 Montserrat Antivero1,2
Montserrat Antivero1,2 Paz Caballero
Paz Caballero Nicolás Leppes
Nicolás Leppes Marco A. Lardies
Marco A. Lardies- 1Facultad de Ciencias Biológicas, Pontifica Universidad Católica de Chile, Santiago, Chile
- 2Departamento de Ciencias, Facultad de Artes Liberales, Universidad Adolfo Ibañez, Santiago, Chile
- 3Facultad de Ciencias del Mar y Geografía, Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile
- 4Instituto Milenio de Socio-Ecología Costera “SECOS”, Santiago, Chile
Soft-sediment habitats are crucial for marine coastal ecosystems, supporting diverse biodiversity both above and below the sediment. Ocean acidification, driven by rising CO2 and nutrient influx, enhances heterotrophic metabolism, raising CO2 levels and lowering pH. These alterations complicate the dynamics of tidal flat, emphasizing the need for further research into their impact on biodiversity. Within these ecosystems, deposit- and suspension-feeding bivalves play crucial roles. Tagelus dombeii, a bivalve mollusc found in soft sediments, exhibits burrowing behavior linked to food supply and is of significant commercial value in southern Chile. This study assessed the response capacity of T. dombeii to key stressors associated with global ocean change, such as ocean acidification and food availability. Our results revealed significant differences in pH levels between the water column and pore water from the sediment in experimental mesocosms. T. dombeii was affected by ocean acidification and food availability in terms of its morphological traits (i.e. length, width, height and growth rate), while oxygen consumption was influenced only by the interaction between acidification and food supply. Notably, heart rate remained constant but increased when food supply was low. Our study suggests that T. dombeii exhibits partial tolerance to variations in seawater pH and carbonate chemistry, possibly due to its natural exposure to acidic pore water, but it is sensitive to food availability. These plastic physiological responses suggest that T. dombeii may be less vulnerable to future global change scenarios, demonstrating potential resilience and ecological success in its natural habitat.
Introduction
The shallow seas found in coastal areas are highly valued ecosystems, ranking higher than open ocean systems, primarily because of their critical role in storing and cycling essential elements and nutrients (biogeochemical cycling) such as Carbon, CaCO3, nitrogen, and phosphorus (Bendell et al., 2014; Kessouri et al., 2021; Iram et al., 2022). Soft-sediment habitats play a vital role in the functioning of marine coastal areas (Clements and Hunt, 2017). These habitats harbor a significant amount of marine biodiversity both above (epifaunal) and below (infaunal) the sediment surface, including various deposit-feeding and suspension-feeding bivalves (Van Colen et al., 2020). Tidal flats are notable for their infaunal inhabitants, which actively burrow into the mud and act as ecosystem engineers by shaping their surrounding microenvironment (Wethey and Woodin, 2022; Salas et al., 2022). These infauna organisms encompass a diverse array of species, including molluscs, worms, and crustaceans (Jaramillo et al., 2007; Singer et al., 2023). According to Murray et al. (2019), global tidal flats have experienced a loss of 16% in their extent from 1984 to 2016, amounting to over 20,000 km2. These impacts are overlapped with global environmental drivers that are undergoing significant changes, including ocean warming (OW), ocean acidification (OA), and deoxygenation (DO).
The progressive escalating levels of atmospheric CO2 driving ocean acidification, combined with the substantial influx of nutrients into coastal regions through upwelling (Torres et al., 2011) or large river systems (Liu et al., 2021) can amplify heterotrophic metabolism. This, in turn, leads to amplified CO2 levels in the water column and exacerbates declines in pH (Carlton et al., 2023). These combined factors contribute to the intricate and ever-changing dynamics of tidal flat ecosystems, emphasizing the ongoing need for research and conservation efforts in these unique habitats. Emerging trends in ocean acidification, driven by high emissions in the 21st century, are projected to exacerbate atmospheric CO2 concentrations, which are expected to surpass 500 parts per million and result in a doubling of ocean heat uptake – levels significantly higher than those observed over the past 420,000 years, a period during which most extant marine organisms evolved (Hoegh-Guldberg et al., 2007; Gleckler et al., 2016).
Shallow-water sediments play crucial roles in the global carbonate cycle as they serve as a significant reservoir of CaCO3 (Andersson and Mackenzie, 2004). These sediments can react to the decreasing saturation state of seawater, releasing alkalinity into the overlying water column. The impact of sediment shell hash on the productivity of infaunal bivalves is still unclear and requires further investigation. The saturation state of calcium carbonate (CaCO3) in seawater is a key driver of shell formation and preservation in marine invertebrates, as it directly influences their ability to deposit calcium carbonate (Barclay et al., 2020). However, this process is also strongly affected by environmental hypercapnia, which reduce carbonate ion availability and thus lower the CaCO3 saturation state. As a result, hypercapnia can indirectly impair calcification. Indeed, long-term exposure to high pCO2 has been associated with reduced growth and/or calcification rates in several marine taxa, including mussels (Michaelidis et al., 2005; Navarro et al., 2013; Leung et al., 2022), echinoderms (Asnicar and Marin, 2022), and coral species (Hoegh-Guldberg et al., 2007; Leung et al., 2022; Rathbone et al., 2022). Species with greater tolerance to such conditions often dis-play high metabolic rates, mobility, and activity levels (Medeiros and Souza, 2023). Overall, ocean acidification has been recognized as a significant threat to marine mollusks, including infaunal bivalves (Gazeau et al., 2013; Martel et al., 2022). However, these effects are often described predominantly within the context of water column acidification, overlooking or with comparatively fewer attention on the drastically different pH and carbonate system conditions that infaunal species experience below the sediment-water interface. Consequently, it is often assumed that infaunal organisms are more resilient to ocean acidification (Widdicombe et al., 2011), and several studies support their apparent tolerance to changes in seawater pH and carbonate chemistry (Vlaminck et al., 2023). However, other research has shown that sediment acidification can negatively affect infaunal organisms, particularly marine bivalves (Hu et al., 2014; Clements et al., 2016; Martel et al., 2022; Vlaminck et al., 2023).
Food supply is a factor that could potentially lead to detrimental effects on marine invertebrates, in addition to other global change effects such as ocean acidification (Lawrence et al., 2015; Flombaum and Martiny, 2021; Kwon et al., 2022). Food supply for bivalves depends on marine phytoplankton which contributes roughly 50% of global net primary production (Field, 1998), while for deposit feeders bivalves obtain their food from organic particle settling from water column, biodeposits and/or detritus (Navarro et al., 2008). A recent analysis of a 26-year time series of spatially averaged monthly mean chlorophyll-a (Chl-a) concentrations in the Southern Pacific Ocean revealed that the largest deviations in Chl-a were closely associated with El Niño–Southern Oscillation (ENSO) events, as evidenced by the strong correspondence between Chl-a anomalies and the Multivariate ENSO Index (Johnson and Lumpkin, 2024). These fluctuations underscore the central role of ENSO-driven variability in modulating nutrient availability, both through altered oceanographic circulation patterns and the disruption of coastal upwelling systems, which are otherwise critical sources of nutrient input sustaining primary productivity in this region. Furthermore, in the tidal flats, the intensified urbanization of coastal habitats has led to a significant degradation of food-web complexity and ecosystem services due to multiple stressors (Eriksson et al., 2010; Christianen et al., 2017). While the combined effect of both factors (OA and food supply) is not widely discussed in the literature, some studies have examined this effect in mollusks. Studies on the effect on oyster larvae (see Hettinger et al., 2013) have shown that a combination of high pCO2 levels and low food supply results in reduced physiological performance. However, this effect occurs independently for each factor and is not additive. It is of vital importance to mention that food supply plays a crucial role in determining an organism’s response to stressful situations, with significant implications for shell-forming mollusks (Ramajo et al., 2016a, 2019; Harayashiki et al., 2020). Therefore, we hypothesized that increased food supply would mitigate the negative effects of acidification in infaunal bivalves.
The tidal flats of southern Chile are characterized by the presence of numerous species of bivalve molluscs, many of which are of commercial importance (Clasing et al., 1994; Navarro et al., 2008). Several species of bivalves coexist at the lower intertidal of large tidal flats located in the enclosed or inland coast of the northern area of the North-Patagonic archipelagos on the Chilean coast (ca. 40–42°S): Tagelus dombeii (Lamarck), Tawera gayi, Ameghinomya antiqua, Semele solida, Gari solida, Mytilus chilensis, and Zemysina insconspicua (see Stead et al., 2002; Jaramillo et al., 2007). In general, studies have shown that sediments with higher densities of bivalves, particularly those where deep burrowers are most abundant, exhibited greater species richness and higher densities of macroinfauna (Stead et al., 2002; Jaramillo et al., 2007). The majority of eulamellibranch bivalves (i.e T. dombeii) engage in suspension feeding while deposit-feeding behavior has only been observed in the Tellinacea and Lucinacea species (Morse and Zardus, 1997; Lardies et al., 2001; Navarro et al., 2008).
In this study, we investigated the effects of ocean acidification and food supply on the morphological traits and physiological performance of the razor clam T. dombeii (Lamarck, 1818), which coexists with the bivalve community in the sandflats of Coihuín, Reloncaví Sound (41°29’S, 72°54’W).
Material and methods
Sampling of organisms and mesocosm setup
During spring, 100 juvenile individuals of Tagelus dombeii (Lamarck, 1818) (Tellinacea: Solecurtidae) with a mean of 18 mm in length, 7 mm in width, and 3 mm in height, which is below the reported size at sexual maturity (40–50 mm in length; Lépez et al., 1997; Sánchez et al., 2003), were collected from the soft sediments of the Coihuín tidal flat (41°29’ S, 72°54’ W), located 8 km southeast of Puerto Montt, Los Lagos Region, Chile (see Supplementary material Figures 1). Sea surface temperature follows a seasonal pattern at Coihuín, with the lowest values occurring during the winter months (8°C), increasing towards the summer maximum in January (17°C; Urrutia et al., 2001; Lardies et al., 2001). Near the sampling site, the spectral exponent (β) of seawater chemical parameters is -1.703 for temperature, -1.584 for salinity and -1.206 for pH indicating high unpredictability dominated by random short-term environmental fluctuations compared with near locations (see Castillo et al., 2024 for details). In contrast, the porewater at a depth of 5 cm in the sediment of the Coihuin tidal flat, during November, has a temperature of 13.19°C, pH of 7.72, and salinity of 30.66 (Lardies et al., unpublished data). The intertidal sediment of Coihuín tidal flat is predominantly sandy (93.81%), and the content of mud (particles < 63 mm diameter) is low in Coihuín, with an annual average of 1.83% (Lardies et al., 2001). Observed organic matter content is low (0.68%), and in the biogenic aggregates (i.e. detritus, phytoplankton, and bacteria) that represented an 0.58% (Lardies et al., 2001). The individuals were transported under humid conditions to the laboratory. No mortalities were recorded during transport. In the laboratory, individuals were acclimated for three days in a “common garden” aquarium containing filtered natural seawater at 12°C (32 ppm salinity, 12D:12N photoperiod), with daily feeding of microalgae, to minimize stress responses associated with transportation. Due to their small size and delicate shells, individuals were not marked to avoid causing harm or shell damage to the animals.
For the mesocosm experiment, random groups of 4 individuals were created and assigned to each 9-liter aquariums for an experimental trial of 67 days. Each aquarium was filled with 6 cm of commercial quartz sand (https://dondecapo.cl/project/cuarzo-32/) across the entire bottom and filled to its total capacity with filtered and UV-sterilized natural seawater (see Martel et al., 2022). It is necessary to use inert material to avoid potential effects of sediment altering the chemical properties of the water, such as pH or alkalinity (Brenner et al., 2016). Water and pore water were measured prior to the experiment for all the 16 aquariums to ensure that the system was within the desire environment condition values.
The assigned treatments included levels of acidification and food supply, simulating CO2 concentration scenarios with a current scenario of 500 µatm and a future scenario of 1500 µatm (pH = 8.0 and 7.5, respectively; described in Navarro et al., 2013; Vargas et al., 2017). The food source used was microalgae (see below) and was assigned as either optimal or restricted food, relative to the individual’s dry weight. As reported by Navarro et al. (2008), the optimal food level corresponds to 4% of individual’s dry weight, while the restricted food level corresponds to 1%. Each aquarium was assigned to one of the 4 treatments, that are: 500 μatm – 1% food supply, 500 μatm – 4% food supply, 1500 μatm – 1% food supply and 1500 μatm – 4% food supply.
During the experimental period, razor clams were fed daily following this relationship with their dry weight (1% and 4%), using Reef Blizzard-O suspension (Brightwell® Aquatics) diluted in 10 mL of filtered and UV-sterilized seawater. All aquariums were monitored daily for mortality without replacement in case of death, reconsidering the food amount per aquarium. Water in all aquariums was renewed twice per week to maintain appropriate conditions. At the end of the experiment, the morphological and physiological measurement were taken from the individuals that survive and then were sacrificed.
Additionally, one aquarium with identical characteristics was subjected to the CO2 exposure for each treatment. In each aquarium, three complete shells were placed on the sand surface, while three additional complete shells were buried approximately 3 cm deep in the sand. The buoyant weight, length, width, and height were measured, and at the end of the experiment, the same parameters were measured for each shell. This was done to estimate the rate of dissolution in the empty shells, which were not affected by biological activity. The buoyant weight (from now on referred as growth rate) served as an indicator of calcification (or decalcification; Lagos et al., 2016). During the experiment, additional individuals collected from the same site were used to analyze the relationship between shell length and dry tissue weight, as well as between oxygen uptake and dry tissue weight, using linear regression.
Seawater pCO2 levels
The pH treatment levels (8.0 and 7.5) were achieved by adding a mixture of dry air with pure CO2 (partial pressure of CO2 [pCO2] = 500 and 1500 μatm, respectively) to each aquarium with seawater in a regular continuous flow using mass flow controllers (Aalborg Instruments & Controls, Inc., Orangeburg, NY, USA; for details see Benítez et al., 2018). The pH of the water (NBS scale) was measured by extracting 60 mL of water from the aquariums with disposable syringes for each aquarium, and then measuring the pH twice with the same water sample. The sediment pH was measured using a 10 mL micropipette (Labnet BioPette Plus), extracting pore water by inserting the micropipette tip around 2–3 cm into the sediment, following the sampling method of Bendell et al. (2014). Water was extracted three times from the same aquarium, transferred to Falcon tubes, and pH was measured twice with each sample. All samples were measured using a pH meter (Mobile 826, Metrohm, Herisau, Switzerland), which was connected to a combined electrode (junction-type). During measurements, temperature was also recorded using a digital thermometer, and seawater salinity was measured using a portable salinometer (Salt6+, Oakton; precision: ± 0.1 PSU and ±0.5°C, respectively). Alkalinity samples were collected twice a week, ensuring that all aquaria were sampled in both the water column and pore water during each sampling week. The samples were fixed with a saturated solution of HgCl2 and stored in 50 mL Falcon bottles in the laboratory under dark and ambient temperature conditions. At the end of the experiment, alkalinity was analyzed using a multiparameter photometer (Hanna HI83303) in the seawater alkalinity mode. For this, 10 mL of aquarium sample was used as a blank, and then 1 mL of reagent (HI755) was added, based on the colorimetric method. The results were expressed in mg/L and subsequently converted to µmol/kgSW to assess the carbonate system.
Temperature, salinity, pH, and alkalinity measurements were used to calculate the carbonate system parameters. These included the partial pressure of CO2 (pCO2) and the saturation states (Ω) of calcite and aragonite, which were estimated using the CO2SYS software in Excel (Pierrot et al., 2006) and the dissociation constants from Mehrbach et al. (1973), refitted by Dickson and Millero (1987), for both the water column and the sediment.
Measurements of biological traits
The measurement of growth rates and morphological changes in individuals during the experiment was obtained by measuring the buoyant weight, length, width, and height of each experimental individual. Buoyant weight corresponds to the weight of the individual in water and is used as a non-invasive estimator of calcification rate, i.e., the growth of individuals (Palmer, 1982). It was measured at the beginning and end of the experiment using an analytical balance (± 0.1 mg, AUX 220, Shimadzu, Kyoto, Japan). For measurements of length, width, and height, an electronic caliper Mitutoyo (Sakado, Japan) was used at the beginning and end of the experiment. Empty shells (without soft tissues) from aquariums under CO2 treatments were measured for buoyant weight, without biological activity, using the same analytical balance used for measuring the buoyant weight of individuals, while for length, width, and height were measured using the electronic caliper.
At the end of the 67 days, the metabolic rate and heart rate were measured to obtain the energy expenditure value of the individuals. Metabolic rate measurements were taken before daily feeding, using 0.067 L glass respirometry chambers with a PreSens Mini Oxy-4 respirometer (PreSens GmbH, Regensburg, Germany). To measure oxygen consumption (mgO2 L-1 h-1), dissolved oxygen in the chambers was quantified every 15 seconds for approximately 1 hour. Finally, the obtained measurement was standardized per gram of weight to obtain the specific metabolic rate of each individual (see Lardies et al., 2021). Background measurements (chambers without animals) were made at the same experimental conditions to quantify microbial oxygen consumption to be subtracted from each experimental measurement. No reduction over 3% of oxygen consumption was recorded (see Osores et al., 2017). Previously, the sensors were calibrated in anoxic water, using a Na2SO3 solution for 0% oxygen and water saturation with air bubbles for 100% oxygen. The obtained measurement was standardized per gram of weight. Heart rate measurements were taken following the methodology of Gaitán-Espitia et al. (2014) and Rodríguez-Romero et al. (2022) using as a proxy of cardiac activity at constant temperature of 12°C for all measurements. Individuals were immobilized with adhesive tape on a plate. One measurement per individual was taken in batches of 4 individuals, each in an individual chamber installed in the temperature-controlled bath (± 0.5°C, LWB-122D, LAB TECH). The protocol included 5 minutes of initial acclimation and 15 minutes of measurement. An AMP 03 heartbeat amplifier (Newshift Lda®) was used, connected to an oscilloscope. Measurements were taken for all the individuals, and the results were expressed in beats per minute.
Statistical analysis
To avoid pseudo-replication errors, variables for each individual were averaged by aquaria. First, homogeneity and normality tests were performed for the data, including Levene and Kolmogorov-Smirnov tests, respectively. To assess the difference in pH levels of both water and sediment water (pore water), a repeated ANOVA measure was performed, considering the variable place (source of water) as a within-subjects factor (within the same aquaria) and sampling time was treated as between-subjects factor.
A Two-Way ANOVA was conducted to assess the effect of CO2 levels, food availability, and the interaction between both factors on morphological traits (length, height, width and growth rates), dissolution rates, oxygen uptake, and heart rate. This determined whether the factors separately or their interaction had significant effects on the individuals. Tukey’s HSD was used as a posteriori test when the main factors indicated significant differences between levels of the corresponding factor (Underwood, 1997). Additionally, the effect size of the treatments on both physiological and morphological variables was calculated using the log response ratio (lnRR), defined as lnRR = treatment/control. This approach allowed for a direct comparison of treatment effects relative to the controls. Bootstrapping was used to estimate the 95% confidence intervals of the lnRR values. All the analyses were conducted in R statistical environment (R Core Team, 2024).
Results
All environmental variables remained stable in all of the treatments throughout the experimental period, with low variations in pH, salinity, and temperature (see Table 1). Based on the analysis of characterization of the population, the relationship between length and dry tissue weight of the individuals showed an R2 value of 0.91, while the relationship between oxygen uptake and dry tissue weight showed an R2 of 0.57 (see Supplementary Figures S2A, B).
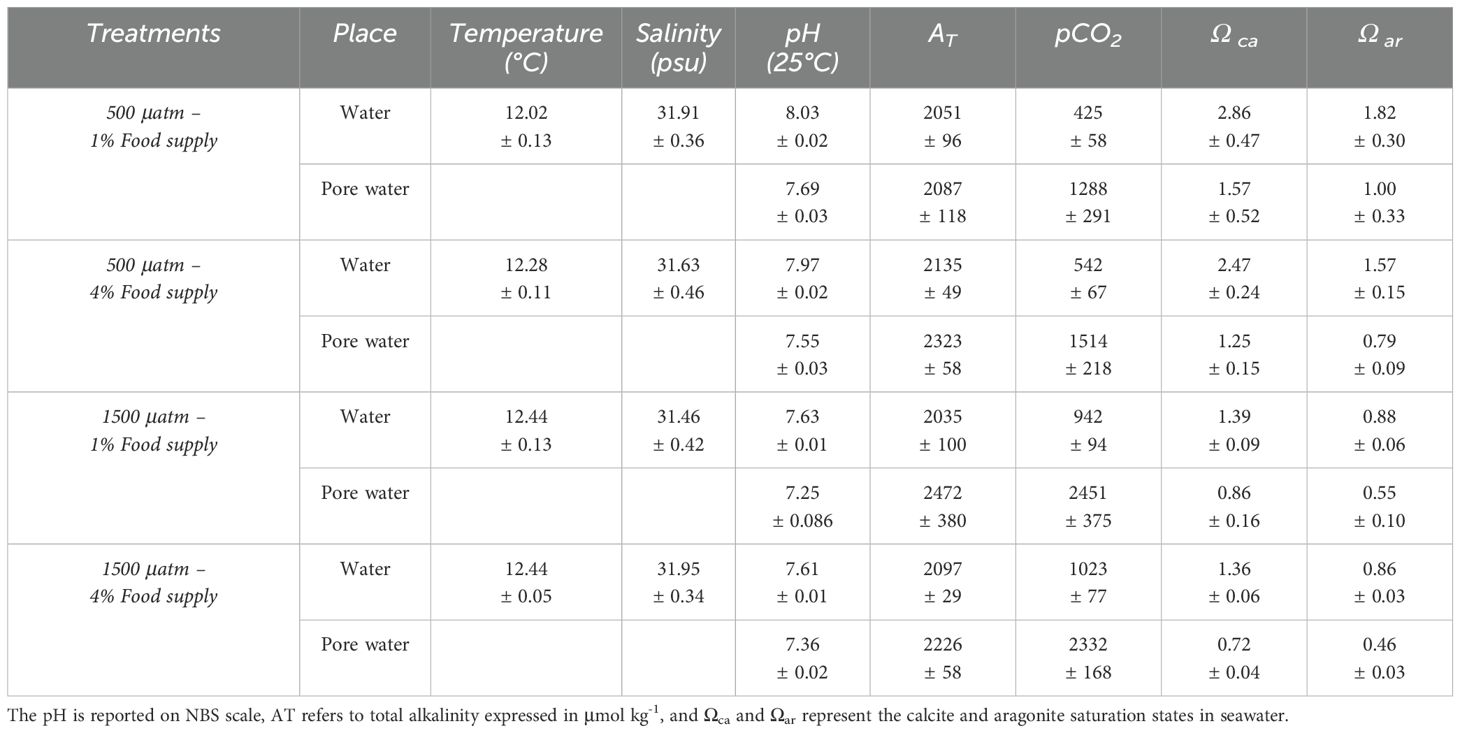
Table 1. Mean values (± SE) of carbonate system parameters for each experimental treatment, including pCO2 levels (500 and 1500 μatm) and food supply (1 and 4% of tissue dry weight), among the two sampling sites within the aquaria (water and pore water).
A significant difference in pH levels was observed between the water column and the pore water (P < 0.01, repeated measures ANOVA, see Supplementary Table S1, Figure S3), but no differences were found based on the sampling time. On average, the pH of the water column was 7.81 ± 0.017 SE, while the pH of the pore water was 7.54 ± 0.018 SE (see Supplementary Figures S3, S4). Particularly, a difference in pH was observed between the overlying water and the sediment, which ranged between 0.28 and 0.57 units under CO2 levels of 500 μatm and between 0.16 and 0.34 units under CO2 levels of 1500 μatm.
Mortality was assessed after the experimental trial. Under the 500 µatm CO2 treatment, individuals exposed to a 4% food supply exhibited a mortality rate of 38.5%, whereas those maintained at a lower food level (1%) showed reduced mortality (14.3%). In contrast, at 1500 µatm CO2, mortality remained at 25.0% under the 4% food condition but increased markedly to 54.0% under the 1% food supply, highlighting an interactive effect between food availability and CO2 levels on survival.
Morphological traits
The interaction between pCO2 and food supply affected the individuals’ morphological characteristics (i.e. length, width, height, growth rate) (Figure 1; Table 2). High food supply in low pH conditions increased total width, with a marginally significant effect in the interaction of the treatments (Figure 1B; Two-Way ANOVA; F1,10 = 4.90, P = 0.051), while for low food supply in low pH conditions there was a decreased in total height, with a marginally significant effect in the interaction of the treatments (Figure 1C; Two-Way ANOVA; F1,10 = 4.52, P= 0.059). Furthermore, pH conditions and food supply affected the total increase length, and growth rate, but this effect was not significant (but see size effect, Supplementary Figure S6). Moreover, shell dissolution recorded for empty shells (see Figure 2A) was higher exposed to the water column and the interaction between the treatments pCO2 and location in the sediment resulted in a significant effect (Two-Way ANOVA; F1,4 = 7.81, P = 0.049). Although shell carbonate content tended to be higher in organisms from both CO2 treatments under 4% food supply, the difference was not statistically significant, being higher in the pCO2 treatment (see size effect, Supplementary Figure S7 and Figure 2C).
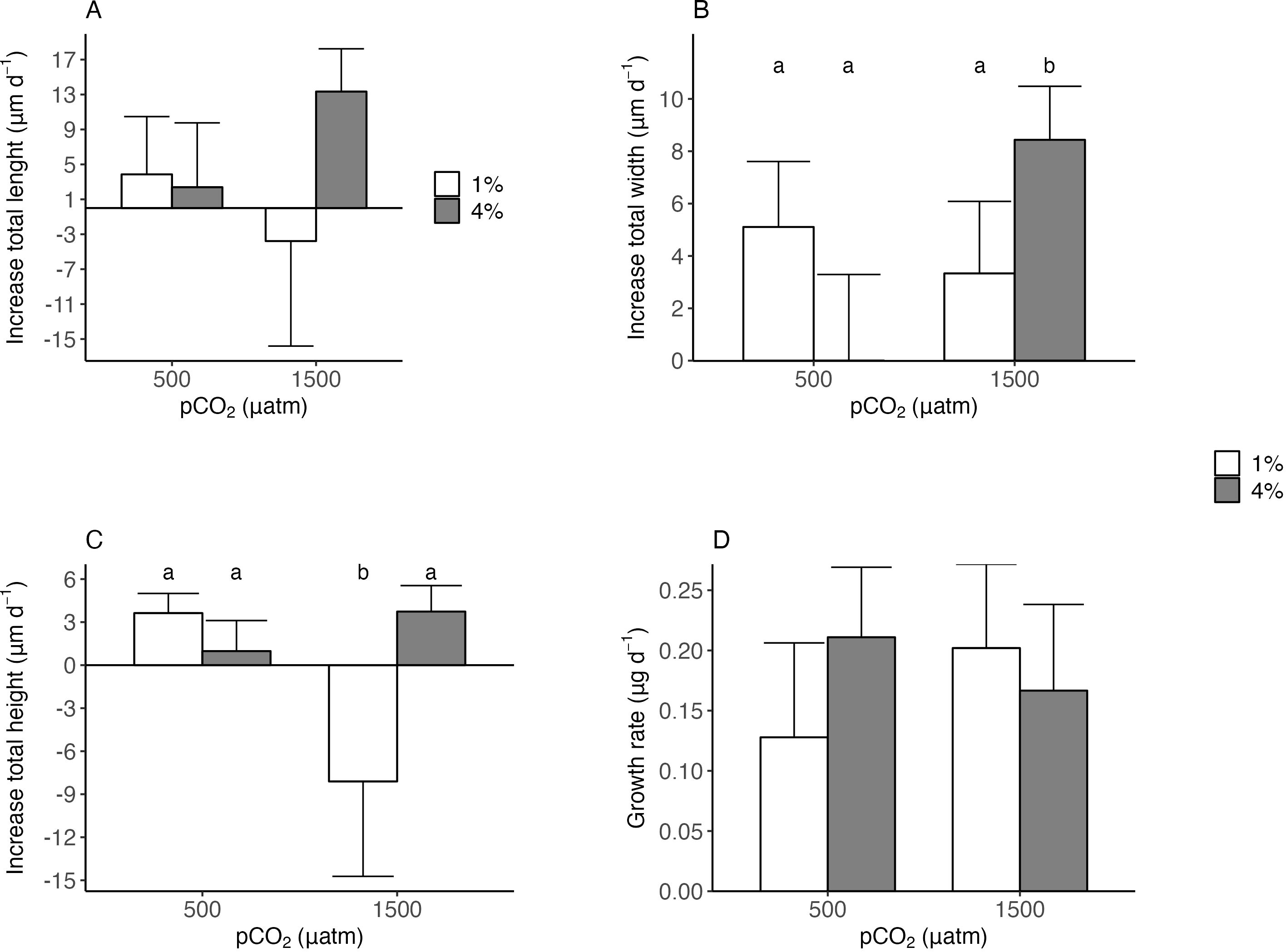
Figure 1. Tagelus dombeii: morphological changes with increase of total length (A), increase of total width (B), increase of total height (C), and growth rate (D) in each experimental condition. The treatments were conditions of pH (500 µatm and 1500 µatm) and food supply (1% and 4% dry weight). The letters in lowercase indicate and mark the significance.
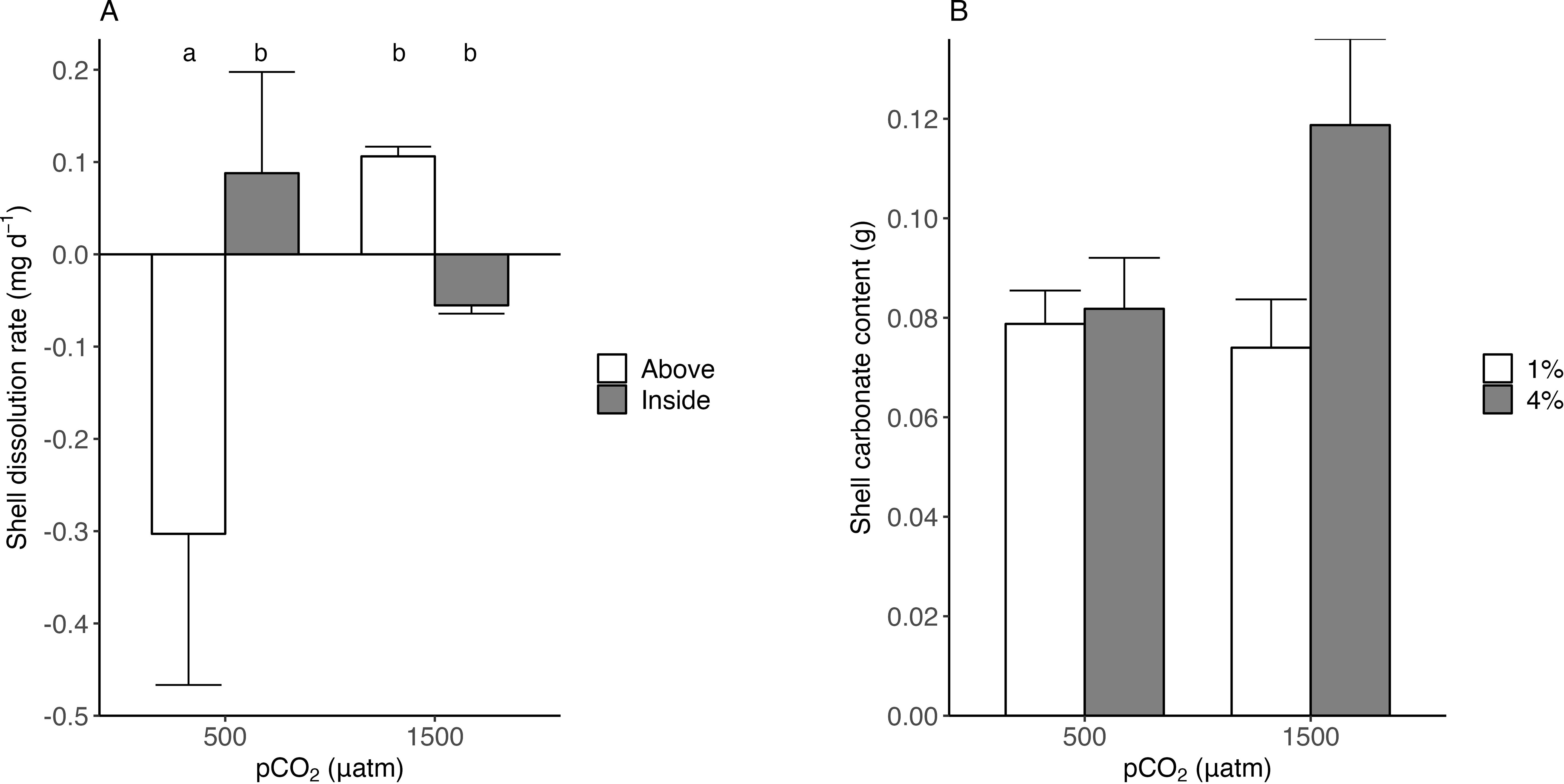
Figure 2. Shell dissolution rate (A), measured above and below the sediment, and shell carbonate content (B) in empty shells of Tagelus dombeii after 67 days of exposure to experimental conditions. The treatments in panel B correspond to pH levels (500 µatm and 1500 µatm) combined with food supply (1% and 4% dry weight). The letters in lowercase indicate and mark the significance.
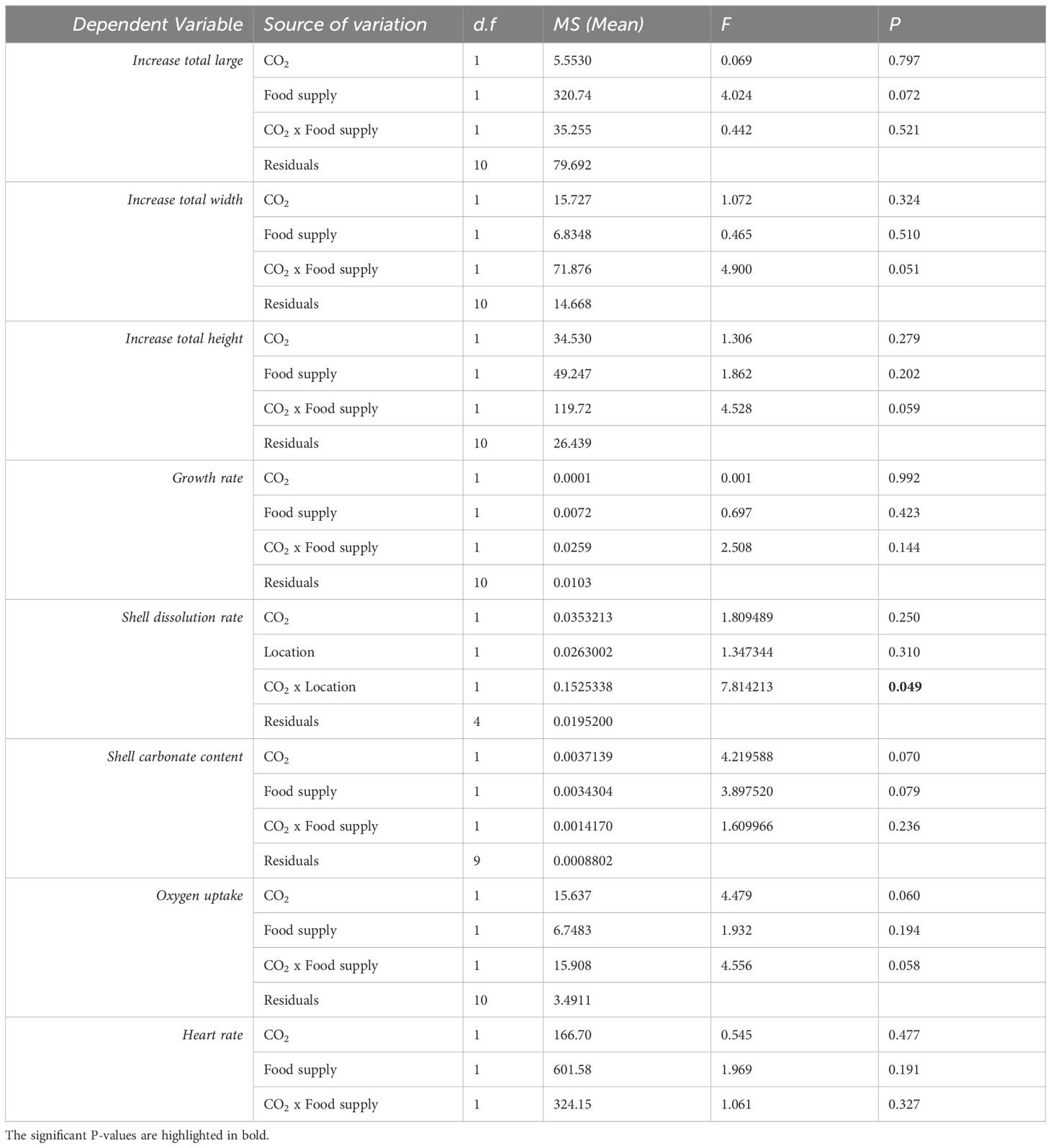
Table 2. Summary of Two-way ANOVA for the effects of pH conditions, food supply, and interaction among both, upon morphological physiological variables (large, width, height, and growth rate).
Physiological traits
The food supply and pCO2 treatment impacted the physiological variables of the razor clams (Table 2). A non significant effect was observed on the oxygen uptake rate, which decreased when the organisms were exposed to low pH (Two-Way ANOVA; F1,10 = 4.47, P = 0.06), and the interaction between CO2 and food supply (Two-Way ANOVA; F1,10 = 4.556, P = 0.058; Figure 3; but see size effect Supplementary Figure S8.). The interaction between the treatments food supply and pCO2 did not influence heart rate (Figure 4). However, the individuals tended to increase their heart rate in conditions of low food supply (see Figure 4).

Figure 3. Tagelus dombeii: metabolic rate as oxygen uptake (mgO2h-1g-1) for razor clams in each experimental condition. The treatments were conditions of pH (500 µatm and 1500 µatm) and food supply (1% and 4% dry weight). The letters in lowercase indicate and mark the significance.
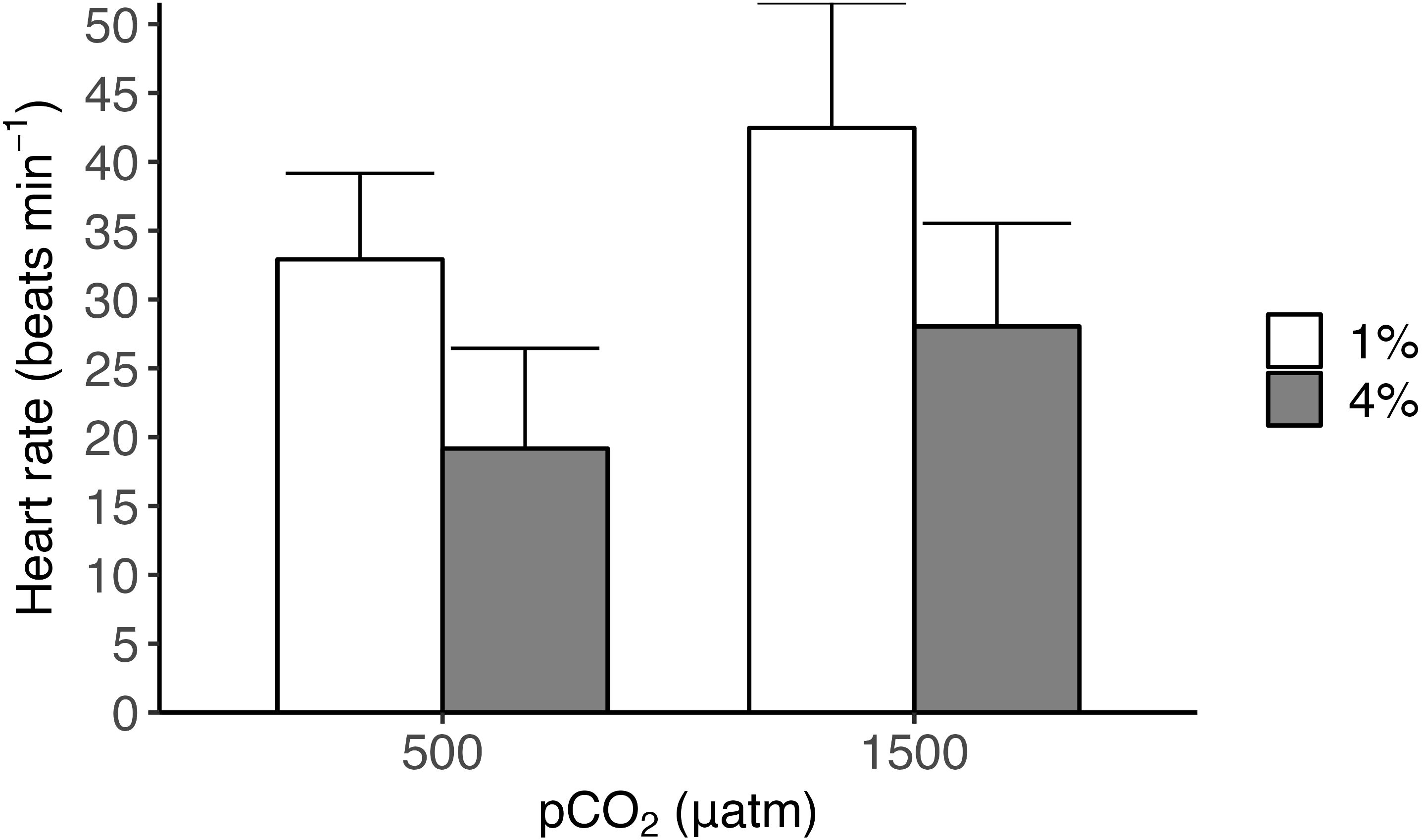
Figure 4. Tagelus dombeii: heart rate as beats min-1 for razor clams in each experimental condition in a mesocosm for 67 days. The treatments for B were conditions of pH (500 µatm and 1500 µatm) and food supply (1% and 4% dry weight).
Discussion
The ongoing rise in CO2 emissions is accelerating both climate change and ocean acidification, significantly impacting various marine invertebrates (Shi and Li, 2024). However, the magnitude and direction of these effects vary according to the specific characteristics of each species (Goethel et al., 2017; Vargas et al., 2017; Harvey et al., 2013). In this context, infaunal bivalves could show greater resilience to these disturbances, attributable, among other factors, to their exposure to pore water, which has an intrinsically lower pH compared to the water column. Our results indicate that T. dombeii possesses adaptive mechanisms that allow it to cope with environmental stress, such as ocean acidification, but not for limited food supply. These mechanisms include morphological modifications and physiological trade-offs that optimize its ability to survive under unfavorable conditions.
The characteristics of pore water are strongly influenced by the dynamics of the overlying water (Zhang et al., 2013; Precht et al., 2004). Various processes, such as the decomposition of organic matter, the sedimentation of materials of terrestrial origin, and the biological activity of both microorganisms and infaunal organisms, can generate a decrease in the pH of sediments (Hohaia et al., 2014) in addition to other processes that can contribute to pH reduction in sediments, such as sulfide oxidation or metal presence (see Bonner et al, 1990). This localized acidification of the benthic environment could have significant implications for key processes in bivalves, such as settlement, recruitment, and survival (Meseck et al., 2018; Clements and Hunt, 2018; Cummings et al., 2009). The pH of pore water tends to be more acidic than that of surface water, a condition attributed to the processes previously described (Hu et al., 2014; Cummings et al., 2009). The observed pH gradient between the overlying water and the sediment highlights the potential for differential buffering capacities at the sediment–water interface. These results are consistent with those reported by McGarrigle and Hunt (2024), who, in controlled experiments, documented that the pH of the water under constant acidification conditions was 7.73 ± 0.13, while that of the sediment was 7.29 ± 0.19, evidencing an average difference of 0.44 units. Such micro-scale heterogeneity is ecologically significant, as it may influence the exposure of benthic organisms to acidified conditions and thus mediate their physiological responses. Although ocean acidification has been a central research topic in recent decades, the carbonate system in sediments has yet to be fully explored. Our results reveal, in addition to a difference in pH between the water column and pore water, variations in the carbonate systems of both environments. The sediments exhibited more acidic conditions, with higher partial pressure of CO2 (pCO2), accompanied by a decrease in the saturation states of calcite and aragonite. Under these undersaturated conditions (Ωca < 1 and Ωar < 1; see Table 1), such as the pore water, the shell’s mineral phase is likely to undergo dissolution, leading to higher energy demands for maintaining its structure and function. In our case, the findings indicate a higher carbonate content in individuals exposed to acidified conditions with 4% food supply, despite the low saturation states of calcite (Ωca = 0.72) and aragonite (Ωar = 0.46). This suggests an increased investment in shell production under ocean acidification, potentially as a compensatory response when food availability is optimal. Such a response may help maintain shell integrity despite the thermodynamically unfavorable conditions for calcification. However, sustained low aragonite/calcite saturation states in sediments may have long-term consequences for bivalve shell integrity and calcification capacity. While significant short-term effects were detected, chronic exposure to undersaturated conditions could progressively impair shell formation.
The outcomes revealed variable morphological effects, highlighting a significant impact on the increase in total height and width, attributed to the interaction between both treatments. However, measurements related to the growth rate in terms of increase in total length, calcification (change in growth rate), and shell carbonate content did not show significant differences. These findings coincide with those reported by Liang et al. (2022), who observed that individuals of the infaunal bivalve Sinonovacula constricta subjected to controlled pH conditions (pH = 8.1) and projected future scenarios (pH = 7.7) did not show significant changes in their growth performance. The fact that the treatments have generated mostly non-significant morphological effects suggests that this species could possess some resistance to acidification. Previous studies have indicated that infaunal organisms, which are regularly exposed to acidified conditions in their natural habitats, such as in our case, where individuals originate from an environment with a pH of 7.7, may exhibit greater tolerance and experience fewer morphological effects under acidification scenarios (Liang et al., 2022; Hu et al., 2014). On the other hand, McGarrigle and Hunt (2024) propose that, during periods of stress, such as exposure to constant elevated levels of CO2, organisms tend to prioritize survival overgrowth, strategically redistributing the available energy. Additionally, it has been suggested that calcifying organisms could overcome the energetic limitations associated with ocean acidification as long as they have sufficient amounts of food (Clements and Darrow, 2018). In this context, food supply could play a key role in physiologically compensating for the stress induced by acidification conditions (Goethel et al., 2017; Ramajo et al., 2016a). Furthermore, individuals of T. dombeii show a dual feeding behavior, using both suspension feeding and deposit feeding strategies (Lardies et al., 2001). This species can alternate between these strategies according to the conditions, obtaining food from both the water column (suspension) and the sediment surface (detritus; Navarro et al., 2008). Additionally, individuals tend to employ suspension feeding when they are at greater depths in the sediment, while they opt for deposit feeding when they are closer to the surface (Lardies et al., 2001; Navarro et al., 2008) thus increasing the likelihood of sustaining feeding activity and coping with adverse conditions, such as environmental stressors like OA and limited food supply.
The results indicate a significant decrease in oxygen consumption under acidification conditions, suggesting metabolic depression in organisms exposed to these conditions. Metabolic depression is a strategy adopted by organisms to extend short-term survival in adverse environments (Ramajo et al., 2016b; Hu et al., 2014). Although this metabolic depression may represent an acute response to environmental stressors, its persistence and long-term consequences remain uncertain. Prolonged exposure to low pH conditions could either lead to physiological acclimation or, conversely, to cumulative energetic deficits that compromise growth, reproduction, or survival. Further studies with longer exposure periods are needed to determine whether this response is transient or maintained over time, and whether it translates into reduced fitness or population-level impacts.
Previous studies have documented this response in various species, including the infaunal brittlestar (Hu et al., 2014), scallops (Ramajo et al., 2016b), and infaunal clams (Martel et al., 2022). Food supply has been shown to enhance resilience to ocean acidification (Ramajo et al., 2016a), as organisms with access to adequate food exhibit higher growth, metabolism, calcification, and ingestion rates compared to those with limited food supply, both under control conditions and acidification scenarios (Navarro et al., 2016). In line with these findings, our study shows that organisms exposed to high food availability and low pH exhibit greater survival than those under low food availability and low pH conditions (see Supplementary Figure S5). However, food availability alone does not always guarantee effective intake and assimilation. Under acidified conditions, metabolic depression can impair feeding and nutrient processing, thereby limiting the benefits of increased food supply. This decoupling between external resource availability and internal energy acquisition has been observed in other bivalves and may compromise energy allocation to growth and reproduction (Ramajo et al., 2016b; Clements and Darrow, 2018).
Heart rate (HR) in mollusks is a fundamental physiological indicator that reflects their health status and their ability to adapt to different environmental conditions (Davis et al., 2023; Fernández et al., 2024). This parameter responds to a variety of stress factors, showing notable plasticity in populations inhabiting variable environments. In the present study, Tagelus dombeii showed a higher (but not significant) heart rate with lower food supply, would indicate an increase in stress experienced by the organism (Davis et al., 2023). Under extreme conditions, the relationship between HR and metabolic rate may decouple (Marshall and McQuaid, 2020); mollusks can maintain a constant HR while decreasing their metabolic rate, adopting alternative strategies to ensure their survival (Marshall and McQuaid, 2020). Consistent with these findings, our results show that T. dombeii exhibited a reduced oxygen consumption rate under low pH conditions, while heart rate increased when food supply was limited. This suggests a decoupling between HR and metabolic rate, likely reflecting an adaptive response to the combined stressors of ocean acidification and food supply. Likewise, ocean acidification can induce metabolic depression, evidenced by a reduction in metabolism, which constitutes an energy-saving strategy against the stress associated with low pH, regardless of changes in heart rate (Martel et al., 2022).
Our results indicate an effect of the interaction between pH and food supply on oxygen consumption but not on food supply. This finding could be explained by the ability of T. dombeii to employ both types of feeding, which allows it to obtain nutrients from two different sources, thus improving its efficiency in food acquisition (Navarro et al., 2008). Although this study did not quantify the time spent on each type of feeding, we suggest that these organisms could alternate between suspension feeding and deposit feeding to minimize the ingestion of low-pH water under acidification conditions (see Vlaminck et al., 2023). This feeding flexibility allows infaunal filter-feeding bivalves to mitigate the effects of acidification, maintaining acid-base balance and avoiding physiological alterations (see Vlaminck et al., 2023), which reinforces the resilience and robustness of these organisms, in line with the results obtained in M. calcarea (Goethel et al., 2017).
The variability in responses to adverse conditions of marine invertebrates depends on the habitat of origin, the scales of environmental variability, and the simultaneous presence of other stress factors (Lefevre, 2016; Castillo et al., 2024; Gaitán-Espitia et al., 2017). These elements influence the physiological responses of organisms to cope with unfavorable conditions. In this study, the razor clams analyzed come from Coihuín, a site near the Reloncaví fjord, which is characterized by high variability in pCO2 and food supply (Vergara-Jara et al., 2019; Lardies et al., 2001; Castillo et al., 2024). This suggests that infaunal organisms in the tidal flat would show greater signs of phenotypic plasticity, given the wide environmental variability of this quasi-estuarine system (see Osores et al., 2017; Castillo et al., 2024).
Global change, particularly ocean acidification, poses significant threats to marine species and fisheries, including T dombeii. The species metabolic depression in response to acidification could reduce its harvestable biomass, thereby affecting the maximum sustainable yield and threatening economic stability in affected regions. The previous plus pressure of artisanal fisheries on T. dombeii, with an annual harvest of 2,203 Mt (100 Mt from the Los Lagos Region), face potential losses. For example, between 1996 and 2007, the selectivity for T. dombeii dropped from a mean size of 76 mm to 54 mm (legal commercial size 65 mm), indicating a shift toward smaller harvestable individuals in the Bio-Bio region (Hernández et al., 2011). This trend could lead to significant economic losses, underlining the need for adaptive fisheries management that accounts for both climate stressors and ecological dynamics.
Tagelus dombeii demonstrates partial resilience to ocean acidification due to its natural exposure to acidic pore water and flexible feeding strategies. This adaptability allows it to optimize nutrient acquisition and maintain physiological balance under stressful conditions. However, its survival and performance are compromised under low food supply, highlighting the importance of trophic conditions. These findings highlight the complexity of infaunal marine invertebrate responses to global change, emphasizing the need to consider multiple stressors in future projections and fisheries.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
MA: Formal Analysis, Investigation, Writing – original draft, Writing – review & editing, Data curation, Methodology, Visualization. NL: Data curation, Formal Analysis, Investigation, Visualization, Writing – original draft, Writing – review & editing, Software. PC: Conceptualization, Data curation, Investigation, Methodology, Writing – original draft. ML: Conceptualization, Formal Analysis, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded by ANID FONDECYT N° 1240367. ML acknowledges the ANID-Millennium Science Initiative Program-Code ICN2019_015 and also thanks the support from PIA ANID ANILLOS ACT240004.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1568035/full#supplementary-material
References
Andersson A. J. and Mackenzie F. T. (2004). Shallow-water oceans: a source or sink of atmospheric CO2? Front. Ecol. Environ. 2, 348–353. doi: 10.1890/1540-9295(2004)002[0348:SOASOS]2.0.CO;2
Asnicar D. and Marin M. G. (2022). Effects of seawater acidification on echinoid adult stage: A review. J. Mar. Sci. Eng. 10, 477. doi: 10.3390/jmse10040477
Barclay K. M., Gingras M. K., Packer S. T., and Leighton L. R. (2020). The role of gastropod shell composition and microstructure in resisting dissolution caused by ocean acidification. Mar. Environ. Res. 162, 105105. doi: 10.1016/j.marenvres.2020.105105
Bendell L. I., Chan K., Crevecoeur S., and Prigent C. (2014). Changes in ammonium and pH within intertidal sediments in relation to temperature and the occurrence of nonindigenous bivalves. Open J. Mar. Sci. 4, 151–162. doi: 10.4236/ojms.2014.43015
Benítez S., Lagos N. A., Osores S., Opitz T., Duarte C., Navarro J. M., et al. (2018). High pCO2 levels affect metabolic rate, but not feeding behavior and fitness, of farmed giant mussel Choromytilus chorus. Aquac. Environ. Interac. 10, 267–278. doi: 10.3354/aei00271
Brenner H., Braeckman U., Le Guitton M., and Meysman F. J. R. (2016). The impact of sedimentary alkalinity release on the water column CO2 system in the North Sea. Biogeosciences 13, 841–863. doi: 10.5194/bg-13-841-2016
Bonner J. S., Autenrieth R. L., and Schreiber L. (1990). Aquatic Sediments. Research Journal of the Water Pollution Control Federation 62(4), 593–614. Available online at: http://www.jstor.org/stable/25043883.
Carlton H., Champlin L., Jeppesen R., Haskins J. C., Rahman F. I., and Watson E. B. (2023). Tidal restrictions in a central Californian estuarine system are associated with higher mean pH, but increased low-pH exposure. Mar. Ecol. Prog. Ser. 703, 177–182. doi: 10.3354/meps14209
Castillo N., Gaitán-Espitia J. D., Quintero-Galvis J. F., Saldías G. S., Martel S. I., Lardies M. A., et al. (2024). Small-scale geographic differences in multiple-driver environmental variability can modulate contrasting phenotypic plasticity despite high levels of gene flow. Sci. Total Environ. 954, 176772. doi: 10.1016/j.scitotenv.2024.176772
Christianen M. J. A., van der Heide T., Holthuijsen S. J., van der Reijden K. J., Borst A. C. W., and Olff H. (2017). Biodiversity and food web indicators of community recovery in intertidal shellfish reefs. Biol. Conserv. 213, 317–324. doi: 10.1016/j.biocon.2016.09.028
Clasing E., Brey T., Stead R., Navarro J., and Asencio G. (1994). Population dynamics of Venus antiqua (Bivalvia: Veneracea) in the Bahía de Yaldad, Isla de Chiloé, southern Chile. J. Exp. Mar. Biol. Ecol. 177, 171–186. doi: 10.1016/0022-0981(94)90235-6
Clements J. C. and Darrow E. S. (2018). Eating in an acidifying ocean: a quantitative review of elevated CO 2 effects on the feeding rates of calcifying marine invertebrates. Hydrobiologia 820, 1–21. doi: 10.1007/s10750-018-3665-1
Clements J. C. and Hunt H. L. (2017). Effects of CO2-driven sediment acidification on infaunal marine bivalves: a synthesis. Mar. pollut. Bull. 117, 6–16. doi: 10.1016/j.marpolbul.2017.01.053
Clements J. C. and Hunt H. L. (2018). Testing for sediment acidification effects on within-season variability in juvenile soft-shell clam (Mya arenaria) abundance on the northern shore of the Bay of Fundy. Estuar. Coasts 41, 471–483. doi: 10.1007/s12237-017-0270-x
Clements J. C., Woodard K. D., and Hunt H. L. (2016). Porewater acidification alters the burrowing behavior and post-settlement dispersal of juvenile soft-shell clams (Mya arenaria). J. Exp. Mar. Biol. Ecol. 477, 103–111. doi: 10.1016/j.jembe.2016.01.013
Cummings V., Vopel K., and Thrush S. (2009). Terrigenous deposits in coastal marine habitats: influences on sediment geochemistry and behaviour of post-settlement bivalves. Mar. Ecol. Prog. Ser. 383, 173–185. doi: 10.3354/meps07983
Davis A. M., Plough L. V., and Paynter K. T. (2023). Intraspecific patterns of mortality and cardiac response to hypoxia in the eastern oyster, Crassostrea virginica. J. Exp. Mar. Biol. Ecol. 566, 151921. doi: 10.1016/j.jembe.2023.151921
Dickson A. G. and Millero F. J. (1987). A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep-Sea Res. 34, 1733–1743. doi: 10.1016/0198-0149(87)90021-5
Eriksson B. K., van der Heide T., van de Koppel J., Piersma T., van der Veer H. W., and Olff H. (2010). Major changes in the ecology of the Wadden Sea: human impacts, ecosystem engineering and sediment dynamics. Ecosystems 13, 752–764. doi: 10.1007/s10021-010-9352-3
Fernández C., Poupin M. J., Lagos N. A., Broitman B. R., and Lardies M. A. (2024). Physiological resilience of intertidal chitons in a persistent upwelling coastal region. Scient. Rep. 14, 21401. doi: 10.1038/s41598-024-72488-8
Field C. B. (1998). Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281, 237–240. doi: 10.1126/science.281.5374.237
Flombaum P. and Martiny A. C. (2021). Diverse but uncertain responses of picophytoplankton lineages to future climate change. Limnology Oceanography 66, 4171–4181. doi: 10.1002/lno.v66.12
Gaitán-Espitia J. D., Bacigalupe L. D., Opitz T., Lagos N. A., Timmermann T., and Lardies M. A. (2014). Geographic variation in thermal physiological performance of the intertidal crab Petrolisthes violaceus along a latitudinal gradient. J. Exp. Biol. 217, 4379–4386. doi: 10.1242/jeb.108217
Gaitán-Espitia J. D., Marshall D., Dupont S., Bacigalupe L. D., Bodrossy L., and Hobday A. J. (2017). Geographical gradients in selection can reveal genetic constraints for evolutionary responses to ocean acidification. Biol. Lett. 13, 20160784. doi: 10.1098/rsbl.2016.0784
Gazeau F., Parker L. M., Comeau S., Gattuso J.-P., O’connor W. A., Martin S., et al. (2013). Impacts of ocean acidification on marine shelled molluscs. Mar. Biol. 160, 2207–2245. doi: 10.1007/s00227-013-2219-3
Gleckler P. J., Durack P. J., Stouffer R. J., Johnson G. C., and Forest C. E. (2016). Industrial-era global ocean heat uptake doubles in recent decades. Nat. Clim. Change 6, 394–398. doi: 10.1038/nclimate2915
Goethel C. L., Grebmeier J. M., Cooper L. W., and Miller T. J. (2017). Implications of ocean acidification in the Pacific Arctic: experimental responses of three arctic bivalves to decreased pH and food availability. Deep Sea Res. Part II: Trop. Stud. Oceanogr. 144, 112–124. doi: 10.1016/j.dsr2.2017.08.01
Harayashiki C. A. Y., Márquez F., Cariou E., and Castro Í.B. (2020). Mollusk shell alterations resulting from coastal contamination and other environmental factors. Environmen. Pollut. 265, 114881. doi: 10.1016/j.envpol.2020.114881
Harvey B. P., Gwynn-Jones D., and Moore P. J. (2013). Meta-analysis reveals complex marine biological responses to the interactive effects of ocean acidification and warming. Ecol. Evol. 3, 1016–1030. doi: 10.1002/ece3.2013.3.issue-4
Hernández A. F., Cubillos L. A., and Quiñones R. A. (2011). Evaluación talla estructurada de los stocks de Ensis macha y Tagelus dombeii en el Golfo de Arauco, Chile. Rev. Biol. Mar. Oceanogr. 46, 157–176. doi: 10.4067/S0718-19572011000200006
Hettinger A., Sanford E., Hill T. M., Hosfelt J. D., Russell A. D., and Gaylord B. (2013). The influence of food supply on the response of Olympia oyster larvae to ocean acidification. Biogeosciences 10, 6629–6638. doi: 10.5194/bg-10-6629-2013
Hoegh-Guldberg O., Mumby P. J., Hooten A. J., Steneck R. S., Greenfield P., Gomez E., et al. (2007). Coral reefs under rapid climate change and ocean acidification. Science 318, 1737–1742. doi: 10.1126/science.1152509
Hohaia A., Vopel K., and Pilditch C. A. (2014). Thin terrestrial sediment deposits on intertidal sandflats: effects on pore-water solutes and juvenile bivalve burial behaviour. Biogeosciences 11, 2225–2235. doi: 10.5194/bg-11-2225-2014
Hu M. Y., Casties I., Stumpp M., Ortega-Martinez O., and Dupont S. (2014). Energy metabolism and regeneration are impaired by seawater acidification in the infaunal brittlestar Amphiura filiformis. J. Exp. Biol. 217, 2411–2421. doi: 10.1242/jeb.100024
Iram N., Maher D. T., Lovelock C. E., Baker T., Cadier C., and Adame M. F. (2022). Climate change mitigation and improvement of water quality from the restoration of a subtropical coastal wetland. Ecol. Appl. 32, e2620. doi: 10.1002/eap.v32.5
Jaramillo E., Contreras H., and Duarte C. (2007). “Community structure of the macroinfauna inhabiting tidal flats characterized by the presence of different species of burrowing bivalves in Southern Chile,” in Biodiversity in Enclosed Seas and Artificial Marine Habitats. Developments in Hydrobiology, vol. 193 . Eds. Relini G. and Ryland J. (Dordrecht: Springer). doi: 10.1007/978-1-4020-6156-1_7
Johnson G. C. and Lumpkin R. L. (2024). Global oceans [in “State of the climate in 2023”. Bull. Amer. Meteor. Soc 105, S156–S213. doi: 10.1175/BAMS-D-24-0100.1
Kessouri F., McWilliams J. C., Bianchi D., Sutula M., Renault L., Deutsch C., et al. (2021). Coastal eutrophication drives acidification, oxygen loss, and ecosystem change in a major oceanic upwelling system. PNAS 118, e2018856118. doi: 10.1073/pnas.2018856118
Kwon E. Y., Sreeush M. G., Timmermann A., Karl D. M., Church M. J., Lee S. S., et al. (2022). Nutrient uptake plasticity in phytoplankton sustains future ocean net primary production. Sci. Adv. 8, eadd2475. doi: 10.1126/sciadv.add2475
Lagos N. A., Benitez S., Duarte C., Lardies M. A., Broitman B. R., Tapia C., et al. (2016). Effects of temperature and ocean acidification on shell characteristics of Argopecten purpuratus: implications for scallop aquaculture in an upwelling-influenced area. Aquacult. Environ. Interact. 8, 357–370. doi: 10.3354/aei00183
Lardies M. A., Caballero P., Duarte C., and Poupin M. J. (2021). Geographical variation in phenotypic plasticity of intertidal sister limpet’s species under ocean acidification scenarios. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.647087
Lardies M. A., Clasing E., Navarro J. M., and Stead R. (2001). Effects of environmental variables on burial depth of two infaunal bivalves inhabiting a tidal flat in southern Chile. J. Mar. Biol. Assoc. UK. 81, 809–816. doi: 10.1017/S0025315401004635
Lawrence J., Popova E., Yool A., and Srokosz M. (2015). On the vertical phytoplankton response to an ice-free Arctic Ocean. J. Geophys. Res. Oceans 120, 8571–8582. doi: 10.1002/2015JC011180
Lefevre S. (2016). Are global warming and ocean acidification conspiring against marine ectotherms? A meta-analysis of the respiratory effects of elevated temperature, high CO2 and their interaction. Conserv. Physiol. 4 (1), cow009. doi: 10.1093/conphys/cow009
Lépez I., Aracena O., Carmona A., Espinoza A., Fuentes L., Sánchez J., et al. (1997). “Caracterización bioeconómica de las pesquerías de huepo (Ensis macha) y navajuela (Tagelus dombeii) en la VIII Región,” in Informe Final Proyecto FIP N°95-20a. (Chile: Subsecretaría de Pesca, Valparaíso).
Leung J. Y., Zhang S., and Connell S. D. (2022). Is ocean acidification really a threat to marine calcifiers? A systematic review and meta-analysis of 980+ studies spanning two decades. Small 18, 2107407. doi: 10.1002/smll.202107407
Liang J., Liu Y., Zhu F., Li Y., Liang S., and Guo Y. (2022). Impact of ocean acidification on the physiology of digestive gland of razor clams Sinonovacula constricta. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.1010350
Liu X., Stock C. A., Dunne J. P., Lee M., Shevliakova E., Malyshev S., et al. (2021). Simulated global coastal ecosystem responses to a half-century increase in river nitrogen loads. Geophysical Res. Lett. 48, e2021GL094367. doi: 10.1029/2021GL094367
Marshall D. J. and McQuaid C. D. (2020). Metabolic regulation, oxygen limitation and heat tolerance in a subtidal marine gastropod reveal the complexity of predicting climate change vulnerability. Front. Physiol. 11, 1106. doi: 10.3389/fphys.2020.01106
Martel S. I., Fernández C., Lagos N. A., Labra F. A., Duarte C., Vivanco J. F., et al. (2022). Acidification and high- temperature impacts on energetics and shell production of the edible clam Ameghinomya antiqua. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.972135
McGarrigle S. A. and Hunt H. L. (2024). Infaunal invertebrate community relationships to water column and sediment abiotic conditions. Mar. Biol. 171, 3. doi: 10.1007/s00227-023-04318-w
Medeiros I. P. M. and Souza M. M. (2023). Acid times in physiology: a systematic review of the effects of ocean acidification on calcifying invertebrates. Environmen. Res. 231, 116019. doi: 10.1016/j.envres.2023.116019
Mehrbach C., Culberson C. H., Hawley J. E., and Pytkowicx R. M. (1973). Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol. Oceanogr. 18, 897–907. doi: 10.4319/lo.1973.18.6.0897
Meseck S. L., Mercaldo-Allen R., Kuropat C., Clark P., and Goldberg R. (2018). Variability in sediment-water carbonate chemistry and bivalve abundance after bivalve settlement in Long Island Sound, Milford, Connecticut. Mar. pollut. Bull. 135, 165–175. doi: 10.1016/j.marpolbul.2018.07.025
Michaelidis B., Ouzounis C., Paleras A., and Pörtner H. O. (2005). Effects of long-term moderate hypercapnia on acid–base balance and growth rate in marine mussels Mytilus galloprovincialis. Mar. Ecol. Prog. Ser. 293, 109–118. doi: 10.3354/meps293109
Morse M. P. and Zardus J. D. (1997). “Bivalvia,” in Microscopic anatomy of invertebrates, vol. 6A . Eds. Harrison F. W. and Kohn A. J. I. (John Wiley & Sons Inc, New York), 7–118. Mollusca.
Murray N. J., Phinn S. R., DeWitt M., Ferrari R., Johnston R., Lyons M. B., et al. (2019). The global distribution and trajectory of tidal flats. Nature 565, 222. doi: 10.1038/s41586-018-0805-8
Navarro J. M., Clasing E., Lardies M. A., and Stead R. A. (2008). Feeding behavior of the infaunal bivalve Tagelus dombeii (Lamarck 1818). Suspension vs. deposit feeding. Rev. Biol. Mar. Oceanogr. 43, 599–605. doi: 10.4067/S0718-19572008000300019
Navarro J. M., Duarte C., Manríquez P. H., Lardies M. A., Torres R., Acuña K., et al. (2016). Ocean warming and elevated carbon dioxide: multiple stressor impacts on juvenile mussels from southern Chile. ICES J. Mar. Sci. 73, 764–771. doi: 10.1093/icesjms/fsv249
Navarro J. M., Duarte C., Manríquez P. H., Torres R., Vargas C. A., Lardies M. A., et al. (2013). Impact of medium-term exposure to elevated pCO2 levels on the physiological energetics of the mussel Mytilus Chilensis. Chemosphere 90, 242–248. doi: 10.1016/j.chemosphere.2012.09.063
Osores S. J., Lagos N. A., San Martín V., Manríquez P. H., Vargas C. A., Torres R., et al. (2017). Plasticity and inter-population variability in physiological and life-history traits of the mussel Mytilus Chilensis: A reciprocal transplant experiment. J. Exp. Mar. Biol. Ecol. 490, 1–12. doi: 10.1016/j.jembe.2017.02.005
Palmer A. R. (1982). Growth in marine gastropods. A non-destructive technique for independently measuring shell and body weight. Malacologia 23, 63–74.
Pierrot D., Lewis E., and Wallace D. W. R. (2006). “MS excel program developed for CO2 system calculations., ORNL/CDIAC-105,” in Carbon Dioxide Information Analysis Center Oak Ridge National Laboratory US Department of Energy(Oak Ridge Tennessee: ORNL Environmental Sciences Division).
Precht E., Franke U., Polerecky L., and Huettel M. (2004). Oxygen dynamics in permeable sediments with wave-driven pore water exchange. Limnol. Oceanogr. 49, 693–705. doi: 10.4319/lo.2004.49.3.0693
Ramajo L., Fernández C., Núñez Y., Caballero P., Lardies M. A., and Poupin M. J. (2019). Physiological responses of juvenile Chilean scallops (Argopecten purpuratus) to isolated and combined environmental drivers of coastal upwelling. ICES J. Mar. Sci. 76, 1836–1849. doi: 10.1093/icesjms/fsz080
Ramajo L., Hendriks I. E., Marbà N., Sejr M. K., Blicher M. E., Lagos N. A., et al. (2016a). Food supply confers calcifiers resistance to ocean acidification. Scient. Rep. 6, 1–6. doi: 10.1038/srep19374
Ramajo L., Marba N., Prado L., Peron S., Lardies M. A., Rodríguez-Navarro A. B., et al. (2016b). Biomineralization changes with food supply confer juvenile scallops (Argopecten purpuratus) resistance to ocean acidification. Glob. Change Biol. 22, 2025–2037. doi: 10.1111/gcb.13179
Rathbone M., Brown K. T., and Dove S. (2022). Tolerance to a highly variable environment does not infer resilience to future ocean warming and acidification in a branching coral. Limnol. Oceanogr. 67, 272–284. doi: 10.1002/lno.11991
R Core Team (2024). A Language and Environment for Statistical Computing. R 4.5.0. version: 2025-04-11. (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Rodríguez-Romero A., Gaitán-Espitia J. D., Opitz T., and Lardies M. A. (2022). Heterogeneous environmental seascape across a biogeographic break influences the thermal physiology and tolerances to ocean acidification in an ecosystem engineer. Divers. Distrib. 28, 1542–1553. doi: 10.1111/ddi.13478
Salas M. C., Defeo O., Firstater F., and Narvarte M. (2022). Impact of a macrofaunal ecosystem engineer on its assemblage and its habitat in mixed sediments as assessed through manipulative experiments. J. Exp. Mar. Biol. Ecol. 554, 151766. doi: 10.1016/j.jembe.2022.151766
Sánchez J., Hernández A., Agüero M., González E., Miranda L., Vásquez C., et al. (2003). “Ordenamiento de la pesquería de huepo y navajuela,” in Informe Final Proyecto FIP N°2002- 26. (Chile: Subsecretaría de Pesca, Valparaíso).
Shi Y. and Li Y. (2024). Impacts of ocean acidification on physiology and ecology of marine invertebrates: a comprehensive review. Aquat. Ecol. 58, 207–226. doi: 10.1007/s10452-023-10058-2
Singer A., Bijleveld A. I., Hahner F., Holthuijsen S. J., Hubert K., Kerimoglu O., et al. (2023). Long-term response of coastal macrofauna communities to de- eutrophication and sea level rise mediated habitat changes, (1980s versus 2018). Front. Mar. Sci. 9, 1–20. doi: 10.3389/fmars.2022.963325
Stead R. A., Clasing E., Lardies M. A., Arratia L. P., and Urrutia G. (2002). The significance of contrasting feeding strategies on the reproductive cycle in two coexisting tellinacean bivalves. J. Mar. Biol. Assoc. United Kingdom 82, 443–453. doi: 10.1017/S0025315402005702
Torres R., Pantoja S., Harada N., González H. E., Daneri G., Frangopulos M., et al. (2011). Air-sea CO2 fluxes along the coast of Chile: From CO2 outgassing in central northern upwelling waters to CO2 uptake in southern Patagonian fjords. J. Geophysical Research: Oceans 116 (C9). doi: 10.1029/2010JC006344
Urrutia G. X., Navarro J. M., Clasing E., and Stead R. A. (2001). The effects of environmental factors on the biochemical composition of the bivalve Tagelus dombeii (Lamarck 1818) (Tellinacea: Solecurtidae) from the intertidal flat of Coihuín, Puerto Montt, Chile. J. Shellfish Res. 20, 1077–1088.
Van Colen C., Ong E. Z., Briffa M., Wethey D. S., Abatih E., Moens T., et al. (2020). Clam feeding plasticity reduces herbivore vulnerability to ocean warming and acidification. Nat. Clim. Change 10, 162–166. doi: 10.1038/s41558-019-0679-2
Vargas C. A., Lagos N. A., Lardies M. A., Duarte C., Manríquez P. H., Aguilera V. M., et al. (2017). Species-specific responses to ocean acidification should account for local adaptation and adaptive plasticity. Nat. Ecol. Evol. 1, 0084. doi: 10.1038/s41559-017-0084
Vergara-Jara M. J., DeGrandpre M. D., Torres R., Beatty C. M., Cuevas L. A., Alarcón E., et al. (2019). Seasonal changes in carbonate saturation state and air-sea CO2 fluxes during an annual cycle in a stratified-temperate fjord (Reloncaví Fjord, Chilean Patagonia). J. Geophys.l Res. Biogeosciences 124, 2851–2865. doi: 10.1029/2019JG005028
Vlaminck E., Moens T., Braeckman U., and Van Colen C. (2023). Ocean acidification and warming modify stimulatory benthos effects on sediment functioning: An experimental study on two ecosystem engineers. Front. Mar. Sci. 10. doi: 10.3389/fmars.2023.1101972
Wethey D. S. and Woodin S. A. (2022). Climate change and Arenicola marina: Heat waves and the southern limit of an ecosystem engineer. Estuar. Coast. Shelf Sci. 276, 108015. doi: 10.1016/j.ecss.2022.108015
Widdicombe S., Spicer J. I., and Kitidis V. (2011). “Effects of ocean acidification on sediment fauna,” in Ocean acidification. Eds. Gattuso J. P. and Hansson L. (Oxford University Press, Oxford), 176–191.
Keywords: razor clam, mollusc, metabolism, heart rate, tidal flat, mesocosm, sediments, global change
Citation: Antivero M, Caballero P, Leppes N and Lardies MA (2025) Infaunal bivalves exhibit resilience to ocean acidification but remain sensitive to food supply. Front. Mar. Sci. 12:1568035. doi: 10.3389/fmars.2025.1568035
Received: 28 January 2025; Accepted: 15 May 2025;
Published: 02 June 2025.
Edited by:
Jose Martin Hernandez-Ayon, Autonomous University of Baja California, MexicoReviewed by:
Liqiang Zhao, Guangdong Ocean University, ChinaFederica Nasi, National Institute of Oceanography and Applied Geophysics, Italy
Araceli Rodriguez-Romero, Spanish National Research Council (CSIC), Spain
Jay Minuti, City University of Hong Kong, Hong Kong SAR, China
Copyright © 2025 Antivero, Caballero, Leppes and Lardies. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marco A. Lardies, bWFyY28ubGFyZGllc0B1YWkuY2w=