Abstract
Shiga-toxigenic Escherichia coli (STEC) O157:H7 uses a myriad of surface adhesive appendages including pili, flagella, and the type 3 secretion system (T3SS) to adhere to and inflict damage to the human gut mucosa. Consumption of contaminated ground beef, milk, juices, water, or leafy greens has been associated with outbreaks of diarrheal disease in humans due to STEC. The aim of this study was to investigate which of the known STEC O157:H7 adherence factors mediate colonization of baby spinach leaves and where the bacteria reside within tainted leaves. We found that STEC O157:H7 colonizes baby spinach leaves through the coordinated production of curli, the E. coli common pilus, hemorrhagic coli type 4 pilus, flagella, and T3SS. Electron microscopy analysis of tainted leaves revealed STEC bacteria in the internal cavity of the stomata, in intercellular spaces, and within vascular tissue (xylem and phloem), where the bacteria were protected from the bactericidal effect of gentamicin, sodium hypochlorite or ozonated water treatments. We confirmed that the T3S escN mutant showed a reduced number of bacteria within the stomata suggesting that T3S is required for the successful colonization of leaves. In agreement, non-pathogenic E. coli K-12 strain DH5α transformed with a plasmid carrying the locus of enterocyte effacement (LEE) pathogenicity island, harboring the T3SS and effector genes, internalized into stomata more efficiently than without the LEE. This study highlights a role for pili, flagella, and T3SS in the interaction of STEC with spinach leaves. Colonization of plant stomata and internal tissues may constitute a strategy by which STEC survives in a nutrient-rich microenvironment protected from external foes and may be a potential source for human infection.
Introduction
Human infections due to Shiga-toxigenic Escherichia coli (STEC) O157:H7 strains range from mild diarrhea to bloody diarrhea or hemorrhagic colitis, which can develop into the potentially fatal hemolytic uremic syndrome or HUS (Karmali et al., 1983; Riley et al., 1983). STEC is a significant dilemma in the realm of food-borne disease, exemplified in recent episodes of diarrheal disease in various regions of the world associated with consumption of contaminated food, water, milk, fruit juices, and leafy green vegetables such as alfalfa, sprouts, arugula, spinach, and lettuce (Michino et al., 1999; Sivapalasingam et al., 2004; Wendel et al., 2009). The precise mechanisms by which STEC is introduced into agricultural products are not fully understood. The bacteria live as commensal organisms in farm and several wild animals and may represent a major source of contamination of water and agricultural produce for human consumption (Solomon et al., 2002; Mootian et al., 2009). For example, recent outbreaks of diarrheal illness associated to spinach consumption were linked to contamination with feces from STEC-harboring wild pigs (CDC, 2006; Jay et al., 2007). Insects (e.g., filth flies) have been implicated as transmission vectors in plant infection by STEC (Talley et al., 2009).
The phyllosphere supplies an important bio-niche for a significant number and wide range of commensal bacterial species as well as for plant and human pathogens (Lindow and Brandl, 2003). Pathogen-associated molecular patterns (PAMPs) such as flagella and lipopolysaccharide (LPS) from Gram-negative bacteria interact with specific target receptors on guard cells to elicit an innate response resulting in the closure of stomata on the Arabidopsis leaf surface (Melotto et al., 2006). Illness-producing bacteria have evolved strategies to overcome plant defenses and to colonize the surface of leaves and deeper tissues of the phyllosphere. They use wounds and natural openings like stomata, hydathodes, and lenticels as natural gateways for passage into internal plant tissues where they grow and begin to cause disease (Hirano and Upper, 2000; Melotto et al., 2006). Pseudomonas syringae, a pathogen of hundreds of plant species utilizes its secretion (type 3 secretion system, T3SS) and an array of effector molecules to successfully suppress plant innate mechanisms of defense and kill its host (Chang et al., 2005; Chisholm et al., 2006). Listeria monocytogenes uses flagella for attachment to radish tissue and Klebsiella, Enterobacter, and Pseudomonas produce fimbriae that mediate attachment to plant surfaces (Korhonen et al., 1986; Vesper, 1987; Gorski et al., 2003). It was suggested that E. coli O157:H7 is able to exist as a component of the phyllosphere microbial community both as epiphytes on leaf surfaces and endophytes within leaf tissues. While some authors suggest that E. coli O157:H7 could be absorbed from soil to internal plant tissues, others suggest entry through plant roots (Wachtel et al., 2002; Islam et al., 2004; Jablasone et al., 2005; Sharma et al., 2009). E. coli O157:H7 can be located and is able to survive in the substomatal cavity and the intercellular spaces of the spongy of leafy greens (Itoh et al., 1998; Solomon et al., 2002; Warriner et al., 2003; Franz et al., 2007). The precise mechanisms of adherence of STEC to plant surfaces, the receptors recognized, and the types of physical interactions participating in this interplay are largely unknown.
Much is known about the mechanisms of interaction of STEC with human epithelial cells. STEC uses its T3SS to inject many effector proteins into intestinal cells that lead to remodeling of the cytoskeleton and consequently to the formation of intestinal attaching and effacing (AE) histopathological lesions. Most of the components required for AE are encoded in the genetic locus of enterocyte effacement (McDaniel and Kaper, 1997). It is well established that the interaction of STEC with human epithelial cells is multi-factorial as it involves the participation of the intimin–Tir complex, secreted proteins, and flagella. Several pili types including, but not limited to, the long polar fimbriae, F9, hemorrhagic coli pilus (HCP), curli, the E. coli laminin-binding fimbriae (ELF), and the E. coli common pilus (ECP), were associated with adherence to human and animal epithelial cells in vitro (Torres et al., 2002; Kaper et al., 2004; Low et al., 2006; Erdem et al., 2007; Rendon et al., 2007; Xicohtencatl-Cortes et al., 2007; Saldana et al., 2009; Samadder et al., 2009). It was reported that the EspA fiber, but not intimin, flagella or pili, was required for attachment of STEC to arugula leaves (Shaw et al., 2008). Recently, we suggested that the T3SS of STEC O157:H7 aids in the colonization of lettuce and spinach (Xicohtencatl-Cortes et al., 2009).
The aim of this study was focused on expanding our knowledge on the interaction of STEC with baby spinach leaves and the role of newly described surface adherence factors in colonization. This effort revealed that STEC uses several surface structures to colonize the leaf surface and gain access to internal tissues and in this way, establish a niche in the plant where bacteria are protected from environmental foes and industrial decontamination. This strategy would ensure survival in the environment and allow transmission to animal reservoirs and to the human host.
Materials and Methods
Bacterial strains and culture conditions
Strains used here are described in Table 1. Bacteria were routinely grown overnight at 37°C with aeration in Luria–Bertani (LB) broth or for induction of virulence factors in Dulbecco's modified Eagle's medium (DMEM) with 0.5% D-mannose (Invitrogen). Except for the intimin (eae), quorum sensing (qseB), and Tir (tir) mutants constructed here, all of the remaining mutants and complemented strains employed have been reported previously by our laboratories or other researchers.
Table 1
| Strains source | Notes | References or source |
|---|---|---|
| EDL933 | Wild type O157:H7 | Riley et al. (1983) |
| EDL933ΔescN | T3SS-ATPase escN::km | Jarvis and Kaper (1996) |
| EDL933Δtir | Tir mutant tir::km | This study |
| EDL933Δeae | Intimin mutant eae::km | This study |
| EDL933ΔespFu | espFu::cm | Campellone et al. (2004) |
| EDL933ΔespP | espP::km | Xicohtencatl-Cortes et al. (2010) |
| EDL933ΔfliC | fliC::km | Erdem et al. (2007) |
| EDL933ΔqseB | qseB::cm | This study |
| EDL933ΔhcpA | hcpA::km | Xicohtencatl-Cortes et al. (2007) |
| EDL933ΔecpA | ecpA::km | Rendon et al. (2007) |
| EDL933ΔelfA | elfA::km | Samadder et al. (2009) |
| EDL933ΔcsgA | csgA::km | Saldana et al. (2009) |
| EDL933ΔcsgA | pCsgA = csgBA | Saldana et al. (2009)(pCsgA) |
| in pBAD-Topo | ||
| EDL933ΔcsgD | csgD::km | Saldana et al. (2009) |
| EDL933ΔcsgD | pCP994 = csgD in | Saldana et al. (2009)(pCP994) |
| pKK233-2 | ||
| EDL933ΔbcsA | bcsA::cm | Saldana et al. (2009) |
| DH5α | K-12 strain | Our collection |
| DH5α (pLEE) | LEE plasmid | This study |
Bacterial strains and plasmids used in this study.
Construction of bacterial mutants
The tir, eae, and qseB genes in STEC O157:H7 strain EDL933 were deleted by the λ Red recombinase mutagenesis approach as described earlier (Datsenko and Wanner, 2000). The primers employed for mutagenesis are described in Table 2. The isogenic mutants were confirmed for the lost genotype by PCR. All mutant derivatives grew at the same rate as the wild-type strain under the conditions tested in this work.
Table 2
| Primer name | Gene | Function | Primer Sequence 5′ → 3′ |
|---|---|---|---|
| K1 f | km | Confirm mutagenesis | GTGTATTGACGGCGTTTATAACTGTGGTATGTGCA |
| ACGTCCAGCGTTATGGTGTAGGCTGGAGCTGCTTC | |||
| K2 r | km | Confirm mutagenesis | TGTTACCGCATTCACATTACCAGAAGTCGCTTTCC |
| CCGTCATATGAATATCCTCCTTAG | |||
| Htir-H1P1 f | tir | Mutagenesis | AATAAAAAGGAGATATTTATGCCTATTGGTAATCTTGGTCATTGTAGGCTGGAGCTGCTTCG |
| Htir-H2P2 r | tir | Mutagenesis | AAATGATTATGGATATATTTAGACGAAACGATGGGATCCCGGCATATGAATATCCTCCTTAG |
| Heae-H1P1f | eae | Mutagenesis | TTGTGGTGGAGCCATAACATGATTACTCATGGTTGTTATACCTGTAGGCTGGAGCTGCTTCG |
| Heae-H2P2 r | eae | Mutagenesis | GCCGGGGTGGTTATGGAATTATTCTACACAAACCGCATAGACCATATGAATATCCTCCTTAG |
| G250 f | qseB | Mutagenesis | TATCGCAGGGATGAAAAAATGCGAATTTTACTGATAGAAGTGTAGGCTGGAGCTGCTTC |
| G251r | qseB | Mutagenesis | CTCACCTAATGTGTAGCCAATACCATGCTCGGTACGAATCATATGAATATCCTCCTTAG |
| Htir f | tir | Confirm mutagenesis | GGTTGCTGCAATTTTATTTAT |
| Htir r | tir | Confirm mutagenesis | TGAAATAGATCCAATACCAATT |
| Heae f | eae | Confirm mutagenesis | AGAATGAAATAGAAGTCGTTG |
| Heae r | eae | Confirm mutagenesis | TTGTGAGCATAGTTGTTGCT |
| G254 f | qseB | Confirm mutagenesis | AGGGCCATTACTGCGATTAC |
| G255 r | qseB | Confirm mutagenesis | GAGCGTACTTAACCGCTTGG |
List of primers used in this study.
F, forward; r, reverse.
Spinach colonization assays
Fresh processed baby spinach leaves (cut into 1 cm-diameter pieces) were washed with sterile PBS and placed into 24-well plates containing DMEM with 0.5% D-mannose. The leaves were then infected with approximately 107 bacteria from an overnight culture grown in LB broth and incubated at 26°C for specific time periods (0–24 h) or as indicated in the text. The supernatant was then removed and the leaves were washed three times with sterile PBS to remove unbound bacteria. For quantitative assessment of bacterial adherence, spinach leaves infected with wild type or isogenic mutants were placed in 1.5 ml Eppendorf tubes containing glass beads in 1 ml of sterile PBS with 0.3% Triton X-100 and vortexed for 5 min. The supernatant was collected and 10-fold serial dilutions were plated out onto MacConkey-Sorbitol agar plates for determination of colony-forming units (CFUs) after overnight incubation (Xicohtencatl-Cortes et al., 2009). When required, the infected leaves where incubated for 2 h with gentamicin (200 μg/ml) to kill surface-exposed bacteria, and the bacteria inside the leaves tissues surviving this treatment, were counted as described above. All experiments were repeated at least three times in triplicate on different days and the results were expressed as percentage of adherence relative to that of the wild-type strain or as CFUs.
Electron microscopy studies
For transmission electron microscopy (TEM) infected and mock-infected leaves were washed as described above and fixed with fixative Trumps (Electron Microscopy Sciences). Fixed tissues were processed with the aid of a PelcoBioWave laboratory microwave (Ted Pella). The samples were washed in 0.1 M sodium cacodylate pH 7.2, post fixed with 2% OsO4, water, washed, and dehydrated in a graded ethanol series 25, 50, 75, 95, 100% followed by 100% acetone. Dehydrated samples were infiltrated in graded acetone/Spurs epoxy resin (30, 50, 70, 100%), and cured at 60°C for 48 h. Cured resin blocks were trimmed, thin sectioned, and collected on Formvar copper slot grids, post-stained with 2% aq. Uranyl acetate and Reynolds’ lead citrate. Sections were examined with a Hitachi H-7000 TEM (Hitachi High Technologies America) and digital images acquired with a Veleta 2k × 2k camera and iTEM software (Olympus Soft-Imaging Solutions Corp.). For scanning electron microscopy (SEM), infected and mock-infected leaves were fixed, dehydrated as mentioned above and critically point dried (Bal-Tec CPD030, Leica Microsystems). Samples were mounted on carbon adhesive tabs on aluminum specimen mount, Au/Pd sputter coated (DeskV, Denton Vacuum), and examined with high-resolution field-emission scanning electron microscope (S-4000, Hitachi High Technologies America).
Antisera
Rabbit polyclonal antibodies raised against STEC antigens, H7 flagella, O157 LPS, ECP, and curli, were available from previous studies and their specificity has been previously described (Rendon et al., 2007; Saldana et al., 2009; Xicohtencatl-Cortes et al., 2009).
Light and immunofluorescence microscopy
Infected leaves were washed three times with sterile PBS to remove unbound bacteria and then fixed with 2% formalin/PBS for 20 min. Peels of the lower surfaces of the leaves (epidermis abaxial) were observed by light microscopy to visualize bacteria associated with or within stomata openings as previously described (Melotto et al., 2006). For immunofluorescence microscopy (IFM), leaves fixed with 2% formalin, were incubated for 1 h with primary rabbit antibodies (listed above) followed by 1-h incubation with secondary anti-rabbit IgG Alexa-conjugated antibodies (Molecular Probes). The specimens were visualized under UV light and phase contrast using an Axio Imager1.0 Zeiss microscope.
Resistance to ozonated water and chlorinated solutions
To determine resistance to chlorine treatment, leaves of approximately the same size were placed in Petri dishes and were infected at 26°C for 9 h with approximately 107 bacteria from overnight cultures grown in DMEM. After the desired incubation period, the spinach leaves were subjected to 10 min-washes in 10-fold dilutions (0.1–10%) of sodium hypochlorite (chlorine), or 10–20 min exposure to ozonated water. Subsequent to this treatment, the spinach leaves were washed three times with sterile PBS, cut into 1 cm-diameter pieces, and placed in 1.5 ml Eppendorf tubes containing 1 ml of sterile PBS with 0.3% Triton X-100 and glass beads and processed as described above for bacterial counting (Xicohtencatl-Cortes et al., 2009). Statistical analysis was done using Student's t-test. The significance level was 5% in all tests (*P < 0.05). The GraphPad software was used.
Results
Role of flagella and T3SS in leaf colonization
We sought to investigate the importance of these surface structures in leaf adherence and colonization. Kinetics studies of spinach infections performed between 0 and 8 h assisted by IFM using specific anti-H7 antibodies revealed the compelling presence of flagella on bacteria adhering to the leaf epidermis. Progressive and increasing amounts of flagella H7 were observed correlating with the increasing number of bacteria adhering with respect to time of incubation (Figure 1). SEM analysis of wild type versus a genetically modified flagella (non-motile) mutant supported these observations (Figure 1G). Comparative quantitative experiments showed that the flagella (fliC) and T3SS (escN) mutants adhered significantly less than the parental strain confirming a role of these appendages in spinach colonization (Figure 2A). We sought to determine if the intimin–Tir complex was important for spinach colonization. This analysis showed that intimin (eae) and Tir (tir) mutants were not significantly abrogated in adherence suggesting that the intimin–Tir interaction is not crucial or necessary for leaf colonization, as it is for human cells. Since the EspP cytotoxin was suggested to play a role in bovine colonization and forms adhesive macrostructures (Dziva et al., 2007; Xicohtencatl-Cortes et al., 2010), we included this mutant in our analysis. The espP mutant was not significantly affected in leaf colonization (Figure 2A).
Figure 1
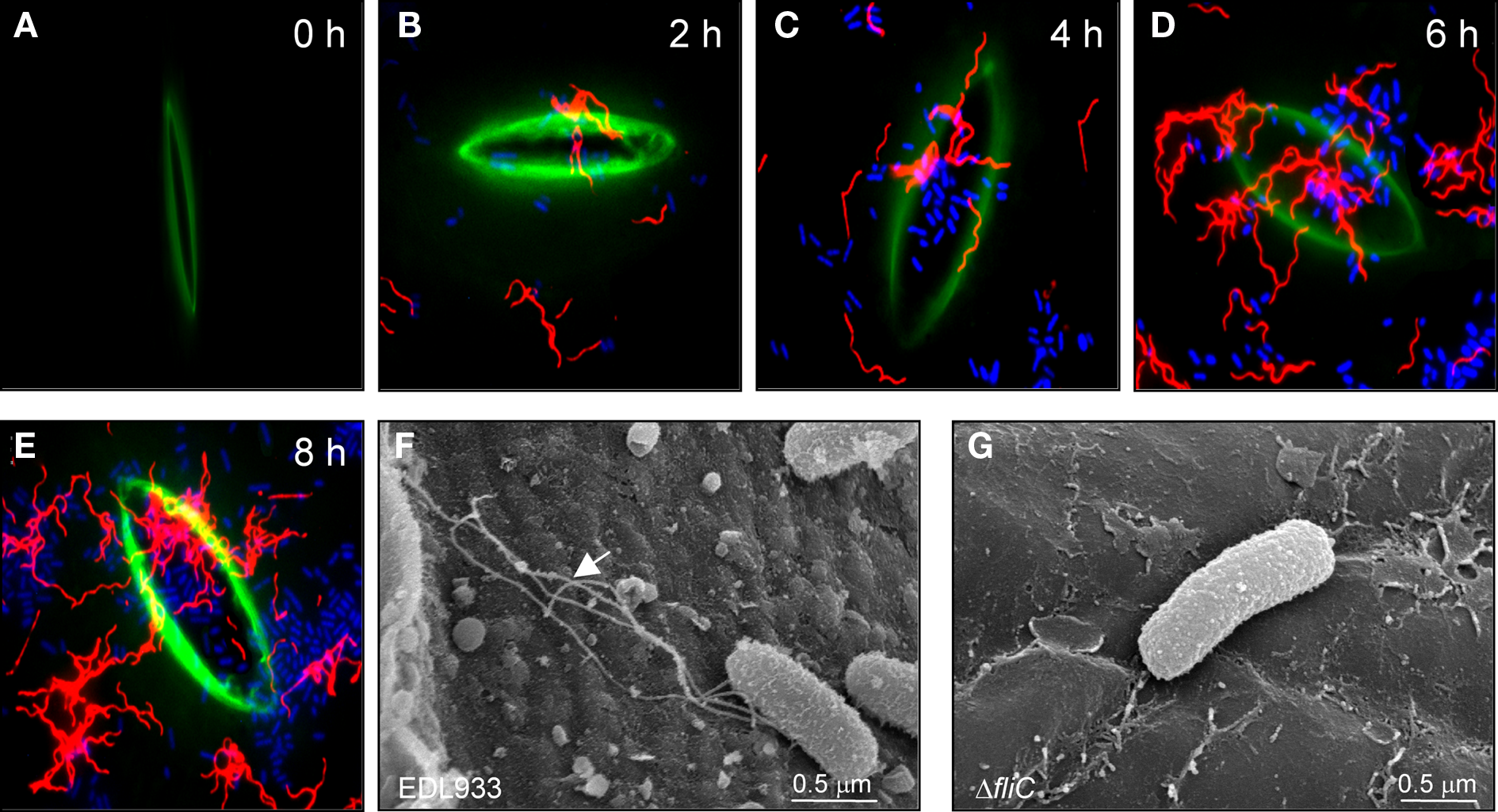
Time-course visualization of flagella on STEC infecting spinach leaves. (A–E), Leaves infected with STEC for 0, 2, 4, 6, and 8 h were processed for IFM using anti-H7 antibodies. Flagella are shown in red, bacteria in blue, and the stomata in green. Images were taken at 60X. (F) Scanning electron micrograph showing EDL933 adhering onto the leaf epidermis and displaying flagella (arrow). (G) The fliC mutant did not show the structures seen in (F).
Figure 2
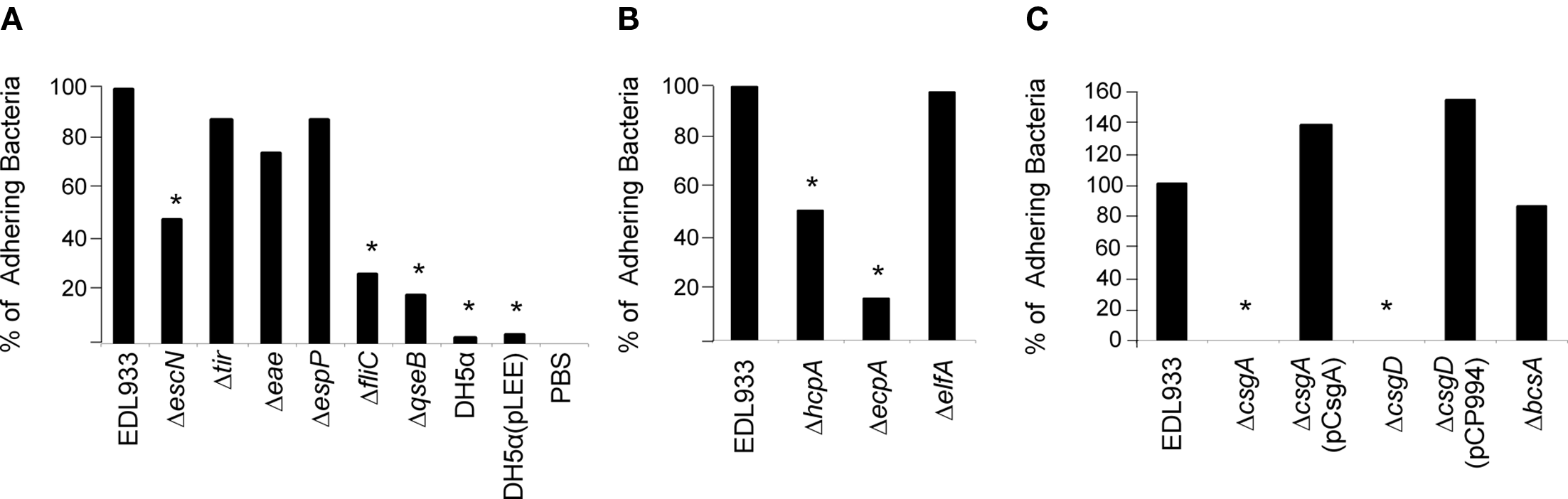
Comparative analysis of spinach colonization by STEC strains after 9 h of infection. (A) EDL933 and isogenic mutants in escN (T3SS), tir (intimin receptor), eae (intimin), espP (toxin/adhesin), fliC (flagellin), and qseB (quorum sensing gene). (B,C) EDL933 and isogenic mutants in pilin genes: hcpA, ecpA, elfA, csgA, csgD (curli regulator), or bcsA (cellulose). All of the strains were used to infect spinach leaves as described in the text. Serial dilutions of homogenized samples were plated out onto MacConkey-Sorbitol plates for bacterial counting. These data represent the average of three experiments repeated on different days in triplicate. *P > 0.05 statistically significant with respect to the wild-type strain.
Role of quorum sensing in leaf colonization
Quorum-sensing regulates many virulence-associated functions in bacteria such as flagellation, motility, and T3S (Sperandio et al., 2001). We found that a quorum-sensing mutant (ΔqseB) was reduced in adherence by 80% as compared to the wild-type strain (Figure 2A) and this result correlates with the observation that the flagella (fliC) and T3SS (escN) mutants were significantly hampered (72 and 52% reduction, respectively) in spinach colonization.
STEC pili types involved in the colonization of baby spinach leaves
The specific adherence molecular mechanisms that mediate phyllosphere colonization by STEC remain elusive. Hence, we investigated the role of recently described surface factors of STEC in adherence to spinach leaves. To this aim, isogenic strains carrying mutations in genes encoding for HCP, ECP, ELF, curli, and cellulose were tested for leaf adherence in comparative quantitative experiments. We found that the csgA and csgD (99% reduction), ecpA (83% reduction), and hcpA (49% reduction) mutants were significantly hampered in adherence to baby spinach leaves in -comparison to the parental strain EDL933 (Figures 2B,C). Notably, when the csgA and csgD mutants were complemented with plasmids (pCsgA and pCP994 respectively) that over express curli (Saldana et al., 2009), the resulting strains attached more than the wild-type strain to the spinach leaves (Figure 2C). The remaining ELF and cellulose mutants were not deficient in spinach colonization (Figure 2C). The presence of ECP and curli on the bacteria colonizing the leaf epidermis was demonstrated by IFM using specific antibodies against these pili structures (Figure 3). Mutants in curli and ECP served as negative controls for production of these pili. In all, the IFM data compellingly indicate that flagella, ECP, and curli are present during the interaction of the colonizing bacteria with the spinach leaves and participate collectively in adherence and colonization.
Figure 3
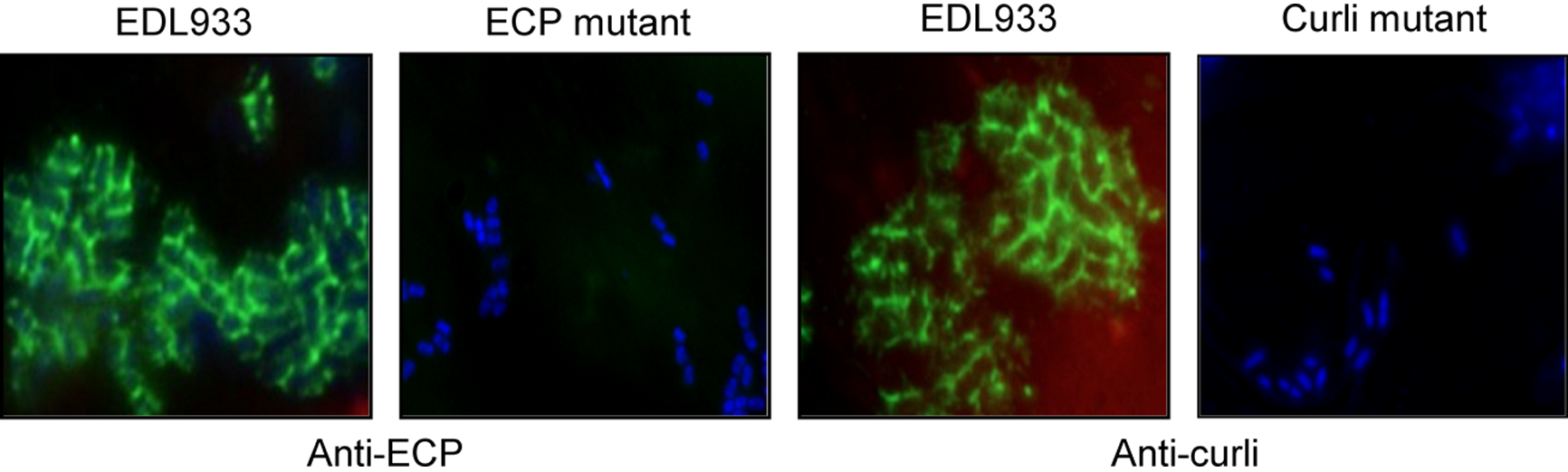
Demonstration of ECP and curli on adhering bacteria. Leaves infected with wild-type strain, ECP or curli mutants (bacteria in blue) were immunostained with specific antibodies against ECP and curli (pili in green). Note the presence of these pili only on EDL933.
Localization of STEC within leaf tissues
Previous studies have shown STEC bacteria on leaf surfaces, on and around stomata, and between cellular spaces and xylem (Itoh et al., 1998; Solomon et al., 2002; Warriner et al., 2003; Franz et al., 2007). We sought to further investigate the ability of STEC to reside within the stomata internal cavity and internal tissues. SEM analysis of spinach leaves incubated from 1 to 24 h with EDL933 revealed the presence of increasing number of bacteria on the stomata and within its internal cavity (Figures 4A–F). Scores of bacteria tethered by filamentous appendages resembling flagella were noted (Figure 4B). Accompanying IFM experiments using anti-O157 LPS antibody showed compelling localization of STEC O157 bacteria on stomata and internalized within stomata cavities (Figures 4G–J). Further, analysis of cross sections of infected leaves confirmed the presence of STEC bacteria within stomata internal cavities, -intercellular spaces of the spongy mesophyll, and notably in the vascular tissue (xylem and phloem; Figure 5).
Figure 4
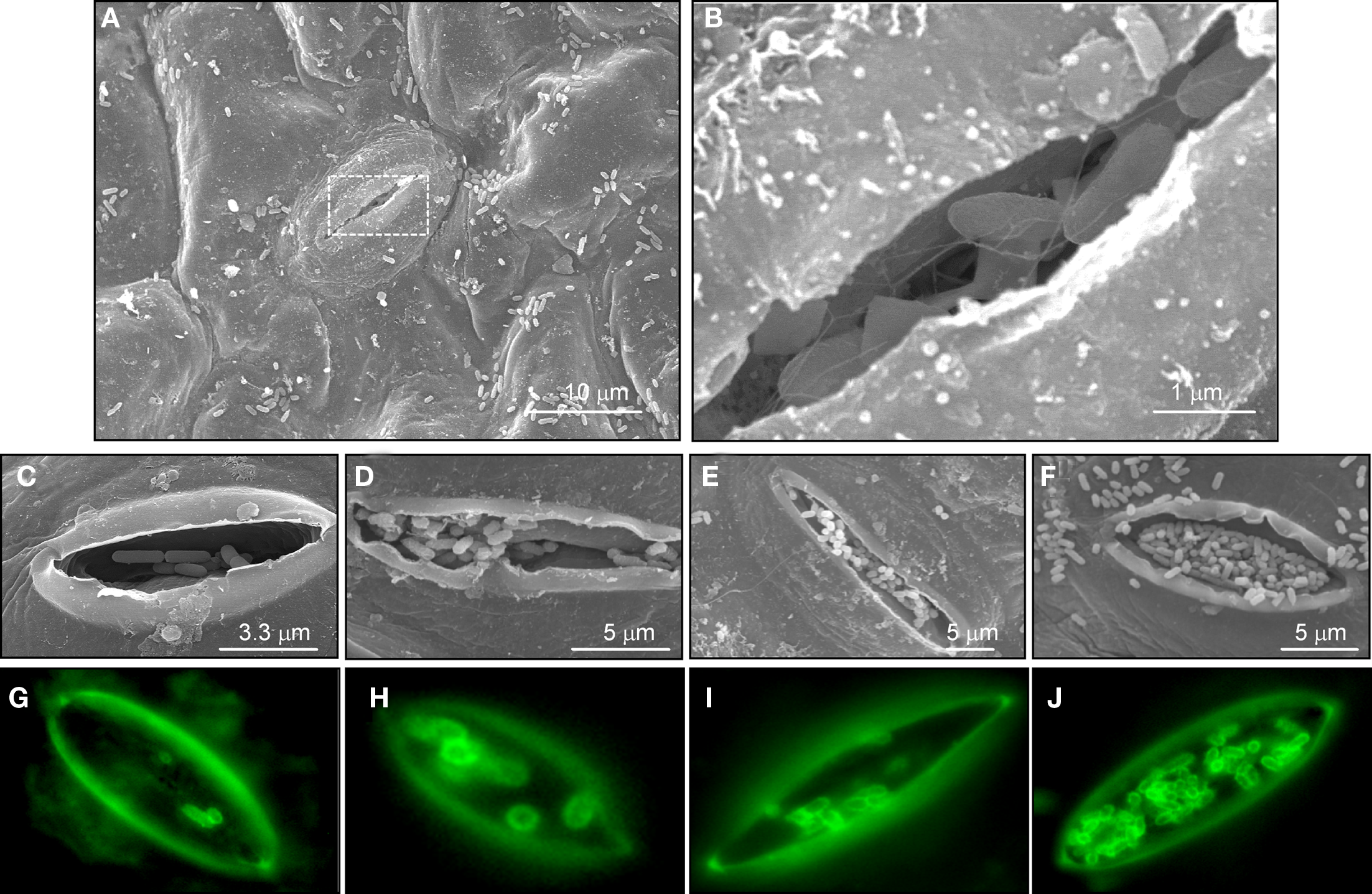
Evidence of STEC in stomata. (A) Scanning electron micrograph showing bacteria on leaf epidermis at 6 h of infection. (B) High magnification of boxed area in (A) showing flagellate bacteria internalized in the stomata. (C–F) Micrographs (60X) of time-course EDL933 infection experiments between 3, 6, 12, and 24 h showing progressive association of bacteria with stomata. (G–J) Same experiment as before employing IFM and anti-0157 antibodies to stain bacteria (green).
Figure 5
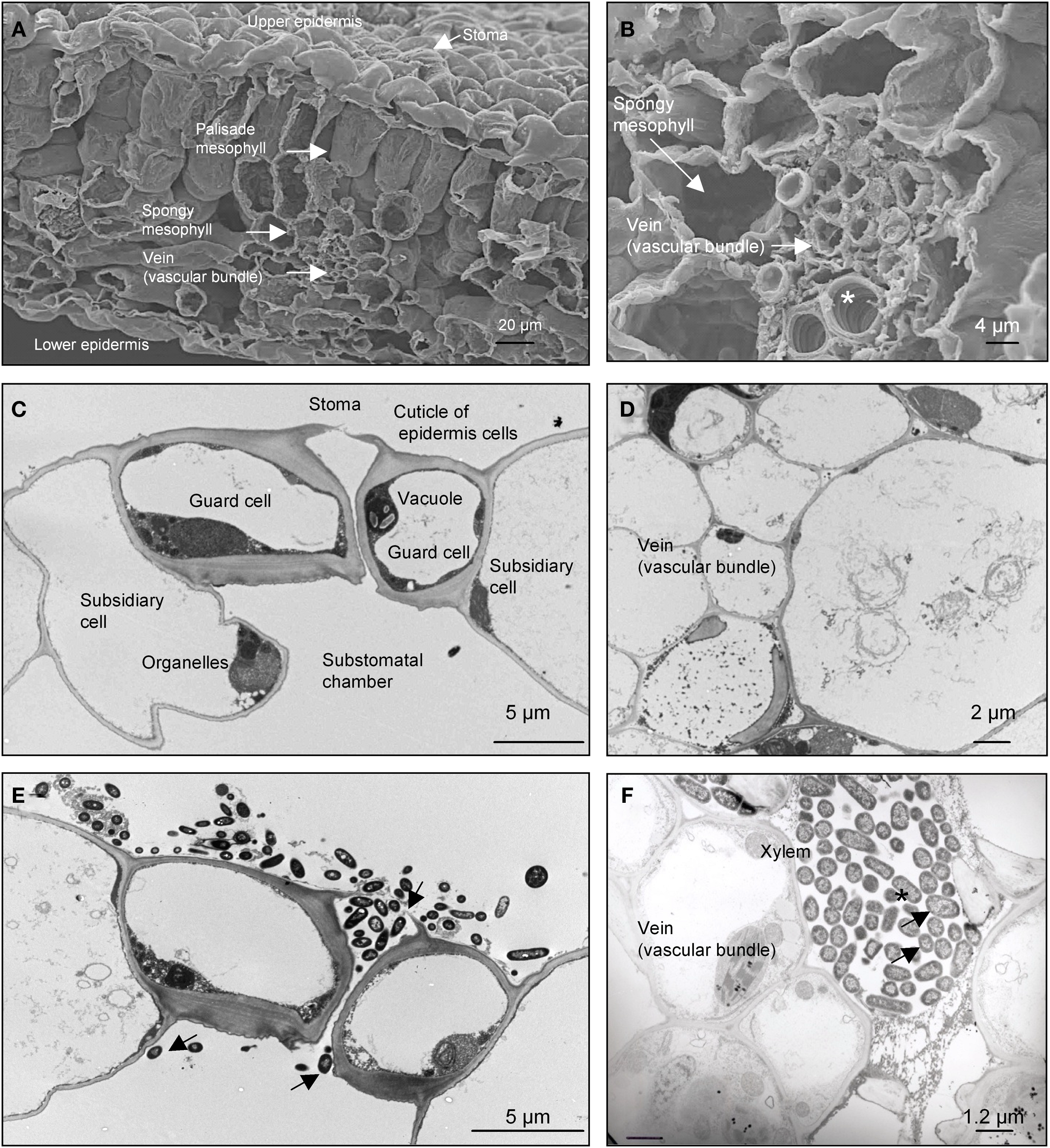
Evidence of STEC present in stomata chambers and in leaf vein. (A) SEM image showing a cross section of uninfected leaf. (B) Close up of the internal tissue of the leaf showing the vascular bundle and spongy mesophyll structures. (C,D) TEM images showing cross sections of uninfected leaf after 24 h in DMEM. Anatomical sites are indicated with arrows. (E) Cross section of leaf infected with EDL933 (arrows) for 24 h revealing colonization on the cuticle of the epidermis, stomata, and the substomatal chamber. (F) Cross section showing internal structures of the spinach leaf and colonization of the vessel elements of the xylem with STEC (arrows).
Based on these observations, we hypothesized a role for T3SS in leaf internalization. Thus, we then compared the ability of EDL933 versus the escN mutant to penetrate the leaf surface using the gentamicin protection assay. We found that EDL933 survived at a rate of 8.3% after gentamicin treatment, while the escN mutant showed 6.1% survival (data not shown). A more clear involvement of T3SS was seen when E. coli K-12 DH5α and DH5α carrying the LEE region, which encodes the T3SS on a plasmid (pLEE), were included in the assay. Leaves infected with DH5α (pLEE) showed 3.2-fold-increase with respect to the DH5α strain (data not shown). Other workers have shown STEC and enteroaggregative E. coli adhering to the leaf epidermis, and on and around stomata but not beyond the substomal chamber (Berger et al., 2009, 2010; Xicohtencatl-Cortes et al., 2009). Based on previous observations and our own, we hypothesized that E. coli O157:H7 are able to provide for themselves with a safe, nutrient-rich niche in the womb of stomata, internal tissues, and intercellular spaces of the leaf, where they are able to replicate and survive, protected from environmental foes.
Leaf internalization provides protection against bactericidal agents
Is stomata colonization a “hide-n-sick” mechanism by which STEC evades industrial decontamination of agricultural products for human consumption? In an attempt to address this important question we subjected experimentally STEC-tainted spinach leaves to decontamination treatments with 0.1–10% of hypochlorite solutions and also to exposure to ozonated water. First, we showed that 10% sodium hypochlorite (equivalent to 6,500 ppm) and 125 ppm of ozonated water killed 100 and 99.9% of the bacteria, respectively (Figures 6B,D). Second, we subjected STEC-infected leaves to the same treatments after which bacteria were still recovered (Figures 6A,C). Although these anti-bacterial agents reduced the bacterial population on the leaves by ∼50% as compared to the mock control, it is clear that a striking number of bacteria residing in the stomata resisted and survived these bactericidal treatments. A reasonable explanation for these results is that internalization of bacteria into stomata and internal compartments of the leaf protected bacteria from bactericidal agents such as antibiotics (e.g., gentamicin), chlorine or ozone.
Figure 6
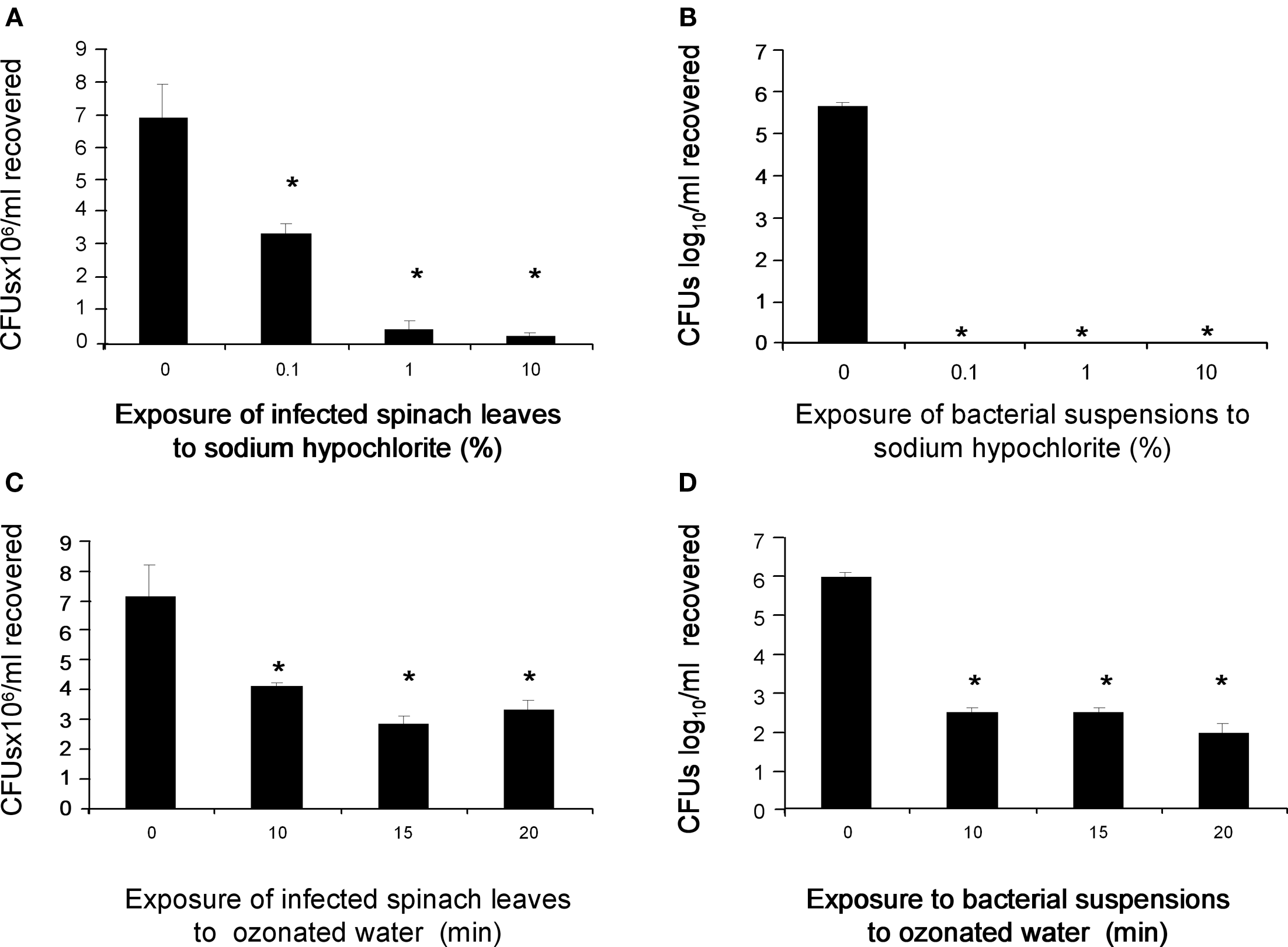
Survival of STEC within stomata to different decontamination processes. (A) STEC infected-spinach leaves for 9 h were treated for 10 min with 0–10% chlorine solutions and then plated out onto MacConkey-Sorbitol agar. Note that even at 10% of chlorine wash, the bacteria were able to grow suggesting that they are protected within the stomata. (B) A suspension of 106 bacteria was treated for 10 min with 0–10% chlorine solutions. (C) Treatment of infected leaves for 9 h with and without ozonated water. (D) A suspension of 106 bacteria was treated with and without ozonated water from 0 to 20 min. These data represent the average of three experiments repeated on different days in triplicate. *P > 0.05 statistically significant.
Discussion
The present study was initiated to address important questions concerning the interaction of STEC with spinach leaves. Specifically, we wanted to identify which of the adhesive factors of STEC, known to be important for human and animal intestinal epithelial cell infection, are also engaged in colonization and survival of these bacteria in baby Spinacia oleracea leaves. Several lines of evidence, obtained through different experimental approaches support the notion of the participation of flagella in baby spinach leaf colonization. We demonstrated in time-course experiments assisted by high-resolution SEM and IFM that the bacteria multiply and colonize the leaf surface and that flagella are produced progressively. In support of these observations, we found that an EDL933 fliC (flagella) isogenic mutant exhibited significant decreased attachment to spinach leaves. In line with our data, Carey et al. (2009) found that fliC was up-regulated during storage of Romaine lettuce at 15°C. It is apparent the flagella display other functions than motility.
A unifying theme in animal and plant bacterial pathogens is the production of specialized secretion mechanisms for direct delivery of effector proteins into host cells to inflict damage. Plant pathogens such as Pseudomonas and Agrobacterium employ flagella, pili, T3SS or T4SS to infect and cause phytopathology, although the nature of their receptors remains unknown (Korhonen et al., 1986; Rahme et al., 1995; McCullen and Binns, 2006). Flagellation and T3S of STEC are regulated by a double component (qseBC) quorum-sensing system (Sperandio et al., 2001). We found that STEC escN (T3SS-ATPase) and qseB mutants were significantly impaired in leaf colonization with respect to the parental strain, indicating that the T3SS and cell-to-cell communication are important in the interaction of STEC with plant cells. While the intimin–Tir complex is crucial for attachment and the characteristic development of actin-rich processes on host cells, neither intimin nor Tir seemed to be required for leaf adherence. This was somewhat expected given the striking differences in chemical composition between human and plant tissue surfaces.
We sought to investigate if pili and cellulose were required for leaf colonization. Comparative adherence studies employing isogenic pili mutants revealed that curli, ECP, and HCP were required for spinach leaf adherence while ELF and cellulose were apparently not. It is well established that pili, in particular curli, are highly hydrophobic appendages that neutralize negative charges on host cells facilitating adherence (Collinson et al., 1991). Recently, it was shown that the level of adherence of E. coli O157:H7 strains to cabbage and iceberg lettuce surfaces correlated with the amount of curli produced and their hydrophobicity (Patel et al., 2011). The demonstration that several redundant adhesive factors are required for spinach leaf colonization provides an immediate conceptual framework to explain the multiple molecular mechanisms of plant infection by a human pathogen. Future studies will be needed to elucidate the nature of the plant receptors involved in the recognition of STEC fimbrial adhesins.
Plant pathogens are able to breach into internal tissues using wounds and natural openings of the phyllosphere (Hirano and Upper, 2000; Melotto et al., 2006), but the molecular mechanisms employed to penetrate through stomata, as a natural gateway for passage into internal tissues and cause phytopathology are unknown. Compelling data from ultrastructural, genetic, and functional studies described here clearly showed the presence of STEC O157 within stomata and electron microscopy analysis of leaf cross sections revealed the presence of aggregates of bacteria in internal compartments of the leaf such stomata chambers, intercellular spaces, xylem (specifically in vessel elements), and phloem. Experiments designed to evaluate the ability of STEC to survive within spinach leaves upon treatment with sodium hypochlorite solutions and ozonated water showed that live bacteria could be recovered suggesting that the internal spaces of spinach leaves protect bacteria against these bactericidal agents. We showed that the T3SS is involved in internalization and survival of STEC in spinach leaves. Transformation of a non-pathogenic E. coli with pLEE (T3SS+) rendered the bacteria capable of adhering to the epidermis, internalizing into leaf tissue allowing themselves to survive the bactericidal treatments. It is possible that STEC possess mechanisms that direct them toward stomata (e.g., through chemotaxis, motility, quorum sensing) and induce guard cells to open these gateways. The low-infectious dose of STEC (10–50 organisms) and its ability to colonize the stoma and internal leaf compartments provide logical explanations by which the bacteria survive man-driven decontamination treatments and are able to cause outbreaks of STEC infections associated with consumption of tainted-agricultural products. Thus, infected leafy greens may represent potential vehicles of infection and transmission to humans. In conclusion, our research shows that STEC, a human pathogen, which uses several pili types, flagella and the T3SS to colonize human intestinal tissue, can also employ these mechanisms to gain access to plant leaf surface, internal tissues and intercellular spaces where the bacteria survive in a protected microenvironment before causing disease.
Statements
Acknowledgments
This work was supported by a grant from Fresh Express Produce and by Grant Number AI66012 from NIAID, NIH to Jorge A. Girón and its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NIH. Jose Luis Puente was supported by CONACyT grant No. 42918Q. We thank Alejandra Vázquez for technical assistance, John M. Leong for kindly providing EDL933ΔespFu strain, James B. Kaper for plasmid pLEE, and Karen Kelley and Kim Backer-Kelley at the Interdisciplinary Center for Biotechnology Research, Electron Microscopy and Bio-Imaging, University of Florida for assistance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1
Berger C. N. Shaw R. K. Ruiz-Perez F. Nataro J. P. Henderson I. R. Pallen M. J. Frankel G. (2009). Interaction of enteroaggregative Escherichia coli with salad leaves. Environ. Microbiol. Rep.1, 234–239.
2
Berger C. N. Sodha S. V. Shaw R. K. Griffin P. M. Pink D. Hand P. Frankel G. (2010). Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ. Microbiol.12, 2385–2397.
3
Campellone K. G. Robbins D. Leong J. M. (2004). EspFU is a translocated EHEC effector that interacts with Tir and N-WASP and promotes Nck-independent actin assembly. Dev. Cell7, 217–228.
4
Carey C. M. Kostrzynska M. Thompson S. (2009). Escherichia coli O157:H7 stress and virulence gene expression on Romaine lettuce using comparative real-time PCR. J. Microbiol. Methods77, 235–242.
5
CDC. (2006). Ongoing multistate outbreak of Escherichia coli serotype O157:H7 infections associated with consumption of fresh spinach – United States, September 2006. MMWR Morb. Mortal. Wkly. Rep.55, 1045–1046.
6
Chang J. H. Urbach J. M. Law T. F. Arnold L. W. Hu A. Gombar S. Grant S. R. Ausubel F. M. Dangl J. L. (2005). A high-throughput, near-saturating screen for type III effector genes from Pseudomonas syringae. Proc. Natl. Acad. Sci. U.S.A.102, 2549–2554.
7
Chisholm S. T. Coaker G. Day B. Staskawicz B. J. (2006). Host-microbe interactions: shaping the evolution of the plant immune response. Cell124, 803–814.10.1016/j.cell.2006.02.008
8
Collinson S. K. Emody L. Muller K. H. Trust T. J. Kay W. W. (1991). Purification and characterization of thin, aggregative fimbriae from Salmonella enteritidis. J. Bacteriol.173, 4773–4781.
9
Datsenko K. A. Wanner B. L. (2000). One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U.S.A.97, 6640–6645.
10
Dziva F. Mahajan A. Cameron P. Currie C. McKendrick I. J. Wallis T. S. Smith D. G. E. Stevens M. P. (2007). EspP, a Type V-secreted serine protease of enterohaemorrhagic Escherichia coli O157:H7, influences intestinal colonization of calves and adherence to bovine primary intestinal epithelial cells. FEMS Microbiol. Lett.271, 258–264.
11
Erdem A. L. Avelino F. Xicohtencatl-Cortes J. Giron J. A. (2007). Host protein binding and adhesive properties of H6 and H7 flagella of attaching and effacing Escherichia coli. J. Bacteriol.189, 7426–7435.10.1128/JB.00464-07
12
Franz E. Visser A. A. Van Diepeningen A. D. Klerks M. M. Termorshuizen A. J. van Bruggen A. H. (2007). Quantification of contamination of lettuce by GFP-expressing Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium. Food Microbiol.24, 106–112.10.1016/j.fm.2006.03.002
13
Gorski L. Palumbo J. D. Mandrell R. E. (2003). Attachment of Listeria monocytogenes to radish tissue is dependent upon temperature and flagellar motility. Appl. Environ. Microbiol.69, 258–266.
14
Hirano S. S. Upper C. D. (2000). Bacteria in the leaf ecosystem with emphasis on Pseudomonas syringae-a pathogen, ice nucleus, and epiphyte. Microbiol. Mol. Biol. Rev.64, 624–653.
15
Islam M. Doyle M. P. Phatak S. C. Millner P. Jiang X. (2004). Persistence of enterohemorrhagic Escherichia coli O157:H7 in soil and on leaf lettuce and parsley grown in fields treated with contaminated manure composts or irrigation water. J. Food Prot.67, 1365–1370.
16
Itoh Y. Sugita-Konishi Y. Kasuga F. Iwaki M. Hara-Kudo Y. Saito N. Noguchi Y. Konuma H. Kumagai S. (1998). Enterohemorrhagic Escherichia coli O157:H7 present in radish sprouts. Appl. Environ. Microbiol.64, 1532–1535.
17
Jablasone J. Warriner K. Griffiths M. (2005). Interactions of Escherichia coli O157:H7, Salmonella typhimurium and Listeria monocytogenes plants cultivated in a gnotobiotic system. Int. J. Food Microbiol.99, 7–18.10.1016/j.ijfoodmicro.2004.06.011
18
Jarvis K. G. Kaper J. B. (1996). Secretion of extracellular proteins by enterohemorrhagic Escherichia coli via a putative type III secretion system. Infect. Immun.64, 4826–4829.
19
Jay M. T. Cooley M. Carychao D. Wiscomb G. W. Sweitzer R. A. Crawford-Miksza L. Farrar J. A. Lau D. K. O'Connell J. Millington A. Asmundson R. V. Atwill E. R. Mandrell R. E. (2007). Escherichia coli O157:H7 in feral swine near spinach fields and cattle, central California coast. Emerging Infect. Dis.13, 1908–1911.
20
Kaper J. B. Nataro J. P. Mobley H. L. (2004). Pathogenic Escherichia coli. Nat. Rev. Microbiol.2, 123–140.
21
Karmali M. A. Petric M. Lim C. Fleming P. C. Steele B. T. (1983). Escherichia coli cytotoxin, haemolytic-uraemic syndrome, and haemorrhagic colitis. Lancet2, 1299–1300.10.1016/S0140-6736(83)91167-4
22
Korhonen T. K. Haahtela K. Romantschuk M. Bamford D. H. (1986). “Role of fimbriae and pili in the attachment of Klebsiella, Enterobacter, and Pseudomonas to plant surfaces,” in Recognition in Microbe-Plant Symbiotic and Pathogenic Interactions, ed. LugtenbergB. (Heidelberg: Springer-Verlag), 229–241.
23
Lindow S. E. Brandl M. T. (2003). Microbiology of the phyllosphere. Appl. Environ. Microbiol.69, 1875–1883.
24
Low A. S. Dziva F. Torres A. G. Martinez J. L. Rosser T. Naylor S. Spears K. Holden N. Mahajan A. Findlay J. Sales J. Smith D. G. Low J. C. Stevens M. P. Gally D. L. (2006). Cloning, expression, and characterization of fimbrial operon F9 from enterohemorrhagic Escherichia coli O157:H7. Infect. Immun.74, 2233–2244.
25
McCullen C. A. Binns A. N. (2006). Agrobacterium tumefaciens and plant cell interactions and activities required for interkingdom macromolecular transfer. Annu. Rev. Cell Dev. Biol.22, 101–127.
26
McDaniel T. K. Kaper J. B. (1997). A cloned pathogenicity island from enteropathogenic Escherichia coli confers the attaching and effacing phenotype on E. coli K-12. Mol. Microbiol.23, 399–407.
27
Melotto M. Underwood W. Koczan J. Nomura K. He S. Y. (2006). Plant stomata function in innate immunity against bacterial invasion. Cell126, 969–980.10.1016/j.cell.2006.06.054
28
Michino H. Araki K. Minami S. Takaya S. Sakai N. Miyazaki M. Ono A. Yanagawa H. (1999). Massive outbreak of Escherichia coli O157:H7 infection in schoolchildren in Sakai City, Japan, associated with consumption of white radish sprouts. Am. J. Epidemiol.150, 787–796.
29
Mootian G. Wu W. H. Matthews K. R. (2009). Transfer of Escherichia coli O157:H7 from soil, water, and manure contaminated with low numbers of the pathogen to lettuce plants. J. Food Prot.72, 2308–2312.
30
Patel J. Sharma M. Ravishakar S. (2011). Effect of curli expression and hydrophobicity of Escherichia coli O157:H7 on attachment to fresh produce surfaces. J. Appl. Microbiol.110, 737–745.
31
Rahme L. G. Stevens E. J. Wolfort S. F. Shao J. Tompkins R. G. Ausubel F. M. (1995). Common virulence factors for bacterial pathogenicity in plants and animals. Science268, 1899–1902.10.1126/science.7604262
32
Rendon M. A. Saldana Z. Erdem A. L. Monteiro-Neto V. Vazquez A. Kaper J. B. Puente J. L. Girón J. A. (2007). Commensal and pathogenic Escherichia coli use a common pilus adherence factor for epithelial cell colonization. Proc. Natl. Acad. Sci. U.S.A.104, 10637–10642.
33
Riley L. W. Remis R. S. Helgerson S. D. McGee H. B. Wells J. G. Davis B. R. Hebert R. J. Olcott E. S. Johnson L. M. Hargrett N. T. Blake P. A. Cohen M. L. (1983). Hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med.308, 681–685.
34
Saldana Z. Xicohtencatl-Cortes J. Avelino F. Phillips A. D. Kaper J. B. Puente J. L. Giron J. A. (2009). Synergistic role of curli and cellulose in cell adherence and biofilm formation of attaching and effacing Escherichia coli and identification of Fis as a negative regulator of curli. Environ. Microbiol.11, 992–1006.
35
Samadder P. Xicohtencatl-Cortes J. Saldana Z. Jordan D. Tarr P. I. Kaper J. B. Giron J. A. (2009). The Escherichia coli ycbQRST operon encodes fimbriae with laminin-binding and epithelial cell adherence properties in Shiga-toxigenic E. coli O157:H7. Environ. Microbiol.11, 1815–1826.
36
Sharma M. Ingram D. T. Patel J. R. Millner P. D. Wang X. Hull A. E. Donnenberg M. S. (2009). A novel approach to investigate the uptake and internalization of Escherichia coli O157:H7 in spinach cultivated in soil and hydroponic medium. J. Food Prot.72, 1513–1520.
37
Shaw R. K. Berger C. N. Feys B. Knutton S. Pallen M. J. Frankel G. (2008). Enterohemorrhagic Escherichia coli exploits EspA filaments for attachment to salad leaves. Appl. Environ. Microbiol.74, 2908–2914.
38
Sivapalasingam S. Friedman C. R. Cohen L. Tauxe R. V. (2004). Fresh produce: a growing cause of outbreaks of foodborne illness in the United States, 1973 through 1997. J. Food Prot.67, 2342–2353.
39
Solomon E. B. Yaron S. Matthews K. R. (2002). Transmission of Escherichia coli O157:H7 from contaminated manure and irrigation water to lettuce plant tissue and its subsequent internalization. Appl. Environ. Microbiol.68, 397–400.
40
Sperandio V. Torres A. G. Giron J. A. Kaper J. B. (2001). Quorum sensing is a global regulatory mechanism in enterohemorrhagic Escherichia coli O157:H7. J. Bacteriol.183, 5187–5197.
41
Talley J. L. Wayadande A. C. Wasala L. P. Gerry A. C. Fletcher J. DeSilva U. Gilliland S. E. (2009). Association of Escherichia coli O157:H7 with filth flies (Muscidae and Calliphoridae) captured in leafy greens fields and experimental transmission of E. coli O157:H7 to spinach leaves by house flies (Diptera: Muscidae). J. Food Prot.72, 1547–1552.
42
Torres A. G. Giron J. A. Perna N. T. Burland V. Blattner F. R. Avelino-Flores F. Kaper J. B. (2002). Identification and characterization of lpfABCC'DE, a fimbrial operon of enterohemorrhagic Escherichia coli O157:H7. Infect. Immun.70, 5416–5427.
43
Vesper S. J. (1987). Production of Pili (Fimbriae) by Pseudomonas fluorescens and correlation with attachment to corn roots. Appl. Environ. Microbiol.53, 1397–1405.
44
Wachtel M. R. Whitehand L. C. Mandrell R. E. (2002). Association of Escherichia coli O157:H7 with preharvest leaf lettuce upon exposure to contaminated irrigation water. J. Food Prot.65, 18–25.
45
Warriner K. Ibrahim F. Dickinson M. Wright C. Waites W. M. (2003). Interaction of Escherichia coli with growing salad spinach plants. J. Food Prot.66, 1790–1797.
46
Wendel A. M. Johnson D. H. Sharapov U. Grant J. Archer J. R. Monson T. Koschmann C. Davis J. P. (2009). Multistate outbreak of Escherichia coli O157:H7 infection associated with consumption of packaged spinach, August-September 2006: the Wisconsin investigation. Clin. Infect. Dis.48, 1079–1086.
47
Xicohtencatl-Cortes J. Chacón E. S. Saldaña Z. Freer E. Girón J. A. (2009). Interaction of Escherichia coli O157:H7 with Leafy Green ProduceJ. Food Prot.72, 1531–1537.
48
Xicohtencatl-Cortes J. Monteiro-Neto V. Ledesma M. A. Jordan D. M. Francetic O. Kaper J. B. Puente J. L. Girón J. A. (2007). Intestinal adherence associated with type IV pili of enterohemorrhagic Escherichia coli O157:H7. J. Clin. Invest.117, 3519–3529.10.1172/JCI30727
49
Xicohtencatl-Cortes J. Saldana Z. Deng W. Castaneda E. Freer E. Tarr P. I. Finlay B. B. Puente J. L. Girón J. A. (2010). Bacterial macroscopic rope-like fibers with cytopathic and adhesive properties. J. Biol. Chem.285, 32336–32342.
Summary
Keywords
plant colonization, stomata, STEC, O157:H7, pathogenesis, spinach, T3SS, pili
Citation
Saldaña Z, Sánchez E, Xicohtencatl-Cortes J, Puente JL and Girón JA (2011) Surface Structures Involved in Plant Stomata and Leaf Colonization by Shiga-Toxigenic Escherichia Coli O157:H7. Front. Microbio. 2:119. doi: 10.3389/fmicb.2011.00119
Received
01 September 2010
Accepted
12 May 2011
Published
27 May 2011
Volume
2 - 2011
Edited by
Adel M. Talaat, University of Wisconsin Madison, USA
Reviewed by
Michael Konkel, Washington State University, USA; Jay C. D. Hinton, Trinity College Dublin, Ireland
Copyright
© 2011 Saldaña, Sánchez, Xicohtencatl-Cortes, Puente and Girón.
This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: Jorge A. Girón, Department of Molecular Genetics and Microbiology, Emerging Pathogens Institute, University of Florida, 2055 Mowry Road, PO Box 100009, Gainesville, FL, 32610, USA. e-mail: jagiron@ufl.edu
This article was submitted to Frontiers in Cellular and Infection Microbiology, a specialty of Frontiers in Microbiology.
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.