 Mauricio Bustamante†
Mauricio Bustamante† Valentina Verdejo†
Valentina Verdejo† Catalina Zúñiga
Catalina Zúñiga Fernanda Espinosa
Fernanda Espinosa Julieta Orlando and Margarita Carú*
Julieta Orlando and Margarita Carú*- Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Santiago, Chile
Water availability is the main limiting factor in arid soils; however, few studies have examined the effects of drying and rewetting on nitrifiers from these environments. The effect of water availability on the diversity of ammonia-oxidizing bacteria (AOB) and archaea (AOA) from a semiarid soil of the Chilean sclerophyllous matorral was determined by microcosm assays. The addition of water every 14 days to reach 60% of the WHC significantly increased nitrate content in rewetted soil microcosms (p < 0.001). This stimulation of net nitrification by water addition was inhibited by acetylene addition at 100 Pa. The composition of AOA and AOB assemblages from the soils microcosms was determined by clone sequencing of amoA genes (A-amoA and B-amoA, respectively), and the 16S rRNA genes specific for β-proteobacteria (beta-amo). Sequencing of beta-amo genes has revealed representatives of Nitrosomonas and Nitrosospira while B-amoA clones consisted only of Nitrosospira sequences. Furthermore, all clones from the archaeal amoA gene library (A-amoA) were related to “mesophilic Crenarchaeota” sequences (actually, reclassified as the phylum Thaumarchaeota). The effect of water availability on both microbial assemblages structure was determined by T-RFLP profiles using the genetic markers amoA for archaea, and beta-amo for bacteria. While AOA showed fluctuations in some T-RFs, AOB structure remained unchanged by water pulses. The relative abundance of AOA and AOB was estimated by the Most Probable Number coupled to Polymerase Chain Reaction (MPN-PCR) assay. AOB was the predominant guild in this soil and higher soil water content did not affect their abundance, in contrast to AOA, which slightly increased under these conditions. Therefore, these results suggest that water addition to these semiarid soil microcosms could favor archaeal contribution to ammonium oxidation.
Introduction
Cycles of drought-rewetting are common in most terrestrial ecosystems, but they are particularly pronounced in arid and semiarid environments mainly with seasonal rainfall (Fierer and Schimel, 2002). The Chilean matorral is a semiarid ecosystem located between the coastal and the Andes mountain ranges, between 32 and 36° south latitude with a temperate Mediterranean climate characterized by dry summers and rainy winters. The matorral is predominantly composed by shrub vegetation, which has been affected by anthropogenic activities resulting in an extensive loss of native vegetation and soil erosion with low fertility and limited nitrogen (Fuentes, 1990). In the Andean foothills some fragments of matorral are composed mainly by Colletia hystrix. These shrubs are considered pioneer species that contribute to primary plant successions stabilizing the soil and increasing its nitrogen content (Silvester et al., 1985) because of their ability to form root nodules with Frankia, a N2-fixing actinomycete (Carú, 1993; Carú et al., 2003). Moreover, this semiarid soil sustained denitrification activity and showed a broad genetic variability of denitrifiers (Orlando et al., 2012). Other major players in the nitrogen cycle are nitrifiers, archaea and/or bacteria, which have not been studied in this particular environment and have generally been addressed in some arid environments only.
In semiarid ecosystems, water availability plays a critical role in the soil affecting the osmotic status, abundance, and composition of microbial cells as well as influencing nutrient cycling (Fierer et al., 2003; Griffiths et al., 2003; Drenovsky et al., 2004; Singh and Kashyap, 2006; Schimel et al., 2007). However, nitrogen is often the most limiting nutrient (Whitford, 1992), and rewetting of dried soils produces an increment in its mineralization (Lundquist et al., 1999). As ammonium oxidation is the limiting step of nitrification and plays a central role in the global nitrogen cycle, the ammonia-oxidizing bacteria (AOB) have been used as indicators of specific environmental conditions in microbial ecology studies (Kowalchuk and Stephen, 2001). Their detection has been mainly performed using the 16S rRNA and amoA (ammonia monooxygenase subunit A) genes specific to AOB. They are affiliated to the beta- and gamma-subdivision of the Proteobacteria (Purkhold et al., 2000). However, to date, gammaproteobacterial AOB have been observed only in marine environments (Kowalchuk and Stephen, 2001; Ward and O'Mullan, 2002).
Until recently, AOB have been considered to be responsible for the majority of ammonium oxidation in soil. However, the recent discovery of the gene responsible for ammonium oxidation (amoA) in the domain archaea, and the detection of putative ammonia-oxidizing archaea (AOA) in different ecosystems, suggests that AOA may play a significant, but previously unrecognized role in the global nitrogen cycle (Francis et al., 2005; Leininger et al., 2006; He et al., 2007; Prosser and Nicol, 2008). These archaea possess genes homologs to those encoding subunits A, B, and C (amoA, amoB, and amoC homologs) of the ammonia monooxygenase (AMO) of autotrophic AOB. During the last few years, crenarchaeota possessing putative amoA genes have been identified in marine and terrestrial environments (Treusch et al., 2005; Leininger et al., 2006), although recently, the AOA have been assigned to the new archaeal phylum Thaumarchaeota (Pester et al., 2011).
Despite the critical importance of nitrification in the nitrogen cycle, our understanding of the role of the different ammonia oxidizers (i.e., AOB and AOA) in different soils and environmental conditions is still limited (Schleper, 2010), mainly because until now there are no selective inhibitors for any of these microbial groups. Nevertheless, there is a growing body of reports addressing questions about AOB and AOA ecological roles today, but it should be considered that to know what factors regulate ammonia-oxidizing populations, AOA and AOB must be studied in a wide variety of habitats. Although ammonia-oxidizers are sensitive to water availability (Stark and Firestone, 1995), especially considering that this is the main limiting factor in arid environments; few studies have examined the effects of drying and rewetting on bacterial nitrification (Stark and Firestone, 1995; Avrahami and Bohannan, 2007; Gleeson et al., 2008). However, to our knowledge, just the work of Gleeson et al. (2010) has determined the direct influence of water availability on AOB and AOA community structures from a semiarid soil, evidencing a clear separation between “wet” and “dry” samples.
The influence of rewetting stress operates at both physiological and community composition levels and these levels interact to link environmental conditions and biogeochemical processes (Schimel et al., 2007). The overall aim of this study has been to evaluate the effect of drying-rewetting treatments, in microcosms assays, on the net nitrification and community structure of AOA and AOB in a semiarid soil from Central Chile. The proposal is that, as water availability is a limiting factor for microbial activity in arid soils, the water addition will produce an increase in nitrification activity. This higher activity could not necessarily be associated with a change in nitrifiers structure due to the long generation times known for some of these microorganisms (Avrahami et al., 2002), and because the applied drying-rewetting treatments were sustained over incubation time although not drastic.
Materials and Methods
Study Site and Soil Sampling
Semiarid soil samples were obtained from the sclerophyllous matorral in the locality of “El Romeral” (33°48′S, 70°14′W), Santiago, Chile. The area presents a dry Mediterranean climate with winter rainfall averaging 350 mm annually and corresponds to vegetation fragments located in the Andean foothills. Ten random sampling points separated from each other between 1 and 2 m within a plot of 15 m × 15 m were selected, soil samples were collected from the upper 10 cm with 6 cm diameter corers and sieved to 2 mm aggregate size. The samples were homogenized to obtain a composite sample to reduce the spatial heterogeneity at a small scale (Webster et al., 2002) and stored at 4°C, for approximately one week, until the microcosms set-up.
Microcosms Design and Sampling Schedule
The microcosms were set-up with 150 g of the soil samples in clean plastic containers in a 1:3 (v:v) soil:air ratio, and kept at 20°C during 56 days. The treatments consisted in the addition of sterile water and were applied in triplicate to reach 60% of the water-holding capacity (WHC) at the beginning of the experiment and every 14 days (tH2O). The control microcosms were not wetted (w/t). In addition, one set of rewetted microcosms was treated with acetylene (100 Pa) to inhibit the autotrophic ammonium oxidation (tH2O-Ac). To maintain aerobic conditions in the microcosms, they were opened and vented every 3 days and acetylene partial pressures were re-established each time. Since in control microcosms without addition of water a notorious increase in nitrate content during a pre-incubation was not observed (data not shown), the addition of acetylene was performed only in the wetted microcosms.
Samples were obtained from each microcosm set every 7 days to determine pH, moisture content (MC), organic matter content (OM), and the ammonium and nitrate concentrations. Samples coinciding with re-wetting were taken 3 h after treatment application. Clone libraries were obtained at the beginning of the incubation (t0), while T-RFLP profiles were determined at 0, 28, and 56 days of incubation. Finally, samples to abundance calculations were taken at the beginning and at the end of the incubation.
Edaphic Factors
The pH was measured from a soil extract in 2 M KCl using potentiometry (pH 500 Benchtop Meter, Oakton® Instruments). The MC and the OM were calculated gravimetrically before and after desiccation and calcination, respectively (Forster, 1995). Ammonium and nitrate concentrations were determined by colorimetric methods from a soil extract in 2M KCl and deionized water, respectively (Nelson, 1983; Yang et al., 1998).
DNA Extraction and PCR Conditions
DNA from each microcosm was obtained from 0.25 g of soil sample using the Ultra Clean Soil DNA kit (MoBio Lab, Inc.) according to the manufacturer's instructions. PCR reactions contained 10–20 ng template DNA, GoTaq® Green Master Mix (GoTaq® DNA Polymerase in 1x Green GoTaq® Reaction Buffer pH 8.5, 200 μM of each dNTP and 1.5 mM MgCl2) (Promega®) and each primer at the appropriate concentration.
The PCR amplification of the 16S rRNA gene specific for beta-proteobacterias (beta-amo) was carried out by a nested PCR strategy. First, 200 nM of the universal primers fD1 (5′-AGAGTTTGATCCTGGCTCAG-3′) and rP2 (5′-ACGGCTACCTTGTTACGACTT-3′) (Weisburg et al., 1991) were used. The second PCR was assessed using 200 nM of primers βAMOf (5′-TGGGGRATAACGCAYCGAAAG-3′) and βAMOr (5′-AGACTCCGATCCGGACTACG-3′) (McCaig et al., 1994).
The amoA gene of AOB (B-amoA) was amplified also using a nested PCR (Yeager et al., 2005). In the first PCR, the forward primer amoA-2F (5′-AARGCGGCSAAGATGCCGCC-3′) and the reverse primer amoA-5R (5′-TTATTTGATCCCCTC-3′) (Webster et al., 2002) each at 230 nM were used. For the second PCR, forward primer amoA-1F (5′-GGGGTTTCTACTGGTGGT-3′) and reverse primer amoA-2R (5′-CCCCTCKGSAAAGCCTTCTTC-3′) (Rotthauwe et al., 1997) were used each at 660 nM.
The amoA gene of AOA (A-amoA) was amplified using primers Crenamo23f (5′-ATGGTCTGGCTWAGACG-3′) and Crenamo616r (5′-GCCATCCATCTGTATGTCCA-3′) (Tourna et al., 2008), each at 200 nM. For T-RFLP analysis the forward primers βAMOf and Crenamo23f 5′end-labeled with FAM were used.
Clone Libraries
The PCR-products were purified using the Wizard® DNA Clean-Up System (Promega®). The cleaned amplicons were ligated to the vector pTZ57R/T and used to transform Escherichia coli XL1B by means of the InsTAclone™ PCR Cloning Kit (Fermentas®) according to manufacturer's guidelines. Transformants were grown in LB medium (1.0% Tryptone; 0.5% yeast extract; 1.0% NaCl; pH 7.0) and selected through their resistance to ampicillin (50 mg ml−1) and their inability to hydrolyze X-gal in presence of IPTG. The presence of the expected insert size was confirmed by PCR using the vector's primers M13F and M13R. As template a cells suspension in buffer TE subject to eight successive cycles of thermal shock 1 min to 98°C/1 minute to 4°C was used. PCR products of amoA and beta-amo genes were digested using 20U of HhaI and HaeIII (Promega®) restriction enzymes, respectively, and a representative clone of each profile was sequenced (Macrogen Inc.) in one direction using a Genetic Analyzer 3730XL (Applied Biosystems). The clone sequences identity was evaluated using the BLASTN program (Altschul et al., 1997). The sequenced clones were stored in 20% glycerol at −80°C.
Terminal Restriction Fragment Length Polymorphism (T-RFLP) Profiles
The beta-amo and the A-amoA amplicons were purified using the Wizard® DNA Clean-Up System (Promega®) according to the manufacturer's instructions. The purified PCR products (100 ng) of beta-amo were digested in separate reactions with 20 U of HhaI and HaeIII (Promega®) restriction enzymes, while 20 U of HhaI and MnlI (Promega®) were used to digest A-amoA products (100 ng). The terminal restriction fragments (T-RFs) were separated with an automated Genetic Analyzer 3100 (Applied Biosystems, Macrogen Inc.). The lengths of fluorescently labeled T-RFs were determined by the comparison with the internal standard GeneScan-500™ LIZ® using the GeneScan 3.71 software (Applied Biosystems).
The peaks with a fluorescence of 30 U and larger than 30 bp were analyzed by peak height. Patterns from different samples were normalized to identical total fluorescence units by an iterative standardization procedure (Dunbar et al., 2001). This normalization procedure is necessary because of the inherent variability in total DNA quantity among samples on the sequencing gel. Relative abundance of T-RFs, as a percentage, was determined by calculating the ratio between height of a given peak and the normalized total peak height of each sample. Manual alignment of T-RFs profiles was necessary because the Genescan software calculates DNA fragment sizes to 1/100 of a base pair, but the error associated with fragment analysis can be up to 0.5 bp. T-RFs of different lengths were assumed to represent distinct operational taxonomic units (OTUs), although not necessarily unique species.
Most Probable Number—Polymerase Chain Reaction (MPN-PCR) Assay
To estimate the abundance of AOA and AOB, as gene units (GU), the MPN-PCR was used. Ten-fold dilution series, in triplicate, of the extracted DNA from w/t and tH2O microcosms at the beginning and at the end of incubation times were performed. Then, 1 μl was used as template in a 25-μl PCR mixture to amplified B-amoA and A-amoA genes as aforementioned; the PCR products visualized on 1.2% agarose gels staining with ethidium bromide (0.5 mg l−1) were used as the positive signal. MPN values were estimated according to Jarvis et al. (2010) based on a MS-Excel spreadsheet freely available from http://www.wiwiss.fuberlin.de/institute/iso/mitarbeiter/wilrich/index.html.
Statistical Analysis
Edaphic factors and T-RFs data from triplicate microcosms of each treatment were analyzed by multiple comparisons through Two-Way analysis of variance (Two-Way ANOVA) followed by Bonferroni's post-test by the GraphPad Prism v4.0 program (GraphPad Software, Inc.). MPN lower and upper confidence limits were estimated according to Jarvis et al. (2010).
Results
This study was designed to determine the effect of drying-rewetting in a semiarid soil on nitrification activity and nitrifiers structure. The experimental approach consisted of monitoring a 14-days rewetting regime during 56 days of soil microcosms incubation. Edaphic factors such as pH (6.3–7.0) and OM (5.7–6.7%) did not vary with respect to the treatment according to Bonferroni's test (p < 0.05). On the other hand, the MC reflected the treatment effect, with significantly different values for wetted (10.9–15.7%) and non-treated microcosms (2.3–4.4%) (Bonferroni's test, p < 0.01) (Table 1).
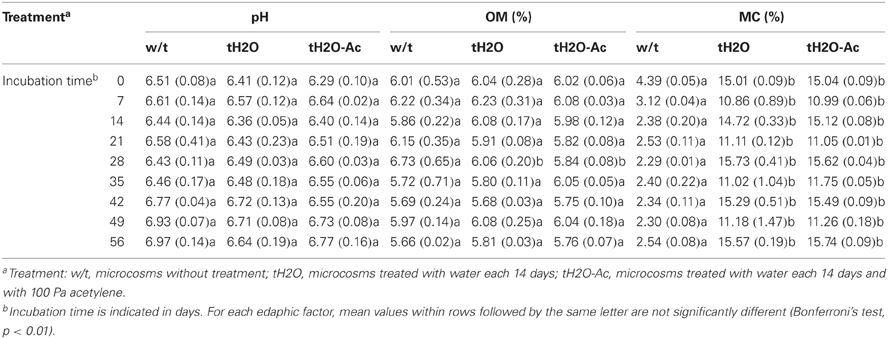
Table 1. Edaphic factors during microcosms incubation time (mean values; numbers in parenthesis are standard deviation).
Net nitrification occurred in rewetted microcosms (tH2O) throughout the incubation period (net nitrification rate: 1.04 μg N-NO3 g−1sdw d−1), in which the nitrate content was above five times higher than in the non-treated samples (w/t) (p < 0.001) (Figure 1). This nitrate accumulation should be the result of nitrate producing and consuming processes. Moreover, this nitrate content increase was inhibited when the wetted soil was incubated in the presence of acetylene (tH2O-Ac) (Figure 1). Acetylene is a potent inhibitor of autotrophic nitrification (Hynes and Knowles, 1982) because the oxidation of acetylene by AMO results in permanent inhibition of the enzyme (Hyman and Wood, 1985). Recently, Offre et al. (2009) also showed that in microcosms containing acetylene, the nitrification of archaeal phylotypes was suppressed. On the other hand, significantly differences (p < 0.05) in ammonium concentration were observed between wetted (tH2O) and control samples (w/t) with respect to wetted microcosms treated with acetylene (tH2O-Ac) (Figure 1). As no ammonium was added to the microcosms, the ammonium is probably continuously regenerated by nitrogen mineralization in a water-dependent process.
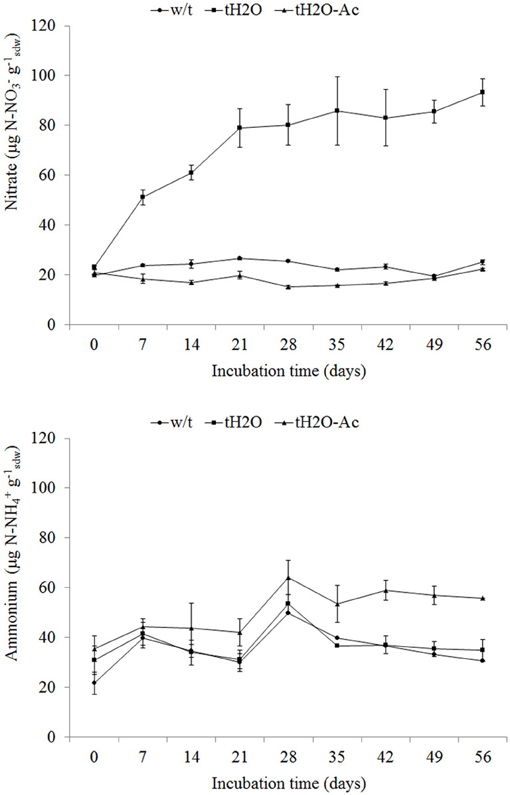
Figure 1. Nitrate (upper panel) and ammonium (lower panel) concentrations (mean values; error bars represent standard deviation) in microcosms without treatment (w/t; circle), microcosms treated with water each 14 days (tH2O; square) and microcosms treated with water each 14 days and 100 Pa acetylene (tH2O-Ac; triangle) during the incubation time. Nitrate concentration data of tH2O microcosms differ from those of w/t and tH2O-Ac microcosms from incubation time of 7 days (p < 0.001; Bonferroni test). Ammonium concentration data of tH2O-Ac microcosms differ from those of w/t and tH2O microcosms from incubation time of 21 days (p < 0.05; Bonferroni's test).
In the next step, the wetted treatment effect on the structure of AOB and/or AOA was assessed. First, the soil AOB composition was determined using two different molecular markers: the bacterial amoA gene (B-amoA) and the beta-proteobacterial 16S rRNA gene (beta-amo). All B-amoA sequences were only related to bacteria of the genus Nitrosospira (Table 2), while clones beta-amo, despite including non-AOB (27.8% of Pseudomonadaceae, 19.4% of Oxalobacteraceae and 8.3% of Comamonadaceae), detected the presence of the two most important groups of AOB (22.2% of Nitrosospira sp. and 5.6% of Nitrosomonas sp.) (Table 2). For this reason, to evaluate the effect of rewetting on the structure of AOB and AOA communities, T-RFLP analysis was assessed using the beta-amo and the A-amoA genes, respectively. The structure of nitrifiers communities was determined from relative abundances of dominant T-RFs. Bacterial and archaeal T-RFLP profiles from wetted and non-treated microcosms obtained with different restriction enzymes, showed similar results with water addition, therefore, only the profiles obtained by digestion with HhaI are showed.
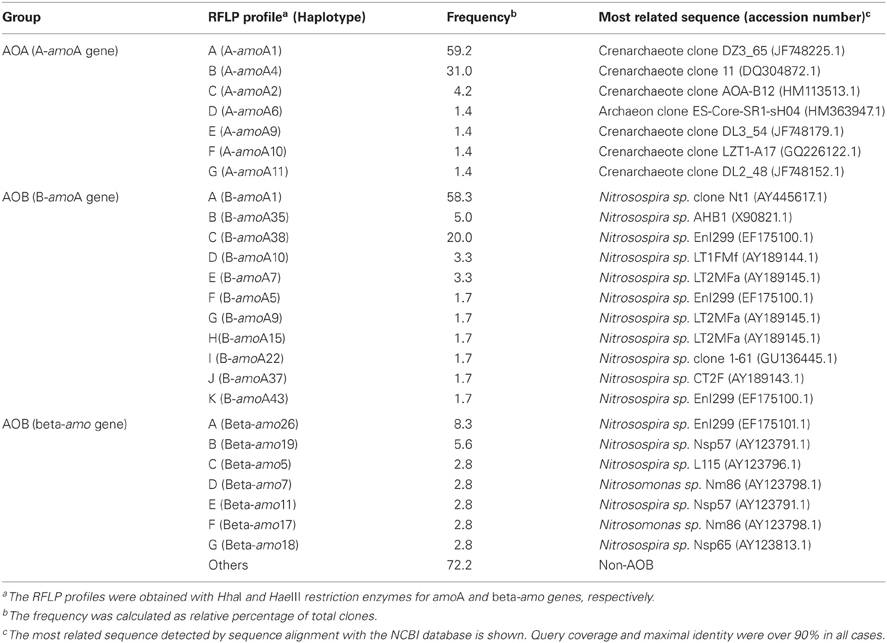
Table 2. Relative frequency of haplotypes in the AOA and AOB gene libraries.
The T-RFLP profiles revealed that the AOA community was significantly affected by treatment. In both microcosms the T-RFs richness was the same at each incubation time, and no significant abundance change was observed for T-RFs of 447, 490, and 596 bp (Figure 2). Among the most significant changes in abundance due to water treatment, are the decline in abundance of T-RFs of 257 and 563 bp from day 28 of incubation, and the increased abundance of the T-RF of 544 bp at the end of incubation. Most T-RFs could not be assigned by in silico restriction to an AOA clone, with the exception of the T-RF of 167 bp which corresponds to the most abundant haplotype A (Table 2). All of haplotypes were related with sequences obtained from soil samples but not with clones derived from extreme environments or marine samples. Summarizing, the AOA structure was affected by the water addition, which could be related to the increase of nitrification activity observed in the treated microcosms.
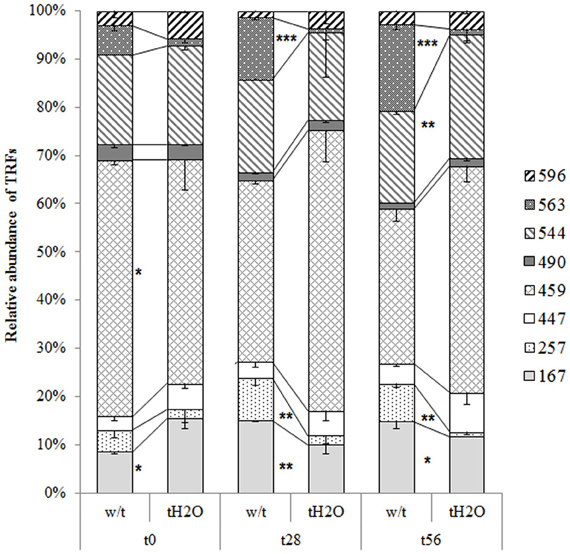
Figure 2. Relative abundances (mean values; error bars represent standard deviation) of the AOA amoA genes T-RFs from microcosms without treatment (w/t) and microcosms treated with water each 14 days (tH2O) during the incubation time. The different textures represent different T-RFs in base pairs (bp). At each incubation time, relative abundances of T-RFs significantly differents are indicated as follows: *p = 0.05; **p = 0.01; ***p = 0.001 (Bonferroni's test).
On the other hand, changes in T-RFLP patterns of AOB between control and treated microcosms could not be observed, indicating that the beta-proteobacterial ammonia-oxidizers community structure remained unchanged under the applied treatment (Figure 3). However, in this case, changes were detected throughout the incubation time when comparing both microcosms; the most notable changes being the increasing T-RF of 71 bp and the decreasing T-RF of 237 bp from day 28 of incubation (Figure 3). The peaks of this profile were identified using the simulation tool MICA (http://mica.ibest.uidaho.edu/) and only two of them could be attributed to ammonia-oxidizers bacteria (T-RF of 437 bp to Nitrosospira sp. and T-RF of 237 bp to Nitrosomonas sp.). However, when the beta-amo genes sequences were digested with HhaI in silico, the T-RF of 71 bp was identified as belonging to Nitrosospira sp., and the association of the T-RF of 237 bp to Nitrosomonas sp. was confirmed, although other non-AOB clones could also produce the same T-RFs.
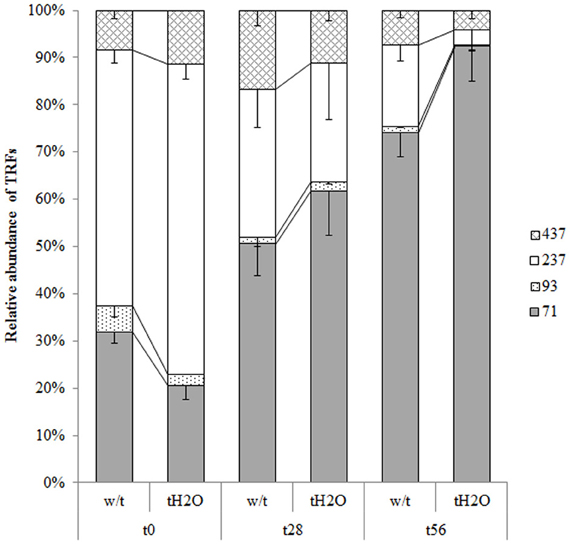
Figure 3. Relative abundances (mean values; error bars represent standard deviation) of the AOB beta-amo genes T-RFs from microcosms without treatment (w/t) and microcosms treated with water each 14 days (tH2O) during the incubation time.The different textures represent different T-RFs in base pairs (bp). At each incubation time, relative abundances of T-RFs are not significantly different (Bonferroni's test, p < 0.05).
Finally, assays were carried out to determine whether the increase of nitrate content in the rewetted soil microcosms was linked to an increase in the number of AOA or AOB. To determine the abundance of AOA and AOB, the MPN-PCR method was used. In this case, the B-amoA molecular marker was used for AOB because of the impossibility of distinguishing the contribution of non-AOB to the gene pool if the beta-amo is used. Therefore, prior to the determination of abundance, the proportion of each haplotype in both groups (AOA and AOB) was calculated (Table 2). All haplotypes of AOA were related to Crenarchaeota clones, the most abundant one (59.2%) was related to an upland red soil under long-term fertilization practices (He et al., 2007). On the other hand, all AOB clones were related to bacteria of the genus Nitrosospira; moreover, several haplotypes were more closely related to sequences from isolated strains. However, the most represented haplotype (58.3%) was associated with a clone from an agriculture soil (Okano et al., 2004).
The MPN-PCR method indicated that the AOB abundances were higher than AOA abundances in both microcosms and incubation times assessed (Table 3). Further, the wetted microcosms (tH2O) showed a significantly higher number of AOA at the end of incubation compared to non-treated microcosms (w/t). However, the abundance of AOB increased with incubation time regardless of treatment and at the end of incubation the number of bacteria was higher in microcosms without treatment (w/t) (Table 3).

Table 3. Abundances of AOA and AOB in the microcosms as determined by MPN-PCR of the corresponding amoA gene (mean values expressed as MPN (UG g−1dws); numbers in parenthesis are lower and upper confidence limits).
Discussion
The aim of this study was to evaluate the effect of periodic rewetting of semiarid soil microcosms from the Chilean sclerophyllous matorral on nitrification and on AOB and AOA communities structure. Episodic water availability affects element cycling in arid and semiarid ecosystems (Schimel et al., 1997; Gebauer and Ehleringer, 2000; Schaeffer and Evans, 2005; Singh and Kashyap, 2006); and many small rain events, as simulated in this study, can cause a rapid increase in soil microorganisms activity (Schwinning and Sala, 2004). Moreover, the increased net nitrification observed in the wetted microcosms (tH2O) agrees with other studies where a positive effect of soil moisture increase on the microbial activity was observed (Stark and Firestone, 1995; Avrahami and Bohannan, 2007; Gleeson et al., 2008, 2010). Additionally, the low nitrate content in the microcosm treated with acetylene supports nitrification observed in the wetted microcosm. Inhibition of soil ammonium oxidation by acetylene has been investigated extensively (McCarty and Bremner, 1986; Garrido et al., 2000) and has been demonstrated for both AOB (Hyman and Wood, 1985) and AOA (Offre et al., 2009). However, it should be noted that net changes in the nitrate pool do not solely reflect nitrifying activity, but it includes the consumption process as the activity carried out by denitrifiers described in this soil (Orlando et al., 2012). Directly linked to the observed nitrification activity, a decrease of ammonium concentrations was observed in the wetted microcosms (tH2O). When acetylene was added to wetted microcosms (tH2O-Ac) to inhibit nitrification activity, ammonium content was higher than in the control microcosms (w/t). As it was not added to the microcosms, the ammonium could be continuously generated by nitrogen mineralization since the frequent soil moisture fluctuations result in pulses of nitrogen mineralization after wetted events as have been described in various reports (Fierer and Schimel, 2002; Austin et al., 2004).
On the other hand, shifts by rewetting of soil in AOA community structure but not in AOB community structure were detected based on T-RFLP profiles. These results agree with those obtained by Offre et al. (2009) which showed that DGGE fingerprint of archaeal amoA gene changed, in the relative abundance of specific archaeal phylotypes, in agricultural soil microcosms with active nitrification incubated during 30 days. In contrast, Jia and Conrad (2009) found no changes in the molecular fingerprint of archaeal amoA gene in fertilized soil microcosms in which ammonium oxidation was stimulated; moreover, the activity changes were correlated with changes in abundance of bacterial amoA gene. In a more recent work, Gleeson et al. (2010) showed that AOA and AOB structures were both affected by changing water filled pore space (WFPS), with the exception of bacterial amoA structure assessed at 25% and 55% WFPS. Previously, Gleeson et al. (2008) showed that water availability affects the AOB structure, and changes in potential nitrification activity were significantly correlated to changes in the structure of nitrifying bacterial communities. In our study, significantly differences in bacterial T-RFLP patterns between control and treated microcosms were not observed, although changes in the relative abundance were detected throughout the incubation time. However, it should be considered that the beta-amo bacterial T-RFs represent not only AOB, but other non-AOB genera; therefore these could be responsible for changes in T-RFLP patterns during incubation. In addition, other studies have suggested that frequent drying and rewetting may select for fast growing microorganisms that are capable of rapid growth on the labile substrates released into the soil during a rewetting event (Lundquist et al., 1999; Denef et al., 2001).
Finally, nitrifiers abundances determined by MPN-PCR have indicated that in this soil AOB GUs per gram were higher than those related to AOA. This prevalence of AOB was also determined by Gleeson et al. (2010) in a semiarid soil of Western Australia, which represents another of the five Mediterranean type environments existing. In contrast, several reports have shown that soil AOA are more abundant than their bacterial counterpart (Leininger et al., 2006; He et al., 2007; Nicol et al., 2008; Shen et al., 2008; Tourna et al., 2008; Offre et al., 2009; Zhang et al., 2010), including another semiarid soils (Adair and Schwartz, 2008); moreover, several reports indicate that the ammonium oxidation is mostly due to the AOA group (Leininger et al., 2006; He et al., 2007; Nicol et al., 2008; Tourna et al., 2008; Offre et al., 2009). In this study, AOB abundance was determined using bacterial amoA gene, and all B-amoA sequences from the clone library were only related to Nitrosospira, which has been detected as the dominant ammonia-oxidizer group in different neutral pH soils (Kowalchuk et al., 2000; Kowalchuk and Stephen, 2001) suggesting that AOB were the main group responsible for ammonium oxidation in some soil types (Jia and Conrad, 2009). This dominance of AOB over AOA could be explained by adaptation to different soil nitrogen conditions, i.e., AOB would be favored by high ammonium concentrations while AOA grow better at very low ammonium concentrations (Di et al., 2010; Pester et al., 2011). The relatively high concentration of ammonium (20–40 μg N-NH+4 per gram of dry soil) detected in the wetted and untreated microcosms of this semiarid soil during the incubation time could explain the higher abundances of AOB compared with AOA. Furthermore, at the end of incubation of both microcosms differences neither in the ammonium content nor in the number of AOB were observed. The source of ammonium may differentially influence AOA and AOB as it was shown that AOA would be favored by ammonium derived from soil OM or in environments with low ammonium concentrations (Stopnišek et al., 2010; Pester et al., 2011; Levičnik-Höfferle et al., 2012) while AOB abundance was shown to increase when soils were amended by mineral nitrogen (Jia and Conrad, 2009). In this work a significant increase in AOB abundance was observed despite rewetting regime, instead AOA increased only slightly in water-treated microcosms at the end of incubation of semiarid soils with low OM but no depletion of ammonium. Likewise, Gleeson et al. (2010) showed that abundance of AOA displayed little response to changes in WFPS availability, although a relationship between water availability and the abundance of bacterial but not archaeal nitrifiers was detected.
In conclusion, the addition of water to these soil microcosms increases the nitrate and ammonium concentration. Results suggest that ammonium was produced through mineralization and was accumulated when nitrification was inhibited by acetylene. Despite water-treatment, communities of AOB had similar structures and abundances, indicating that AOB had little contribution to the nitrification process due to the rewetting regime assessed.
Moreover, the ammonium concentration in dry and wetted soil microcosms could be sufficient to sustain a water-independent nitrification that leads to an increased abundance of AOB at the end of the incubation time. By contrast, changes in structure and abundance of AOA communities in the wetted microcosms suggest that most of the oxidation of the mineralized ammonium, upon water addition, could be carried out by archaea. However, the role of AOA and AOB, their contribution to the nitrification process (Offre et al., 2009) and the environmental factors that define the ecological niche of each group (Tuba et al., 2009; Schleper, 2010) are under discussion at present.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by FONDECYT project N°1080280.
References
Adair, K. L., and Schwartz, E. (2008). Evidence that ammonia-oxidizing archaea are more abundant than ammonia-oxidizing bacteria in semiarid soils of northern Arizona, USA. Microb. Ecol. 56, 420–426.
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., and Lipman, D. J. (1997). Gapped BLAST y PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402.
Austin, A. T., Yahdjian, L., Stark, J. M., Belnap, J., Porporato, A., Norton, U., Ravetta, D. A., and Schaeffer, S. M. (2004). Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 141, 221–235.
Avrahami, S., and Bohannan, B. J. M. (2007). Response of Nitrosospira sp. strain AF-like ammonia oxidizers to changes in temperature, soil moisture content, and fertilizer concentration. Appl. Environ. Microbiol. 73, 1166–1173.
Avrahami, S., Conrad, R., and Braker, G. (2002). Effect of soil ammonium concentration on N2O release and on the community structure of ammonia oxidizers and denitrifiers. Appl. Environ. Microbiol. 68, 5685–5692.
Carú, M. (1993). Characterization of native Frankia strains isolated from Chilean shrubs (Rhamnaceae). Plant Soil 157, 137–145.
Carú, M., Mosquera, G., Bravo, L., Guevara, R., Sepúlveda, D., and Cabello, A. (2003). Infectivity and effectivity of Frankia strains from the Rhamnaceae family on different actinorhizal plants. Plant Soil 251, 219–225.
Denef, K., Six, J., Bossuyt, H., Frey, S. D., Elliott, E. T., Merckx, R., and Paustian, K. (2001). Influence of dry–wet cycles on the interrelationship between aggregate, particulate organic matter, and microbial community dynamics. Soil Biol. Biochem. 33, 1599–1611.
Di, H. J., Cameron, K. C., Shen, J. P., Winefield, C. S., O'Callaghan, M., Bowatte, S., and He, J. Z. (2010). Ammonia oxidizing bacteria and archaea grow under contrasting soil nitrogen conditions. FEMS Microbiol. Ecol. 72, 386–394.
Drenovsky, R. E., Vo, D., Graham, K. J., and Scow, K. M. (2004). Soil water content and organic carbon availability are major determinants of soil microbial community composition. Microb. Ecol. 48, 424–430.
Dunbar, J., Ticknor, L. O., and Kuske, C. R. (2001). Phylogenetic specificity and reproducibility and new method for analysis of terminal restriction fragment profiles of 16S rRNA genes from bacterial communities. Appl. Environ. Microbiol. 67, 190–197.
Fierer, N., and Schimel, J. (2002). Effects of drying–rewetting frequency on soil carbon and nitrogen transformations. Soil Biol. Biochem. 34, 777–787.
Fierer, N., Schimel, J. P., and Holden, P. A. (2003). Influence of drying-rewetting frequency on soil bacterial community structure. Microb. Ecol. 45, 63–71.
Forster, J. C. (1995). “Soil Nitrogen,” in Methods in Applied Soil Microbiology and Biochemistry, eds K. Alef and P. Nannipieri (San Diego, CA: Academic Press Inc.), 79–87.
Francis, C. A., Roberts, K. J., Beman, J. M., Santoro, A. E., and Oakley, B. B. (2005). Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. U.S.A. 102, 14683–14688.
Fuentes, E. R. (1990). “Landscape change in Mediterranean-type habitats of Chile: patterns and processes,” in Changing Landscapes: An Ecological Perspective, eds I. S. Zonneveld and R. T. T. Forman (Berlin, Germany: Springer-Verlag), 165–190.
Garrido, F., Hénault, C., Gaillard, H., and Germon, J. C. (2000). Inhibitory capacities of acetylene on nitrification in two agricultural soils. Soil Biol. Biochem. 32, 1799–1802.
Gebauer, R. L., and Ehleringer, J. R. (2000). Water and nitrogen uptake patterns following moisture pulses in a cold desert community. Ecology 81, 1415–1424.
Gleeson, D. B., Herrmann, A. M., Livesley, S. J., and Murphy, D. V. (2008). Influence of water potential on nitrification and structure of nitrifying bacterial communities in semiarid soils. Appl. Soil Ecol. 40, 189–194.
Gleeson, D. B., Müller, C., Banerjee, S., Ma, W., Siciliano, S. D., and Murphy, D. V. (2010). Response of ammonia oxidizing archaea and bacteria to changing water filled pore space. Soil Biol. Biochem. 42, 1888–1891.
Griffiths, R. I., Whiteley, A. S., O'Donnell, A. G., and Bailey, M. J. (2003). Physiological and community responses of established grassland bacterial populations to water stress. Appl. Environ. Microbiol. 69, 6961–6968.
He, J. Z., Shen, J. P., Zhang, L. M., Zhou, Y. G., Zheng, Y. M., Xu, M. G., and Di, H. (2007). Quantitative analyses of the abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term fertilization practices. Environ. Microbiol. 9, 2364–2374.
Hyman, M. R., and Wood, P. M. (1985). Suicidal inactivation and labelling of ammonia mono-oxygenase by acetylene. Biochem. J. 227, 719–725.
Hynes, R. K., and Knowles, R. (1982). Effect of acetylene on autotrophic and heterotrophic nitrification. Can. J. Microbiol. 28, 334–340.
Jarvis, B., Wilrich, C., and Wilrich, P. T. (2010). Reconsideration of the derivation of Most Probable Numbers, their standard deviations, confidence bounds and rarity values. J. Appl. Microbiol. 109, 1660–1667.
Jia, Z., and Conrad, R. (2009). Bacteria rather than Archaea dominate microbial ammonia oxidation in an agricultural soil. Environ. Microbiol. 11, 1658–1671.
Kowalchuk, G. A., Stienstra, A. W., Heilig, G. H. J., Stephen, J. R., and Woldendorp, J. W. (2000). Composition of communities of ammonium-oxidising bacteria in wet, slightly acid grassland soils using 16S rDNA-analysis. FEMS Microbiol. Ecol. 31, 207–215.
Kowalchuk, G. A., and Stephen, J. R. (2001). Ammonia-oxidizing bacteria: a model for molecular microbial ecology. Annu. Rev. Microbiol. 55, 485–529.
Leininger, S., Urich, T., Schloter, M., Scwark, L., Qi, J., Nicol, G. W., Prosser, J. I., Schuster, S. C., and Schleper, C. (2006). Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nat. Lett. 442, 806–809.
Levičnik-Höfferle, S., Nicol, G. W., Ausec, L., Mandić-Mulec, I., and Prosser, J. I. (2012). Stimulation of thaumarchaeal ammonia oxidation by ammonia derived from organic nitrogen but not added inorganic nitrogen. FEMS Microbiol. Ecol. 80, 114–123.
Lundquist, E. J., Scow, K. M., Jackson, L. E., Uesugi, S. L., and Johnson, C. R. (1999). Rapid response of soil microbial communities from conventional, low input, and organic farming systems to a wet/dry cycle. Soil Biol. Biochem. 31, 1661–1675.
McCaig, A. E., Embley, T. M., and Prosser, J. I. (1994). Molecular analysis of enrichment cultures of marine ammonia oxidizers. FEMS Microbiol. Lett. 120, 363–368.
McCarty, G. W., and Bremner, J. M. (1986). Inhibition of nitrification in soil by acetylenic compounds. Soil Sci. Soc. Am. J. 50, 1198–1201.
Nelson, D. W. (1983). Determination of ammonium in KCl extracts by the salicylate method. Commun. Soil Sci. Plant Anal. 14, 1051–1062.
Nicol, G. W., Leininger, S., Schleper, C., and Prosser, J. I. (2008). The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ. Microbiol. 10, 2966–2978.
Offre, P., Prosser, J. I., and Nicol, G. W. (2009). Growth of ammonia-oxidizing archaea in soil microcosms is inhibited by acetylene. FEMS Microbiol. Ecol. 70, 99–108.
Okano, Y., Hristova, K. R., Leutenegger, C. M., Jackson, L. E., Denison, R. F., Gebreyesus, B., Lebauer, D., and Scow, K. M. (2004). Application of real-time PCR to study effects of ammonium on population size of ammonia-oxidizing bacteria in soil. Appl. Environ. Microbiol. 70, 1008–1016.
Orlando, J., Carú, M., Pommerenke, B., and Braker, G. (2012). Diversity and activity of denitrifiers of Chilean arid soil ecosystems. Front. Microbio. 3:101. doi: 10.3389/fmicb.2012.00101
Pester, M., Schleper, C., and Wagner, M. (2011). The Thaumarchaeota: an emerging view of their phylogeny and ecophysiology. Curr. Opin. Microbiol. 14, 300–306.
Prosser, J. I., and Nicol, G. W. (2008). Relative contributions of archaea and bacteria to aerobic ammonia oxidation in the environment. Environ. Microbiol. 10, 2931–2941.
Purkhold, U., Pommerening-Röser, A., Juretschko, S., Schmid, M. C., Koops, H. P., and Wagner, M. (2000). Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and amoA sequence analysis: implications for molecular diversity surveys. Appl. Environ. Microbiol. 66, 5368–5382.
Rotthauwe, J. H., Witzel, K. P., and Liesack, W. (1997). The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 63, 4704–4712.
Schaeffer, S. M., and Evans, R. D. (2005). Pulse additions of soil carbon and nitrogen affect soil nitrogen dynamics in an arid Colorado Plateau shrubland. Oecologia 145, 425–433.
Schimel, D. S., Braswell, B. H., and Parton, W. J. (1997). Equilibration of terrestrial water, nitrogen and carbon cycles. Proc. Natl. Acad. Sci. U.S.A. 94, 8280–8283.
Schimel, J., Balser, T. C., and Wallenstein, M. (2007). Microbial stress-response physiology and its implications for ecosystem function. Ecology 88, 1386–1394.
Schleper, C. (2010). Ammonia oxidation: different niches for bacteria and archaea? ISME J. 4, 1092–1094.
Schwinning, S., and Sala, O. E. (2004). Hierarchical organization of resource pulse responses in arid and semiarid ecosystems. Oecologia 141, 211–220.
Shen, J. P., Zhang, L. M., Zhu, Y. G., Zhang, J. B., and He, J. Z. (2008). Abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea communities of an alkaline sandy loam. Environ. Microbiol. 10, 1601–1611.
Silvester, W. B., Balboa, O. Y., and Martinez, J. A. (1985). Nodulation and nitrogen fixation in members of the Rhamnaceae (Colletia, Retanilla, Talguenea, Trevoa) growing in the Chilean matorral. Symbiosis 1, 29–38.
Singh, J. S., and Kashyap, A. (2006). Dynamics of viable nitrifier community N-mineralization and nitrification in seasonally dry tropical forests and savanna. Microbiol. Res. 161, 169–179.
Stark, J. M., and Firestone, M. K. (1995). Mechanisms for soil moisture effects on activity of nitrifying bacteria. Appl. Environ. Microbiol. 61, 218–221.
Stopnišek, N., Rangin, C. G., Hoffëler, S., Nicol, G. W., Mandić-Mulec, I., and Prosser, J. I. (2010). Thaumarchaeal ammonia oxidation in an acidic forest peat soil is not influenced by ammonium amendment. Appl. Environ. Microbiol. 76, 7626–7634.
Tourna, M., Freitag, T. E., Nicol, G. W., and Prosser, J. I. (2008). Growth, activity and temperature responses of ammonia-oxidizing archaea and bacteria in soil microcosms. Environ. Microbiol. 10, 1357–1364.
Treusch, A. H., Leininger, S., Schleper, C., Kietzin, A., Klenk, H. P., and Schuster, S. C. (2005). Novel genes for nitrite reductase and Amo-related proteins indicate a role of uncultivated mesophilic crenarchaeota in nitrogen cycling. Environ. Microbiol. 7, 1985–1995.
Tuba, H., Boon, N., Wittebolle, L., and Marzorati, M. (2009). Environmental factors shaping the ecological niches of ammonia-oxidizing archaea. FEMS Microbiol. Rev. 33, 855–869.
Ward, B. B., and O'Mullan, G. D. (2002). Worldwide distribution of Nitrosococcus oceani, a marine ammonia-oxidizing gamma-proteobacterium, detected by PCR and sequencing of 16S rRNA and amoA genes. Appl. Environ. Microbiol. 68, 4153–4157.
Webster, G., Embley, T. M., and Prosser, J. I. (2002). Grassland management regimens reduce small scale heterogeneity and species diversity of α-protobacteria ammonia oxidizer populations. Appl. Environ. Microbiol. 68, 20–30.
Weisburg, W. G., Barns, S. M., Pelletier, D. A., and Lane, D. J. (1991). 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 173, 697–703.
Whitford, W. G. (1992). Biogeochemical consequences of desertification. ACS Symp. Ser. 483, 352–359.
Yang, J. E., Skogley, E. O., Schaff, B. E., and Kim, J. J. (1998). Simple spectrophotometric determination of nitrate in water, resin, and soil extracts. Soil Sci. Soc. Am. J. 62, 1108–1115.
Yeager, C. M., Northup, D. E., Grow, C. C., Barns, S. M., and Kuske, C. R. (2005). Changes in nitrogen-fixing and ammonia-oxidizing bacterial communities in soil of a mixed conifer forest after wildfire. Appl. Environ. Microbiol. 71, 2713–2722.
Keywords: ammonia-oxidizing archaea, ammonia-oxidizing bacteria, water pulses, semiarid soil, microcosms
Citation: Bustamante M, Verdejo V, Zúñiga C, Espinosa F, Orlando J and Carú M (2012) Comparison of water availability effect on ammonia-oxidizing bacteria and archaea in microcosms of a Chilean semiarid soil. Front. Microbio. 3:282. doi: 10.3389/fmicb.2012.00282
Received: 26 January 2012; Accepted: 18 July 2012;
Published online: 08 August 2012.
Edited by:
Lisa Y. Stein, University of Alberta, CanadaReviewed by:
Ines Mandic-Mulec, University of Ljubljana, SloveniaChristoph Mueller, Justus-Liebig-University Giessen, Germany
Copyright © 2012 Bustamante, Verdejo, Zúñiga, Espinosa, Orlando and Carú. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Margarita Carú, Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Casilla 653, Santiago, Chile. e-mail:bWFyZ2FyaXRhX2NhcnVAeWFob28uY29t
† These authors have equally contributed to this work.