


- Unité des Aspergillus, Département de Parasitologie et Mycologie, Institut Pasteur, Paris, France
In Aspergillus fumigatus like in other filamentous ascomycetes, β-1,3-glucan constitutes a prominent cell wall component being responsible for rigidity of the cell wall structure. In filamentous fungi, softening of the cell wall is absolutely required during conidial germination and hyphal branching. Because of the central structure of β-1,3-glucans, it is expected that β-1,3-glucanases play a major role in cell wall softening. Based on in silico and experimental data, this review gives an overview of β-1,3-glucan modifying enzymes in A. fumigatus genome and their putative role during morphogenesis.
Introduction
The cell wall of Aspergillus fumigatus is predominantly composed of polysaccharides. The central fibrillar core of the cell wall is composed of a polymer of β-1,3-glucan which is a branched glucan with 4% β-1,6 branch points, to which the chitin, the galactomannan and β-1,3-1,4-glucan are covalently bound (Fontaine et al., 2000). All these polysaccharides are specific of fungal cells, and constitute good antifungal target.
β-1,3-glucans of A. fumigatus are synthesized by a plasma membrane-bound glucan synthase complex, which uses UDP-glucose as a substrate and extrudes linear β-1,3-glucan chains through the membrane into the periplasmic space (Beauvais et al., 1993, 2001). Upon arrival in the cell wall space, newly synthesized polysaccharides are remodeled in order to be incorporated in the pre-existing cell wall. An example of the modifications that occur is given in Figure 1. Steps involved in the remodeling process are structural modifications of the linear glucan chains, produced by the glucan synthase complex, followed by branching, elongation, and degradation of the β-1,3-glucan of the cell wall. Morphogenetic events like conidial swelling (the isodiametral growth phase of germination) germ tube emergence and the production of lateral hypha during mycelial growth require that the cell wall loses its rigidity to allow the emergence of an “additional cell.” Two different biochemical events have been suggested to be associated with germination: the first one is an increased intracellular osmotic pressure (D'Enfert, 1997) and the second one is the softening of the existing cell wall by glycosylhydrolases.
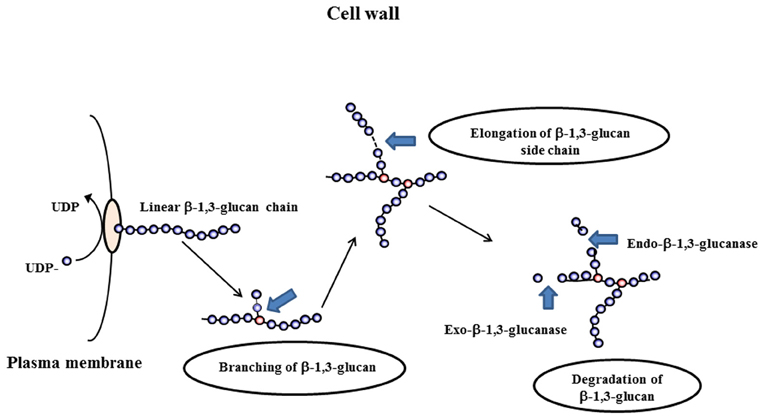
Figure 1. Differents steps occurring during the remodeling process of the cell wall: branching of β(1–3)glucan, elongation and degradation of β(1–3)glucan chains.
Osmotic pressure is high in resting conidia due to their high content of trehalose and polyol. The molecules are, however, fully degraded during the swelling of the conidium which suggests the occurrence of another biochemical structural modification event to facilitate the remodeling of the cell wall during germination. Glycosylhydrolases are the candidates of choice for these cell wall modifications.
Here we present a survey of potential endo-β-1,3-glucanases and exo-β-1,3-glucanases found in the A. fumigatus genome as well as the already identified glucanases and glucanosyltransferase activities described in A. fumigatus (Table 1). A survey of β-1,3-glucanases was undertaken in the genomes of A. fumigatus strain Af293 (Nierman et al., 2005) using the cadre genome (http://www.cadre-genomes.org.uk/Aspergillus_fumigatus) and the Carbohydrate-Active enZYme (CAZy) Database (http://www.cazy.org/).
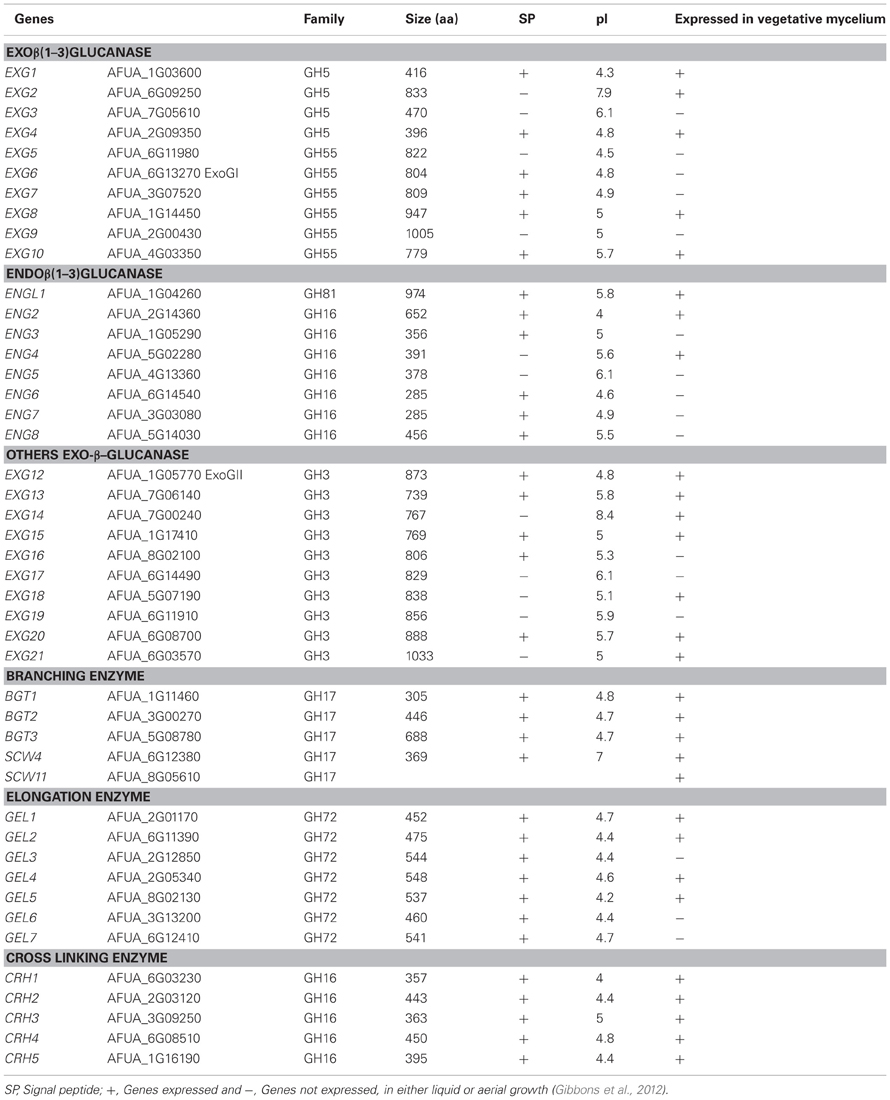
Table 1. AFUA-number of A. fumigatus genes from Af293 strain (http://www.cadre-genomes.org.uk/Aspergillus_fumigatus) and characteristics of modifying cell wall β-1,3-glucanases of A. fumigatus.
Are β–Glucanase Involved in the Cell Wall Modification?
β-1,3-Glucan hydrolyzing enzymes can be divided into exo-β-1,3-glucanases and endo-β-1,3-glucanases. Endo-β-1,3-glucanase activities cleave inside a glucan chain in a more or less random fashion, while the exo-β-1,3-glucanase activities release glucose residues from the non-reducing end (Figure 1). Theoretically, the cleavage of long chains of β-1,3-glucan should cause a softening of the cell wall. Therefore, we expect this type of enzyme to be involved in cell wall remodeling that occur during the swelling and germination of conidia or the branching of hyphae.
In order to identify all the putative β-1,3-glucanases in the A. fumigatus genome sequenced strain Af293 (Nierman et al., 2005), we used the classification of the Carbohydrates enZYmes database [CAZy database (http://www.cazy.org/)]. This database describes the families of enzymes with structurally related catalytic and carbohydrate binding modules (CBM) that degrade, modify or create glycosidic bonds. In this classification, we selected and further investigated the families which could contain putative β-1,3-glucanases and each potential endo/exo β-1,3-glucanase identified in the A. fumigatus genome was used as a query for BLAST searches to identify possible homologs using cadre genome (http://www.cadre-genomes.org.uk/Aspergillus_fumigatus). This search has led to the identification of twenty nine potential exo-β-1,3-glucanases and endo-β-1,3-glucanases that belong to the CAZy GH families 3, 5, 16, 55, and 81 (Table 1). The expression of these different genes in mycelial growth either in liquid or aerial conditions (Gibbons et al., 2012) has been shown in Table 1. The molecular characteristics and the phylogenetic tree of each of the proteins belonging to these families have been investigated.
Exo-β-1,3-Glucanase Families
In the CAZy database, family GH5 represent a group of hydrolases with different substrate specificities such as for example exo β-1,3-glucanases, endo β-1,4-glucanases, endo β-1,6-glucanases. Four proteins belonging to GH5 family in A. fumigatus have been identified in the genome (AfExg1p = AFUA_1G03600, AfExg2p = AFUA_6G09250, AfExg3p = AFUA_7G05610, and AfExg4p = AFUA_2G09350). The characteristics of these proteins are presented in Figure 2A and Table 1.
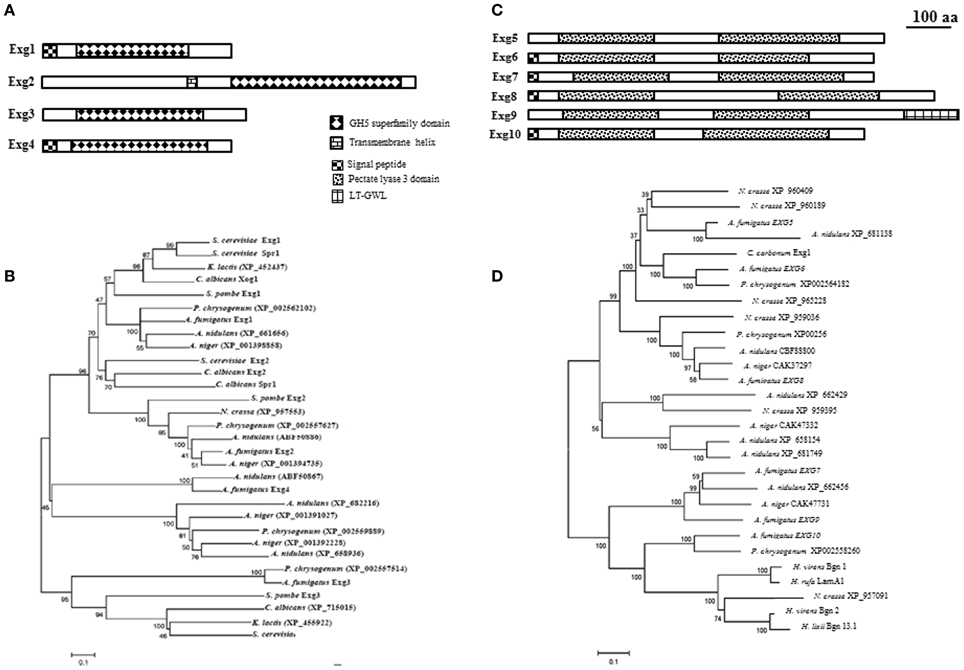
Figure 2. Domain organization and phylogeny of A. fumigatus exo β-1,3-glucanases. Protein domains identified using SignalP 3.0, big-PI, TMHMM v. 2.0, and CD search (100 aa: protein length proportional to 100 amino acids). Phylogenetic trees of A. fumigatus were built by Neighbor Joining. Numbers next to nodes represent bootstrap values calculated using 1000 repetitions. A. fumigatus proteins were aligned with orthologs identified in the filamentous fungi Aspergillus nidulans, Aspergillus niger, Penicillium chrysogenum, Neurospora crassa, Magnaporthe grisea, and the yeasts Saccharomyces cerevisiae, Schizosaccharomyces pombe, Kluyveromyces lactis, and Candida albicans. (A) GH5 family domain organization. (B) Phylogeny of GH5 family, (C) GH55 family domain organization, and (D) Phylogeny of GH55 glucanases additionally contains sequences of Cochliobolus carbonum EXG1 (Schaeffer et al., 1994), Hypocrea rufa LamA1 (Nobe et al., 2004), Hypocrea virens Bgn1 and Bgn2 (Kim et al., 2002), and Hypocrea lixii Bgn13.1 (De la Cruz et al., 1995).
AfExg1p and AfExg4p have a signal peptide in contrast to the others AfExg2p and AfExg3p. The phylogenetic tree of GH5 proteins is presented in Figure 2B. Recently the respective orthologs in S. pombe have been described (Duenas-Santero et al., 2010). SpExg1p is secreted, SpExg2p is bound to the membrane via a GPI anchor and SpExg3p is a cytoplasmic protein. Interestingly, in S. cerevisiae and C. albicans, it has been shown that Exg1p and Xog1p are able to degrade β-1,3-glucan (Vazquez de Aldana et al., 1991; Chambers et al., 1993) and display exo β-1,3-glucanase activities but Suzuki et al. (2001) showed also that the recombinant ScExg1p was able to cleave also β-1,6-glucan. SpExg1 and SpExg3 are only acting on β-1,6-glucans, while no activity was detected for SpExg2 (Duenas-Santero et al., 2010). In S. cerevisiae, the third ortholog SSG1 has been shown to code for a sporulation specific exo-β-1,3-glucanase activity (San Segundo et al., 1993). In S. cerevisiae, the deletion of SSG1 showed a significant delay in the appearance of mature asci (San Segundo et al., 1993). In contrast, no growth phenotype for the single exg1 and exg2 deletion strains of S. cerevisiae has been observed (Larriba et al., 1995). Moreover, the triple ssg1 exg1 exg2 mutants behave like the simple ssg1 mutant. In S. pombe, the phenotype of the triple exg mutants behave like the parental strain (Duenas-Santero et al., 2010). None of the EXG1–4 gene has been deleted or otherwise characterized in A. fumigatus.
In the CAZy database, family GH55 includes exo-β-1,3-glucanases (EC 3.2.1.58) and endo-β-1,3-glucanases (EC 3.2.1.39). In A. fumigatus, six proteins belonging to this family are present: AfExg5p = AFUA_6G11980, AfExg6p = AFUA_6G13270, AfExg7p = AFUA_3G07520, AfExg8p = AFUA_1G14450, AfExg9p = AFUA_2G00430, AfExg10 = AFUA_4G03350, (Table 1).
It has been shown that AfEXG6 encodes a 82 kDa exo β-1,3-glucanase enzyme named ExoGI which hydrolyze exclusively a β-1,3-glucan chain with a minimal substrate size of four glucose residues (Fontaine et al., 1997a). The characteristics of the A. fumigatus proteins are shown in Figure 2C and Table 1. AfExg6p, AfExg7p, AfExg8p and AfExg10p feature a signal peptide, while AfExg5p and AfExg9p are intracellular proteins. AfExg9p features an additional LT_GEWL (Lytic Transglycosylase and Goose Egg White Lysozyme) domain. The exoglucanases AfExg5p to AfExg10p all feature two domains with similarity to the pectate lyase 3 domain. GH55 exoglucanases are not found in yeasts like S. cerevisiae, C. albicans and S. pombe. The protein sequences of the orthologous genes in Cochliobolus carbonum Exg1 (Schaeffer et al., 1994), Hypocrea rufa LamA1 (Nobe et al., 2004), Hypocrea virens Bgn1 and Bgn2 (Kim et al., 2002), and Hypocrea lixii Bgn13.1 (De la Cruz et al., 1995) were added to the alignment of the GH55 family to construct a phylogenetic tree (Figure 2D). It has been shown that C. carbonum Exg1p and H. rufa LamA1p encode for specific exo-β-1,3-glucanases (Nobe et al., 2004) in contrast to Bgn1p, Bgn2p and Bgn13.1p which encode for specific endo-β-1,3-glucanases (De la Cruz et al., 1995; Kim et al., 2002). To date, the biological function of the A. fumigatus GH55 glycosyl-hydrolases is unknown since none of these genes have been deleted yet.
Endo-β-1,3-Glucanase Families
The specificity of the members of the family GH16 in the CAZy database is quite wide since members of this GH family can act as xyloglucan xyloglucosyltransferases (EC 2.4.1.207), keratan-sulfate endo-1,4-β-galactosidases (EC 3.2.1.103), endo-1,3-β-glucanase (EC 3.2.1.39), endo-1,3(4)-β-glucanase (EC 3.2.1.6); licheninase (EC 3.2.1.73), β-agarase (EC 3.2.1.81), κ-carrageenase (EC 3.2.1.83), or xyloglucanase (EC 3.2.1.151). The GH16 superfamily has been divided into nine sub-families based on a phylogenetic analysis of conserved domains of members of this superfamily (cd00413; http://www.ncbi.nlm.nih.gov/Structure/cdd/cddsrv.cgi?uid=29534): GH16_lichenases (cd02175), GH16_XETs (cd02176), GH16_kappa_carrageenases (cd02177), GH16_beta_agarases (cd02178), GH16_beta_GRPs (cd02179), GH16_laminarinases (cd02180), GH16_MLG1_glucanases (mixed-linked glucanases, cd02181), GH16_laminarinase_like proteins (cd02182), and GH16_GPI_glucanosyltransferases (cd02183).
In A. fumigatus, seven members AfENG2 to AfENG8 are present in the genome (Table 1, Figure 3A). While AfEng2p to AfEng5p belong to the subfamily GH16_MLG1_glucanases, and AfEng6p and AfEng7p to the subfamily GH16_laminarinase_like proteins, AfEng8p was not assigned to a GH16 subfamily. The cluster of GH16_MLG1_glucanases from AfEng2p to AfEng5p contains four proteins which all share the same catalytic region but aside from that feature different protein domains. AfEng2p and AfEng3p both exhibit an N-terminal signal peptide but only AfEng2p, which is considerably larger than its three homologs, holds a GPI signal sequence. AfEng4p and AfEng5p on the other hand are not predicted to contain a signal peptide but both feature a transmembrane helix, suggesting a localization at the plasma membrane with the active domain pointing toward the cell wall. So while these proteins vary in terms of protein domains all four of them might still be localized close to the cell wall, either by being secreted or by being bound to the plasma membrane by a GPI anchor or a transmembrane domain.
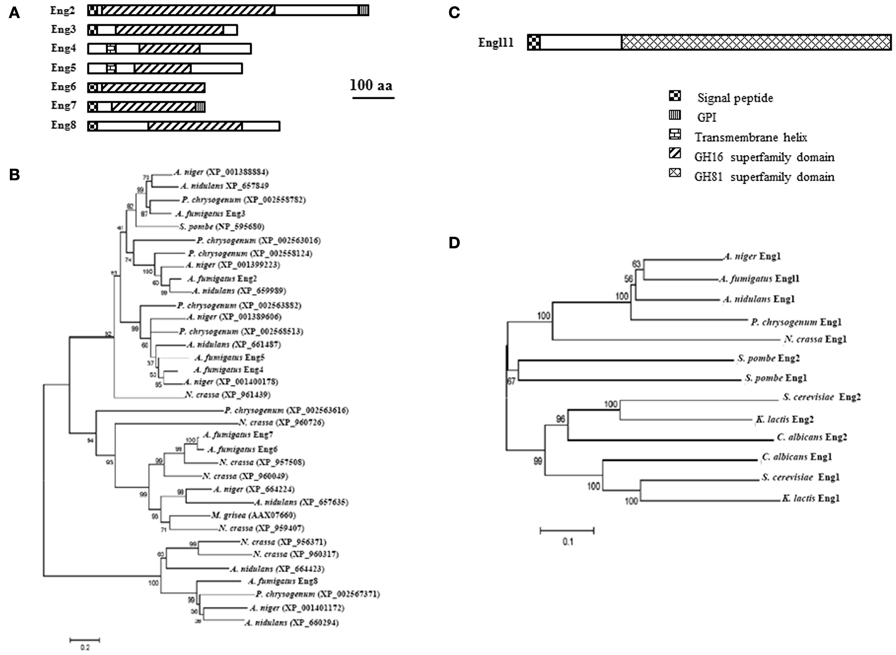
Figure 3. Domain organization and phylogenetic tree of A. fumigatus endo-β-1,3-glucanases. Protein domains identified using SignalP 3.0, big-PI, TMHMM v. 2.0, and CD search (100 aa: protein length proportional to 100 amino acids). (A) GH16 family domain organization and (B) phylogeny of GH16 family (C) GH81 family domain organization and (D) Phylogenetic tree of GH81 family.
It has been shown that AfENG2 encode for a endo β-1,3-glucanase protein (Hartl et al., 2011). These enzyme acts on β-1,3-glucans and lichenan but no degradation were observed with β(1–6)substrate. Eng2p preferentially acts on soluble polymers like laminarin and shorter β-1,3- linked oligosaccharides and have a slight transferase activity with substrates of low degree of polymerization (DP4 and DP6). The eng2 mutant is similar to the parental strain suggesting that AfEng2p alone does not play a morphogenetic role (Hartl et al., 2011). The two GH16_laminarinase_like proteins AfEng6p and AfEng7p exhibit the same protein length and almost identical molecular weights. Both feature a signal peptide followed by the catalytic domain. AfEng7p additionally features a C-terminal GPI signal sequence. While the bacterial members of this sub-family often contain a CBM such as CBM6 (Hong et al., 2002) or CBM13 (ricin-type β-trefoil) (Asano et al., 2002; Ferrer, 2006; Shi et al., 2010), no such CBMs were identified in AfEng6p and AfEng7p.
Although AfEng8p could not be assigned to the licheninase subfamily with high significance, part of the GH16 conserved domain of AfEng8p showed some strong homology to GH16_lichenases (Planas, 2000), which have been shown to be essential for the degradation of extracellular lichenan in Bacillus subtilis (Wolf et al., 1995). The fact that AfEng8p also features a signal peptide suggests a similar role in A. fumigatus.
In the GH16 family, three clades were identified (Figure 3B): the predicted GH16_MLG1_glucanases (top), GH16_laminarinase_like proteins (middle), and the probable lichenases (bottom). S. pombe is the only yeast that possesses a GH16 glucanase (NP_595680). In the filamentous fungi the number of GH16 glucanases ranges from one in M. grisea (AAX07660) to eight in A. fumigatus. When looking at fungi from the same genus (A. fumigatus, A. nidulans, and A. niger), the total number as well as the number of enzymes present in each clade varies. Because of their endoglucanase activity, multiple deletion of the eng gene is ongoing using the beta six rec cassette allowing repetitive round of gene deletion after excision of the resistance marker (Hartmann et al., 2010).
Only endo-β-1,3-glucanase (EC 3.2.1.39) belong to the GH81 family. A single member of this family has been found in the A. fumigatus genome. It is a glycosylated 74 kDa endo-β-1,3-glucanases protein that was isolated from A. fumigatus autolysate (supernatant of the broken mycelium grown 3 days at 37°C) (Fontaine et al., 1997b). This enzyme represents 10–15% of the total β-1,3-glucanase activity found in the A. fumigatus cell wall autolysate (supernatant of the cell wall extract incubated 3 days at 37°C). The enzyme recognized at least five glucose units linked by a β-1,3-bond. The gene encoding this activity has been cloned and named ENGL1 (AFUA_1G04260) (Mouyna et al., 2002). AfEngl1p has an N-terminal signal peptide and a large GH81 domain responsible for the endo-1,3-glucanase activity (Figure 3C). In contrast to A. fumigatus, two orthologs are present in the S. serevisiaie, S. pombe, and C. albicans genome. The phylogenetic tree of this GH81 family is presented in Figure 3D. Like for AfEngl1p, it has been shown that SpEng1p, SpEng2p (Martin-Cuadrado et al., 2008a) as well as ScEng1p (Baladron et al., 2002) exclusively hydrolyze linear β-1,3-glucan chains and display the same enzymatic activity as AfEngl1p. Moreover the four conserved aspartic and glutamic acid residues necessary for enzymatic activity has been determined by site-directed mutagenesis in ScEng2p corresponding to D518, D588, E609, and E613 (Martin-Cuadrado et al., 2008a).
In S. pombe, Martin-Cuadrado et al. (2003) showed that SpEng1p encodes protein with detectable endo β-1,3-glucanase activity whose deletion interfere in cell separation, because cells fails to degrade the primary septum that separates the two sisters cells. SpEng1p localizes to the septum region at the time of cell separation. SpEng2p also exhibits endo-β-1,3-glucanase activity but mutant Δeng2 do not have a cell separation defect. Δeng2 of S. pombe on the other hand is involved in ascus wall degradation after sporulation (Encinar del Dedo et al., 2009). Esteban et al. (2005) showed in C. albicans that CaEng1p is also involved in cell separation. Similar results has been obtained in S. cerevisiae (Baladron et al., 2002) where eng1 deletion mutants show defects in cell separation. In contrast, the lack of phenotype for the Δengl1 mutant suggests that this endo-β-1,3-glucanase activity does not play a morphogenetic role in A. fumigatus.
It has been shown that SpEng1p features a C-terminal CBM consisting of three repeats of about 50 amino acids each (Martin-Cuadrado et al., 2008b). The deletion of this CBM termed SpEng1CBM caused a reduction of catalytic activity against insoluble substrates and a defect in targeting of Eng1p to the septum. No such CBM module has been found in other proteins of the GH81 family in A. fumigatus, S. cerevisiae, or C. albicans.
Others Exo-β-Glucanases
In the CAZy database, family GH3 contains exo-hydrolases able to degrade different substrates. They are classified as β-glucosidase (EC 3.2.1.21); xylan 1,4-β-xylosidase (EC 3.2.1.37); β-N-acetylhexosaminidase (EC 3.2.1.52); glucan 1,3-β-glucosidase (EC 3.2.1.58); glucan 1,4-β-glucosidase (EC 3.2.1.74); exo-1,3-1,4-glucanase (EC 3.2.1.-); α-L-arabinofuranosidase (EC 3.2.1.55). Ten members of GH3 are present in the genome of A. fumigatus (Table 1): AFUA_1G05770, AFUA_7G06140, AFUA_7G00240, AFUA_1G17410, AFUA_8G02100, AFUA_6G14490, AFUA_5G07190, AFUA_6G11910, AFUA_6G08700 and AFUA_6G03570. The characteristics of the proteins encoded by these genes are summarized in Figure 4A and Table 1. Half of the proteins are secreted. All the proteins of A. fumigatus share conserved amino acids residues corresponding to the GH3 domain of the protein. Eight proteins have a Fibronectin like domain3 at the N-terminus. Fibronectin (FN) is a multidomain protein with the ability to bind simultaneously to cell surface receptors, collagen, proteoglycans, and other FN molecules (Schwarzbauer and De Simone, 2011). The phylogenetic tree is described in Figure 4B. Two main groups are observed. For most of the filamentous fungus, we observed a high redundancy of gene (from 5 to 11) in contrast to C. albicans (2) and S. pombe (only one). No orthologs have been found in the S. cerevisiae genome. To date, the only enzymatic activity characterized for this family in yeast or filamentous fungus correspond to ExoGII encoded by AFUA_1G05770 of A. fumigatus (Fontaine et al., 1997a). ExoGII has been isolated from the cell wall autolysate and is a 230 kDa protein able to degrade β-1,3-glucan as well as β-1,6-glucan and p-nitrophenyl-glucose (pNP-Glc). Such hydrolytic activity is in agreement with the classification of this enzyme in the GH3 family.
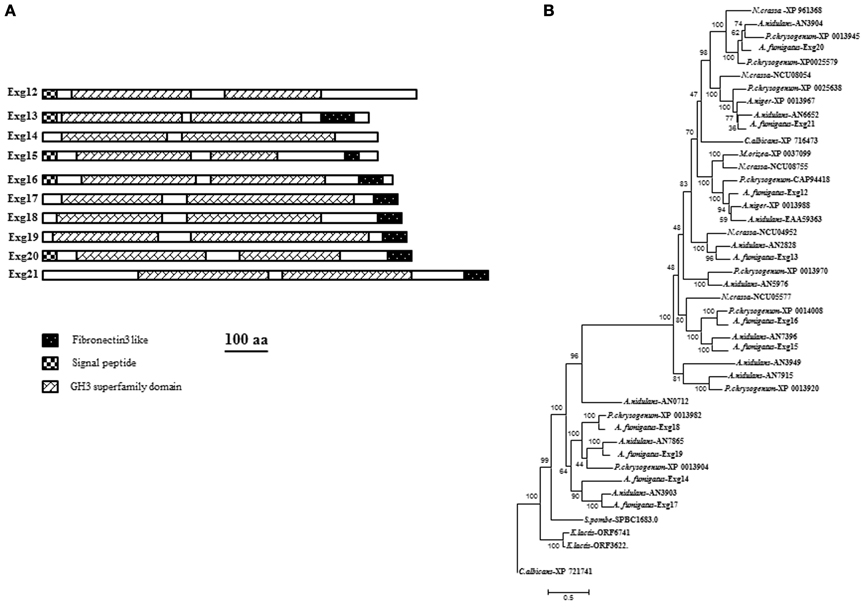
Figure 4. (A) Domain organization of GH3 families of A. fumigatus. Protein domains identified using SignalP 3.0, big-PI, TMHMM v. 2.0, and CD search (100 aa: protein length proportional to 100 amino acids). and (B) Phylogenetic tree of β(1–3)glucanases GH3 family.
What Morphogenetic Role Plays the β(1,3)-Glucanosyltransferase?
For many years, glucanosyltransferases were classified as glycosylhydrolases because their transglycosidase function was unknown. These glucanosyltransferases have indeed a glucanase activity because the first step in their sequential activity is to cleave linear β-1,3-glucans before the structural modification of the glucan (branching or elongation) in the second step. Because of their action on already established β-1,3-glucans, these enzymes could play a role in the structural modification of the cell wall and favor the emergence of a germ tube or an accessory hypha. Such roles have been suggested but never demonstrated.
Branching Enzyme
A glucanosyltransferase activity encoded by the BGT1 gene has been identified biochemically in the A. fumigatus cell wall autolysate, Bgt1p belongs to the Carbohydrate-Active Enzyme (CAZy) database Glycosyl hydrolase GH17 (http://www.cazy.org/) (Table 1). Bgt1p cleaves a laminaribiose from the reducing end of linear β-(1,3)-glucans and transfers the remaining glucan to the end of another β-1,3-glucan acceptor with a β-(1,6)-linkage (Mouyna et al., 1998). Such activity had been described originally in S. cerevisiae (Klebl and Tanner, 1989). We recently characterized Bgt2p in A. fumigatus. This protein presents homology with Bgt1p (Gastebois et al., 2010a). Using a recombinant protein produced in P. pastoris, we showed that the enzymatic activity of Bgt2p is a new branching activity, able to cleave two residues of a β-1,3-glucan chain and transfer them to the inside of another chain of β-1,3-glucan with β-1,6-linkage. However, the single and double bgt1/bgt2 mutants do not display a phenotype distinct from the parental strain (Mouyna et al., 1998; Gastebois et al., 2010a). Moreover, the amount of the β-1,3-glucan branching point has not changed in these mutants in comparison to the parental strain. Three other orthologs (AFUA_6G12380, AFUA_8G05610 and AFUA_5G08780) belonging to the GH17 family are currently investigated for their role as branching enzymes in the A. fumigatus cell wall.
Elongation Enzyme
Elongation of β-1,3-glucan is performed through the action of a β-1,3-glucanosyltransferase, which were originally discovered in A. fumigatus. This enzyme Gel1p splits a β-1,3-glucan molecule internally and transfers the newly generated reducing end to the non-reducing end of another β-1,3-glucan molecule, resulting in the elongation of the glucan chain (Hartland et al., 1996). Gel1p belongs to a family of seven members specifically recognized as the unique GH72 family in the Carbohydrate-Active Enzyme (CAZy) database (http://www.cazy.org/) (Mouyna et al., 2000a) (Table 1). This enzyme activity has been confirmed among paralogs of Gelp in A. fumigatus (Mouyna et al., 2000b; Gastebois et al., 2010b) and orthologs in other species like the GAS family in S. cerevisiae (Vai et al., 1991; Ragni et al., 2007), the GAS family in S. pombe (De Medina-Redondo et al., 2008, 2010) and PHR family in C. albicans (Saporito-Irwin et al., 1995; Muhlschlegel and Fonzi, 1997).
This GH72 family is subdivided in two families corresponding to the presence or absence of a Carbohydrate Binding Domain (CBM43) at the C-terminus. The Gel-family proteins are glycosylated and attached to the plasma membrane through a glycosylphosphatidylinositol (GPI) anchor. The recent crystallization of Gas2p, a Gelp homolog in S. cerevisiae, allows a better understanding of the enzyme's activity (Hurtado-Guerrero et al., 2009). Gas2p is composed of two interacting domains, a β(α)8 catalytic domain and a cystein-rich domain of the CBM43 family. The catalytic domain contains three disulfide bridges that are involved in the formation of the acceptor-saccharide binding site. The active site is defined by two catalytic residues, Glu176 and Glu275, and three tyrosine residues, Tyr107, Tyr244 and Tyr307, all conserved in the GH72 family. Moreover, although that seven homologs are present in the A. fumigatus genome, only GEL1, GEL2, and GEL4 are expressed at each stage of growth in the conditions tested (Gastebois et al., 2010b). Deletion of each gene has a different effect on the phenotype. The gel1 mutant behaves like the wild type, in contrast to the gel2 mutant which showed reduced growth with several phenotypical changes and cell wall modification (Mouyna et al., 2005). In addition, it was shown that GEL4 is an essential gene (Gastebois et al., 2010b). The morphogenetic role of these transglycosidases does not seem, however, associated to their glucanase function since these activity continue until the formation of an insoluble β-1,3-glucan.
Cross-Linking Enzyme?
The CRH family proteins which are classified in GH16 were originally studied in S. cerevisiae and they have been suggested biochemically to be involved in the linkage between β-1,6-glucan and chitin (Cabib et al., 2007, 2008). Although there is no β-1,6-glucan in A. fumigatus cell wall, 5 CRH orthologous genes have been identified in the A. fumigatus genome, AFUA_6G03230, AFUA_2G03120, AFUA_3G09250, AFUA_6G08510 AFUA_1G16190, and belonging to the GH16_GPI_glucanosyltransferase subfamily (Table 1). Their biochemical function remains unknown in this fungus. In addition, single deletion of these genes was not associated to any growth phenotype modification (Chabane S, Reichard U, unpublished). Their role in morphogenesis will be only understood when the quintuple crh mutant will be obtained.
Conclusions
Our understanding of the biochemical events responsible for conidial germination or hyphal branching remains incomplete. This review summarized the β-1,3-glucan modifying enzymes which have been described and the putative β-glucanases identified in A. fumigatus genome. Although the role of glycosylhydrolases during morphogenetic events has been suggested in filamentous fungus for many years, there is still no real proof of their importance during cell wall softening. The corresponding β-glucanases mutants obtained did not show any morphological differences compared to the wild type. In addition, if modifications of the glucanase activity have been seen in these mutants, the quantification of the activity is always performed in toto and never takes into account the localization in situ and their degrading capacity at a very specific cellular site. Such importance of the localization of the glycosylhydrolases has been already demonstrated in S. pombe (Martin-Cuadrado et al., 2008b). The redundancy of the putative exo β-or endo-glucanase in the genome (at least 29) make their analysis difficult because of putative compensatory mechanisms. The expression level of each of these putative β-glucanase during germination by RNA seq analysis is undergoing. Other hydrolases like chitinases that can have also similar function as glucanases will be jointly investigated. It should could give us some clue in the future to focus and study the genes upregulated during these morphogenesis events.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The research leading to these results has received funding from the European Union's Seventh Framework Programme [FP7/2007–2013] under grant agreement n° HEALTH-2010-260338 (ALLFUN), ERA-Net Pathogenomics Biomarkers for prevention, diagnosis and response to therapy of invasive aspergillosis (AspBIOmics), and ESF (European science Foundation) Fuminomics.
References
Asano, T., Taki, J., Yamamoto, M., and Aono, R. (2002). Cloning and structural analysis of bglM gene coding for the fungal cell wall-lytic beta-1,3-glucan-hydrolase BglM of Bacillus circulans IAM1165. Biosci. Biotechnol. Biochem. 66, 1246–1255.
Baladron, V., Ufano, S., Duenas, E., Martin-Cuadrado, A. B., del Rey, F., and Vazquez de Aldana, C. R. (2002). Eng1p, an endo-1,3-beta-glucanase localized at the daughter side of the septum, is involved in cell separation in Saccharomyces cerevisiae. Eukaryot. Cell 1, 774–786.
Beauvais, A., Bruneau, J. M., Mol, P. C., Buitrago, M. J., Legrand, R., and Latge, J. P. (2001). Glucan synthase complex of Aspergillus fumigatus. J. Bacteriol. 183, 2273–2279.
Beauvais, A., Drake, R., Ng, K., Diaquin, M., and Latge, J. P. (1993). Characterization of the 1,3 beta-glucan synthase of Aspergillus fumigatus. J. Gen. Microbiol. 139, 3071–3078.
Cabib, E., Blanco, N., Grau, C., Rodriguez-Pena, J. M., and Arroyo, J. (2007). Crh1p and Crh2p are required for the cross-linking of chitin to beta(1-6)glucan in the Saccharomyces cerevisiae cell wall. Mol. Microbiol. 63, 921–935.
Cabib, E., Farkas, V., Kosik, O., Blanco, N., Arroyo, J., and McPhie, P. (2008). Assembly of the yeast cell wall. Crh1p and Crh2p act as transglycosylases in vivo and in vitro. J. Biol. Chem. 283, 29859–29872.
Chambers, R. S., Broughton, M. J., Cannon, R. D., Carne, A., Emerson, G. W., and Sullivan, P. A. (1993). An exo-beta-(1,3)-glucanase of Candida albicans: purification of the enzyme and molecular cloning of the gene. J. Gen. Microbiol. 139, 325–334.
D'Enfert, C. (1997). Fungal spore germination: insights from the molecular genetics of Aspergillus fumigatus and Neurospora crassa. Fungal Genet. Biol. 21, 163–172.
De la Cruz, J., Pintor-Toro, J. A., Benitez, T., Llobell, A., and Romero, L. C. (1995). A novel endo-beta-1,3-glucanase, BGN13.1, involved in the mycoparasitism of Trichoderma harzianum. J. Bacteriol. 177, 6937–6945.
De Medina-Redondo, M., Arnáiz-Pita, Y., Clavaud, C., Fontaine, T., del Rey, F., Latgé, J. P., et al. (2010). β(1,3)-glucanosyl-transferase activity is essential for cell wall integrity and viability of Schizosaccharomyces pombe. PLoS ONE 18:e14046. doi: 10.1371/journal.pone.0014046
De Medina-Redondo, M., Arnaiz-Pita, Y., Fontaine, T., del Rey, F., Latge, J. P., and Vazquez de Aldana, C. R. (2008). The beta-1,3-glucanosyltransferase gas4p is essential for ascospore wall maturation and spore viability in Schizosaccharomyces pombe. Mol. Microbiol. 68, 1283–1299.
Duenas-Santero, E., Martin-Cuadrado, A. B., Fontaine, T., Latge, J. P., del Rey, F., and Vazquez de Aldana, C. (2010). Characterization of glycoside hydrolase family 5 proteins in Schizosaccharomyces pombe. Eukaryot. Cell 9, 1650–1660.
Encinar del Dedo, J., Duenas, E., Arnaiz, Y., del Rey, F., and Vazquez de Aldana, C. R. (2009). beta-glucanase Eng2 is required for ascus wall endolysis after sporulation in the fission yeast Schizosaccharomyces pombe. Eukaryotic Cell 8, 1278–1286.
Esteban, P. F., Rios, I., Garcia, R., Duenas, E., Pla, J., Sanchez, M., et al. (2005). Characterization of the CaENG1 gene encoding an endo-1,3-beta-glucanase involved in cell separation in Candida albicans. Curr. Microbiol. 51, 385–392.
Ferrer, P. (2006). Revisiting the Cellulosimicrobium cellulans yeast-lytic beta-1,3-glucanases toolbox: a review. Microb. Cell Fact. 5:10. doi: 10.1186/1475-2859-5-10
Fontaine, T., Hartland, R. P., Diaquin, M., Simenel, C., and Latgé, J. P. (1997a). Differential patterns of activity displayed by two exo-beta-1,3-glucanases associated with the Aspergillus fumigatus cell wall. J. Bacteriol. 179, 3154–3163.
Fontaine, T., Hartland, R. P., Beauvais, A., Diaquin, M., and Latge, J. P. (1997b). Purification and characterization of an endo-1,3-beta-glucanase from Aspergillus fumigatus. Eur. J. Biochem. 243, 315–321.
Fontaine, T., Simenel, C., Dubreucq, G., Adam, O., Delepierre, M., Lemoine, J., et al. (2000). Molecular organization of the alkali-insoluble fraction of Aspergillus fumigatus cell wall. J. Biol. Chem. 275, 27594–27607.
Gastebois, A., Mouyna, I., Simenel, C., Clavaud, C., Coddeville, B., Delepierre, M., et al. (2010a). Characterization of a new beta(1-3)-glucan branching activity of Aspergillus fumigatus. J. Biol. Chem. 285, 2386–2396.
Gastebois, A., Fontaine, T., Latge, J. P., and Mouyna, I. (2010b). The β(1-3)glucanosyltransferase Gel4p is essential for Aspergillus fumigatus. Eukaryot. Cell 9, 1294–1298.
Gibbons, J. G., Beauvais, A., Beau, R., McGary, K. L., Latgé, J. P., and Rokas, A. (2012). Global transcriptome changes underlying colony growth in the opportunistic human pathogen Aspergillus fumigatus. Eukaryot. Cell 11, 58–68.
Hartl, L., Gastebois, A., Aimanianda, V., and Latge, J. P. (2011). Characterization of the GPI-anchored endo beta-1,3-glucanase Eng2 of Aspergillus fumigatus. Fungal Genet. Biol. 48, 185–191.
Hartland, R. P., Fontaine, T., Debeaupuis, J. P., Simenel, C., Delepierre, M., and Latge, J. P. (1996). A novel beta-(1-3)-glucanosyltransferase from the cell wall of Aspergillus fumigatus. J. Biol. Chem. 271, 26843–26849.
Hartmann, T., Dumig, M., Jaber, B. M., Szewczyk, E., Olbermann, P., Morschhauser, J., et al. (2010). Validation of a self-excising marker in the human pathogen Aspergillus fumigatus by employing the β-rec/six site specific recombination system. Appl. Environ. Microbiol. 76, 6313–6317.
Hong, T. Y., Cheng, C. W., Huang, J. W., and Meng, M. (2002). Isolation and biochemical characterization of an endo-1,3-beta-glucanase from Streptomyces sioyaensis containing a C-terminal family 6 carbohydrate-binding module that binds to 1,3-beta-glucan. Microbiology 148, 1151–1169.
Hurtado-Guerrero, R., Schuttelkopf, A. W., Mouyna, I., Ibrahim, A. F., Shepherd, S., Fontaine, T., et al. (2009). Molecular mechanisms of yeast cell wall glucan remodeling. J. Biol. Chem. 284, 8461–8469.
Kim, D. J., Baek, J. M., Uribe, P., Kenerley, C. M., and Cook, D. R. (2002). Cloning and characterization of multiple glycosyl hydrolase genes from Trichoderma virens. Curr. Genet. 40, 374–384.
Klebl, F, and Tanner, W. (1989). Molecular cloning of a cell wall exo-beta-1,3-glucanase from Saccharomyces cerevisiae. J. Bacteriol. 171, 6259–6264.
Larriba, G., Andaluz, E., Cueva, R., and Basco, R. D. (1995). Molecular biology of yeast exoglucanases. FEMS Microbiol. Lett. 125, 121–126.
Martin-Cuadrado, A. B., Duenas, E., Sipiczki, M., Vazquez de Aldana, C. R., and del Rey, F. (2003). The endo-beta-1,3-glucanase eng1p is required for dissolution of the primary septum during cell separation in Schizosaccharomyces pombe. J. Cell. Sci. 116, 1689–1698.
Martin-Cuadrado, A. B., Fontaine, T., Esteban, P. F., Encinar del Dedo, J., de Medina-Redondo, M., del Rey, F., et al. (2008a). Characterization of the endo-beta-1,3-glucanase activity of S. cerevisiae Eng2 and other members of the GH81 family. Fungal Genet. Biol. 45, 542–553.
Martin-Cuadrado, A. B., Encinar del Dedo, J., de Medina-Redondo, M., Fontaine, T., del Rey, F., Latge, J. P., et al. (2008b). The Schizosaccharomyces pombe endo-1,3-beta-glucanase Eng1 contains a novel carbohydrate binding module required for septum localization. Mol. Microbiol. 69, 188–200.
Mouyna, I., Hartland, R. P., Fontaine, T., Diaquin, M., Simenel, C., Delepierre, M., et al. (1998). A 1,3-beta-glucanosyltransferase isolated from the cell wall of Aspergillus fumigatus is a homologue of the yeast Bgl2p. Microbiology 144, 3171–3180.
Mouyna, I., Monod, M., Fontaine, T., Henrissat, B., Lechenne, B., and Latge, J. P. (2000a). Identification of the catalytic residues of the first family of beta(1-3)glucanosyltransferases identified in fungi. Biochem. J. 347, 741–747.
Mouyna, I., Fontaine, T., Vai, M., Monod, M., Fonzi, W. A., Diaquin, M., et al. (2000b). Glycosylphosphatidylinositol-anchored glucanosyltransferases play an active role in the biosynthesis of the fungal cell wall. J. Biol. Chem. 275, 14882–14889.
Mouyna, I., Morelle, W., Vai, M., Monod, M., Lechenne, B., Fontaine, T., et al. (2005). Deletion of GEL2 encoding for a beta(1-3)glucanosyltransferase affects morphogenesis and virulence in Aspergillus fumigatus. Mol. Microbiol. 56, 1675–1688.
Mouyna, I., Sarfati, J., Recco, P., Fontaine, T., Henrissat, B., and Latgé, J. P. (2002). Molecular characterization of a cell wall-associated beta(1-3)endoglucanase of Aspergillus fumigatus. Med. Mycol. 40, 455–464.
Muhlschlegel, F. A., and Fonzi, W. A. (1997). PHR2 of Candida albicans encodes a functional homolog of the pH-regulated gene PHR1 with an inverted pattern of pH-dependent expression. Mol. Cell. Biol. 17, 5960–5967.
Nierman, W. C., Pain, A., Anderson, M. J., Wortman, J. R., Kim, H. S., Arroyo, J., et al. (2005). Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 438, 1151–1156.
Nobe, R., Sakakibara, Y., Ogawa, K., and Suiko, M. (2004). Cloning and expression of a novel Trichoderma viride laminarinase AI gene (lamAI). Biosci. Biotechnol. Biochem. 68, 2111–2119.
Planas, A. (2000). Bacterial 1,3-1,4-beta-glucanases: structure, function and protein engineering. Biochim. Biophys. Acta 1543, 361–382.
Ragni, E., Fontaine, T., Gissi, C., Latge, J. P., and Popolo, L. (2007). The Gas family of proteins of Saccharomyces cerevisiae: characterization and evolutionary analysis. Yeast 24, 297–308.
San Segundo, P., Correa, J., Vazquez de Aldana, C. R., and del Rey, F. (1993). SSG1, a gene encoding a sporulation-specific 1,3-beta-glucanase in Saccharomyces cerevisiae. J. Bacteriol. 175, 3823–3837.
Saporito-Irwin, S. M., Birse, C. E., Sypherd, P. S., and Fonzi, W. A. (1995). PHR1, a pH-regulated gene of Candida albicans, is required for morphogenesis. Mol. Cell. Biol. 15, 601–613.
Schaeffer, H. J., Leykam, J., and Walton, J. D. (1994). Cloning and targeted gene disruption of EXG1, encoding exo-beta 1,3-glucanase, in the phytopathogenic fungus Cochliobolus carbonum. Appl. Environ. Microbiol. 60, 594–598.
Schwarzbauer, J. E., and De Simone, D. W. (2011). Fibronectins, their fibrillogenesis, and in vivo functions. Cold Spring Harb. Perspect. Biol. 16, 1–19.
Shi, P., Yao, G., Yang, P., Li, N., Luo, H., Bai, Y., et al. (2010). Cloning, characterization, and antifungal activity of an endo-1,3-beta-D:-glucanase from Streptomyces sp. Appl. Microbiol. Biotechnol. 85, 1483–1490.
Suzuki, K., Yabe, T., Maruyama, Y., Abe, K., and Nakajima, T. (2001). Characterization of recombinant yeast exo-beta-1,3-glucanase (Exg1p) expressed in Escherichia coli cells. Biosci. Biotechnol. Biochem. 65, 1310–1314.
Vai, M., Gatti, E., Lacanà, E., Popolo, L., and Alberghina, L. (1991). Isolation and deduced amino acid sequence of the gene encoding gp115, a yeast glycophospholipid-anchored protein containing a serine-rich region. J. Biol. Chem. 266, 12242–12248.
Vazquez de Aldana, C. R., Correa, J., San Segundo, P., Bueno, A., Nebreda, A. R., Mendez, E., et al. (1991). Nucleotide sequence of the exo-1,3-beta-glucanase-encoding gene, EXG1, of the yeast Saccharomyces cerevisiae. Gene 97, 173–182.
Keywords: Aspergillus fumigatus, cell wall, β-1,3-glucan, β-1,3-glucan modifying enzyme, endo-β-1,3-glucanase, exo-β-1,3-glucanase
Citation: Mouyna I, Hartl L and Latgé J-P (2013) β-1,3-glucan modifying enzymes in Aspergillus fumigatus. Front. Microbiol. 4:81. doi: 10.3389/fmicb.2013.00081
Received: 27 November 2012; Accepted: 21 March 2013;
Published online: 17 April 2013.
Edited by:
Elaine M. Bignell, Imperial College London, UKReviewed by:
Carol Munro, University of Aberdeen, UKSven Krappmann, Friedrich-Alexander-Universität Erlangen-Nürnberg, Germany
Copyright © 2013 Mouyna, Hartl and Latgé. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Jean-Paul Latgé, Unité des Aspergillus, Département de Parasitologie et Mycologie, Institut Pasteur, 25 rue du Docteur Roux, 75015 Paris, France. e-mail:amVhbi1wYXVsLmxhdGdlQHBhc3RldXIuZnI=
†Present address: Lukas Hartl, Microsynth Austria GmbH, Leberstrasse 20, Vienna, Austria
‡These authors have contributed equally to this work.