 Tomoyuki Yoshida1,2
Tomoyuki Yoshida1,2 Akatsuki Saito2,3 Yuki Iwasaki1,4 Sayuki Iijima1 Terue Kurosawa1 Yuko Katakai5
Akatsuki Saito2,3 Yuki Iwasaki1,4 Sayuki Iijima1 Terue Kurosawa1 Yuko Katakai5 Yasuhiro Yasutomi6
Yasuhiro Yasutomi6 Keith A. Reimann7 Toshiyuki Hayakawa2
Keith A. Reimann7 Toshiyuki Hayakawa2 Hirofumi Akari1,2*
Hirofumi Akari1,2*
- 1 Tsukuba Primate Research Center, National Institute of Biomedical Innovation, Tsukuba, Ibaraki, Japan
- 2 Primate Research Institute, Kyoto University, Inuyama, Aichi, Japan
- 3 International Research Center for Infectious Diseases, The Institute of Medical Science, The University of Tokyo, Minato-ku, Tokyo, Japan
- 4Graduate School of Medicine and Dentistry, Tokyo Medical and Dental University, Bunkyo-ku, Tokyo, Japan
- 5Corporation for Production and Research of Laboratory Primates, Tsukuba, Ibaraki, Japan
- 6Laboratory of Immunoregulation and Vaccine Research, Tsukuba Primate Research Center, National Institute of Biomedical Innovation, Tsukuba, Ibaraki, Japan
- 7Division of Viral Pathogenesis, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA, USA
Natural killer (NK) cells are capable of regulating viral infection without major histocompatibility complex restriction. Hepatitis C is caused by chronic infection with hepatitis C virus (HCV), and impaired activity of NK cells may contribute to the control of the disease progression, although the involvement of NK cells in vivo remains to be proven. GB virus B (GBV-B), which is genetically most closely related to HCV, induces acute and chronic hepatitis upon experimental infection of tamarins. This non-human primate model seems likely to be useful for unveiling the roles of NK cells in vivo. Here we characterized the biological phenotypes of NK cells in tamarins and found that depletion of the CD16+ subset in vivo by administration of a monoclonal antibody significantly reduced the number and activity of NK cells.
Introduction
Natural killer (NK) cells are a component of the innate immune system that play a central role in host defense against viral infection and tumor cells. Much of the evidence for a role for NK cells in controlling viral infections has come from experiments with mice that were genetically modified (Lian and Kumar, 2002) or were treated with NK cell-depleting antibodies (Kasai et al., 1980) or from the study of humans with inherited NK cell deficiencies (Biron et al., 1989; Orange, 2002).
NK cells can be rapidly recruited into infected organs and tissue by chemoattractant factors produced by virus-infected cells and activated resident macrophages, which are also a major source of interferon (IFN) that induces NK cell proliferation, NK cell-mediated cytolysis of virus-infected cells, and the secretion of chemokines (Robertson, 2002). NK cells can kill virus-infected cells by using cytotoxic granules or by recognizing and inducing lysis of antibody-coated target cells (antibody-dependent cell cytotoxicity) via antibody binding receptor CD16. For instance, human blood NK cells are cytotoxic against dengue virus-infected cells in target organs via direct cytolysis and antibody-dependent cell-mediated cytotoxicity (reviewed by Navarro-Sánches et al., 2005). Early activity of NK cells may be important for clearing acute infections such as that of dengue virus. However, the effect that NK cells may exert on chronic infections with viruses such as hepatitis C virus (HCV) is less clear.
HCV is the causative agent of chronic hepatitis C, cirrhosis, and finally liver cancer. In general, acquired and innate immunity induced by acute HCV infection is not sufficient for the viral clearance, and persistent HCV infection frequently leads to progression to chronic hepatitis (reviewed by Cheent and Khakoo, 2010). It was reported that dendritic cells (DCs) in HCV infection were not responsive to IFN-α, and thus failed to promote subsequent activation of NK cells as a primary innate immune response (reviewed by Kanto, 2008). This is in agreement with the finding that the killing activity of NK cells in patients with chronic hepatitis C is inactivated in in vitro studies (Deignan et al., 2002; Golden-Mason et al., 2008). These data suggest that the dysfunction of NK cells contributes to the persistent infection of HCV and chronic hepatitis. On the other hand, it was suggested that inappropriately activated NK cells caused liver injury after the viral infection (Liu et al., 2000). The population of NK cells is relatively minor in peripheral lymphoid organs but is abundant in liver, raising a question as to their function in the innate immune response to acute and chronic HCV infection in the liver. It is possible that NK cells partially regulate the replication of HCV in this organ during early infection whereas they promote the liver dysfunction in chronic HCV infection. To examine these possibilities, it is necessary to clarify the involvement of NK cells in vivo in HCV infection. However, it is questionable whether the results of ex vivo analyses of NK cells would reflect their actual roles in vivo. Therefore, it might be more informative to study the function of NK cells directly by means of in vivo depletion technique in animal models.
A chimpanzee model of HCV infection has frequently been employed to evaluate the role of acquired antiviral immune responses, although the involvement of NK cells has not been fully evaluated because of the limitations on the use of chimpanzees due to ethical and financial restrictions (Cohen and Lester, 2007). Accordingly, New World monkeys infected with GB virus B (GBV-B) appear to be a promising model because (i) among viruses so far known, GBV-B is genetically most closely related to HCV and can infect New World monkeys, including tamarins, marmosets and owl monkeys, but not Old World monkeys (reviewed by Akari et al., 2009), (ii) tamarins develop acute and chronic hepatitis after experimental GBV-B infection (Bukh et al., 1999; Sbardellati et al., 2001; Lanford et al., 2003; Martin et al., 2003; Ishii et al., 2007; Takikawa et al., 2010), (iii) the infection induces antiviral cellular immune responses (Woollard et al., 2008), and (iv) tamarins and marmosets are commercially available and easily handled, reared and bred. Moreover, tamarins, being primates, may have a similar immune system to humans, and therefore they may be useful for studying the function of NK cells against the hepatitis virus in this tamarin model.
Our final goal is to study the role of NK cells as a major player in innate immunity during the course of the progression of viral hepatitis. Since some basic information regarding the biological characteristics of NK cells still remains unclear, we initially sought to characterize NK cells in tamarins to provide a technical basis for further studies.
Materials and Methods
Animals
Five red-handed tamarins (Saguinus midas) and five cynomolgus monkeys (Macaca fascicularis) were used in this study. The animals were cared for in accordance with National Institute of Biomedical Innovation rules and guidelines for experimental animal welfare, and all protocols were approved by our Institutional Animal Study Committee.
Flow Cytometry
Flow cytometry was performed as previously described (Akari et al., 1997) with a slight modification. Fifty microliters of whole blood from cynomolgus monkeys and tamarins was stained with combinations of fluorescence-conjugated monoclonal antibodies (mAb): anti-CD3 (SP34-2; Becton Dickinson), anti-CD4 (L200; BD Pharmingen), anti-CD8 (CLB-T8/4H8; Sanquin), anti-CD16 (3G8; BD Pharmingen), and anti-CD16 (DJ130c; Dako). Then, erythrocytes were lysed with FACS lysing solution (Becton Dickinson). After having been washed with sample buffer containing phosphate-buffered saline (PBS), 1% fetal calf serum (FCS), and 1% formaldehyde, the labeled cells were resuspended in the sample buffer. The expression of the immunolabeled molecules on the lymphocytes was analyzed with a FACSCanto II flow cytometer (Becton Dickinson). Peripheral blood mononuclear cells (PBMCs) were separated from the blood of these monkeys by a Ficoll-Paque gradient method. The cells were resuspended in complete medium composed of RPMI-1640 medium supplemented with 10% FCS, 1% penicillin/streptomycin, 2 mM HEPES and 55 μM 2-mercaptoethanol at 4°C until use. Fluorochrome-labeled mouse mAbs were reacted with 2 × 105 PBMCs at 4°C for 30 min. The labeled cells were washed with PBS containing 1% FCS, and resuspended in the sample buffer. The expression of the immunolabeled molecules on the lymphocytes was analyzed as mentioned above.
Flow Cytometric 5-(and 6)-Carboxyfluorescein Diacetate Succinimidyl Ester (CfSE)/7-Amino Actinomycin D (7-AAD) Cytotoxic Assay
Peripheral blood mononuclear cells were separated from the blood of these monkeys by a Ficoll-Paque gradient method. These PBMCs were then resuspended in complete medium at 37°C until use. The flow cytometric CFSE/7-AAD cytotoxicity assay was performed as previously described (Lecoeur et al., 2001) with slight modifications. K562 cells (3 × 106) were labeled with 500 nM CFSE (from a 1 mM stock solution in dimethyl sulfoxide [Sigma] stored at −20°C) in Hanks’ Balanced Salt Solution for 8 min at 37°C in total of 2 ml. The cells were then washed twice in complete medium and used immediately for the cytotoxicity assay. The CFSE-labeled target cells (20,000 cells) were used at different E (effector):T (target) ratios (0:1, 3:1, and 9:1). After 24 h incubation, the cells were stained with 0.25 μg/ml of 7-AAD and incubated for 10 min at 37°C in a CO2 incubator. The cells were washed twice with 1% FCS–PBS, resuspended in sample buffer and analyzed immediately by flow cytometry.
Magnetic Cell Separation
Magnetic cell separation (MACS) was performed as previously described (Tenorio and Saavedra, 2005) with slight modifications. PBMCs (1 × 107) were washed with 3 ml of MACS buffer composed of PBS with 2 mM EDTA and 0.5% bovine serum albumin, and resuspended in 100 μl of the same buffer. Ten microliters of fluorescein isothiocyanate (FITC)-labeled anti-CD16 mAb (3G8) was added. The cells with or without the mAb were incubated for 10 min at 4°C, washed with 1 ml of MACS buffer, and resuspended in 80 μl of the same buffer. They were mixed with 20 μl of anti-FITC MicroBeads and incubated for 15 min at 4°C, washed with 1 ml of MACS buffer, and resuspended in 500 μl of the same buffer. The CD16-positive cells were separated by negative selection using LD columns and a MACS separation unit following the instructions provided by the manufacturer (Miltenyi Biotec). CD16-negative cells were resuspended in complete medium and co-cultured with K562 cells at 37°C for the NK cytotoxicity assay immediately.
Detection of Circulating Anti-CD16 Mab (3G8)
Concentrations of an anti-CD16 antibody (3G8) in plasma samples were assessed using a mouse IgG1 Quantitative ELISA Kit (Bethyl Laboratory, Inc.). The assay was performed according to the manufacturer’s instruction with a slight modification. To detect the mAb in monkey plasma, 96-well enzyme-linked immunosorbent assay (ELISA) plates were coated with a capture antibody and incubated for 1 h at 37°C and washed with wash solution (50 mM Tris, 0.14 M NaCl, 0.05% Tween 20, pH 8.0) three times. The plates were blocked with blocking solution (Postcoat) for 30 min at 37°C. Plasma samples from antibody-treated monkeys were diluted in dilution buffer (50 mM Tris, 0.14 M NaCl, 1% bovine serum albumin, 0.05% Tween 20, pH 8.0), applied to the wells in serial dilutions, incubated for 1 h at 37°C and washed with the wash solution five times. Goat anti-mouse IgG1 conjugated with horseradish peroxidase and diluted 1:50000 in dilution buffer was added to each well and incubated for 1 h at 37°C. Each well was washed with the wash solution five times. Substrate solution was added to each well and incubated for 10–15 min at room temperature, and then the reaction was stopped with H2SO4. Optical density was measured using an ELISA reader at 450 nm.
In vivo Depletion of CD16 Positive Cells
Mouse anti-human CD16 (3G8) mAb (Fleit et al., 1982) was produced in serum-free medium and purified using protein A affinity chromatography. Endotoxin levels were lower than 1 EU/mg. The antibody was administered to tamarins (Tm 05-003, Tm 06-020) and cynomolgus monkeys (Mf 00-005, Mf 99-110) intravenously at 50 mg/kg at a rate of 18 ml/min using a syringe pump. Lymphocyte subsets were monitored for 3 weeks after the administration.
Statistical Analysis
Statistical analyses of lymphocyte ratios were performed using Student’s t-test and single-factor ANOVA, followed by Fisher’s protected least-significant difference post hoc test by using StatView software (SAS Institute, NC, USA). The results were confirmed in more than three independent experiments in tamarins and cynomolgus monkeys.
Results
Lymphocyte Subsets in Tamarins
First, we examined the lymphocyte subsets in tamarins as compared with cynomolgus monkeys (Figure 1). The percentages of T and B lymphocytes indicated as CD20−CD3+ and CD20+CD3− subsets in the total lymphocytes were found to be 68.8% (range 41.9–68.8%) and 12.3% (range 11.8–12.6%) in tamarins and 68.4% (range 42.6–68.4%) and 10.2% (range 9.1–11.4%) in cynomolgus monkeys, respectively. The percentage of CD4+ T cells in the CD3+ subset was 45.5% (range 41.9–52.5%) and 55.3% (range 42.6–64.4%) while that of CD8+ T cells was 41.0% (range 35.8–44.5%) and 31.2% (range 29.3–34.6%) in tamarins and cynomolgus monkeys, respectively. Next, the NK cell subset was determined as CD3− CD16+ lymphocytes in this study. The percentage of NK cells was 30.5% (range 16.9–52.5%) and 18.9% (range 13.7–22.4%) in tamarins and cynomolgus monkeys, respectively. We analyzed statistically whether these lymphocyte ratios were different between tamarins and cynomolgus monkeys, and found that there were no significant differences of the lymphocyte ratios between them. We therefore concluded that the proportions of the major lymphocyte subsets in tamarins were relatively similar to those in cynomolgus monkeys.
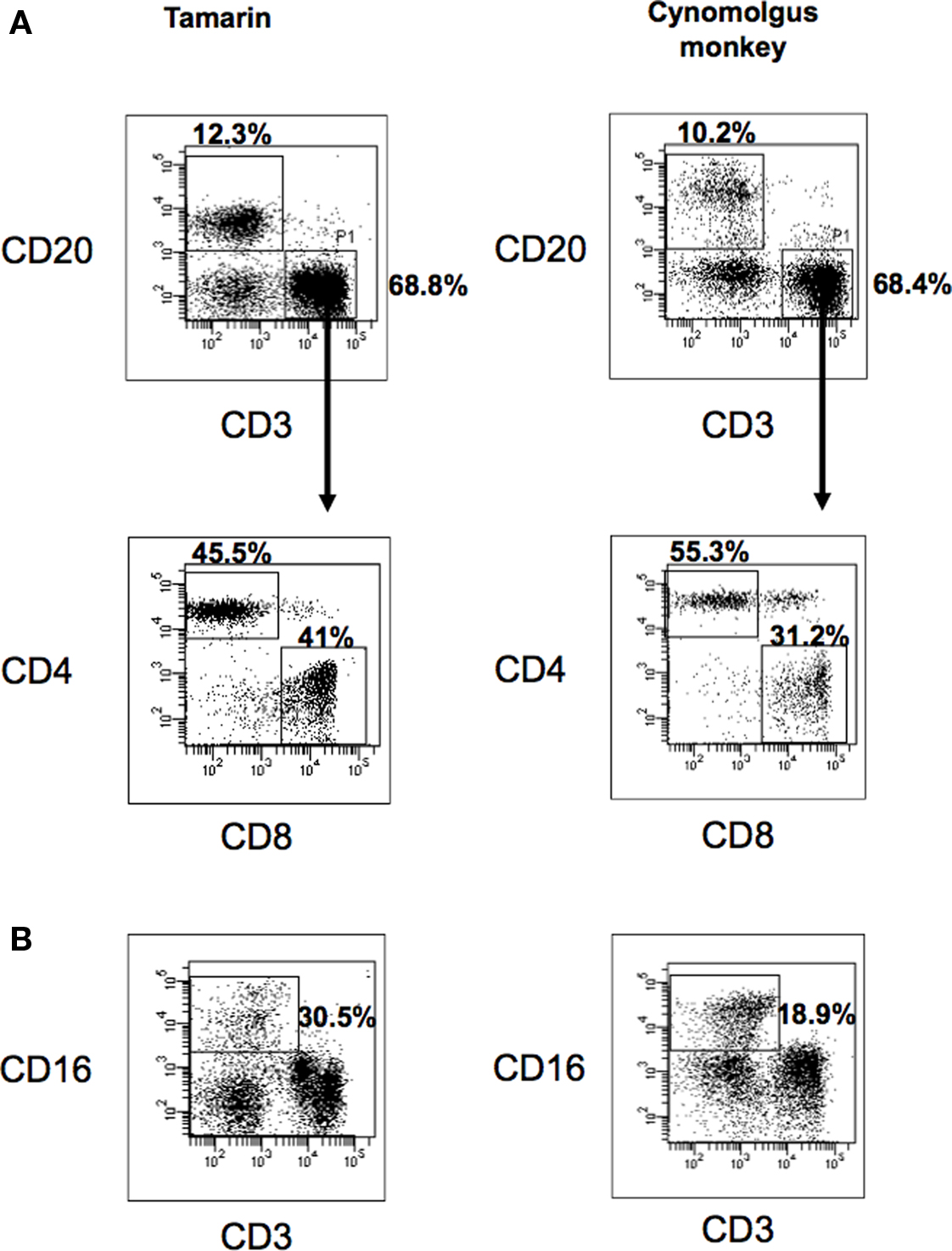
Figure 1. Flow cytometric analysis of CD3, CD20, CD4, CD8, and CD16 expression on lymphocytes in cynomolgus monkeys and tamarins. Representative results in tamarins and cynomolgus monkeys are shown. (A) Flow cytometric analysis of CD3 and CD20 expression in lymphocytes (top panels) and CD4 and CD8 expression in CD3+ population (bottom panels) of both monkeys. (B) Flow cytometric analysis of CD3 and CD16 expression in both monkey species. The numbers denote the percentages of lymphocytes. We analyzed statistically whether these lymphocyte ratios were different between tamarins and cynomolgus monkeys by using StatView software. There were no significant differences of the lymphocyte ratios between them.
Fluorescence-Based in Vitro Assay for Quantitatively Evaluating Natural Killer Activity
Natural killer cell cytotoxic assays conventionally require considerable numbers of PBMCs, and this has been a major hurdle for analyzing the NK activity in small New World monkeys due to the limited availability of their blood. Therefore, we employed an alternative method using a fluorescence-based assay to assess the activity of NK cells in tamarins as previously described (Lecoeur et al., 2001) with slight modifications. When CFSE-stained K562 target cells were incubated with the effector PBMCs obtained from tamarins at an effector/target (E/T) ratio of 9:1, 42% of the K562 cells were positive for 7-AAD, which stains apoptotic cells (Figure 2A). We confirmed that the killing activity of NK cells was dose-dependent, and that the level in tamarins was higher than that in cynomolgus monkeys (Figures 2B and 3).
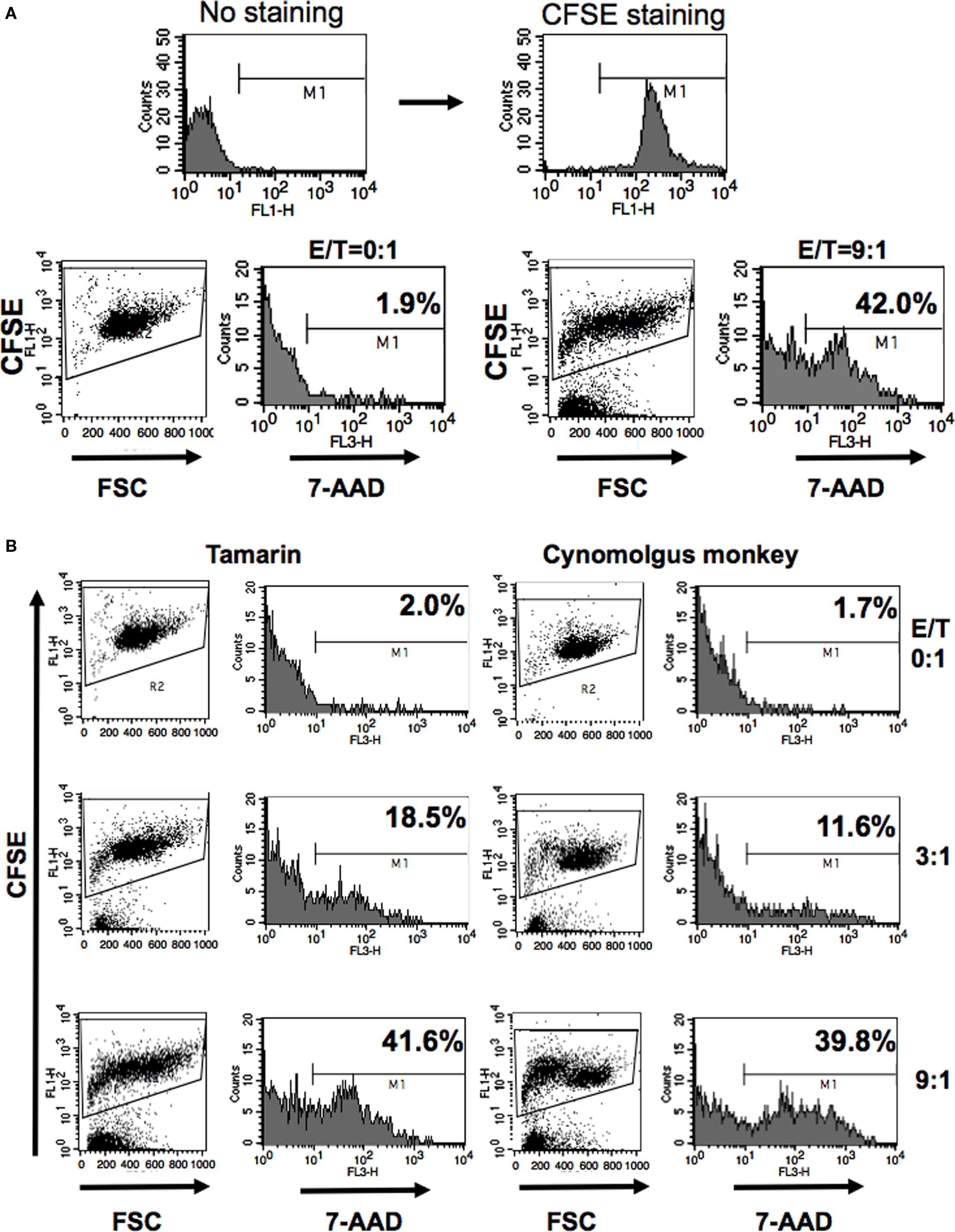
Figure 2. Cytotoxicity assay by CFSE and 7-AAD staining. (A) CFSE-stained K562 cells were co-cultured with PBMCs at E/T ratios of 9:1 and 0:1. The CFSE-stained K562 cells were first gated by FSC and SSC characteristics, and 3000 events were captured. (B) CFSE-stained K562 target cells were killed by tamarin PBMCs using E/T ratios of 9:1, 3:1, and 0:1. The numbers in the panels denoted the percentages of 7-AAD positive cells. We analyzed statistically whether these killing activities were different between tamarins and cynomolgus monkeys by using StatView software.

Figure 3. Dose-dependency of killing activity of NK cells in tamarins. (A,B) K562 target cells were stained with CFSE and co-cultured with PBMCs as described in Section “Materials and Methods”. CFSE-stained K562 target cells were killed by PBMCs of tamarins and cynomolgus monkeys in a dose-dependent manner. For all experiments, the number of observations used to calculate the mean were n = 5. We analyzed statistically whether these killing activities were different between tamarins and cynomolgus monkeys by using StatView software.
Next, in order to examine if CD16+ lymphocytes represent a major population with NK activity, CD16− PBMCs were obtained by negative selection using MACS (Figure 4A) in both tamarins and cynomolgus monkeys. We found that depletion of CD16+ cells greatly attenuated the killing activity in both tamarins and cynomolgus monkeys (Figure 4B), indicating that CD16+ lymphocytes are a major population with NK activity.
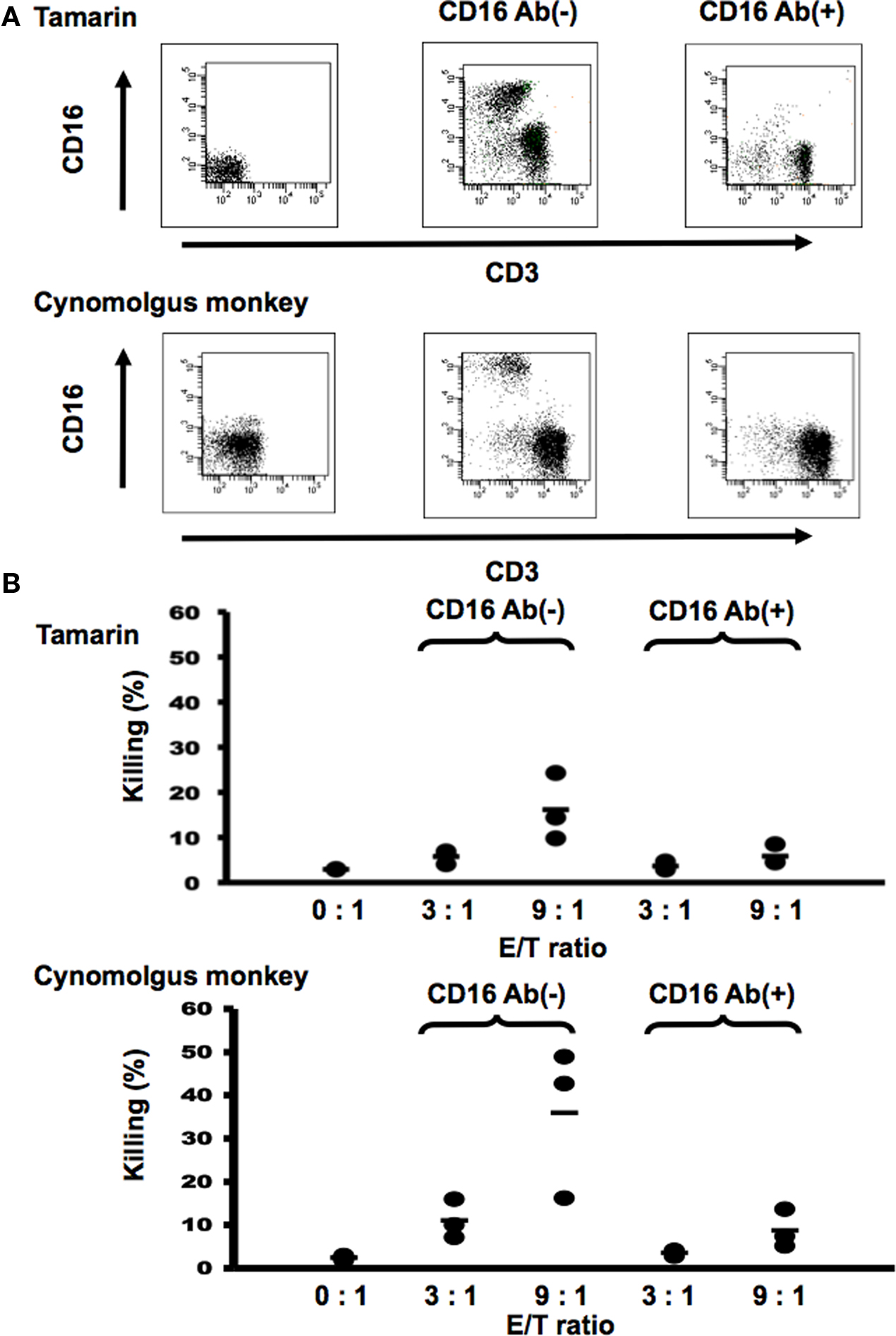
Figure 4. CD16+ cells were a major population with natural killer activity in tamarins. (A) CD16+ cells were depleted from PBMCs by MACS as described in Section “Materials and Methods”. CD16− PBMCs were obtained by negative selection using MACS. (B) K562 cells were stained with CFSE and co-cultured with CD16-treated or untreated PBMCs as described in Section “Materials and Methods”. Results shown are representative of three independent experiments.
In vivo Depletion of CD16+ NK Cells Using a Murine Anti-CD16 MAb
We next sought to establish a system to directly evaluate the role of NK cells in tamarins. We asked if the administration of an anti-CD16 (3G8) mAb could deplete CD16+ lymphocytes in vivo. Tamarins were intravenously administered 3G8 or control mAb (MOPC-21) at a dose of 50 mg/kg. Using an anti-CD16 antibody that is not cross-blocked by 3G8 (clone DJ130c), it was found that at 1–3 days after the treatment CD16+ cells were completely depleted, followed by recovery to the initial levels at around 2 weeks after the administration, which was consistent with the results in cynomolgus monkeys (Figure 5B). It is noteworthy that the numbers of CD4+/CD8+ T and B lymphocytes were not affected by the treatment and that administration of control antibody did not deplete CD16+ cells during the period tested (data not shown), showing that the effect of 3G8 on CD16+ cells was specific (data not shown). We also measured the concentration of the 3G8 mAb in the plasma of antibody-treated monkeys. As shown in Figure 6, the concentration of 3G8 reached a plateau at day 1, followed by a gradual decrease in both tamarins and cynomolgus monkeys, which was consistent with the kinetics of CD16+ cells. In the case of MOPC-21 administration to tamarins, similar kinetics of its concentration with that of 3G8 were observed (data not shown).
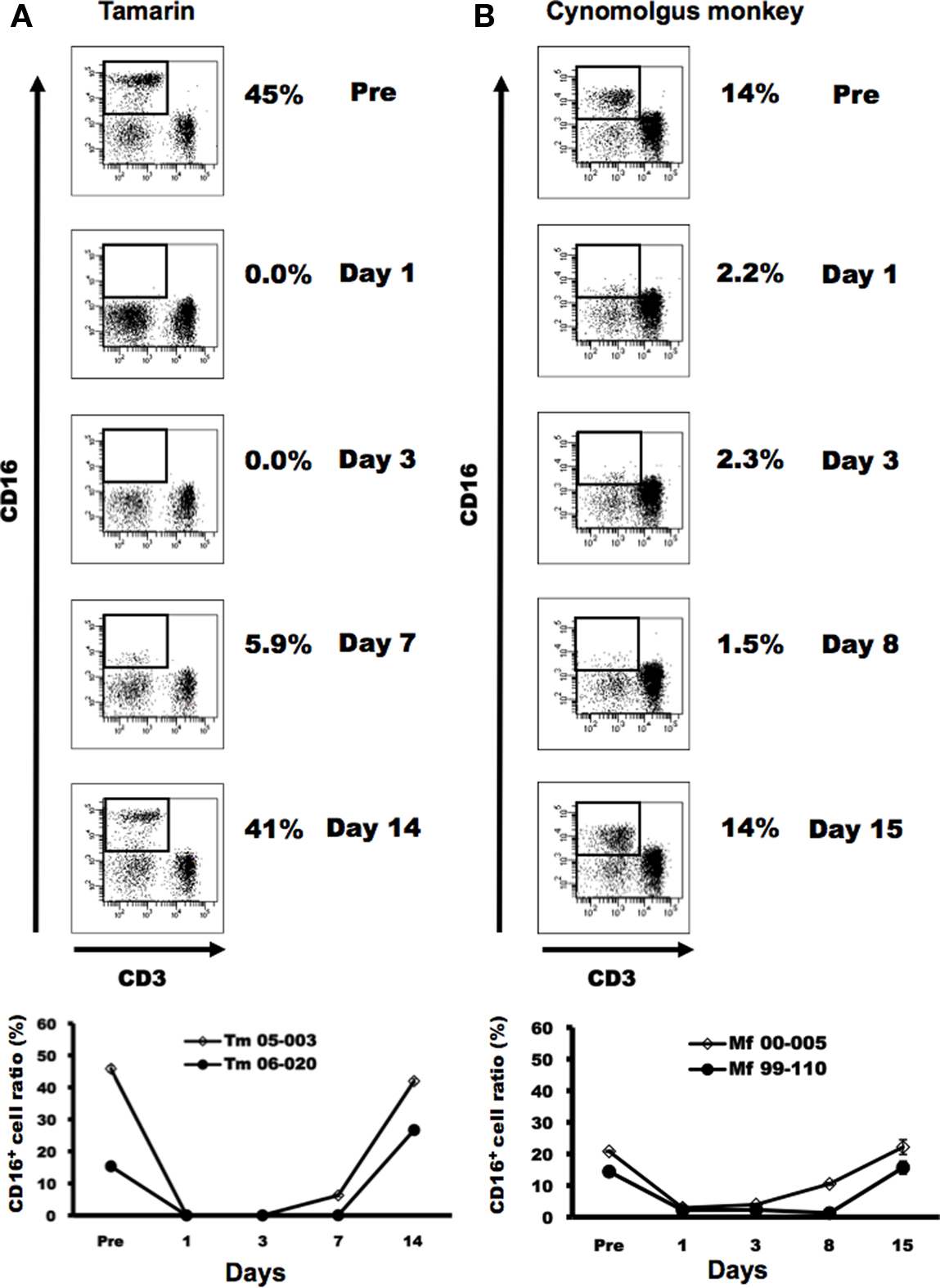
Figure 5. In vivo depletion of CD16+ NK cells using a murine anti-CD16 (3G8) mAb in tamarins. (A,B) Tamarins and cynomolgus monkeys were administered with 50 mg/kg of the 3G8 mAb. CD16+ NK cell numbers were determined in whole blood specimens. Tamarins: Tm 05-003 and Tm 06-020. Cynomolgus monkeys: Mf 00-005 and Mf 99-110.
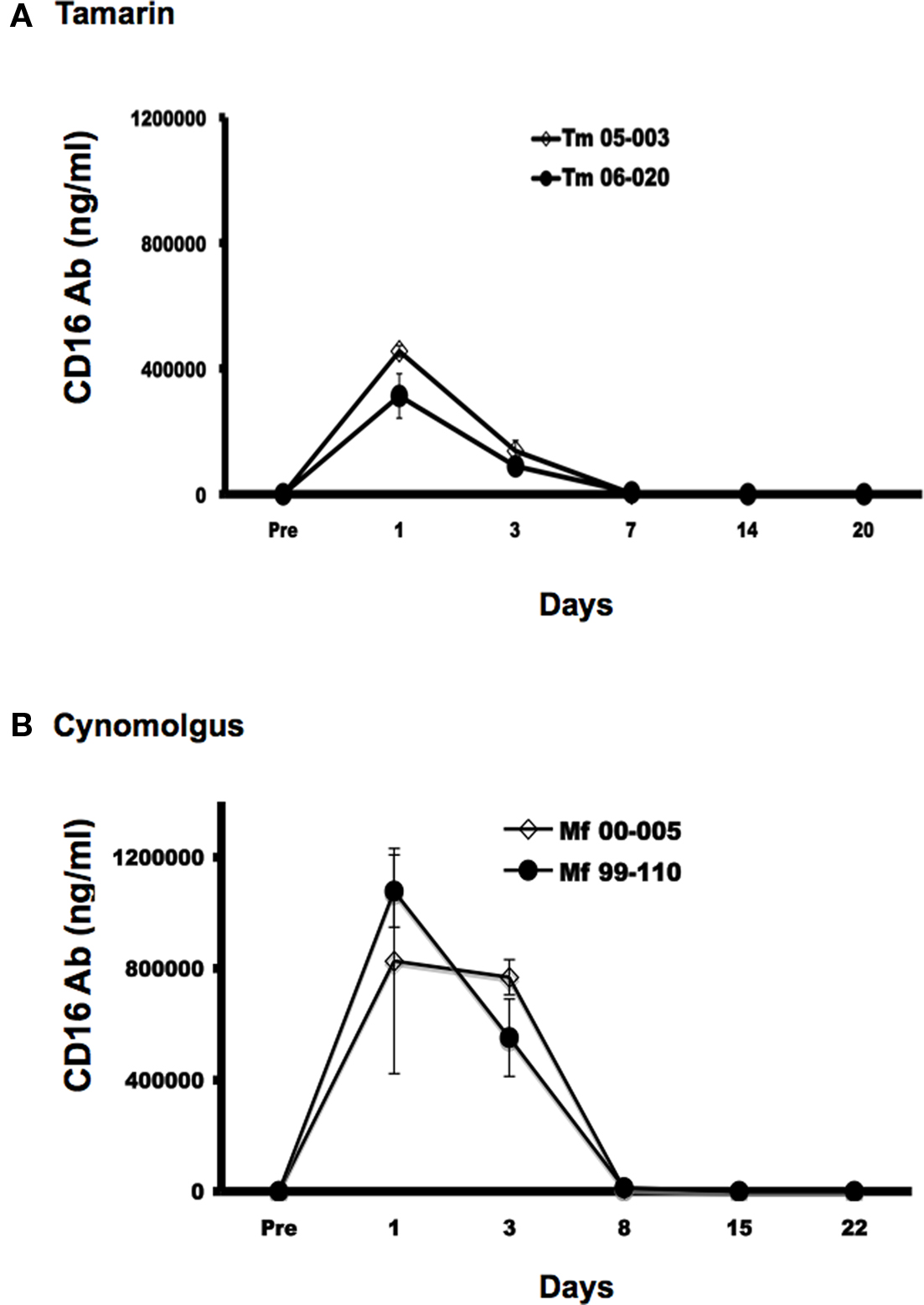
Figure 6. Kinetics of concentration of CD16 (3G8) mAb in vivo. The concentration of CD16 (3G8) mAb in plasma was measured by ELISA as described in Section “Materials and Methods”. Results shown are representative of three independent experiments. (A) Tamarins: Tm 05-003 and Tm 06-020. (B) Cynomolgus monkeys: Mf 00-005 and Mf 99-110.
Attenuation of Cd16+ NK Cell Function By in Vivo Depletion of CD16+ Cells
Finally, we tested whether depletion of the CD16+ subset could attenuate the NK activity in PBMCs. The killing activity was reduced at day 1 and the reduction persisted for 1 week post-treatment in the 3G8-treated monkeys (Figure 7). These results showed that the administration of the 3G8 mAb significantly influenced the number and activity of CD16+ lymphocytes in both tamarins and cynomolgus monkeys.
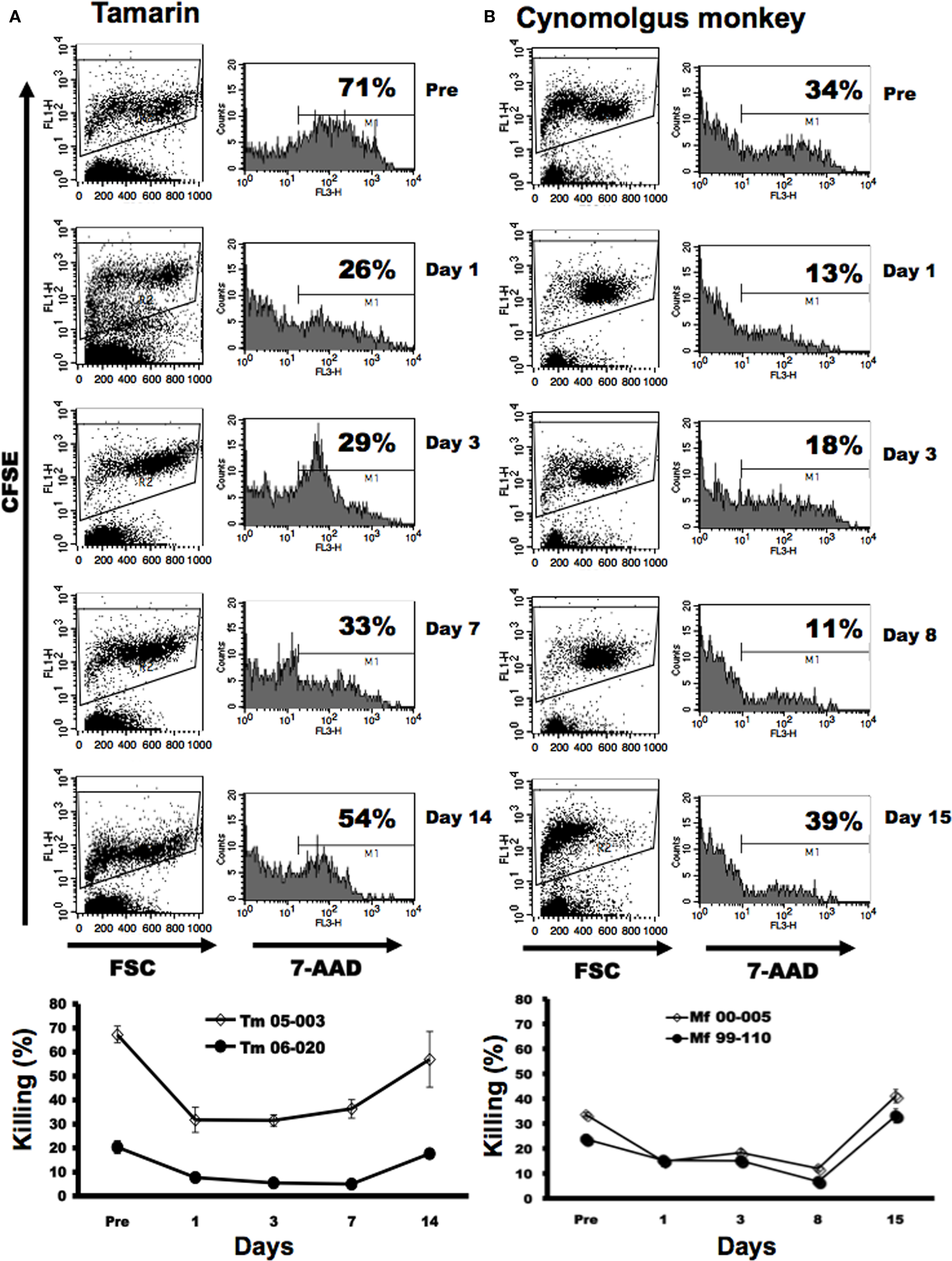
Figure 7. Attenuation of natural killer activity by in vivo depletion of CD16+ cells. (A,B) Tamarins and cynomolgus monkeys were administered 50 mg/kg of murine 3G8 mAb. The activity was determined in PBMCs of both monkeys. Tamarins: Tm 05-003 and Tm 06-020. (B) Cynomolgus monkeys: Mf 00-005 and Mf 99-110.
Discussion
In this study, we attempted to establish a technical basis for the study of NK cells in tamarins. First, we characterized the NK cells in tamarins and showed that the anti-CD16 (3G8) mAb, an NK marker, cross-reacted with the PBMCs (Figure 1). Second, we assessed the killing activity of the CD16+ NK cells in tamarins using our improved method (Figures 2–4) and demonstrated that CD16+ NK cells were likely to be a major population with the killing activity in tamarins. Finally, to directly examine the role of CD16+ NK cells in vivo, we assessed the effect of anti-CD16 (3G8) mAb in vivo. After administration of the mAb, CD16+ NK cells were completely depleted and the killing activity was substantially attenuated in the treated monkeys (Figures 5 and 7). Our results suggest that our method for depletion of CD16+ NK cells in vivo is useful for investigating the pivotal role of NK cells in the response against hepatitis viruses.
Generally it is impossible to monitor the immunological status in humans pre- and post-infection with hepatitis viruses and to deplete specific subsets such as NK cells in vivo. Non-human primates have immune systems similar to that of humans and are suitable for the evaluation of innate and adaptive immune responses against hepatitis viruses (Woollard et al., 2008). GBV-B is most closely related to HCV. Since experimental infection with GBV-B induces acute and chronic hepatitis in tamarins, this model may be useful for the study of antiviral immunity. Moreover, we have also been developing a chimeric virus between HCV and GBV-B. Therefore, if the HCV/GBV-B chimeric virus is able to infect and replicate in tamarins, our method for in vivo depletion of CD16+ NK cell in tamarins is very useful tool to understand the relationship between the chimeric virus and CD16+ NK cells. Moreover, it is still unclear whether NK cells might play a pivotal role at the acute or chronic phase in hepatitis. Analyses to address this issue are in progress using our GBV-B model.
Unexpectedly, almost complete in vivo depletion of CD16+ NK cells was not able to completely remove the NK activity in PBMCs, i.e., about one-third of the NK activity remained as compared with that before mAb treatment. This indicates that the CD16− subpopulation represents a substantial component of NK cells in monkeys. So far, mAbs recognizing tamarin’s NK-specific CD markers other than CD16, such as CD56 and CD159A (Choi et al., 2008), are not available. Further characterization and phenotyping of NK cells in tamarins will be necessary to selectively and totally deplete NK cells in vivo.
Interestingly, tamarins have been used for the study of experimental infection with Rabies virus, Epstein–Barr virus, Hepatitis A virus and Herpesvirus as well as GBV-B (Mackett et al., 1996; Batista-Morais et al., 2000; Purcell et al., 2002; de Thoisy et al., 2003; Martin et al., 2003; Takikawa et al., 2010). Therefore we hope that our system may be useful for examining the role of NK cells in the control of viral infection as well as to develop novel antiviral strategies.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to give special thanks to Dr. Hiroaki Shibata and members of Corporation for Production and Research of Laboratory Primates for technical assistance. Reagents used in this study were provided by the NIH Non-human Primate Reagent Resource (R24 RR016001, NIAID contract HHSN272200900037C). This work was supported by grants from the Ministry of Health, Labor and Welfare of Japan (to Hirofumi Akari) and Ministry of Education, Culture, Sports, Science and Technology of Japan (to Tomoyuki Yoshida and Hirofumi Akari) and Ministry of the Environment of Japan (to Tomoyuki Yoshida and Hirofumi Akari) and by Global COE Program A06 of Kyoto University.
References
Akari, H., Iwasaki, Y., Yoshida, T., and Iijima, S. (2009). Non-human primate surrogate model of hepatitis C virus infection. Microbiol. Immunol. 53, 53–57.
Akari, H., Terao, K., Murayama, Y., Nam, K. H., and Yoshikawa, Y. (1997). Peripheral blood CD4+ CD8+ lymphocytes in cynomolgus monkeys are of resting memory T lineage. Int. Immunol. 9, 591–597.
Batista-Morais, N., Neilson-Rolim, B., Matos-Chaves, H. H., de Brito-Neto, J., and Maria-da-Silva, L. (2000). Rabies in tamarins (Callithrix jacchus) in the state of Ceara, Brazil, a distinct viral variant? Mem. Inst. Oswaldo Cruz 95, 609–610.
Biron, C. A., Byron, K. S., and Sullivan, J. L. (1989). Severe herpesvirus infections in an adolescent without natural killer cells. N. Engl. J. Med. 320, 1731–1735.
Bukh, J., Apgar, C. L., and Yanagi, M. (1999). Toward a surrogate model for hepatitis C virus: an infectious molecular clone of the GB virus-B hepatitis agent. Virology 262, 470–478.
Cheent, K., and Khakoo, S. I. (2010). Natural killer cells and hepatitis C: action and reaction. Gut. doi:10.1136/gut.2010.212555. [Epub ahead of print].
Choi, E. I., Wang, R., Peterson, L., Letvin, N. L., and Reimann, K. A. (2008). Use of an anti-CD16 antibody for in vivo depletion of natural killer cells in rhesus macaques. Immunology 124, 215–222.
Cohen, J., and Lester, B. (2007). AIDS research. Trials of NIH’s AIDS vaccine get a yellow light. Science 318, 1852.
de Thoisy, B., Pouliquen, J. F., Lacoste, V., Gessain, A., and Kazanji, M. (2003). Novel gamma-1 herpesviruses identified in free-ranging new world monkeys (golden-handed tamarin [Saguinus midas], squirrel monkey [Saimiri sciureus], and white-faced saki [Pithecia pithecia]) in French guiana. J. Virol. 77, 9099–9105.
Deignan, T., Curry, M. P., Doherty, D. G., Golden-Mason, L., Volkov, Y., Norris, S., Nolan, N., Traynor, O., McEntee, G., Hegarty, J. E., and O’Farrelly, C. (2002). Decrease in hepatic CD56(+) T cells and V alpha 24(+) natural killer T cells in chronic hepatitis C viral infection. J. Hepatol. 37, 101–108.
Fleit, H. B., Wright, S. D., and Unkeless, J. C. (1982). Human neutrophil Fc gamma receptor distribution and structure. Proc. Natl. Acad. Sci. U.S.A. 79, 3275–3279.
Golden-Mason, L., Madrigal-Estebas, L., McGrath, E., Conroy, M. J., Ryan, E. J., Hegarty, J. E., O’Farrelly, C., and Doherty, D. G. (2008). Altered natural killer cell subset distributions in resolved and persistent hepatitis C virus infection following single source exposure. Gut 57, 1121–1128.
Ishii, K., Iijima, S., Kimura, N., Lee, Y. J., Ageyama, N., Yagi, S., Yamaguchi, K., Maki, N., Mori, K., Yoshizaki, S., Machida, S., Suzuki, T., Iwata, N., Sata, T., Terao, K., Miyamura, T., and Akari, H. (2007). GBV-B as a pleiotropic virus: distribution of GBV-B in extrahepatic tissues in vivo. Microbes Infect. 9, 515–521.
Kasai, M., Iwamori, M., Nagai, Y., Okumura, K., and Tada, T. (1980). A glycolipid on the surface of mouse natural killer cells. Eur. J. Immunol. 10, 175–180.
Lanford, R. E., Chavez, D., Notvall, L., and Brasky, K. M. (2003). Comparison of tamarins and marmosets as hosts for GBV-B infections and the effect of immunosuppression on duration of viremia. Virology 311, 72–80.
Lecoeur, H., Fevrier, M., Garcia, S., Riviere, Y., and Gougeon, M. L. (2001). A novel flow cytometric assay for quantitation and multiparametric characterization of cell-mediated cytotoxicity. J. Immunol. Methods 253, 177–187.
Lian, R. H., and Kumar, V. (2002). Murine natural killer cell progenitors and their requirements for development. Semin. Immunol. 14, 453–460.
Liu, Z. X., Govindarajan, S., Okamoto, S., and Dennert, G. (2000). NK cells cause liver injury and facilitate the induction of T cell-mediated immunity to a viral liver infection. J. Immunol. 164, 6480–6486.
Mackett, M., Cox, C., Pepper, S. D., Lees, J. F., Naylor, B. A., Wedderburn, N., and Arrand, J. R. (1996). Immunisation of common marmosets with vaccinia virus expressing Epstein–Barr virus (EBV) gp340 and challenge with EBV. J. Med. Virol. 50, 263–271.
Martin, A., Bodola, F., Sangar, D. V., Goettge, K., Popov, V., Rijnbrand, R., Lanford, R. E., and Lemon, S. M. (2003). Chronic hepatitis associated with GB virus B persistence in a tamarin after intrahepatic inoculation of synthetic viral RNA. Proc. Natl. Acad. Sci. U.S.A. 100, 9962–9967.
Navarro-Sánches, E., Desprès, P., and Cedillo-Barrón, L. (2005). Innate immune responses to dengue virus. Arch. Med. Res. 36, 425–435.
Orange, J. S. (2002). Human natural killer cell deficiencies and susceptibility to infection. Microbes Infect. 4, 1545–1558.
Purcell, R. H., Wong, D. C., and Shapiro, M. (2002). Relative infectivity of hepatitis A virus by the oral and intravenous routes in 2 species of non-human primates. J. Infect. Dis. 185, 1668–1671.
Robertson, M. J. (2002). Role of chemokines in the biology of natural killer cells. J. Leukoc. Biol. 71, 173–183.
Sbardellati, A., Scarselli, E., Verschoor, E., De Tomassi, A., Lazzaro, D., and Traboni, C. (2001). Generation of infectious and transmissible virions from a GB virus B full-length consensus clone in tamarins. J. Gen. Virol. 82, 2437–2448.
Takikawa, S., Engle, R. E., Faulk, K. N., Emerson, S. U., Purcell, R. H., and Bukh, J. (2010). Molecular evolution of GB virus B hepatitis virus during acute resolving and persistent infections in experimentally infected tamarins. J. Gen. Virol. 91, 727–733.
Tenorio, E. P., and Saavedra, R. (2005). Differential effect of sodium arsenite during the activation of human CD4+ and CD8+ T lymphocytes. Int. Immunopharmacol. 5, 1853–1869.
Keywords: CD16, cynomolgus monkey, tamarin, NK cell
Citation: Yoshida T, Saito A, Iwasaki Y, Iijima S, Kurosawa T, Katakai Y, Yasutomi Y, Reimann KA, Hayakawa T and Akari H (2010) Characterization of natural killer cells in tamarins: a technical basis for studies of innate immunity. Front. Microbio. 1:128. doi: 10.3389/fmicb.2010.00128
Received: 04 August 2010;
Accepted: 28 October 2010;
Published online: 08 December 2010.
Edited by:
Yasuko Yokota, National Institute of Infectious Diseases, JapanReviewed by:
Koji Ishii, National Institute of Infectious Diseases, JapanIkuo Shoji, Kobe University Graduate School of Medicine, Japan
Copyright: © 2010 Yoshida, Saito, Iwasaki, Iijima, Kurosawa, Katakai, Yasutomi, Reimann, Hayakawa and Akari. This is an open-access article subject to an exclusive license agreement between the authors and the Frontiers Research Foundation, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are credited.
*Correspondence: Hirofumi Akari, Primate Research Institute, Kyoto University, Inuyama, Aichi 484-8506, Japan. e-mail:YWthcmlAcHJpLmt5b3RvLXUuYWMuanA=