 Joana Vasconcelos1,2,3*†
Joana Vasconcelos1,2,3*† Ricardo Sousa3,4
Ricardo Sousa3,4 João Ferro5
João Ferro5 Ana Rita Pinto4Mafalda Freitas3,4
Ana Rita Pinto4Mafalda Freitas3,4 Rodrigo Riera2,6
Rodrigo Riera2,6- 1Faculdade de Ciências da Vida, Universidade da Madeira, Campus Universitário da Penteada, Funchal, Portugal
- 2Grupo en Biodiversidad y Conservación (BIOCON), Instituto Universitario de Investigación en Acuicultura Sostenible y Ecosistemas Marinos (IU-ECOAQUA), Universidad de Las Palmas de Gran Canaria, Marine Scientific and Technological Park, Telde, Spain
- 3MARE - Marine and Environmental Sciences Centre/Aquatic Research Network (ARNET), Agência Regional para o Desenvolvimento da Investigação Tecnologia e Inovação (ARDITI), Funchal, Portugal
- 4Direção de Serviços de Monitorização, Estudos e Investigação do Mar, Direção Regional do Mar, Funchal, Portugal
- 5Faculdade de Ciências Exatas e da Engenharia, Universidade da Madeira, Campus Universitário da Penteada, Funchal, Portugal
- 6Departamento de Ecología, Facultad de Ciencias, Universidad Católica de la Santísima Concepción, Concepción, Chile
Gonadal development in limpets is well-known. However, the integration of fecundity data on coastal management actions remains overlooked. Here, we use fecundity of two exploited limpets (Patella ordinaria and P. aspera) to demonstrate its potential as a conservation tool to preserve the harvested populations of both species. From October 2021 to June 2022, 158 females of P. ordinaria and 70 of P. aspera were collected from the coastal rocky shores of the archipelago of Madeira (NE Atlantic Ocean). The fecundity in both species appears to be determinate, and estimated batch fecundity 365,638 ± 204,462 oocytes for P. ordinaria and 73,029 ± 43,496 oocytes for P. aspera. The presence of spawning individuals of both species until May is of significant relevance to their management and conservation. A harvest-ban (November-March) is currently implemented in the coastal governance actions of P. ordinaria and P. aspera. Based on the fecundity results, small adjustments can be made to further improve the conservation of adult individuals, which are important for the continuity of future generations. So far, fecundity data constitute an overlooked life trait of key importance to preserve populations of exploited species, primarily those with limited mobility such as semi-sessile littoral limpets.
1 Introduction
The extensive human footprint has underpinned drastic changes to ecosystems all over the globe (Steffen et al., 2015; Venter et al., 2016). Marine, freshwater and terrestrial realms have been, and still are, subject to habitat loss and fragmentation (Dubois et al., 2018; Jones et al., 2018; Ellis et al., 2021). Conservation actions have been developed to halt this destructive trend and also to preserve scarcely-touched areas on our planet (Geldmann et al., 2013; Ward et al., 2022), such as the spatial management with the creation of no-take marine reserves. The main aim of a high portion of protected areas is to preserve certain endangered species, e.g. large mammals in Africa (Pacifici et al., 2020), or commercially valuable species, e.g. large-sized demersal fish (Hackradt et al., 2014). The establishment of these protected areas is mainly based on conventional biodiversity measures such as, abundance of individuals, densities per surface area, and also species richness in biodiversity hotspots (Blowes et al., 2020; Davies et al., 2021). Their assessment is typically based on management actions that follow up the previous parameters used to declare these spaces as protected areas (Dudley and Stolton, 2008). Other parameters remain overlooked such as, functional traits, phylogenetic diversity or those related to fecundity or reproductive potential (see exceptions Coleman et al., 2015; Guilhaumon et al., 2015). The integration of information on fecundity and reproductive potential would be an asset for conservation assessment studies. However, most of the studies are limited to the harvest control measures catch and bag limits, gear restrictions, closed seasons and minimum size limit (Worm et al., 2009; Gwinn et al., 2015; Liu et al., 2016). The latter has been used as a threshold to separate reproductively active adults and pre-adults or juveniles (e.g. Lavin et al., 2021), as it occurs in a wide range of commercial marine species, i.e. fish, crustaceans and molluscs (Ellis and Cowan, 2001; Alós et al., 2014; Sousa et al., 2019b). Fecundity studies may provide a more accurate picture of the reproductive cycle of a species throughout the year. There are management actions that target this temporal variability on the reproductive potential of the species such as, the implementation of seasons; yet, they are mostly based on maturity stages data classified according only to the progression of the gonads volume in the haemocoel, and only focused on the months where reproductive individuals are reported (de Mitcheson et al., 2020). An exhaustive analysis of fecundity data is expected to identify nuances and subtle variations of gonad development among individuals that may result in accurate management actions that ensure the continuity of the species under protection (Lambert, 2008).
Chronic fishing and harvesting activities have substantial effects on the reproductive parameters of the targeted species (Walsh et al., 2006; Wright and Trippel, 2009; Pellowe and Leslie, 2020). Reproductive dynamics are thought to suffer shifts due to long-term exploitation (Murphy et al., 1994; Sousa et al., 2019b; Pellowe and Leslie, 2020). In harvested molluscs, such as intertidal and shallow subtidal limpets, several reproductive parameters have been used as a proxy to establish the exploitation status of their populations (Riera et al., 2016; Sousa et al., 2019a). The impact of harvesting on littoral limpets from Madeira has been extensively studied in recent years (e.g., Sousa et al., 2019a; Sousa et al., 2019b; Sousa et al., 2020a; Sousa et al., 2020b). Reproductive parameters, including size and age at first maturity, have traditionally been used to assess the level of exploitation of P. ordinaria and P. aspera in this region. The populations of these two exploited species have been drastically reduced, with a more pronounced decrease in accessible sites (Sousa et al., 2019a). The persistent exploitation of this coastal resource has underpinned changes in life history traits, reproductive output, genetic diversity, among others (Henriques et al., 2017). These shifts are still occurring despite the implementation of management actions such as a closed season during the reproductive season (November-March) (Portaria n. 151/2022; Sousa et al., 2019a) and a minimum harvesting size of 40 mm shell length (Sousa et al., 2019b). This size limit management action was implemented to prevent the harvest of a portion of the spawning stock to maintain replenishment rates (as size at first maturity for P. ordinaria and P. aspera are 37.5 and 37.4 mm SL, respectively; Sousa et al., 2019b). This leaves a portion of the spawning stock vulnerable to fishing pressure, diminishing the reproductive output (Liu et al., 2016; Lavin et al., 2021) that increases hyperallometrically with size and age (Hixon et al., 2014; Barneche et al., 2018; Vasconcelos et al., 2023). This size-selective harvesting is worrisome, especially for the protandrous hermaphrodite P. aspera. In this species, smaller and younger limpets reach sexual maturation as males, with a fraction of them changing to females afterwards (Orton et al., 1956; Espinosa et al., 2006; Espinosa et al., 2009; Martins et al., 2017; Sousa et al., 2019b). This increase of females in larger size classes as the result of sex change, make them extremely vulnerable and a target to harvest (Espinosa et al., 2006), with overfishing affecting the more fecund females and thus reducing the reproductive output of populations (Espinosa et al., 2009). In this sense, we hypothesize that the potential fecundity of P. aspera is lower when compared with P. ordinaria, as a consequence of P. aspera being a protandrous hermaphrodite. Also, asynchronous spawning seasons may be expected between the northern and southern populations of both species. Preliminary results on the fecundity of one of these species were published recently (Vasconcelos et al., 2023). While studying P. ordinaria sampled along the Madeiran archipelago during the months of spawning, the authors found differences in the fecundity of northern and southern populations of this species, which may be indicative of this asynchrony.
We here used these two limpets as a model of study to explore the potential of fecundity data to be integrated into coastal management actions. The main goals of the present study are to: (i) analyze at a microscopic level, the gametogenic stages of P. ordinaria and P. aspera females, (ii) study the four main criteria applied for fecundity type determination for both patellid limpets, and (iii) compare the number and size of vitellogenic oocytes for the northern and southern populations of Madeira. The integration of the former parameters may result in management actions that would potentially be the cornerstone of the sustainable exploitation of these molluscs in the medium and long-term.
2 Material and methods
2.1 Data sampling
For this study, 158 specimens of Patella ordinaria (49.34 6.86 mm total shell length, SL) and 70 P. aspera (45.65 6.83 mm SL), two species that coexist in the lower intertidal to the subtidal from 0 to 6 m deep (Côrte-Real et al., 1996; Sousa et al., 2017; Sousa et al., 2020b), were randomly collected between October 2021 and June 2022 in four coastal settlements in the Madeira archipelago, two on the North coast (Porto Moniz and São Vicente) and two on the South coast (Funchal and Porto Santo) (Table 1; Figure 1). Due to the fact that these species are winter breeders (Henriques et al., 2012; Sousa et al., 2017), there were some limitations in the monthly (e.g., December) and site (as the North coast of Porto Santo Island) sampling of the reproductive stock, mainly due to the sea adverse conditions, particularly on the north coast of the archipelago (Castejón et al., 2022; Vasconcelos et al., 2023).
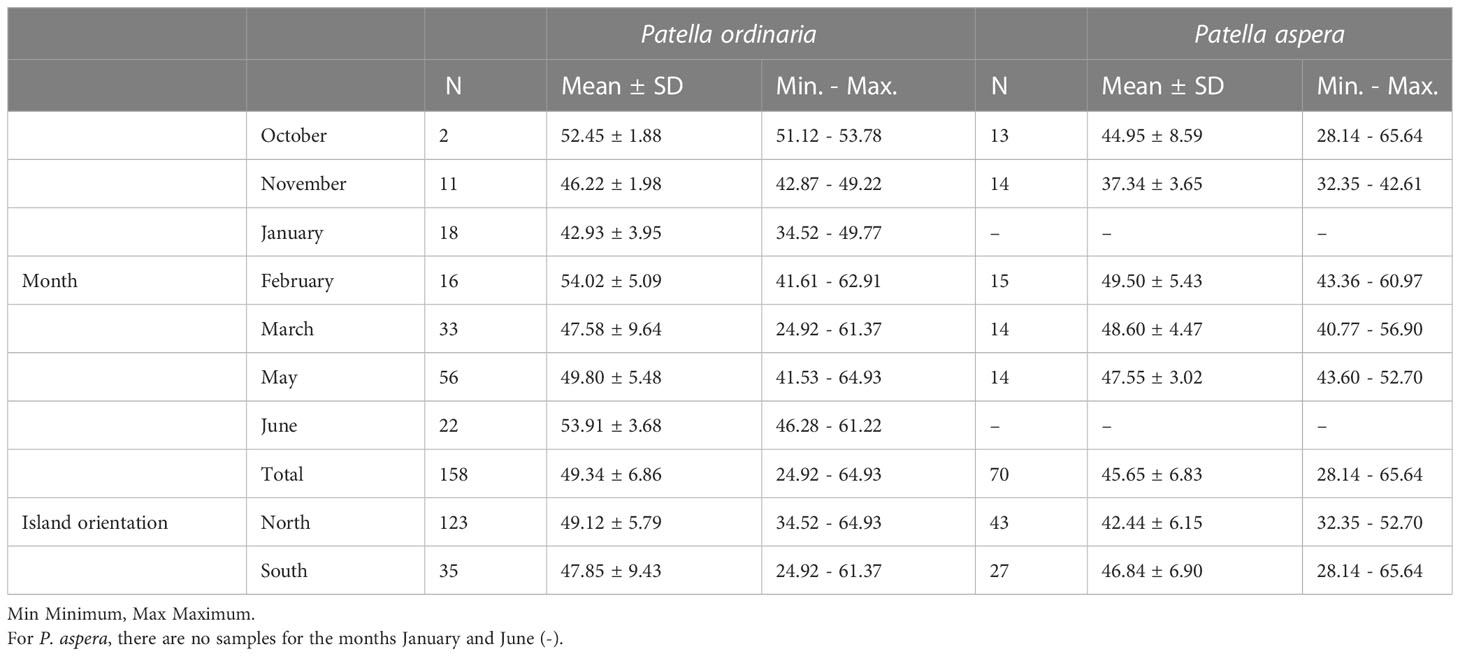
Table 1 Number (N) of individuals and the average total length (TL, mm) and respective standard deviation (SD) of Patella ordinaria and P. aspera from the archipelago of Madeira (NE Atlantic Ocean) sampled between October 2021 and June 2022.
Figure 1 Map showing the sampling areas (Funchal, Porto Moniz, Porto Santo and São Vicente) of Patella ordinaria and P. aspera collected from October 2021 to June 2022 in the Madeira archipelago (NE Atlantic Ocean).
In the laboratory, fresh individuals were sorted by species, measured for total shell length (SL, 0.01 mm accuracy) and weighted for total body wet weight (TW, 0.01 g accuracy). A small cut was made in the mantle at the posterior part of the foot (Cañizares et al., 2021) the gonads gently removed and weighted (GW, to the nearest 0.01 g). Sex was determined according to gonad pigmentation Orton et al. (1956), and female gonads preserved as a whole in Roti-Histofix ECO PLUS for histological analysis (Vasconcelos et al., 2023).
2.2 Histological analysis and sexual cycle
Histological sections were assembled from small portions of gonad dehydrated with ethanol at different concentrations (70, 90 and 95%) and embedded in Technovit 7100 resin (Vasconcelos et al., 2017). The homogeneity of oocyte distribution within ovaries was checked from histological sections (5 m) made on the anterior, middle, and posterior regions of the ovary. As the middle area was representative of the entire gonad, sections were made on this part of the ovary, stained with methylene blue and digitized using a visual image analysis system (Leica DM2700 P microscope coupled to the Leica DFC 450 C camera and the software Leica Application Suite X – LAS 3.7, Leica Microsystems). Classification of oogenesis stages and oocytes into previtellogenic (PO), vitellogenic (VO) and atresic oocytes (AO) (Supplementary Figure 1) were based on Prusina et al. (2014) (adopted and modified from McCarthy et al., 2008; Belkhodja et al., 2011): inactive; early development; late active; ripe; atresic; spawning; and spent.
The gonadosomatic index (GSI) was calculated according to the equation:
where GW is the gonad weight (in g) and TW the total body wet weight (in g).
2.3 Fecundity analysis
Fecundity of P. ordinaria and P. aspera, was assessed based on: (i) the presence or absence of an hiatus between PO and VO; (ii) the number of VO during the spawning period; (iii) average size of VO throughout the spawning period; and (iv) the incidence of atresia throughout the spawning period (Hunter, 1992; Greer Walker et al., 1994; Murua et al., 2003). For the first three criteria, 95 P. ordinaria (62 in the maturity stage ripe and 33 at the beginning of spawning) and 30 P. aspera (27 in ripe and 3 at the beginning of spawning) were analyzed. For the analysis of the first criterion, histological sections were used, while for the second and third criteria, the gravitometric method was applied (Hunter et al., 1989) following the protocol used previously for P. ordinaria (Vasconcelos et al., 2023). The number of oocytes per individual was registered and the diameter measured on 2,766 PO (1,815 P. ordinaria and 951 P. aspera) and 5,365 VO (3,614 P. ordinaria and 1,751 P. aspera) using the ImageJ software v1.53r (http://imagej.nih.gov/ij/) (Schneider et al., 2012). For the fourth criteria, the relative intensity of atresia, defined as the percentage of atretic vitellogenic oocytes in relation to total number of normal oocytes present in the ovary (Hunter and Macewicz, 1985), was estimated by examining histological sections of 104 gonads of P. ordinaria (16 in late active, 60 in ripe and 28 in spawning stage) and 48 of P. aspera (19 in late active, 26 in ripe and 3 in the spawning stage). Gonads in the atresic state of maturation were not considered as this could cause a miscalculation because the vast majority of oocytes are in atresia. The prevalence of atresia (defined as the proportion of females with atresia stage oocytes in the total number of females) was also investigated.
Batch fecundity measurements were based on the direct gravimetric method. From the histological examination, 46 females in the ripe stage (25 of P. ordinaria and 16 of P. aspera) were selected and the number of VO counted throughout the spawning season. Counts performed per specimen may be underestimated due to the loss of some oocytes during the gravimetric process. This problem was encountered by Guallart et al. (2020) and Vasconcelos et al. (2023).
2.4 Data analysis
Differences in the average number and size of oocytes across months were tested. As ANOVA assumptions were not met, the non-parametric Kruskal-Wallis rank sum test (stats R package, R Core Team, 2022) was used to investigate trends and differences in oocyte size and number across months. Kruskal-Wallis test does not require sample sizes to be the same in all groups. The effect size for Kruskal-Wallis test was computed as the eta squared based on the H-statistic (η 2, rstatix R package, Tomczak and Tomczak, 2014; Kassambara, 2022b). The eta squared estimate ranges from 0 to 1, where values closer to 1 indicate a higher proportion of variance in the dependent variable explained by the independent variable. Values from 0.01 to 0.06 represent a small effect, 0.06 to 0.14 moderate effect and 0.14 a large effect (Tomczak and Tomczak, 2014; Kassambara, 2022b). The post-hoc Pairwise Wilcoxon Test was applied to calculate pairwise comparisons across months with corrections for multiple testing (stats R package, R Core Team, 2022). The method selected for adjusting p values was the “BH” (Benjamini and Hochberg, 1995), a more powerful method than the family-wise error rate. The relative intensity of atresia throughout months was compared with the Kruskal-Wallis rank sum test (stats R package, R Core Team, 2022). The correlation between limpet size and the absolute batch fecundity was investigated using the Pearson correlation test (ggpubr R package, Kassambara, 2022a).
3 Results
3.1 Histological analysis and sexual cycle
The frequency distribution of different gametogenic stages observed throughout the study period is illustrated in Figure 2. Mature limpets were represented in all sampling months. Gametogenesis occurred prior to October, with 50% of the females in the Late active and 50% in Ripe stages in P. ordinaria whilst only 39% in the Early active and Late Active stage in P. aspera. Ripe gonads were more common from October to May in P. ordinaria and from February to March in P. aspera. During ripening, an average of 24% of female gonads of P. ordinaria were in the atresic stage, with most of oocytes undergoing atresia. In P. aspera, the atresic stage was observed in October, November and February with an average of 19%.

Figure 2 Monthly variation of different gametogenic stages in Patella ordinaria and P. aspera collected from October 2021 to June 2022 in the Madeira archipelago (NE Atlantic Ocean).
The gonadosomatic index increased from October-November to March for females of P. aspera and P. ordinaria respectively (Figure 3). Monthly values revealed an increase from November to March for both species (Kruskal-Wallis test (K-W); P. ordinaria: H = 107.98, p 0.001, = 0.687; P. aspera: H = 48.484, p 0.001, = 0.684; Figure 3), supporting the maturity frequency distributions based on the microscopic analysis of females gonads (Figure 2). The maximum GSI values were found in March for both species, where females of P. ordinaria attained 21.9% (10.9 6.32, mean ± SD) and the females of P. aspera 9.12% (5.14 2.65).
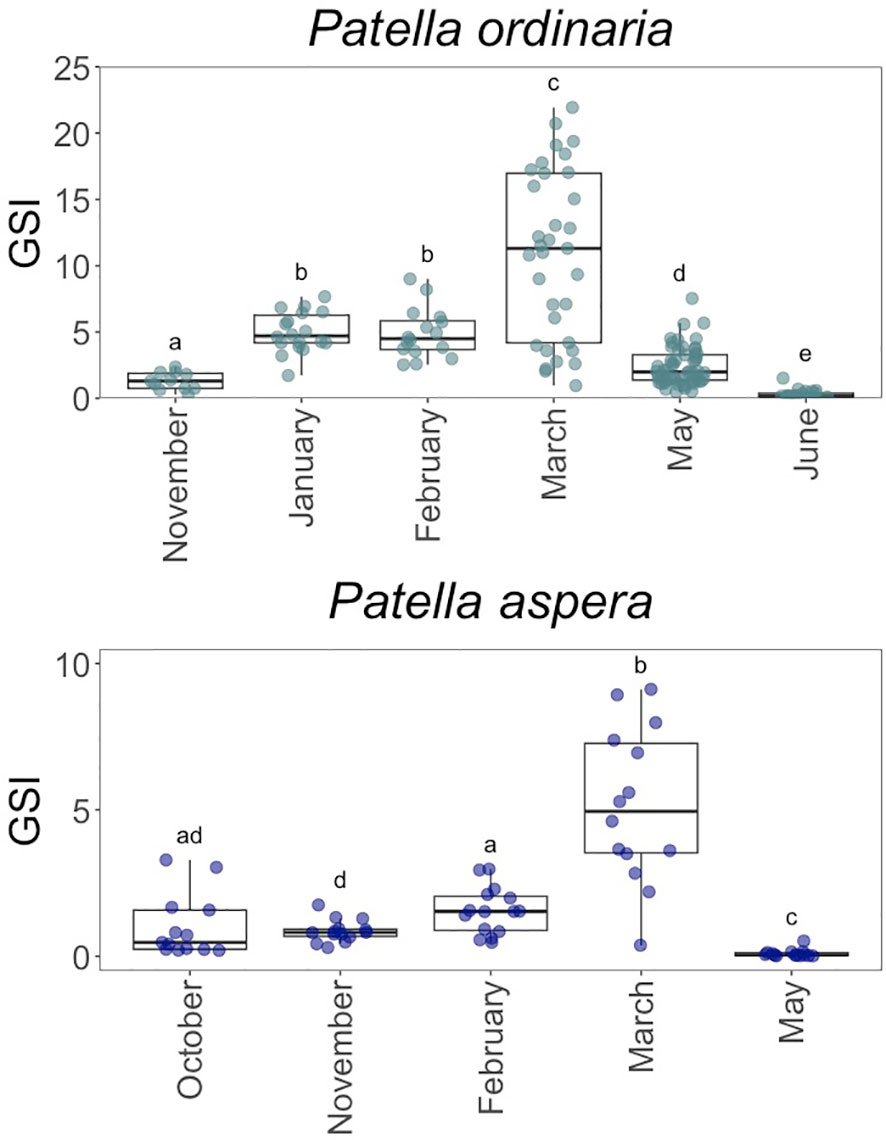
Figure 3 Gonadosomatic index (GSI) for Patella ordinaria and P. aspera sampled from October 2021 to June 2022 in the archipelago of Madeira (NE Atlantic Ocean). Boxplot showing the minimum and maximum values in the data, the 25th percentile, the median and the 75th percentile. Different letters indicate statistically significant differences between months (p < 0.05).
3.2 Fecundity estimates
The homogeneity of variance and the normality of the distribution of the number and size of PO and VO were tested for P. ordinaria and P. aspera. Regarding the number (Levene’s test: W = 1.1629, p > 0.05) and size (W = 1.7854, p 0.05) of PO in P. ordinaria, we can assume the homogeneity of variances across months. Number (W = 8.2877, p 0.001) and size (W = 6.5546, p 0.001) of VO across months did not exhibit homogeneous variance. For P. aspera, the number (Levene’s test: W =7.1013, p 0.001) and size (W = 3.1174, p 0.05) of PO and the number of VO (W = 3.6633, p 0.05) did not exhibit homogeneous variance across months. The size of VO (W = 2.1727, p = 0.05466) exhibits homogeneity of the variances across months. The size and number of PO and VO departed significantly from normality (Shapiro-Wilk test: p 0.001) in both species.
Fecundity was analyzed under the four above-mentioned criteria. The analysis of the oocytes size frequency distribution showed a distinct interruption between PO and VO in most of the sampled months for both Patella species (Figure 4). A progressing dominant cohort in the oocyte size frequency distribution can be observed during the sampling period. The average size of PO varied between 45 18 m (mean SD; 10 - 98 m) for P. ordinaria and 47 17 m (15 - 100) for P. aspera. The size of VO ranged from 80 – 248 m (149 m 23) for P. ordinaria and from 83 – 216 m (136 m 30) for P. aspera (Figure 4). For P. ordinaria, the average number of PO decreased significantly from November till March (Kruskal-Wallis test (K-W): H = 28.132, p 0.05, = 0.228; Figure 5A). By contrast, for the same period there was an increase in the number of VO (K-W: H = 40.799, p 0.05, = 0.359; Figure 5B). For P. aspera, the Kruskal-Wallis test showed significant differences in the number of PO (H = 26.802, p 0.05, = 0.519; Figure 5E) and VO (H = 30.138, p 0.05, = 0.599; Figure 5F). Yet, there was no trend in the number of PO (Figure 5E) such as the increase seen in VO from October to March (Figure 5F). The average size of PO of both P. ordinaria (H = 8.2708, p 0.05, = 0.00862; Figure 5C) and P. aspera (H = 8.9925, p 0.05, = 0.00576; Figure 5G) did not vary significantly across months. The size of VO of P. ordinaria decreased significantly from November till June (H = 141.89, p 0.05, = 0.0320; Figure 5D). During the same period, the average size of VO of P. aspera increased significantly (H = 189.3, p 0.05, = 0.112; Figure 5H).
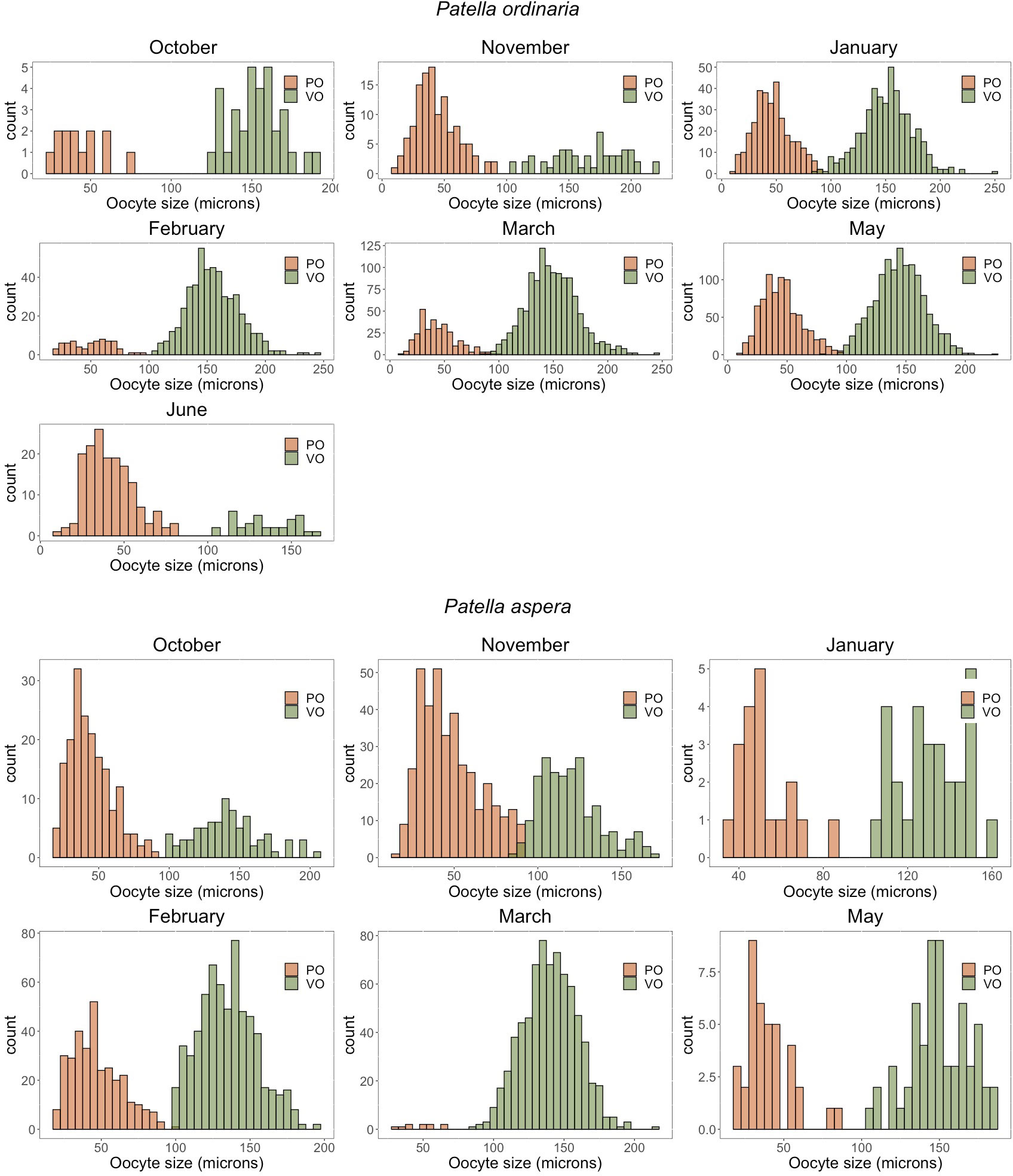
Figure 4 Oocyte size frequency distribution in the limpets Patella ordinaria and P. aspera sampled from October 2021 to June 2022 in the archipelago of Madeira (NE Atlantic Ocean). PO Previtellogenic oocytes, VO vitellogenic oocytes.
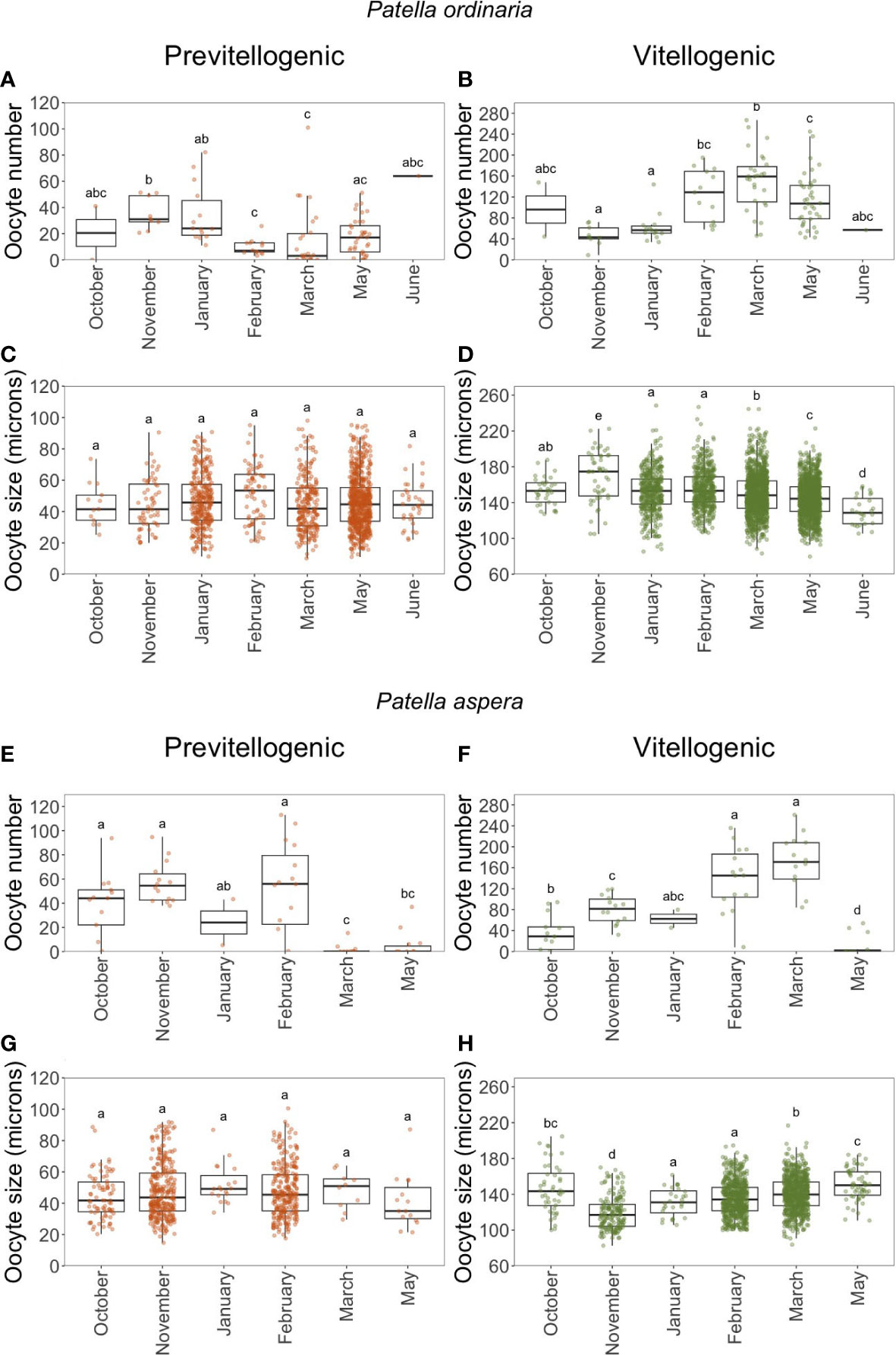
Figure 5 Monthly variation in the number (A, B, E, F) and size (C, D, G, H) of previtellogenic and vitellogenic oocytes of Patella ordinaria and P. aspera sampled in the archipelago of Madeira (NE Atlantic Ocean). Boxplot shows the minimum and maximum values in the data, the 25th percentile, the median and the 75th percentile. Different letters indicate statistically significant differences between months (p < 0.05).
The number and size of PO and VO was analyzed per maturity stage (Figure 6). For the VO, an increase in their number from Late active to Ripe stage, followed by a decrease during spawning stage was observed for both P. ordinaria (H = 30.774, p 0.001, = 0.285; Figure 6B) and P. aspera (H= 20.94, p 0.001, = 0.421; Figure 6F). Regarding their size, there was an increased from Late active to the Spawning stage in both P. ordinaria (H = 338.23, p 0.001, = 0.102; Figure 6D) and P. aspera (H = 172.83, p 0.001, = 0.103; Figure 6H). PO attained their lowest number in the Ripe stage (P. ordinaria: H = 26.406, p 0.001, = 0.242, Figure 6A; P. aspera: H = 17.625, p 0.001, = 0.347; Figure 6E), associated with an increase in their size from Late active to the Ripe stage (P. ordinaria: H = 14.86, p 0.001, = 0.00965; Figure 6C; P. aspera: H = 6.4793, p 0.05, = 0.00786; Figure 6G) followed by a decrease in spawning stage in P. ordinaria.
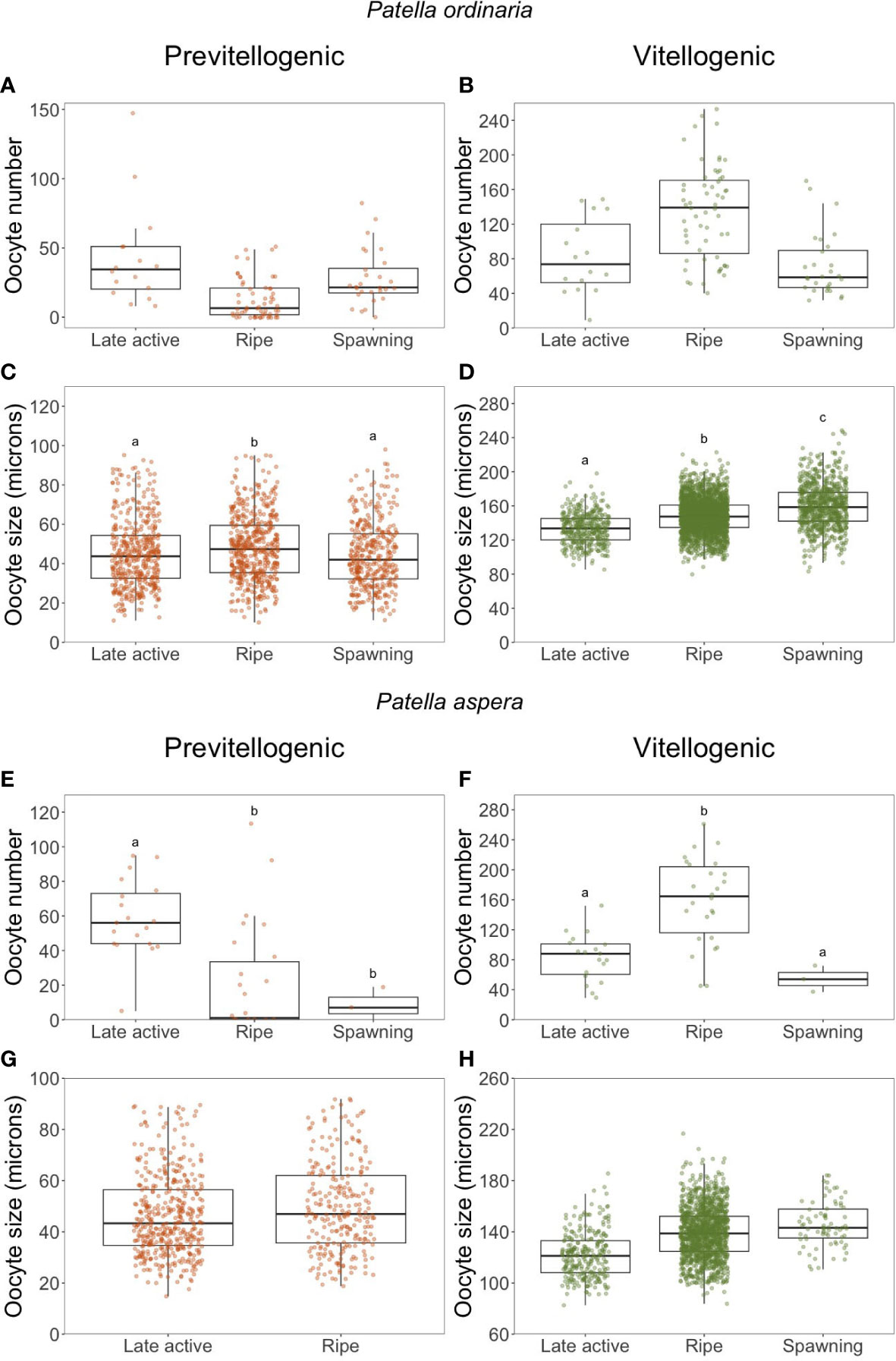
Figure 6 Number (A, B, E, F) and size (C, D, G, H) of previtellogenic and vitellogenic oocytes per maturity stages of Patella ordinaria and P. aspera sampled in the archipelago of Madeira (NE Atlantic Ocean). Boxplot showing the minimum and maximum values in the data, the 25th percentile, the median and the 75th percentile. Different letters indicate statistically significant differences between maturity stages (p < 0.05).
As no clear trends were observed in the number and size of PO and VO of both species (Figure 5), samples were spatially separated into northern and southern populations. For the northern populations of both species, the average number of PO decreased from November to May (P. ordinaria: H = 14.143, p 0.05, = 0.192; Figure 7A; P. aspera: H = 8.5234, p 0.05, = 0.502; Figure 7E). However, no trends in their size were observed (Figures 7C, G). For P. ordinaria, the average number of VO increase significantly from November to May (H = 25.093, p 0.05, = 0.381; Figure 7B) associated with a decrease in their size (H = 87.476, p 0.05, = 0.0506; Figure 7D). For P. aspera, the opposite was observed. The number of VO decreased significantly (H = 6.9579, p 0.05, = 0.381; Figure 7F), with the increase in their size (H = 86.021, p 0.05, = 0.322; Figure 7H). The southern populations of P. ordinaria and P. aspera were analyzed for number and size of VO (Supplementary Figure 2). From the analysis of the only available months of sampling in the southern part of the archipelago (October and March), there is an increasing trend in the number of VO, although significant only for P. aspera (P.ordinaria: H = 1.8968, p 0.05, = 0.0332; P. aspera: H = 10.531, p 0.05, = 0.561). No trend was observed in the average size of VO (P. ordinaria: H = 1.0521, p 0.05, = 0.0000461; P. aspera: H = 1.4769, p 0.05, = 0.000619).
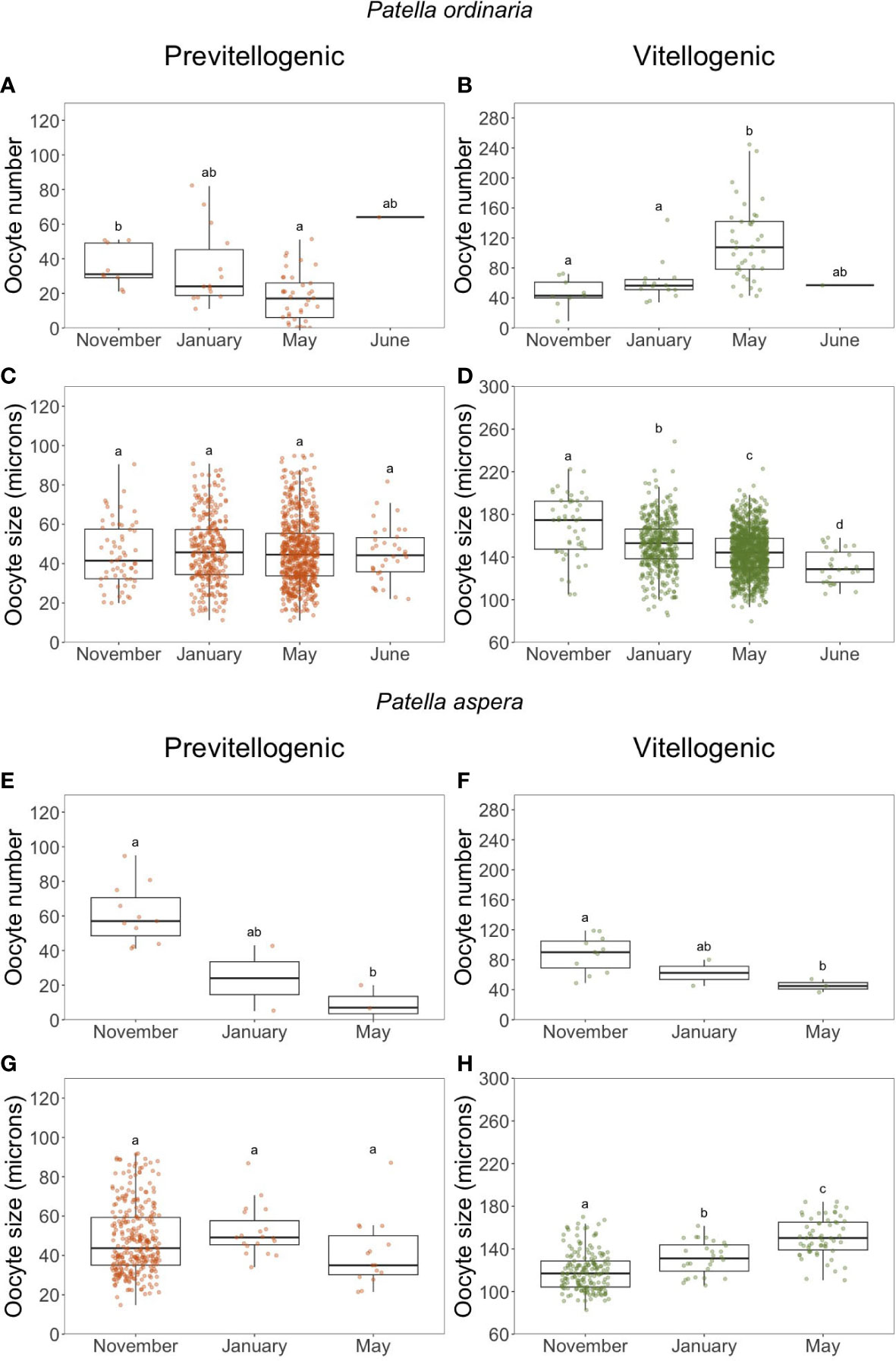
Figure 7 Monthly variation in the number (A, B, E, F) and size (C, D, G, H) of previtellogenic and vitellogenic oocytes of Patella ordinaria and P. aspera sampled in the north coast of the archipelago of Madeira (NE Atlantic Ocean). Boxplot shows the minimum and maximum values in the data, the 25th percentile, the median and the 75th percentile. Different letters indicate statistically significant differences between months (p < 0.05).
There was an increase from 9.5 2.12% (mean SD) in November to 24.4 15.4% in May in the relative intensity of the atresia for P. ordinaria, although at the limit of significance (H = 12.154, p 0.05, = 0.0746; Figure 8). For P. aspera, no significant differences were found in the relative intensity of atresia across months (H = 8.6772, p 0.05, = 0.0876; Figure 8), although atresia varied between 10.3 1.53% in May to 44 4.24% in January (Figure 8). The prevalence of atresia, i.e. the proportion of females with oocytes in atresia, was high for both species (P. ordinaria = 97%; P. aspera = 98%).
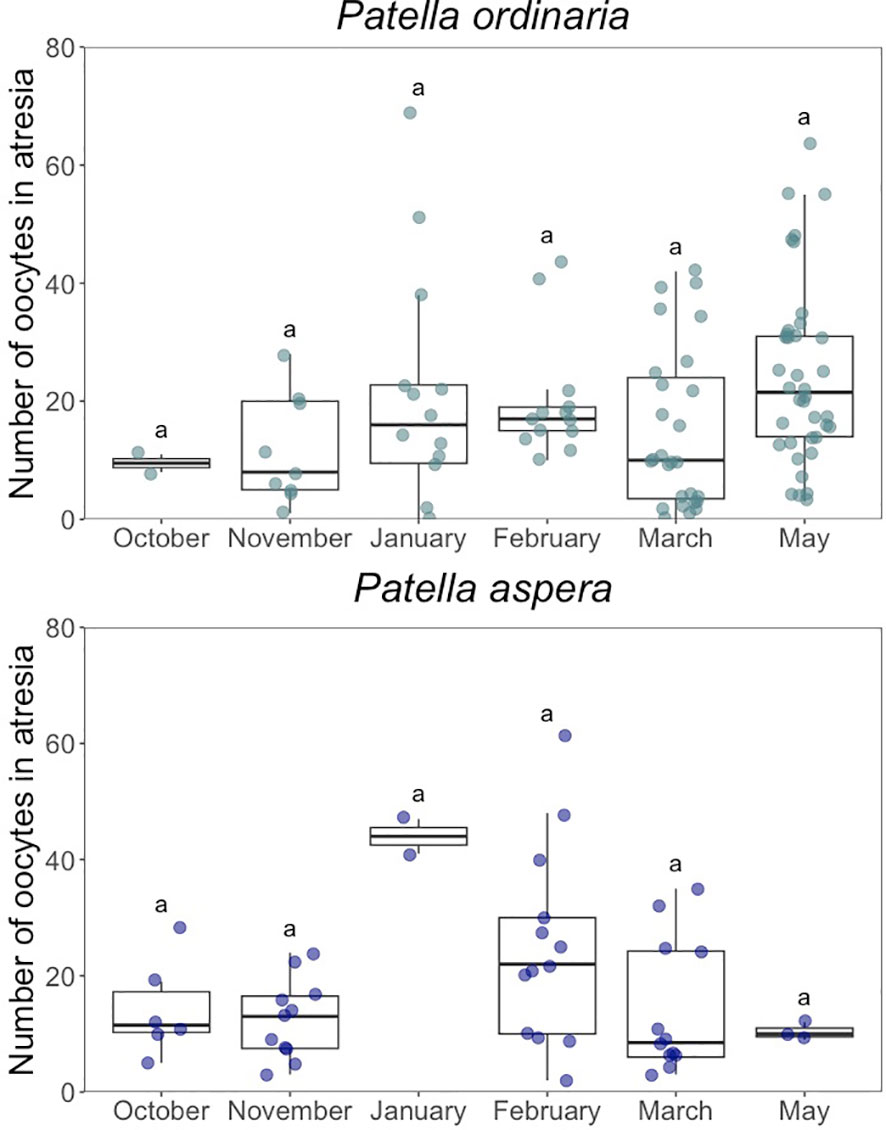
Figure 8 Relative intensity of the atresia estimated for Patella ordinaria and P. aspera sampled in the archipelago of Madeira (NE Atlantic Ocean) across the spawning season. Boxplot showing the minimum and maximum values in the data, the 25th percentile, the median and the 75th percentile. Different letters indicate statistically significant differences between months (p < 0.05).
Lastly, batch fecundity, estimated based on females in the ripe stage, ranged between 80,006 and 774,504 oocytes for females of P. ordinaria with 45.72 mm SL and 61.37 mm SL, respectively (354,572 189,883, mean SD). For P. aspera, batch fecundity was on average 77,404 43,910 oocytes, ranging from 22,371 (44.71 mm SL) to 177,202 oocytes (53.38 mm SL). A significant positive correlation was found between the number of oocytes and the shell size of P. ordinaria (Pearson’s correlation: R = 0.884, p 0.001; Supplementary Figure 3). This relationship was not that clear in P. aspera (R = 0.272, p 0.05; Supplementary Figure 3).
4 Discussion
Monitoring species’ life histories, particularly their reproductive strategy, is crucial for their effective and long-term management. However, much of the available information is based on the macroscopic identification of the maturity stages, which can lead to errors (Neves et al., 2022). Until recently, all works published on the reproductive biology of the limpets P. ordinaria and P. aspera to determine their exploitation status in Madeira archipelago were based on the macroscopic analysis of gonads (Henriques et al., 2012; Sousa et al., 2017; Sousa et al., 2019b; Sousa et al., 2020a). In this analysis, maturity stages are classified according only to the progression of gonad volume in the haemocoel, going from either the rudimentary virgin stage or resting after discharge of gametes (inactive or neuter) to fully developed occupying the entire haemocoel (stage V) (Orton et al., 1956). Only two recent works refer to preliminary data on the fecundity of P. aspera (assays on females fecundity to develop methodologies for the culture of both limpet species, Castejón et al., 2022) and P. ordinaria (Vasconcelos et al., 2023). In this regard, the present work fills the existing gaps by characterizing the fecundity of both limpet species based on microscopic analysis of female gonads to support its management in Madeira archipelago. Reproduction of both patellid limpets has been considered to occur during winter, from November to April (Henriques et al., 2012; Sousa et al., 2017), with spawning most likely to occur from January till April (Pinto et al., 2010; Henriques et al., 2012; Sousa et al., 2017) and with maximum GSI values reported in January and February for P. aspera and P. ordinaria, respectively (Pinto et al., 2010; Henriques et al., 2012; Sousa et al., 2017). When using the microscopic analysis of females’ gonad in the present work, adults in the maturity stages of ripe and spawning were observed until May, despite the decrease in the GSI values after the peak in March. This proves the importance of the microscopic analysis in the determination of the spawning and resting periods.
All lines of evidence to assess fecundity were analyzed for the limpets P. ordinaria and P. aspera. During the spawning season, a hiatus and a trend can be observed in the oocyte size frequency, with two recognizable cohorts of oocytes, a smaller one of PO that progresses across the sampling period to a larger one that represents the VO. This is indicative of a disruption in the oocyte recruitment with the fecundity well set before the onset of spawning (Hunter, 1992). This bimodal distribution has been previously observed for P. ordinaria from Madeira archipelago (Vasconcelos et al., 2023), S. granularis from the SE coast of South Africa (Vat, 2000) and P. rustica from the SE Adriatic (Prusina et al., 2014). When analyzing the number of oocytes across months and maturity stages, the average number of PO decreased while the VO increased for both limpet species. Also, an expected decrease in the number of VO at the end of the spawning season, more precisely in May, was detected for both species, corroborating the presence of the hiatus between PO and VO and that no new VO are recruited to replace those that have been shed during spawning. In a previous work on the fecundity of P. ordinaria from the same region (Vasconcelos et al., 2023), this predicted decrease could not be observed, most likely due to the sampling period that lasted only till March. And the mean diameter of VO increased significantly over the spawning period, though more evident for P. aspera, corroborating the presence of a dominant size class of oocytes that progresses across months. Given the increase in the average size of VO and the decrease in their number, we can suggest that this is the case of determined fecundity. For indeterminate fecundity, the size of VO might remain constant or decrease (Murua et al., 2003). The evidences indicates that spawning, though to a lesser extent, may occur until May in both species, two months later than the end of the harvest-ban season (November-March). For P. ordinaria (9.5% - 24.4%) and P. aspera (10.3% - 44%), the low levels of the relative intensity of atresia detected across months, may be also indicative of determinate spawners (Hunter, 1992). The higher value found in January for P. aspera (44%) is a consequence of the lower number of individuals analyzed. Low levels of atresia were reported previously for P. ordinaria of Madeira archipelago (7.9% - 20.8% Vasconcelos et al., 2023).
Fecundity parameters also showed some variability between the northern and southern populations for P. ordinaria and P. aspera. Unfortunately, due to the absence of a greater number of samples throughout the spawning season, the expected trends regarding the number and size of VO were not as evident as when analyzed along the stages of maturation (Figure 6). For the northern populations of P. ordinaria, there was an increase in the number of VO till May, followed by a decrease. For P. aspera an increase in the size of VO was observed from November to May. The fact that the samples are too separated in time, i.e., without specimens between January and May, it is hardly difficult to infer whether there is an increase or decrease in the average size of VO of P. ordinaria and in the number of VO of P. aspera. However, for P. ordinaria, a large number of vitellogenic oocytes and an average GSI of 2.40% was still detected in May, giving the suspicion that this population extends its reproductive cycle beyond the currently established closed season. The harsher hydrodynamic and higher speed wind, typical conditions of the north coast of Madeira, induce spawning (Orton et al., 1956). It is also very likely the extension of the breeding season, when compared to the populations inhabiting the south coasts of the archipelago. Though the environmental conditions are the same for P. aspera, after the closed season the largest individuals are the target of exploitation leading to a reduction of females, as a fraction of males changes to females after reaching sexual maturation. Hence, the removal of larger animals will largely target females (Martins et al., 2017) that may still be spawning, greatly affecting the reproductive potential of this species. Regarding southern populations (Supplementary Figure 2), as only the months of October and March are available, it is difficult to infer trends throughout the spawning season. However, for both limpets, there was an expected decrease in the number of PO and a consequent increase in the number of VO, though with no significant increase in their average size.
The estimated fecundity (354,572 ± 189,883) was similar to that previously obtained for P. ordinaria in the same area (average of 385,613 194,902 oocytes; 46 - 59 mm SL Vasconcelos et al., 2023) but much higher than the one estimated in an assay for gametes and larval production for P. ordinaria aquaculture which averaged 186,000 oocytes (37 - 57 mm SL, Castejón et al., 2022). The fecundity in P. aspera females was on average 77,404 ± 43,910 oocytes, very similar to the one estimated in the same assay for P. aspera aquaculture which averaged 59,000 oocytes (34 - 53 mm SL, Castejón et al., 2022). Although the size range is similar for both species, the lower average size of P. aspera explains, in part, the clear differences in batch fecundity with P. ordinaria, as fecundity increases hyperallometrically with size. Moreover, the size-selective harvesting targets mainly females of the protandric hermaphrodite species P. aspera. These differences are of particular concern and should be taken in consideration when creating possible new management measures for limpet harvesting. P. ferruginea, currently considered at risk of extinction (Guallart et al., 2020), shows greater fecundity for shell sizes smaller than those sampled in the present work. For instance, while female fecundity of P. ferruginea with 40.0 mm SL is around 189,200 oocytes (Chafarinas Islands; Guallart et al., 2020), P. ordinaria from Madeira presents around 80,006 (present study) to 92,098 oocytes (Vasconcelos et al., 2023) in females with 46 mm SL. For P. ordinaria, Castejon et al. (2022) estimated an even lower fecundity for specimens with 37 mm SL, which was around 12,000 oocytes. When considering the maximum sizes analyzed, P. vulgata from the SW of England (500,000 eggs with 52 mm SL; Ballantine, 1961), and Cellana ornata from southern New Zealand (200,000 to 360,000 eggs for females > 40 mm SL; Dunmore and Schiel, 2000), showed fecundities lower than those estimated for P. ordinaria, but still higher than the ones verified for P. aspera (177,202 oocytes for 53.38 mm SL). For females of P. ferruginea with sizes higher than 80 mm SL, the number of oocytes ranged from 2.3 to 5.0 million eggs (Espinosa et al., 2006; Guallart et al., 2020). Since individuals of P. ordinaria and P. aspera larger than 60 mm SL are more common in the marine protected areas (MPAs) of the Madeira archipelago (with maximum shell lengths of 79.63 mm SL for P. ordinaria and 84.22 mm SL for P. aspera; Sousa et al., 2020a), and that all specimens used were captured in areas of considerable anthropogenic exploitation (capture only ceases during the closed season between November and March), we could not access or find evidence that these species can attain a level of fecundity as high as that reported for P. ferruginea. Moreover, as egg production is sensitive to selective pressures (Ramirez Llodra, 2002), this may explain the lower levels observed especially for P. aspera, the most popular limpet species consumed by the Madeiran population. Like the taxa P. ferruginea (Espinosa et al., 2006; Guallart et al., 2020), Cellana ornata (Dunmore and Schiel, 2000) and C. sandwicensis (Mau et al., 2018), P. ordinaria from Madeira exhibits size-dependent fecundity. Whilst this relationship was not that clear for P. aspera (present study and Castejón et al., 2022), limpets at MPAs may exhibit higher batch fecundity.
Fecundity is one of the major cornerstones of population biology (Bradshaw and McMahon, 2008). Temporal variations in fecundity are influenced by a wide range of factors such as, age, body size relationships, population density, mate choice, and environmental variability. Reproductive and fecundity data have traditionally been overlooked in fisheries management regardless of their importance to leverage the reproductive potential of large adult fish (Marshall et al., 2021). The integration of size-fecundity relationships into stock assessment models have been shown to be of utmost importance to achieve sustainable management goals, and their potential to establish spatio-temporal closures (STCs) and harvest slots (HSs) (Marshall et al., 2021). These limpets have allometric growth (Henriques et al., 2012; Sousa et al., 2017); hence, identifying and protecting size-age groups with maximum reproductive potential has been previously shown as a pivotal step to halt overfishing through managing growth of individuals and recruitment of stocks (Brown-Peterson et al., 2011). In the present study, the four criteria analyzed suggest that both exploited limpets may have determinate fecundity. It also showed the presence of spawning adults beyond the closed season (November-March), during the months of April and May. Furthermore, a spatial variability was observed between northern and southern populations, where the former ones showed vitellogenic oocytes till May in P. ordinaria whilst P. aspera showed a size increase of the vitellogenic oocytes from November to May. This means that some adults may be still spawning in May for both species, and hence to get an effective conservation of adult specimens and to ensure offspring of next generations, a more prolonged closed season till May is urgently needed. As evidenced by this study, fecundity data constitute a tool of key importance to preserve populations of exploited species, primarily those with limited mobility such as semi-sessile limpets.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
JV: Conceptualization, data acquisition, data curation, methodology, formal analysis, validation, critical analysis, resources, project administration, supervision, writing—original draft preparation, and writing—review and editing. RS: biological sampling, methodology, validation, critical analysis and revision of the paper. JF: data acquisition, data curation and interpretation. AP: biological sampling, validation. MF: resources, revision of the paper. RR: data interpretation, critical analysis, project administration, supervision, writing—original draft preparation, writing—review and editing. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the Portuguese national funds through Fundação para a Ciência e a Tecnologia I.P. (FCT) within the project “Metacommunity dynamics of complex life cycle species in exploited ecosystems” (LAPACOM) grant number EXPL/MAT-APL/0262/2021, MARE grant number UIDB/04292/2020, Associate Laboratory ARNET grant number LA/P/0069/2020, Interreg MAC within the project RASPA grant number MAC2/1.1a/305, the National Data Collection Framework, and the University of Las Palmas of Grancanaria (ULPGC). JV was supported by the ‘Grants for the Viera y Clavijo training program for researchers’ of the Government of the Canary Islands (VIERA Y CLAVIJO-2022-CIENCIAS-1).
Acknowledgments
We would like to express our gratitude to Dr. Manfred Kaufmann (University of Madeira) and Dr. Pedro Neves (OOM/ARDITI) and Dr. Cláudia Ribeiro (IFCN) for kindly facilitating their image analysis systems. We extend our gratitude to DRM, for providing all the facilities and material needed for the development of this work, and to the students José Mateus and Ricardo Mota, for having participated in the various stages of fecundity determination during their 1-month “Summer internships for University Students” funded by the Regional Youth Directorate of the Government of Madeira.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1173629/full#supplementary-material
References
Alós J., Palmer M., Catalan I. A., Alonso-Fernández A., Basterretxea G., Jordi A., et al. (2014). Selective exploitation of spatially structured coastal fish populations by recreational anglers may lead to evolutionary downsizing of adults. Mar. Ecol. Prog. Ser. 503, 219–233. doi: 10.3354/meps10745
Ballantine W. J. (1961). The population dynamics of patella vulgata and other limpets (Queen Mary University of London).
Barneche D. R., Robertson D. R., White C. R., Marshall D. J. (2018). Fish reproductive-energy output increases disproportionately with body size. Science 360, 642–644. doi: 10.1126/science.aao6868
Belkhodja H., Jaafoura M. H., Missaoui H., Romdhane M. S. (2011). Histological investigation of the reproductive cycle of the limpet Patella caerulea linnaeu. Cahiers biol. Mar. 52, 279–290. doi: 10.21411/CBM.A.C8C14E76
Benjamini Y., Hochberg Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc.: Ser. B (Methodological) 57, 289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x
Blowes S. A., Chase J. M., Di Franco A., Frid O., Gotelli N. J., Guidetti P., et al. (2020). Mediterranean Marine protected areas have higher biodiversity via increased evenness, not abundance. J. Appl. Ecol. 57, 578–589. doi: 10.1111/1365-2664.13549
Brown-Peterson N. J., Wyanski D. M., Saborido-Rey F., Macewicz B. J., Lowerre-Barbieri S. K. (2011). A standardized terminology for describing reproductive development in fishes. Mar. Coast. Fish. 3, 52–70. doi: 10.1080/19425120.2011.555724
Cañizares J. M., Castejón D., Haroun R., Nogueira N., Andrade C. A. (2021). Enhancing oocyte maturation and fertilisation in the black-foot limpet Patella candei d′Orbigny 1840 (patellidae, mollusca). Aquacul. Rep. 21, 100856. doi: 10.1016/j.aqrep.2021.100856
Castejón D., García L., Cañizares J. M., De Girolamo M., Nunes C., Isidro E., et al. (2022). Methodologies for patellid limpets’ aquaculture: from broodstock management to juveniles. Front. Mar. Sci. 9, 884262. doi: 10.3389/fmars.2022.884262
Coleman M., Bates A., Stuart-Smith R., Malcolm H., Harasti D., Jordan A., et al. (2015). Functional traits reveal early responses in marine reserves following protection from fishing. Diversity Distrib. 21, 876–887. doi: 10.1111/ddi.12309
Côrte-Real H., Hawkins S., Thorpe J. (1996). An interpretation of the taxonomic relationship between the limpets Patella rustica and P. piperata. J. Mar. Biol. Assoc. United Kingdom 76, 717–732. doi: 10.1017/S0025315400031416
Davies B. F., Holmes L., Rees A., Attrill M. J., Cartwright A. Y., Sheehan E. V. (2021). Ecosystem approach to fisheries management works–how switching from mobile to static fishing gear improves populations of fished and non-fished species inside a marine-protected area. J. Appl. Ecol. 58, 2463–2478. doi: 10.1111/1365-2664.13986
de Mitcheson Y. J. S., Linardich C., Barreiros J. P., Ralph G. M., Aguilar-Perera A., Afonso P., et al. (2020). Valuable but vulnerable: over-fishing and under-management continue to threaten groupers so what now? Mar. Policy 116, 103909. doi: 10.1016/j.marpol.2020.103909
Dubois N., Saulnier-Talbot É., Mills K., Gell P., Battarbee R., Bennion H., et al. (2018). First human impacts and responses of aquatic systems: a review of palaeolimnological records from around the world. Anthropocene Rev. 5, 28–68. doi: 10.1177/2053019617740365
Dudley N., Stolton S. (2008). Defining protected areas: an international conference in almeria, spain (Gland: IUCN).
Dunmore R., Schiel D. (2000). Reproduction in the intertidal limpet Cellana ornata in southern new zealand. New Z. J. Mar. Freshw. Res. 34, 653–660. doi: 10.1080/00288330.2000.9516966
Ellis S. L., Cowan D. F. (2001). Volunteer-based monitoring of juvenile american lobster, Homarus americanus. Mar. Freshw. Res. 52, 1103–1112. doi: 10.1071/MF01194
Ellis E. C., Gauthier N., Klein Goldewijk K., Bliege Bird R., Boivin N., Díaz S., et al. (2021). People have shaped most of terrestrial nature for at least 12,000 years. Proc. Natl. Acad. Sci. 118, e2023483118. doi: 10.1073/pnas.2023483118
Espinosa F., Guerra-Garcia J. M., FA D., Garcia-Gomez J. C. (2006). Aspects of reproduction and their implications for the conservation of the endangered limpet, Patella ferruginea. Invertebrate Reprod. Dev. 49, 85–92. doi: 10.1080/07924259.2006.9652197
Espinosa F., Rivera-Ingraham G., García-Gómez J. C. (2009). Gonochorism or protandrous hermaphroditism? evidence of sex change in the endangered limpet Patella ferruginea. Mar. Biodiversity Records 2, e153. doi: 10.1017/S1755267209990790
Geldmann J., Barnes M., Coad L., Craigie I. D., Hockings M., Burgess N. D. (2013). Effectiveness of terrestrial protected areas in reducing habitat loss and population declines. Biol. Conserv. 161, 230–238. doi: 10.1016/j.biocon.2013.02.018
Greer Walker M., Witthames P., Bautista De Los Santos I. (1994). Is the fecundity of the atlantic mackerel (Scomber scombrus: scombridae) determinate? Sarsia 79, 13–26. doi: 10.1080/00364827.1994.10413543
Guallart J., Peña J. B., Pérez-Larruscaín J., Luque Á.A., Templado J. (2020). Filling gaps: closing the life cycle of the endangered mediterranean limpet patella ferruginea gmelin 1791 (gastropoda, patellidae). Mediterr. Mar. Sci. 21, 400–419. doi: 10.12681/mms.22508
Guilhaumon F., Albouy C., Claudet J., Velez L., Ben Rais Lasram F., Tomasini J.-A., et al. (2015). Representing taxonomic, phylogenetic and functional diversity: new challenges for mediterranean marine-protected areas. Diversity Distrib. 21, 175–187. doi: 10.1111/ddi.12280
Gwinn D. C., Allen M. S., Johnston F. D., Brown P., Todd C. R., Arlinghaus R. (2015). Rethinking length-based fisheries regulations: the value of protecting old and large fish with harvest slots. Fish Fish. 16, 259–281. doi: 10.1111/faf.12053
Hackradt C. W., García-Charton J. A., Harmelin-Vivien M., Perez-Ruzafa A., Le Direach L., Bayle-Sempere J., et al. (2014). Response of rocky reef top predators (serranidae: epinephelinae) in and around marine protected areas in the western mediterranean sea. PloS One 9, e98206. doi: 10.1371/journal.pone.0098206
Henriques P., Delgado J., Sousa R., Ray S. (2017). Patellid limpets: an overview of the biology and conservation of keystone species of the rocky shores. Organismal Mol. Malacol., 71–95. doi: 10.5772/67862
Henriques P., Sousa R., Pinto A., Delgado J., Faria G., Alves A., et al. (2012). Life history traits of the exploited limpet Patella candei (mollusca: patellogastropoda) of the north-eastern atlantic. J. Mar. Biol. Assoc. United Kingdom 92, 1379–1387. doi: 10.1017/S0025315411001068
Hixon M. A., Johnson D. W., Sogard S. M. (2014). Boffffs: on the importance of conserving old-growth age structure in fishery populations. ICES J. Mar. Sci. 71, 2171–2185. doi: 10.1093/icesjms/fst200
Hunter J. R. (1992). Fecundity, spawning, and maturity of female dover sole Microstomus pacificus, with an evaluation of assumptions and precision. Fish. Bull.(Wash. DC) 90, 101–128.
Hunter J. R., Macewicz B. J. (1985). Rates of atresia in the ovary of captive and wild northern anchovy, Engraulis mordax. Fishery Bull. 83, 119–136.
Hunter J., Macewicz B. J., Kimbrell C. A. (1989). Fecundity and other aspects of the reproduction of sablefish, Anoplopoma fimbria, in central california waters. CalCOFI Rep. 30, 61–72.
Jones K. R., Klein C. J., Halpern B. S., Venter O., Grantham H., Kuempel C. D., et al. (2018). The location and protection status of earth’s diminishing marine wilderness. Curr. Biol. 28, 2506–2512. doi: 10.1016/j.cub.2018.06.010
Lambert Y. (2008). Why should we closely monitor fecundity in marine fish populations? J. Northwest Atlantic fishery Sci. 41, 93–106. doi: 10.2960/J.v41.m628
Lavin C. P., Jones G. P., Williamson D. H., Harrison H. B. (2021). Minimum size limits and the reproductive value of numerous, young, mature female fish. Proc. R. Soc. B 288, 20202714. doi: 10.1098/rspb.2020.2714
Liu O. R., Thomas L. R., Clemence M., Fujita R., Kritzer J. P., McDonald G., et al. (2016). An evaluation of harvest control methods for fishery management. Rev. Fish. Sci. Aquacul. 24, 244–263. doi: 10.1080/23308249.2016.1161002
Marshall D. J., Bode M., Mangel M., Arlinghaus R., Dick E. (2021). Reproductive hyperallometry and managing the world’s fisheries. Proc. Natl. Acad. Sci. 118, e2100695118. doi: 10.1073/pnas.2100695118
Martins G. M., Borges C. D., Vale M., Ribeiro P. A., Ferraz R. R., Martins H. R., et al. (2017). Exploitation promotes earlier sex change in a protandrous patellid limpet, Patella aspera rödin. Ecol. Evol. 7, 3616–3622. doi: 10.1002/ece3.2925
Mau A., Bingham J.-P., Soller F., Jha R. (2018). Maturation, spawning, and larval development in captive yellowfoot limpets (Cellana sandwicensis). Invertebrate Reprod. Dev. 62, 239–247. doi: 10.1080/07924259.2018.1505670
McCarthy M., Woosnam P., Culloty S. (2008). Histological investigation of the reproductive cycles of the limpets Patella vulgata and Patella ulyssiponensis. Mar. Biol. 153, 871–877. doi: 10.1007/s00227-007-0859-x
Murphy E., Rodhouse P., Nolan C. (1994). Modelling the selective effects of fishing on reproductive potential and population structure of squid. ICES J. Mar. Sci. 51, 299–313. doi: 10.1006/jmsc.1994.1031
Murua H., Kraus G., Saborido-Rey F., Witthames P., Thorsen A., Junquera S. (2003). Procedures to estimate fecundity of marine fish species from field samples in relation to reproductive strategy. J. Northwest Atlantic fishery Sci. 33, 33–54. doi: 10.2960/J.v33.a3
Neves A., Sequeira V., Vieira A. R., Silva E., Silva F D. A., Mendes S., et al. (2022). Reproduction of the blue jack mackerel, Trachurus picturatus, in western portugal: microscopic gonad analysis reveals indeterminate fecundity and skipped spawning patterns. Zool. Stud. 61, 41. doi: 10.6620/ZS.2022.61-41
Orton J., Southward A., Dodd J. (1956). Studies on the biology of limpets: ii. the breeding of Patella vulgata l. in britain. J. Mar. Biol. Assoc. United Kingdom 35, 149–176. doi: 10.1017/S0025315400009036
Pacifici M., Di Marco M., Watson J. E. (2020). Protected areas are now the last strongholds for many imperiled mammal species. Conserv. Lett. 13, e12748. doi: 10.1111/conl.12748
Pellowe K. E., Leslie H. M. (2020). Size-selective fishing leads to trade-offs between fishery productivity and reproductive capacity. Ecosphere 11, e03071. doi: 10.1002/ecs2.3071
Pinto A. R., Alves A., Faria G., Reis R., Delgado J. (2010). “Reproductive biology of Patella candei d’Orbigny 1840 in madeira archipelago,” in XVI Simposio Iberico de Estudios de Biologia Marina (Alicante: Departamento de Ciencias del Mar y Biología Aplicada de la Universidad de Alicante).
Prusina I., Ezgeta-Balić D., Ljubimir S., Dobroslavić T., Glamuzina B. (2014). On the reproduction of the mediterranean keystone limpet Patella rustica: histological overview. J. Mar. Biol. Assoc. United Kingdom 94, 1651–1660. doi: 10.1017/S0025315414000976
Ramirez Llodra E. (2002). Fecundity and life-history strategies in marine invertebrates. Adv. Mar. Biol. 43, 87—–170. doi: 10.1016/S0065-2881(02)43004-0
R Core Team (2022). R: a language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing).
Riera R., Pérez Ó., Álvarez O., Simón D., Díaz D., Monterroso Ó., et al. (2016). Clear regression of harvested intertidal mollusks. a 20-year, (1994–2014) comparative study. Mar. Environ. Res. 113, 56–61. doi: 10.1016/j.marenvres.2015.11.003
Schneider C. A., Rasband W. S., Eliceiri K. W. (2012). Nih image to imagej: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Sousa R., Delgado J., Pinto A. R., Henriques P. (2017). Growth and reproduction of the north-eastern atlantic keystone species Patella aspera (mollusca: patellogastropoda). Helgoland Mar. Res. 71, 1–13. doi: 10.1186/s10152-017-0488-9
Sousa R., Henriques P., Vasconcelos J., Pinto A. R., Delgado J., Riera R. (2020a). The protection effects of marine protected areas on exploited molluscs from an oceanic archipelago. Aquat. Conservat.: Mar. Freshw. Ecosyst. 30, 717–729. doi: 10.1002/aqc.3285
Sousa R., Riera R., Vasconcelos J., Gouveia L., Pinto A. R., Delgado J., et al. (2020b). Artisanal harvest of shellfish in the northeastern atlantic: the example of limpet and topshell fisheries in the archipelago of Madeira. In Ray S., Diarte-Plata G., Escamilla-Montes R. Invertebrates: ecophysiology and management (Rijeka: IntechOpen), 147–164.
Sousa R., Vasconcelos J., Henriques P., Pinto A. R., Delgado J., Riera R. (2019a). Long-term population status of two harvested intertidal grazers (Patella aspera and Patella candei), before, (1996–2006) and after, (2007–2017) the implementation of management measures. J. Sea Res. 144, 33–38. doi: 10.1016/j.seares.2018.11.002
Sousa R., Vasconcelos J., Riera R., Pinto A. R., Delgado J., Henriques P. (2019b). Potential impact of harvesting management measures on the reproductive parameters of the limpets Patella aspera and Patella candei from madeira island. Estuar. Coast. Shelf Sci. 226, 106264. doi: 10.1016/j.ecss.2019.106264
Steffen W., Broadgate W., Deutsch L., Gaffney O., Ludwig C. (2015). The trajectory of the anthropocene: the great acceleration. Anthropocene Rev. 2, 81–98. doi: 10.1177/2053019614564785
Tomczak M., Tomczak E. (2014). The need to report effect size estimates revisited. an overview of some recommended measures of effect size. Trends Sport Sci. 21, 19–25.
Vasconcelos J., Faria G., Freitas R., Gordo L. S. (2017). Fecundity regulation strategy of the blue jack mackerel, Trachurus picturatus (bowdich 1825), off madeira island (ne atlantic). Fish. Res. 190, 150–156. doi: 10.1016/j.fishres.2017.02.009
Vasconcelos J., Sousa R., Ferro J., Pinto A. R., Riera R. (2023). Fecundity strategy of the highly exploited limpet Patella ordinaria, from an oceanic archipelago. Reg. Stud. Mar. Sci. 60, 102849. doi: 10.1016/j.rsma.2023.102849
Vat L. S. (2000). The growth and reproduction of patella granularis (Mollusca: patellogastropoda) on the south-east coast of south Africa (Rhodes University).
Venter O., Sanderson E. W., Magrach A., Allan J. R., Beher J., Jones K. R., et al. (2016). Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun. 7, 1–11. doi: 10.1038/ncomms12558
Walsh M. R., Munch S. B., Chiba S., Conover D. O. (2006). Maladaptive changes in multiple traits caused by fishing: impediments to population recovery. Ecol. Lett. 9, 142–148. doi: 10.1111/j.1461-0248.2005.00858.x
Ward D., Melbourne-Thomas J., Pecl G. T., Evans K. ,. M., McCormack P. C., et al. (2022). Safeguarding marine life: conservation of biodiversity and ecosystems. Rev. Fish Biol. Fish. 32, 65–100. doi: 10.1007/s11160-022-09700-3
Worm B., Hilborn R., Baum J. K., Branch T. A., Collie J. S., Costello C., et al. (2009). Rebuilding global fisheries. Science 325, 578–585. doi: 10.1126/science.1173146
Keywords: reproduction, commercial species, conservation, limpet, intertidal, North-eastern Atlantic Ocean
Citation: Vasconcelos J, Sousa R, Ferro J, Pinto AR, Freitas M and Riera R (2023) Fecundity, an overlooked life-history trait for coastal management of commercial molluscs? Front. Mar. Sci. 10:1173629. doi: 10.3389/fmars.2023.1173629
Received: 24 February 2023; Accepted: 02 May 2023;
Published: 22 May 2023.
Edited by:
Diego Castejón Bueno, Centro Interdisciplinar de Investigação Marinha e Ambiental da Madeira (CIIMAR-Madeira), PortugalReviewed by:
Alan Hodgson, Rhodes University, South AfricaSher Khan Panhwar, University of Karachi, Pakistan
Copyright © 2023 Vasconcelos, Sousa, Ferro, Pinto, Freitas and Riera. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joana Vasconcelos, am9hbmEudmFzY29uY2Vsb3NAc3RhZmYudW1hLnB0; am9hbmFwYXRyaWNpYS5yZWlzQHVscGdjLmVz
†Present address: Joana Vasconcelos, Grupo en Biodiversidad y Conservación (BIOCON), IU-ECOAQUA, Universidad de Las Palmas de Gran Canaria, Marine Scientific and Technological Park, Telde, Spain