


- 1 Department of Microbiology and Immunology, Dalhousie University, Halifax, NS, Canada
- 2 Division of Infectious Diseases, Department of Medicine, Dalhousie University, Halifax, NS, Canada
- 3 Laboratory of Intracellular Parasites, National Institute of Allergy and Infectious Diseases, National Institute of Health Rocky Mountain Laboratories, Hamilton, MT, USA
The Legionella pneumophila chaperonin, high temperature protein B (HtpB), was discovered as a highly immunogenic antigen, only a few years after the identification of L. pneumophila as the causative agent of Legionnaires’ disease. As its counterparts in other bacterial pathogens, HtpB did not initially receive further attention, particularly because research was focused on a few model chaperonins that were used to demonstrate that chaperonins are essential stress proteins, present in all cellular forms of life and involved in helping other proteins to fold. However, chaperonins have recently attracted increasing interest, particularly after several reports confirmed their multifunctional nature and the presence of multiple chaperonin genes in numerous bacterial species. It is now accepted that bacterial chaperonins are capable of playing a variety of protein folding-independent roles. HtpB is clearly a multifunctional chaperonin that according to its location in the bacterial cell, or in the L. pneumophila-infected cell, plays different roles. HtpB exposed on the bacterial cell surface can act as an invasion factor for non-phagocytic cells, whereas the HtpB released in the host cell can act as an effector capable of altering organelle trafficking, the organization of actin microfilaments and cell signaling pathways. The road to discover the multifunctional nature of HtpB has been exciting and here we provide a historical perspective of the key findings linked to such discovery, as well as a summary of the experimental work (old and new) performed in our laboratory. Our current understanding has led us to propose that HtpB is an ancient protein that L. pneumophila uses as a key molecular tool important to the intracellular establishment of this fascinating pathogen.
Background
Chaperonins and Their Essential Protein Folding Function
Chaperonins are a family of structurally and functionally conserved, essential proteins, present in virtually all prokaryotic and eukaryotic forms of life. Intuitively, then, contemporary chaperonins must be related to one of the first proteins present in the common ancestor of all organisms currently known. The striking amino acid sequence and structural conservation of the chaperonin groups clearly suggests that these proteins must be very important. The primary function of chaperonins, recognized to be important enough to explain their essential nature, is in helping other proteins to fold properly and reach their native (functional) state.
Because this review is focused on the protein folding-independent functions of the Legionella chaperonin, a discussion on the protein folding ability of chaperonins is not forthcoming. Therefore, we provide the following key references for the benefit of those with further interests in this topic (Braig et al., 1994; Lund, 1995; Houry et al., 1999; Kerner et al., 2005; Sigler et al., 1998). In particular, recent comprehensive reviews that cover various aspects of the fascinating structure, biochemistry, and physiology of these formidable protein folding molecular machines (or nanoboxes in which proteins can fold) are those of England et al. (2008), Horwich et al. (2007), Lin and Rye (2006), and Lund (2011).
Classification of Chaperonins
It seems that Hemmingsen et al. (1988) were the first to coin the term “chaperonins” to describe a small group of related proteins involved in “post-translational assembly of oligomeric protein structures.” Since then, investigators have recognized the existence of different chaperonin types, which are currently classified into two groups based on their structure and evolutionary origin. Group I chaperonins are found in bacteria and in endosymbiotic organelles of eukaryotes (e.g., mitochondria and chloroplasts), have a mass of ∼60-kDa and are typically induced under stress, e.g., heat shock. Therefore, group I chaperonins are also known as heat shock proteins 60 (Hsp60s; Zeilstra-Ryalls et al., 1991). These proteins form homo-oligomeric rings that consist of seven chaperonin subunits (Braig et al., 1994). Two of these 7-mer rings come together to form the 14-mer barrel complex that mediates protein folding in association with a third homo-oligomeric ring, comprised of seven subunits of co-chaperonin, a protein of ∼10-kDa also known as Hsp10. Association with the 10-kDa co-chaperonins is an exclusive feature of Group I chaperonins. Other designations for Hsp10/Hsp60, are GroES/GroEL, Cpn10/Cpn60, and HtpA/HtpB. The intensively investigated Escherichia coli GroEL chaperonin constitutes the paradigm of Group I chaperonins.
Group II chaperonins, also known as TriC (TCP-1 ring complex) or CCT (chaperonin-containing TCP-1), are found in archaea, and the cytoplasm of eukaryotes (Lund, 1995). Group II chaperonins form eight- or nine-membered hetero-oligomeric rings with subunits that may have different masses (Kim et al., 1994; Klumpp and Baumeister, 1998). CCTs mediate the specialized folding of proteins (many of which are linked to the cytoskeleton), but do not team with 10 kDa co-chaperonins, although the protein prefoldin (Ohtaki et al., 2010) has been identified as a co-chaperone for CCTs. Group II chaperonins have an extended apical domain thought to cap the central cavity of the double-ringed complex, which replaces the need for the 7-mer co-chaperonin ring of Group I chaperonins (Fenton et al., 1996; Gutsche et al., 1999; Horwich and Saibil, 1998). Group II chaperonins are heterogeneous and are thought to have evolved by gene duplication and subsequent mutation (Archibald et al., 2000). While conserved within their respective groups, Group I and Group II chaperonins are only distantly related, but thought to share a common protein ancestor (Gupta, 1995).
A third chaperonin group has been recently reported in bacteria (Techtmann and Robb, 2010). Its representative chaperonin is that of the bacterium Carboxydothermus hydrogenoformans, which forms a 16-mer structure capable of refolding denatured proteins in an ATP-dependent manner. Group III chaperonins are distantly related to both Group I and Group II chaperonins, and thus they might represent an ancient horizontal transfer event from archaea to bacteria.
Protein Folding-Independent Functions of Group I Chaperonins
The Hsp60 of the bacterial endosymbiont Buchnera aphidicola (also called symbionin) acts as a histidine kinase (Morioka et al., 1994), whereas the GroEL of symbiotic Enterobacter aerogenes is a potent insect toxin (Yoshida et al., 2001), and the chaperonin of Mycobacterium leprae, is a protease (Portaro et al., 2002). Two views could be advanced to explain this functional diversity. In the first view, functional diversity is a preserved characteristic of chaperonins. That is, Group I chaperonins started as jacks-of-all-trades and gradually evolved toward specialization in protein folding. Thus, the contemporary examples of diversity mentioned above, represent evolutionary remnants of original functions preserved after specialization. In the second view, functional diversity is a newly emerged characteristic. That is, ancient chaperonins started as specialized proteins that gradually evolved toward functional diversity.
Two cases of functional chaperonin diversity resulting from few amino acid changes seem to favor the second view of “newly emerged functions.” Only 11 amino acids are different between the toxic chaperonin from endosymbiotic E. aerogenes, and the non-toxic chaperonin of E. coli, of which four amino acid positions are critical for toxicity. When the non-toxic E. coli chaperonin was engineered at the four critical residues to resemble the E. aerogenes chaperonin, it too became a potent insect toxin (Yoshida et al., 2001). In the case of the Hsp65 chaperonin of M. leprae, only three amino acids (Thr-375, Lys-409, and Ser-502) comprise the threonine catalytic group responsible for protease activity (Portaro et al., 2002).
In a recent article based on the analysis of 669 complete bacterial genomes, Lund proposed that one of the mechanisms responsible for functional diversity in Group I chaperonins relies on gene duplication followed by unconstrained mutation of the duplicated gene sequences (Lund, 2009). The analysis showed that 467/669 genomes contained a single chaperonin gene, 183/669 genomes contained multiple chaperonin genes (from 2 to a maximum of 7), and 13 Mycoplasma genomes contained no discernable chaperonin genes. Lund (2009) thus argued that the essential protein folding needs of a bacterial cell are met by a single chaperonin (whose gene would be constrained for change), while the other chaperonins would be free to mutate and acquire functional specializations. At least in the case of Mycobacterium tuberculosis, this notion has been experimentally substantiated. M. tuberculosis has two chaperonin genes encoding the chaperonins Cpn60.1 and Cpn60.2, where cpn60.2 is essential whereas cpn60.1 can be deleted from the genome (Hu et al., 2008). These two chaperonins are functionally different (Cehovin et al., 2010) supporting the idea of functional diversity afforded by gene duplication.
However, there are other cases in which functional diversity rests on a single chaperonin. As it will be discussed below in detail, one of these cases is the chaperonin of Legionella pneumophila. Other examples include those bacterial pathogens that typically use their chaperonins as adherence factors, or immune-modulators. In this capacity, chaperonins have been recently added to the list of “moonlighting” proteins (Jeffery, 2009). The term moonlighting is defined in the Webster’s Dictionary of the English Language as “working at a job in addition to one’s regular one,” and was introduced in the biochemical field to describe those proteins that perform a well-recognized function by day (regular job in a given environment or cellular location), and a not so obvious yet important function by night (other jobs in a different environment or cellular location).
Actinobacillus actinomycetemcomitans (Goulhen et al., 1998; Paju et al., 2000), Borrelia burgdorferi (Scorpio et al., 1994), Chlamydia spp. (Lund, 2009), Clostridium difficile (Hennequin et al., 2001), Helicobacter pylori (Huesca et al., 1996), Haemophilus ducreyi (Frisk et al., 1998), Listeria monocytogenes (Trost et al., 2005), and Salmonella enterica sv. Typhimurium (Ensgraber and Loos, 1992), are but some examples of bacterial pathogens that display their chaperonin in extracytoplasmic locations, and where the surface-associated, periplasmic, or released/secreted chaperonin seems to play alternate functional roles. For instance, the chaperonin of some of the aforementioned pathogens acts as an adhesion factor, but there are many that interact with mammalian cell surface receptors to initiate signaling events that result in cytokine production (reviewed by Ranford et al., 2000), phosphorylation of signaling molecules (Zhang et al., 2001, 2004), or other physiological outputs (Galdiero et al., 1997).
Group I chaperonins of endosymbiotic organelles are also functionally diverse, but given the nature of our review and its focus on a bacterial pathogen, we will not discuss here organellar chaperonins. Therefore, readers interested in the prominent role of chaperonins in immunity and autoimmunity are referred to a recent scholar review (Henderson, 2010) that includes details on the immune-modulatory ability of these proteins. In summary, chaperonins are ancient proteins, essential for the life of eukaryotic and prokaryotic cells. Their essential nature seemingly rests on their protein folding ability, but in several cases chaperonins appear to be multifunctional.
The Chaperonin of Legionella pneumophila, HtpB
The remaining portion of this review will be devoted to a discussion of the L. pneumophila chaperonin as a multifunctional (“moonlighting”) protein (Figure 1), including a presentation of our recent experimental findings. To facilitate the distinction between the chaperonins that we will be discussing, and to respect current nomenclature, the L. pneumophila chaperonin will be subsequently referred to as high temperature protein B (for HtpB). The designation HtpA is used for the L. pneumophila co-chaperonin, which is encoded by the first gene in the L. pneumophila htpAB operon. The chaperonin/co-chaperonin system of E. coli will be referred to as GroEL/GroES. Other chaperonins will be referred to as Hsp60 or Cpn60.
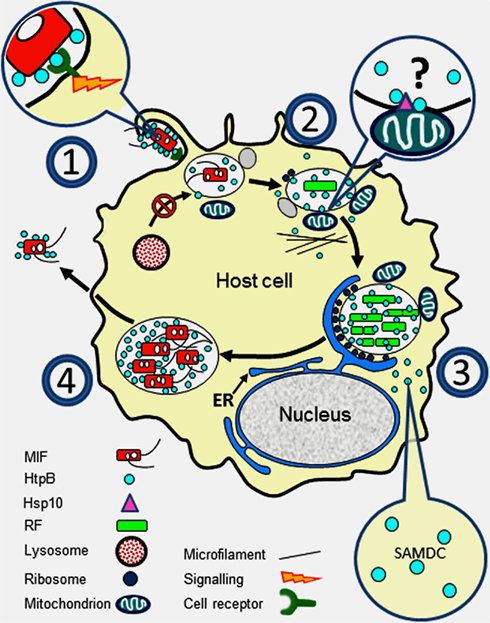
Figure 1. Surface exposed or released HtpB accompanies L. pneumophila along its growth cycle in host cells. (1) Extracellular L. pneumophila upregulates expression of HtpB in the presence of host cells (see Links Between HtpB and L. pneumophila Virulence), and the interaction of surface-exposed HtpB with cell receptors (Inset 1) triggers a signal leading to internalization (see Surface-Exposed HtpB Acts as an Invasion Factor). (2) Internalized legionellae associate with ER-derived vesicles, attracts mitochondria, and inhibit fusion with lysosomes. HtpB bound to beads is sufficient to mimic the last two events (see Surface-Exposed HtpB Alters Organelle Traffic). HtpB reaches the cytoplasm of the host cell where it could alter the actin cytoskeleton (Inset 2). The mechanism by which HtpB attracts mitochondria is unknown, but alteration of actin fibers and tethering via mitochondrial Hsp10 could be involved (see HtpB in the Eukaryotic Cytoplasm has Several Protein Targets). (3) During replication, released HtpB accumulates in the LCV from which it could reach the host cell cytoplasm (see Links Between HtpB and L. pneumophila Virulence and HtpB is Found in Extracytoplasmic Locations). Inset 3: HtpB in the cytoplasm of host cells (mammalian and amebal) interacts with SAMDC to potentially increase the intracellular pool of polyamines (see HtpB in the eukaryotic cytoplasm has several protein targets). (4) As L. pneumophila differentiates into MIFs, the amount of HtpB associated with the cell envelope and bacterial cell surface increases (see HtpB is Found in Extracytoplasmic Locations). As the LCV ruptures, large amounts of HtpB are likely released together with MIFs. Immunomodulatory effects (see Immunological Studies with HtpB) can be triggered by HtpB at any stage of the cycle. Key: ER, endoplasmic reticulum; RF, replicative form; MIF, mature infectious form; SAMDC, S-adenosyl methionine decarboxylase.
Historical Perspective of HtpB Research before 1998
Discovery and initial characterization
Between the mid-1980s and early 1990s, a number of publications reported the existence of a common antigen of about 60-kDa in many bacterial species. Sompolinsky et al. (1980a,b) referred to it as the “common antigen,” and Yamaguchi et al. (1989) used the term “cross-reacting protein antigen.” These antigens were eventually identified as chaperonins. Similarly, HtpB was first spotted as a 58-kDa common antigen cross-reactive with 60-kDa antigens from several Legionella species and other bacteria (Sampson et al., 1986; Plikaytis et al., 1987). This antigen prominently reacted with sera from patients diagnosed with Legionnaires’ disease (LD) and was used to confirm, by serology, culture-positive cases of LD (Sampson et al., 1986). This study also showed that when a rabbit serum raised against L. pneumophila serogroup 1 was pre-absorbed with whole L. pneumophila Philadelphia-1 cells, the 58-kDa antigen was no longer recognized by immunoblot. This is an interesting result because implies that the common antigen was surface exposed on the whole L. pneumophila cells used for cross-absorption. Plikaytis et al. (1987) were the first to purify HtpB and raise a rabbit hyperimmune serum against the purified protein, and shortly after, Pau et al. (1988) reported an optimized method for the purification of HtpB. A modification of this optimized method, which involves a combination of ammonium sulfate precipitation, size-exclusion, and ion-exchange chromatography, is the one used in the Garduño lab for the purification of HtpB.
We will close this section by mentioning that Gabay and Horwitz (1985) characterized HtpB as the major cytoplasmic membrane protein of L. pneumophila. Their studies are important because they established the ability of HtpB to interact with the bacterial cytoplasmic membrane, a trait that we believe is important in both the translocation of HtpB into the L. pneumophila periplasm (see HtpB is Found in Extracytoplasmic Locations below), and across the Legionella-containing vacuole (LCV) membrane into the host cell cytosol (see HtpB is Found in Extracytoplasmic Locations and Intracellularly Released HtpB Alters the Actin Cytoskeleton of Host Cells).
Monoclonal antibodies and the unique epitopes of HtpB
Several monoclonal antibodies were raised against HtpB once it was available as a purified protein (Helsel et al., 1988; Sampson et al., 1991; Steinmetz et al., 1991). These early monoclonal antibodies demonstrated that HtpB possesses epitopes cross-reactive with many other Group I chaperonins, as well as HtpB-specific epitopes. Monoclonal antibody GW2X4B8B2H6 (Helsel et al., 1988) does not cross-react with many Group I chaperonins (except for a few, including the Bordetella Cpn60), and recognizes the C-terminus of HtpB (Hoffman et al., 1989). We have widely used this antibody to monitor expression of recombinant HtpB. Monoclonal antibody 2125 (Steinmetz et al., 1991) is highly specific for HtpB and does not cross-react with any other bacterial chaperonin tested. Therefore, 2125 has been used as a tool for the rapid identification of Legionella spp. (Steinmetz et al., 1992). But our interest here is focused on the screening method used by Steinmetz et al. (1991) to identify their monoclonal antibodies, because they used whole, live, non-permeabilized, non-fixed cells attached to wells of 96-well ELISA plates, which, again, implied that HtpB was surface exposed in (or easily released by) Legionella. However, these investigators could not detect Legionella whole cells by immunofluorescence microscopy. Another interesting finding of Steinmetz et al. (1991) is that not all the L. pneumophila strains tested had surface-exposed HtpB, in spite of showing abundant HtpB after sonication. In conclusion, experimentation with monoclonal antibodies against HtpB has clearly shown that HtpB has unique structural regions not found in other Group I chaperonins, and also suggested that HtpB is surface exposed in some strains of L. pneumophila.
Early molecular biology experiments with HtpB
Paul S. Hoffman’s lab was the first to clone and express the L. pneumophila htpAB operon in E. coli (Hoffman et al., 1989) and a year later, the nucleotide sequence of htpB was published almost simultaneously by Sampson et al. (1990) and Hoffman et al. (1990). There was good agreement between the two published DNA sequences of htpB, but only Hoffman et al. (1990) reported the sequence and gene organization of the htpAB operon. The expression of ectopic HtpB in E. coli also allowed Hoffman et al. (1989) to determine that HtpB could not complement a temperature-sensitive GroEL defect in E. coli strain CG218 [groEL100(Ts)]. This is an important experimental result because it indicated, at the molecular level, that GroEL and HtpB are not functionally equivalent.
Links between HtpB and L. pneumophila virulence
Hoffman et al. (1990) showed by immunofluorescence microscopy that HtpB is detectable on the surface of the virulent L. pneumophila Philadelphia-1 strain SVir suspended in Dulbecco-modified Eagle’s medium (DMEM). In contrast, surface-exposed HtpB was only weakly detectable on the salt-tolerant avirulent derivative AVir suspended in DMEM. Clearly, only virulent legionellae suspended in DMEM had the ability to display HtpB on their cell surface, an observation that provided the first link between HtpB and virulence. These investigators also showed that HtpB is abundantly expressed (and released) in L. pneumophila-infected HeLa cells, which were immuno-labeled with an intense diffuse pattern (rather than a particulate one), suggesting that HtpB was free in the LCV where this bacterium replicates (Hoffman et al., 1990). Its abundant release in the LCV also suggested that HtpB might play a role in the intracellular establishment of L. pneumophila.
An early response of L. pneumophila strain 2064 to the presence of host cells involves de novo synthesis of increasing amounts of HtpB (Fernandez et al., 1996; see Induction of HtpB Expression by Heat Shock and Presence of Host Cells below). However, an isogenic, salt-tolerant, avirulent derivative of 2064 was unable to respond, and showed no de novo synthesis of HtpB in the same experimental conditions used for 2064 (Fernandez et al., 1996). This observation provided an additional link between HtpB and L. pneumophila virulence, and suggested that HtpB might be required at an early stage of the infection process, even before L. pneumophila is internalized. In conclusion, the abilities to produce new HtpB in response to host mammalian cells, and display HtpB on the bacterial cell surface, are lost in avirulent legionellae.
Induction of HtpB expression by heat shock and presence of host cells
High temperature protein B is induced by heat shock. Increased levels of HtpB were detected in L. pneumophila (Lema et al., 1988) and in L. pneumophila and E. coli (Hoffman et al., 1989) upon temperature increases. However, the maximum increase in HtpB expression upon heat shock was ∼twofold, and at all temperatures tested HtpB remained as one of the most abundant proteins in L. pneumophila. This constitutes a pattern of heat shock that is different from the pattern typically seen in other bacteria (e.g., E. coli as shown in Hoffman et al., 1989), where the basal levels of chaperonin are low and a sharp increase is observed at high temperatures. Clearly, HtpB is not a typical Hsp in L. pneumophila. In addition, HtpB seems to be induced in virulent L. pneumophila by the presence of mammalian host cells (monocytes and L929 cells), as demonstrated by Fernandez et al. (1996) using pulse radiolabeling in cycloheximide-treated, Legionella-infected cells. The induced synthesis of new HtpB did not require bacterial internalization (inhibited with cytochalasin D), suggesting that contact with host cells was sufficient to trigger the response. Finally, Fernandez et al. (1996) determined by immunoelectron microscopy that HtpB epitopes were present on the phagosomal membrane and the cytoplasm of the infected cell.
Immunological studies with HtpB
From its very discovery, HtpB was regarded as strongly antigenic. Thus, investigators focused on establishing whether HtpB was a protective antigen, potentially applicable for vaccination against LD. Immunization with HtpB protected guinea pigs from a lethal aerosol challenge with L. pneumophila, and the protection was mediated by a strong cellular response (Blander and Horwitz, 1993). These authors wondered how HtpB is released intracellularly to elicit a cellular response, and performed immunoelectron microscopy localization studies (reported as unpublished data) indicating that HtpB was abundantly released into phagosomes of infected human monocytes. Finally these authors also mentioned that HtpB is released into the supernatant of liquid L. pneumophila cultures, suggesting it could be a secreted protein.
Weeratna et al. (1994) also immunized guinea pigs with HtpB, but contrary to the results of Blander and Horwitz (1993), they did not record a strong protective effect. The response to HtpB immunization was mainly humoral. However, guinea pigs that recovered from a L. pneumophila infection showed strong cutaneous delayed-type hypersensitivity, as well as strong lymphocyte proliferative responses to HtpB, suggesting that the presentation of HtpB during infection differs from the presentation of soluble HtpB during vaccination. To date, the experimental differences observed in the protective abilities of HtpB between these two immunization studies have not been resolved.
Purified bacterial chaperonins, including HtpB, are capable of triggering the secretion of interleukin (IL)-1 and the transcription of several cytokine genes in antigen presenting cells (Retzlaff et al., 1994), an effect demonstrated to be LPS-independent. In particular, HtpB was shown to interact with macrophage cell receptors and trigger a signaling cascade that involved PKC (Retzlaff et al., 1996). The IL-1β response was greatly reduced by heat inactivation of HtpB, a treatment that would not affect LPS-induced effects (Retzlaff et al., 1996).
In summary, HtpB is highly immunogenic, capable of interacting with cell surface receptors on macrophages, and able to elicit immunological responses via activation of signaling cascades. These early studies with HtpB resonate with those that recognized chaperonins as an important danger signal easily recognized by antigen presenting cells (Bethke et al., 2002), as part of an immune surveillance mechanism (Zügel and Kaufmann, 1999).
Are there multiple copies of HtpB in L. pneumophila?
We would like to end this historical perspective with a brief discussion of the puzzling notion advanced by Lema and Brown (1995) that L. pneumophila has two HtpB chaperonins, encoded by two copies of the htpB gene. By SDS-PAGE, these authors showed that L. pneumophila has two HtpB species of different mass and protease-digestion patterns. Southern blot analysis of DNA hybridized with an htpAB probe showed two distinct bands. These results are in sharp conflict with those of Hoffman et al. (1989), who by Southern blot showed only one htpAB locus. In addition, the completed genome sequences of five different L. pneumophila strains (D’Auria et al., 2010), indicate that there is only one copy of the htpAB operon in the common lab strains of L. pneumophila. Our own results (see HtpB Exists in Different Forms and HtpB is Essential for L. pneumophila Viability below) also confirm the presence of only one htpAB locus in two L. pneumophila strains.
HtpB Research – 1998 to Date
The evidence presented above, reveals HtpB as an intriguing L. pneumophila protein that potentially moonlights as a virulence factor. There is only one copy of the htpAB operon in the L. pneumophila chromosome, which shows the typical gene organization of Group 1 chaperonins (Figure 2), where a single regulatory region with one σ32 stress promoter (recognized by RpoH) and a housekeeping σ70 promoter, is present upstream of the co-chaperonin gene htpA. The putative htpAB transcripts produced from each of the promoters are bicistronic. Dr. K. Brassinga (currently at the University of Manitoba, Canada) mapped three integration host factor (IHF) binding sites in the regulatory region of the htpAB operon. One of these IHF binding sites overlaps an UP element immediately upstream of the σ32 stress promoter, and has been hypothesized to be responsible for the high basal level of HtpB expression in L. pneumophila (unpublished results). Interestingly, the expression of L. pneumophila IHF is developmentally regulated (Morash et al., 2009), with the highest levels being present in the differentiated mature infectious forms that emerge from host cells. What follows is an account of the HtpB research performed in our lab, which has confirmed the virulence functions of this intriguing chaperonin.

Figure 2. Upstream regulatory region and gene organization of the L. pneumophila htpAB operon. Diagram (not at scale) showing the known regulatory elements in the promoter region and the putative bicistronic transcripts (thin, right angle arrows) produced from the σ32 stress promoter and the housekeeping σ70 promoter. The dotted thick line represents an UP element, and the solid thick lines represent integration host factor binding sites. SD, Shine–Dalgarno sequence. The regulatory mechanism that controls the expression of the htpAB operon is not well understood.
HtpB is found in extracytoplasmic locations
To substantiate previous (mostly anecdotal) suggestions that HtpB is found on the cell surface of L. pneumophila (see Discovery and Initial Characterization, Monoclonal Antibodies and the Unique Epitopes of HtpB, and Links Between HtpB and L. pneumophila Virulence), Garduño et al. (1998a) undertook a detailed ultrastructural study based on immunoelectron microscopy, to define the localization of HtpB in L. pneumophila. Using a polyclonal antibody raised against the purified ectopic HtpB expressed in E. coli, and the monoclonal antibody GW2X4B8B2H6 (Helsel et al., 1988), it was found that ∼58% of the HtpB epitopes detected by immunoelectron microscopy were extracytoplasmic. An additional ∼16% of the epitopes were found in the cytoplasmic membrane. Among the extracytoplasmic HtpB epitopes, ∼30–40% were associated with the outer membrane or on the bacterial cell surface. In addition, the polyclonal antibody labeled the surface of whole, unfixed L. pneumophila cells, confirming the presence of surface-exposed HtpB. To date, similar results have been obtained with the Philadelphia-1 strains Svir, Lp02, and JR32, and the Olda clinical isolate 2064. Garduño et al. (1998a) also demonstrated that L. pneumophila abundantly releases HtpB in the LCV while replicating in HeLa cells, confirming the previous suggestion of Blander and Horwitz (1993) that HtpB accumulates in phagosomes, and explaining the diffuse labeling pattern observed in L. pneumophila-infected HeLa cells by Hoffman et al. (1990).
This immunolocalization study also showed that in E. coli the GroEL and HtpB chaperonins largely reside in the cytoplasm. Thus, we hypothesized that L. pneumophila must have a translocation mechanism, not present in E. coli, which allows the mobilization of HtpB to extracytoplasmic locations, including the bacterial cell surface. Using a combined experimental approach involving immunoelectron microscopy, protease-sensitivity, osmotic shock, and immunoblotting we have determined that ∼1% of the total cell-associated HtpB is present in the periplasm of L. pneumophila, and that a functional Dot/Icm type IV secretion system is required for the surface localization of HtpB (Chong et al., 2006). That is, loss-of-function dot mutations led to absence of surface-exposed HtpB and its accumulation in the periplasm of L. pneumophila. In particular, an Lp02 ΔdotB mutant accumulated up to fourfold more HtpB in the periplasmic space than the parent strain Lp02 (Chong et al., 2006). We still do not know how HtpB reaches the periplasm of L. pneumophila, but from the periplasm it reaches the bacterial cell surface in a Dot/Icm-dependent manner (unpublished results). It is possible that the strong association of HtpB with the inner membrane of L. pneumophila (Gabay and Horwitz, 1985) results in its passage to the periplasm, by a mechanism similar to that described for the cell-penetrating peptides (Zorko and Langel, 2005). A similar mechanism could be invoked for the passage of HtpB across the LCV membrane (see Intracellularly Released HtpB Alters the Actin Cytoskeleton of Host Cells).
Structural changes of the bacterial cell envelope during the morphological differentiation of L. pneumophila, correlate with an increased level of periplasmic HtpB and its association with the outer membrane, as detected by immunogold electron microscopy (Garduño et al., 1998b) and cell fractionation (Garduño et al., 2002). Finally, Galka et al. (2008) found small amounts of HtpB among the secreted proteins of L. pneumophila, and a larger amount in outer membrane vesicles (OMVs). We were also able to detect HtpB in purified OMVs by immunoblot, but detection had to rely on our polyclonal HtpB-specific antibody, because monoclonal antibody GW2X4B8B2H6 was not reactive with this material, suggesting that in OMVs the C-terminus of HtpB is hidden.
Collectively, the experimental results presented in this section suggest that HtpB is clearly present in extracytoplasmic locations, and that extracytoplasmic HtpB appears to be important for L. pneumophila biology, including its morphological differentiation.
HtpB exists in different forms
The notion advanced by Lema and Brown (1995) that L. pneumophila has two HtpB chaperonins is appealing, not at the gene level, but at the protein level, mainly because in our own investigations we have often seen in SDS-PAGE gels two distinct protein bands clearly labeled with HtpB-specific antibodies. In addition, under non-reducing conditions, an additional species of HtpB with an apparent mass of 80-kDa is shown (unpublished data). This 80-kDa band is only labeled with polyclonal antibody and is not recognized by monoclonal antibody GW2X4B8B2H6, suggesting that the C-terminus of this form of HtpB is not accessible. However, when this band is excised from the non-reducing gel and then re-run in a reducing SDS-PAGE gel, a single 60-kDa HtpB band is observed, which can now be labeled with monoclonal antibody GW2X4B8B2H6. Additional evidence for the existence of post-translational modifications in HtpB, comes from the analysis of our various preparations of purified HtpB. When HtpB is purified as a recombinant protein from E. coli, it runs in 2-D protein gels as a series of clustered spots of slightly different isoelectric points (pI). This pattern is common in bacterial chaperonins, particularly GroEL, where the differences in pI are likely due to different levels of phosphorylation (Sherman and Goldberg, 1992). It should be considered here that, inevitably, this preparation of recombinant HtpB is mixed with GroEL, which would increase the heterogeneity of the sample. However, 2-D protein gels of the highly purified HtpB from L. pneumophila show a series of scattered spots of different mass and pI, all of which yield identity to HtpB by mass spectrometry (unpublished data). Thus, it is clear that HtpB experiences post-translational modifications in L. pneumophila, which might involve crosslinking via disulfide bonds, phosphorylation, cleavage, and(or) altered binding abilities. Some of these modifications have been documented in other bacterial chaperonins. For instance, the phosphorylated chaperonins of E. coli (Sherman and Goldberg, 1994), M. tuberculosis (Kumar et al., 2009), and Streptomyces granaticolor (Bobek et al., 2004) have altered binding properties, and the secreted chaperonin of M. tuberculosis Cpn60.2 is cleaved by the surface anchored protease Rv2224c (Rengarajan et al., 2008). We currently do not know whether the differentially processed HtpB forms are meant to have different locations or perform particular functions, but homologs of Rv2224c are not found in L. pneumophila.
We discovered that overexpression of HtpB in L. pneumophila correlates with filamentation (unpublished results). That L. pneumophila forms long filaments is a widely known fact, and filamentation has been previously linked to the ability of L. pneumophila to survive in the environment and form biofilms (Piao et al., 2006). Thus, we have identified htpB as the first L. pneumophila gene implicated in filamentation. Furthermore, HtpB expressed alone from an IPTG-induced promoter, or in combination with HtpA from its own promoter, is sufficient to induce filamentation in E. coli. Expression of HtpA alone from its own promoter does not induce filamentation in E. coli (unpublished results) The molecular mechanism that links HtpB and filamentation remains to be elucidated, but we hypothesize that it is mediated by one of the HtpB forms present in the bacterial cytoplasm (simply because in E. coli HtpB is confined to the cytoplasm). That is, excess HtpB could result in either sequestration or misfolding of a protein involved in cell division (Kerner et al., 2005), or stabilization/activation of a cell division inhibitor, e.g., MinD (Houry et al., 1999). Alternatively, excess htpB transcript could interact with other transcripts or with RNA-binding factors, modifying the expression of components of the cell division machinery. Interestingly, impairment of the E. coli GroEL function by temperature-sensitive mutations (Horwich et al., 1993), and severe heat shock in some bacterial species, e.g., Aeromonas salmonicida (Garduño et al., 1992) results in filamentation, but the mechanism involved is unknown. Since HtpB is upregulated during the interaction of L. pneumophila with mammalian cells (refer to Links Between HtpB and L. pneumophila Virulence and Induction of HtpB Expression by Heat Shock and Presence of Host Cells above, and Fernandez et al., 1996) it would be expected that the interacting legionellae would become filamentous, a phenomenon that we have observed in human macrophage lines.
HtpB is essential for L. pneumophila viability
Attempts to replace htpB with a kanamycin- or a gentamicin-resistance cassette repeatedly yielded negative results (Chong et al., 2009). We recovered numerous putative post-allelic replacement clones with the correct antibiotic selection phenotype, but in all clones tested we still detected HtpB by immunoblot and htpAB by PCR. This is not surprising because chaperonins are essential and bacteria harboring a single chaperonin gene cannot afford to lose it. However, in bacteria with multiple chaperonin genes, usually one of the genes can be deleted (refer to Protein Folding-Independent Functions of Group I Chaperonins above, and Hu et al., 2008). Therefore, we attempted to delete htpAB in a L. pneumophila mutant carrying the groELS operon of E. coli in its chromosome. Immunoblot confirmed that recombinant GroEL was expressed in the mutant at levels comparable to those of HtpB. Nonetheless, allelic exchange of htpAB with a gentamicin-resistance cassette was still unsuccessful, suggesting that groELS could not genetically complement the htpAB operon. Interestingly, Southern blot analysis of putative post-allelic replacement clones showing the correct antibiotic-resistance phenotype (from L. pneumophila carrying or not a chromosomal groELS operon) indicated the presence of two htpAB loci, one apparently intact and another with the integrated gentamicin-resistance cassette.
In summary, the htpAB locus is essential for the viability of L. pneumophila, cannot be genetically complemented by the groELS operon of E. coli, and attempts to delete it result in genetic rearrangements that seem to involve gene duplication. Not being able to obtain a ΔhtpB mutant, and being convinced that the use of temperature-sensitive htpB mutants is not useful to study the protein folding-independent functions of HtpB (mainly because chaperonins fold so many important proteins in bacterial cells (Fujiwara et al., 2010) and would thus be impossible to ascribe phenotypes to either HtpB or its obligate folding substrates), we have relied on functional tests, which involve purified or recombinant HtpB, to determine whether HtpB is a bona fide moonlighting protein.
HtpB meets the defining characteristics of a moonlighting protein
As explained in Section “Protein Folding-Independent Functions of Group I Chaperonins” above, a moonlighting protein performs two different roles when it is in different cellular locations or in different molecular environments. If HtpB is found in cytoplasmic and extracytoplasmic locations, as well as associated with the cytoplasmic membrane of L. pneumophila, we wondered whether it would play different functional roles according to its location. In the following subsections we will describe HtpB as a multifunctional protein that according to its location and molecular environment plays different roles.
Surface-exposed HtpB acts as an invasion factor. The HtpB found on the legionellae surface (as confirmed by its susceptibility to trypsin and neutralization by antibodies) turned out to play the role of an invasion factor, mediating the internalization of L. pneumophila by HeLa cells (Garduño et al., 1998b). Five different lines of experimental evidence collectively indicated that surface-exposed HtpB interacts with specific receptors on HeLa cells promoting both attachment and internalization of L. pneumophila (or inert HtpB-coated latex microbeads). We attempted to identify the HeLa cell receptor for HtpB, and focused upon an ∼70-kDa HeLa cell membrane protein pulled down by HtpB-coated beads. In addition, a protein band of the same molecular size was labeled in an overlay membrane assay where HeLa cell membrane proteins separated by SDS-PAGE were transferred to nitrocellulose, incubated with purified HtpB, and subsequently washed and labeled with an HtpB-specific antibody (unpublished data). Although we were not able to unequivocally identify this protein, others have reported a number of receptors for Group I chaperonins, which include Toll-like receptor (TLR)-4 (Ohashi et al., 2000), TLR-2 (Nussbaum et al., 2006; Vabulas et al., 2001), the β2 integrin CD18 (Long et al., 2003), and cellular prion protein (Watarai et al., 2003). Regardless of the identity of the HeLa cell receptor for HtpB, a signaling event was clearly involved in the phagocytosis of HtpB-coated beads into a tight phagosome (Garduño et al., 1998b).
Surface-exposed HtpB alters organelle traffic. In HeLa cells, the internalized HtpB-coated beads appeared to traffic differently than bovine serum albumin (BSA)-coated beads, so we engaged in the characterization of trafficking events that followed the internalization of HtpB-coated beads. It took several years to complete a series of experiments that substantiated the notion that HtpB-coated beads indeed have a unique trafficking in relation to beads coated with GroEL or BSA. These experiments showed that internalized HtpB-coated beads attract mitochondria in CHO cells and macrophages, delay the fusion of phagosomes with Texas red-ovalbumin-labeled lysosomes in CHO cells and bone marrow-derived mouse macrophages, and induce a transient disappearance of stress fibers in CHO cells (Chong et al., 2009). Therefore, the purified HtpB attached to inert microbeads is capable of mimicking 3 post-internalization events that typify the early trafficking of L. pneumophila, and constitutes the first L. pneumophila protein that alone is sufficient to recruit mitochondria.
Outer membrane vesicles purified from L. pneumophila cultures and attached to microbeads via antibodies that recognize the L. pneumophila lipopolysaccharide, were able to transiently inhibit phagosome–lysosome fusion (Fernandez-Moreira et al., 2006). Since HtpB is present in OMVs in a unique form (see HtpB is Found in Extracytoplasmic Locations above), and HtpB-coated beads also transiently inhibit phagosome–lysosome fusion, we are tempted to speculate here that the HtpB present in OMVs might moonlight as a factor that delays fusion with lysosomes.
Intracellularly released HtpB alters the actin cytoskeleton of host cells. Since our intention was to conduct a direct comparison between the effects of HtpB from without (as it would be presented by extracellular L. pneumophila) and its effects from within (as it would be presented by intracellular L. pneumophila during infection), we needed a host cell type that would interact well with, and internalize, exogenously added protein-coated beads while being also amenable for genetic manipulation to express ectopic HtpB in their cytoplasm. CHO cells met these requirements, and therefore our experiments were focused on the stably transfected CHO-AA8 Tet-Off cells (Clontech-BD, Palo Alto, CA, USA) carrying an integrated vector (pTRE2hyg) containing the htpB gene. These cells are subsequently referred to as CHO-htpB cells (Chong et al., 2009). The aforementioned HtpB effects from without (see Surface-Exposed HtpB Alters Organelle Traffic above), were investigated in CHO-htpB cells not expressing ectopic HtpB to which we added beads coated with HtpB, or the control proteins BSA and GroEL.
The first experiment conducted with CHO-htpB cells to address effects from within, was to determine whether or not HtpB is indeed presented from within as a protein that reaches the infected cell’s cytoplasm. Using fusions with the translocation reporter gene cyaA (encoding the calmodulin-dependent Bordetella pertussis adenylate cyclase subunit) we were able to determine that during infection of CHO-htpB cells with L. pneumophila strains Lp02 and JR-32, HtpB reaches the cytoplasm of the infected cell. These results were confirmed in U937-derived macrophages (unpublished data). Therefore, we confidently proceeded to investigate the effects of HtpB from within, which required induction of ectopic HtpB in CHO-htpB cells in the absence of doxycycline.
The ectopically expressed HtpB in CHO-htpB cells (presented from within as the HtpB released from the LCV during infection) induced the disappearance of stress fibers and the relocalization of polymerized actin at the periphery of the cell. The same effect (but transiently) was produced by HtpB presented from without (see Surface-Exposed HtpB Alters Organelle Traffic), indicating the ability of HtpB to trigger the same effect from opposite sides of a membrane. The most convincing explanation for this observation is that HtpB is capable of triggering a signaling pathway by interacting with membrane receptors, and that this interaction involves the integration of HtpB in the membrane. Alternatively, it is possible that the HtpB present in the eukaryotic cytoplasm acts as a foreign protein folding machine that could trigger conformational changes in specific host factors and initiate signaling cascades. In this respect, it should be recalled that (i) several chaperonin receptors do exist (see Surface-Exposed HtpB Acts as an Invasion Factor), (ii) chaperonins, in general, have demonstrated their ability to act as signaling molecules (Ranford et al., 2000), (iii) chaperonins can integrate into membranes (Török et al., 1997), and (iv) chaperonins can interact with small GTP-binding proteins like Ras (Ikawa and Weinberg, 1992). We have hypothesized that the alteration of actin microfilaments could be involved in the altered trafficking of mitochondria in L. pneumophila-infected cells, and in cells with internalized HtpB-coated beads (Chong et al., 2009).
HtpB in the eukaryotic cytoplasm has several protein targets. To search for eukaryotic proteins that could potentially interact with the intracellularly released HtpB, we expressed HtpB in the genetically tractable eukaryote Saccharomyces cerevisiae, and also conducted a series of yeast two-hybrid assays.
In S. cerevisiae, HtpB (but not GroEL nor the yeast Hsp60) induced pseudohyphal growth, a yeast phenotype assumed during sexual reproduction that is tightly regulated by a Ras2-controlled signaling cascade (Chong et al., 2006). That HtpB uses this signaling cascade was demonstrated by showing that a S. cerevisiae Δras2 mutant does not filament upon expression of ectopic HtpB. These observations were followed by a series of yeast 2-hybrid assays against a yeast genomic library and a HeLa cell cDNA library, where HtpB (bait) was shown to interact with yeast S-adenosyl methionine decarboxylase (SAMDC), mammalian merlin-associated protein, and mitochondrial Hsp10 (Chong et al., 2006, and unpublished results). The hit with SAMDC was particularly meaningful in relation to pseudohyphal growth, mainly because alterations in intracellular levels of polyamines had been previously correlated with fungal filamentation (Herrero et al., 1999). We cloned SPE.2, the yeast gene that encodes SAMDC, and determined that its overexpression in S. cerevisiae also induced pseudohyphal growth, a result that validated SAMDC as a target of HtpB, and linked polyamines to HtpB and pseudohyphal growth signaling in S. cerevisiae. It was puzzling, however, that SAMDC was not identified in the yeast 2-hybrid screening of the HeLa cDNA library, but we have recently obtained evidence for the interaction of HtpB with mammalian and amebal SAMDC, by far western and dot blot (unpublished results). The fact that SAMDC is part of the mechanism by which HtpB effects intracellular signaling and filamentation in yeast, clearly established a link between HtpB and polyamines. Therefore, we wondered whether polyamines have a physiological impact on L. pneumophila.
It turns out that polyamines enhance the intracellular growth of L. pneumophila, whereas the inhibition of their synthesis impairs such growth. In addition, according to our bioinformatics analysis of the L. pneumophila genomes, L. pneumophila lacks 10 of the 12 enzymes described so far that are involved in the biosynthesis of polyamines in bacteria. This was a striking finding suggesting that L. pneumophila is incapable of synthesizing all polyamines, and that it might acquire them directly from its hosts. Therefore, we have hypothesized that one of the functions performed by the HtpB released into the cytoplasm of host cells could be to (through its interaction with SAMDC) increase the intracellular pool of polyamines, which L. pneumophila subsequently takes up. We are currently testing this hypothesis by: (i) measuring the levels of polyamines in CHO-htpB cells expressing and not expressing HtpB, as well as in L. pneumophila-infected cells, and (ii) determining whether HtpB extends the half-life of mammalian or amebal SAMDC, protecting it from early natural degradation. For now the role of polyamines on the physiology of L. pneumophila, and the hypothetical role of HtpB in the process constitutes an unfolding story.
As for the interactions with merlin-associated protein and Hsp10, future investigation awaits to elucidate their meaning. However, both interactions could have potential implications for the already identified effects of HtpB in mammalian cells. That is, merlin-associated protein is a member of the band 4.1 superfamily (Takeuchi et al., 1994) considered microfilament reorganizers. The protein Merlin itself is closely related to ezrin, radixin, and moesin, which are involved in the organization of cortical actin (McClatchey and Fehon, 2009). An HtpB interaction with these proteins is certainly relevant to the redistribution of actin filaments in CHO cells exposed to HtpB-coated beads and in CHO-htpB cells expressing HtpB (see Intracellularly Released HtpB Alters the Actin Cytoskeleton of Host Cells above). However, any specific involvement is yet to be demonstrated. On the other hand, an interaction with mitochondrial Hsp10 could be relevant to the recruitment of mitochondria by HtpB-coated beads (see Surface-Exposed HtpB Alters Organelle Traffic above) simply because Hsp10 has been detected on the surface of mitochondria, as well as in other extra-mitochondrial locations where Hsp10 moonlights as the early pregnancy factor (Sadacharan et al., 2001). This is not entirely surprising since Hsp10 is a mitochondrial protein whose encoding gene resides in the cell nucleus, and it is synthesized in the eukaryotic cytosol, from where Hsp10 needs to be imported into the mitochondria (Ryan et al., 1997). While mitochondrial protein import is mostly co-translational, it is entirely possible that some Hsp10 molecules could stay on the mitochondrial surface (bound to the import apparatus) after translation, and therefore be available to interact with HtpB.
An Integrated Functional Model for HtpB
The identified functions of HtpB (both confirmed and hypothetical) are summarized in Table 1. Based on these functions we have envisioned the following model to explain how HtpB moonlighting activities might impact the biology and pathogenesis of L. pneumophila (Figure 1): HtpB in the bacterial cytoplasm meets the essential protein folding needs of L. pneumophila helping in adaptation to stress and mounting responses to potential hosts. At the same time, elevated levels of HtpB in the bacterial cytoplasm correlate with filamentation, a phenotype that seems to favor the survival of L. pneumophila in the aquatic environment. As the major cytoplasmic membrane protein of L. pneumophila, HtpB could fulfill a lipochaperonin function (Török et al., 1997). Surface-exposed HtpB, which increases in the presence of mammalian host cells, as well as during the morphological differentiation of L. pneumophila into mature infectious forms, interacts with eukaryotic cell receptors and mediates attachment to and invasion of host cells. The abundantly released HtpB in the lumen of early phagosomes and LCV has no identified functions, as yet, but possibly it is from this compartment that HtpB reaches the cytoplasm of host cells, either via OMVs (see HtpB is Found in Extracytoplasmic Locations), or by direct passage through the LCV membrane (Zorko and Langel, 2005). It is in the cytoplasm of host cells (either free in the cytosol, or bound to the LCV membrane) that HtpB mediates recruitment of mitochondria, alters the actin cytoskeleton of the host cell, and putatively increases the intracellular pool of polyamines.
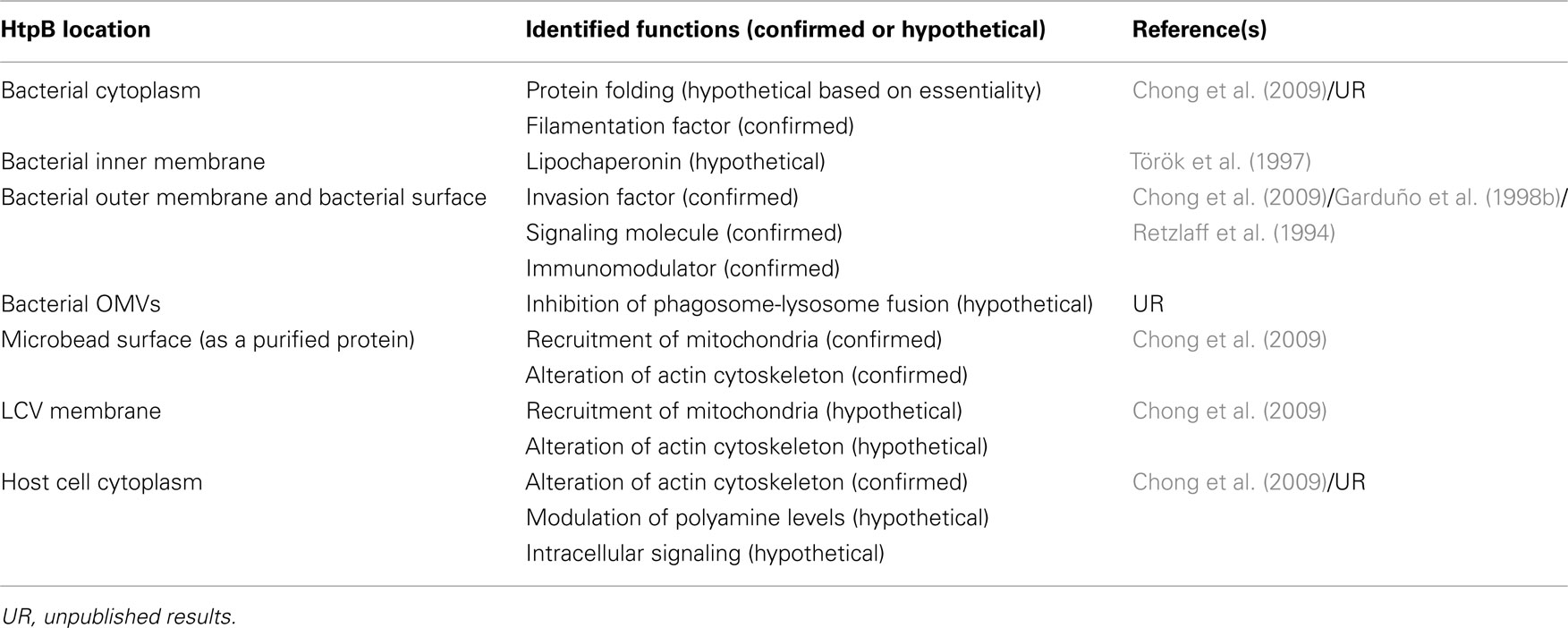
Table 1. Identified functions of the L. pneumophila chaperonin, HtpB, according to its location in the bacterial cell and in the host cell.
The study of HtpB functions, which are not seemingly shared by other Group 1 chaperonins, promises to increase our general understanding of chaperonin biology and the evolution of intracellular pathogens that have adapted to the human host by using an ancient protein tool.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The work performed in the Garduño lab, has been funded by the Canadian Natural Sciences and Engineering Research Council (NSERC). Rafael A. Garduño holds a Canada Research Chair, Tier II, in Foodborne and Waterborne Bacterial Pathogens. We acknowledge the valuable suggestions received from the anonymous reviewers of our original manuscript, which resulted in a much improved revised version.
References
Archibald, J. M., Logsdon, J. M., and Doolittle, W. F. (2000). Origin and evolution of eukaryotic chaperonins: phylogenetic evidence for ancient duplications in CCT genes. Mol. Biol. Evol. 17, 1456–1466.
Bethke, K., Staib, F., Distler, M., Schmitt, U., Jonuleit, H., Enk, A. H., Galle, P. R., and Heike, M. (2002). Different efficiency of heat shock proteins (HSP) to activate human monocytes and dendritic cells: superiority of Hsp60. J. Immunol. 169, 6141–6148.
Blander, S. J., and Horwitz, M. A. (1993). Major cytoplasmic membrane protein of Legionella pneumophila, a genus common antigen and member of the hsp 60 family of heat shock proteins, induces protective immunity in a guinea pig model of Legionnaires’ disease. J. Clin. Invest. 91, 717–723.
Bobek, J., Halada, P., Angelis, J., Vohradský, J., and Mikulík, K. (2004). Activation and expression of proteins during synchronous germination of aerial spores of Streptomyces granaticolor. Proteomics 4, 3864–3880.
Braig, K., Otwinowski, Z., Hegde, R., Boisvert, D. C., Joachimiak, A., Horwich, A. L., and Sigler, P. B. (1994). The crystal structure of the bacterial chaperonin GroEL at 2.8 Å. Nature 371, 578–586.
Cehovin, A., Coates, A. R. M., Hu, Y., Riffo-Vasquez, Y., Tormay, P., Botanch, C., Altare, F., and Henderson, B. (2010). Comparison of the moonlighting actions of the two highly homologous chaperonin 60 proteins of Mycobacterium tuberculosis. Infect. Immun. 78, 3196–3206.
Chong, A., Lima, C. A., Allan, D. S., Nasrallah, G. K., and Garduño, R. A. (2009). The purified and recombinant Legionella pneumophila chaperonin alters mitochondrial trafficking and microfilament organization. Infect. Immun. 77, 4724–4739.
Chong, A., Riveroll, A., Allan, D. S., Garduño, E., and Garduño, R. A. (2006). “TheHsp60 chaperonin of Legionella pneumophila: an intriguing player in infection of host cells,” in Legionella: State of the Art 30 Years after Its Recognition, eds N. P. Cianciotto, Y. Abu Kwaik, P. H. Edelstein, B. S. Fields, D. F. Geary, T. G. Harrison, C. A. Joseph, R. M. Ratcliff, J. E. Stout, and M. S. Swanson (Washington, DC: ASM Press), 255–260.
D’Auria, G., Jimenez-Hernandez, N., Peris-Bondia, F., Moya, A., and Latorre, A. (2010). Legionella pneumophila pangenome reveals strain-specific virulence factors. BMC Genomics 11, 181.
England, J., Lucent, D., and Pande, V. (2008). Rattling the cage: computational models of chaperonin-mediated protein folding. Curr. Opin. Struct. Biol. 18, 163–169.
Ensgraber, M., and Loos, M. (1992). A 66-kilodalton heat shock protein of Salmonella typhimurium is responsible for binding of the bacterium to intestinal mucus. Infect. Immun. 60, 3072–3078.
Fenton, W. A., Weissman, J. S., and Horwich, A. L. (1996). Putting a lid on protein folding: structure and function of the co-chaperonin, GroES. Chem. Biol. 3, 157–161.
Fernandez, R. C., Logan, S. M., Lee, S. H., and Hoffman, P. S. (1996). Elevated levels of Legionella pneumophila stress protein Hsp60 early in infection of human monocytes and L929 cells correlate with virulence. Infect. Immun. 64, 1968–1976.
Fernandez-Moreira, E., Helbig, J. H., and Swanson, M. S. (2006). Membrane vesicles shed by Legionella pneumophila inhibit fusion of phagosomes with lysosomes. Infect. Immun. 74, 3285–3295.
Frisk, A., Ison, C. A., and Lagergärd, T. (1998). GroEL heat shock protein of Haemophilus ducreyi: association with cell surface and capacity to bind to eukaryotic cells. Infect. Immun. 66, 1252–1257.
Fujiwara, K., Ishihama, Y., Nakahigashi, K., Soga, T., and Taguchi, H. (2010). A systematic survey of in vivo obligate chaperonin-dependent substrates. EMBO J. 29, 1552–1564.
Gabay, J. E., and Horwitz, M. A. (1985). Isolation and characterization of the cytoplasmic and outer membranes of the Legionnaires’ disease bacterium (Legionella pneumophila). J. Exp. Med. 161, 409–422.
Galdiero, M., de l’Ero, G. C., and Marcatili, A. (1997). Cytokine and adhesion molecule expression in human monocytes and endothelial cells stimulated with bacterial heat shock proteins. Infect. Immun. 65, 699–707.
Galka, F., Wai, S. N., Kusch, H., Engelmann, S., Hecker, M., Schmeck, B., Hippenstiel, S., Uhlin, B. E., and Steinert, M. (2008). Proteomic characterization of the whole secretome of Legionella pneumophila and functional analysis of outer membrane vesicles. Infect. Immun. 76, 1825–1836.
Garduño, R. A., Faulkner, G., Trevors, M. A., Vats, N., and Hoffman, P. S. (1998a). Immunolocalization of Hsp60 in Legionella pneumophila. J. Bacteriol. 180, 505–513.
Garduño, R. A., Garduño, E., and Hoffman, P. S. (1998b). Surface-associated Hsp60 chaperonin of Legionella pneumophila mediates invasion in a HeLa cell model. Infect. Immun. 66, 4602–4610.
Garduño, R. A., Garduño, E., Hiltz, M., and Hoffman, P. S. (2002). Intracellular growth of Legionella pneumophila gives rise to a differentiated form dissimilar to stationary phase forms. Infect. Immun. 70, 6273–6283.
Garduño, R. A., Lee, E. J. Y., and Kay, W. W. (1992). S-layer mediated association of Aeromonas salmonicida with murine macrophages. Infect. Immun. 60, 4373–4382.
Goulhen, F., Hafezi, A., Uitto, V.-J., Hinode, D., Nakamura, R., Grenier, D., and Mayrand, D. (1998). Subcellular localization and cytotoxic activity of the GroEL-like protein isolated from Actinobacillus actinomycetemcomitans. Infect. Immun. 66, 5307–5313.
Gupta, R. S. (1995). Evolution of the chaperonin families (Hsp60, Hsp10, and Tcp-1) of proteins and the origin of eukaryotic cells. Mol. Microbiol. 15, 1–11.
Gutsche, I., Essen, L. O., and Baumeister, W. (1999). Group II chaperonins: new TRiC(k)s and turns of a protein folding machine. J. Mol. Biol. 293, 295–312.
Helsel, L. O., Bibb, W. F., Butler, C. A., Hoffman, P. S., and McKinney, R. M. (1988). Recognition of a genus-wide antigen of Legionella by a monoclonal-antibody. Curr. Microbiol. 16, 201–208.
Hemmingsen, S. M., Woolford, C., van der Vies, S. M., Tilly, K., Dennis, D. T., Georgopoulos, C. P., Hendrix, R. W., and Ellis, R. J. (1988). Homologous plant and bacterial proteins chaperone oligomeric protein assembly. Nature 333, 330–334.
Henderson, B. (2010). Integrating the cell stress response: a new view of molecular chaperones as immunological and physiological homeostatic regulators. Cell Biochem. Funct. 28, 1–14.
Hennequin, C., Porcheray, F., Waligora-Dupriet, A.-J., Collignon, A., Barc, M.-C., Bourlioux, P., and Karjalainen, T. (2001). GroEL (Hsp60) of Clostridium difficile is involved in cell adherence. Microbiology 147, 87–96.
Herrero, A. B., Lopez, M. C., Garcia, S., Schmidt, A., Spaltmann, F., Ruiz-Herrera, J., and Dominguez, A. (1999). Control of filament formation in Candida albicans by polyamine levels. Infect. Immun. 67, 4870–4878.
Hoffman, P. S., Butler, C. A., and Quinn, F. D. (1989). Cloning and temperature-dependent expression in Escherichia coli of a Legionella pneumophila gene coding for a genus-common 60-kDa antigen. Infect. Immun. 57, 1731–1739.
Hoffman, P. S., Houston, L., and Butler, C. A. (1990). Legionella pneumophila htpAB heat shock operon: nucleotide sequence and expression of the 60 kilodalton antigen in L. pneumophila-infected HeLa cells. Infect. Immun. 58, 3380–3387.
Horwich, A. L., Fenton, W. A., Chapman, E., and Farr, G. W. (2007). Two families of chaperonin: physiology and mechanism. Annu. Rev. Cell Dev. Biol. 23, 115–145.
Horwich, A. L., Low, K. B., Fenton, W. A., Hirshfield, I. N., and Furtak, K. (1993). Folding in vivo of bacterial cytoplasmic proteins: role of GroEL. Cell 74, 909–917.
Horwich, A. L., and Saibil, H. R. (1998). The thermosome: chaperonin with a built-in lid. Nat. Struct. Biol. 5, 333–336.
Houry, W. A., Frishman, D., Eckerskorn, C., Lottspeich, F., and Hartl, F. U. (1999). Identification of in vivo substrates of the chaperonin GroEL. Nature 402, 147–154.
Hu, Y., Henderson, B., Lund, P. A., Tormay, P., Ahmed, M. T., Gurcha, S. S., Besra, G. S., and Coates, A. R. (2008). A Mycobacterium tuberculosis mutant lacking the groEL homologue cpn60.1 is viable but fails to induce an inflammatory response in animal models of infection. Infect. Immun. 76, 1535–1546.
Huesca, M., Borgia, S., Hoffman, P. S., and Lingwood, C. A. (1996). Acidic pH changes receptor binding specificity of Helicobacter pylori: a binary adhesion model in which surface heat shock (stress) proteins mediate sulfatide recognition in gastric colonization. Infect. Immun. 64, 2643–2648.
Ikawa, S., and Weinberg, R. A. (1992). An interaction between p21ras and heat shock protein hsp60, a chaperonin. Proc. Natl. Acad. Sci. U.S.A. 89, 2012–2016.
Kerner, M. J., Naylor, D. J., Ishihama, Y., Maier, T., Chang, H.-C., Stines, A. P., Georgopoulos, C., Frishman, D., Hayer-Hartl, M., Mann, M., and Hartl, F. U. (2005). Proteome-wide analysis of chaperonin-dependent protein folding in Escherichia coli. Cell 122, 209–220.
Kim, S., Willison, K. R., and Horwich, A. L. (1994). Cystosolic chaperonin subunits have a conserved ATPase domain but diverged polypeptide-binding domains. Trends Biochem. Sci. 19, 543–548.
Klumpp, M., and Baumeister, W. (1998). The thermosome: archetype of group II chaperonins. FEBS Lett. 430, 73–77.
Kumar, C. M. S., Khare, G., Srikanth, C. V., Tyagi, A. K., Sardesai, A. A., and Mande, S. C. (2009). Facilitated oligomerization of mycobacterial GroEL: evidence for phosphorylation-mediated oligomerization. J. Bacteriol. 191, 6525–6538.
Lema, M. W., Brown, A., Butler, C. A., and Hoffman, P. S. (1988). Heat shock response in Legionella pneumophila. Can. J. Microbiol. 34, 1148–1153.
Lema, M. W., and Brown, A. (1995). Legionella pneumophila has two 60-kilodalton heat-shock proteins. Curr. Microbiol. 31, 332–335.
Lin, Z., and Rye, H. S. (2006). GroEL-mediated protein folding: making the impossible, possible. Crit. Rev. Biochem. Mol. Biol. 41, 211–239.
Long, K. H., Gomez, F. J., Morris, R. E., and Newman, S. L. (2003). Identification of heat shock protein 60 as the ligand on Histoplasma capsulatum that mediates binding to CD18 receptors on human macrophages. J. Immunol. 170, 487–494.
Lund, P. (2011). Insights into chaperonin function from studies on archaeal thermosomes. Biochem. Soc. Trans. 39, 94–98.
Lund, P. A. (2009). Multiple chaperonins in bacteria – why so many? FEMS Microbiol. Rev. 33, 785–800.
McClatchey, A. I., and Fehon, R. G. (2009). Merlin and the ERM proteins – regulators of receptor distribution and signaling at the cell cortex. Trends Cell Biol. 19, 198–206.
Morash, M. G., Brassinga, A. K. C., Warthan, M., Gourabathini, P., Garduño, R. A., Goodman, S. D., and Hoffman, P. S. (2009). Reciprocal expression of integration host factor and HU in the developmental cycle and infectivity of Legionella pneumophila. Appl. Environ. Microbiol. 75, 1826–1837.
Morioka, M., Muraoka, H., Yamamoto, K., and Ishikawa, H. (1994). An endosymbiont chaperonin is a novel type of histidine protein kinase. J. Biochem. 116, 1075–1081.
Nussbaum, G., Zanin-Zhorov, A., Quintana, F., Lider, O., and Cohen, I. R. (2006). Peptide p277 of HSP60 signals T cells: inhibition of inflammatory chemotaxis. Int. Immunol. 18, 1413–1419.
Ohashi, K., Burkhart, V., Flohe, S., and Kolb, H. (2000). Cutting edge: heat shock protein 60 is a putative endogenous ligand of the Toll-like receptor-4 complex. J. Immunol. 164, 558–561.
Ohtaki, A., Noguchi, K., and Yohda, M. (2010). Structure and function of archaeal prefoldin, a co-chaperone of group II chaperonin. Front. Biosci. 15, 708–717.
Paju, S., Goulhen, F., Asikainen, S., Grenier, D., Mayrand, D., and Uitto, V.-J. (2000). Localization of heat shock proteins in clinical Actinobacillus actinomycetemcomitans strains and their effects on epithelial cell proliferation. FEMS Microbiol. Lett. 182, 231–235.
Pau, C.-P., Plikaytis, B. B., Carlone, G. M., and Warner, I. M. (1988). Purification, partial characterization, and seroreactivity of a genuswide 60-kilodalton Legionella protein antigen. J. Clin. Microbiol. 26, 67–71.
Piao, Z., Sze, C. C., Barysheva, O., Iida, K.-I., and Yoshida, S.-I. (2006). Temperature-regulated formation of mycelial mat-like biofilms by Legionella pneumophila. Appl. Environ. Microbiol. 72, 1613–1622.
Plikaytis, B. B., Carlone, G. M., Pau, C.-P., and Wilkinson, H. W. (1987). Purified 60-kilodalton Legionella protein antigen with Legionella-specific and non-specific epitopes. J. Clin. Microbiol. 25, 2080–2084.
Portaro, F. C., Hayashi, M. A., De Arauz, L. J., Palma, M. S., Assakura, M. T., Silva, C. L., and de Camargo, A. C. (2002). The Mycobacterium leprae hsp65 displays proteolytic activity. Mutagenesis studies indicate that the M. leprae hsp65 proteolytic activity is catalytically related to the HslVU protease. Biochemistry 41, 7400–7406.
Ranford, J. C., Coates, A. R., and Henderson, B. (2000). Chaperonins are cell-signalling proteins: the unfolding biology of molecular chaperones. Expert Rev. Mol. Med. 2, 1–17.
Rengarajan, J., Murphy, E., Park, A., Krone, C. L., Hett, E. C., Bloom, B. R., Glimcher, L. H., and Rubin, E. J. (2008). Mycobacterium tuberculosis Rv2224c modulates innate immune responses. Proc. Natl. Acad. Sci. U.S.A. 105, 264–269.
Retzlaff, C., Yamamoto, Y., Hoffman, P. S., Friedman, H., and Klein, T. W. (1994). Bacterial heat shock proteins directly induce cytokine mRNA and interleukin-1 secretion in macrophage cultures. Infect. Immun. 62, 5689–5693.
Retzlaff, C., Yamamoto, Y., Okubo, S., Hoffman, P. S., Friedman, H., and Klein, T. W. (1996). Legionella pneumophila heat-shock protein-induced increase of interleukin-1β mRNA involves protein kinase C signalling in macrophages. Immunology 89, 281–288.
Ryan, M. T., Naylor, D. J., Høj, P. B., Clark, M. S., and Hoogenraad, N. J. (1997). The role of molecular chaperones in mitochondrial protein import and folding. Int. Rev. Cytol. 174, 127–193.
Sadacharan, S. K., Cavanagh, A. C., and Gupta, R. S. (2001). Immunoelectron microscopy provides evidence for the presence of mitochondrial heat shock 10-kDa protein (chaperonin 10) in red blood cells and a variety of secretory granules. Histochem. Cell Biol. 116, 507–517.
Sampson, J. S., O’Connor, S. P., Holloway, B. P., Plikaytis, B. B., Carlone, G. M., and Mayer, L. W. (1990). Nucleotide sequence of htpB, the Legionella pneumophila gene encoding the 58-kilodalton (kDa) common antigen, formerly designated the 60-kDa common antigen. Infect. Immun. 58, 3154–3157.
Sampson, J. S., Plikaytis, B. B., Aloisio, C. H., Carlone, G. M., Pau, C.-P., and Stinson, A. R. (1991). Immunologic characterization and specificity of three monoclonal antibodies against the 58-kilodalton protein of Legionella pneumophila. J. Clin. Microbiol. 29, 836–841.
Sampson, J. S., Plikaytis, B. B., and Wilkinson, H. W. (1986). Immunologic response of patients with legionellosis against major protein-containing antigens of Legionella pneumophila serogroup 1 as shown by immunoblot analysis. J. Clin. Microbiol. 23, 92–99.
Scorpio, A., Johnson, P., Laquerre, A., and Nelson, D. R. (1994). Subcellular localization and chaperone activities of Borrelia burgdorferi Hsp60 and Hsp70. J. Bacteriol. 176, 6449–6456.
Sherman, M. Y., and Goldberg, A. L. (1992). Heat shock in Escherichia coli alters the protein-binding properties of the chaperonin groEL by inducing its phosphorylation. Nature 357, 167–169.
Sherman, M., and Goldberg, A. L. (1994). Heat shock-induced phosphorylation of GroEL alters its binding and dissociation from unfolded proteins. J. Biol. Chem. 269, 31479–31483.
Sigler, P. B., Xu, Z., Rye, H. S., Burston, S. G., Fenton, W. A., and Horwich, A. L. (1998). Structure and function in GroEL-mediated protein folding. Annu. Rev. Biochem. 67, 581–608.
Sompolinsky, D., Hertz, J. B., Hoiby, N., Jensen, K., Mansa, B., Pedersen, V. B., and Samra, Z. (1980a). An antigen common to a wide range of bacteria. 2. A biochemical study of a “common antigen” from Pseudomonas aeruginosa. Acta Pathol. Microbiol. Scand. B 88, 253–260.
Sompolinsky, D., Hertz, J. B., Hoiby, N., Jensen, K., Mansa, B., and Samra, Z. (1980b). An antigen common to a wide range of bacteria. 1. The isolation of a “common antigen” from Pseudomonas aeruginosa. Acta Pathol. Microbiol. Scand. B 88, 143–149.
Steinmetz, I., Rheinheimer, C., and Bitter-Suermann, D. (1992). Rapid identification of legionellae by a colony blot assay based on a genus-specific monoclonal antibody. J. Clin. Microbiol. 30, 1016–1018.
Steinmetz, I., Rheinheimer, C., Hübner, I., and Suermann, D. B. (1991). Genus-specific epitope on the 60-kilodalton Legionella heat shock protein recognized by a monoclonal antibody. J. Clin. Microbiol. 29, 346–354.
Takeuchi, K., Kawashima, A., Nagafuchi, A., and Tsukita, S. (1994). Structural diversity of band 4.1 superfamily members. J. Cell Sci. 107, 1921–1928.
Techtmann, S. M., and Robb, F. T. (2010). Archaeal-like chaperonins in bacteria. Proc. Natl. Acad. Sci. U.S.A. 107, 20269–20274.
Török, Z., Horváth, I., Goloubinoff, P., Kovács, E., Glatz, A., Balogh, G., and Vigh, L. (1997). Evidence for a lipochaperonin: association of active protein-folding GroESL oligomers with lipids can stabilize membranes under heat shock. Proc. Natl. Acad. Sci. U.S.A. 94, 2192–2197.
Trost, M., Wehmhöner, D., Kärst, U., Dieterich, G., Wehland, J., and Jänsch, L. (2005). Comparative proteome analysis of secretory proteins from pathogenic and non-pathogenic Listeria species. Proteomics 5, 1544–1557.
Vabulas, R. M., Ahmad-Nejad, P., da Costa, C., Miethke, T., Kirschning, C. J., Häcker, H., and Wagner, H. (2001). Endocytosed HSP60s use Toll-like receptor 2 (TLR2) and TLR4 to activate the toll/interleukin-1 receptor signaling pathway in innate immune cells. J. Biol. Chem. 276, 31332–31339.
Watarai, M., Kim, S., Erdenebaatar, J., Makino, S.-I., Horiuchi, M., Shirahata, T., Sakeguchi, S., and Katamine, S. (2003). Cellular prion protein promotes Brucella infection into macrophages. J. Exp. Med. 198, 5–17.
Weeratna, R., Stamler, D. A., Edelstein, P. H., Ripley, M., Marrie, T., Hoskin, D., and Hoffman, P. S. (1994). Human and guinea pig immune responses to Legionella pneumophila protein antigens OmpS and Hsp60. Infect. Immun. 62, 3454–3462.
Yamaguchi, H., Taguchi, H., Katura, T., Kumada, J., Uekusa, T., and Ogata, S. (1989). Purification of cross-reacting protein antigen shared by Yersinia enterocolitica and other Gram-negative bacteria with monoclonal antibody. Microbiol. Immunol. 33, 683–688.
Yoshida, N., Oeda, K., Watanabe, E., Mikami, T., Fukita, Y., Nishimura, K., Komai, K., and Matsuda, K. (2001). Protein function. Chaperonin turned insect toxin. Nature 411, 44.
Zeilstra-Ryalls, J., Fayet, O., and Georgopoulos, C. (1991). The universally conserved GroE (Hsp60) chaperonins. Annu. Rev. Microbiol. 45, 301–325.
Zhang, L., Pelech, S. L., Mayrand, D., Grenier, D., Heino, J., and Uitto, V. J. (2001). Bacterial heat shock protein-60 increases epithelial cell proliferation through the ERK1/2 MAP kinases. Exp. Cell Res. 266, 11–20.
Zhang, L., Pelech, S., and Uitto, V. J. (2004). Long-term effect of heat shock protein 60 from Actinobacillus actinomycetemcomitans on epithelial cell viability and mitogen-activated protein kinases. Infect. Immun. 72, 38–45.
Zorko, M., and Langel, Ü. (2005). Cell penetrating peptides: mechanism and kinetics of cargo delivery. Adv. Drug Deliv. Rev. 57, 529–545.
Keywords: HtpB, Hsp60, GroEL, pathogenesis, mitochondria, microfilaments, polyamines
Citation: Garduño RA, Chong A, Nasrallah GK and Allan DS (2011) The Legionella pneumophila chaperonin – an unusual multifunctional protein in unusual locations. Front. Microbio. 2:122. doi: 10.3389/fmicb.2011.00122
Received: 29 January 2011; Paper pending published: 22 February 2011;
Accepted: 17 May 2011; Published online: 10 June 2011.
Edited by:
Carmen Buchrieser, Pasteur Institute, FranceReviewed by:
Philippe Mazodier, Pasteur Institute, FranceHelene Bierne, French National Institute for Agricultural Research, France
Copyright: © 2011 Garduño, Chong, Nasrallah and Allan. This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: Rafael A. Garduño, Department of Microbiology and Immunology, Sir Charles Tupper Medical Building, 7th floor, 5850 College Street, Halifax, NS, Canada B3H-1X5. e-mail:cmFmYWVsLmdhcmR1bm9AZGFsLmNh