 Dominik Hanke1,2*
Dominik Hanke1,2* Zahady D. Velásquez2
Zahady D. Velásquez2 Kathrin Büttner3
Kathrin Büttner3 Andreas Krueger4
Andreas Krueger4 Ralf Ross4
Ralf Ross4 Andreas Hecker5
Andreas Hecker5 Sybille Mazurek1
Sybille Mazurek1 Veronika Grau5
Veronika Grau5 Anja Taubert2
Anja Taubert2 Carlos Hermosilla2
Carlos Hermosilla2 Katrin Richter5,6†
Katrin Richter5,6† Iván Conejeros2†
Iván Conejeros2†- 1Institute of Veterinary Physiology and Biochemistry, Justus Liebig University Giessen, Giessen, Germany
- 2Institute of Parasitology, Biomedical Research Center Seltersberg, Justus Liebig University Giessen, Giessen, Germany
- 3Unit for Biomathematics and Data Processing, Justus Liebig University Giessen, Giessen, Germany
- 4Institute of Molecular Immunology, Biomedical Research Center Seltersberg, Justus Liebig University Giessen, Giessen, Germany
- 5Laboratory of Experimental Surgery, Department of General and Thoracic Surgery, German Center for Lung Research (DZL), Cardiopulmonary Institute (CPI), Justus Liebig University Giessen, Giessen, Germany
- 6Department of Natural Sciences, Bonn-Rhein-Sieg University of Applied Sciences, Rheinbach, Germany
Introduction: Different innate immune cell types are known to release extracellular traps (ETs) in response to invasive pathogens, including parasites. These ETs function to trap, immobilize, and eventually kill pathogens. In line with this, monocytes and macrophages have been shown to release ETs, known as monocyte/macrophage extracellular traps (METs). Toxoplasma gondii (T. gondii) is an apicomplexan zoonotic parasite that infects humans and homeothermic animals. While most studies have focused on prolonged exposure of immune cells to T. gondii, this study characterized the early innate immune reaction of mononuclear phagocytes to vital T. gondii tachyzoites.
Methods: Primary human and bovine monocytes, monocytic THP-1 cells, and THP-1 cell-derived macrophages (M0-, M1-, and M2-like) were exposed to T. gondii tachyzoites for 4 h. Scanning electron microscopy (SEM), transmission electron microscopy (TEM), immunofluorescencemicroscopy, and confocal microscopy were used to visualize cell activation and the presence of METs. Additionally, the release of pro-inflammatory cytokines interleukin (IL)-1β and IL-6, and expression of Toll-like receptor (TLR) 2 and TLR4 were analyzed.
Results and discussion: Microscopic analysis illustrated the activation of all cell types tested within 4 h of exposure to T. gondii tachyzoites. Numerous tachyzoites were found intracellularly in THP-1 cell-derived M1-like macrophages. Furthermore, the co-localization of extracellular DNA (extDNA) and histones in extracellular web-like fibers proved classical characteristics of extruded T. gondii-induced METs, although this was a rare event. In primary human monocytes, an increased release of IL-1β and IL-6 was observed following exposure to T. gondii tachyzoites. When co-stimulated with lipopolysaccharide (LPS), primary human monocytes showed an enhanced release of IL-1β and IL-6 in response to T. gondii. In contrast to monocytic THP-1 cells, THP-1 cell-derived M1-like macrophages released IL-1β in response to T. gondii tachyzoite exposure. When additionally stimulated by LPS, all THP-1 cell-derived macrophages showed an enhanced release of IL-1β, and monocytic THP-1 cells an increased release of IL-6 in response to T. gondii tachyzoites. This study provides insights into the early innate immune response of human and bovine mononuclear phagocytes to T. gondii. While cytokine secretion was prominent, MET formation was rare in the early response (i.e. < 4 h of exposure) to T. gondii tachyzoites.
Introduction
Toxoplasma gondii (T. gondii) is a polyxenous zoonotic apicomplexan parasite able to infect warm-blooded mammals and birds worldwide (1). T. gondii is the etiological agent of human toxoplasmosis and it is estimated that one-third of the global population is seropositive for T. gondii (2). In humans, toxoplasmosis is particularly problematic in children after congenital transmission (i.e. prenatal toxoplasmosis) and in opportunistic infections of immunocompromised patients (i.e. postnatal toxoplasmosis) (3, 4). T. gondii is the only representative species of its genus and belongs to the class Sporozoa (subclass Coccidia, family Sarcocystidae). The life cycle of T. gondii comprises distinct parasitic stages. Infective parasitic stages are sporozoites within sporulated oocysts found in felid feces. Moreover, male and female gametocytes (exclusively form in the intestine of felines, acting as definitive hosts), fast replicating tachyzoites and bradyzoites in tissue cysts of intermediate hosts are found. During the acute phase of toxoplasmosis, fast replicating tachyzoites proliferate within all nucleated cell types, whereas bradyzoites, i.e. the slow replicating stages, develop within cysts and are primarily found in muscles, spleen, liver and brain. These stages propagate the chronic persistence of T. gondii. The main transmission routes of T. gondii to humans are: i) ingestion of undercooked meat containing tissue cysts, ii) consumption of food- and/or water contaminated with sporulated oocysts, iii) congenital transmission, iv) administration of blood transfusions containing tachyzoites, and v) organ transplants containing either tissue cysts or tachyzoites (2, 5).
Within the mammalian host innate immune system, mononuclear phagocytes like monocytes and macrophages play a crucial role in the initiation of adaptive immunity by acting as first responders to infection in the vasculature and tissue (6, 7). After polymorphonuclear neutrophils (PMN), monocytes are the second most abundant phagocytes in the blood, comprising 10% of the peripheral white blood cells in humans and 4% in cattle (8, 9). Besides macrophages and dendritic cells (DC), monocytes form part of the mononuclear phagocyte system (MPS) and are characterized by their rapid recruitment in response to pathogen infection or tissue injury (9, 10). Classical monocyte- or macrophage-derived effector mechanisms, include phagocytosis, chemokine- [e.g. chemoattractant protein-1 (CCL2)] and cytokine secretion [e.g. interleukin (IL)-1β and IL-6)], reactive oxygen species (ROS) production, and expression of a wide range of Toll-like receptors (TLRs) and purinergic receptors (e.g. P2X7) (11–16). Additionally, mammalian monocytes and macrophages are capable to release extracellular traps (METs) (17–24), an early defense mechanism which was first observed in PMN and named neutrophil extracellular traps (NETs) (25). As shown for extruded NETs, METs are mainly composed of extracellular DNA (extDNA) and proteins like citrullinated histones (H1, H2A/H2B, H3, H4) and myeloperoxidase (MPO). METs function to trap, immobilize, and eventually kill pathogens, including parasites such as Besnoitia besnoiti (26), Cryptosporidium parvum (23), Eimeria ninakohlyakimovae (19) and Neospora caninum (21). In addition, the release of METs has been described in response to human sperm in vitro and ex vivo in male patients with leukocytospermia (27). Nonetheless, only one study so far reported on T. gondii-induced MET release, namely in monocytes of harbour seals (Phoca vitulina) (18), thereby proving that this defense mechanism seems to be highly conserved, as also assumed for NET formation (28). MET release is also induced by chemical activators like A23187 (calcium ionophore), phorbol 12-myristate 13-acetate (PMA), platelet activating factor (PAF) (20, 24), zymosan (18–21, 26), or gold nanoparticles (29).
Monocytic THP-1 cells and THP-1 cell-derived-macrophages are a widely accepted model for human mononuclear phagocytes to study immunological mechanisms in vitro (30). THP-1 cells were originally isolated from a patient with acute myeloid leukemia (30–32). Monocytic THP-1 cells can be differentiated into non-polarized M0-like macrophages, pro-inflammatory M1-like macrophages and anti-inflammatory M2-like macrophages, providing the opportunity to directly compare all four cell types in vitro (16, 30, 33).
Monocytic THP-1 cells and THP-1 cell-derived macrophages have been shown to release METs in response to a variety of chemical activators and stimuli [e.g. tunicamycin (34), PMA (35), A23187, HOCl, nigericin, tumor necrosis factor (TNF)-α (36), zymosan (37), murine extracellular cold-inducible RNA-binding protein (eCIRP) (38), and aflatoxin B1 (39)] as well as parasite [e.g. Trichomonas vaginalis (37)] and bacteria [e.g. Escherichia coli (17), Streptococcus agalactiae (40), and Mycobacterium massiliense (41)].
An important function of mononuclear phagocytes is the release of cytokines like IL-1β and IL-6 in response to diverse danger- or pathogen-associated molecular patterns (DAMPs, PAMPs) (42–45). IL-1β and IL-6 have distinct roles in the inflammatory process. IL-1β is a primary initiator of innate immune reaction and key player in host defense against infections, while the pro- and anti-inflammatory IL-6 amplifies and modulates inflammatory responses (44). IL-1β is processed from a precursor by caspase cleavage to become biologically active and is then secreted. The mere induction of pro-IL-1β expression (priming step) is inefficient to induce secretion (44, 46). Consecutive second PAMPs or DAMPs are required to induce the NOD- LRR- and pyrin domain-containing protein 3 (NLRP3) inflammasome-dependent processing, and secretion of mature IL-1β (45). IL-6 is secreted by mononuclear phagocytes in response to PAMPs (44, 46). These PAMPs bind to pattern recognition receptors (PRRs) such as TLR2.
In mammals, 13 TLRs are known. These are located on the plasma membrane and in intracellular compartments, where they activate signaling cascades to produce molecules and cytokines to fight against pathogens including T. gondii (47, 48). TLR2 and TLR4 have been shown to be involved in immune responses against T. gondii, with TLR2 recognizing the parasitic surface profilin or glycosylphosphatidylinositol (GPI) proteins, and stimulating the production of IL-12, while TLR4 plays a supportive role, especially at high infectious doses, as shown for several mouse models (47).
The role of IL-1β and IL-6 in T. gondii-induced immune responses is controversial, possibly due to the use of different parasite strains (e.g. type I [RH], type II [Prugniaud, ME49], type III [CEP], or atypical strains such as TgChBrUD2), reflecting the considerable genetic variability of T. gondii. This strain diversity is reflected in pronounced differences in virulence and immune modulation among strains (49, 50), which may explain the inconsistent findings. A key factor contributing to this variability is partly driven by strain-specific polymorphic effector proteins that differentially modulate host immune signaling pathways, thereby influencing cytokine responses (51, 52). In line with this, Gov et al., 2013 and Guimaraes Gois et al., 2022 demonstrated strain-specific cytokine profiles (53, 54). While some studies on primary human monocytes showed a T. gondii tachyzoite-induced increased release of IL-1β after 4, 16 and 24 h (53, 55–58), Liam et al., 2018 found no impact of tachyzoite exposure on lipopolysaccharide (LPS)-induced IL-1β release (56). The same applies for IL-6. Pelloux et al., 1994 and Tombácz et al., 2018 found no impact of T. gondii tachyzoites or profilin on the release of IL-6 by primary monocytes after 24 h exposure (59, 60), whereas Salazar Gonzalez et al., 2014 have found an impact of T. gondii profilin (61). Further studies provide evidence for a parasite-induced release of macrophage migration inhibitory factor (MIF), IL-1β, IL-6 and IL-12 by monocytic THP-1 cells (53, 55, 62, 63), and a fostered IL-1β and MIF secretion by M0-like macrophages (54, 64).
Of note, the above described studies either focused on the release of METs (18), production of IL-1β and IL-6 (53–61, 63, 64) or the expression of TLR2 and TLR4 (47, 48).
Here we studied in a comprehensive systematic manner the effect of T. gondii tachyzoites on the acute and early phase response of primary human and bovine monocytes as well as monocytic THP-1 cells and THP-1 cell-derived macrophages, including the release of METs, the production of pro-inflammatory cytokines IL-1β and IL-6, and the expression of the surface receptors TLR2 and TLR4.
Materials and methods
THP-1 cell culture
Monocytic THP-1 cells (Leibniz Institute, DSMZ-German Collection of Microorganisms and Cell Cultures GmbH, Braunschweig, Germany, Cat#ACC16) were cultured in T-75 cell culture flasks (Sarstedt, Nümbrecht, Germany, Cat#83.3911.002) in Roswell Park Memorial Institute (RPMI) 1640 medium (Sigma-Aldrich, Merck, Darmstadt, Germany, Cat#R8758) supplemented with 10% fetal calf serum (FCS) Xtra (Capricorn Scientific, Ebsdorfergrund, Germany, Cat#FBS-16A), 1% penicillin-streptomycin [50 U/ml penicillin, 50 µg/ml streptomycin] (Gibco™, Thermo Fisher Scientific, Waltham, Massachusetts, USA, Cat#151401222), and transferred to 12-well plates (Greiner Bio-One, Kremsmünster, Austria, Cat#655180) or 96-well plates (Greiner Bio-One, Cat#655180) on the day of the experiment. To generate THP-1 cell-derived macrophages of M0-, M1-, and M2-like phenotypes, cells were seeded in T-75 cell culture flasks (6 x 106 cells/flask) with RPMI-1640 medium. Cells were differentiated using 50 nM PMA (Thermo Fisher Scientific, Cat#P1585) according to a previously described protocol (16) resulting in cells with macrophage-like properties, such as increased cell adherence and a reduced proliferation rate compared to monocytic THP-1 cells (65). The protocol for the polarization to M1-like macrophages has also been published (16). Briefly, cells were cultured in medium supplemented with 10 ng/ml LPS (Escherichia coli, O111:B4, Sigma-Aldrich, Merck, Cat#L2630) and 10 ng/ml interferon (IFN)-γ (R&D Systems, Minneapolis, USA, Cat#285-IF) for 48 h (16). For polarization to M2-like macrophages, the medium was supplemented with 20 ng/ml IL-4 (Preprotec, Thermo Fisher Scientific, Cat#200-04) and 20 ng/ml IL-13 (Gibco™, Preprotec, Thermo Fisher Scientific, Cat#200-13). On day 5 of differentiation, cells in T-75 cell culture flasks were washed with Dulbecco’s phosphate buffered saline (PBS, Sigma-Aldrich, Merck, Cat#D8537), detached using TrypLE™ Express (Gibco™, Thermo Fisher Scientific, Cat#12605010) according to the manufacturer’s protocol, transferred to Eppendorf tubes (Sarstedt, Cat#72.691), 8-well cell culture chambers (Sarstedt, Cat#94.6170.802), 12-well plates or 96-well plates and cultured in Dulbecco’s modified Eagle’s medium (DMEM) without glucose, glutamine and phenol red (Gibco™, Thermo Fisher Scientific, Cat#A1443001) supplemented with 10% heat-inactivated FCS Xtra, 2 mM D-glucose (Carl Roth, Karlsruhe, Germany, Cat#X997.1), 1 mM L-glutamine (Sigma-Aldrich, Merck, Cat#G3126), 1% penicillin-streptomycin. For all experiments the same batch of FCS Xtra was used and heat-inactivation was done according to the manufacturer’s protocol, as FCS contains active deoxyribonuclease (DNases) that can degrade extracellular traps (66).
Isolation of primary human and bovine monocytes
Primary human monocytes were obtained by positive and negative selection protocols. For positive selection, buffy coats from healthy female and male donors (n = 6) of the Center of Transfusion Medicine and Hemotherapy (JLU Giessen) and the University Hospital of Giessen and Marburg (UKGM), were used. The protocol for positive selection of primary human monocytes was based on a previously published method (67). Briefly, 15 ml of buffy coats were diluted 1:1 with RPMI-1640 medium (Sigma-Aldrich, Cat#R5886), supplemented with 5% FCS (Biochrom GmbH, Berlin, Germany, Cat#S0115), 2 mM L-glutamine (Sigma-Aldrich, Merck, Darmstadt, Germany, Cat#G7513), 1 mM sodium pyruvate (Sigma-Aldrich, Merck, Cat#S8636), 1 mM minimum essential medium (MEM) non-essential amino acid solution (Sigma-Aldrich, Merck, Cat#M7145) and 1% penicillin-streptomycin (Sigma-Aldrich, Merck, Cat#P4333), transferred to 15 ml Biocoll® separation solution (Biochrom GmbH, Cat#L6115, density: 1.077 g/ml), and centrifuged [2500 x g, without brake, 30 min, room temperature (RT)]. The ring containing peripheral blood mononuclear cells (PBMCs) was carefully collected and cells were washed three times by filling up to a volume of 30 ml with MACS® separation buffer (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany, Cat#130-091-222) containing 2% bovine serum albumin (BSA; Miltenyi Biotec GmbH, Cat#130-091-376) followed by centrifugation (1500 x g, 7 min, 4 °C). The supernatant was discarded and the PBMC pellet was resuspended in 5 ml MACS® separation buffer containing 2% BSA and subjected to positive selection for CD14+ monocytes using immunomagnetic human CD14+ MicroBeads® (Miltenyi Biotec GmbH, Cat#130-050-201). Briefly, 2.5 x 108 PBMCs and 500 µl CD14+ MicroBeads® were incubated in a total volume of 2.5 ml MACS® separation buffer containing 2% BSA for 15 min at 4 °C. CD14+ monocytes were isolated using LS® columns (Miltenyi Biotec GmbH, Cat#130-042-401) and QuadroMACS™ separators (Miltenyi Biotec GmbH, Cat#130-090-976) according to the manufacturer’s protocol. Isolated human monocytes were resuspended in cold RPMI-1640 medium, supplemented with 5% FCS, 2 mM L-glutamine, 1 mM sodium pyruvate, 1 mM MEM non-essential amino acid solution and 1% penicillin-streptomycin (all Sigma-Aldrich) and stored on ice until experiments.
For negative selection, human peripheral blood was obtained from healthy female and male adult donors (n = 6) via antecubital venipuncture and collected in a heparinized syringe (Heparin-Natrium-5000-ratiopharm®, ratiopharm, Ulm, Germany) containing 1 mM ethylenediaminetetraacetic acid (EDTA; Applichem, Darmstadt, Germany, Cat#A-1105) per one ml blood. Monocytes were isolated from whole blood using the RosetteSep™ human Monocyte Enrichment Cocktail (Stemcell Technologies, Cologne, Germany, Cat#15068) and Lymphoprep™ density gradient (Stemcell Technologies, Cat#18060), following to the manufacturer’s protocol. Monocytes were resuspended in cold RPMI-1640 medium (Capricorn Scientific, Cat#RPMI-ADV), supplemented with 1% penicillin-streptomycin (Gibco™, Thermo Fisher Scientific, Cat#15140-122) and stored on ice until performing experiments. The purity of monocytes was confirmed to be > 70% by flow cytometry analysis (FCA) as previously described (16).
For the isolation of primary bovine monocytes, healthy adult dairy cows (n = 6) served as blood donors. Peripheral blood was obtained by jugular venipuncture and collected in heparinized tubes (Kabe Labortechnik GmbH, Nümbrecht-Elsenroth, Germany, Cat#041551). The modified protocol for the isolation of primary bovine monocytes was based on previously published methods (23, 68, 69). In brief, 50 ml blood was centrifuged (1260 x g, without brake, 30 min, RT). Then 10 ml plasma plus the ring containing PBMCs was collected. The plasma phase was filtered through a 0.45 µm filter (Sarstedt, Cat#83.1826) and incubated for 1 h in a 2% gelatin-coated T-75 flask at 37 °C and 5% CO2. For gelatin coating, the T-75 flask was incubated the day before with 2% gelatin solution (Sigma-Aldrich, Merck, Cat#G1890) for 2 h at 37 °C and 5% CO2, the supernatant was removed, and the T-75 flask was dried overnight in the same conditions. The PBMCs were resuspended in 30 ml PBS containing 0.02% EDTA, transferred to 15 ml Histopaque®-1077 separation solution (Sigma-Aldrich, Merck, Cat#10771, density: 1.077 g/ml) and centrifuged (1500 x g, without brake, 30 min, 4 °C). The ring containing PBMCs was collected, and cells were washed two times by filling up to a volume of 50 ml with PBS containing 0.02% EDTA followed by centrifugation (400 x g, 7 min, 4 °C). The supernatant was discarded and PBMCs were resuspended in 10 ml of pre-warmed RPMI-1640 medium (Sigma-Aldrich, Merck, Cat#R8758) supplemented with 1% penicillin and streptomycin (Gibco™, Thermo Fisher Scientific, Cat#151401222). The supernatant of incubated plasma in the T-75 flask was removed and the T-75 flask was washed once with pre-warmed RPMI-1640 medium supplemented with 1% penicillin-streptomycin. The PBMC suspension was transferred to the washed T-75 flask and incubated for 1 h at 37 °C and 5% CO2. Non-adherent cells were removed by rinsing the flask once with pre-warmed RPMI-1640 medium supplemented with 1% penicillin-streptomycin. Monocytes were detached by addition of 10 ml 10 mM EDTA suspended in Hanks’ Balanced Salt Solution (HBSS) without sodium bicarbonate, calcium and magnesium (Corning®, USA, New York, Cat#20-021-CVR) for 10 min at 37 °C and 5% CO2, centrifuged (400 x g, 10 min, 4°C), resuspended in cold RPMI-1640 medium supplemented with 1% penicillin-streptomycin, and stored on ice. A commercially available mouse anti-bovine CD172a antibody (Bio-Rad Laboratories, Feldkirchen, Germany, Cat#MCA2041GA) was used to stain primary bovine monocytes, resulting in a purity of > 90% for CD172+ monocytes (Supplementary Figure S1).
In vitro culture of Toxoplasma gondii tachyzoites
Vital tachyzoites of T. gondii (RH strain) were isolated from continuous passages on African green monkey kidney epithelial (MARC)-145 cells using DMEM (Sigma-Aldrich, Merck, Cat#D6429) supplemented with 5% FCS, 1% penicillin-streptomycin (Gibco™, Thermo Fisher Scientific, Cat#151401222). T. gondii tachyzoites released from host cell cultures were seperated from cell detritus by centrifugation (460 x g, 1 min, RT). Supernatants containing T. gondii tachyzoites, were filtered (5 µm pore sized filters, Satorius, Göttingen, Germany, Minisart®, Cat#17594-K) and pelleted (460 x g, 5 min, RT). The harvested T. gondii tachyzoites were counted in a Neubauer chamber, suspended in medium, and used for co-incubation with primary human or bovine monocytes as well as monocytic THP-1 cells or THP-1 cell-derived M0-, M1-, and M2-like macrophages. All experiments were performed at 37 °C and 5% CO2 atmosphere, with a multiplicity of infection (MOI) of 1:4 (cells: parasites), and a co-incubation period of 4 h.
Scanning electron microscopy
In total, 2.5 x 105 primary human or bovine monocytes as well as monocytic THP-1 cells or THP-1 cell-derived M0-, M1-, and M2-like macrophages were either left untreated or co-cultured for 4 h with 1 x 106 T. gondii tachyzoites on 10 mm coverslips (Carl Roth, Cat#KHX4.1) pre-coated with 0.01% poly-L-lysine (Sigma-Aldrich, Cat#4707) in 12-well plates. After incubation, cells were fixed in 2.5% glutaraldehyde (Sigma-Aldrich, Merck, Cat#G6257), post-fixed in 1% osmium tetroxide (Polysciences Inc., Warrington, Pennsylvania, USA, Cat#0223B-10), washed in purified water critical point-dried using CO2 treatment, and sputtered with gold particles. The samples were analyzed using a Philips XL30 microscope (Amsterdam, Netherlands) at the Institute of Anatomy and Cell Biology (JLU Giessen). Brightness and contrast were processed in a standardized way using Image J software, Fiji Is Just ImageJ (FIJI) version (70). The percentage of protrusion positive cells was calculated for each primary monocyte donor, using a cell range of 282 to 796 for human cells and 465 to 1871 for bovine cells. Cells were classified as protrusion positive by the presence of filopodia and cytonemes.
Pappenheim staining
1 x 105 THP-1 cell-derived M0-, M1-, and M2-like macrophages were either left untreated or co-cultured with 4 x 105 T. gondii tachyzoites in 8-well cell culture chambers for 4 h and fixed with 4% formaldehyde (Carl Roth, Cat#CP10.4) diluted in PBS for 15 min at RT. Samples were gently washed three times with PBS. For staining, the cells were treated with May-Grünwald’s eosine-methylene blue solution (Sigma-Aldrich, Merck, Cat#1.01424) for 2 min, distilled water for 5 min, Giemsa’s azur eosin methylene blue solution (1:20, Sigma-Aldrich, Merck, Cat#1.09204) for 15 min, distilled water for 10 min, and air dried at RT. Specimens were documented using a Leica DM IL LED microscope equipped with a Leica Flexcam C3 (Wetzlar, Germany). Brightness and contrast were processed in a standardized way using Image J software, FIJI version (70).
Transmission electron microscopy
1 x 107 M1-like macrophages were either left untreated or co-cultured with 4 x 107 T. gondii tachyzoites in an Eppendorf tube for 4 h and centrifuged (1000 × g, 10 min, RT). The cell pellet was fixed in 2.5% glutaraldehyde and 1.5% formaldehyde diluted in 0.3 M 4-(2-hydroxyethyl)piperazine-1-ethanesulfonic acid buffer (HEPES, pH 7.35, Sigma-Aldrich, Merck, Cat#H4034) and stored at 4 °C according to Brinkmann et al., 2004 and Behrendt et al., 2008 (25, 71). The cell pellet was washed and post-fixed in 0.15 M HEPES buffer containing 1% osmium tetroxide. After thorough washing in distilled water, the samples were incubated overnight in 2% aqueous uranyl acetate dihydrate (Merck, Cat#8473) at RT, dehydrated in ethanol (Sigma-Aldrich, Merck, Cat#32205-M), and embedded in an epoxy resin using the Agar 100 Resin Kit (Agar Scientific, Essex, UK Cat#AGR1031). Ultrathin sections were mounted on grids and examined using a TEM Zeiss 902 (Oberkochem, Germany) at the Institute of Anatomy and Cell Biology (JLU Giessen). Brightness and contrast were processed in a standardized way using Image J software, FIJI version (70).
Visualization of monocyte/macrophage extracellular traps via conventional and confocal immunofluorescence microscopy
1.5 x 105 primary human or bovine monocytes as well as monocytic THP-1 cells or THP-1 cell-derived M0-, M1-, and M2-like macrophages were either left untreated, co-cultured for 4 h with 6 x 105 T. gondii tachyzoites or stimulated with A23187 [5 µM] (Sigma-Aldrich, Merck, Cat#C7522) or nigericin [0.5 µM] (sodium salt, Sigma-Aldrich, Merck, Cat#N7143) on 15 mm coverslips (Carl Roth, Cat#P232.1) pre-coated with 0.01% poly-L-lysine in 12-well plates, fixed in 4% formaldehyde diluted in PBS, and stored at 4°C until further use. The fixed samples were washed three times with PBS, blocked in blocking/permeabilization solution [PBS containing 3% BSA (Sigma-Aldrich, Merck, Cat#A7906) and 0.3% Triton X-100 (Sigma-Aldrich, Merck, Cat#T8787)] for 1 h at RT and reacted with primary antibodies in blocking/permeabilization solution overnight at 4 °C. Anti-histone antibodies (clone H11-4, Sigma-Aldrich, Merck, Cat#MAB3422) and anti-myeloperoxidase (MPO) antibodies (Biorbyt, Cambridge, UK, Cat#orb16003) were used to detect corresponding proteins on extruded METs. After three washings with PBS, samples were incubated in a secondary antibody blocking/permeabilization solution (Alexa Fluor 594 goat anti-mouse IgG, Invitrogen™, Thermo Fisher Scientific, Cat#A11005) for 30 min at RT. Finally, the samples were washed three times with PBS. Coverslips were mounted on microscope slides (Glaswarenfabrik KarlHecht GmbH & Co. KG, Sondheim, Germany, Cat#42400010) with Fluoromount-G™ (Invitrogen™, Thermo Fisher Scientific, Cat#00-4959-52) to stain DNA with 4′,6-diamidino-2-phenylindole (DAPI) and incubated for 48 h at RT. To visualize METs, specimens were analyzed using an Olympus IX81 microscope (Tokyo, Japan) equipped with an Excelitas X-Cite 120PC Q fluorescence illuminator (Pittsburgh, Pennsylvania, USA) and an Olympus XM-10-R digital camera. Confocal microscopy was performed using a Nikon Eclipse Ti2-A microscope (Tokyo, Japan) equipped with a ReScan® confocal unit (RCM 1.1 Visible, Confocal.nl, Amsterdam, Netherlands). The confocal microscope was equipped with a 50 μm pinhole, a Nikon motorized Z-stage (DI1500), and a Toptica CLE laser (Graefelfing, Germany) with the following excitation modes: blue/DAPI/405 nm, green/fluorescein-5-isothiocyanate (FITC)/488 nm, and red/AlexaFluor594/561 nm. An Excelitas sCMOS® camera (PCO edge) was used for documentation and the instrument was operated using NIS-Elements software (version 5.11, Nikon). Z-series were displayed as maximum z-projections and gamma, brightness and contrast were processed in a standardized way using Image J software, FIJI version (70).
Quantification of extracellular DNA
5 x 104 primary human or bovine monocytes as well as 1 x 105 monocytic THP-1 cells or THP-1 cell-derived M0-, M1-, and M2-like macrophages were either left untreated, co-cultured for 4 h with 2 x 105 or 4 x 105 T. gondii tachyzoites or stimulated with A23187 [5 µM] or nigericin [0.5 µM] in 96-well plates (Greiner Bio-One, Cat#655180). After incubation, Sytox Green [5 µM] (Invitrogen™, Thermo Fisher Scientific, Cat#S7020) was added to all samples to stain extDNA followed by centrifugation (400 x g, 5 min, RT). The supernatants were collected carefully to fresh 96-well plates and measured at an excitation wavelength of 488 nm using a Clariostar Plus plate reader and MARS data analysis software (version 4.20, both BMG Labtech, Ortenberg, Germany).
Quantification of nuclear area expansion and MET positive cells
1.5 x 105 primary human or bovine monocytes as well as monocytic THP-1 cells or THP-1 cell-derived M0-, M1-, and M2-like macrophages were either left untreated, co-cultured for 4 h with 6 x 105 T. gondii tachyzoites or stimulated with A23187 [5 µM] or nigericin [0.5 µM] on 15 mm coverslips pre-coated with 0.01% poly-L-lysine in 12-well plates, fixed in 4% formaldehyde in PBS, and stored at 4 °C until further use. Fixed samples were washed three times with PBS, blocked in blocking/permeabilization solution for 1 h at RT, and washed three times with PBS. Coverslips were mounted on microscope slides with Fluoromount-G™ to stain DNA with DAPI an incubated for 48 h at RT. Cell nuclei were visualized using a Keyence BZ-X800 microscope (Osaka, Japan) and measurements were performed using the Keyence BZ-X800 analyzer software (version 1.1.2.4). The average nuclear area of DAPI-stained DNA was calculated for each cell donor/passage using a field of 5.290 x 3.968 mm. The percentage of MET positive cells was determined by analyzing 1250 cells per condition. Cells were classified as MET positive by the presence of extracellular, fibre-like DNA structures outside the cell area.
Measurement of cytokine release via ELISA
1 x 105 primary human or bovine monocytes as well as monocytic THP-1 cells or THP-1 cell-derived M0-, M1-, and M2-like macrophages were seeded into 96-well plates. Cells were either left untreated, co-cultured with 4 x 105 T. gondii tachyzoites in the presence or absence of LPS [1 µg/ml] or treated with the known chemical activators of MET release A23187 [5 µM] or nigericin [0.5 µM] for 4 h. To proof that the used cells responded with IL-1β release to classical NLRP3 activation stimuli, cells were first primed with LPS for 3.5 h followed by exposure for 40 min to 2’/3’-O-(4-benzoylbenzoyl)adenosine-5’-triphosphate, tri(triethylammonium) salt (BzATP) [200 µM] (Jena Bioscience, Jena, Germany, Cat#NU-1620) or nigericin [25 µM] in the absence of T. gondii tachyzoites. Thereafter, the cells were centrifuged (500 x g, 8 min, 4 °C) and the cell-free supernatants were collected and stored at -20 °C for further use. IL-1β and IL-6 concentrations were measured in cell supernatants using the human IL-1 beta/IL-1F2 DuoSet enzyme-linked immunosorbent assay (ELISA; R&D Systems, Cat#DY201; assay range: 3.9 – 250 pg/ml) and the human IL-6 DuoSet ELISA (R&D Systems, Cat#DY206; assay range: 9.4 – 600 pg/ml) or the bovine IL-1 beta Uncoated ELISA Kit (Invitrogen™, Thermo Fisher Scientific, Cat#ESS0027) and the bovine IL-6 Uncoated ELISA Kit (Invitrogen™, Thermo Fisher Scientific, Cat#ESS0029).
To estimate the proportion of dead primary human or bovine monocytes as well as monocytic THP-1 cells or THP-1 cell-derived M0-, M1-, and M2-like macrophages, lactate dehydrogenase (LDH) activity was measured in the cell culture supernatants according to the supplier’s instructions of the CytoTox 96® Non-Radioactive Cytotoxicity Assay (Promega, Madison, Wisconsin, USA; Cat#G1780) using a Epoch™ Microplate Spectrophotometer (BioTek Instruments, Inc., Vermont, USA). The LDH activities determined in the cell-free supernatants are expressed as a percentage of the total LDH activity of the lysed control cells.
Detection of monocytic TLR2, TLR4, CD11b and F-actin by flow cytometry
5 x 105 primary human and bovine monocytes were left in plain medium or co-cultured for 4 h with 2 x 106 T. gondii tachyzoites in cell culture Petri dishes (Sarstedt, Cat#83.3902.500) (primary human monocytes) or in Eppendorf tubes (primary bovine monocytes), and thereafter fixed in 0.7% formaldehyde (primary human monocytes) or BD Cytofix (Franklin Lakes, New Jersey, USA, Cat#554722) (primary bovine monocytes). In case of primary human monocytes, the samples were washed in PBS containing 2% FCS and incubated for 10 min on ice with 10% heat-inactivated human serum to block Fc-receptors. Samples were washed with PBS containing 2% FCS and labelled with antibodies raised against CD11b (1:50, ImmunoTools GmbH, Friesoythe, Germany, Cat#21279116X3), TLR2 (1:10, BioLegend®, San Diego, USA, Cat#309708) or TLR4 (1:10, BioLegend®, Cat#312806) according to the manufacturer’s protocol. F-actin was detected by phalloidin coupled with Alexa 488 (abcam®, Cambridge, UK, Cat#ab176753). The stained cells were washed twice in PBS containing 2% FCS, centrifuged (1500 x g, 7 min, 4°C), resuspended in 0.5 ml PBS containing 2% FCS, and analyzed using a BD Accuri™ C6 flow cytometer (BD Biosciences, Heidelberg, Germany). In case of primary bovine monocytes, the samples were washed in PBS containing 1% BSA and labelled with antibodies raised against CD11b (BD, Cat#553312), TLR2 (Bio-Rad Laboratories, Hercules, California, USA, Cat#HCA152A647) or TLR4 (Bio-Rad Laboratories, Cat#MCA2061A647) in a dilution of 1:100 according to the manufacturer’s protocol. F-actin was detected by phalloidin coupled with Alexa 488. The stained cells were washed in PBS containing 1% BSA, centrifuged (600 x g, 10 min, 4°C), resuspended in 0.4 ml PBS containing 1% BSA, analyzed using a BD Accuri™ C6 flow cytometer. In the cell/parasite suspension, no discrimination between infected cells vs. uninfected cells was performed. The gating strategy and fluorescence intensity profiles for primary human and bovine monocytes are shown in Supplementary Figure S2 and Supplementary Figure S3.
Statistical analyses
Data were presented as mean ± standard deviation (SD) or median ± interquartile range (75th and 25th percentiles) using GraphPad Prism® (version 10.2.3, GraphPad Software, Boston, Massachusetts, USA). Data analysis was performed using SAS® statistical software (version 9.4, Statistical Analysis System Institute, Cary, North Carolina, USA). For all experiments, the number (n) refers to independent experiments performed on different days and different cell passages/blood donors. Outliers were not excluded from the analyses. The statistical tests used for individual data sets are indicated in the legends of the respective figures.
Results
Differential microscopic analyses illustrate activation of mononuclear phagocytes within 4 h of exposure to T. gondii tachyzoites
A panel of microscopic analyses was performed to investigate early responses of mononuclear phagocytes to T. gondii tachyzoites. To compare findings obtained from primary human and bovine monocytes, the human monocytic THP-1 cell line was analyzed in parallel.
In SEM analyses, primary untreated (control) human and bovine monocytes revealed typical spherical shapes and surfaces with little cytoplasmic spreading and ruffling (26, 72) (Figure 1A).
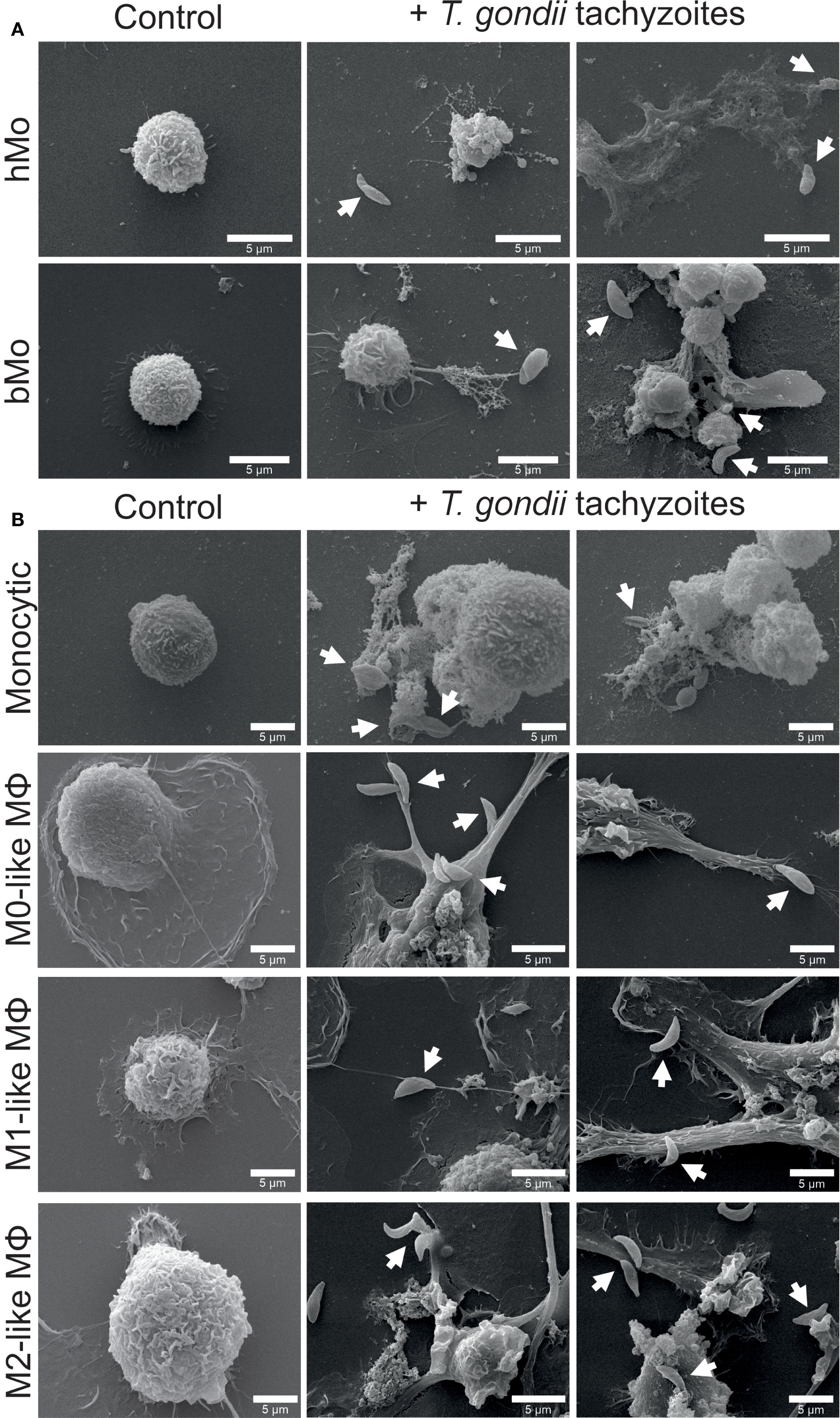
Figure 1. Exposure to Toxoplasma gondii tachyzoites triggers phenotypic changes in primary human and bovine monocytes, monocytic THP-1 cells and THP-1 cell-derived macrophages. Untreated (control) and parasite-exposed (+ T. gondii tachyzoites) primary human (hMo) and bovine monocytes (bMo) (A), monocytic THP-1 cells (monocytic) as well as THP-1 cell-derived M0-, M1-, and M2-like macrophages (MΦ) (B) were fixed after 4 h of co-incubation with T. gondii tachyzoites (MOI 1:4) and analyzed by scanning electron microscopy (SEM). Interactions of cells with T. gondii tachyzoites are indicated by white arrows. Representative pictures from n = 3 independent experiments are shown.
SEM images show that both primary human and bovine monocytes exhibit morphological changes (e.g. protrusion) upon exposure to the parasite, similar to those observed in other mononuclear phagocytes (Figure 1A, Supplementary Figure S4) (73–75). Compared with non-stimulated primary human and bovine monocytes, T. gondii tachyzoites induced cytoplasmic spreading, cell surface ruffling, formation of membrane protrusions by filopodia and cytonemes, and MET formation. The release of METs was accompanied by an extrusion of a large amount of cell debris as well as parasite entrapment, thereby probably corresponding to ‘suicidal METosis’ (Figure 1A). Untreated monocytic THP-1 cells presented a round-shaped and loosely attached phenotype, while THP-1 cell-derived macrophages had an irregular expanded surface and showed pronounced adhesion capacity (Figure 1B). Exposure to T. gondii tachyzoites induced membrane protrusions, mainly consisting of filopodia (Figure 1B). Similar results were found in Pappenheim-stained THP-1 cell-derived macrophages (Supplementary Figure S5).
In addition, TEM analyses were performed and confirmed the typical ultrastructure of THP-1 cell-derived macrophages (Figure 2) (76). THP-1 cell-derived M1-like control macrophages exhibited an ameboid shape and a large number of cytoplasmic granules, vesicles and vacuoles (Figure 2A). T. gondii exposure induced cell protrusions (Figure 2B). Moreover, ET-like structures, which contained and eventually trapped T. gondii tachyzoites, were observed in juxtaposition to M1-like macrophages (Figures 2B, C). Besides, several intracellular T. gondii tachyzoites were found, partially enclosed by a distinct membrane, resembling that of phagolysosomes or parasitophorous vacuoles (PV) (Figure 2D).
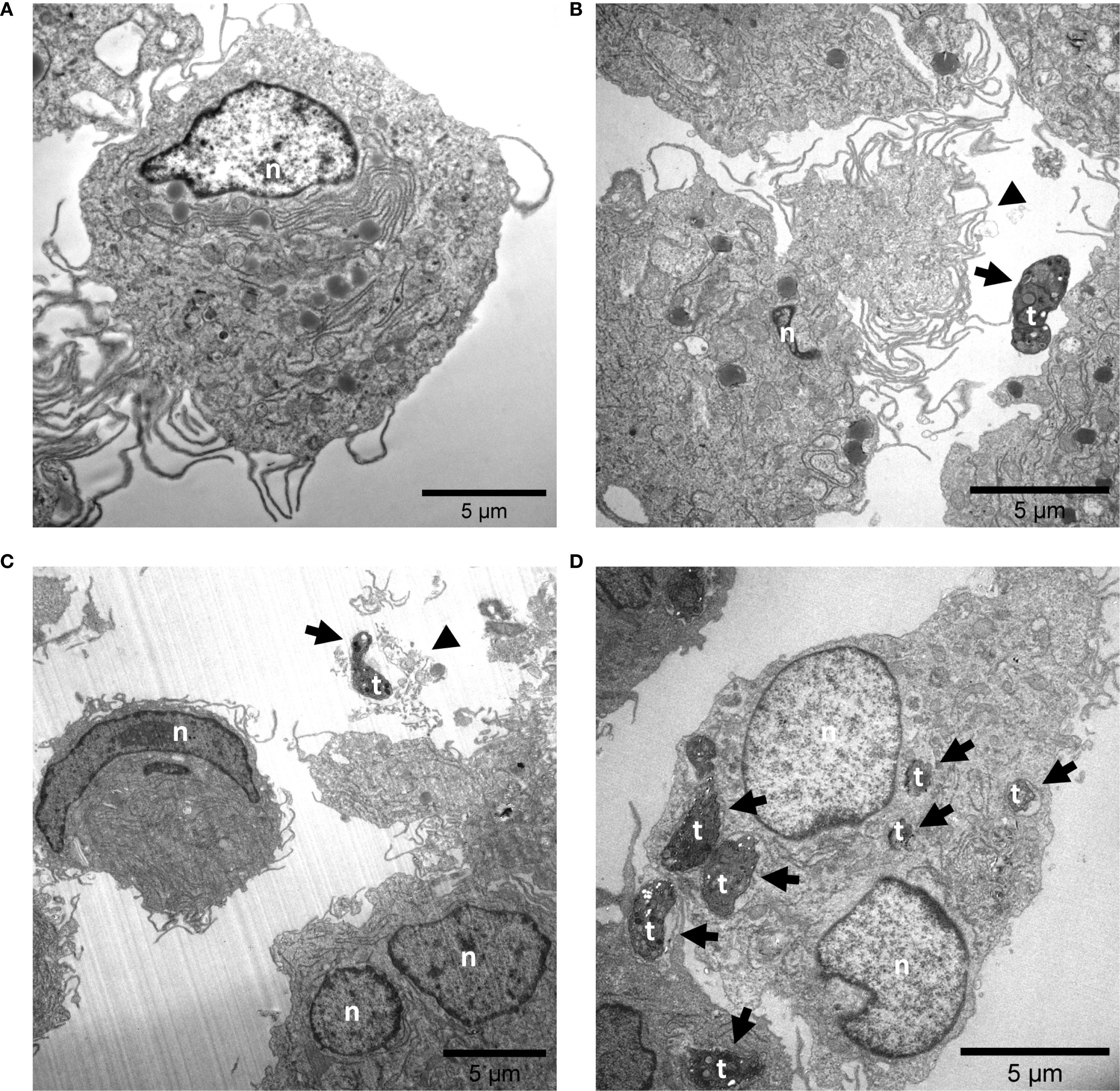
Figure 2. Exposure to Toxoplasma gondii tachyzoite causes the release of ET-like structures by M1-like macrophages. Untreated (A) and T. gondii tachyzoite-exposed M1-like macrophages (MOI 1:4) (B-D) were fixed after 4 h of co-incubation and analyzed by transmission electron microscopy (TEM). The black arrowheads points at regions that might correspond to extracellular trap release from M1-like macrophages (B, C). In addition, several T. gondii tachyzoites were observed in intracellular position (D). Interactions of M1-like macrophages with T. gondii tachyzoites are indicated by black arrows. n, nuclear; t, tachyzoite.
Immunofluorescence microscopy illustrated filamentous structures showing the co-localization of extDNA with classical ET markers like histone and MPO and thereby confirmed METs being released in response to T. gondii tachyzoites from all, primary human and bovine monocytes, monocytic THP-1 cells, and THP-1 cell-derived macrophages (Figures 3A, B; Supplementary Figure S6; Supplementary Figures S7A, B). When quantifying % of MET positive cells, only minor tachyzoite-driven effects were detected (Figures 3A, B). The weak or lack of a significant induction of parasite-driven METs indicated that MET release from primary human and bovine monocytes as well as all kinds of THP-1 cells, was a rare event (Figures 3A, B). Similar results were obtained by using the MET inducers A23187 (5 µM, 4 h) and nigericin (0.5 µM, 4 h) (Supplementary Figures S8A, B) (20, 36).
![Fluorescence microscopy images showing control and T. gondii tachyzoite-treated cells. Panel A compares human- and bovine monocytes (hMo, bMo) under control and treated conditions with arrows indicating extracellular traps (ETs). Panel B features monocytic THP-1 cells and THP-1 cell-derived macrophages (M0-like, M1-like, M2-like) similarly treated, with arrows marking ETs. Adjacent bar graphs quantify MET positive cells [%]. Scale bars indicate 20 micrometers.](https://www.frontiersin.org/files/Articles/1683634/fimmu-16-1683634-HTML/image_m/fimmu-16-1683634-g003.jpg)
Figure 3. Extracellular trap marker detection in co-cultures of Toxoplasma gondii tachyzoites with primary human and bovine monocytes, monocytic THP-1 cells and THP-1 cell-derived macrophages. Primary human (hMo) and bovine monocytes (bMo) (A), monocytic THP-1 cells (monocytic) as well as THP-1 cell-derived M0-, M1-, and M2-like macrophages (MΦ) (B) were left untreated (control) or exposed to T. gondii tachyzoites (+ T. gondii tachyzoites, T.g., MOI 1:4). The cells were fixed after 4 h of incubation and stained for DNA (blue) and histone (red). Extracellular traps, which are characterized by the co-localization of DNA and histones, are indicated by white arrows. Percentage of monocyte/macrophage extracellular trap (MET) positive cells was determined (n = 5). The graphs show mean with standard deviation (SD). Statistical analysis was performed by comparing T. gondii-exposed cells with controls using the non-parametric Wilcoxon signed-rank test; no p-values below p ≤ 0.05 were observed.
In line with the fluorescence microscopic data, neither nuclear area expansion (NAE) estimation (Supplementary Figure S9) nor fluorometric determination and quantification of extDNA (Figure 4) showed a tachyzoite-driven upregulation of METosis in primary human and bovine monocytes, monocytic THP-1 cells, and THP-1 cell-derived macrophages. As expected, stimulation with the positive controls A23187 [5 µM] and nigericin [0.5 µM] (20, 36) significantly induced extDNA release by monocytic THP-1 cells and THP-1 cell-derived macrophages (p ≤ 0.05; n = 5; Figures 4C-F).
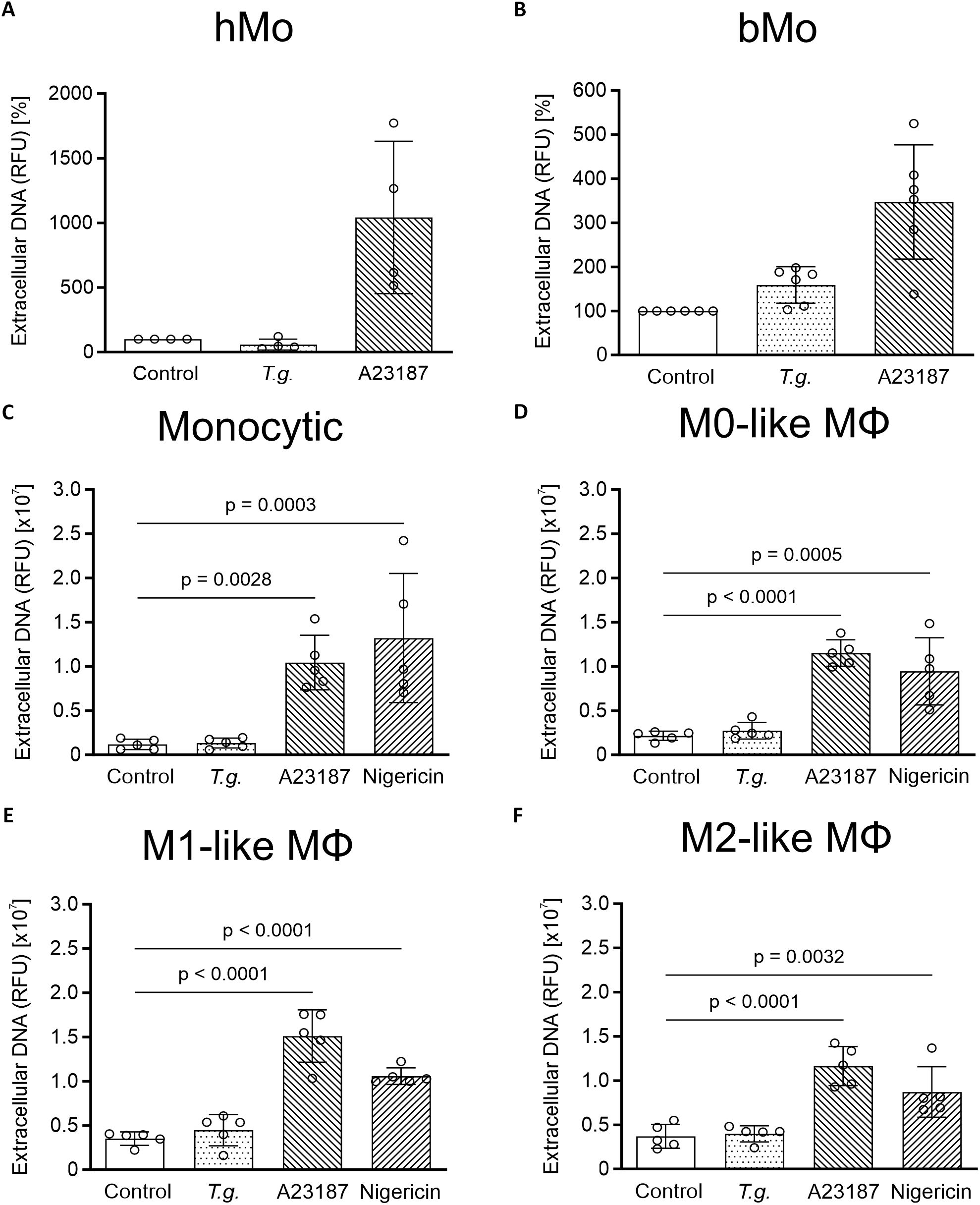
Figure 4. Extracellular DNA in supernatants from Toxoplasma gondii-exposed primary human and bovine monocytes, monocytic THP- cells and THP-1 cell-derived macrophages. Supernatants of untreated (control) primary human (hMo) (A) and bovine monocytes (bMo) (B), monocytic THP-1 cells (monocytic) (C) as well as THP-1 cell-derived M0- (D), M1- (E), and M2-like macrophages (MΦ) (F), T. gondii-exposed cells (T.g., MOI 1:4) or treated cells with A23187 [5 µM] or nigericin [0.5 µM] were collected after 4 h of incubation. Sytox Green was added to the supernatants, followed by spectrophotometric analysis. The amount of extracellular DNA is indicated as relative fluorescence units (RFU) (n = 4 – 6). For both hMo and bMo, controls were normalized to 100% and treatment values were adjusted accordingly to account for inter-donor differences. The graphs show mean with standard deviation (SD) and p-values below 0.05 (p ≤ 0.05). Statistical analysis was performed by comparing T.g.-exposed, A23187- or nigericin-treated cells for primary monocytes using the Friedman test, followed by pairwise comparisons via the Wilcoxon signed-rank test, followed by the Bonferroni correction post-hoc pairwise comparison, and for all kinds of THP-1 cells using a two-way analysis of variance (ANOVA) with residual normality assessed to validate model assumptions, followed by the Bonferroni correction post-hoc pairwise comparison.
Effects of T. gondii tachyzoite exposure on release of IL-1β and IL-6
Activation of mononuclear phagocytes by pathogens can lead to the release of pro-inflammatory cytokines (13). Here, we investigated if the early response of monocytes and macrophages involves secretion of pro-inflammatory cytokines. To determine the release of the pro-inflammatory cytokines IL-1β and IL-6 by primary human monocytes, monocytic THP-1 cells, and THP-1 cell-derived M0-, M1-, and M2-like macrophages were exposed to T. gondii tachyzoites in absence or presence of LPS (Figure 5). We here focused on human mononuclear phagocytes, since commercial ELISAs for the bovine system did not produce plausible results and were therefore discarded (data not shown). Untreated primary human monocytes released low amounts of IL-1β ranging from < 3.9 to 7 pg/ml and IL-6 < 9.4 pg/ml (n = 6) (Figures 5A, B). Similar results were found for untreated monocytic THP-1 cells with IL-1β levels from < 3.9 pg/ml and IL-6 from < 9.4 to 29 pg/ml (n = 9) (Figures 5C, D). T. gondii tachyzoites enhanced the release of IL-1β (from < 3.9 to 98 pg/ml) and of IL-6 (from < 9.4 to 406 pg/ml) by primary human monocytes (p ≤ 0.05; n = 6) (Figures 5A, B) but had no effect on IL-1β and IL-6 secretion by monocytic THP-1 cells (Figures 5C, D). Only in M1-like macrophages, exposure to T. gondii tachyzoites enhanced the release of IL-1β ranging from 13 to 125 pg/ml (p ≤ 0.05; n = 6) but had no effect on IL-6 release (Figures 5G, H).
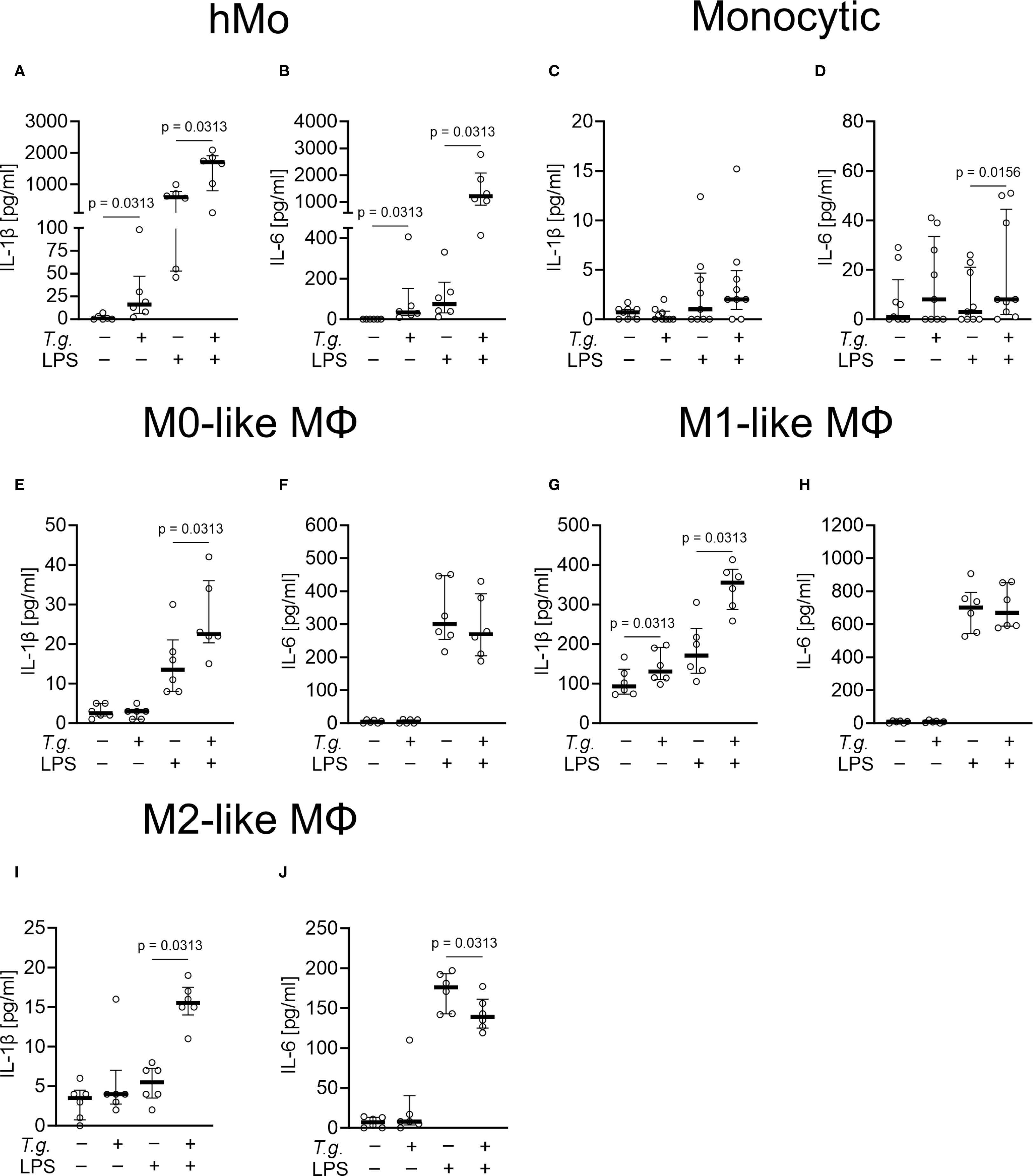
Figure 5. Release of interleukin (IL)-1β and IL-6 by Toxoplasma gondii tachyzoite-exposed primary human monocytes, monocytic THP-1 cells and THP-1 cell-derived macrophages. Primary human monocytes (hMo) (A, B), monocytic THP-1 cells (monocytic) (C, D) as well as THP-1 cell-derived M0- (E, F), M1- (G, H), and M2-like macrophages (MΦ) (I, J) were left untreated (control), exposed to T. gondii tachyzoites (T.g., MOI 1:4) in the absence or presence of LPS [1 µg/ml] or primed with LPS [1 µg/ml] alone for 4 h each (n = 6 – 9). The concentrations of IL-1β (A, C, E, G ,I) and IL-6 (B, D, F, H, J) were measured in cell culture supernatants by ELISA. The graphs show median with the interquartile range (75th and 25th percentiles) and p-values below 0.05 (p ≤ 0.05). Statistical analysis was performed by comparing T.g.-exposed cells with controls or T.g.-exposed/LPS-treated cells with LPS-treated cells using the Friedman test, followed by pairwise comparisons via the Wilcoxon signed-rank test.
Interestingly, exposure to T. gondii tachyzoites induced additive effects in LPS-primed cells. While LPS-primed primary human monocytes released IL-1β ranging from 46 to 996 pg/ml and IL-6 ranging from 11 to 331 pg/ml, T. gondii tachyzoites exposure increased IL-1β release (from 105 to 2087 pg/ml) and IL-6 (from 413 to 2772 pg/ml) (p ≤ 0.05; n = 6) (Figures 5A, B). Similarly, the LPS-induced release of IL-1β by THP-1 cell-derived macrophages (M0-like: ranging from 8 to 30 pg/ml; M1-like: ranging from 105 to 305 pg/ml; M2-like: ranging from < 3.9 to 8 pg/ml) was increased by tachyzoites exposure with the highest levels detected in supernatants of M1-like macrophages (ranging from 156 to 329 pg/ml; p ≤ 0.05; n = 6) (Figures 5E, G, I). Interestingly, no impact of T. gondii tachyzoites exposure was detected on LPS-induced IL-6 release by M0- and M1-like macrophages (Figures 5F, H). A moderate increase of IL-6 release was detected in LPS-primed monocytic THP-1 cells (ranging from < 9.4 to 26 pg/ml; p ≤ 0.05; n = 9) (Figure 5D), whereas a diminished IL-6 secretion was detected in LPS-primed M2-like macrophages (p ≤ 0.05; n = 6) (Figure 5J). Of note, the parasite-driven enhancement of IL-1β release was not due to monocyte or macrophage cell death since LDH values remained low (< 10% increase) in all experimental settings (n = 6 – 9) Supplementary Table S1).
To prove that the used cells responded with IL-1β release to classical NLRP3 activation, stimuli BzATP [200 µM] or nigericin [25 µM] were used as positive controls. For this purpose, the cells were first primed with LPS for 3.5 h followed by exposure for 40 min to BzATP [200 µM] or nigericin [25 µM]. As expected (77), all tested human mononuclear phagocytes responded with an increased release of IL-1β with the highest levels detected in supernatants of M1-like macrophages (ranging from 427.0 to 2899.0 pg/ml; n = 6) (Supplementary Table S2). In addition, we tested the effects of the MET inducers A23187 [1 µM] and nigericin [0.5 µM] after 4 h of incubation (Supplementary Table S2). Of note, high values of cell death were estimated in the presence of A23187 and nigericin with a maximum of 80% cell death in M1-like macrophages treated with A23187 (Supplementary Table S2).
Taken together, the current data suggest that exposure to T. gondii tachyzoites activates primary human monocytes and fosters the release of pro-inflammatory cytokines. We found subtype-specific differences in the THP-1 cells, with THP-1 cell-derived M0- and M1-like macrophages as responding cells to T. gondii tachyzoites exposure.
Tachyzoite exposure does not affect TLR2, TLR4, F-actin, and CD11b expression in primary human and bovine monocytes
It was shown before, that TLR2 and TLR4 expression on bovine neutrophils increased when confronted with the apicomplexan parasite Eimeria bovis (78). In addition, T. gondii-derived GPI proteins showed TNF-α production dependent on TLR2 and TLR4 on mice macrophages (79). Thus, we here evaluated the changes in TLR2 and TLR4 in primary human- and bovine monocytes exposed to T. gondii tachyzoites for 4 h. The results showed no significant differences (p ≥ 0.05, n = 4 – 6) when compared with the unexposed controls (Supplementary Figure S10). In addition, the cellular membrane expression of CD11b and F-actin polymerization, known factors involved in parasite phagocytosis and immune cell migration was analyzed (80). In the context of primary human and bovine monocyte activation by T. gondii exposure no significant changes were observed between the unexposed and the T. gondii-exposed cells (p ≥ 0.05; n = 4 – 6) (Supplementary Figure S10).
Discussion
In the present study, we focused on early immune responses of human and bovine mononuclear phagocytes against vital T. gondii tachyzoites on the level of morphological changes, MET release, pro-inflammatory cytokine secretion and surface receptor expression. In addition to freshly isolated primary human and bovine monocytes, we used monocytic THP-1 cells and THP-1 cell-derived macrophages, that are widely used in research and represent well-accepted in vitro models for human mononuclear phagocytes (30, 32). Despite the critical role of monocytes and macrophages in early innate reactions, investigation on monocyte-derived immune response against protozoan and metazoan parasites is still an understudied topic in both human and veterinary medicine. Thus, their role in the context of toxoplasmosis remains unclear (81). Of note, one report on T. gondii tachyzoite-driven MET formation in harbour seal (Phoca vitulina) exists (18).
Exposure to T. gondii tachyzoites induced morphological changes in investigated monocytes and macrophages, including cytoplasmic spreading, cell surface ruffling, and the formation of membrane protrusions by filopodia and cytonemes, as well as MET formation.
These findings are in line with previous research showing that T. gondii infection triggers significant morphological and functional changes in various mononuclear phagocytes. Across different cell types, including dendritic cells, microglia, and macrophages, tachyzoite invasion consistently induces rapid cytoskeletal remodeling, characterized by podosome loss, cytoplasmic spreading, and the formation of membrane protrusions, which collectively contribute to a hypermigratory phenotype (73–75). The parasite effector TgWIP has been identified as a central mediator of these processes by modulating host cell migration (82), suggesting a conserved strategy by which T. gondii manipulates host immune cell behavior.
Like NETosis (83), the ability of primary monocytes to release METs in response to a variety of stimuli is a well-established effector mechanism (18–21, 23, 24, 26, 27, 29). As key players in the immune defense against toxoplasmosis, macrophages influence the host’s ability to control the infection through their functional polarization (54, 84, 85). MET formation was already reported for monocytic THP-1 cells and THP-1 cell-derived M0-, and M1-like macrophages in response to a range of different stimuli e.g. chemicals, parasite and bacteria (17, 34–41). This is in principle in line with our findings in microscopic analyses (SEM, TEM, and Pappenheim-staining), where amorphous extracellular material was associated with T. gondii tachyzoites exposure, eventually corresponding to MET formation. However, the nuclear membranes of T. gondii-exposed M1-like macrophages remained intact, suggesting that MET formation did not occur in the majority of these cells (86, 87). THP-1 cell-derived macrophages are capable of phagocytosis (30, 88–91) and we found beside extracellular presence, several tachyzoites also intracellularly in M1-like macrophages. However, our analyses cannot distinguish between phagocytosis and active tachyzoite infection. In future studies heat-inactivated T. gondii tachyzoites could be useful to further clarify this.
To unambiguously identify MET formation, we stained extracellular structures in immune cell-parasite co-cultures for classical ET markers e.g. DNA, histones and MPO. For positive controls, the ET-inducing stimulants A23187 and nigericin (20, 36) were used. Immunostaining experiments confirmed the presence of MET positive cells but in contrast to previous findings (20), this was an extremely rare event, even when the known MET inducers A23187 or nigericin were used as positive controls. By using another established spectrofluorometric method for measuring extDNA in cell culture supernatants (92), exposure to T. gondii showed no changes in all tested cells. Furthermore, A23187 treatment resulted in elevated levels of cell death, as indicated by increased LDH activity in the corresponding cell-free supernatants. Consequently, it is not possible to clearly distinguish between the effects attributable to MET formation and those resulting from cell death. In this context, Matsubara et al., 1994 have shown in six human leukemia cell lines that exposure to A23187 [1 μM] induced cell apoptosis within 3 h (93), indicating that at least A23187 should be used as a positive control for MET formation with caution. We and others (20, 24, 36) used even higher concentrations and a longer exposure time. To further substantiate our findings, we next measured NAE, an established method to estimate the very early phase of ETosis (61, 94). Again, neither T. gondii exposure nor stimulation with A23187 or nigericin enhanced NAE and, thus, no significant induction of METosis was quantifiable.
In summary, our findings indicate that MET formation is a rather rare event and therefore most probably of minor importance in the early response of mononuclear phagocytes to T. gondii tachyzoites. Obviously, these data underline immune cell type-specific responses, since T. gondii tachyzoites were recently proven as potent ET inducers in primary neutrophils of human, bovine, murine, cat, dog, sheep, goat, bottlenose dolphin, and harbour seal origin (18, 87, 95–99).
As mentioned above, the role of IL-1β and IL-6 in T. gondii-driven immune responses is controversial. In previous reports, the exposure of primary human monocytes to T. gondii tachyzoites after 4, 16 and 24 h induced the release of IL-1β (53, 55, 57, 58) but not of IL-6 (59). T. gondii tachyzoite-induced release of IL-1β (53, 55, 64) or IL-6 (62, 63) after 4, 8, 12, 24 h by monocytic THP-1 cells and THP-1 cell-derived M0-like macrophages was consistently demonstrated.
To gain further insights into monocyte- and macrophage-mediated immune responses during early T. gondii infection, we next examined the release of IL-1β and IL-6. In addition to primary human monocytes, we performed experiments on the THP-1 cell line, allowing controlled culture and treatment conditions. In line with previous studies (53, 55, 62–64), we found that exposure to T. gondii tachyzoites activates human mononuclear phagocytes and fosters the release of pro-inflammatory cytokines. Importantly, we found differences within the THP-1 cell subtypes. The THP-1 cell-derived M1-like macrophages reacted by an increased release of IL-1β following exposure to T. gondii, and this effect was enhanced by addition of LPS. These data might indicate that parasite phagocytosis may result in a more pronounced cytokine response in M1-like macrophages, which is in accordance to previous studies (100, 101). In LPS-primed M2-like macrophages, a subtle but statistically significant decrease in IL-6 release was observed. A similar effect was described before for LPS-primed M0-like macrophages after stimulation with T. gondii lysate (102). Moreover, Salazar Gonzalez et al., 2014 showed that T. gondii profilin induced the release of IL-6 by primary human monocytes (61). A possible explanation for the contrasting findings could be the different experiment conditions (e.g. exposure times and MOI) used in different studies.
In line with previous studies (64, 98), here, we investigated the early responses of mononuclear phagocytes after 4 h of parasite exposure applying MOI of 1:4. Other studies observed an increased release of IL-1β by monocytic THP-1 cells after 12 h (MOI 1:2) and 18 h (MOI 1:3) (53, 55) or by M0-like macrophages after 4 h (MOI 1:10) (64) exposure to the parasite. The use of different parasite strains is not expected to account for these differences, as the same T. gondii RH strain was used (55, 64). Investigating higher MOIs in combination with variable exposure times could, however, in future studies provide more comprehensive insight into innate host responses. As mentioned before, the release of IL-1β typically requires a priming phase (to induce pro-IL-1β expression) followed by a second stimulus (DAMPs, PAMPs) for cleavage and release of the mature IL-1β (42–45). This could be an explanation for the observed enhanced IL-1β release in LPS-primed M1-like macrophages. Studies using longer exposure times (53, 55) or higher infection rates (64) might favor the release of other DAMPs, which eventually results in activated IL-1β secretion. Contamination of mononuclear phagocytes with LPS or damage-induced priming during the isolation process of primary cells might also pre-activate mononuclear phagocytes (103) and, thus, result in differences in their sensitivity to T. gondii tachyzoites. To validate the immunological activity of mononuclear phagocytes, we here included control experiments that assessed their responsiveness to well-characterized stimuli. By priming cells with LPS and subsequently treating them with a second stimulus (BzATP or nigericin), we induced cytokine secretion and evaluated the functional capacity of the cells. One of the key control experiments involved the use of the P2X7 receptor agonist BzATP and the pore-forming reagent nigericin to induce NLRP3 inflammasome activation. The NLRP3 inflammasome is a multi-protein complex that plays a crucial role in the innate immune response by promoting the maturation and secretion of pro-inflammatory cytokines, such as IL-1β (104). Activation of the NLRP3 inflammasome requires two signals: a priming signal, typically provided by PRRs agonists like LPS, and a second signal, which can be delivered by various DAMPs or PAMPs (42–45). BzATP, a stable adenosine triphosphate (ATP) analog, acts as an agonist for the P2X7 receptor, which is known to mediate NLRP3 inflammasome activation through potassium efflux (105). Nigericin, on the other hand, is a potassium ionophore that disrupts intracellular potassium homeostasis, leading to NLRP3 inflammasome assembly and activation (106). In current control experiments, both BzATP and nigericin reliably induced NLRP3 inflammasome-dependent IL-1β release, demonstrating the functional integrity of the mononuclear phagocytes used in our study. The successful induction of IL-1β secretion by LPS and BzATP thereby confirmed the immunological activity of the cells and the validity of the current experimental approach. Furthermore, in all experiments LDH levels were determined to test for cell death.
TLR2 and TLR4 were both shown to be involved in the immune response against T. gondii. While TLR2 was shown to recognize the parasitic surface profilin or GPI proteins and stimulating the production of IL-12, TLR4 rather plays a supportive role, especially at high infectious doses, as summarized for several mouse models (47, 48, 79). We further determined the expression levels of F-actin and CD11b. F-actin is part of the cytoskeleton and known to be involved in phagocytosis (106). CD11b plays a critical role in the migration and adhesion of immune cells, including monocytes (80). In T. gondii-infected primary human monocytes, a significant increase in the expression of CD11b in the high-affinity form of macrophage-1 antigen (Mac-1) was observed, which promotes increased crawling distance and transmigration across endothelial barriers, thus supporting the spread of the parasite in the host (80). In the current study, we did, however, not find any effects of T. gondii tachyzoite exposure on TLR2, TLR4, F-actin or CD11b expression by primary human and bovine monocytes. However, the lack of distinction between infected and uninfected cells complicates the interpretation of surface marker expression and represents a limitation of this study.
Taken together, we here characterized the early innate immune responses of human and bovine mononuclear phagocytes to vital T. gondii tachyzoites, providing valuable insights into the initial interactions between this zoonotic parasite and professional phagocytes. Our findings demonstrate that exposure to T. gondii tachyzoites very early activates monocytes and macrophages, leading to the release of pro-inflammatory cytokines IL-1β and IL-6, particularly when co-stimulated with LPS. This cytokine response appeared more important than the formation of METs, which was observed as a rare event. The absence of significant changes in the expression of key markers (TLR2, TLR4, F-actin, and CD11b) by primary monocytes upon exposure to T. gondii suggests that the early immune response is primarily mediated through cytokine secretion rather than by changes on cell surface receptor expression or cytoskeletal rearrangements. These results highlight the importance of cytokine signaling in the early innate immune response to T. gondii and provide a foundation for further investigations into the molecular mechanisms underlying this response. Future studies should focus on elucidating the signaling pathways involved in cytokine production and exploring the potential therapeutic targeting of these pathways to modulate the immune response against T. gondii infection. Further studies using standardized methods are needed to clarify the role of METs in advanced toxoplasmosis and their possible contribution to immune-related tissue damage in vivo, as reported for NET extrusion.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
All procedures on primary human cells were performed in accordance with the Helsinki Declaration and in compliance with relevant laws and institutional guidelines and have been approved by the local ethics committee at the Justus Liebig University (JLU) Giessen (Giessen, Germany, approval No. 90/18, No. 98/23 and No. 05/2000). The privacy rights of the volunteers have been observed and written informed consent was given by each volunteer. Regarding cattle blood sampling, all procedures were conducted following the guidelines of the JLU Giessen Animal Care Committee. Protocols were approved by the Ethics Commission for Experimental Animal Studies of the Federal State of Hesse (Regierungspräsidium Giessen; GI 18/10 Nr. V 2/2022; JLU-No. 0002_V) and are in accordance with European Animal Welfare Legislation: ART13TFEU, and currently applicable German Animal Protection Laws.
Author contributions
DH: Investigation, Methodology, Writing – original draft, Visualization, Formal analysis, Data curation. ZV: Writing – review & editing, Visualization, Methodology. KB: Validation, Writing – review & editing, Formal analysis. AK: Writing – review & editing, Resources. RR: Writing – review & editing, Visualization, Validation, Methodology. AH: Writing – review & editing, Resources. SM: Funding acquisition, Supervision, Writing – review & editing. VG: Writing – review & editing, Resources, Conceptualization. AT: Conceptualization, Supervision, Writing – review & editing, Resources, Funding acquisition. CH: Supervision, Conceptualization, Writing – review & editing, Resources, Funding acquisition. KR: Formal analysis, Visualization, Data curation, Validation, Methodology, Supervision, Conceptualization, Investigation, Writing – original draft. IC: Visualization, Data curation, Writing – original draft, Formal analysis, Conceptualization, Methodology, Validation, Investigation, Supervision.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. KR was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation, Project-ID 515250365 and Project-ID 528562393-FIP 26).
Acknowledgments
We are grateful to Bianca Kulik, Gabriele Fuchs-Moll, Hannah Salecker, Sabine Stumpf, Sandra Ruehl, Stefan Scheldt and Kathrin Petri (Justus Liebig University Giessen, Giessen, Germany) for excellent technical support; Annika Seipp and Ulrich Gaertner (Justus Liebig University Giessen, Giessen, Germany) for their excellent technical support in scanning- and transmission electron microscopic analyses; Ulrich J. Sachs (Institute of Clinical Immunology, Transfusion Medicine, and Hemostasis, Justus Liebig University Giessen, Giessen, Germany) for providing buffy coats from healthy blood donors; and to Jens Arne Bodenbender as well as the staff of the Upper Hardthof Livestock Teaching and Research Facility (Justus Liebig University Giessen, Germany) for their diligent routine dairy cattle maintenance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1683634/full#supplementary-material
References
1. Hill DE and Dubey JP. Toxoplasma gondii as a parasite in food: analysis and control. Microbiol Spectr. (2016) 4. doi: 10.1128/microbiolspec.PFS-0011-2015
2. Dubey JP. Outbreaks of clinical toxoplasmosis in humans: five decades of personal experience, perspectives and lessons learned. Parasit Vectors. (2021) 14:263. doi: 10.1186/s13071-021-04769-4
3. Dubey JP, Lindsay DS, and Speer CA. Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clin Microbiol Rev. (1998) 11:267–99. doi: 10.1128/CMR.11.2.267
4. Ehmen HG and Lüder CGK. Long-term impact of Toxoplasma gondii infection on human monocytes. Front Cell Infect Microbiol. (2019) 9:235. doi: 10.3389/fcimb.2019.00235
5. Attias M, Teixeira DE, Benchimol M, Vommaro RC, Crepaldi PH, and De Souza W. The life-cycle of Toxoplasma gondii reviewed using animations. Parasit Vectors. (2020) 13:588. doi: 10.1186/s13071-020-04445-z
6. Masek KS and Hunter CA. Pro-inflammatory responses in macrophages during Toxoplasma gondii infection. In: Madame Curie Bioscience Database. Austin, TX, USA: Landes Bioscience (2013). Available online at: https://www.ncbi.nlm.nih.gov/books/NBK5976/.
7. Orchanian SB and Lodoen MB. Monocytes as primary defenders against Toxoplasma gondii infection. Trends Parasitol. (2023) 39:837–49. doi: 10.1016/j.pt.2023.07.007
8. Bitzmann D. Ultrastrukturelle, immunzytochemische und glykohistochemische Untersuchungen am Blut des Rindes (Bos tauris). München, Germany: Ludwig-Maximilians-Universität München (2009).
9. Guilliams M, Mildner A, and Yona S. Developmental and functional heterogeneity of monocytes. Immunity. (2018) 49:595–613. doi: 10.1016/j.immuni.2018.10.005
10. Guilliams M, Ginhoux F, Jakubzick C, Naik SH, Onai N, Schraml BU, et al. Dendritic cells, monocytes and macrophages: a unified nomenclature based on ontogeny. Nat Rev Immunol. (2014) 14:571–8. doi: 10.1038/nri3712
11. Deshmane SL, Kremlev S, Amini S, and Sawaya BE. Monocyte chemoattractant protein-1 (MCP-1): an overview. J Interferon Cytokine Res. (2009) 29:313–26. doi: 10.1089/jir.2008.0027
12. Jiang W and Gilkeson G. Sex differences in monocytes and TLR4 associated immune responses; implications for systemic lupus erythematosus (SLE). J Immunother Appl. (2014) 1:1. doi: 10.7243/2055-2394-1-1
13. Chiu S and Bharat A. Role of monocytes and macrophages in regulating immune response following lung transplantation. Curr Opin Organ Transplant. (2016) 21:239–45. doi: 10.1097/MOT.0000000000000313
14. Jiménez-Dalmaroni MJ, Gerswhin ME, and Adamopoulos IE. The critical role of Toll-like receptors - from microbial recognition to autoimmunity: a comprehensive review. Autoimmun Rev. (2016) 15:1–8. doi: 10.1016/j.autrev.2015.08.009
15. Torres-Ruiz J, Carrillo-Vazquez DA, Padilla-Ortiz DM, Vazquez-Rodriguez R, Nuñez-Alvarez C, Juarez-Vega G, et al. TLR expression in peripheral monocyte subsets of patients with idiopathic inflammatory myopathies: association with clinical and immunological features. J Transl Med. (2020) 18:125. doi: 10.1186/s12967-020-02290-3
16. Richter K, Asci N, Singh VK, Yakoob SH, Meixner M, Zakrzewicz A, et al. Activation of endothelial NO synthase and P2X7 receptor modification mediates the cholinergic control of ATP-induced interleukin-1β release by mononuclear phagocytes. Front Immunol. (2023) 14:1140592. doi: 10.3389/fimmu.2023.1140592
17. Aulik NA, Hellenbrand KM, and Czuprynski CJ. Mannheimia haemolytica and its leukotoxin cause macrophage extracellular trap formation by bovine macrophages. Infect Immun. (2012) 80:1923–33. doi: 10.1128/IAI.06120-11
18. Reichel M, Muñoz-Caro T, Sanchez Contreras G, Rubio García A, Magdowski G, Gärtner U, et al. Harbour seal (Phoca vitulina) PMN and monocytes release extracellular traps to capture the apicomplexan parasite Toxoplasma gondii. Dev Comp Immunol. (2015) 50:106–15. doi: 10.1016/j.dci.2015.02.002
19. Pérez D, Muñoz MC, Molina JM, Muñoz-Caro T, Silva LMR, Taubert A, et al. Eimeria ninakohlyakimovae induces NADPH oxidase-dependent monocyte extracellular trap formation and upregulates IL-12 and TNF-α, IL-6 and CCL2 gene transcription. Vet Parasitol. (2016) 227:143–50. doi: 10.1016/j.vetpar.2016.07.028
20. Granger V, Faille D, Marani V, Noël B, Gallais Y, Szely N, et al. Human blood monocytes are able to form extracellular traps. J Leukoc Biol. (2017) 102:775–81. doi: 10.1189/jlb.3MA0916-411R
21. Yang Z, Wei Z, Hermosilla C, Taubert A, He X, Wang X, et al. Caprine monocytes release extracellular traps against Neospora caninum in vitro. Front Immunol. (2018) 8:2016. doi: 10.3389/fimmu.2017.02016
22. Weng W, Hu Z, and Pan Y. Macrophage extracellular traps: current opinions and the state of research regarding various diseases. J Immunol Res. (2022) 2022:7050807. doi: 10.1155/2022/7050807
23. Hasheminasab SS, Conejeros I, Gärtner U, Kamena F, Taubert A, and Hermosilla CR. MCT-dependent Cryptosporidium parvum-induced bovine monocyte extracellular traps (METs) under physioxia. Biology. (2023) 12:961. doi: 10.3390/biology12070961
24. Ibrahim N, Knöbl V, Hayden H, Bauer WM, Worel N, Neumayer C, et al. Human monocyte subsets differ in their capacity to form extracellular traps. Cell Death Discov. (2024) 10:1–3. doi: 10.1038/s41420-024-02034-y
25. Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, et al. Neutrophil extracellular traps kill bacteria. Science. (2004) 303:1532–5. doi: 10.1126/science.1092385
26. Muñoz-Caro T, Silva LMR, Ritter C, Taubert A, and Hermosilla C. Besnoitia besnoiti tachyzoites induce monocyte extracellular trap formation. Parasitol Res. (2014) 113:4189–97. doi: 10.1007/s00436-014-4094-3
27. Schulz M, Zambrano F, Schuppe H-C, Wagenlehner F, Taubert A, Gaertner U, et al. Monocyte-derived extracellular trap (MET) formation induces aggregation and affects motility of human spermatozoa. vitro Syst Biol Reprod Med. (2019) 65:357–66. doi: 10.1080/19396368.2019.1624873
28. Neumann A, Brogden G, and von Köckritz-Blickwede M. Extracellular traps: an ancient weapon of multiple kingdoms. Biology. (2020) 9:34. doi: 10.3390/biology9020034
29. Bartneck M, Keul HA, Zwadlo-Klarwasser G, and Groll J. Phagocytosis independent extracellular nanoparticle clearance by human immune cells. Nano Lett. (2010) 10:59–63. doi: 10.1021/nl902830x
30. Chanput W, Mes JJ, and Wichers HJ. THP-1 cell line: an in vitro cell model for immune modulation approach. Int Immunopharmacol. (2014) 23:37–45. doi: 10.1016/j.intimp.2014.08.002
31. Genin M, Clement F, Fattaccioli A, Raes M, and Michiels C. M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer. (2015) 15:577. doi: 10.1186/s12885-015-1546-9
32. Mohd Yasin ZN, Mohd Idrus FN, Hoe CH, and Yvonne-Tee GB. Macrophage polarization in THP-1 cell line and primary monocytes: a systematic review. Differentiation. (2022) 128:67–82. doi: 10.1016/j.diff.2022.10.001
33. Murray PJ and Wynn TA. Protective and pathogenic functions of macrophage subsets. Nat Rev Immunol. (2011) 11:723. doi: 10.1038/nri3073
34. Nakayama T and Saitoh H. Tunicamycin-induced neutrophil extracellular trap (NET)-like structures in cultured human myeloid cell lines. Cell Biol Int. (2015) 39:355–9. doi: 10.1002/cbin.10396
35. Henthorn CR, Chris Minion F, and Sahin O. Utilization of macrophage extracellular trap nucleotides by Mycoplasma hyopneumoniae. Microbiol Read Engl. (2018) 164:1394–404. doi: 10.1099/mic.0.000717
36. Jensen M, Thorsen NW, Hallberg LAE, Hägglund P, and Hawkins CL. New insight into the composition of extracellular traps released by macrophages exposed to different types of inducers. Free Radic Biol Med. (2023) 202:97–109. doi: 10.1016/j.freeradbiomed.2023.03.025
37. Fei L, Zhengkai W, Weina J, Lili C, Yuhang G, Zhengtao Y, et al. Trichomonas vaginalis triggers the release of THP-1 extracellular traps. Parasitol Res. (2019) 118:267–74. doi: 10.1007/s00436-018-6139-5
38. Lee Y, Reilly B, Tan C, Wang P, and Aziz M. Extracellular CIRP induces macrophage extracellular trap formation via gasdermin D activation. Front Immunol. (2021) 12:780210. doi: 10.3389/fimmu.2021.780210
39. An Y, Shi X, Tang X, Wang Y, Shen F, Zhang Q, et al. Aflatoxin B1 induces reactive oxygen species-mediated autophagy and extracellular trap formation in macrophages. Front Cell Infect Microbiol. (2017) 7:53. doi: 10.3389/fcimb.2017.00053
40. Doster RS, Sutton JA, Rogers LM, Aronoff DM, and Gaddy JA. Streptococcus agalactiae induces placental macrophages to release extracellular traps loaded with tissue remodeling enzymes via an oxidative burst-dependent mechanism. mBio. (2018) 9:e02084–18. doi: 10.1128/mBio.02084-18
41. Je S, Quan H, Yoon Y, Na Y, Kim B-J, and Seok SH. Mycobacterium massiliense induces macrophage extracellular traps with facilitating bacterial growth. PloS One. (2016) 11:e0155685. doi: 10.1371/journal.pone.0155685
42. Beurel E and Jope RS. Lipopolysaccharide-induced interleukin-6 production is controlled by glycogen synthase kinase-3 and STAT3 in the brain. J Neuroinflamm. (2009) 6:9. doi: 10.1186/1742-2094-6-9
43. Kawai T and Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. (2010) 11:373–84. doi: 10.1038/ni.1863
44. Dinarello CA. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol Rev. (2018) 281:8–27. doi: 10.1111/imr.12621
45. Mantovani A, Dinarello CA, Molgora M, and Garlanda C. Interleukin-1 and related cytokines in the regulation of inflammation and immunity. Immunity. (2019) 50:778–95. doi: 10.1016/j.immuni.2019.03.012
46. Broz P and Dixit VM. Inflammasomes: mechanism of assembly, regulation and signalling. Nat Rev Immunol. (2016) 16:407–20. doi: 10.1038/nri.2016.58
47. Yarovinsky F and Sher A. Toll-like receptor recognition of Toxoplasma gondii. Int J Parasitol. (2006) 36:255–9. doi: 10.1016/j.ijpara.2005.12.003
48. Walseng E, Furuta K, Goldszmid RS, Weih KA, Sher A, and Roche PA. Dendritic cell activation prevents MHC class II ubiquitination and promotes MHC class II survival regardless of the activation stimulus. J Biol Chem. (2010) 285:41749–54. doi: 10.1074/jbc.M110.157586
49. Saeij JPJ, Boyle JP, and Boothroyd JC. Differences among the three major strains of Toxoplasma gondii and their specific interactions with the infected host. Trends Parasitol. (2005) 21:476–81. doi: 10.1016/j.pt.2005.08.001
50. Sanchez SG and Besteiro S. The pathogenicity and virulence of Toxoplasma gondii. Virulence. (2021) 12:3095–114. doi: 10.1080/21505594.2021.2012346
51. Saeij JPJ, Coller S, Boyle JP, Jerome ME, White MW, and Boothroyd JC. Toxoplasma co-opts host gene expression by injection of a polymorphic kinase homologue. Nature. (2007) 445:324–7. doi: 10.1038/nature05395
52. Rosowski EE, Lu D, Julien L, Rodda L, Gaiser RA, Jensen KDC, et al. Strain-specific activation of the NF-κB pathway by GRA15, a novel Toxoplasma gondii dense granule protein. J Exp Med. (2011) 208:195–212. doi: 10.1084/jem.20100717
53. Gov L, Karimzadeh A, Ueno N, and Lodoen MB. Human innate immunity to Toxoplasma gondii is mediated by host caspase-1 and ASC and parasite GRA15. mBio. (2013) 4:e00255–13. doi: 10.1128/mBio.00255-13
54. Guimaraes Gois PS, Franco PS, Cota Teixeira S, Guirelli PM, de Araujo TE, da Fonseca Batistao DW, et al. Polarisation of human macrophages towards an M1 subtype triggered by an atypical Brazilian strain of Toxoplasma gondii results in a reduction in parasite burden. Folia Parasitol (Praha). (2022) 69:2022.020. doi: 10.14411/fp.2022.020
55. Gov L, Schneider CA, Lima TS, Pandori W, and Lodoen MB. NLRP3 and potassium efflux drive rapid IL-1β release from primary human monocytes during Toxoplasma gondii infection. J Immunol Baltim Md 1950. (2017) 199:2855. doi: 10.4049/jimmunol.1700245
56. Lima TS, Gov L, and Lodoen MB. Evasion of human neutrophil-mediated host defense during Toxoplasma gondii infection. mBio. (2018) 9:e02027–17. doi: 10.1128/mBio.02027-17
57. Pandori WJ, Lima TS, Mallya S, Kao TH, Gov L, and Lodoen MB. Toxoplasma gondii activates a Syk-CARD9-NF-κB signaling axis and gasdermin D-independent release of IL-1β during infection of primary human monocytes. PloS Pathog. (2019) 15:e1007923. doi: 10.1371/journal.ppat.1007923
58. Pandori WJ, Matsuno SY, Shin J-H, Kim SC, Kao TH, Mallya S, et al. Role for caspase-8 in the release of IL-1β and active caspase-1 from viable human monocytes during Toxoplasma gondii infection. J Immunol. (2024) 212:1161–71. doi: 10.4049/jimmunol.2200513
59. Pelloux H, Pernod G, Ricard J, Renversez TC, and Ambroise-Thomas P. Interleukin-6 is secreted by human monocytes after stimulation with anti-Toxoplasma gondii sera. J Infect Dis. (1994) 169:1181–2. doi: 10.1093/infdis/169.5.1181
60. Tombácz K, Burgess G, Holder A, Werners A, and Werling D. Toxoplasma gondii profilin does not stimulate an innate immune response through bovine or human TLR5. Innate Immun. (2018) 24:422–9. doi: 10.1177/1753425918798450
61. Salazar Gonzalez RM, Shehata H, O’Connell MJ, Yang Y, Moreno-Fernandez ME, Chougnet CA, et al. Toxoplasma gondii-derived profilin triggers human Toll-like receptor 5-dependent cytokine production. J Innate Immun. (2014) 6:685–94. doi: 10.1159/000362367
62. Friedland JS, Shattock RJ, Johnson JD, Remick DG, Holliman RE, and Griffin GE. Differential cytokine gene expression and secretion after phagocytosis by a human monocytic cell line of Toxoplasma gondii compared with Mycobacterium tuberculosis. Clin Exp Immunol. (1993) 91:282–6. doi: 10.1111/j.1365-2249.1993.tb05896.x
63. Castro AS, Alves CMOS, Angeloni MB, Gomes AO, Barbosa BF, Franco PS, et al. Trophoblast cells are able to regulate monocyte activity to control Toxoplasma gondii infection. Placenta. (2013) 34:240–7. doi: 10.1016/j.placenta.2012.12.006
64. Chu J-Q, Shi G, Fan Y-M, Choi I-W, Cha G-H, Zhou Y, et al. Production of IL-1β and inflammasome with up-regulated expressions of NOD-like receptor related genes in Toxoplasma gondii-infected THP-1 macrophages. Korean J Parasitol. (2016) 54:711–7. doi: 10.3347/kjp.2016.54.6.711
65. Daigneault M, Preston JA, Marriott HM, Whyte MKB, and Dockrell DH. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PloS One. (2010) 5:e8668. doi: 10.1371/journal.pone.0008668
66. Neubert E, Senger-Sander SN, Manzke VS, Busse J, Polo E, Scheidmann SEF, et al. Serum and serum albumin inhibit in vitro formation of neutrophil extracellular traps (NETs). Front Immunol. (2019) 10:12. doi: 10.3389/fimmu.2019.00012
67. Ross R, Hasheminasab SS, Conejeros I, Gärtner U, Kamena F, Krueger A, et al. Human dendritic cell interactions with the zoonotic parasite Cryptosporidium parvum result in activation and maturation. Front Immunol. (2024) 15:1388366. doi: 10.3389/fimmu.2024.1388366
68. Goddeeris BM, Baldwin CL, ole-MoiYoi O, and Morrison WI. Improved methods for purification and depletion of monocytes from bovine peripheral blood mononuclear cells. Functional evaluation of monocytes in responses to lectins. J Immunol Methods. (1986) 89:165–73. doi: 10.1016/0022-1759(86)90354-6
69. Ceciliani F, Ávila Morales G, De Matteis G, Grandoni F, Furioso Ferreira R, Roccabianca P, et al. Methods in isolation and characterization of bovine monocytes and macrophages. Methods San Diego Calif. (2021) 186:22–41. doi: 10.1016/j.ymeth.2020.06.017
70. Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, et al. Fiji: an open-source platform for biological-image analysis. Nat Methods. (2012) 9:676–82. doi: 10.1038/nmeth.2019
71. Behrendt JH, Hermosilla C, Hardt M, Failing K, Zahner H, and Taubert A. PMN-mediated immune reactions against Eimeria bovis. Vet Parasitol. (2008) 151:97–109. doi: 10.1016/j.vetpar.2007.11.013
72. Bumol TF and Douglas SD. Human monocyte cytoplasmic spreading in vitro: early kinetics and scanning electron microscopy. Cell Immunol. (1977) 34:70–8. doi: 10.1016/0008-8749(77)90230-1
73. Weidner JM, Kanatani S, Hernández-Castañeda MA, Fuks JM, Rethi B, Wallin RPA, et al. Rapid cytoskeleton remodelling in dendritic cells following invasion by Toxoplasma gondii coincides with the onset of a hypermigratory phenotype. Cell Microbiol. (2013) 15:1735–52. doi: 10.1111/cmi.12145
74. Bhandage AK, Kanatani S, and Barragan A. Toxoplasma-induced hypermigration of primary cortical microglia implicates GABAergic signaling. Front Cell Infect Microbiol. (2019) 9:73. doi: 10.3389/fcimb.2019.00073
75. García-Sánchez M, Jiménez-Pelayo L, Horcajo P, Regidor-Cerrillo J, Ólafsson EB, Bhandage AK, et al. Differential responses of bovine monocyte-derived macrophages to infection by Neospora caninum isolates of high and low virulence. Front Immunol. (2019) 10:915. doi: 10.3389/fimmu.2019.00915
76. Perrotta I, Carito V, Russo E, Tripepi S, Aquila S, and Donato G. Macrophage autophagy and oxidative stress: an ultrastructural and immunoelectron microscopical study. Oxid Med Cell Longev. (2011) 2011:282739. doi: 10.1155/2011/282739
77. Murray RZ and Stow JL. Cytokine secretion in macrophages: SNAREs, Rabs, and membrane trafficking. Front Immunol. (2014) 5:538. doi: 10.3389/fimmu.2014.00538
78. Muñoz-Caro T, Gibson AJ, Conejeros I, Werling D, Taubert A, and Hermosilla C. The role of TLR2 and TLR4 in recognition and uptake of the apicomplexan parasite Eimeria bovis and their effects on NET formation. Pathogens. (2021) 10:118. doi: 10.3390/pathogens10020118
79. Debierre-Grockiego F, Campos MA, Azzouz N, Schmidt J, Bieker U, Resende MG, et al. Activation of TLR2 and TLR4 by glycosylphosphatidylinositols derived from Toxoplasma gondii. J Immunol Baltim Md 1950. (2007) 179:1129–37. doi: 10.4049/jimmunol.179.2.1129
80. Ueno N, Harker KS, Clarke EV, McWhorter FY, Liu WF, Tenner AJ, et al. Real-time imaging of Toxoplasma-infected human monocytes under fluidic shear stress reveals rapid translocation of intracellular parasites across endothelial barriers. Cell Microbiol. (2014) 16:580–95. doi: 10.1111/cmi.12239
81. Geissmann F, Manz MG, Jung S, Sieweke MH, Merad M, and Ley K. Development of monocytes, macrophages, and dendritic cells. Science. (2010) 327:656–61. doi: 10.1126/science.1178331
82. Sangaré LO, Ólafsson EB, Wang Y, Yang N, Julien L, Camejo A, et al. In vivo CRISPR screen identifies TgWIP as a Toxoplasma modulator of dendritic cell migration. Cell Host Microbe. (2019) 26:478–492.e8. doi: 10.1016/j.chom.2019.09.008
83. Silva LMR, Muñoz-Caro T, Burgos RA, Hidalgo MA, Taubert A, and Hermosilla C. Far beyond phagocytosis: phagocyte-derived extracellular traps act efficiently against protozoan parasites in vitro and in vivo. Mediators Inflammation. (2016) 2016:5898074. doi: 10.1155/2016/5898074
84. Jensen KDC, Wang Y, Wojno EDT, Shastri AJ, Hu K, Cornel L, et al. Toxoplasma polymorphic effectors determine macrophage polarization and intestinal inflammation. Cell Host Microbe. (2011) 9:472–83. doi: 10.1016/j.chom.2011.04.015
85. Park J and Hunter CA. The role of macrophages in protective and pathological responses to Toxoplasma gondii. Parasite Immunol. (2020) 42:e12712. doi: 10.1111/pim.12712
86. von Köckritz-Blickwede M, Goldmann O, Thulin P, Heinemann K, Norrby-Teglund A, Rohde M, et al. Phagocytosis-independent antimicrobial activity of mast cells by means of extracellular trap formation. Blood. (2008) 111:3070–80. doi: 10.1182/blood-2007-07-104018
87. Velásquez ZD, Peixoto R, Gärtner U, Hermosilla C, Taubert A, and Conejeros I. Dynamics of cell cycle proteins involved in Toxoplasma gondii-induced bovine NET formation. Front Immunol. (2023) 14:1125667. doi: 10.3389/fimmu.2023.1125667
88. Kurynina AV, Erokhina MV, Makarevich OA, Sysoeva VY, Lepekha LN, Kuznetsov SA, et al. Plasticity of human THP-1 cell phagocytic activity during macrophagic differentiation. Biochem Biokhimiia. (2018) 83:200–14. doi: 10.1134/S0006297918030021
89. Herdoiza Padilla E, Crauwels P, Bergner T, Wiederspohn N, Förstner S, Rinas R, et al. mir-124-5p regulates phagocytosis of human macrophages by targeting the actin cytoskeleton via the ARP2/3 complex. Front Immunol. (2019) 10:2210. doi: 10.3389/fimmu.2019.02210
90. Liu T, Huang T, Li J, Li A, Li C, Huang X, et al. Optimization of differentiation and transcriptomic profile of THP-1 cells into macrophage by PMA. PloS One. (2023) 18:e0286056. doi: 10.1371/journal.pone.0286056
91. Lu C, Wu Z, Gao H, Li H, Deng R, Luo N, et al. Sperm induce macrophage extracellular trap formation via phagocytosis-dependent mechanism. Biol Reprod. (2023) 109:319–29. doi: 10.1093/biolre/ioad068
92. von Köckritz-Blickwede M, Chow O, Ghochani M, and Nizet V. Visualization and functional evaluation of phagocyte extracellular traps. San Diego (CA, USA): Methods in Microbiology. Immunol Infection. (2010) 37:139–60. doi: 10.1016/S0580-9517(10)37007-3
93. Matsubara K, Kubota M, Adachi S, Kuwakado K, Hirota H, Wakazono Y, et al. Different mode of cell death induced by calcium ionophore in human leukemia cell lines: possible role of constitutive endonuclease. Exp Cell Res. (1994) 210:19–25. doi: 10.1006/excr.1994.1003
94. Grob D, Conejeros I, Velásquez ZD, Preußer C, Gärtner U, Alarcón P, et al. Trypanosoma brucei brucei induces polymorphonuclear neutrophil activation and neutrophil extracellular traps release. Front Immunol. (2020) 11:559561. doi: 10.3389/fimmu.2020.559561
95. Abi Abdallah DS, Lin C, Ball CJ, King MR, Duhamel GE, and Denkers EY. Toxoplasma gondii triggers release of human and mouse neutrophil extracellular traps. Infect Immun. (2012) 80:768–77. doi: 10.1128/IAI.05730-11
96. Yildiz K, Gokpinar S, Gazyagci AN, Babur C, Sursal N, and Azkur AK. Role of NETs in the difference in host susceptibility to Toxoplasma gondii between sheep and cattle. Vet Immunol Immunopathol. (2017) 189:1–10. doi: 10.1016/j.vetimm.2017.05.005
97. Lacerda LC, dos Santos JL, Wardini AB, da Silva AN, Santos AG, Silva Freire HP, et al. Toxoplasma gondii induces extracellular traps release in cat neutrophils. Exp Parasitol. (2019) 207:107770. doi: 10.1016/j.exppara.2019.107770
98. Imlau M, Conejeros I, Muñoz-Caro T, Zhou E, Gärtner U, Ternes K, et al. Dolphin-derived NETosis results in rapid Toxoplasma gondii tachyzoite ensnarement and different phenotypes of NETs. Dev Comp Immunol. (2020) 103:103527. doi: 10.1016/j.dci.2019.103527
99. Wei Z, Wang Z, Liu X, Wang C, Han Z, Wu D, et al. Toxoplasma gondii triggers neutrophil extracellular traps release in dogs. Front Cell Infect Microbiol. (2020) 10:429. doi: 10.3389/fcimb.2020.00429
100. Wu T-T, Chen T-L, and Chen R-M. Lipopolysaccharide triggers macrophage activation of inflammatory cytokine expression, chemotaxis, phagocytosis, and oxidative ability via a Toll-like receptor 4-dependent pathway: validated by RNA interference. Toxicol Lett. (2009) 191:195–202. doi: 10.1016/j.toxlet.2009.08.025
101. Tosh KW, Mittereder L, Bonne-Annee S, Hieny S, Nutman TB, Singer SM, et al. The IL-12 response of primary human dendritic cells and monocytes to Toxoplasma gondii is stimulated by phagocytosis of live parasites rather than host cell invasion. J Immunol Baltim Md 1950. (2016) 196:345–56. doi: 10.4049/jimmunol.1501558
102. Lee E-J, Heo Y-M, Choi J-H, Song H-O, Ryu J-S, and Ahn M-H. Suppressed production of pro-inflammatory cytokines by LPS-activated macrophages after treatment with Toxoplasma gondii lysate. Korean J Parasitol. (2008) 46:145. doi: 10.3347/kjp.2008.46.3.145
103. Botting RA, Bertram KM, Baharlou H, Sandgren KJ, Fletcher J, Rhodes JW, et al. Phenotypic and functional consequences of different isolation protocols on skin mononuclear phagocytes. J Leukoc Biol. (2017) 101:1393–403. doi: 10.1189/jlb.4A1116-496R
104. He Y, Hara H, and Núñez G. Mechanism and regulation of NLRP3 inflammasome activation. Trends Biochem Sci. (2016) 41:1012–21. doi: 10.1016/j.tibs.2016.09.002
105. Oken AC, Lisi NE, Krishnamurthy I, McCarthy AE, Godsey MH, Glasfeld A, et al. High-affinity agonism at the P2X7 receptor is mediated by three residues outside the orthosteric pocket. Nat Commun. (2024) 15:6662. doi: 10.1038/s41467-024-50771-6
Keywords: Toxoplasma gondii, mononuclear phagocytes, extracellular traps, cytokines, early innate immunity
Citation: Hanke D, Velásquez ZD, Büttner K, Krueger A, Ross R, Hecker A, Mazurek S, Grau V, Taubert A, Hermosilla C, Richter K and Conejeros I (2025) Early responses of primary human and bovine monocytes, monocytic THP-1 cells and THP-1 cell-derived macrophages to vital Toxoplasma gondii tachyzoites. Front. Immunol. 16:1683634. doi: 10.3389/fimmu.2025.1683634
Received: 11 August 2025; Accepted: 26 September 2025;
Published: 30 October 2025.
Edited by:
Debanjan Mukhopadhyay, Presidency University, IndiaReviewed by:
Arne Lucas ten Hoeve, University Medical Center Freiburg, GermanyLouis-Philippe Leroux, Université du Québec, Canada
Copyright © 2025 Hanke, Velásquez, Büttner, Krueger, Ross, Hecker, Mazurek, Grau, Taubert, Hermosilla, Richter and Conejeros. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dominik Hanke, RG9taW5pay5IYW5rZUB2ZXRtZWQudW5pLWdpZXNzZW4uZGU=
†These authors have contributed equally to this work and share senior authorship