

- 1Institute of Functional Microbiology, Department of Pathobiology, University of Veterinary Medicine Vienna, Vienna, Austria
- 2Bavarian Health and Food Safety Authority, Oberschleißheim, Germany
The highly heterogeneous genus Bacillus comprises the largest species group of endospore forming bacteria. Because of their ubiquitous nature, Bacillus spores can enter food production at several stages resulting in significant economic losses and posing a potential risk to consumers due the capacity of certain Bacillus strains for toxin production. In the past, food microbiological diagnostics was focused on the determination of species using conventional culture-based methods, which are still widely used. However, due to the extreme intra-species diversity found in the genus Bacillus, DNA-based identification and typing methods are gaining increasing importance in routine diagnostics. Several studies showed that certain characteristics are rather strain-dependent than species-specific. Therefore, the challenge for current and future Bacillus diagnostics is not only the efficient and accurate identification on species level but also the development of rapid methods to identify strains with specific characteristics (such as stress resistance or spoilage potential), trace contamination sources, and last but not least discriminate potential hazardous strains from non-toxic strains.
Introduction
Proper diagnostic tools are of utmost importance, not only in the field of clinical but also in the field of food and veterinary microbiology diagnostics. Since the first isolation, purification, and cultivation of a pathogenic bacterium, namely Bacillus anthracis, by Koch (1876), the use of solid plating media has become the “golden standard” in classical microbiology and is still the “method of choice” for identification and enumeration of bacteria in routine diagnostic labs. However, general drawbacks of conventional culture-based methods, such as low specificity (poor inclusivity/exclusivity) and low discriminatory power, question their suitability to cope with today’s diagnostic needs. In addition, it has been show that certain characteristics, such as the presence of toxin genes, are rather strain than species-specific. For instance, botulinum neurotoxins are not only produced by C. botulinum but also by some C. baratii and C. butyricum strains, Staphylococcus aureus enterotoxin genes have also been found in other Staphylococcus spp. (e.g., Tsukamoto et al., 2002; De Medici et al., 2009; Oliveira et al., 2010). There are also some studies reporting on the detection of B. cereus enterotoxins in non-B. cereus group Bacillus spp. (Rowan et al., 2001; Phelps and McKillip, 2002) and, more recently, a heat stable toxin, structural related to the B. cereus emetic toxin cereulide, has been found in a Paenibacillus tundrae strain (Rasimus et al., 2012). The capacity for the production of spoilage-associated enzymes, such as proteases and lipases, may also vary significantly among strains of the same species (see, e.g., De Jonghe et al., 2010). Generally, members of the genus Bacillus and related genera show a high inter- and intra-species heterogeneity, which confronts diagnostic labs with various challenges and the urgent need for novel diagnostic concepts.
The most prominent member of the genus Bacillus is the B. cereus group that comprises several genetically closely related species, which are summarized under the term Bacillus cereus sensu lato (Kolsto et al., 2009; Ehling-Schulz et al., 2011a). With the increase of genetic information and the description of more and more so-called “borderline” strains (Klee et al., 2006; Fricker et al., 2008), a refinement of the current nomenclature might become necessary. Due to their different risk potentials, ranging from risk group 1 to risk group 3, the members of the B. cereus group are still handled as separate species. According to their major virulence characteristics, the following species are discriminated: B. anthracis that causes the fatal human and animal disease anthrax, B. thuringiensis that is commercially used as biopesticide (Mock and Fouet, 2001; Bravo et al., 2011), and B. cereus sensu stricto, the name giving species of the group. The latter one is an opportunistic human pathogen that can cause two forms of food poisoning: emesis and diarrhea. The diarrheal form of B. cereus food poisoning resembles the symptoms of a C. perfringens infection and the emetic form parallels the symptoms of intoxications related to S. aureus enterotoxins (for review, see Ehling-Schulz et al., 2004a; Stenfors Arnesen et al., 2008). Normally, both forms of the food borne disease are self-limited but reports on more severe cases, requiring hospitalization and including even death, are currently increasing (e.g., Lund et al., 2000; Dierick et al., 2005; Naranjo et al., 2011; Ehling-Schulz and Messelhaeusser, 2012). In addition, B. cereus can also cause non-gastrointestinal diseases, such as local eye or wound infection, as well as systemic infections, such as bacteraemia and endocarditis (for review, see Bottone, 2010). B. mycoides and B. weihenstephanensis are two psychrotolerant members of the group, which can grow even under refrigerator conditions. The latter two are known for their spoilage potential but their pathogenic potential seems to be rather low (Guinebretière et al., 2008, 2010). Very recently, B. cytotoxicus, a novel thermotolerant member of the group has been described, which may also possess food poisoning potential (Guinebretière et al., 2013).
Because of the medical, food safety and food quality relevance as well as economic importance, several methods for typing of B. cereus s.l. have been developed and comprehensive genome sequencing projects are underway, opening new avenues for diagnostics. This review will provide an overview on state of the art methods for identification and typing of this interesting group of spore formers and will discuss future trends in Bacillus diagnostics, focusing rather on strain characterization than on species identification.
Who is Out there?
The B. cereus group (Figure 1) comprises bacteria that can grow over a wide temperature range and showing quite variable pathogenic potentials, ranging from strains used as plant growth promoters and biopesticides to strains causing fatal diseases.
FIGURE 1. Who is who? – a mixed culture of B. cereus group strains, namely: B. thuringiensis IEBC-T066001 (serotype entomocidus biovar subtoxicus, Canada), B. weihenstephanensis WSBC 10207 (isolated from pasteurized milk, Germany), and B. cereus F4430 (isolated from pea soup connected to a foodborne outbreak in Belgium).
The Classical Way of Bacillus Diagnostic: Cultural Detection, Enumeration, and Differentiation
Cultural detection and enumeration of presumptive B. cereus (B. cereus s.l.), following internationals standards (such as the ISO 7932; Anonymous, 2004), is still the standard procedure for Bacillus diagnostic in food microbiology laboratories. Because of their close genetic relatedness, B. cereus group members cannot be differentiated by classical cultural detection methods or 16S rDNA sequencing and are therefore subsumed as “presumptive B. cereus.”
Isolation and enumeration of presumptive B. cereus from different food matrices is routinely performed using selective plating media. Two standard plating media, the polymyxin-egg yolk-mannitol-bromothymol blue agar (PEMBA) and the mannitol-egg yolk-polymyxin (MYP) agar, are currently recommended by the International Organization for Standardization (ISO) or the Food and Drug Administration (FDA). However, these media bear the risk of substantial misidentifications since various strains, especially if food matrices are analyzed, are showing atypical reaction on these media (Fricker et al., 2008). In the last few years, new chromogenic media have been designed for the detection of B. cereus group members. All these media are based on enzymes that are under regulatory control of the pleiotropic regulator PlcR. However, since molecular polymorphisms in the plcR gene have been found in all strains showing atypical growth characteristics, the concept of the selective plating media using PlcR-regulated enzymatic activities must be generally questioned and should be reconsidered (Fricker et al., 2008).
For the enumeration of presumptive B. cereus two main methods are widely used in food microbiology, colony-count-techniques on different solid agars and the most probable number (MPN)-technique. For colony count methods employing solid media the detection limit is routinely assessed according to international standards, such as the ISO 7932 (Anonymous, 2004). Since routinely 0.1 mL of the sample (fluid material) or the first serial dilution step (solid material) are plated, the theoretical detection limit is about 10 or 100 CFU/g, respectively, but the detection limit can be lowered by the power of ten by plating 1 mL of the sample or first dilution step on three plates of the solid medium. Enumeration procedures can also be combined with a molecular-based differentiation of the isolates based on their toxin gene profiles, enabling a rough estimate of the number of pathogenic B. cereus in the food sample investigated (see following section for details). Since B. cereus is a ubiquitous spore former, its presence cannot totally be avoided in many food products and, from a consumer safety perspective, the determination of the presence and prevalence of toxigenic strains is of special importance. First assays for enumeration of presumptive and/or toxigenic B. cereus strains employing molecular methods, such as qPCR have been described (Martínez-Blanch et al., 2009; Ceuppens et al., 2010; Dzieciol et al., 2013) but their applicability in route diagnostic is still hampered by the fact that the current systems do not allow a differentiation of live and dead bacteria, vegetative cells or spores.
The Good, The Bad, The Ugly: Molecular Tool Box for Typing and Profiling of Strains
Molecular diagnostic tools for B. cereus focus more and more on detection of toxin genes rather than on the differentiation between B. cereus sensu stricto and other members of the B. cereus group. Although for other purposes the determination of the species might be required. Due to the bioterrorism potential B. anthracis various PCR systems for its specific detection and differentiation from the other members of the B. cereus group have been developed. For instance, Leski et al. (2009) used the Bacillus collagen-like protein bcl genes as target sequence whereas Wielinga et al. (2011) integrated a chromosomal marker sequence, target genes located on the two B. anthracis virulence plasmids and an internal amplification control in a probe-based multiplex real-time PCR assay. Since the latter assay targets, beside a B. anthracis-specific chromosomal marker, the coding region of the edema factor gene (cya; encoding an anthrax toxin component) located on pXO1, and the coding region of the capsule synthesis gene capB, located on pXO2, this assay allows a one-step detection and discrimination of different B. anthracis virulence types (Wielinga et al., 2011). Because of the economic importance of B. thuringiensis as biopesticide, PCR systems targeting different parts of the cry insecticidal toxin genes, have been developed during the last two decades (see, e.g., Bourque et al., 1993).
More recently, PCR systems for toxin gene profiling of B. cereus group strains have been developed. The detection of the different toxin genes can either be performed using gel-based PCR or real-time PCR systems (e.g., Guinebretière et al., 2002; Ehling-Schulz et al., 2004b, 2006b; Fricker et al., 2007; Wehrle et al., 2010). Because members of the B. cereus group frequently possess the ability to produce more than one toxin, suitable diagnostic tools for toxin gene profiling should cover the genes encoding the three main enterotoxins, namely the non-hemolytic enterotoxin (Nhe), the hemolysin BL (Hbl), and cytotoxin K (CytK) as well as the emetic toxin cereulide synthetase genes ces. Nhe and Hbl are related three component toxins whereas CytK is single component protein toxin, belonging to the group of β-barrel toxins (for review, see Stenfors Arnesen et al., 2008). The emetic toxin cereulide is a cyclic heat stable depsipeptide produced by the non-ribosomal cereulide peptide synthetase (Ehling-Schulz et al., 2005a, 2006a). Current studies indicate that literarily all B. cereus group strains carry the nhe genes and most of the strains are also able to produce Nhe, although the levels of toxin production very significantly from strains to strains (Moravek et al., 2006; Stenfors Arnesen et al., 2008). Between 44 and 60% of B. cereus strains are able to produce the Hbl toxin (Ehling-Schulz et al., 2005b; Moravek et al., 2006). The ability for CytK production was found in about 40–85% of B. cereus isolates investigated so far (Ngamwongsatit et al., 2008; Stenfors Arnesen et al., 2008). Generally, it seems that certain toxin gene profiles are predominant in specific groups of strains derived from different origin (Ehling-Schulz et al., 2005b, 2006b). For instance, cytK was found in 70% of strains connected to diarrheal food borne outbreaks but it was only rarely found in emetic strains (8%). Recent studies from different continents including isolates from diverse origins indicate the progressive emergence of pathotypes with novel toxin gene profiles (Thorsen et al., 2006; Ehling-Schulz et al., 2011a; Chon et al., 2012), confronting food industry and food microbiology labs with potential novel hazards.
For outbreak investigations of B. cereus s.l., the toxin gene profile might be much more important than the exact species determination. It is well known that not only B. cereus sensu stricto can harbor the toxin genes described above; instead, enterotoxin genes are broadly distributed within the B. cereus group (e.g., Prüss et al., 1999; Guinebretière et al., 2008) and B. thuringiensis has also been described as the possible cause of foodborne outbreaks and other infections, such as local wound and eye infection as well as pulmonary infections (Jackson et al., 1995; Hernandez et al., 1998; Callegan et al., 2006; Ghelardi et al., 2007). Thorsen et al. (2006) reported on a B. weihenstephanensis strain carrying the cereulide synthetase genes and possessing the capability for cereulide toxin formation. Therefore, future developments in food microbiology diagnostics should be more focused on the determination of toxins and virulence factors than on the differentiation of species. Nevertheless, one should bear in mind that the sole presence or absence of an individual toxin gene does not fully explain the pathogenic potential of a certain strain and molecular methods should always be accompanied by sensitive and accurate toxin quantification systems (see, e.g., Bauer et al., 2010). For instance, it has been shown that the toxigenic potential among emetic as well as enterotoxic strains can vary substantially (see, e.g., Moravek et al., 2006; Stark et al., 2013).
Population Studies and Contamination Route Analysis
Digging into Bacillus Populations: PCR-Based Typing Systems
For molecular typing of members of the B. cereus group various PCR-based methods are currently available. Beside random amplification of polymorphic DNA (RAPD)-PCR, REP (repetitive extragenic palindromic)-, ERIC (enterobacterial repetitive intergenic consensus)-, or BOX-PCR can be used for genomic fingerprinting of isolates. For instance, BOX-PCR genomic fingerprinting and also variable-number tandem repeats (VNTR) analysis show the close relationship between B. anthracis and some B. cereus strains (e.g., Kim et al., 2002; Chaves et al., 2011). Since several years, the RAPD-PCR is an established method for molecular typing of different members of Bacillus spp. For instance, RAPD was applied for epidemiological subtyping of B. cereus and B. lentus and for differentiation between B. anthracis and other members of the B. cereus group (Stephan, 1996; Daffonchio et al., 1999). RAPD may also represent an interesting screening method for emetic B. cereus strains also on a routine laboratory basis (Ehling-Schulz et al., 2005b). In summary, RAPD is a valuable and widely used tool for molecular typing of different Bacillus spp. Nevertheless, in contrast to other molecular typing methods, such as multilocus sequence typing (MLST), the interlaboratory reproducibility of data frequently causes difficulties, which might have been one of the reasons why MLST-based systems gradual became the “golden standard” during the last years.
The “Golden Standards” for Population Studies of B. cereus s.l.: Multilocus Sequence Typing and Amplified Fragment Length Polymorphism
Due to high genomic plasticity, the population structure of B. cereus s.l. is quite dynamic and there is potential within this group of bacteria for emergence of new pathogenic lineages with increased or new virulence, or increased ability to survive in adverse environmental conditions (Kolsto et al., 2009; Ehling-Schulz et al., 2011a; Tourasse et al., 2011). An in-depth knowledge of the population structure of B. cereus s.l. is therefore not only of general academic interest but also of great importance for clinical and food microbiology diagnostics. During the last decade various MLST-based schemes for typing of B. cereus s.l. strains have been developed (for overview, see Tourasse and Kolsto, 2008), which have been successfully applied for inferring genetic relationships among B. cereus s.l. strains of different origin, such as soil, insects, food, and humans (Ehling-Schulz et al., 2005b; Vassileva et al., 2006; Cardazzo et al., 2008; Hoffmaster et al., 2008; Raymond et al., 2010). Although the different MLST schemes employ different house keeping genes, all of them revealed three major clades. Interestingly, the same major clusters were found by Fourier transform infrared (FTIR) spectroscopic analysis, pointing toward conserved phenotypic traits of genetic-related strains (Ehling-Schulz et al., 2005b). However, regardless of the typing method used, the different B. cereus group species are interspersed within the different clusters, questioning the suitability of diagnostics solely based on species identification.
One major drawback of MLST-based approaches is the requirement of substantial hands-on-time for sequencing of seven genes per strain and subsequent data analysis, which limits its applicability for high throughput studies. The development of microfluidic biochips might simplify MLST analysis in the future (Read et al., 2010). Currently, the use of the sporulation stage III AB gene (spoIIIAB) as a single genetic marker might represent an alternative to obtain a rough snapshot of genetic relations among B. cereus s.l. strains under study (Ehling-Schulz et al., 2005b). This genetic marker resembles the structure of MLST-derived clusters and its suitability for sequence typing was recently reconfirmed by comparing clusters derived form hierarchical cluster analysis of spoIIIAB sequences with the clusters obtained by whole genome sequencing using a sliding window approach (Fricker et al., 2011; Segerman et al., 2011).
When high throughput capacities are needed, amplified fragment length polymorphism (AFLP) might be the method of choice because it does not require laborious sequencing efforts. For instance, Guinebretière et al. (2008) used AFLP for typing of a comprehensive collection of 425 well-characterized B. cereus group strains derived from very different ecological niches. Seven major clusters (denoted I–VII) were identified, which correlate with physiological properties of the strains. Interestingly, the potential of strains for causing food poisoning correlated with certain phylogenetic groups (Guinebretière et al., 2010). To assign strains to different genetic groups an online tool has been developed, which is available at https://www.tools.symprevius.org/Bcereus/english.php.
In addition, Tourasse et al. (2010) have developed a database called HyperCat, allowing the integration of data from the two different typing systems (MLST, AFLP) described above as well as data derived from multilocus enzyme electrophoresis (MEE), to calculate super trees. HyperCat was applied to carry out a multi-data type analysis on 2213 strains of different origin, including 450 food and dairy production strains. This integrative approach confirmed the major clusters but also revealed some novel phylogenetic branches, including a putative new lineage of B. anthracis (Tourasse et al., 2011). The next step toward a more holistic understanding of this evolutionary interesting and economical important group of microorganisms would be now to include data from functional genomics (transcriptomics, proteomics, and metabolomics).
Bacterial IDs: Fingerprinting Techniques
Molecular Fingerprints
The main fingerprinting technique, the pulsed field gel electrophoresis (PFGE) is used as one of the most important typing method for a wide field of foodborne pathogens, especially for epidemiological studies in outbreak situations. In principle, PFGE can be used for typing of B. cereus (Carlson et al., 1994; Liu et al., 1997; Ohsaki et al., 2007) but PulseNet International, a network for tracking foodborne infections worldwide, does not provide a protocol for molecular typing of B. cereus so far (Swaminathan et al., 2006). However, for epidemiological studies, especially in case of foodborne outbreaks, standard protocols would be mandatory for generating comparable data worldwide. In addition, there are technical difficulties in attaining sufficient chromosomal DNA for macrorestriction of certain strains, especially from food-derived ones. Generally, MLST and AFLP are more commonly used for the differentiation and epidemiological investigations of B. cereus group members than PFGE, and multilocus VNTR analysis (MLVA) and single-nucleotide polymorphism (SNP) analysis are the current “methods of choice” for typing of isolates belonging to the highly monomorphic species B. anthracis (e.g., Keim et al., 2000; Kuroda et al., 2010).
Metabolic Fingerprints
Fourier transform infrared spectroscopy is a powerful tool for microbial diagnostics and epidemiological studies and has already been successfully used to type B. cereus group strains (Ehling-Schulz et al., 2005b; Mietke et al., 2010). Basically, FTIR is a vibrational spectroscopic technique, which is able to distinguish microbial cells at different taxonomic levels (Naumann et al., 1991; Wenning et al., 2008). The entire biochemical composition of whole cells is recorded by the absorbance of mid-infrared light by the molecules present in the cells. The resulting spectra are used as fingerprints and analyzed by pattern recognition techniques. The same spectrum from a microbial sample can be used for identification purposes as well as for typing below the species level. This enables an application in contamination route analysis, epidemiological studies and for determination of specific properties of B. cereus s.l. (Ehling-Schulz et al., 2005b, 2011b). Due to its cost efficiency and high throughput capacities, FTIR spectroscopy represents an interesting alternative to genetic methods for B. cereus subtyping and for tracing contamination sources.
Third Generation Sequencing for Next Generation Diagnostics?
The introduction of massive parallel sequencing in the mid-2000s was a hallmark in genome sequencing, allowing rapid sequencing of DNA on a gigabase scale. The advances in high throughput sequencing technologies during the last years enable the sequencing of microbial genomes in less than 1 day. Concurringly with the upscaling of sequencing capacities the costs per base for sequencing are constantly dropping, thereby opening new perspectives for genome-based diagnostics.
The role of B. anthracis as a potent bioterror agent has lead to a renewed interest in its close relative B. cereus, resulting in several genome sequencing projects. Currently (January 2013), genomic sequence information is available for about 225 B. cereus group strains (Table 1). The list of strains in the sequencing pipeline is steadily growing but the lack of bioinformatic tools is still the bottleneck for a broader application of genome sequence-based diagnostics. Especially for bacteria, such as B. cereus, showing a high rate of genome rearrangements and genomic repeats and transposable elements (Tourasse et al., 2006; Kolsto et al., 2009; Ehling-Schulz et al., 2011a) de novo sequence assembly is laborious and time consuming. Therefore, more and more genomes are left unfinished as permanent draft sequences. Third generation sequencing may help to overcome this obstacle to a certain degree by generating longer reads, facilitating sequence assembly. However, the development of appropriate, user-friendly bioinformatic tools will be the major challenge for implementation of genomics in microbial diagnostics in the upcoming years.
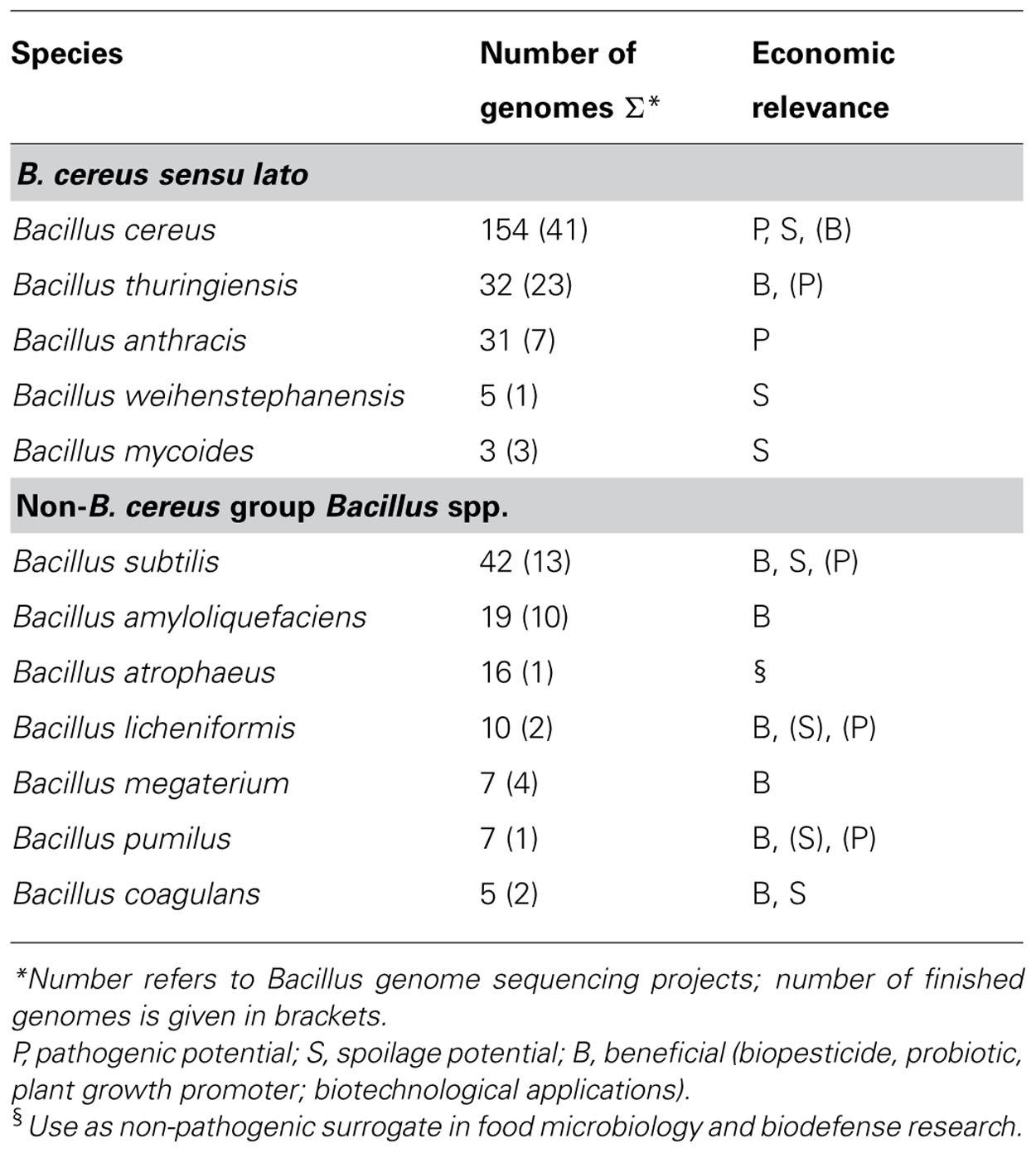
TABLE 1. Overview on Bacillus species for which multiple genomes are public available (http://www.ncbi.nlm.nih.gov/genome, last accessed January 2013)
First tools to minimize post-sequencing data processing have already been developed. For instance, Segerman et al. (2011) used a method that defines orthologous sequence reads instead of orthologous genes for subtyping B. anthracis strains and for obtaining a general overview of the phylogenomic structure of the genus Bacillus. Very recently, Agren et al. (2012) presented a software tool, which uses fragmented alignments to analyze multiple genomes. This software, named after the Greek Argonauts six-armed giant tribe “Gegenees,” is designed as an open platform and can be accessed at http://gegenees.org/index.html. “Gegenees” was successfully used to search for unique signatures for B. anthracis, by analyzing 134 Bacillus genomes. Based on the identified signatures, target group specific primers were designed (Agren et al., 2012).
For comparative genomotyping DNA microarray-based analysis might also be an interesting route to follow. The huge amount of genetic information from recent and ongoing Bacillus sequencing projects makes it feasible to design high-density whole genome microarrays to gain in-depth insights into genetic footprints of strains. For instance, Papazisi et al. (2011) used a multi-genome DNA array to study the genomic diversity of B. cereus s.l. and evolutionary traits of B. anthracis. Such arrays are not only useful to gain insights into the pathophysiology of Bacilli but might also be valuable tools to search for specific strain characteristics, such as stress resistance genes and spoilage-associated genetic determinants.
Conclusion and Future Perspectives
The pathogenic potential among B. cereus group strains ranges from probiotics to highly toxic strains, causing fatal diseases. The discrimination of hazardous strains from harmless, or even beneficial, isolates is therefore the major challenge in future B. cereus diagnostics. It is expected that the currently taxonomic focused diagnostics will gradually be replaced/or complemented by more risk orientated diagnostics. The diagnostic tools, developed during the last decade, for toxin gene profiling and for the determination of specific molecular characteristics as well as for the detection of specific patho- and ecotypes and for quantification of toxins are gaining increasing importance and will lead to a significant improvement of B. cereus diagnostics.
However, the classical cultural methods for detection and enumeration of members of the B. cereus group are still important tools in the field of food microbiology and could complement and cross-validate results from molecular analyses (Figure 2). Because current methods exploited for identification and subtyping of B. cereus s.l. require the isolation of single strains, culture-based methods will still be an intrinsic part of food microbiology diagnostics during the next years.
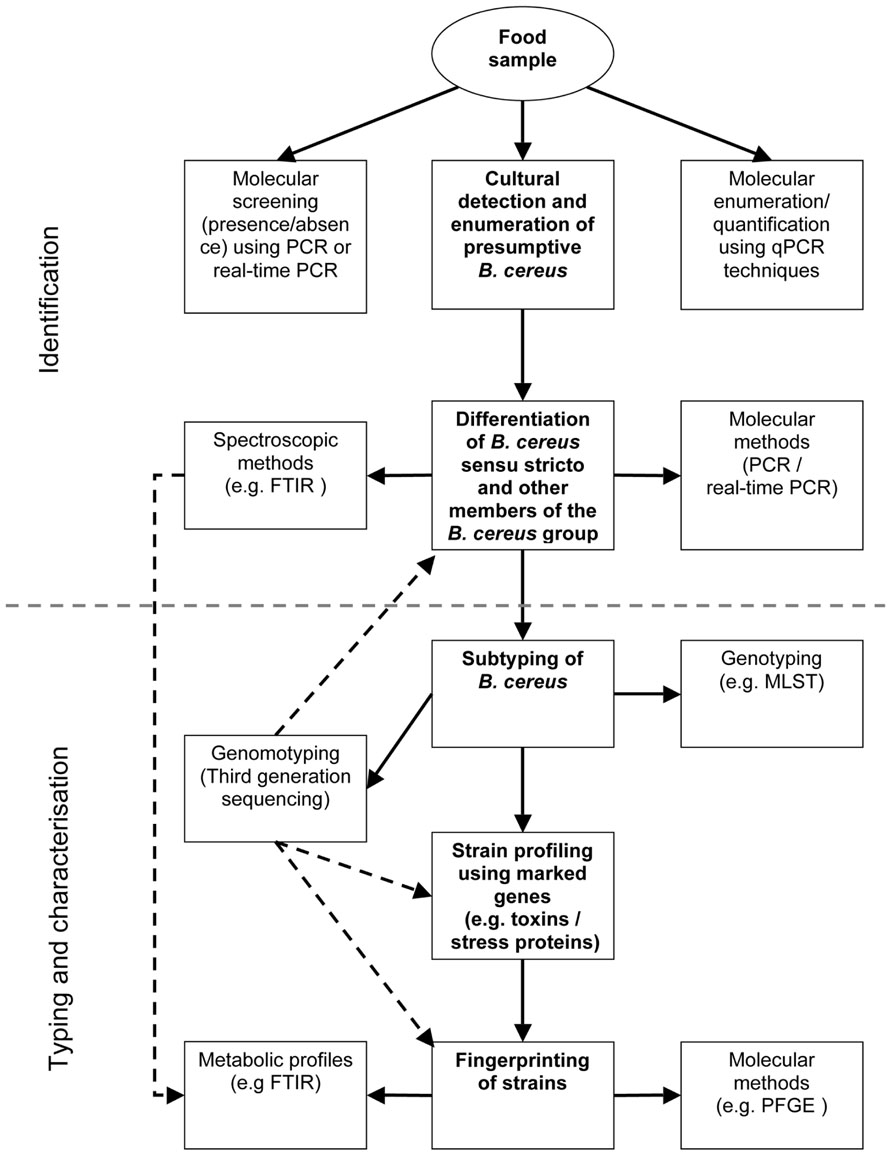
FIGURE 2. Work flow for differential diagnostic of B. cereus s.l. in food samples. - - - - : spectral/sequence data generated once can be used for multiple analyses.
Recent developments in genomotyping are also opening new perspectives for food microbiology diagnostics and are expected to (i) help to decipher specific molecular characteristics of highly pathogenic, food spoiling or beneficial strains and provide biomarkers for a new generation of diagnostics, (ii) foster rapid contamination route analyses, which is getting increasingly important due to globalization of food production, and (iii) facilitate tracing of sources of food borne outbreak by supporting linkage of patient isolates with food-derived isolates. However, before the full potential of next generation sequence-based genomics, or even a part of it, can be exploited for food microbiology diagnostic, appropriate user-friendly bioinformatic tools need to be developed.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Irene Klein for her help with mixed cultures preparations of B. cereus s.l., and acknowledge Stefan Schulz’s assistance in manuscript preparation. This work was supported by the German Ministry of Economics and Technology (via AiF) and the FEI (Forschungskreis der Ernährungsindustrie e.V., Bonn); project AiF 16845N and AiF 17506N.
References
Agren, J., Sundström, A., Håfström, T., and Segerman, B. (2012). Gegenees: fragmented alignment of multiple genomes for determining phylogenomic distances and genetic signatures unique for specified target groups. PLoS ONE 7:e39107. doi: 10.1371/journal.pone.0039107
Anonymous. (2004). Microbiology of Food and Animal Feeding Stuffs – Horizontal Method for the Enumeration of Presumptive Bacillus cereus – Colony-count Technique at 30 Degrees C. Geneva: International Organization for Standardization (ISO).
Bauer, T., Stark, T., Hofmann, T., and Ehling-Schulz, M. (2010). Development of a stable isotope dilution analysis for the quantification of the Bacillus cereus toxin cereulide in foods. J. Agric. Food Chem. 58, 1420–1428.
Bottone, E. J. (2010). Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 23, 382–438.
Bourque, S. N., Valéro, J. R., Mercier, J., Lavoie, M. C., and Levesque, R. C. (1993). Multiplex polymerase chain reaction for detection and differentiation of the microbial insecticide Bacillus thuringiensis. Appl. Environ. Microbiol. 59, 523–527.
Bravo, A., Likitvivatanavong, S., Gill, S. S., and Soberón, M. (2011). Bacillus thuringiensis: a story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 41, 423–431.
Callegan, M. C., Cochran, D. C., Kane, S. T., Ramadan, R. T., Chodosh, J., McLean, C., et al. (2006). Virulence factor profiles and antimicrobial susceptibilities of ocular Bacillus isolates. Curr. Eye Res. 31, 693–702.
Cardazzo, B., Negrisolo, E., Carraro, L., Alberghini, L., Patarnello, T., and Giaccone, V. (2008). Multiple-locus sequence typing and analysis of toxin genes in Bacillus cereus food-borne isolates. Appl. Environ. Microbiol. 74, 850–860.
Carlson, C. R., Caugant, D. A., and Kolstø, A. B. (1994). Genotypic diversity among Bacillus cereus and Bacillus thuringiensis strains. Appl. Environ. Microbiol. 60, 1719–1725.
Ceuppens, S., Boon, N., Rajkovic, A., Heyndrickx, M., Van de Wiele, T., and Uyttendaele, M. (2010). Quantification methods for Bacillus cereus vegetative cells and spores in the gastrointestinal environment. J. Microbiol. Methods 83, 202–210.
Chaves, J. Q., Pires, E. S., and Vivoni, A. M. (2011). Genetic diversity, antimicrobial resistance and toxigenic profiles of Bacillus cereus isolated from food in Brazil over three decades. Int. J. Food Microbiol. 147, 12–16.
Chon, J. W., Kim, J. H., Lee, S. J., Hyeon, J. Y., and Seo, K. H. (2012). Toxin profile, antibiotic resistance, and phenotypic and molecular characterization of Bacillus cereus in Sunsik. Food Microbiol. 32, 217–222.
Daffonchio, D., Borin, S., Frova, G., Gallo, R., Mori, E., Fani, R., et al. (1999). A randomly amplified polymorphic DNA marker specific for the Bacillus cereus group is diagnostic for Bacillus anthracis. Appl. Environ. Microbiol. 65, 1298–1303.
De Jonghe, V., Coorevits, A., De Block, J., Van Coillie. E., Grijspeerdt, K., Herman, L., et al. (2010). Toxinogenic and spoilage potential of aerobic spore-formers isolated from raw milk. Int. J. Food Microbiol. 136, 318–325.
De Medici, D., Anniballi, F., Wyatt, G. M., Lindström, M., Messelhäusser, U., Aldus, C. F., et al. (2009). Multiplex PCR for detection of botulinum neurotoxin-producing clostridia in clinical, food, and environmental samples. Appl. Environ. Microbiol. 75, 6457–6461.
Dierick, K., Van Coillie, E., Swiecicka, I., Meyfroidt, G., Devlieger, H., Meulemans, A., et al. (2005). Fatal family outbreak of Bacillus cereus-associated food poisoning. J. Clin. Microbiol. 43, 4277–4279.
Dzieciol, M., Fricker, M., Wagner, M., Hein, I., and Ehling-Schulz, M. (2013). A diagnostic real-time PCR assay for quantification and differentiation of emetic and non-emetic Bacillus cereus in milk. Food Control 32, 176–185.
Ehling-Schulz, M., Fricker, M., Grallert, H., Rieck, P., Wagner, M., and Scherer, S. (2006a). Cereulide synthetase gene cluster from emetic Bacillus cereus: structure and location on a mega virulence plasmid related to Bacillus anthracis toxin plasmid pXO1. BMC Microbiol. 6:20. doi: 10.1186/1471-2180-6-20
Ehling-Schulz, M., Guinebretiere, M. H., Monthan, A., Berge, O., Fricker, M., and Svensson, B. (2006b). Toxin gene profiling of enterotoxic and emetic Bacillus cereus. FEMS Microbiol. Lett. 260, 232–240.
Ehling-Schulz, M., Fricker, M., and Scherer, S. (2004a). Bacillus cereus, the causative agent of an emetic type of food borne illness. Mol. Nutr. Food Res. 48, 479–487.
Ehling-Schulz, M., Fricker, M., and Scherer, S. (2004b). Identification of emetic toxin producing Bacillus cereus strains by a novel molecular assay. FEMS Microbiol. Lett. 232, 189–195.
Ehling-Schulz, M., Knutsson, R., and Scherer, S. (2011a). “Bacillus cereus” in Genomes of Food- and Water-Borne Pathogens, eds S. Kathariou, P. Fratamico, and Y. Liu (Washington, DC: ASM Press), 147–164.
Ehling-Schulz, M., Messelhäusser, U., and Granum, P. E. (2011b). “Bacillus cereus in milk and dairy production,” in Rapid Detection, Characterization and Enumeration of Food-borne Pathogens, ed. J. Hoorfar (Washington, DC: ASM Press), 275–289.
Ehling-Schulz, M., and Messelhaeusser, U. (2012). “One pathogen but two different types of food borne outbreaks; Bacillus cereus in catering facilities in Germany,” in Case Studies in Food Safety and Quality Management: Lessons from Real-life Situations, ed. J. Hoorfar (Cambridge: Woodhead Publishing), 63–70.
Ehling-Schulz, M., Vukov, N., Schulz, A., Shaheen, R., Andersson, M., Märtlbauer E., et al. (2005a). Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 71, 105–113.
Ehling-Schulz, M., Svensson, B., Guinebretiere, M.-H., Lindbäck, T., Andersson, M., Schulz, A., et al. (2005b). Emetic toxin formation of Bacillus cereus is restricted to a single evolutionary lineage of closely related strains. Microbiology 151, 183–197.
Fricker, M., Ågren, J., Segerman, B., Knutsson, R., and Ehling-Schulz, M. (2011). Evaluation of Bacillus strains as model systems for the work on Bacillus anthracis spores. Int. J. Food Microbiol. 145, S129–S136.
Fricker, M., Messelhäußer, U., Busch, U., Scherer, S., and Ehling-Schulz, M. (2007). Diagnostic real-time PCR assays for the detection of emetic Bacillus cereus strains in foods and recent foodborne outbreaks. Appl. Environ. Microbiol. 73, 1892–1898.
Fricker, M., Reissbrodt, R., and Ehling-Schulz, M. (2008). Evaluation of standard and new chromogenic selective plating media for isolation and identification of Bacillus cereus. Int. J. Food Microbiol. 121, 27–34.
Ghelardi, E., Celandroni, F., Salvetti, S., Fiscarelli, E., and Senesi, S. (2007). Bacillus thuringiensis pulmonary infection: critical role for bacterial membrane-damaging toxins and host neutrophils. Microbes Infect. 9, 591–598.
Guinebretière, M. H., Auger, S., Galleron, N., Contzen, M., De Sarrau, B., De Buyser, M. L., et al. (2013). Bacillus cytotoxicus sp. nov. is a new thermotolerant species of the Bacillus cereus group occasionally associated with food poisoning. Int. J. Syst. Evol. Microbiol. 6, 31–40.
Guinebretière, M. H., Broussolle, V., and Nguyen-The, C. (2002). Enterotoxigenic profiles of food-poisoning and food-borne Bacillus cereus strains. J. Clin. Microbiol. 40, 3053–3056.
Guinebretière, M. H., Thompson, F. L., Sorokin, A., Normand, P., Dawyndt, P., Ehling-Schulz, M., et al. (2008). Ecological diversification in the Bacillus cereus group. Environ. Microbiol. 10, 851–865.
Guinebretière, M. H., Velge, P., Couvert, O., Carlin, F., Debuyser, M. L., and Nguyen-The, C. (2010). Ability of Bacillus cereus group strains to cause food poisoning varies according to phylogenetic affiliation (groups I to VII) rather than species affiliation. J. Clin. Microbiol. 48, 3388–3391.
Hernandez, E., Ramisse, F., Ducoureau, J. P., Cruel, T., and Cavallo, J. D. (1998). Bacillus thuringiensis subsp. konkukian (serotype H34) superinfection: case report and experimental evidence of pathogenicity in immunosuppressed mice. J. Clin. Microbiol. 36, 2138–2139.
Hoffmaster, A. R., Novak, R. T., Marston, C. K., Gee, J. E., Helsel, L., Pruckler, J. M., et al. (2008). Genetic diversity of clinical isolates of Bacillus cereus using multilocus sequence typing. BMC Microbiol. 8:191. doi: 10.1186/1471-2180-8-191
Jackson, S. G., Goodbrand, R. B., Ahmed, R., and Kasatiya, S. (1995). Bacillus cereus and Bacillus thuringiensis isolated in a gastroenteritis outbreak investigation. Lett. Appl. Microbiol. 21, 103–105.
Keim, P., Price, L. B., Klevytska, A. M., Smith, K. L., Schupp, J. M., Okinaka, R., et al. (2000). Multiple-locus variable-number tandem repeat analysis reveals genetic relationships within Bacillus anthracis. J. Bacteriol. 182, 2928–2936.
Kim, W., Hong, Y. P., Yoo, J. H., Lee, W. B., Choi, C. S., and Chung, S. I. (2002). Genetic relationships of Bacillus anthracis and closely related species based on variable-number tandem repeat analysis and BOX-PCR genomic fingerprinting. FEMS Microbiol. Lett. 207, 21–27.
Klee, S. R., Ozel, M., Appel, B., Boesch, C., Ellerbrok, H., Jacob, D., et al. (2006). Characterization of Bacillus anthracis-like bacteria isolated from wild great apes from Cote d’Ivoire and Cameroon. J. Bacteriol. 188, 5333–5344.
Koch, R. (1876). Untersuchungen über Bakterien: V. Die Ätiologie der Milzbrand-Krankheit, begründet auf die Entwicklungsgeschichte des Bacillus anthracis. Cohns Beitr. Biol. Pflanz. 2, 277–310.
Kolsto, A. B., Tourasse, N. J., and Okstad, O. A. (2009). What sets Bacillus anthracis apart from other Bacillus species? Annu. Rev. Microbiol. 63, 451–476.
Kuroda, M., Serizawa, M., Okutani, A., Sekizuka, T., Banno, S., and Inoue, S. (2010). Genome-wide single nucleotide polymorphism typing method for identification of Bacillus anthracis species and strains among B. cereus group species. J. Clin. Microbiol. 48, 2821–2829.
Leski, T. A., Caswell, C. C., Pawlowski, M., Klinke, D. J., Bujnicki, J. M., Hart, S. J., et al. (2009). Identification and classification of bcl genes and proteins of Bacillus cereus group organisms and their application in Bacillus anthracis detection and fingerprinting. Appl. Environ. Microbiol. 75, 7163–7172.
Liu, P. Y., Ke, S. C., and Chen, S. L. (1997). Use of pulsed-field gel electrophoresis to investigate a pseudo-outbreak of Bacillus cereus in a pediatric unit. J. Clin. Microbiol. 35, 1533–1535.
Lund, T., De Buyser, M. L., and Granum, P. E. (2000). A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Mol. Microbiol. 8, 254–261.
Martínez-Blanch, J. F., Sánchez, G., Garay, E., and Aznar, R. (2009). Development of a real-time PCR assay for detection and quantification of enterotoxigenic members of Bacillus cereus group in food samples. Int. J. Food Microbiol. 35, 15–21.
Mietke, H., Beer, W., Schleif, J., Schabert, G., and Reissbrodt, R. (2010). Differentiation between probiotic and wild-type Bacillus cereus isolates by antibiotic susceptibility test and Fourier transform infrared spectroscopy (FT-IR). Int. J. Food. Microbiol. 140, 57–60.
Moravek, M., Dietrich, R., Buerk, C., Broussolle, V., Guinebretière, M. H., Granum, P. E., et al. (2006). Determination of the toxic potential of Bacillus cereus isolates by quantitative enterotoxin analyses. FEMS Microbiol. Lett. 257, 293–298.
Naranjo, M., Denayer, S., Botteldoorn, N., Delbrassinne, L., Veys, J., Waegenaere, J., et al. (2011). Sudden death of a young adult associated with Bacillus cereus food poisoning. J. Clin. Microbiol. 249, 4379–4381.
Naumann, D., Helm, D., and Labischinski, H. (1991). Microbiological characterizations by FT-IR spectroscopy. Nature 351, 81–82.
Ngamwongsatit, P., Buasri, W., Pianariyanon, P., Pulsrikarn, C., Ohba, M., Assavanig, A., et al. (2008). Broad distribution of enterotoxin genes (hblCDA, nheABC, cytK, and entFM) among Bacillus thuringiensis and Bacillus cereus as shown by novel primers. Int. J. Food Microbiol. 121, 352–356.
Ohsaki, Y., Koyano, S., Tachibana, M., Shibukawa, K., Kuroki, M., Yoshida, I., et al. (2007). Undetected Bacillus pseudo-outbreak after renovation work in a teaching hospital. J. Infect. 54, 617–622.
Oliveira, A. M., Miya, N. T., Sant’Ana, A. S., and Pereira, J. L. (2010). Behavior and enterotoxin production by coagulase negative Staphylococcus in cooked ham, reconstituted skimmed milk, and confectionery cream. J. Food Sci. 75, M475–M481.
Papazisi, L., Rasko, D. A., Ratnayake, S., Bock, G. R., Remortel, B. G., Appalla, L., et al. (2011). Investigating the genome diversity of B. cereus and evolutionary aspects of B. anthracis emergence. Genomics 298, 26–39.
Phelps, R. J., and McKillip, J. L. (2002). Enterotoxin production in natural isolates of Bacillaceae outside the Bacillus cereus group. Appl. Environ. Microbiol. 68, 3147–3151.
Prüss, B. M., Francis, K. P., von Stetten, F., and Scherer, S. (1999). Correlation of 16S ribosomal DNA signature sequences with temperature-dependent growth rates of mesophilic and psychrotolerant strains of the Bacillus cereus group. J. Bacteriol. 181, 2624–2630.
Rasimus, S., Mikkola, R., Andersson, M. A., Teplova, V. V., Venediktova, N., Ek-Kommonen, C., et al. (2012). Psychrotolerant Paenibacillus tundrae isolates from barley grains produce new cereulide-like depsipeptides (paenilide and homopaenilide) that are highly toxic to mammalian cells. Appl. Environ. Microbiol. 78, 3732–3743.
Raymond, B., Wyres, K. L., Sheppard, S. K., Ellis, R. J., and Bonsall, M. B. (2010). Environmental factors determining the epidemiology and population genetic structure of the Bacillus cereus group in the field. PLoS Pathog 6:e1000905. doi: 10.1371/journal.ppat.1000905
Read, T. D., Turingan, R. S., Cook, C., Giese, H., Thomann, U. H., Hogan, C. C., et al. (2010). Rapid multi-locus sequence typing using microfluidic biochips. PLoS ONE 5:e10595. doi: 10.1371/journal.pone.0010595
Rowan, N. J., Deans, K., Anderson, J. G., Gemmell, C. G., Hunter, I. S., and Chaithong, T. (2001). Putative virulence factor expression by clinical and food isolates of Bacillus spp. after growth in reconstituted infant milk formulae. Appl. Environ. Microbiol. 67, 3873–3881.
Segerman, B., DeMedici, D., Ehling-Schulz, M., Fach, P., Fenicia, L., Fricker, M., et al. (2011). Bioinformatic tools for using whole genome sequencing as a rapid high resolution diagnostic typing tool when tracing bio-terror organisms in the food and feed chain. Int. J. Food Microbiol. 145, S167–S176.
Stark, T., Marxen, S., Rütschle, A., Lücking, G., Scherer, S., Ehling-Schulz, M., et al. (2013). Mass spectrometric profiling of Bacillus cereus strains and quantitation of the emetic toxin cereulide by means of stable isotope dilution analysis and HEp-2 bioassay, Anal. Bioanal. Chem. 405, 191–201.
Stenfors Arnesen, L. P., Fagerlund, A., and Granum, P. E. (2008). From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 32, 579–606.
Stephan, R. (1996). Randomly amplified polymorphic DNA (RAPD) assay for genomic fingerprinting of Bacillus cereus isolates. Int. J. Food Microbiol. 31, 311–316.
Swaminathan, B., Gerner-Smidt, P., Ng, L. K., Lukinmaa, S., Kam, K. M., Rolando, S., et al. (2006). Building PulseNet International: an interconnected system of laboratory networks to facilitate timely public health recognition and response to foodborne disease outbreaks and emerging foodborne diseases. Foodborne Pathog. Dis. 3, 36–50.
Thorsen, L., Hansen, B. M., Nielsen, K. F., Hendriksen, N. B., Phipps, R. K., and Budde, B. B. (2006). Characterization of emetic Bacillus weihenstephanensis, a new cereulide-producing bacterium. Appl. Environ. Microbiol. 72, 5118–5121.
Tourasse, N. J., Helgason, E., Klevan, A., Sylvestre, P., Moya, M., Haustant, M., et al. (2011). Extended and global phylogenetic view of the Bacillus cereus group populationj by combination of MLST, AFLP, and MLEE genotyping data. Food Microbiol. 28, 236–244.
Tourasse, N. J., Helgason, E., Økstad, O. A., Hegna, I. K., and Kolstø, A. B. (2006). The Bacillus cereus group: novel aspects of population structure and genome dynamics. J. Appl. Microbiol. 101, 579–593.
Tourasse, N. J., and Kolsto, A. B. (2008). SuperCAT: a supertree database for combined and integrative multilocus sequence typing analysis of the Bacillus cereus group of bacteria (including B. cereus, B. anthracis and B. thuringiensis). Nucleic Acids Res. 36, D461–D468.
Tourasse, N. J., Okstad, O. A., and Kolstø, A. B. (2010). HyperCAT: an extension of the SuperCAT database for global multi-scheme and multi-datatype phylogenetic analysis of the Bacillus cereus group population. Database (Oxford) 2010, baq017.
Tsukamoto, K., Mukamoto, M., Kohda, T., Ihara, H., Wang, X., Maegawa, T., et al. (2002). Characterization of Clostridium butyricum neurotoxin associated with food-borne botulism. Microb. Pathog. 33, 177–184.
Vassileva, M., Torii, K., Oshimoto, M., Okamoto, A., Agata, N., Yamada, K., et al. (2006). Phylogenetic analysis of Bacillus cereus isolates from severe systemic infections using multilocus sequence typing scheme. Microbiol. Immunol. 50, 743–749.
Wehrle, E., Didier, A., Moravek, M., Dietrich, R., and Märtlbauer, E. (2010). Detection of Bacillus cereus with enteropathogenic potential by multiplex real-time PCR based on SYBR Green I. Mol. Cell. Probes 24, 124–130.
Wenning, M., Scherer, S., and Naumann, D. (2008). “Infrared spectroscopy in the identification of microorganisms,” in Vibrational Spectroscopy for Medical Diagnosis, eds M. P. Diem, R. Griffiths, and J. M. Chalmers (Chichester: John Wiley & Sons Ltd), 71–96.
Keywords: Bacillus cereus, Bacillus toxin, enterotoxins, cereulide, toxin gene profiling, MLST, fingerprinting, food poisoning
Citation: Ehling-Schulz M and Messelhäusser U (2013) Bacillus “next generation” diagnostics: moving from detection toward subtyping and risk-related strain profiling. Front. Microbiol. 4:32. doi: 10.3389/fmicb.2013.00032
Received: 20 October 2012; Paper pending published: 02 January 2013;
Accepted: 05 February 2013; Published online: 22 February 2013.
Edited by:
Danilo Ercolini, Università degli Studi di Napoli Federico II, ItalyReviewed by:
Folarin Anthony Oguntoyinbo, University of Lagos, NigeriaLouis Coroller, Université de Brest, France
Copyright: © 2013 Ehling-Schulz and Messelhäusser. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Monika Ehling-Schulz, Institute of Functional Microbiology, Department of Pathobiology, University of Veterinary Medicine Vienna, Veterinaerplatz 1, 1210 Vienna, Austria. e-mail:bW9uaWthLmVobGluZy1zY2h1bHpAdmV0bWVkdW5pLmFjLmF0