 Linda López-Abuchar1*
Linda López-Abuchar1* Christian Peralta-Madriz1Carolina Soto-Navarro2Gabriela Vinueza-Hidalgo1
Christian Peralta-Madriz1Carolina Soto-Navarro2Gabriela Vinueza-Hidalgo1 Andrew Whitworth1Christopher Beirne1
Andrew Whitworth1Christopher Beirne1- 1Mangrove Restoration and Conservation Program, Asociación Conservación Osa, Puerto Jiménez, Costa Rica
- 2Wilder Nature, Rewilding Europe, Heilig Landstichting, Netherlands
Mangrove forests are biodiversity hotspots that provide critical ecosystem services, from coastal protection to carbon storage. Yet, these ecosystems are disappearing at alarming rates, and while restoration efforts are expanding globally, long-term monitoring—especially in the Americas—remains scarce and often narrowly focused on vegetation structure, overlooking biological recovery. In this study, we evaluate whether gastropod community assemblages can serve as functional indicators of ecological recovery across different stages of mangrove restoration in Costa Rica. Using a space-for-time approach, we compared tree structure and gastropod assemblages across restored sites of varying ages, unrestored areas, and mature mangrove forests. We applied linear mixed models to examine how restoration stages influenced structure (tree height and DBH) and biodiversity metrics (gastropod abundance and composition). Tree structure improved consistently with restoration age, with five-year-old trees reaching nearly half the height and DBH of mature forest counterparts. Gastropod abundance exhibited a non-linear response—initially declining post-planting, then peaking by year five. Community composition also shifted: Melampus dominated early stages, while older sites supported Vitta, Cerithideopsis, and Littoraria. The exclusive presence of Thaisella in mature forests suggests its potential as a bioindicator of late-stage ecological recovery. Our results underscore the value of integrating biological indicators into mangrove monitoring. Gastropod assemblages offer a powerful lens through which to monitor ecological functionality, providing a low-cost, scalable tool to enhance adaptive management and guide future restoration efforts in tropical coastal ecosystems.
1 Introduction
Mangroves are among the world’s most productive ecosystems while occupying only 0.12% of the world’s total land area (Dodd and Ong, 2008; Nagelkerken et al., 2008). Mangroves provide multiple ecological services primarily related to coastal protection and carbon storage (Ong and Gong, 2013; Del Valle et al., 2019; Hilmi et al., 2022), while also sustaining rich assemblages of species, serving as breeding, refuge, and feeding zones for terrestrial and marine animals (Holguin et al., 2001; Hutchison et al., 2014; Wibowo et al., 2022). However, they are being destroyed at an alarming rate due to multiple anthropogenic factors (Kathiresan and Bingham, 2001; Hutchison et al., 2014; de Lacerda et al., 2019). Approximately 524,500 hectares of global mangrove area have been lost from 1996 to 2020 (Global Mangrove Watch, 2025). The loss of these ecosystems translates into detrimental effects on their ecological services and the livelihoods of coastal populations (Ellison et al., 2020; Lovelock et al., 2022). As a result, many mangrove restoration programs have been established worldwide to address this issue and reestablish mangroves in areas where they have been degraded or eradicated completely.
Reforestation programs often emphasize tree planting over restoring ecosystem functions, neglecting key ecological processes (Ellison, 2008; Kodikara et al., 2017; Lovelock et al., 2022). Monitoring is frequently under-prioritized during planning and, when conducted, is usually short-term and focused only on vegetation structure (Bosire et al., 2003; Ellison et al., 2020) This lack of long-term monitoring hinders the ability to assess project success or failure (Lovelock et al., 2022).
Robust ecological monitoring, community involvement, and transparent reporting are essential for adaptive management, which supports restoration success across ecological, political, and financial dimensions (Stokes et al., 2016; Ellison et al., 2020; Lovelock et al., 2022). However, monitoring often focuses only on vegetation structural parameters—such as growth rates, recruitment, and succession—while overlooking functional recovery, a key indicator of restoration success (Bosire et al., 2003; Thornton and Johnstone, 2015).
While assessing structural parameters of restored forests is essential for understanding early stabilization, it is critical to evaluate restoration success at the ecosystem level by monitoring biodiversity dynamics such as the recolonization of macrobenthic fauna (Bosire et al., 2008; Blanco and Castaño, 2012; Basyuni et al., 2022). Gastropods, key macrobenthic fauna of mangrove ecosystems (Cannicci et al., 2008; Lee, 2008; Ortiz and Blanco, 2012; Salmo et al., 2017), serve as ecosystem engineers and keystone species (Cannicci et al., 2008; Isroni et al., 2023), playing a vital role in food webs and nutrient recycling (Macintosh et al., 2002; Ghasemi et al., 2011; Batvari et al., 2016). Their limited mobility makes them highly sensitive to environmental changes (Baderan et al., 2019), positioning them as effective biotic indicators (Macintosh et al., 2002; Nordhaus et al., 2009; Blanco and Castaño, 2012; Salmo et al., 2017; Basyuni et al., 2022; Sujarta et al., 2022). Additionally, they hold economic value for local communities (Dewiyanti and Sofyatuddin, 2012; Awang et al., 2022). Despite their ecological and economic significance, studies on gastropod abundance, community dynamics, and ecological roles remain scarce (Ellison, 2008; Zvonareva et al., 2015).
In response to a past focus on short-term structural assessments and the growing recognition of the need to understand biological and ecological functions in mangroves, recent studies have increasingly evaluated macrobenthic communities to assess ecological recovery in restored mangroves, particularly in the Indo-pacific region (Ashton et al., 2003; Dewiyanti and Sofyatuddin, 2012; Zvonareva et al., 2015; Salmo et al., 2017; Baderan et al., 2019; Yadav et al., 2019; Adharyan Islamy and Hasan, 2020; Basyuni et al., 2022). Gastropod diversity, composition and distribution has also been studied in the Americas, including Colombia (Cantera et al., 1999; Vilardy and Polanía, 2002; Ortiz and Blanco, 2012), Mexico (Cruz-Abrego et al., 1994; Hernandez-Alcántara and Solis-Weiss, 1994), Costa Rica (Pomareda and Zanella, 2006; Vargas-Zamora and Sibaja-Cordero, 2011; Vargas et al., 2015), Venezuela (Balbas Acosta et al., 2013), Brazil (Collin R et al., 2005; Gorman and Turra, 2016) and Belize (Ellison and Farnsworth, 1990, 1992). Despite these efforts, there is still limited information on using gastropods as indicators to assess ecological rehabilitation and functional recovery in mangrove restoration, particularly in Central America and Costa Rica.
Our study evaluates for the first time in Central America, changes in gastropod abundance and community assemblages over time during a five-year mangrove restoration initiative in the Térraba-Sierpe National Wetland, the largest mangrove system in Costa Rica. Using a space-for-time approach that considers the age of the restored sites (planted between 2019 and 2023), we assess temporal changes in both structural and biological characteristics, comparing them to unrestored areas and mature mangrove forests. We hypothesize that older restoration sites will exhibit greater gastropod abundance and distinct community assemblages across different restoration stages. We hope this study serves as a guide to enhance mangrove restoration projects and ecological assessments across the Americas.
2 Materials and methods
2.1 Study site
The study was conducted at a series of community-driven restoration sites, with technical and funding support from locally based conservation NGO Osa Conservation, in the Térraba-Sierpe National Wetland (TSNW), Costa Rica (Figure 1). The TSNW is a protected area of approximately 30,000 hectares in the northwestern region of the Osa Peninsula, declared as Ramsar site in 1995 (Acuña-Piedra and Quesada-Román, 2017). This wetland hosts the largest mangrove forest in the country with an area of 17,736 hectares (Naranjo Loría, 2014; Barrantes and Peralta-Madriz, 2021). The TSNW is formed by two river basins: the Río Grande de Térraba and the Río Sierpe. The wetland is located between 0 and 5 m.a.s.l. and has an average temperature of 26.7 C° and average annual precipitation of 4000 mm (Solano and Villalobos; Jimenez-R and Soto, 1984; Naranjo Loría, 2014).
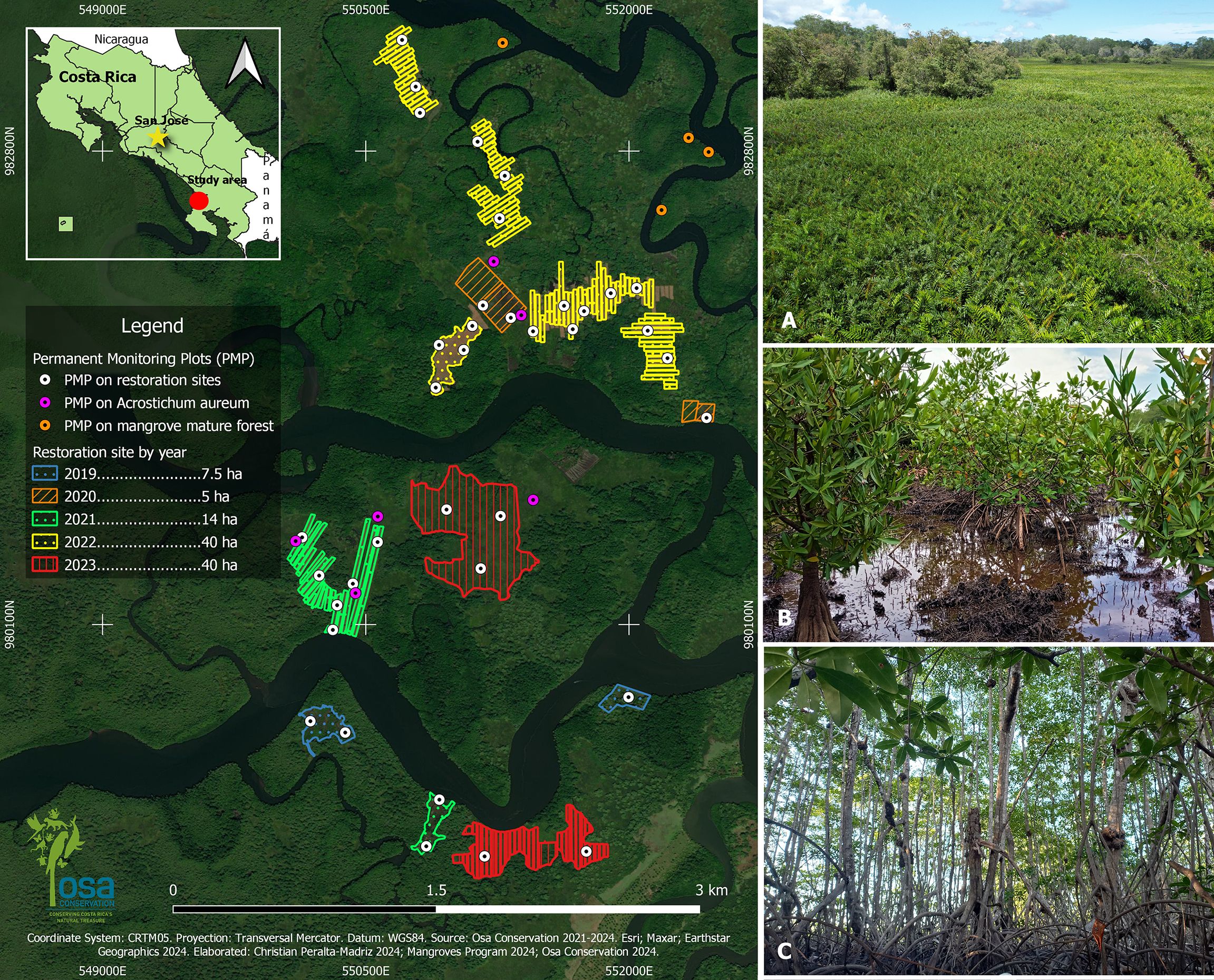
Figure 1. Osa Conservation restoration sites and monitoring plots within the Térraba- Sierpe National Wetland, located in the Osa Peninsula, southern Pacific Costa Rica (left) - as identified by the red circle on the country inlay. Color depicts restoration per year. Negraforra sites where mangrove was removed (A), restoration area depicting a 4 year old mangrove recovery site (B), mature mangrove forest (C).
Between 1972 and 1992, the wetland experienced its highest deforestation rate. Changes in economic activities led local communities to extract timber and charcoal, drastically reducing mangrove coverage (Acuña-Piedra and Quesada-Román, 2017). The reforested sites, initially devoid of tree cover, became dominated by the opportunistic fern Acrostichum aureum. While this fern is part of the local mangrove flora, its high density and the morphology of its rhizomes prevent the natural regeneration of mangrove trees and the establishment of propagules (Barrantes and Peralta-Madriz, 2021). Although there was a slight increase in mangrove forest area after the protected area was established in 1994, A. aureum still covered approximately 2,758 hectares of the wetland by 2012 (Leiva Barrantes and Cerdas, 2015; Acuña-Piedra and Quesada-Román, 2017).
Based on the above, SINAC, Costa Rica’s agency in charge of the administration and surveillance of protected areas, designated priority sites for active restoration within the wetland. Osa Conservation joined this national effort in 2018. To date, 171 hectares of mangroves area have been actively restored, where three species of mangroves have been planted (Pelliciera rhizophorae, Rhizophora mangle and Rhizophora racemosa), favouring the natural regeneration of other mangrove species (Avicennia germinans, Laguncularia racemosa) when the fern is cleared.
2.2 Gastropod sampling
We use a space-for-time approach to determine the influence of restoration age on gastropod communities, using plots of different ages to reflect progression of ecological restoration after planting. Restoration sites planted in 2019, 2020, 2022 and 2023 were included in the present study, as well as control sites in mature mangrove forest (the ultimate target of the restoration) and in sites dominated by Acrostichum aureum (common name Negraforra; as a negative control reflecting no regeneration has occurred). We established 100m2 Permanent Monitoring Plots (PMP) using a systematic randomized design in both control sites and restoration sites planted between 2019 and 2023. We conducted gastropod sampling in 41 PMPs (Figure 1). Each PMP consisted of three subplots of 50cm x 50cm located equidistantly along a diagonal. The sampling was done in 2023 and 2024 during February and March (dry season) following the methodology of the Ecological Mangrove Monitoring Protocol developed by SINAC-UNA (2020). Gastropods were sampled on all exposed roots, accessible trunk and branches, as well as the substrate and organic matter present in each subplot. Almost all individuals were identified in situ and unidentified individuals were collected and transferred to Osa Conservation laboratory facilities in Puerto Jimenez. Identifications were made to genus level with the taxonomic keys of Keen and Cruz (Keen, 1958; Cruz Soto and Jiménez Ramón, 1994).
2.3 Restoration structural characteristics
In each PMP the species, height and diameter of each tree was recorded. Height was measured using a tape measure from the base of the plant to the beginning of the apical meristem of the last leaf in formation. The diameter of the stem was measured with a vernier graduated in millimeters. Diameter measurement point varied according to the size of the individual: trees shorter than 50 cm were not measured; for trees between 50 and 150 cm in height with a diameter less than 10 cm, diameter was measured at 50 cm above the ground; and for trees taller than 150 cm with a diameter greater than 10 cm, diameter was measured at 130 cm. In the case of mature forest trees, height estimation was done with a Haglöf electronic clinometer (model ECII D) considering the base of the trunk at the level of the sediment and the highest visible part of the crown, and Diameter at Breast Height (DBH) was measured using a diametric taper, considering the highest root, as the basis for taking the measurement.
2.4 Statistical analysis
For each of the response terms outlined below, we implemented generalized linear mixed effects models using the ‘glmmTMB’ package (Brooks et al., 2017) in R.4.3.3 (R Core Team, 2023). In each model we included site as a random intercept term to account for the fact that multiple subsamples were taken within the same restoration site. For each response term, we compare the null model (a model with no fixed effects), to the full model (a model containing restoration sites according to the planting year as a categorical effect). We assess the strength of evidence that restoration treatment influences the response term in question through comparing the full model to the null model using AICc, and take any model which improves on the null model by a ΔAICc > 6 units as ‘strong’ statistical support for restoration age. We compare effect sizes between strata of interest to determine the biological significance of the changes observed. As a measure of goodness of fit, we use marginal (fixed effects only) and conditional (full model) pseudo-R2 (Nakagawa and Schielzeth, 2013). All models were checked using standard residual plotting techniques. We assessed 1) How do mangrove structural characteristics change with restoration age? 2) How does the restoration stage affect gastropods’ general abundance? 3) Is there a genus-specific response of gastropods to the mangrove restoration stage?
To assess how tree structure changed over time, we modeled average tree height and diameter at breast height (DBH) as response variables, with Restoration Year as a categorical fixed effect. This included restoration plots from 0.5 to 5 years post-planting, plus control treatments from mature mangroves and Negraforra (Acrostichum aureum); the latter was excluded from modeling due to the absence of trees. To examine how restoration influenced overall gastropod abundance, we fitted a generalized linear mixed model with the total count of individuals across all genera as the response. Finally, to assess shifts in community composition over time, we ran separate genus-specific models with each genus’ abundance as the response to restoration stage, grouping them into abundant and rare genera to improve model fit and enhance interpretability. All models mentioned above were fitted using a negative binomial distribution to account for overdispersion and the prevalence of zeros in our response variables.
To test for significant differences in gastropod community composition across restoration stages and sampling periods (monitoring years), we performed a permutational multivariate analysis of variance (PERMANOVA). To further explore patterns in community composition across restoration stages, we conducted a Non-metric Multidimensional Scaling (NMDS) analysis using the metaMDS function from the vegan package in R, using Euclidean distances. The abundance matrix included five genera (Melampus, Vitta, Littoraria, Cerithideopsis, and Thaisella). Restoration stages were represented by convex hulls to visualize clustering patterns. The final stress value of the solution was reported as a measure of model fit. To examine relationships between community structure and vegetation attributes, we applied vector fitting (envfit) using average tree height and diameter at breast height (DBH).
3 Results
3.1 How do mangrove structural characteristics change with restoration age?
Our results provide compelling statistical evidence for both tree height (ΔAICc = -83 from the null model; marginal R2 = 0.90; conditional R2 = 0.96) and DBH (ΔAICc = -88; marginal R2 = 0.89; conditional R2 = 0.97) increasing over time (Figure 2; Appendix 1 in Supplementary Material; Supplementary Table 1). After five years the predicted tree height reached 3.61 m (L95% = 221 cm; U95% = 591 cm), roughly half that of a mature mangrove stand and predicted DBH was 5.7 cm (L95% = 3.5 cm; U95%= 9.2 cm), more than half that of the average tree DBH in mature mangrove forest (8.7 cm).
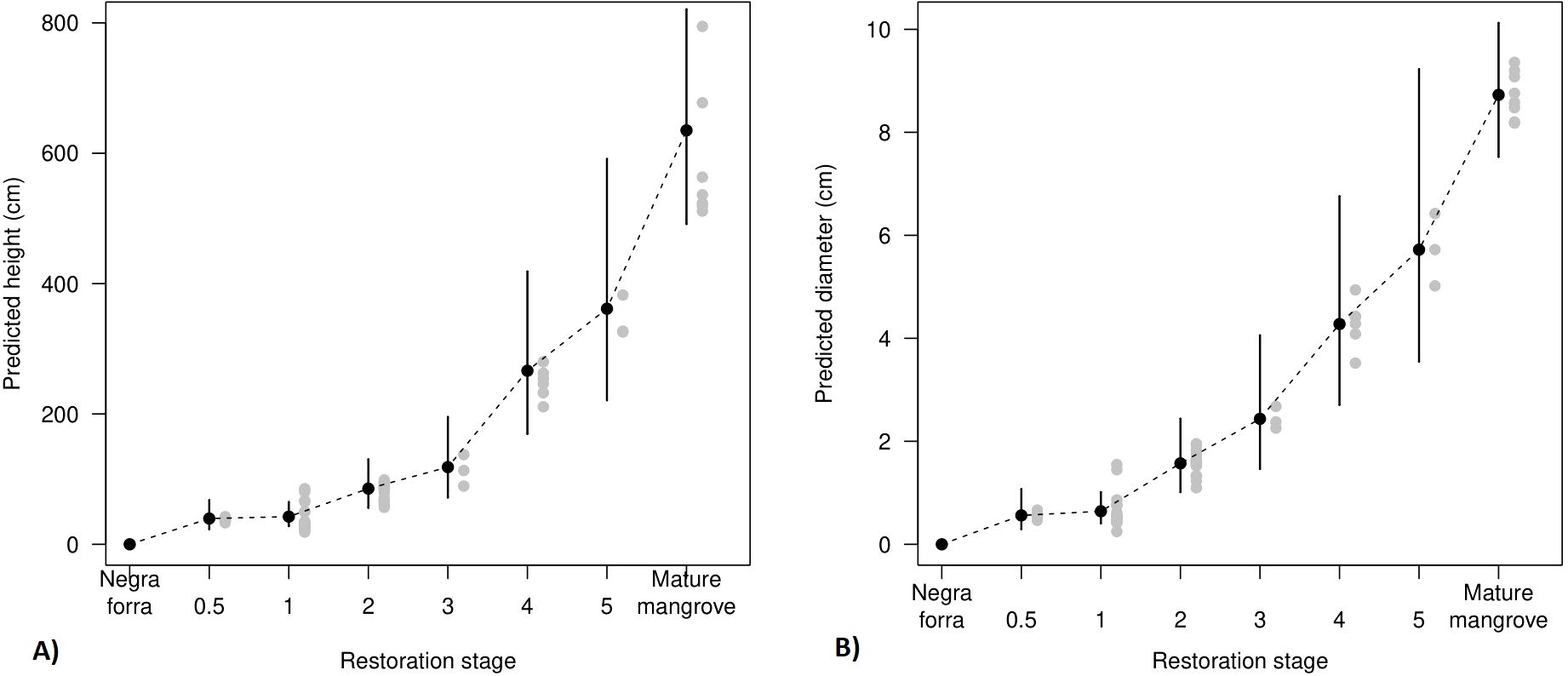
Figure 2. Predicted tree height (A) and DBH (B) increasing with time since restoration. Black points represent predicted mean population level; gray dots represent partial residuals (variance left over after accounting for all the predictors in the models); and vertical lines denote the 95% confidence intervals around the fixed effect for each restoration stage. As mangrove plants were completely absent from the Negraforra plots, this category was completely absent from the modelling process and is shown as zero height and DBH for illustrative purposes.
3.2 How does the restoration stage affect gastropods’ general abundance?
We recorded a total of 526 individuals representing five different genera: Melampus, Littoraria, Vitta, Cerithideopsis and Thaisella. Strong statistical evidence indicated restoration stage influencing gastropod general abundance (ΔAICc= -38.5; marginal R2 = 0.67; conditional R2 = 0.67) with non-linear patterns (Figure 3; Appendix 2 in Supplementary Material; Supplementary Table 3). Gastropod abundance dropped from 3 to 0.6 individuals per plot one year after restoration, then steadily increased, peaking at nearly 30 individuals by year 5. Although abundance declined in mature mangroves (7.25), it remained higher than in the Negraforra control.
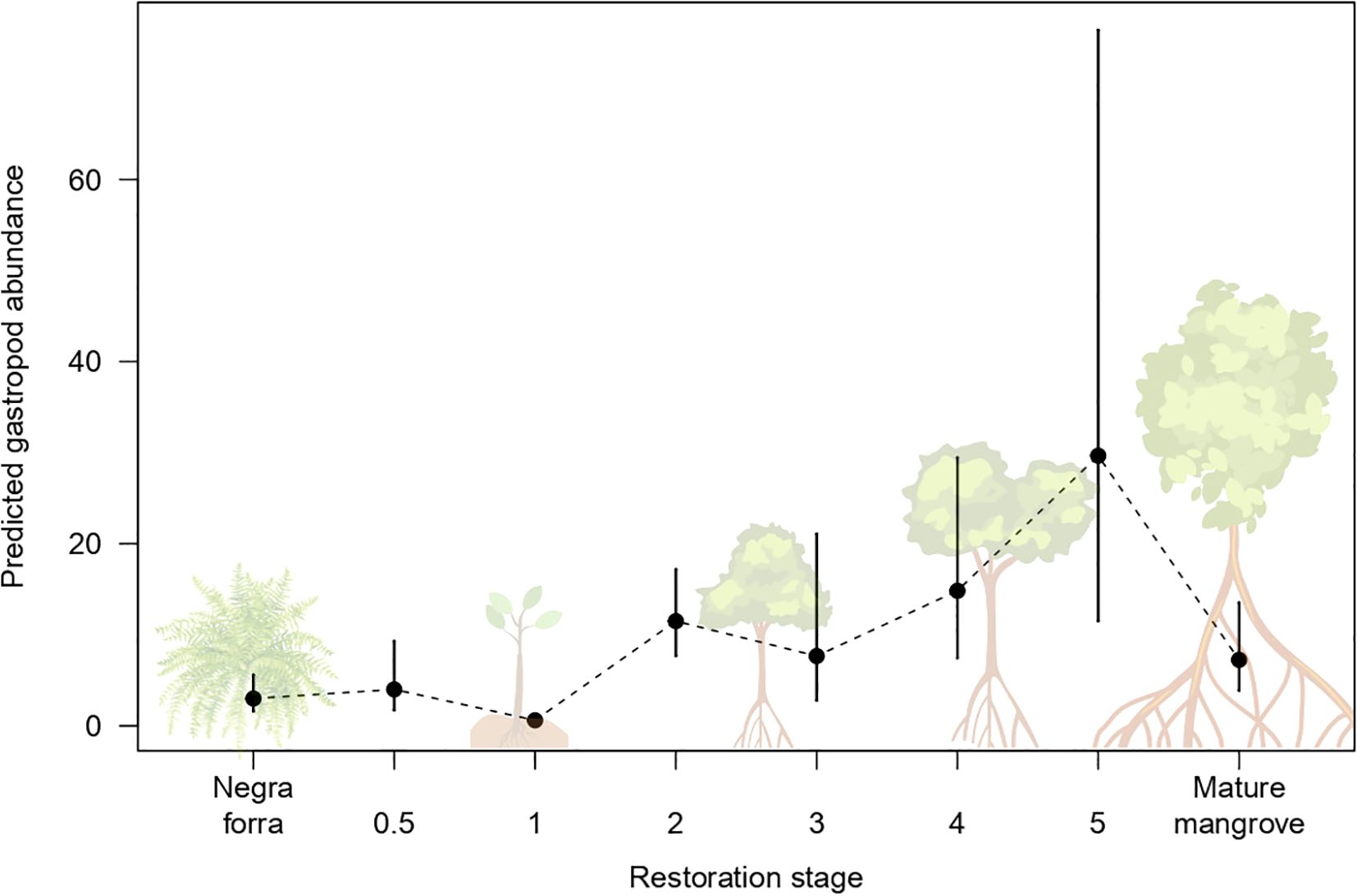
Figure 3. Predicted overall gastropod abundance vs restoration stages (years since restoration). Black points represent predicted mean population level and lines denote the 95% confidence intervals around the fixed effects.
3.3 Is there a genus-specific response of gastropods to the mangrove restoration stage?
Genus-specific assemblages varied notably across restoration stages (Figure 4; Appendix 4.1-4.5 in Supplementary Material; Supplementary Tables 4-8) with three general patterns observed: 1) an increase with restoration progression in time (Vitta, Littoraria, and Cerithideopsis), 2) a decrease with restoration temporal progression (Melampus), and 3) presence observed only in mature mangrove (Thaisella). However, the patterns of change are not consistent between the genera, which increase with the restoration stage. While Littoraria abundance generally increases with time in restoration sites, reaching its peak in mature mangrove, Vitta and Cerithideopsis initially increase with restoration progression, being higher in the oldest restoration site (year five) but then significantly decreasing in mature mangroves. Thaisella, only present in mature mangroves, was absent from all restored and unrestored sites.
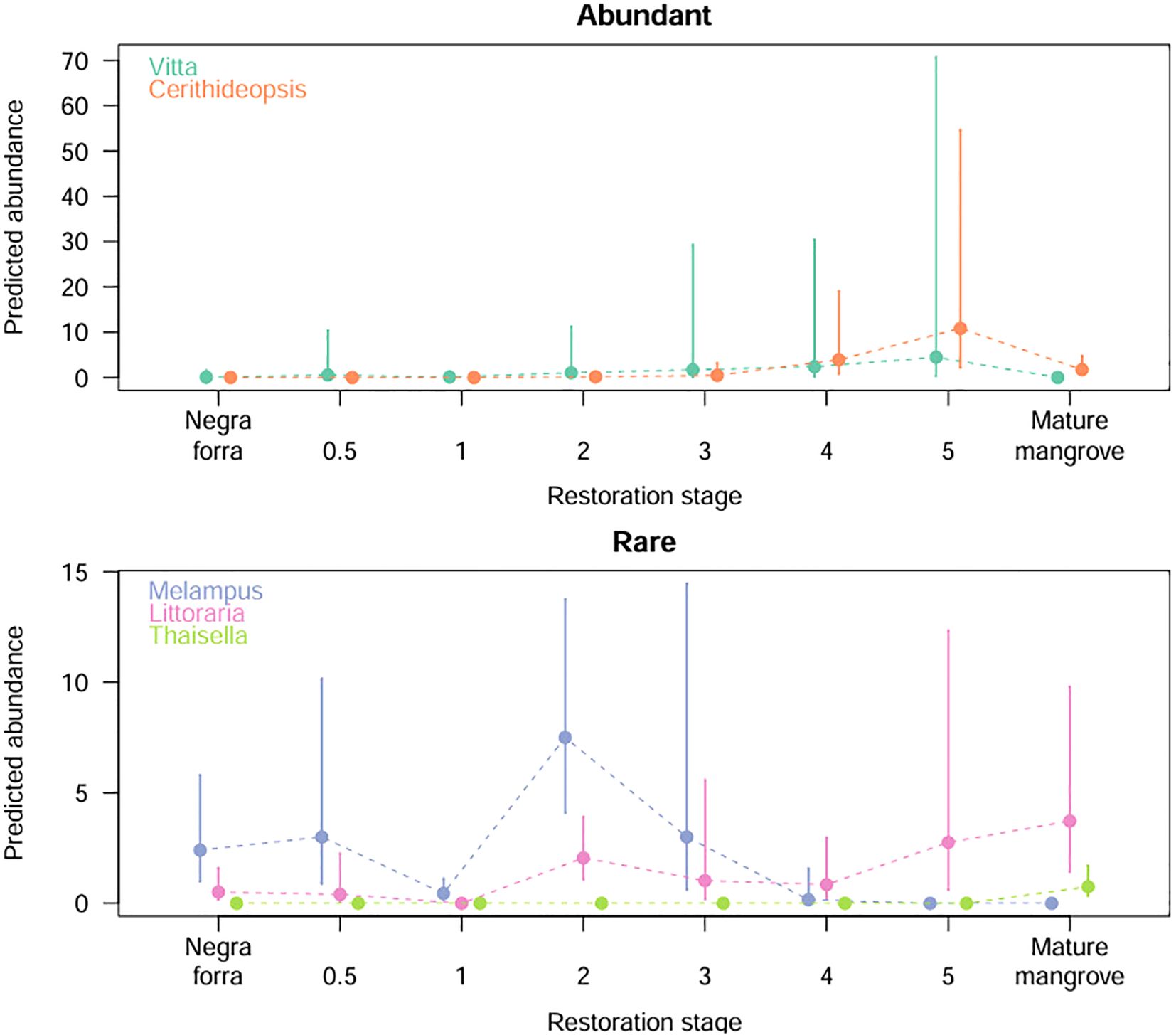
Figure 4. Predicted genus-specific abundance changes with restoration progression, where Vitta and Cerithideopsis were pooled as abundant taxa and Melampus, Littoraria and Thaisella as rare. Colored lines confidence intervals around the mean abundance estimate (note - confidence intervals for restoration stages with zero individuals were excluded from the plots, as there was no variation in the reference category).
We confirmed significant and consistent differences in genus-specific compositions between restoration years, as indicated by PERMANOVA results (R² = 0.39, P = 0.001) (Appendix 6 in Supplementary Material; Supplementary Table 10). We also assessed sampling year as an explanatory variable in the PERMANOVA but as it only explained less than 1% of variation, we do not consider it here. Community-level compositional shifts became more pronounced as restoration progressed, as visualized through the Non-metric Multidimensional Scaling (NMDS) (Figure 5). Sites from Negraforra and early restoration stages (years 0.5 to 3) show overlapping compositions, although sites from three years post-restoration began to diverge slightly. In contrast, sites from years 4 and 5 cluster closely together, indicating a distinct community composition likely driven by the increased prevalence of Vitta and Cerithideopsis. Mature forest sites are clearly separated from all restoration stages, reflecting their unique and established assemblage structure, characterized by the exclusive presence of Thaisella. Both tree height and diameter were highly correlated with the NMDS axes (P = 0.001) and accounted for 26% and 30% of the variation respectively (Appendix 5 in Supplementary Material; Supplementary Table 9).
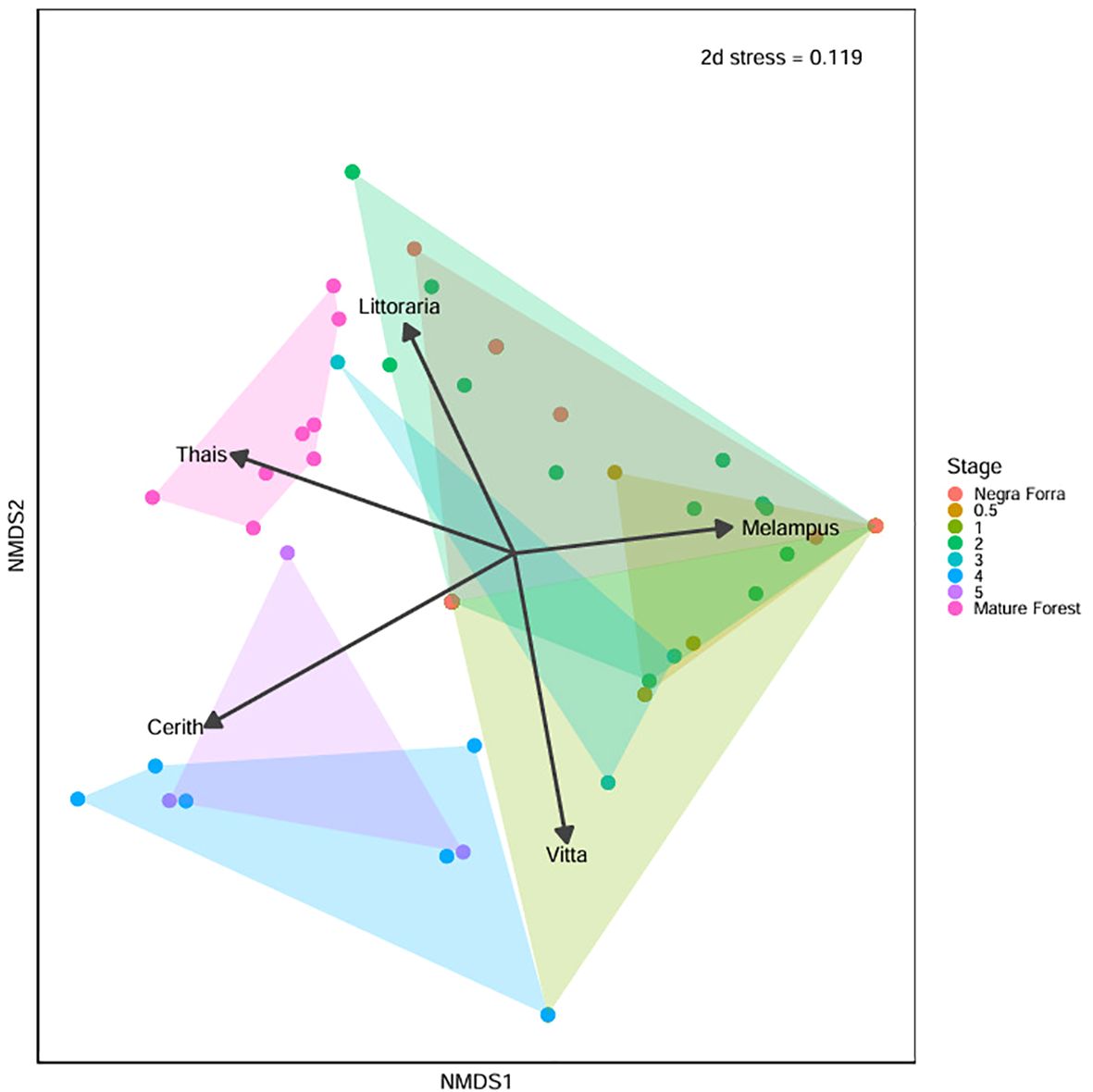
Figure 5. Plot resulting from the NMDS analysis of the gastropod communities based on Euclidean dissimilarity to visualize differences in gastropod community composition across restoration stages. The final 2D solution had a stress value of 0.119, indicating a good representation of the data.
4 Discussion
4.1 Restored mangrove structure improves rapidly through time
We found that, after just five years, the height and DBH of actively planted mangroves averaged about half of those in mature mangroves. These structural parameters, which increase with stand age are commonly used to assess ecosystem recovery (Ferreira et al., 2015; Azman et al., 2021), proved effective for monitoring short-term changes in mangrove forests. Interestingly, the average tree height in our study aligned with that observed in other five-year-old actively restored mangroves in the region, while the average DBH was significantly higher.
For comparison, Ferreira et al (2015) reported a 4.12m height and 1.85cm DBH in northeastern Brazil, whereas in southern Brazil, the average height was 1.23 m (DBH not reported). Similarly low values were recorded in Cuba 1.78 m height and 1.85 cm DBH. These other sites typically involved monoculture plantings of Rhizophora mangle, which may limit restoration outcomes, as monocultures are often less effective for ecological rehabilitation (Bosire et al., 2008). In contrast, our restoration sites included three species, potentially enhancing forest heterogeneity, improving structural development, and reducing mortality due to propagule predation by crabs (Ellison, 2008). Growth variations are also influenced by site-specific factors such as soil texture, tidal amplitude, salinity, and density and diversity of planted species (Zvonareva et al., 2015; Ellison, 2008), especially across intertidal zones.
We also compared planted mangrove structure with two other habitat types: unrestored areas (Negraforra) and mature mangroves. As expected, no mangrove trees or seedlings were found in Negraforra sites, where the fern Acrostichum aureum inhibits natural mangrove regeneration (Dahdouh-Guebas et al., 2004; Blanco and Castaño, 2012; Censkowsky, 2022). These findings highlight the necessity of active restoration to recover ecological function in degraded mangrove areas colonized by A. aureum in the Térraba-Sierpe National Wetland.
Interestingly, the average height of mature mangrove forests in our study was lower than the 10.8 m reported for mangroves in the TSNW (Jimenez-R and Soto, 1984) and the 10 m recorded for intact forests in Costa Rica’s North Pacific region (Cordero-Murillo et al., 2023). It was also shorter than the 15.4 m average height and 13.4 cm DBH observed in nearby Golfo Dulce mangroves (Samper-Villarreal and Silva-Benavides, 2015). This discrepancy likely results from historical clearing in TSNW for tannin extraction prior to its protection in 1994. Mature forests that have not been degraded are generally more structurally complex than younger stands (Samper-Villarreal and Silva-Benavides, 2015), suggesting that Golfo Dulce mangrove may be older than those in the TSNW. Despite these differences, we are confident in our data, as the mature mangrove sites were selected based on local knowledge of undisturbed areas over the last decade.
Globally, mangrove restoration outcomes remain poorly documented particularly in Latin America and the Caribbean (O’Connell et al., 2021; Friess et al., 2022; Lovelock et al., 2022). Nevertheless, several evaluations of restored and naturally regenerated mangrove forests have been conducted in the Indo-Pacific. Azman et al. (2021), in a comparative study in Malaysia, found that while naturally regenerated forests had higher tree species diversity, it was the restored mangroves which rapidly increased biomass and more closely resembled intact forests in structure. Similarly, Luo et al. (2010) observed no significant structural differences between planted and natural forests after 50 years. In Kenya, Bosire et al. (2003) found that restored stands support seedling recruitment by providing protective structural development, unlike unrestored mangroves. Given the positive growth of all three planted species in our study, continued restoration and maintenance in the TSNW is strongly recommended to support the recovery of habitat functionality.
4.2 Overall gastropod abundance increases with restoration time, and community assemblages differ across restoration stages
Our results also show that gastropod abundance in restored mangroves increased non-linearly, initially decreasing in the first year after planting, then rising over time. By year five, gastropod abundance exceeded that of mature mangrove forests. This trend—characterized by an initial rise followed by a plateau or decline—mirrors patterns observed in other restoration efforts (Dewiyanti and Sofyatuddin, 2012; Zvonareva et al., 2015; Salmo et al., 2017; Chen et al., 2021).
This increase in abundance is closely linked to the development of vegetation structure, which provides shelter, predator protection, and food for macrobenthic fauna (Skilleter and Warren, 2000; Dewiyanti and Sofyatuddin, 2012; Gorman and Turra, 2016; Harefa et al., 2024). Additional factors—including mangrove species diversity, organic carbon content, sediment parameters, salinity, pH, tidal regime, and soil moisture—also significantly influence gastropod recruitment (Macintosh et al., 2002; Dewiyanti and Sofyatuddin, 2012; Hookham et al., 2014; Salmo et al., 2017). For instance, Dewiyanti and Sofyatuddin (2012) found the highest gastropod abundance in four-year-old restoration sites with elevated organic carbon, while Salmo et al. (2017) linked shifts in species composition to increased forest cover and organic matter. Although we did not measure these abiotic factors, we highly recommend their inclusion in future studies to better explain gastropod community changes.
Community-level trends identified through the NMDS analysis were marked by overlap in early restoration stages but began to clearly diverge by years 4 and 5, ultimately showing marked differences from mature forest composition, highlighting the importance of assessing changes in community assemblages on a long term basis. This approach provides valuable insights for tracking restoration progress and evaluating the ecological success of mangrove rehabilitation efforts.
Our findings, in line with studies across the Indo-Pacific, support the use of gastropod abundance and community composition as early indicators of successional progress in mangrove restoration (Dewiyanti and Sofyatuddin, 2012; Zvonareva et al., 2015; Awang et al., 2022; Basyuni et al., 2022). We expect that, as our sites mature, gastropod abundance will stabilize and community assemblages will more closely resemble those of mature forests. As noted by Zvonareva et al. (2015) the dominance of opportunistic organisms over mangrove-associated taxa may indicate that the ecosystem has not yet reached a state of maturity.
Opportunistic epifaunal gastropods are commonly found in young or intermediate forests, which offer habitat heterogeneity, combining open areas with canopy-covered zones (Macintosh et al., 2002; Salmo et al., 2017; Harefa et al., 2024). In contrast, mature forests are typically dominated by arboreal mangrove-associated gastropods (Basyuni et al., 2022; Salmo et al., 2017). In our study, Melampus was more abundant in early restoration sites, and declined thereafter, while Vitta and Cerithideopsis abundance increased after year 3. Littoraria was present across all sampled sites, (except in year 1), highlighting its adaptability. Thaisella kiosquiformis (identified to species level) was exclusively observed in mature forest sites.
Both Melampus and Vitta -detritivorous grazers- were abundant during early restoration stages. Melampus, which thrives in shaded areas with detritus (Proffitt and Devlin, 2005; Lee and Silliman, 2006), peaked in two-year-old sites, where tree growth was limited, and decaying Negraforra rhizomes were common (personal observation). Its decline over time may be due to increasing competition from more dominant grazers like Littoraria. Vitta, on the other hand, became more common after year three, likely benefiting from increased canopy cover and reduced A. aureum biomass, which improved habitat conditions for larval recruitment (Blanco and Castaño, 2012). Vitta was absent from mature mangrove sites, possibly due to reduced soil litter (its main food source (Cantera et al., 1983) and increased interspecific competition (Lee and Silliman, 2006).
Littoraria was found across all restoration stages (except in year one), Negraforra and mature forests. This genus, known for its high mobility and dietary adaptability - feeding on algae, fungi and leaf sprouts (Banco and Cantera, 1999; Macintosh et al., 2002; Ellison, 2008) - includes many mangrove-associated species (Reid, 1985; Ohgaki, 1992; Zvonareva et al., 2015; Chen et al., 2021). Given the ecological differences among species, species-level identification is essential for accurate ecological assessments - something we recommend for future studies, as it was a limitation in ours.
Cerithideopsis, a genus of epifaunal detritivores from the Potamididae family, was recorded only in sites older than three years, with peak abundance observed in year five-year-old sites. These gastropods, adapted to shaded mud substrates (Cantera et al., 1999; Pomareda and Zanella, 2006) and tree climbing, thrive in mature forest conditions (Reid et al., 2008). Their presence aligns with increased canopy cover and substrate complexity in older sites (Cantera et al., 1999).
Thaisella kiosquiformis, a predatory species of sessile mollusks and barnacles, was found only in mature forests and it was typically observed on fallen wood (Banco and Cantera, 1999; Cantera et al., 1999; Simone, 2017). Its absence from restoration sites likely reflects that these ecosystems have not yet reached the stage of ecological maturity required for its establishment. Its presence could serve as a useful bioindicator for advanced forest development; however, further understanding of reproduction dynamics, recruitment, and the proximity of adjacent mature forests as potential source populations is needed.
We anticipate that further structural development will enable colonization by additional mangrove-associated species, including T. kiosquiformis. Based on Salmo et al. (2017), this shift may become apparent 11–15 years post-restoration, signaling functional recovery through stabilized gastropod abundance and mature community assemblage, resembling that of natural mature mangrove stands.
Our study faced limitations in assessing gastropod diversity due to the lack of comparable data from other restored mangrove sites in Costa Rica, as well as seasonal constraints, since sampling was conducted only during the dry season. We recommend that future studies include multiple seasons to capture potential temporal variations in gastropod communities. However, the presence of genera typical of intermediate forest stages, even after only five years, indicates successful habitat rehabilitation (Macintosh et al., 2002). This suggests that the restored mangrove ecosystem is already providing the essential resources and habitat conditions necessary for these taxa to thrive, while also reflecting specific community-level trends that align with the restoration trajectory.
In line with our hypothesis, we found that older restoration sites—particularly those five years post-planting—exhibit greater gastropod abundance. Community assemblages also varied significantly across restoration stages, with early sites (up to three years) showing similar compositions, and a clear shift occurring in restoration years four and five.
We strongly recommend future research on Thaisella kiosquiformis to establish its utility as a bioindicator of mature, stable restored mangroves ecosystems. Long-term monitoring -15 to 20 years- is essential to fully track and understand ecosystem development and inform adaptive management. Identifying gastropods to species level will further clarify their ecological roles and improve restoration assessments. Our results provide compelling evidence for the effectiveness of active restoration initiatives, which, informed by prior assessments of degradation causes, aid in rehabilitating the ecological functionality of mangroves, particularly in promoting macrobenthic recolonization (Gorman and Turra, 2016; Salmo et al., 2017; Harefa et al., 2024).
As the first study of its kind in Central American mangroves, our findings highlight the importance of incorporating faunal indicators, such as gastropods, into monitoring plans. Our results show that mangrove tree height and DBH increase with restoration age -comparable in height to other regional projects, and with higher average DBH. Gastropod abundance also increased with restoration age, and shifts in community assemblages reflect ecological succession. We hope this study encourages further research in Latin America and the Caribbean on gastropods as ecological indicators of mangrove restoration trajectories.
Data availability statement
Database and Rscript used for developing this paper are available in the Zenodo repository under DOI: 10.5281/zenodo.15787013 (https://doi.org/10.5281/zenodo.15787013).
Ethics statement
Animal sampling was approved by National System of Conservation Areas (SINAC). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
LL-A: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. CP-M: Conceptualization, Data curation, Investigation, Methodology, Resources, Visualization, Writing – review & editing. CS-N: Conceptualization, Funding acquisition, Investigation, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing. GV-H: Funding acquisition, Project administration, Validation, Writing – review & editing. AW: Conceptualization, Funding acquisition, Validation, Writing – review & editing. CB: Data curation, Formal Analysis, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declared financial support was received for the research and/or publication of this article. This work was supported by BAND Foundation, The Overbrook Foundation, One Tree Planted and Plant-for-the Planet US.
Acknowledgments
We thank the National System of Protected Areas-SINAC for the permits to conduct this restoration project and this research. We thank the local associations APREMAA and APEAHTS whose dedicated work has been pivotal in achieving our results in mangrove restoration within the Térraba Sierpe National Wetland.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be constructed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1563965/full#supplementary-material
References
Acuña-Piedra J. F. and Quesada-Román A. (2017). Cambios en el uso y cobertura de la tierra entre 1948 y 2012 en el Humedal Nacional Térraba-Sierpe, Costa Rica. Rev. Cienc. Marinas y Costeras 9, 9. doi: 10.15359/revmar.9-2.1
Adharyan Islamy R. and Hasan V. (2020). Checklist of mangrove snails (Mollusca: Gastropoda) in south coast of pamekasan, Madura Island, East Java, Indonesia. Biodiversitas 21, 3127–3134. doi: 10.13057/biodiv/d210733
Ashton E. C., Macintosh D. J., and Hogarth P. J. (2003). A baseline study of the diversity and community ecology of crab and molluscan macrofauna in the Sematan mangrove forest, Sarawak, Malaysia. J. Trop. Ecol. 19, 127–142. doi: 10.1017/S0266467403003158
Awang U. N., Jasni N. A. A., Sahari M. S. I., Nor S. M. M., Baharuddin N., Deraman M. Y., et al. (2022). Abundance of marine gastropods (Mollusca: Gastropoda) in replanted mangrove forests along. Available online at: http://www.bioflux.com.ro/aacl. (Accessed February 04, 2024).
Azman M. S., Sharma S., Shaharudin M. A. M., Hamzah M. L., Adibah S. N., Zakaria R. M., et al. (2021). Stand structure, biomass and dynamics of naturally regenerated and restored mangroves in Malaysia. For Ecol. Manage 482. doi: 10.1016/j.foreco.2020.118852
Baderan D. W. K., Hamidun M. S., Utina R., Rahim S., and Dali R. (2019). The abundance and diversity of mollusks in mangrove ecosystem at coastal area of North Sulawesi, Indonesia. Biodiversitas 20, 987–993. doi: 10.13057/biodiv/d200408
Balbas Acosta V., Betancourt R., and Arcas Prieto A. (2013). Estructura comunitaria de bivalvos y gasterópodos en raíces del mangle rojo Rhizophora mangle (Rhizophoraceae) en isla Larga, bahía de Mochima, Venezuela. Rev. Biología Tropical. 62 (2), 551–565.
Banco J. F. and Cantera J. R. (1999) The Vertical Distribution of Mangrove Gastropods and Environmental Factors Relative to Tide Level at Buenaventura Bay, Pacific Coast of Colombia. Available online at: https://www.researchgate.net/publication/233490929. (Accessed October 10, 2024).
Barrantes M. and Peralta-Madriz C. (2021). Fortalecimiento efectivo de los ecosistemas mangles en Costa Rica (Térraba-Sierpe) y mejoramiento de la calidad de vida de la población costera local. (Estrategia Nacional de Biodiversidad de Costa Rica & SINAC)
Basyuni M., Bimantara Y., Cuc N. T. K., Balke T., and Vovides A. G. (2022). Macrozoobenthic community assemblage as key indicator for mangrove restoration success in North Sumatra and Aceh, Indonesia. Restor. Ecol. 30 (7), e13614. doi: 10.1111/rec.13614
Batvari B. P. D., Sivakumar S., Shanthi K., Lee K. J., Oh B. T., Krishnamoorthy R., et al. (2016). Heavy metals accumulation in crab and shrimps from Pulicat lake, north Chennai coastal region, southeast coast of India. Toxicol. Ind. Health 32, 1–6. doi: 10.1177/0748233713475500
Blanco J. F. and Castaño M. C. (2012). Efecto de la conversión del manglar a potrero sobre la densidad y tallas de dos gasterópodos en el delta del río Turbo (golfo de Urabá, Caribe Colombiano). (Revista Biologia Tropical)
Bosire J. O., Dahdouh-Guebas F., Kairo J. G., and Koedam N. (2003). Colonization of non-planted mangrove species into restored mangrove stands in Gazi Bay, Kenya. Aquat Bot. 76, 267–279. doi: 10.1016/S0304-3770(03)00054-8
Bosire J. O., Dahdouh-Guebas F., Walton M., Crona B. I., Lewis R. R., Field C., et al. (2008). Functionality of restored mangroves: A review. Aquat Bot. 89, 251–259. doi: 10.1016/j.aquabot.2008.03.010
Brooks M. E., Kristensen K., van Benthem K. J., Magnusson A., Berg C. W., Nielsen A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/rj-2017-066
Cannicci S., Burrows D., Fratini S., Smith T. J., Offenberg J., and Dahdouh-Guebas F. (2008). Faunal impact on vegetation structure and ecosystem function in mangrove forests: A review. Aquat Bot. 89, 186–200. doi: 10.1016/j.aquabot.2008.01.009
Cantera J., Arnaud P. M., and Thomassin B. A. (1983). Biogeographic and ecological remarks on molluscan distribution in mangrove biotopes. 1. Gastropods. J. moll. Stud. 12, 10–26.
Cantera J. R., Thomassin B. A., and Arnaud P. M. (1999). Faunal zonation and assemblages in the Pacific Colombian mangroves. Hydrobiologia 413, 17–33. doi: 10.1023/A:1003890826741
Censkowsky U. (2022). Conserving mangrove ecosystems, securing livelihoods-the seafood sector can play its part. The international journal for rural development. 01/22.
Chen S., Xing B., Yu W., Chen B., Liao J., An W., et al. (2021). Distribution of a newly recorded gastropod species, mainwaringia leithii (Gastropoda, littorinidae), in young, rehabilitated mangroves in China. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.770963
Collin R B., Fernandes C. M., Gomes C. P., Brito B. A., Lima dos Santos S., and Tagliaro C. H. (2005). Molluscan diversity and abundance among coastal habitats of Northern Brazil. Ecotropica. 11, 9–20.
Cordero-Murillo A. L., Acosta-Vargas L. G., Pineda-Gómez J. A., and Torres-Gómez D. (2023). Estructura, composición y contenido de carbono de manglares en los Humedales Níspero y San Buenaventura-Colorado, Golfo de Nicoya, Costa Rica. Rev. Forestal Mesoamericana Kurú.
Cruz-Abrego F. M., Toledano-Granados A., and Flores-Andolais F. (1994). Ecología comunitaria de los gasterópodos marinos (MoUusca: Gastropoda) en Isla Contoy, México. Revista Biologia Tropical. 439–441.
Cruz Soto R. A. and Jiménez Ramón J. A. (1994). Moluscos asociados a las áreas de manglar de la Costa Pacífica de América Central: Guía (Heredia, Costa Rica: Editorial Fundación UNA).
Dahdouh-Guebas F., Van Hiel E., Chan J. C. W., Jayatissa L. P., and Koedam N. (2004). Qualitative distinction of congeneric and introgressive mangrove species in mixed patchy forest assemblages using high spatial resolution remotely sensed imagery (IKONOS). Syst. Biodivers 2, 113–119. doi: 10.1017/S1477200004001422
de Lacerda L. D., Borges R., and Ferreira A. C. (2019). Neotropical mangroves: Conservation and sustainable use in a scenario of global climate change. Aquat Conserv. 29, 1347–1364. doi: 10.1002/aqc.3119
Del Valle A., Eriksson M., Ishizawa O. A., and Miranda J. J. (2019). Mangroves for Coastal Protection Evidence from Hurricanes in Central America. Available online at: http://www.worldbank.org/research. (Accessed February 10, 2024).
Dewiyanti I. and Sofyatuddin K. (2012). Diversity of Gastropods and Bivalves in mangrove ecosystem rehabilitation areas in Aceh Besar and Banda Aceh districts, Indonesia. AACL Bioflux. 5(2), 55–59
Dodd R. S. and Ong J. E. (2008). "Future of mangrove ecosystems to 2025," in Aquatic ecosystems: Trends and global prospects, ed. N. V. C. Polunin. (Cambridge: Foundation for Environmental Conservation, Cambridge University Press), 172–187. doi: 10.1017/CBO9780511751790.017
Ellison A. M. (2008). Managing mangroves with benthic biodiversity in mind: Moving beyond roving banditry. J. Sea Res. 59, 2–15. doi: 10.1016/j.seares.2007.05.003
Ellison A. M. and Farnsworth E. J. (1990). The ecology of Belizean mangrove-root fouling communities. I. Epibenthic fauna are barriers to isopod attack of red mangrove roots. Journal of Experimental Marine Biology and Ecology 247, 87–98.
Ellison A. M. and Farnsworth E. J. (1992). The ecology of Belizean mangrove-root fouling communities: patterns of epibiont distribution and abundance, and effects of root growth. Hydrobiologia. doi: 10.1007/BF00008207
Ellison A. M., Felson A. J., and Friess D. A. (2020). Mangrove rehabilitation and restoration as experimental adaptive management. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00327
Ferreira A. C., Ganade G., and Luiz de Attayde J. (2015). Restoration versus natural regeneration in a neotropical mangrove: Effects on plant biomass and crab communities. Ocean Coast. Manag 110, 38–45. doi: 10.1016/j.ocecoaman.2015.03.006
Friess D. A., Gatt Y. M., Ahmad R., Brown B. M., Sidik F., and Wodehouse D. (2022). Achieving ambitious mangrove restoration targets will need a transdisciplinary and evidence-informed approach. One Earth 5, 456–460. doi: 10.1016/j.oneear.2022.04.013
Ghasemi S., Zakaria M., and Hoveizeh Mola N. (2011). Abundance of molluscs (Gastropods) at mangrove forests of Iran. J. Am. Sci.
Global Mangrove Watch (2025). *Global Mangrove Watch*. Available at: https://www.globalmangrovewatch.org/?bounds=[[-333.7198759692009,-73.42852308178541],[146.5144990308034,74.68315820119335]] (Accessed July 1, 2025).
Gorman D. and Turra A. (2016). The role of mangrove revegetation as a means of restoring macrofaunal communities along degraded coasts. Sci. Total Environ. 566–567, 223–229. doi: 10.1016/j.scitotenv.2016.05.089
Harefa M. S., Nasution Z., Tuhono E., Susilowati A., and Wulandari W. (2024). “Diversity of bivalve and gastropod species in mangrove restoration areas,” in IOP Conference Series: Earth and Environmental Science. doi: 10.1088/1755-1315/1352/1/012062
Hernandez-Alcántara P. and Solis-Weiss V. (1994). Algunas comunidades macrobénticas asociadas al manglar (Rhizophora mangle) en laguna de Términos, Golfo de México. Rev. Biología Tropical. 43(1-3), 117–129.
Hilmi E., Sari L. K., Cahyo T. N., Mahdiana A., Soedibya P. H. T., and Sudiana E. (2022). Survival and growth rates of mangroves planted in vertical and horizontal aquaponic systems in North Jakarta, Indonesia. Biodiversitas 23, 687–694. doi: 10.13057/biodiv/d230213
Holguin G., Vazquez P., and Bashan Y. (2001). The role of sediment microorganisms in the productivity, conservation, and rehabilitation of mangrove ecosystems: An overview. Biol. Fertil Soils 33, 265–278. doi: 10.1007/s003740000319
Hookham B., Tan Shau-Hwai A., Dayrat B., and Hintz W. (2014). A baseline measure of tree and gastropod biodiversity in replanted and natural mangrove stands in Malaysia: Langkawi Island and Sungai Merbok. Trop. Life Sci. Res. 25, 1–12.
Hutchison J., Spalding M., and Zu Ermgassen P. (2014). The role of mangroves in fisheries enhancement. Available online at: www.nature.org. (Accessed November 08, 2024).
Isroni W., Sari P. D. W., Sari L. A., Daniel K., South J., Islamy R. A., et al. (2023). Checklist of mangrove snails (Gastropoda: Mollusca) on the coast of Lamongan District, East Java, Indonesia. Biodiversitas 24, 1676–1685. doi: 10.13057/biodiv/d240341
Jimenez-R J. A. and Soto S. R. (1984). Patrones regionales en la estructura y composición florística de los manglares de la Costa Pacífica de Costa Rica. Revista Biologia Tropical.
Kathiresan K. and Bingham B. L. (2001). “Biology of Mangroves and Mangrove Ecosystems,” in Advances in Marine Biology, (Elsevier) 81–251.
Keen A. M. (1958). Sea Shells of Tropical West America: Marine Mollusks from Lower California to Colombia Vol. 39 (London: Stanford University Press), 151. doi: 10.1017/S0025315400013199
Kodikara K. A. S., Mukherjee N., Jayatissa L. P., Dahdouh-Guebas F., and Koedam N. (2017). Have mangrove restoration projects worked? An in-depth study in Sri Lanka. Restor. Ecol. 25, 705–716. doi: 10.1111/rec.12492
Lee S. Y. (2008). Mangrove macrobenthos: Assemblages, services, and linkages. J. Sea Res. 59, 16–29. doi: 10.1016/j.seares.2007.05.002
Lee S. C. and Silliman B. R. (2006). Competitive displacement of a detritivorous salt marsh snail. J. Exp. Mar. Biol. Ecol. 339, 75–85. doi: 10.1016/j.jembe.2006.07.012
Leiva Barrantes M. R. and Cerdas S. A. (2015). Distribución espacial de las especies de mangle y su asociación con los tipos de sedimentos del sustrato, en el sector estuarino del Humedal Nacional Térraba-Sierpe, Costa Rica. Revista Biologia Tropical. 63(Suppl. 1), 47–60.
Lovelock C. E., Barbier E., and Duarte C. M. (2022). Tackling the mangrove restoration challenge. PloS Biol. 20. doi: 10.1371/journal.pbio.3001836
Luo Z., Sun O. J., and Xu H. (2010). A comparison of species composition and stand structure between planted and natural mangrove forests in Shenzhen Bay, South China. J. Plant Ecol. 3, 165–174. doi: 10.1093/jpe/rtq004
Macintosh D. J., Ashton E. C., and Havanon S. (2002). Mangrove rehabilitation and intertidal biodiversity: A study in the Ranong mangrove ecosystem, Thailand. Estuar. Coast. Shelf Sci. 55, 331–345. doi: 10.1006/ecss.2001.0896
Nagelkerken I., Blaber S. J. M., Bouillon S., Green P., Haywood M., Kirton L. G., et al. (2008). The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat Bot. 89, 155–185. doi: 10.1016/j.aquabot.2007.12.007
Nakagawa S. and Schielzeth H. (2013). A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 4, 133–142. doi: 10.1111/j.2041-210x.2012.00261.x
Naranjo Loría M. (2014). Descomposición de la hojarasca de Rhizophora racemosa en el manglar de Estero Caballo y Boca Nueva Humedal Nacional Térraba-Sierpe, Costa Rica (Universidad de Costa Rica).
Nordhaus I., Hadipudjana F. A., Janssen R., and Pamungkas J. (2009). Spatio-temporal variation of macrobenthic communities in the mangrove-fringed Segara Anakan lagoon, Indonesia, affected by anthropogenic activities. Reg. Environ. Change 9, 291–313. doi: 10.1007/s10113-009-0097-5
O’Connell D. P., Fusi M., Djamaluddin R., Rajagukguk B. B., Bachmid F., Kitson J. J. N., et al. (2021). Assessing mangrove restoration practices using species-interaction networks. Restor. Ecol. 30(4), e13546. doi: 10.1111/rec.13546
Ohgaki S.-I. (1992). The malacological society of Japan NII-Electronic Library Service Distribution and Movement of the Mangrove LittoTaria (Gastropoda) on Ishigaki Island, Okinawa. malacological Soc. Japan 51, 269–278.
Ortiz L. F. and Blanco J. F. (2012). Distribución de los gasterópodos del manglar, Neritina virginea (Neritidae) y Littoraria angulifera (Littorinidae) en la Ecorregión Darién, Caribe Colombiano. Rev. Biol. Trop. (Int. J. Trop. Biol. ISSN 60, 219–232.
Pomareda E. and Zanella I. (2006). Diversidad de moluscos asociados a manglares en isla San Lucas. Rev. Cienc. Ambientales 32, 11–13. doi: 10.15359/rca.32-1.3
Proffitt C. E. and Devlin D. J. (2005). Long-term growth and succession in restored and natural mangrove forests in southwestern Florida. Wetl Ecol. Manag 13, 531–551. doi: 10.1007/s11273-004-2411-9
R Core Team (2023). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/. (Accessed February 18, 2024).
Reid D. G. (1985). Habitat and zonation patterns of Littoraria species (Gastropoda: Littorinidae) in Indo-Pacific mangrove forests. doi: 10.1111/j.1095-8312.1985.tb01551.x
Reid D. G., Dyal P., Lozouet P., Glaubrecht M., and Williams S. T. (2008). Mudwhelks and mangroves: The evolutionary history of an ecological association (Gastropoda: Potamididae). Biological Journal of the Linnean Socieo Mol. Phylogenet Evol. 47, 680–699. doi: 10.1016/j.ympev.2008.01.003
Salmo S. G., Tibbetts I., and Duke N. C. (2017). Colonization and shift of mollusc assemblages as a restoration indicator in planted mangroves in the Philippines. Biodivers Conserv. 26, 865–881. doi: 10.1007/s10531-016-1276-6
Samper-Villarreal J. and Silva-Benavides A. M. (2015). Complejidad estructural de los manglares de Playa Blanca, Escondido y Rincón de Osa, Golfo Dulce, Costa Rica. Rev. Biol. Trop. (Int. J. Trop. Biol. ISSN 63, 199–208.
Simone L. R. L. (2017). A new species of Thaisella (Neogastropoda: Muricidae) from Caribbean Guatemala, with accounts on the anatomy and taxonomy of the genus in the Western Atlantic. Arch. Molluskenkd 146, 111–120. doi: 10.1127/arch.moll/146/111-120
SINAC-UNA (2020). Protocolo PRONAMEC: Protocolo para el monitoreo ecológico de manglares en Costa Rica (San José, Costa Rica: Universidad Nacional de Costa Rica).
Skilleter G. A. and Warren S. (2000). Effects of habitat modification in mangroves on the structure of mollusc and crab assemblages. Available online at: www.elsevier.nl/locate/jembe. (Accessed February 08, 2024).
Solano J. and Villalobos R. (2005) Regiones y Subregiones Climáticas de Costa Rica (Instituto Meteorológico Nacional).
Stokes D. J., Bulmer R. H., and Lundquist C. J. (2016). Addressing the mismatch between restoration objectives and monitoring needs to support mangrove management. Ocean Coast. Manag 134, 69–78. doi: 10.1016/j.ocecoaman.2016.09.024
Sujarta P., Mailissa M. G., Keiluhu H. J., Hadisusanto S., and Yuliana S. (2022). Community distribution and utilization of gastropods in the Coastal Area of Liki Island, Sarmi District, Papua, Indonesia. Biodiversitas 23, 5001–5011. doi: 10.13057/biodiv/d231006
Thornton S. R. and Johnstone R. W. (2015). Mangrove rehabilitation in high erosion areas: Assessment using bioindicators. Estuar. Coast. Shelf Sci. 165, 176–184. doi: 10.1016/j.ecss.2015.05.013
Vargas J. A., Acuña-González J., Gómez E., and Molina J. (2015). Metals in coastal mollusks of Costa Rica. Revista Biología Tropical. 63 (Suppl. 1), 209–218. doi: 10.15517/rbt.v63i4.18143
Vargas-Zamora J. A. and Sibaja-Cordero J. A. (2011). Molluscan assemblage from a tropical intertidal estuarine sand-mud flat, Gulf of Nicoya, Pacific, Costa Rica, (1984-1987). (Revista Biologia Tropical)
Vilardy S. and Polanía J. (2002). Mollusc fauna of the mangrove root-fouling community at the Colombian archipelago of San Andrés and Old Providence. Wetl Ecol. Manag 10, 273–282. doi: 10.1023/A:1020167428154
Wibowo A. A., Basukiadi A., Nurdin E., and Nasution N. S. (2022). Benthic infaunal spatial biodiversity, coexistence, and availability for shorebird communities in the Jakarta Coastal Wetlands, Indonesia. Int. J. Bonorowo Wetlands 12(1), 23–32. doi: 10.13057/bonorowo/w120103
Yadav R., Malla P. K., Dash D., Bhoi G., Patro S., and Mohapatra A. (2019). Diversity of gastropods and bivalves in the mangrove ecosystem of Paradeep, east coast of India: a comparative study with other Indian mangrove ecosystems. Molluscan Res. 39, 325–332. doi: 10.1080/13235818.2019.1644701
Keywords: mangrove restoration, adaptive management, gastropod community assemblage, ecological indicators, Costa Rica
Citation: López-Abuchar L, Peralta-Madriz C, Soto-Navarro C, Vinueza-Hidalgo G, Whitworth A and Beirne C (2025) Temporal changes in habitat structure and gastropod community assemblage in response to active restoration of a Central American mangrove. Front. Mar. Sci. 12:1563965. doi: 10.3389/fmars.2025.1563965
Received: 20 January 2025; Accepted: 23 June 2025;
Published: 18 July 2025.
Edited by:
Heliana Teixeira, University of Aveiro, PortugalReviewed by:
Arnaldo Marín, University of Murcia, SpainXin Liao, Guangxi Mangrove Research Center, China
Copyright © 2025 López-Abuchar, Peralta-Madriz, Soto-Navarro, Vinueza-Hidalgo, Whitworth and Beirne. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Linda López-Abuchar, bGlsb2FidWNoQGdtYWlsLmNvbQ==