Two of these SBAs, deoxycholic acid (DCA) and lithocholic acid (LCA), are the most abundant metabolites within the human microbiome (172), as well as important immunomodulators (171). In poultry, multiple studies have demonstrated that dietary supplementation of one-day-old broiler chicks with DCA reduced clinical histopathology in the ileum and the resultant weight loss following Eimeria maxima and C. perfringens-induced necrotic enteritis (NE). These phenotypic changes following DCA supplementation in chickens, were likely due to reduced C. perfringens and E. maxima colonization in the ilium (96, 97). DCA alone inhibited C. perfringens growth in vitro (97); however, DCA supplementation resulted in a reduced pro-inflammatory innate response to NE, including lower expression of IFN-γ and MMP9 cytokines (96), Lipopolysaccharide-Induced TNF Factor (LITAF), and Cyclooxygenases-2 (COX-2) (97). This attenuation of the inflammatory response may contribute to reduced clinical signs after NE infection. Supplementation of day-old broiler diets with DCA has also been effective at protecting against C. jejuni infection, preventing colonization, and improving bird performance following infection (98). Interestingly, DCA had no impact on C. jejuni viability in vitro, suggesting that modulation of the microbiota contributes to protection. Similar results have also been observed in mice, with oral DCA administration resulting in the prevention of C. jejuni colitis. However, we observed that DCA inhibited proinflammatory gene expression of IL1β, Cxcl2, and IL17a, but also led to downregulation of the mammalian target of rapamycin (mTOR) pathway within primary splenocytes following exposure (99). This suggests a potential modulatory effect of DCA on innate cells and correlates with a reduction in proinflammatory responses (173). Furthermore, mTOR downregulation has also been associated with enhanced antimicrobial activity in macrophages, with exposure of primary murine monocytes to the short-chain fatty acid butyrate during differentiation, resulting in significantly increased bacterial intracellular killing, alongside decreased mTOR (174).
Organic acids have been frequently use as food preservatives and exert antimicrobial effects on the growth of pathogenic bacteria (33). Many organic acids, particularly short-chain fatty acids (SCFAs), are produced by the fermentation of undigested carbohydrates by members of the gut microflora. Some of these organic acids have been included in animal feed and function as fungistats (33). However, there is growing evidence to suggest that SCFAs may have a potent immunomodulatory effect, with butyrate being associated with enhanced host defense peptide (HDP) expression in the HD11 chicken macrophage cell line, primary chicken monocytes, and intestinal explants. Furthermore, chicken monocyte antibacterial activity against S. enteritis was seen to increase with butyrate exposure in a dose-dependent manner, with dietary supplementation of chicken feed with sodium butyrate resulting in protection against challenge (100). Butyrate also significantly reduced the expression of pro-inflammatory cytokines IL-1β, IL-8, and MMP9 following S. enteritis challenge (101). Supplementation with butyrate attenuated NE-induced weight loss and reductions in FCR, as well as the severity of clinical lesions (102). Butyrate has also been shown to modulate the response of innate immune cells associated with physical barriers to infection, with exposure of primary chicken enterocytes to butyrate resulting in significantly reduced adhesion and invasion of S. enteritis (101).
Propionate is a three-carbon SCFAs produced by the Bacteroidetes, Firmicutes, and Lachnospiraceae families within the GI tract (175). Dietary supplementation with sodium propionate has been demonstrated to dramatically alter the composition of the cecal microbiota and inhibit fat deposition within broilers by downregulating fat synthesis genes and affecting feed consumption (176). Moreover, dietary supplementation with propionate associated with chromium has been observed to significantly improve final body weight, weight gain, feed efficiency, and carcass characteristics in broilers compared to control diets (177). In mammals, propionate has been shown to inhibit S. typhimurium colonization, likely through the inhibition of growth and modulation of intracellular pH (178). There is also extensive evidence in mammals suggesting that propionate has immunomodulatory properties. This includes the inhibition of inflammatory responses following stimulation with S. aureus in vitro and in vivo, with murine macrophages pre-incubated with propionate demonstrating reduced nitric oxide production and decreased pro-inflammatory cytokine production in mice treated with propionate following S. aureus challenge (179, 180). Propionate also modulates AMP expression in pig intestinal epithelial cells and macrophages (103). Indeed, this has also been observed in poultry exposed to chicken HD11 cells and primary monocytes exposed to propionate, resulting in significantly increased AvBD9 and cathelicidin B1 expression. Moreover, supplementation with propionate resulted in significant reductions in cecal S. Enteritidis following the challenge of four-day-old broiler chicks (103).
3.5 Vitamins
Vitamins, which are essential organic compounds, can also be produced by the microbiota or attained through the diet. Several vitamins have been found to be crucial for immunological functions of immune cells including macrophages and lymphocytes (181). The effects of dietary vitamins on the immune system of poultry have been reviewed by Shojadoost et al., who focused on the effects of vitamins on the innate immune system.
Vitamin D has been identified as a key contributor to innate immunity, promoting antibacterial responses, and pattern recognition receptors (PRR) and cytokine gene expression in humans (182). In poultry, supplementation with active vitamin D3, 25-hydroxycholecalciferol (25-OH-D3) improved survival and ameliorated inflammatory stress (104, 183, 184). Furthermore, a dietary deficiency of active vitamin D3, 25-hydroxycholecalciferol (25-OH-D3), reduced the performance and egg quality in laying hens, and this condition was resolved with 25-OH-D3 supplementation (104). Vitamin D3 supplementation significantly improved FCR in broiler chicks (185), increased villus length/crypt depth ratios within the first 21 days of growth in broiler duodenum and jejunum (186), and significantly improved body weight following LPS challenge (105). Collectively, these studies suggested that 25-OH-D3 is important for poultry productivity. There is also great interest in the ability of 25-OH-D3 to act as an anti-inflammatory modulator. 25-OH-D3 supplementation reduces pro-inflammatory responses following LPS challenge. This included a reduction in pro-inflammatory IL-1β and IL-6 expression, as well as the IFN-γ:IL-4 ratio after LPS challenge (104, 105). It is hypothesized that modulation of the NF-κB pathway contributes to the effects of 25-OH-D3 (104). Similarly, inhibition of proinflammatory cytokines, such as IL-1β and CXCL8, was observed in the MQ-NCSU chicken macrophage cell line pre-treated with 25-OH-D3 (106). Furthermore, pretreatment with 25-OH-D3 also increased NO production from macrophages after LPS or Pam3-CSK stimulation. Moreover, 25-OH-D3 reduced the inflammatory response of chicken T lymphocytes but did not affect their degranulation response (107). Modulation of innate effector cells after dietary 25-OH-D3 supplementation in broilers was observed, as demonstrated by improved ex vivo monocyte phagocytosis, chemotaxis, and bacterial killing (108). Together, the improvement in innate immune cell function may confer protection against a range of bacterial infections and reduce immunopathology, which can reduce productivity by reducing tissue damage (187) in poultry. However, further investigations are needed to determine the effectiveness of 25-OH-D3 supplementation in preventing or reducing bacterial infections.
After hatching, the inclusion of Vitamin A group of fat-soluble micronutrients, inhibits production losses associated with vitamin A deficiency, which is known to cause poor growth, reduced egg production, and impaired immune development and function (188, 189). The inclusion of vitamin A also improved performance, as measured by significantly higher body weights observed in broiler chickens supplemented with the standard (1,500 IU/kg) and high 1(5,000 IU/kg) vitamin A-supplemented diets compared to unsupplemented and pair-fed controls (190). Vitamin A supplementation also improved the wing weight of broilers in a dose-dependent manner. This regime also reduced the occurrence and severity of wooden breasts and white striping within meat (191). In mammals, vitamin A has been linked to the development of physiological barriers, and its deficiency results in impaired respiratory, gastrointestinal, and urogenital mucosal barrier function, which is partially attributed to the loss of mucus-secreting goblet cells (192). In poultry, vitamin A deficiency decreases respiratory mucin and IgA production (109) and gastrointestinal goblet cell proliferation (110), and this condition is associated with the development of squamous metaplasia and epithelial damage in turkeys (111). Taken together, these results suggest that the loss of the protective mucosal barrier on epithelial surfaces increases the risk of infection (189, 192). Vitamin A is also essential for the development of innate immune effector cells, and its deficiency diminishes the ability of mammalian phagocytes to engulf and lyse bacteria after infection (192). In poultry, vitamin A deficiency significantly reduced phagocytosis of heterophils in broiler chickens at 49 days post-hatching. Furthermore, addition of vitamin A exceeding the recommended dosage increased the phagocytosis index of heterophils at 28- and 49-day post-hatch compared to both vitamin A-deficient birds and birds at the recommended dose (112). Vitamin A supplementation increased the phagocytosis index of chicken intra-abdominal macrophage isolates from white-leghorn chickens compared to those from vitamin A-deficient birds (193), suggesting a vital role for vitamin A in the functional abilities of immune cells. The presence of vitamin A has been linked to the regulation of inflammatory processes, and its deficiency upregulates the expression of pro-inflammatory cytokines and immunopathology in a rat model (192). Furthermore, pre-incubation of murine macrophages with vitamin A in the form of retinoids inhibits the expression of inflammation-associated IL-12 and IFN-γ following activation both in vitro and in vivo (115). Therefore, it was unexpected that the in ovo administration of 90 μmol/egg vitamin A in broiler chicks enhanced IL-12 and IFN-γ expression, suggesting a differential response in birds (113). However, an earlier study by the same group observed that the immunomodulatory effects of vitamin A were dose-dependent; in ovo administration of 90 μmol/egg retinoids upregulated IFN-α, IFN-γ, IL-1β, IL-2, IL-8, IL-12, and IL-13 in chicken embryos, but 270 μmol/egg downregulated these cytokines (114).
Vitamin E is a group of lipid-soluble molecules, well known for their antioxidant properties and ability to modulate immune cells, as they are found in higher concentrations in these cells than in other cells in the blood (194). Vitamin E exists in four functional forms α, β, γ, and δ, with α being the most abundant and functionally active form (189). Poultry feed supplementation is believed to improve bird performance, with higher broiler body weight and improved FCR observed each week for five weeks post-hatching (195). However, these findings were not confirmed in any of these studies (196, 197). Vitamin E deficiency greatly impairs the ability of the adaptive immune system to respond to infection and is associated with reduced antibody production and lymphocyte proliferation. Vitamin E deficiency results in increased inflammatory cytokine production and impaired antibacterial and phagocytic activities of neutrophils in infants (194). Furthermore, in vivo supplementation of excess vitamin E increased the phagocytic activity of rat alveolar macrophages (198), suggesting that vitamin E ability can enhance innate immune responses. In poultry, excess dietary vitamin E increased phagocytosis of opsonized sheep red blood cells by macrophages isolated from the abdominal exudate of three-week-old broilers. However, no such increase was observed in the phagocytosis of non-opsonized cells or in broilers at five- and seven-week post-hatch (116). Vitamin E has also been studied for its potential protective effect against bacterial infection. The administration of vitamin E to the drinking water of turkeys resulted in a reduction in mortality and airsacculitis following E. coli challenge as well as a reduction in bacterial colonization of the liver. The addition of vitamin E has also been associated with a shift in the heterophil-to-lymphocyte ratio, indicating an enhancement in innate responses (117). Moreover, vitamin E supplementation has been observed to improve laying performance and reduce mortality following S. enteritidis infections (118).
Unlike the other vitamins discussed above, vitamin C is water-soluble and synthesized from glucose. Moreover, vitamin C is not stored in the body; however excess dietary intake is excreted by the kidneys. Unlike humans, poultry can produce endogenously but require supplementation during times of stress (189). Indeed, vitamin C supplementation has been observed to improve poultry performance, egg weight, and quantity as well as enhance FCR and carcass weight in layers and broilers, respectively (199). Vitamin C is a potent antioxidant that functions as a cofactor for the enzymes involved in biosynthesis and regulation. Vitamin has also been observed to play a vital role in the function of the innate immune system, helping maintain barrier integrity, enhancing activity, acting as an electron donor for phagocytes, and modulating cytokine expression (119). Indeed, heterophils isolated from broilers had improved bacterial killing ability in vitro following vitamin C exposure, although no increase in phagocytosis was detected (120). This increased antimicrobial activity may be because of vitamin C on the production of reactive oxygen species (ROS) from heterophiles. This notion is confirmed by results showing that vitamin C deficiency reduces human neutrophils to produce ROS, which can be rectified by vitamin C supplementation (200). This suggests that vitamin C facilitates ROS production in the innate effector cells. Supplementation with vitamin C has also improved protection against bacterial infection, which is associated with a reduction in the translocation of S. enteritidis to the liver, maintaining intestinal morphology, and the composition of the cecal microbiota after infection. However, no significant reduction in mortality was observed in these animals (121). This is in accordance with the observation that prophylactic supplementation of vitamin C, for six days after hatching led to a significant reduction in S. enteritidis colonization in broiler crops after infection. Moreover, therapeutic administration of vitamin C, in one-day old broiler chicks significantly reduced bacterial colonization of the crop and cecal tonsils at three- and ten days post-infection (122).
3.6 Plant derived compounds
In human medicine, growing evidence suggests that plant extracts may have potent immunomodulatory potential, affecting neutrophils (201, 202), leukocyte function (203), and intestinal barrier function (204). This has also been observed in poultry, with intestinal intraepithelial lymphocytes isolated from broilers fed carvacrol, cinnamaldehyde, or capsicum oleoresin, demonstrating altered metabolic gene expression and a shift towards lipid metabolism (123). Additionally, dietary supplementation of day-old broilers with thymol significantly increased trans-epithelial electric resistance (124), suggesting reduced susceptibility to infection, as loss of membrane integrity is associated with bacterial pathogenesis (205). In the same study, thymol supplementation was also observed to increase isolated blood phagocyte engulfment of microspherical hydrophilic particles, indicating additional immunomodulatory effects on innate effector cells (124). Dietary supplementation with cranberry extract has also been demonstrated to significantly enhance chicken heterophil antibacterial activity, increasing phagocytosis, and intracellular killing of S. aureus in vitro (125). Meanwhile, enhancement of phagocytosis of both Indian ink and sheep erythrocytes was also reported in peripheral blood phagocytes isolated from laying hens following stimulation with ethanolic sea buckthorn extract. However, garlic extract diminished the phagocytic ability of these cells, suggesting an extract-dependent effect (126). Milk thistle, turmeric, shiitake, and reishi mushroom extracts, as well as cinnamaldehyde have all been demonstrated to significantly enhance the proliferation of chicken spleen lymphocytes as well as modulate cellular responses, increasing NO production and expression of pro-inflammatory cytokines in the HD11 cell line following stimulation (127, 128). Hesperidin and genistein, flavonoids derived from citrus and soy, respectively, also increased lymphocyte number within the intestinal epithelium following dietary supplementation of 21-day-old-broiler chicks (129).
4 Discussion
Bacterial diseases can have a devastating effect on poultry welfare and productivity in poultry rearing systems (206). Antibiotics have been widely used to treat or prevent bacterial infections and to promote growth in poultry farms. However, this use has led to the emergence of antibiotic-resistant bacteria, which can cause serious health issues in both humans and animals. It is crucial for the poultry industry to take proactive steps to reduce the use of antibiotics and use alternative strategies to control and treat bacterial infections. These factors, combined with the need to expand food production to accommodate the growing global population collectively highlight the potential value of effective manipulation of the avian innate immune system by immunostimulatory compounds. Here, we have highlighted several ‘classes’ of immunomodulatory compounds that can enhance poultry performance and immune responses to bacterial pathogenic challenges. The use of these compounds as either feed additives or through direct application in vivo or in ovo presents an attractive management strategy owing to the non-specific responsiveness and ubiquity of the innate effector cells and physiological barriers within birds (34). Thus, effective innate immunomodulation and/or induction of trained immunity within birds may provide effective protection against multiple distinct pathogens following the application of a single stimulant (207). The ability to protect against a range of pathogens is highly beneficial and difficult to achieve using traditional vaccination strategies (25). Moreover, the ability to induce protection against multiple bacterial pathogens may potentially facilitate a reduction in antibiotic use or a shift towards narrow-spectrum antibiotics, a key recommendation in poultry antibiotic stewardship programs (208).
The reported capacity of immunomodulators to improve bird performance, health, and welfare is also highly desired and reminiscent of the widespread use antibiotic growth promoters without the risk of AMR selection (209). Indeed, supplementation of prebiotics, such as S. cerevisiae derived MOS (210, 211) and beta glucan have been shown to be effective at increasing bird performance, leading to enhanced FCR and weight gain (80, 146–150). Several probiotics have also led to increased weight gain in birds following supplementation (212) in addition to plant bioactive compounds (213–215) and essential oils (216). Collectively, this suggests that the immunomodulators discussed may provide the combined benefit of inducing effective protection against bacterial infection, in addition to enhancing bird performance, resulting in greater yields. Additionally, the favorable modulation of the innate immune system may also reduce production costs, as currently within the UK, and broilers routinely receive over a dozen separate vaccine treatments within the first 24 days of life (217). These treatments, as well as additional vaccines that may also be included due to seasonal factors or pathogenic outbreaks, all present financial and labor costs to purchase and implement (217). Effective modulation of the innate immune system may provide protection against multiple vaccine targets, thereby reducing costs. The ability of immunomodulatory compounds to effectively impact bird health when included as dietary additives may also avoid potential issues associated with spray vaccination, such as sudden drops in body temperature during application at one-day post-hatch within hatcheries and poultry units (218). Moreover, there remains the potential for the repurposing of waste products from other industries as stimulants, such as the use of beta-glucans from spent yeast being extracted for technological applications (219). In addition to reducing the costs associated with the production of immunostimulatory compounds, this also increases sustainability and minimizes environmental impact. Owing to the non-antibiotic nature of the compounds examined, a limited environmental impact would also be likely due to the reduced presence of antibiotic compounds in animal-derived compounds and animal waste, reducing the likelihood of AMR selection within the local environment (220). Proper modulation of the innate immune system may also enhance vaccine effectiveness, increase responsiveness, and promote crosstalk between the innate and adaptive immune systems, which is an essential factor within effective vaccination (221).
However, despite the many potential benefits associated with manipulation of the innate immune system, a greater understanding of the potential side effects is required before widespread adoption. One central issue that may detrimentally impact immunomodulation in poultry is the disadvantageous induction of inflammation. Indeed, in mammals, the induction of trained immunity has been found to be strongly associated with atherosclerosis following training with both β-glucan and BCG (222, 223). Furthermore, inappropriate activation of trained immunity has also been suggested to be associated with the development of autoimmune and autoinflammatory conditions, which would be deleterious to bird welfare and performance if this occurs in poultry (224). It can also be postulated that the risk of adverse inflammatory responses may increase because of the intensive vaccination programs currently associated with poultry production. Thus, there is a need to investigate the effectiveness of immune stimulants in birds under a range of conditions, including different farming setups, birds that have undergone vaccination, and different bird breeds, to accurately evaluate their effectiveness in situ.
In summary, our review indicates that innate immune modulation could be a promising approach for controlling bacterial infections in poultry and for improving their performance. Based on previously published literature, we identified a range of compounds that have shown potential in enhancing the innate immune response to bacterial challenges. However, further work is needed to (i) determine whether this approach is effective against all bacterial infections in chickens, (iii) evaluate the efficacy and safety of using specific compounds to control bacterial infection, and (iii) explore the potential benefits and risks of combining these compounds with vaccines against bacterial infections to enhance vaccine effectiveness without inducing an inflammatory response leading to immunopathology in birds. The optimal compounds are those that control bacterial infections, improve growth performance and gut health barriers, and reduce the severity of inflammation.
Author contributions
JA and SB contributed to the conception and structure of the review. JA performed the literature search and wrote the first draft of the manuscript. SB and JA wrote sections of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This review was supported by the U.K. Research and Innovation Biotechnology and Biological Sciences Research Council Grants BBS/E/I/00007030, BBS/E/I/00007031, BB/S01506X/1, BBS/E/I/00002529, BBS/E/I/00007039, BBS/E/I/00007032, BB/N002598/1 and BB/V019031/1, in addition to funding from the British Egg Marketing Board Research and Education Trust and the University of Surrey.
Acknowledgments
Permission was granted for adaption of Figure 1 from Divangahi et al., 2020. License number 5507690448896.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. WRI. Creating a sustainable food future: interim findings. In: World Resources Report. World Resources Institute Washington, DC (2013).
2. Kleyn FJ, Ciacciariello M. Future demands of the poultry industry: will we meet our commitments sustainably in developed and developing economies? World's Poultry Sci J (2021) 77(2):267–78. doi: 10.1080/00439339.2021.1904314
3. Godfray HCJ, Aveyard P, Garnett T, Hall Jim W, Key Timothy J, Lorimer J, et al. Meat consumption, health, and the environment. Science (2018) 361(6399):eaam5324. doi: 10.1126/science.aam5324
4. Hafez HM, Attia YA. Challenges to the poultry industry: current perspectives and strategic future after the COVID-19 outbreak. Front Veterinary Sci (2020) 7:516. doi: 10.3389/fvets.2020.00516
5. Bessei W. Impact of animal welfare on worldwide poultry production. World's Poultry Sci J (2018) 74(2):211–24. doi: 10.1017/s0043933918000028
6. Jones PJ, Niemi J, Christensen J-P, Tranter RB, Bennett RM. A review of the financial impact of production diseases in poultry production systems. Anim Production Sci (2019) 59(9):1585. doi: 10.1071/an18281
7. Meunier M, Guyard-Nicodème M, Dory D, Chemaly M. Control strategies against Campylobacter at the poultry production level: biosecurity measures, feed additives and vaccination. J Appl Microbiol (2016) 120(5):1139–73. doi: 10.1111/jam.12986
8. Stanaway JD, Atuhebwe PL, Luby SP, Crump JA. Assessing the feasibility of typhoid elimination. Clin Infect Dis (2020) 71(Supplement_2):S179–84. doi: 10.1093/cid/ciaa585
9. Shane SM, Faust A. Evaluation of sanitizers for hatching eggs. J Appl Poultry Res (1996) 5(2):134–8. doi: 10.1093/japr/5.2.134
10. Coufal CD, Chavez C, Knape KD, Carey JB. Evaluation of a method of ultraviolet light sanitation of broiler hatching eggs. Poultry Sci (2003) 82(5):754–9. doi: 10.1093/ps/82.5.754
11. Ekperigin HE, McCapes RH, Redus R, Ritchie WL, Cameron WJ, Nagaraja KV, et al. Research note: microcidal effects of a new pelleting process. Poultry Sci (1990) 69(9):1595–8. doi: 10.3382/ps.0691595
12. Dhillon AS, Jack OK. Two outbreaks of colibacillosis in commercial caged layers. Avian Dis (1996) 40(3):742–6. doi: 10.2307/1592290
13. Davis M, Morishita TY. Relative ammonia concentrations, dust concentrations, and presence of Salmonella species and Escherichia coli inside and outside commercial layer facilities. Avian Dis (2005) 49(1):30–5. doi: 10.1637/0005-2086(2005)49[30:RACDCA]2.0.CO;2
14. Roth N, Käsbohrer A, Mayrhofer S, Zitz U, Hofacre C, Domig KJ. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: a global overview. Poult Sci (2019) 98(4):1791–804. doi: 10.3382/ps/pey539
15. Mund MD, Khan UH, Tahir U, Mustafa B-E, Fayyaz A. Antimicrobial drug residues in poultry products and implications on public health: a review. Int J Food Properties (2017) 20(7):1433–46. doi: 10.1080/10942912.2016.1212874
16. Butaye P, Devriese LA, Haesebrouck F. Antimicrobial growth promoters used in animal feed: effects of less well known antibiotics on gram-positive bacteria. Clin Microbiol Rev (2003) 16(2):175–88. doi: 10.1128/cmr.16.2.175-188.2003
17. Castanon JIR. History of the use of antibiotic as growth promoters in European poultry feeds. Poultry Sci (2007) 86(11):2466–71. doi: 10.3382/ps.2007-00249
18. More SJ. European Perspectives on efforts to reduce antimicrobial usage in food animal production. Irish veterinary J (2020) 73:2–2. doi: 10.1186/s13620-019-0154-4
19. Scott HM, Acuff G, Bergeron G, Bourassa MW, Gill J, Graham DW, et al. Critically important antibiotics: criteria and approaches for measuring and reducing their use in food animal agriculture. Ann New York Acad Sci (2019) 1441(1):8–16. doi: 10.1111/nyas.14058
20. Founou LL, Founou RC, Essack SY. Antibiotic resistance in the food chain: a developing country-perspective. Front Microbiol (2016) 7. doi: 10.3389/fmicb.2016.01881
21. Prestinaci F, Pezzotti P, Pantosti A. Antimicrobial resistance: a global multifaceted phenomenon. Pathog Global Health (2015) 109(7):309–18. doi: 10.1179/2047773215y.0000000030
22. Dadgostar P. Antimicrobial resistance: implications and costs. Infect Drug Resist (2019) 12:3903–10. doi: 10.2147/idr.s234610
23. O'Neill J. Tackling drug-resistant infections globally: final report and recommendations. London, UK: Wellcome Trust and HM Government. (2016).
24. Ravikumar R, Chan J, Prabakaran M. Vaccines against major poultry viral diseases: strategies to improve the breadth and protective efficacy. Viruses (2022) 14(6):1195. doi: 10.3390/v14061195
25. Redweik GAJ, Jochum J, Mellata M. Live bacterial prophylactics in modern poultry. Front Veterinary Sci (2020) 7. doi: 10.3389/fvets.2020.592312
26. Grein K, Jungbäck C, Kubiak V. Autogenous vaccines: quality of production and movement in a common market. Biologicals (2022) 76:36–41. doi: 10.1016/j.biologicals.2022.01.003
27. Rabie NS, Amin Girh ZMS. Bacterial vaccines in poultry. Bull Natl Res Centre (2020) 44(1):15. doi: 10.1186/s42269-019-0260-1
28. Dimitrov KM, Afonso CL, Yu Q, Miller PJ. Newcastle Disease vaccines-a solved problem or a continuous challenge? Vet Microbiol (2017) 206:126–36. doi: 10.1016/j.vetmic.2016.12.019
29. Marangon S, Busani L. The use of vaccination in poultry production. Rev Sci Tech (2007) 26(1):265–74. doi: 10.20506/rst.26.1.1742
30. Żbikowska K, M. Michalczuk and B. Dolka: the use of bacteriophages in the poultry industry. Animals (2020) 10(5):872. doi: 10.3390/ani10050872
31. Patterson JA, Burkholder KM. Application of prebiotics and probiotics in poultry production. Poult Sci (2003) 82(4):627–31. doi: 10.1093/ps/82.4.627
32. Al-Mnaser A, Dakheel M, Alkandari F, Woodward M. Polyphenolic phytochemicals as natural feed additives to control bacterial pathogens in the chicken gut. Arch Microbiol (2022) 204(5):253. doi: 10.1007/s00203-022-02862-5
33. Ricke SC. Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult Sci (2003) 82(4):632–9. doi: 10.1093/ps/82.4.632
34. Wigley P. Immunology of birds. In: eLS Chichester, UK:John Wiley and Sons, Ltd (Ed.). (2017). doi: 10.102/9780470015902.a0026259
35. Kaiser P. Advances in avian immunology–prospects for disease control: a review. Avian Pathol (2010) 39(5):309–24. doi: 10.1080/03079457.2010.508777
36. Qureshi M. Avian macrophage: effector functions in health and disease. Dev Comp Immunol (2000) 24(2-3):103–19. doi: 10.1016/s0145-305x(99)00067-1
37. Sabet T, Hsia W-C, Stanisz M, El-Domeiri A, Van Alten P. A simple method for obtaining peritoneal macrophages from chickens. J Immunol Methods (1977) 14(2):103–10. doi: 10.1016/0022-1759(77)90001-1
38. Mutua MP, Muya S, Gicheru MM. Protective roles of free avian respiratory macrophages in captive birds. Biol Res (2016) 49(1):29. doi: 10.1186/s40659-016-0090-7
39. Toth TE, Siegel PB. Cellular defense of the avian respiratory system: dose-response relationship and duration of response in intratracheal stimulation of avian respiratory phagocytes by a Pasteurella multocida bacterin. Avian Dis (1993) 37(3):756–62. doi: 10.2307/1592025
40. Toth TE, Veit H, Gross WB, Siegel PB. Cellular defense of the avian respiratory system: protection against Escherichia coli airsacculitis by Pasteurella multocida-activated respiratory phagocytes. Avian Dis (1988) 32(4):681–7. doi: 10.2307/1590985
41. Chaplin DD. Overview of the immune response. J Allergy Clin Immunol (2010) 125(2 Suppl 2):S3–S23. doi: 10.1016/j.jaci.2009.12.980
42. Kawasaki T, Kawai T. Toll-like receptor signaling pathways. Front Immunol (2014) 5:461. doi: 10.3389/fimmu.2014.00461
43. Temperley ND, Berlin S, Paton IR, Griffin DK, Burt DW. Evolution of the chicken toll-like receptor gene family: a story of gene gain and gene loss. BMC Genomics (2008) 9(1):62. doi: 10.1186/1471-2164-9-62
44. Rehman MS-U, Rehman S. U., Yousaf W, Hassan F-u, Ahmad W, Liu Q, et al. The potential of toll-like receptors to modulate avian immune system: exploring the effects of genetic variants and phytonutrients. Front Genet (2021) 12. doi: 10.3389/fgene.2021.671235
45. Fellah JS, Jaffredo T, Nagy N, Dunon D. Chapter 3 - development of the avian immune system. In: Schat KA, Kaspers B, Kaiser P, editors. Avian immunology, 2nd ed. Boston: Academic Press (2014). doi: 10.1016/B978-0-12-396965-1.00003-0
46. Boodhoo N, Behboudi S. Marek’s disease virus-specific T cells proliferate, express antiviral cytokines but have impaired degranulation response. Front Immunol (2022) 13. doi: 10.3389/fimmu.2022.973762
47. Smith AL, Göbel TW. Chapter 5 - avian T cells: antigen recognition and lineages. In: Schat KA, Kaspers B, Kaiser P, editors. Avian immunology, 2nd ed. Boston: Academic Press (2014). doi: 10.1016/B978-0-12-396965-1.00005-4
48. Matsuyama-Kato A, Shojadoost B, Boodhoo N, Raj S, Alizadeh M, Fazel F, et al. Activated chicken gamma delta T cells are involved in protective immunity against marek’s disease. Viruses (2023) 15(2):285. doi: 10.3390/v15020285
49. Matsuyama-Kato A, Boodhoo N, Iseki H, Abdul-Careem MF, Plattner BL, Behboudi S, et al. Differential activation of chicken gamma delta T cells from different tissues by toll-like receptor 3 or 21 ligands. Dev Comp Immunol (2022) 131:104391. doi: 10.1016/j.dci.2022.104391
50. Matsuyama-Kato A, Iseki H, Boodhoo N, Bavananthasivam J, Alqazlan N, Abdul-Careem MF, et al. Phenotypic characterization of gamma delta (γδ) T cells in chickens infected with or vaccinated against marek's disease virus. Virology (2022) 568:115–25. doi: 10.1016/j.virol.2022.01.012
51. Boodhoo N, Behboudi S. Differential virus-specific IFN-gamma producing T cell responses to marek’s disease virus in chickens with B19 and B21 MHC haplotypes. Front Immunol (2022) 12. doi: 10.3389/fimmu.2021.784359
52. Boodhoo N, Gurung A, Sharif S, Behboudi S. Marek’s disease in chickens: a review with focus on immunology. Veterinary Res (2016) 47(1):119. doi: 10.1186/s13567-016-0404-3
53. Ratcliffe MJH, Härtle S. Chapter 4 - b cells, the bursa of fabricius and the generation of antibody repertoires. In: Schat KA, Kaspers B, Kaiser P, editors. Avian immunology, 2nd ed. Boston: Academic Press (2014). doi: 10.1016/B978-0-12-396965-1.00004-2
54. McCormack WT, Tjoelker LW, Thompson CB. Avian b-cell development: generation of an immunoglobulin repertoire by gene conversion. Annu Rev Immunol (1991) 9(1):219–41. doi: 10.1146/annurev.iy.09.040191.001251
55. Sadeyen J-R, Wu Z, Davies H, van Diemen PM, Milicic A, La Ragione RM, et al. Immune responses associated with homologous protection conferred by commercial vaccines for control of avian pathogenic Escherichia coli in turkeys. Veterinary Res (2015) 46(1):5. doi: 10.1186/s13567-014-0132-5
56. Buchmann K. Evolution of innate immunity: clues from invertebrates via fish to mammals. Front Immunol (2014) 5:459. doi: 10.3389/fimmu.2014.00459
57. Flajnik MF, Kasahara M. Origin and evolution of the adaptive immune system: genetic events and selective pressures. Nat Rev Genet (2010) 11(1):47–59. doi: 10.1038/nrg2703
58. Dubuffet A, Zanchi C, Boutet G, Moreau J, Teixeira M, Moret Y. Trans-generational immune priming protects the eggs only against gram-positive bacteria in the mealworm beetle. PloS Pathog (2015) 11(10):e1005178. doi: 10.1371/journal.ppat.1005178
60. Loker ES, Adema CM, Zhang S-M, Kepler TB. Invertebrate immune systems–not homogeneous, not simple, not well understood. Immunol Rev (2004) 198:10–24. doi: 10.1111/j.0105-2896.2004.0117.x
61. Melillo D, Marino R, Italiani P, Boraschi D. Innate immune memory in invertebrate metazoans: a critical appraisal. Front Immunol (2018) 9. doi: 10.3389/fimmu.2018.01915
62. Sánchez-Ramón S, Conejero L, Netea MG, Sancho D, Palomares Ó, Subiza JL. Trained immunity-based vaccines: a new paradigm for the development of broad-spectrum anti-infectious formulations. Front Immunol (2018) 9:2936(2936). doi: 10.3389/fimmu.2018.02936
63. Netea MG, Domínguez-Andrés J, Barreiro LB, Chavakis T, Divangahi M, Fuchs E, et al. Defining trained immunity and its role in health and disease. Nat Rev Immunol (2020) 20(6):375–88. doi: 10.1038/s41577-020-0285-6
64. Verwoolde MB, van den Biggelaar RHGA, van Baal J, Jansen CA, Lammers A. Training of primary chicken monocytes results in enhanced pro-inflammatory responses. Veterinary Sci (2020) 7(3):115. doi: 10.3390/vetsci7030115
65. Goonewardene KB, Karu N, Ahmed KA, Popowich S, Chow-Lockerbie B, Ayalew LE, et al. CpG-ODN induced antimicrobial immunity in neonatal chicks involves a substantial shift in serum metabolic profiles. Sci Rep (2021) 11(1):9028. doi: 10.1038/s41598-021-88386-2
66. Divangahi M, Aaby P, Khader SA, Barreiro LB, Bekkering S, Chavakis T, et al. Trained immunity, tolerance, priming and differentiation: distinct immunological processes. Nat Immunol (2021) 22(1):2–6. doi: 10.1038/s41590-020-00845-6
67. Dominguez-Andres J, Netea MG. Long-term reprogramming of the innate immune system. J Leukocyte Biol (2019) 105(2):329–38. doi: 10.1002/jlb.mr0318-104r
68. Cheled-Shoval SL, Amit-Romach E, Barbakov M, Uni Z. The effect of in ovo administration of mannan oligosaccharide on small intestine development during the pre- and posthatch periods in chickens. Poultry Sci (2011) 90(10):2301–10. doi: 10.3382/ps.2011-01488
69. Pourabedin M, Xu Z, Baurhoo B, Chevaux E, Zhao X. Effects of mannan oligosaccharide and virginiamycin on the cecal microbial community and intestinal morphology of chickens raised under suboptimal conditions. Can J Microbiol (2014) 60(5):255–66. doi: 10.1139/cjm-2013-0899
70. Chee SH, Iji PA, Choct M, Mikkelsen LL, Kocher A. Characterisation and response of intestinal microflora and mucins to manno-oligosaccharide and antibiotic supplementation in broiler chickens. Br Poultry Sci (2010) 51(3):368–80. doi: 10.1080/00071668.2010.503477
71. Yitbarek A, Echeverry H, Brady J, Hernandez-Doria J, Camelo-Jaimes G, Sharif S, et al. Innate immune response to yeast-derived carbohydrates in broiler chickens fed organic diets and challenged with clostridium perfringens. Poult Sci (2012) 91(5):1105–12. doi: 10.3382/ps.2011-02109
72. Ibuki M, Kovacs-Nolan J, Fukui K, Kanatani H, Mine Y. Analysis of gut immune-modulating activity of β-1,4-mannobiose using microarray and real-time reverse transcription polymerase chain reaction. Poultry Sci (2010) 89(9):1894–904. doi: 10.3382/ps.2010-00791
73. Stefaniak T, Madej JP, Graczyk S, Siwek M, Łukaszewicz E, Kowalczyk A, et al. Selected prebiotics and synbiotics administered in ovo can modify innate immunity in chicken broilers. BMC Veterinary Res (2019) 15(1):105. doi: 10.1186/s12917-019-1850-8
74. Xie S, Zhao S, Jiang L, Lu L, Yang Q, Yu Q. Lactobacillus reuteri stimulates intestinal epithelial proliferation and induces differentiation into goblet cells in young chickens. J Agric Food Chem (2019) 67(49):13758–66. doi: 10.1021/acs.jafc.9b06256
75. Nii T, Jirapat J, Isobe N, Yoshimura Y. Effects of oral administration of Lactobacillus reuteri on mucosal barrier function in the digestive tract of broiler chicks. J Poultry Sci (2020) 57(1):67–76. doi: 10.2141/jpsa.0190035
76. Wu Y, Zhen W, Geng Y, Wang Z, Guo Y. Pretreatment with probiotic Enterococcus faecium NCIMB 11181 ameliorates necrotic enteritis-induced intestinal barrier injury in broiler chickens. Sci Rep (2019) 9(1):10256. doi: 10.1038/s41598-019-46578-x
77. Wang B, Hussain A, Zhou Y, Zeng Z, Wang Q, Zou P, et al. Saccharomyces boulardii attenuates inflammatory response induced by clostridium perfringens via TLR4/TLR15-MyD8 pathway in HD11 avian macrophages. Poultry Sci (2020) 99(11):5356–65. doi: 10.1016/j.psj.2020.07.045
78. Lee K-W, Kim DK, Lillehoj HS, Jang SI, Lee S-H. Immune modulation by Bacillus subtilis-based direct-fed microbials in commercial broiler chickens. Anim Feed Sci Technol (2015) 200:76–85. doi: 10.1016/j.anifeedsci.2014.12.006
79. Meijerink N, Kers JG, Velkers FC, van Haarlem DA, Lamot DM, de Oliveira JE, et al. Early life inoculation with adult-derived microbiota accelerates maturation of intestinal microbiota and enhances NK cell activation in broiler chickens. Front Veterinary Sci (2020) 7. doi: 10.3389/fvets.2020.584561
80. Zhang B, Guo Y, Wang Z. The modulating effect of β-1, 3/1, 6-glucan supplementation in the diet on performance and immunological responses of broiler chickens. Asian-Australas J Anim Sci (2008) 21(2):237–44. doi: 10.5713/ajas.2008.70207
81. Huff GR, Huff WE, Jalukar S, Oppy J, Rath NC, Packialakshmi B. The effects of yeast feed supplementation on turkey performance and pathogen colonization in a transport stress/Escherichia coli challenge . Poultry Sci. (2013) 92(3):655–62. doi: 10.3382/ps.2012-02787
82. Shao Y, Guo Y, Wang Z. β-1,3/1,6-Glucan alleviated intestinal mucosal barrier impairment of broiler chickens challenged with Salmonella enterica serovar typhimurium. Poultry Sci (2013) 92(7):1764–73. doi: 10.3382/ps.2013-03029
83. Lowry VK, Farnell MB, Ferro PJ, Swaggerty CL, Bahl A, Kogut MH. Purified β-glucan as an abiotic feed additive up-regulates the innate immune response in immature chickens against Salmonella enterica serovar enteritidis. Int J Food Microbiol (2005) 98(3):309–18. doi: 10.1016/j.ijfoodmicro.2004.06.008
84. Huff GR, Huff WE, Farnell MB, Rath NC, Solis de los Santos F, Donoghue AM. Bacterial clearance, heterophil function, and hematological parameters of transport-stressed turkey poults supplemented with dietary yeast extract1. Poultry Sci (2010) 89(3):447–56. doi: 10.3382/ps.2009-00328
85. Guo Y, Ali RA, Qureshi MA. The influence of beta-glucan on immune responses in broiler chicks. Immunopharmacol Immunotoxicol (2003) 25(3):461–72. doi: 10.1081/iph-120024513
86. Zhuge X, Sun Y, Jiang M, Wang J, Tang F, Xue F, et al. Acetate metabolic requirement of avian pathogenic Escherichia coli promotes its intracellular proliferation within macrophage. Vet Res (2019) 50(31). doi: 10.1186/s13567-019-0650-2
87. Gomis S, Babiuk L, Godson DL, Allan B, Thrush T, Townsend H, et al. Protection of chickens against Escherichia coli infections by DNA containing CpG motifs. Infect Immun (2003) 71(2):857–63. doi: 10.1128/iai.71.2.857-863.2003
88. Gomis S, Babiuk L, Allan B, Willson P, Waters E, Ambrose N, et al. Protection of neonatal chicks against a lethal challenge of Escherichia coli using DNA containing cytosine-Phosphodiester-Guanine motifs. Avian Dis (2004) 48(4):813–22. doi: 10.1637/7194-041204R
89. Taghavi A, Allan B, Mutwiri G, Van Kessel A, Willson P, Babiuk L, et al. Protection of neonatal broiler chicks against Salmonella typhimurium septicemia by DNA containing CpG motifs. Avian Dis (2008) 52(3):398–406. doi: 10.1637/8196-121907-Reg
90. He H, Genovese KJ, Swaggerty CL, Nisbet DJ, Kogut MH. In vivo priming heterophil innate immune functions and increasing resistance to Salmonella enteritidis infection in neonatal chickens by immune stimulatory CpG oligodeoxynucleotides. Veterinary Immunol Immunopathol (2007) 117(3):275–83. doi: 10.1016/j.vetimm.2007.03.002
91. Kalhari Bandara G, Shelly P, Thushari G, Ashish G, Shanika K, Ruwani K, et al. Intrapulmonary delivery of CpG-ODN microdroplets provides protection against Escherichia coli septicemia in neonatal broiler chickens. Avian Dis (2017) 61(4):503–11. doi: 10.1637/11684-060617-Reg.1
92. Gunawardana T, Foldvari M, Zachar T, Popowich S, Chow-Lockerbie B, Ivanova MV, et al. Protection of neonatal broiler chickens following in ovo delivery of oligodeoxynucleotides containing CpG motifs (CpG-ODN) formulated with carbon nanotubes or liposomes. Avian Dis (2015) 59(1):31–7. doi: 10.1637/10832-032814-Reg
93. Brenda A, Colette W, Wolfgang K, Mishal S, Andy P, Volker G, et al. In ovo administration of innate immune stimulants and protection from early chick mortalities due to yolk sac infection. Avian Dis (2018) 62(3):316–21. doi: 10.1637/11840-041218-Reg.1
94. Komori T, Saito K, Sawa N, Shibasaki Y, Kohchi C, Soma G, et al. Innate immunity activated by oral administration of LPSp is phylogenetically preserved and developed in broiler chickens. Anticancer Res (2015) 35(8):4461–6.
95. Hebishima T, Matsumoto Y, Watanabe GEN, Soma G-I, Kohchi C, Taya K, et al. Recovery from immunosuppression-related disorders in humans and animals by IP-PA1, an edible lipopolysaccharide. Anticancer Res (2010) 30(8):3113.
96. Bansal M, Alenezi T, Fu Y, Almansour A, Wang H, Gupta A, et al. Specific secondary bile acids control chicken necrotic enteritis. Pathogens (2021) 10(8):1041. doi: 10.3390/pathogens10081041
97. Wang H, Latorre JD, Bansal M, Abraha M, Al-Rubaye B, Tellez-Isaias G, et al. Microbial metabolite deoxycholic acid controls clostridium perfringens-induced chicken necrotic enteritis through attenuating inflammatory cyclooxygenase signaling. Sci Rep (2019) 9(1):14541. doi: 10.1038/s41598-019-51104-0
98. Alrubaye B, Abraha M, Almansour A, Bansal M, Wang H, Kwon YM, et al. Microbial metabolite deoxycholic acid shapes microbiota against campylobacter jejuni chicken colonization. PloS One (2019) 14(7):e0214705. doi: 10.1371/journal.pone.0214705
99. Sun X, Winglee K, Gharaibeh RZ, Gauthier J, He Z, Tripathi P, et al. Microbiota-derived metabolic factors reduce campylobacteriosis in mice. Gastroenterology (2018) 154(6):1751–1763.e2. doi: 10.1053/j.gastro.2018.01.042
100. Sunkara LT, Achanta M, Schreiber NB, Bommineni YR, Dai G, Jiang W, et al. Butyrate enhances disease resistance of chickens by inducing antimicrobial host defense peptide gene expression. PloS One (2011) 6(11):e27225. doi: 10.1371/journal.pone.0027225
101. Gupta A, Bansal M, Wagle B, Sun X, Rath N, Donoghue A, et al. Sodium butyrate reduces Salmonella enteritidis infection of chicken enterocytes and expression of inflammatory host genes in vitro. Front Microbiol (2020) 11. doi: 10.3389/fmicb.2020.553670
102. Bortoluzzi C, Rothrock MJ, Vieira BS, Mallo JJ, Puyalto M, Hofacre C, et al. Supplementation of protected sodium butyrate alone or in combination with essential oils modulated the cecal microbiota of broiler chickens challenged with coccidia and clostridium perfringens. Front Sustain Food Syst (2018) 2. doi: 10.3389/fsufs.2018.00072
103. Sunkara LT, Jiang W, Zhang G. Modulation of antimicrobial host defense peptide gene expression by free fatty acids. PloS One (2012) 7(11):e49558. doi: 10.1371/journal.pone.0049558
104. Geng Y, Ma Q, Wang Z, Guo Y. Dietary vitamin D3 supplementation protects laying hens against lipopolysaccharide-induced immunological stress. Nutr Metab (2018) 15:58. doi: 10.1186/s12986-018-0293-8
105. Morris A, Shanmugasundaram R, Lilburn MS, Selvaraj RK. 25-hydroxycholecalciferol supplementation improves growth performance and decreases inflammation during an experimental lipopolysaccharide injection. Poult Sci (2014) 93(8):1951–6. doi: 10.3382/ps.2014-03939
106. Shojadoost B, Behboudi S, Villanueva AI, Brisbin JT, Ashkar AA, Sharif S. Vitamin D3 modulates the function of chicken macrophages. Res Veterinary Sci (2015) 100:45–51. doi: 10.1016/j.rvsc.2015.03.009
107. Boodhoo N, Sharif S, Behboudi S. 1α,25(OH)2 vitamin D3 modulates avian T lymphocyte functions without inducing CTL unresponsiveness. PloS One (2016) 11(2):e0150134. doi: 10.1371/journal.pone.0150134
108. Chou P-C, Lin P-C, Wu S-W, Wang C-K, Chung T-K, Walzem RL, et al. Differential modulation of 25-hydroxycholecalciferol on innate immunity of broiler breeder hens. Animals (Basel) (2021) 11(6):1742. doi: 10.3390/ani11061742
109. Fan X, Liu S, Liu G, Zhao J, Jiao H, Wang X, et al. Vitamin a deficiency impairs mucin expression and suppresses the mucosal immune function of the respiratory tract in chicks. PloS One (2015) 10(9):e0139131. doi: 10.1371/journal.pone.0139131
110. Uni Z, Zaiger G, Gal-Garber O, Pines M, Rozenboim I, Reifen R. Vitamin a deficiency interferes with proliferation and maturation of cells in the chicken small intestine. Br Poultry Sci (2000) 41(4):410–5. doi: 10.1080/713654958
111. Aye PP, Morishita TY, Saif YM, Latshaw JD, Harr BS, Cihla FB. Induction of vitamin a deficiency in turkeys. Avian Dis (2000) 44(4):809–17. doi: 10.2307/1593053
112. Raza A, Khan SA, Raza FK, Saeed MA, Bashir IN. Effects of vitamin-a on growth traits, immunoregulatory organs and immune response in broiler chicken. J Appl Anim Res (1997) 12(1):81–8. doi: 10.1080/09712119.1997.9706189
113. Alizadeh M, Astill J, Alqazlan N, Shojadoost B, Taha-Abdelaziz K, Bavananthasivam J, et al. In ovo co-administration of vitamins (A and d) and probiotic lactobacilli modulates immune responses in broiler chickens. Poultry Sci (2022) 101(4):101717. doi: 10.1016/j.psj.2022.101717
114. Shojadoost B, Alizadeh M, Taha-Abdelaziz K, Shoja Doost J, Astill J, Sharif S. In ovo inoculation of vitamin a modulates chicken embryo immune functions. J Interferon Cytokine Res (2021) 41(1):20–8. doi: 10.1089/jir.2020.0212
115. Kang BY, Chung SW, Kim SH, Kang SN, Choe YK, Kim TS. Retinoid-mediated inhibition of interleukin-12 production in mouse macrophages suppresses Th1 cytokine profile in CD4+T cells. Br J Pharmacol (2000) 130(3):581–6. doi: 10.1038/sj.bjp.0703345
116. Konjufca VK, Bottje WG, Bersi TK, Erf GF. Influence of dietary vitamin e on phagocytic functions of macrophages in broilers. Poultry Sci (2004) 83(9):1530–4. doi: 10.1093/ps/83.9.1530
117. Huff GR, Huff WE, Balog JM, Rath NC, Izard RS. The effects of water supplementation with vitamin e and sodium salicylate (Uni-sol®) on the resistance of turkeys to Escherichia coli respiratory infection. Avian Dis (2004) 48(2):324–31. doi: 10.1637/7112
118. Liu YJ, Zhao LH, Mosenthin R, Zhang JY, Ji C, Ma QG. Protective effect of vitamin e on laying performance, antioxidant capacity, and immunity in laying hens challenged with Salmonella enteritidis. Poultry Sci (2019) 98(11):5847–54. doi: 10.3382/ps/pez227
119. Carr A, Maggini S. Vitamin c and immune function. Nutrients (2017) 9(11):1211. doi: 10.3390/nu9111211
120. Andreasen CB, Frank DE. The effects of ascorbic acid on in vitro heterophil function. Avian Dis (1999) 43(4):656–63. doi: 10.2307/1592734
121. Gan L, Fan H, Mahmood T, Guo Y. Dietary supplementation with vitamin c ameliorates the adverse effects of Salmonella enteritidis-challenge in broilers by shaping intestinal microbiota. Poult Sci (2020) 99(7):3663–74. doi: 10.1016/j.psj.2020.03.062
122. Hernandez-Patlan D, Solis-Cruz B, Pontin KP, Latorre JD, Hernandez-Velasco X, Merino-Guzman R, et al. Evaluation of ascorbic acid or curcumin formulated in a solid dispersion on Salmonella enteritidis infection and intestinal integrity in broiler chickens. Pathogens (2019) 8(4):229. doi: 10.3390/pathogens8040229
123. Kim DK, Lillehoj HS, Lee SH, Jang SI, Bravo D. High-throughput gene expression analysis of intestinal intraepithelial lymphocytes after oral feeding of carvacrol, cinnamaldehyde, or capsicum oleoresin. Poult Sci (2010) 89(1):68–81. doi: 10.3382/ps.2009-00275
124. Placha I, Takacova J, Ryzner M, Cobanova K, Laukova A, Strompfova V, et al. Effect of thyme essential oil and selenium on intestine integrity and antioxidant status of broilers. Br Poultry Sci (2014) 55(1):105–14. doi: 10.1080/00071668.2013.873772
125. Islam MR, Oomah DB, Diarra MS. Potential immunomodulatory effects of non-dialyzable materials of cranberry extract in poultry production. Poultry Sci (2017) 96(2):341–50. doi: 10.3382/ps/pew302
126. Dorhoi A, Dobrean V, Zăhan M, Virag P. Modulatory effects of several herbal extracts on avian peripheral blood cell immune responses. Phytother Res (2006) 20(5):352–8. doi: 10.1002/ptr.1859
127. Lee SH, Lillehoj HS, Hong YH, Jang SI, Lillehoj EP, Ionescu C, et al. In vitro effects of plant and mushroom extracts on immunological function of chicken lymphocytes and macrophages. Br Poultry Sci (2010) 51(2):213–21. doi: 10.1080/00071661003745844
128. Lee SH, Lillehoj HS, Jang SI, Lee KW, Park MS, Bravo D, et al. Cinnamaldehyde enhances in vitro parameters of immunity and reduces in vivo infection against avian coccidiosis. Br J Nutr (2011) 106(6):862–9. doi: 10.1017/s0007114511001073
129. Kamboh AA, Hang SQ, Khan MA, Zhu WY. In vivo immunomodulatory effects of plant flavonoids in lipopolysaccharide-challenged broilers. Animal (2016) 10(10):1619–25. doi: 10.1017/S1751731116000562
130. Gibson GR, Roberfroid MB. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J Nutr (1995) 125(6):1401–12. doi: 10.1093/jn/125.6.1401
131. Ricke SC. Impact of prebiotics on poultry production and food safety. Yale J Biol Med (2018) 91(2):151–9.
132. Takiishi T, Fenero CIM, Câmara NOS. Intestinal barrier and gut microbiota: shaping our immune responses throughout life. Tissue Barriers (2017) 5(4):e1373208. doi: 10.1080/21688370.2017.1373208
133. Gadde U, Kim WH, Oh ST, Lillehoj HS. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: a review. Anim Health Res Rev (2017) 18(1):26–45. doi: 10.1017/s1466252316000207
134. Yan F, Polk DB. Probiotics and immune health. Curr Opin Gastroenterol (2011) 27(6):496–501. doi: 10.1097/mog.0b013e32834baa4d
135. Kabir SML. The role of probiotics in the poultry industry. Int J Mol Sci (2009) 10(8):3531–46. doi: 10.3390/ijms10083531
136. Sakaridis I, Ellis RJ, Cawthraw SA, van Vliet AHM, Stekel DJ, Penell J, et al. Investigating the association between the caecal microbiomes of broilers and campylobacter burden. Front Microbiol (2018) 9. doi: 10.3389/fmicb.2018.00927
137. Dang AT, Marsland BJ. Microbes, metabolites, and the gut–lung axis. Mucosal Immunol (2019) 12(4):843–50. doi: 10.1038/s41385-019-0160-6
138. Synodinou KD, Nikolaki MD, Triantafyllou K, Kasti AN. Immunomodulatory effects of probiotics on COVID-19 infection by targeting the gut–lung axis microbial cross-talk. Microorganisms (2022) 10(9):1764. doi: 10.3390/microorganisms10091764
139. Harris V, Ali A, Fuentes S, Korpela K, Kazi M, Tate J, et al. Rotavirus vaccine response correlates with the infant gut microbiota composition in Pakistan. Gut Microbes (2018) 9(2):93–101. doi: 10.1080/19490976.2017.1376162
140. Jordan A, Carding SR, Hall LJ. The early-life gut microbiome and vaccine efficacy. Lancet Microbe (2022) 3(10):e787–94. doi: 10.1016/s2666-5247(22)00185-9
141. Yitbarek A, Astill J, Hodgins DC, Parkinson J, Nagy É, Sharif S. Commensal gut microbiota can modulate adaptive immune responses in chickens vaccinated with whole inactivated avian influenza virus subtype H9N2. Vaccine (2019) 37(44):6640–7. doi: 10.1016/j.vaccine.2019.09.046
142. Kim HS, Hong JT, Kim Y, Han SB. Stimulatory effect of β-glucans on immune cells. Immune Netw (2011) 11(4):191–5. doi: 10.4110/in.2011.11.4.191
143. Park J-H, Lee S-I, Kim I-H. Effect of dietary β-glucan supplementation on growth performance, nutrient digestibility, and characteristics of feces in weaned pigs. J Appl Anim Res (2018) 46(1):1193–7. doi: 10.1080/09712119.2018.1481855
144. Ma T, Tu Y, Zhang N-f, Guo J-p, Deng K-d, Zhou Y, et al. Effects of dietary yeast β-glucan on nutrient digestibility and serum profiles in pre-ruminant Holstein calves. J Integr Agric (2015) 14(4):749–57. doi: 10.1016/S2095-3119(14)60843-1
145. Dawood MAO, Koshio S, Ishikawa M, Yokoyama S, El Basuini MF, Hossain MS, et al. Dietary supplementation of β-glucan improves growth performance, the innate immune response and stress resistance of red sea bream, pagrus major. Aquaculture Nutr (2017) 23(1):148–59. doi: 10.1111/anu.12376
146. Ding B, Zheng J, Wang X, Zhang L, Sun D, Xing Q, et al. Effects of dietary yeast beta-1,3-1,6-glucan on growth performance, intestinal morphology and chosen immunity parameters changes in haidong chicks. Asian-Australas J Anim Sci (2019) 32(10):1558–64. doi: 10.5713/ajas.18.0962
147. Moon SH, Lee I, Feng X, Lee HY, Kim J, Ahn DU. Effect of dietary beta-glucan on the performance of broilers and the quality of broiler breast meat. Asian-Australas J Anim Sci (2016) 29(3):384–9. doi: 10.5713/ajas.15.0141
148. Chen L, Jiang T, Li X, Wang Q, Wang Y, Li Y. Immunomodulatory activity of beta-glucan and mannan-oligosaccharides from Saccharomyces cerevisiae on broiler chickens challenged with feed-borne aspergillus fumigatus. Pakistan Veterinary J (2016) 36:297–301.
149. Santin E, Maiorka A, Macari M, Grecco M, Sanchez JC, Okada TM, et al. Performance and intestinal mucosa development of broiler chickens fed diets containing saccharomyces cerevisiae cell wall. J Appl Poultry Res (2001) 10(3):236–44. doi: 10.1093/japr/10.3.236
150. Gao J, Zhang HJ, Yu SH, Wu SG, Yoon I, Quigley J, et al. Effects of yeast culture in broiler diets on performance and immunomodulatory functions. Poultry Sci (2008) 87(7):1377–84. doi: 10.3382/ps.2007-00418
151. Vancamelbeke M, Vermeire S. The intestinal barrier: a fundamental role in health and disease. Expert Rev Gastroenterol Hepatol (2017) 11(9):821–34. doi: 10.1080/17474124.2017.1343143
152. Brown GD, Gordon S. A new receptor for β-glucans. Nature (2001) 413(6851):36–7. doi: 10.1038/35092620
153. Rodrigues MV, Zanuzzo FS, Koch JFA, De Oliveira CAF, Sima P, Vetvicka V. Development of fish immunity and the role of β-glucan in immune responses. Molecules (2020) 25(22):5378. doi: 10.3390/molecules25225378
154. Gantner BN, Simmons RM, Canavera SJ, Akira S, Underhill DM. Collaborative induction of inflammatory responses by dectin-1 and toll-like receptor 2. J Exp Med (2003) 197(9):1107–17. doi: 10.1084/jem.20021787
155. Verwoolde MB, van den Biggelaar RHGA, de Vries Reilingh G, Arts JAJ, van Baal J, Lammers A, et al. Innate immune training and metabolic reprogramming in primary monocytes of broiler and laying hens. Dev Comp Immunol (2021) 114:103811. doi: 10.1016/j.dci.2020.103811
156. Santecchia I, Vernel-Pauillac F, Rasid O, Quintin J, Gomes-Solecki M, Boneca IG, et al. Innate immune memory through TLR2 and NOD2 contributes to the control of leptospira interrogans infection. PloS Pathog (2019) 15(5):e1007811–e1007811. doi: 10.1371/journal.ppat.1007811
157. Salem AK, Weiner GJ. CpG oligonucleotides as immunotherapeutic adjuvants: innovative applications and delivery strategies. Advanced Drug Delivery Rev (2009) 61(3):193–4. doi: 10.1016/j.addr.2008.12.003
158. Sherbet GV. Chapter 26 - notable approaches to cancer immunotherapy. In: Sherbet GV, editor. Molecular approach to cancer management. Cambridge, Massachusetts, US: Academic Press (2017). doi: 10.1016/B978-0-12-812896-1.00026-X
159. Shirota H, Klinman DM. Chapter 9 - CpG oligodeoxynucleotides as adjuvants for clinical use. In: Schijns VEJC, O'Hagan DT, editors. Immunopotentiators in modern vaccines, 2nd ed. Cambridge, Massachusetts, US: Academic Press (2017). doi: 10.1016/B978-0-12-804019-5.00009-8
160. Krieg AM. Chapter 53 - CpG oligodeoxynucleotides for mucosal vaccines. In: Mestecky J, Lamm ME, McGhee JR, Bienenstock J, Mayer L, Strober W, editors. Mucosal immunology, 3rd ed. Burlington: Academic Press (2005). doi: 10.1016/B978-012491543-5/50057-7
161. Ferreira AV, Domiguéz-Andrés J, Netea MG. The role of cell metabolism in innate immune memory. J Innate Immun (2022) 14(1):42–50. doi: 10.1159/000512280
162. Ribes S, Meister T, Ott M, Redlich S, Janova H, Hanisch UK, et al. Intraperitoneal prophylaxis with CpG oligodeoxynucleotides protects neutropenic mice against intracerebral Escherichia coli K1 infection. J Neuroinflamm (2014) 11:14. doi: 10.1186/1742-2094-11-14
163. Rosadini CV, Kagan JC. Early innate immune responses to bacterial LPS. Curr Opin Immunol (2017) 44:14–9. doi: 10.1016/j.coi.2016.10.005
164. Kohchi C, Inagawa H, Nishizawa T, Yamaguchi T, Nagai S, Soma G-I. Applications of lipopolysaccharide derived from pantoea agglomerans (IP-PA1) for health care based on macrophage network theory. J Bioscience Bioengineering (2006) 102(6):485–96. doi: 10.1263/jbb.102.485
165. Takahashi Y, Kondo M, Itami T, Honda T, Inagawa H, Nishizawa T, et al. Enhancement of disease resistance against penaeid acute viraemia and induction of virus-inactivating activity in haemolymph of kuruma shrimp, Penaeus japonicus, by oral administration of pantoea agglomerans lipopolysaccharide (LPS). Fish Shellfish Immunol (2000) 10(6):555–8. doi: 10.1006/fsim.2000.0268
166. Kadowaki T, Yasui Y, Nishimiya O, Takahashi Y, Kohchi C, Soma G, et al. Orally administered LPS enhances head kidney macrophage activation with down-regulation of IL-6 in common carp (Cyprinus carpio). Fish Shellfish Immunol (2013) 34(6):1569–75. doi: 10.1016/j.fsi.2013.03.372
167. Chow J, Lee SM, Shen Y, Khosravi A, Mazmanian SK. Chapter 8 - host–bacterial symbiosis in health and disease. In: Fagarasan S, Cerutti A, editors. Advances in immunology. Academic Press (2010). doi: 10.1016/B978-0-12-381300-8.00008-3
168. Rodrigues DR, Wilson KM, Bielke LR. Proper immune response depends on early exposure to gut microbiota in broiler chicks. Front Physiol (2021) 12. doi: 10.3389/fphys.2021.758183
169. Levy M, Thaiss CA, Elinav E. Metabolites: messengers between the microbiota and the immune system. Genes Dev (2016) 30(14):1589–97. doi: 10.1101/gad.284091.116
170. Yeşilyurt N, Yılmaz B, Ağagündüz D, Capasso R. Involvement of probiotics and postbiotics in the immune system modulation. Biologics (2021) 1(2):89–110. doi: 10.3390/biologics1020006
171. Godlewska U, Bulanda E, Wypych TP. Bile acids in immunity: bidirectional mediators between the host and the microbiota. Front Immunol (2022) 13. doi: 10.3389/fimmu.2022.949033
172. Wu J, Wang K, Wang X, Pang Y, Jiang C. The role of the gut microbiome and its metabolites in metabolic diseases. Protein Cell (2021) 12(5):360–73. doi: 10.1007/s13238-020-00814-7
173. Weichhart T, Hengstschläger M, Linke M. Regulation of innate immune cell function by mTOR. Nat Rev Immunol (2015) 15(10):599–614. doi: 10.1038/nri3901
174. Schulthess J, Pandey S, Capitani M, Rue-Albrecht KC, Arnold I, Franchini F, et al. The short chain fatty acid butyrate imprints an antimicrobial program in macrophages. Immunity (2019) 50(2):432–445.e7. doi: 10.1016/j.immuni.2018.12.018
175. Langfeld LQ, Du K, Bereswill S, Heimesaat MM. A review of the antimicrobial and immune-modulatory properties of the gut microbiota-derived short chain fatty acid propionate - what is new? Eur J Microbiol Immunol (Bp) (2021) 11(2):50–6. doi: 10.1556/1886.2021.00005
176. Li H, Zhao L, Liu S, Zhang Z, Wang X, Lin H. Propionate inhibits fat deposition via affecting feed intake and modulating gut microbiota in broilers. Poultry Sci (2021) 100(1):235–45. doi: 10.1016/j.psj.2020.10.009
177. Van Hoeck V, Sonawane M, Gonzalez Sanchez AL, Van Dosselaer I, Buyens C, Morisset D. Chromium propionate improves performance and carcass traits in broilers. Anim Nutr (2020) 6(4):480–7. doi: 10.1016/j.aninu.2020.03.005
178. Jacobson A, Lam L, Rajendram M, Tamburini F, Honeycutt J, Pham T, et al. A gut commensal-produced metabolite mediates colonization resistance to Salmonella infection. Cell Host Microbe (2018) 24(2):296–307.e7. doi: 10.1016/j.chom.2018.07.002
179. Park JW, Kim HY, Kim MG, Jeong S, Yun CH, Han SH. Short-chain fatty acids inhibit staphylococcal lipoprotein-induced nitric oxide production in murine macrophages. Immune Netw (2019) 19(2):e9. doi: 10.4110/in.2019.19.e9
180. Jeong S, Kim HY, Kim AR, Yun CH, Han SH. Propionate ameliorates staphylococcus aureus skin infection by attenuating bacterial growth. Front Microbiol (2019) 10:1363. doi: 10.3389/fmicb.2019.01363
181. Mora JR, Iwata M, Von Andrian UH. Vitamin effects on the immune system: vitamins a and d take centre stage. Nat Rev Immunol (2008) 8(9):685–98. doi: 10.1038/nri2378
182. Ismailova A, White JH, Vitamin D. Infections and immunity. Rev Endocrine Metab Disord (2022) 23(2):265–77. doi: 10.1007/s11154-021-09679-5
183. Chou PC, Chen YH, Chung TK, Walzem RL, Lai LS, Chen SE. Supplemental 25-hydroxycholecalciferol alleviates inflammation and cardiac fibrosis in hens. Int J Mol Sci (2020) 21(21):8379. doi: 10.3390/ijms21218379
184. Lin H-Y, Chung TK, Chen Y-H, Walzem RL, Chen S-E. Dietary supplementation of 25-hydroxycholecalciferol improves livability in broiler breeder hens. Poultry Sci (2019) 98(11):6108–16. doi: 10.3382/ps/pez330
185. Vazquez JR, Gómez GV, López CC, Cortés AC, Díaz AC, Fernández SRT, et al. Effects of 25-hydroxycholecalciferol with two D3 vitamin levels on production and immunity parameters in broiler chickens. J Anim Physiol Anim Nutr (2018) 102(1):e493–7. doi: 10.1111/jpn.12715
186. Chou SH, Chung TK, Yu B. Effects of supplemental 25-hydroxycholecalciferol on growth performance, small intestinal morphology, and immune response of broiler chickens. Poultry Sci (2009) 88(11):2333–41. doi: 10.3382/ps.2009-00283
187. Broom LJ, Kogut MH. Inflammation: friend or foe for animal production? Poult Sci (2018) 97(2):510–4. doi: 10.3382/ps/pex314
188. Huang Z, Liu Y, Qi G, Brand D, Zheng SG. Role of vitamin a in the immune system. J Clin Med (2018) 7(9):258. doi: 10.3390/jcm7090258
189. Shojadoost B, Yitbarek A, Alizadeh M, Kulkarni RR, Astill J, Boodhoo N, et al. Centennial review: effects of vitamins a, d, e, and c on the chicken immune system. Poult Sci (2021) 100(4):100930. doi: 10.1016/j.psj.2020.12.027
190. Lessard M, Hutchings D, Cave NA. Cell-mediated and humoral immune responses in broiler chickens maintained on diets containing different levels of vitamin a. Poultry Sci (1997) 76(10):1368–78. doi: 10.1093/ps/76.10.1368
191. Savaris VDL, Broch J, de Souza C, Rohloff Junior N, de Avila AS, Polese C, et al. Effects of vitamin a on carcass and meat quality of broilers. Poult Sci (2021) 100(12):101490. doi: 10.1016/j.psj.2021.101490
192. Stephensen CB, Vitamin A. Infection, and immune function. Annu Rev Nutr (2001) 21:167–92. doi: 10.1146/annurev.nutr.21.1.167
193. Sijtsma SR, Rombout JHWM, Dohmen MJW, West CE, van der Zijpp AJ. Effect of vitamin a deficiency on the activity of macrophages in Newcastle disease virus-infected chickens. Veterinary Immunol Immunopathol (1991) 28(1):17–27. doi: 10.1016/0165-2427(91)90039-F
194. Lewis ED, Meydani SN, Wu D. Regulatory role of vitamin e in the immune system and inflammation. IUBMB Life (2019) 71(4):487–94. doi: 10.1002/iub.1976
195. Khalifa OA, Al Wakeel RA, Hemeda SA, Abdel-Daim MM, Albadrani GM, El Askary A, et al. The impact of vitamin e and/or selenium dietary supplementation on growth parameters and expression levels of the growth-related genes in broilers. BMC Veterinary Res (2021) 17(1):251. doi: 10.1186/s12917-021-02963-1
196. Habibian M, Ghazi S, Moeini MM. Effects of dietary selenium and vitamin e on growth performance, meat yield, and selenium content and lipid oxidation of breast meat of broilers reared under heat stress. Biol Trace Element Res (2016) 169(1):142–52. doi: 10.1007/s12011-015-0404-6
197. Pečjak M, Leskovec J, Levart A, Salobir J, Rezar V, Dietary Vitamin E, et al. Selenium and their combination on carcass characteristics, oxidative stability and breast meat quality of broiler chickens exposed to cyclic heat stress. Animals (2022) 12(14):1789. doi: 10.3390/ani12141789
198. Moriguchi S, Kobayashi N, Kishino Y. High dietary intakes of vitamin e and cellular immune functions in rats. J Nutr (1990) 120(9):1096–102. doi: 10.1093/jn/120.9.1096
199. Hieu TV, Guntoro B, Qui NH, Quyen NTK, Al Hafiz FA. The application of ascorbic acid as a therapeutic feed additive to boost immunity and antioxidant activity of poultry in heat stress environment. Vet World (2022) 15(3):685–93. doi: 10.14202/vetworld.2022.685-693
200. Bozonet S, Carr A, Pullar J, Vissers M. Enhanced human neutrophil vitamin c status, chemotaxis and oxidant generation following dietary supplementation with vitamin c-rich SunGold kiwifruit. Nutrients (2015) 7(4):2574–88. doi: 10.3390/nu7042574
201. Gierlikowska B, Stachura A, Gierlikowski W, Demkow U. Phagocytosis, degranulation and extracellular traps release by neutrophils-the current knowledge, pharmacological modulation and future prospects. Front Pharmacol (2021) 12:666732. doi: 10.3389/fphar.2021.666732
202. Yuandani M, Jantan I, Mohamad HF, K. Husain and AF. Abdul Razak: inhibitory effects of standardized extracts of Phyllanthus amarus and Phyllanthus urinaria and their marker compounds on phagocytic activity of human neutrophils. Evidence-Based Complementary Altern Med (2013) 2013:603634. doi: 10.1155/2013/603634
203. Harun NH, Septama AW, Jantan I. Immunomodulatory effects of selected Malaysian plants on the CD18/11a expression and phagocytosis activities of leukocytes. Asian Pacific J Trop Biomedicine (2015) 5(1):48–53. doi: 10.1016/S2221-1691(15)30170-2
204. Yang HS, Haj FG, Lee M, Kang I, Zhang G, Lee Y. Laminaria japonica extract enhances intestinal barrier function by altering inflammatory response and tight junction-related protein in lipopolysaccharide-stimulated caco-2 cells. Nutrients (2019) 11(5):1001. doi: 10.3390/nu11051001
205. Chelakkot C, Ghim J, Ryu SH. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp Mol Med (2018) 50(8):1–9. doi: 10.1038/s12276-018-0126-x
206. Swelum AA, Elbestawy AR, El-Saadony MT, Hussein EOS, Alhotan R, Suliman GM, et al. Ways to minimize bacterial infections, with special reference to Escherichia coli, to cope with the first-week mortality in chicks: an updated overview. Poult Sci (2021) 100(5):101039. doi: 10.1016/j.psj.2021.101039
207. Ciarlo E, Heinonen T, Théroude C, Asgari F, Le Roy D, Netea MG, et al. Trained immunity confers broad-spectrum protection against bacterial infections. J Infect Dis (2020) 222(11):1869–81. doi: 10.1093/infdis/jiz692
208. Gray P, Jenner R, Norris J, Page S, Browning G. Antimicrobial prescribing guidelines for poultry. Aust Vet J (2021) 99(6):181–235. doi: 10.1111/avj.13034
209. Huang CM, Lee TT. Immunomodulatory effects of phytogenics in chickens and pigs [[/amp]]mdash; a review. Asian-Australasian J Anim Sci (2018) 31(5):617–27. doi: 10.5713/ajas.17.0657
210. Rehman A, Arif M, Sajjad N, Al-Ghadi MQ, Alagawany M, Abd El-Hack ME, et al. Dietary effect of probiotics and prebiotics on broiler performance, carcass, and immunity. Poultry Sci (2020) 99(12):6946–53. doi: 10.1016/j.psj.2020.09.043
211. Froebel LK, Jalukar S, Lavergne TA, Lee JT, Duong T. Administration of dietary prebiotics improves growth performance and reduces pathogen colonization in broiler chickens. Poultry Sci (2019) 98(12):6668–76. doi: 10.3382/ps/pez537
212. Kabir SML, Rahman MM, Rahman MB, Ahmed SU. The dynamics of probiotics on growth performance and immune response in broilers. Int J Poult. Sci (2004) 3(5):361–4. doi: 10.3923/ijps.2004.361.364
213. Guo FC, Kwakkel RP, Soede J, Williams BA, Verstegen MWA. Effect of a Chinese herb medicine formulation, as an alternative for antibiotics, on performance of broilers. Br Poultry Sci (2004) 45(6):793–7. doi: 10.1080/00071660400012741
214. Florou-Paneri P, Giannenas I, Christaki E, Botsoglou N. Performance of chickens and oxidative stability of the produced meat as affected by feed supplementation with oregano, vitamin c, vitamin e and their combinations. Archiv fur Geflugelkunde (2006) 70:232–40.
215. Atteh JO, Onagbesan OM, Tona K, Decuypere E, Geuns JMC, Buyse J. Evaluation of supplementary stevia (Stevia rebaudiana, bertoni) leaves and stevioside in broiler diets: effects on feed intake, nutrient metabolism, blood parameters and growth performance. J Anim Physiol Anim Nutr (2008) 92(6):640–9. doi: 10.1111/j.1439-0396.2007.00760.x
216. Zhu N, Wang J, Yu L, Zhang Q, Chen K, Liu B. Modulation of growth performance and intestinal microbiota in chickens fed plant extracts or virginiamycin. Front Microbiol (2019) 10. doi: 10.3389/fmicb.2019.01333
217. Pattison M, McMullin P, Bradbury JM, Alexander D. Poultry diseases. Elsevier Health Sci (2007) Hardback ISBN: 9780702028625. 6th Edition.
218. Legnardi M, Baranyay H, Simon C, Molnár J, Bijlsma T, Cecchinato M, et al. Infectious bronchitis hatchery vaccination: comparison between traditional spray administration and a newly developed gel delivery system in field conditions. Veterinary Sci (2021) 8(8):145. doi: 10.3390/vetsci8080145
219. Gautério GV, Silvério SIDC, Egea MB, Lemes AC. β-glucan from brewer’s spent yeast as a techno-functional food ingredient. Front Food Sci Technol 2(2022). doi: 10.3389/frfst.2022.1074505
220. Manyi-Loh C, Mamphweli S, Meyer E, Okoh A. Antibiotic use in agriculture and its consequential resistance in environmental sources: potential public health implications. Molecules (2018) 23(4):795. doi: 10.3390/molecules23040795
221. Coffman RL, Sher A, Seder RA. Vaccine adjuvants: putting innate immunity to work. Immunity (2010) 33(4):492–503. doi: 10.1016/j.immuni.2010.10.002
222. Lamb DJ, Eales LJ, Ferns GA. Immunization with bacillus calmette-guerin vaccine increases aortic atherosclerosis in the cholesterol-fed rabbit. Atherosclerosis (1999) 143(1):105–13. doi: 10.1016/s0021-9150(98)00284-6
223. Quintin J, Saeed S, Martens JHA, Giamarellos-Bourboulis EJ, Ifrim DC, Logie C, et al. Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microbe (2012) 12(2):223–32. doi: 10.1016/j.chom.2012.06.006
Keywords: immune modulators, chickens, avian bacterial pathogens, prebiotics and probiotics, pathogen associated molecular patterns (PAMPs), microbial metabolites and derivatives, vitamins
Citation: Adams JRG, Mehat J, La Ragione R and Behboudi S (2023) Preventing bacterial disease in poultry in the post-antibiotic era: a case for innate immunity modulation as an alternative to antibiotic use. Front. Immunol. 14:1205869. doi: 10.3389/fimmu.2023.1205869
Received: 14 April 2023; Accepted: 12 June 2023;
Published: 03 July 2023.
Edited by:
Felix Ngosa Toka, Ross University School of Veterinary Medicine, Saint Kitts and NevisReviewed by:
Juan D. Latorre, University of Arkansas, United StatesPatrick Rik Butaye, Ghent University, Belgium
Copyright © 2023 Adams, Mehat, La Ragione and Behboudi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: James R. G. Adams, amEwMTMyOUBzdXJyZXkuYWMudWs=; Shahriar Behboudi, U2hhaHJpYXIuYmVoYm91ZGlAcGlyYnJpZ2h0LmFjLnVr