


- 1National Reference Laboratory of Veterinary Drug Residues, College of Veterinary Medicine, South China Agricultural University, Guangzhou, China
- 2Key Laboratory of Fishery Drug Development, Ministry of Agriculture, P. R. China, Pearl River Fisheries Research Institute, Chinese Academy of Fishery Science, Guangzhou, China
16S rRNA methylase and QepA, a fluoroquinolone efflux pump, are new mechanisms of resistance against aminoglycosides and fluoroquinolone, respectively. One of 16S rRNA methylase genes, rmtB, was found to be associated with qepA, were both located on the same transposable element. In this study, we intended to determine the current prevalence and characteristics of the 16S rRNA methylase genes and qepA, and to study the association between rmtB and qepA. A total of 892 Escherichia coli isolates were collected from various diseased food-producing animals in China from 2004 to 2008 and screened by PCR for 16S rRNA methylase genes and qepA. About 12.6% (112/892) and 0.1% (1/892) of isolates that were highly resistant to amikacin were positive for rmtB and armA, respectively. The remaining five 16S rRNA methlyase genes were not detected. Thirty-six (4.0%) strains carried qepA. About 32.1% of rmtB-positive strains harbored qepA, which was not detected in rmtB-negative strains. Most strains were clonally unrelated, while identical PFGE profiles of rmtB-positive isolates were found in the same farm indicating clonal transmission. Conjugation experiments showed that rmtB was transferred to the recipients, and qepA also cotransferred with rmtB in some cases. The spread of E. coli of food animal origin harboring both rmtB and qepA suggests that surveillance for antimicrobial resistance of animal origin as well as the study of the mechanisms of resistance should be undertaken.
Introduction
In China, aminoglycosides and quinolone were commonly used for treating severe infections caused by Gram-negative bacteria in animal husbandry. As a result, multiple resistance determinants to these antimicrobial agents have emerged in various pathogenic microbes. Recently, a new type of mechanism, post-transcriptional methylation of the 16S rRNA, has been reported, and this results in high-level resistance to aminoglycoside antibiotics. At present, ten 16S rRNA methylase genes have been identified, including armA, rmtA, rmtB, rmtC, rmtD, rmtE, rmtF, rmtG, rmtH, and npmA, which are capable of conferring extraordinarily high levels of resistance to most clinically important aminoglycosides, including amikacin, gentamicin, kanamycin, and tobramycin (Galimand et al., 2003; Yokoyama et al., 2003; Doi and Arakawa, 2007; Wachino et al., 2007; Davis et al., 2010; Galimand et al., 2012; Bueno et al., 2013; O'Hara et al., 2013). RmtB and ArmA are the most frequently identified methylases in Enterobacteriaceae isolated in East Asia, Europe, and South America (Yan et al., 2004; Bogaerts et al., 2007; Berçot et al., 2008; Kang et al., 2009; Yu et al., 2010). The plasmid mediated efflux pump gene, qepA, which confers resistance to hydrophilic fluoroquinolones by efflux, has also been reported recently to be frequently associated with the rmtB gene (Périchon et al., 2007; Yamane et al., 2007; Deng et al., 2011b; Yao et al., 2011).
The emergence of 16S rRNA methylases in bacteria of animal origin was first discovered in E. coli isolates of pig origin harboring the armA gene in Spain in 2005 (González-Zorn et al., 2005). Since then, 16S rRNA methylase-producing E. coli isolates have been increasingly detected in pigs, chicken, cows, and companion animals (dogs and cats) in different countries (Chen et al., 2007; Liu et al., 2008; Du et al., 2009; Davis et al., 2010; Hopkins et al., 2010; Deng et al., 2011a). Stains from food-producing animals colonized with 16S rRNA methylase conferring high level of resistance to aminoglycosides have been considered a potential source of resistant E. coli causing infection in the community (Chen et al., 2007). A combination of factors has contributed to the rapid dissemination of 16S rRNA methylase genes, such as co-selection, which is mainly due to conjugative plasmids and other mobile genetic elements rather than clonal expansion. Some studies have reported that the dissemination of the rmtB gene involves IS26, Tn3, Tn1721, ISCR1, and ISCR3 (Yamane et al., 2007; Berçot et al., 2008; Périchon et al., 2008; Du et al., 2009). In this study, we intended to investigate the distribution of 16S rRNA methylase genes and qepA among E. coli isolates originating from various food-producing animals in China from 2004 to 2008. In addition, in order to understand how the rmtB and qepA genes disseminated, molecular typing and conjugation experiments were conducted to determine the mechanisms of resistance and mobility of the rmtB and qepA genes in these isolates.
Materials and Methods
Bacterial Isolates
E. coli isolates were recovered from feces, livers, lungs, or milk samples of diseased food-producing animals with diarrhea, respiratory diseases or other diseases, including 360 pigs, 261 chickens, 179 ducks, 34 geese, 9 pigeons, 12 partridges, and 37 cows in six provinces of China from July 2004 to October 2008. Each isolate was from a separate animal, and a total of 892 isolates were collected from 150 farms. All samples were seeded on MacConkey agar plates and incubated at 37°C for 24 h. The isolates were identified with conventional biochemical tests and confirmed using an API 20E system (bioMérieux, Marcy l'Étoile, France). E. coli ATCC25922 was used as the MIC reference strain and E. coli J53 AzideR as the recipient for conjugation experiments.
Detection of 16S rRNA Methylases Genes and qepA
A multiplex-PCR method was used to detect the genes enconcoding 16S rRNA methylases (armA, rmtA, rmtB, rmtC, rmtD) with primer sets described previously (Doi and Arakawa, 2007). The primers used to amplify npmA were 5′-AGG GCT ATC TAA TGT GGT G-3′ and 5′-TAT TTC CGC TTC TTC GTA T-3′. The qepA gene was amplified with primers 5′-GCA GGT CCA GCA GCG GGT AG-3′ and 5′-CTT CCT GCC CGA GTA TCG TG-3′. The PCR products were confirmed by sequencing.
Antimicrobial Susceptibility Testing
The minimal inhibitory concentrations (MICs) of ampicillin, ceftiofur, nalidixic acid, ciprofloxacin, norfloxacin, gentamicin, amikacin, streptomycin, neomycin, apramycin, chloramphenicol, florfenicol, and sulfamethoxazole/trimethoprim were determined by the agar dilution method according to Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2008, 2010). E. coli ATCC25922 was used as a quality control strain.
Molecular Typing
Pulsed-field gel electrophoresis (PFGE) analysis of Xbal-digested genomic DNA was performed to determine the genetic relatedness of 16S rRNA methylase-producing E. coli isolates using a CHEF-II System (Bio-Rad Laboratories, Hercules, CA, USA) with PFGE separation conditions of 0.5–63.8 s for 20.3 h. PFGE patterns were interpreted according to well established criteria (Tenover et al., 1995). Isolates that had PFGE patterns with no more than six different bands were considered to be clonally related.
All 16S rRNA methylase-positive isolates were classified into phylogenetic groups (A, B1, B2, or D) according to the presence of chuA, yjaA, and TSPE4, as determined by an established multiplex PCR-based method described previously (Clermont et al., 2000).
Conjugation Experiment
In order to test the transferability of the amikacin resistance determinant to the azide-resistant strain J53, the rmtB-positive stains were employed as putative donors in a conjugation assay using the broth mating method. The transconjugants were selected on LB agar plates supplemented with amikacin (200 μg/mL) and sodium azide (200 μg/mL). From the transconjugants, PCRs for rmtB and qepA were performed using primers reported previously. The MICs for donors, transconjugants, and recipients were measured by the agar dilution method in accordance with CLSI guidelines. The antimicrobials tested were amikacin, gentamicin, nalidixic acid, ciprofloxacin, norfloxacin, ampicillin and other antimicrobials.
Results
Detection of 16S rRNA Methylases Genes and qepA
All isolates, which were derived from various diseased food-producing animals, were screened for 16S rRNA methylase genes by PCR. One armA and 112 rmtB genes were found to be present in these isolates, representing 0.1 and 12.6% of the total 892 E. coli isolates, respectively. None of other five 16S rRNA methylase genes were detected in any of the isolates. As shown in Table 1, the distribution of 16S rRNA methylase genes in different animal species was varied. Details of armA and rmtB-positive isolates were listed in Table A1.
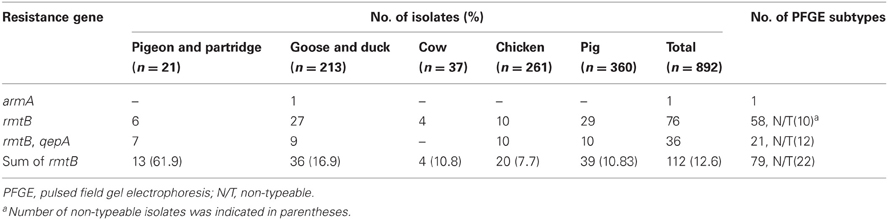
Table 1. Distribution of the 16S rRNA methylase genes and qepA among all isolates from different sources.
The qepA gene was detected in 36 of 892 isolates (4.0%). About 32.1% of rmtB-positive strains harbored qepA, while qepA was not detected in rmtB-negative strains.
Susceptibility Testing Results
The susceptibility testing to 14 antimicrobial agents was conducted for the 113 isolates found to be 16S rRNA methylase-positive. All the strains producing 16S rRNA methylases displayed high-level resistance (MIC > 128 μg/ml) to amikacin and gentamicin, as well as to ampicillin, tetracycline, streptomycin, and nalidixic acid. In addition, there was also a very high frequency of resistance to sulfamethoxazole/trimethoprim, florfenicol, and chloramphenicol among these isolates: 97.2, 93.7, and 92.4%, respectively. Of the 113 isolates, all were multi-resistant and demonstrated resistance to ciprofloxacin (88.6%), norfloxacin (82.3%), neomycin (75.9%), apramycin (59.5%), and ceftiofur (42.3%) (Table 2).
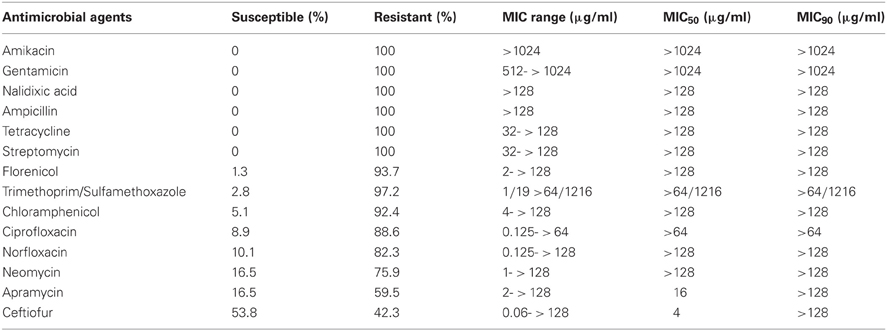
Table 2. Susceptibility of 113 E. coli isolates carrying 16S rRNA methylases to 14 antimicrobial agents.
Molecular Typing
In order to characterize the clonality of all 16S rRNA methylase-positive strains, molecular typing was performed by PFGE analysis and phylogenetic group assignment was carried out. PFGE was performed on 113 16S rRNA methylase-producers, and 91 isolates were found to be typeable while 22 were non-typeable. Seventy-nine major profiles were obtained among the 90 rmtB-positive strains, and 60 of them were represented by a single isolate (Table 3).
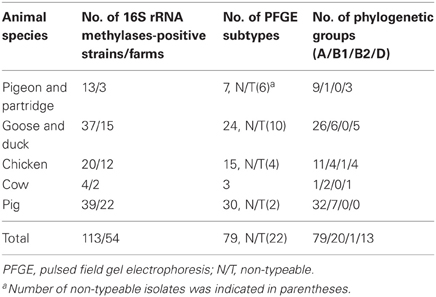
Table 3. Distribution of 16S rRNA methylase- positive strains, their clonal relationship, and phylogenetic groups from different animal species.
The majority of armA- and rmtB-producing isolates were found to belong to phylogenetic group A (N = 79, 69.9%). Twenty (17.7%), and 13 (11.5%) of the strains were found to belong to groups B1 and D, respectively. Only one rmtB-positive strain from chicken belonged to group B2.
Conjugation Experiment
Plasmid transfer of high-level aminoglycoside resistance to E. coli J53 was successful for 43 of the 65 rmtB-carrying strains. The qepA gene also co-transferred with the rmtB gene from 17 rmtB-qepA-positive donor strains, and its presence was confirmed in all of 17 transconjugants by PCR. The MICs of amikacin and gentamicin for all rmtB-carrying transconjugants were 256- to 512-fold higher than those for the recipients, indicating that the rmtB gene contributes to high-level resistance to aminoglycosides. Furthermore, the plasmids in some transconjugants also confered resistance to ampicillin (100%), tetracycline (6.8%), chloramphenicol (25.0%), nalidixic acid (9.12%), trimethoprim/sulfamethoxazole (40.9%) and ceftiofur (29.5%). Compared to the qepA-negative transconjugants, which displayed no changes in susceptibility to fluoroquinolone, the MICs of ciprofloxacin and norfloxacin for qepA-harboring transconjugants were 8- to 64-fold higher than those found for the recipients. These results suggest that qepA contributes to the decrease in hydrophilic fluoroquinolone susceptibility.
Discussion
Detection of 16S rRNA Methylases Genes and qepA
In this study, 892 E. coli isolates originating from various diseased food-producing animals, including 360 pigs, 261 chickens, 179 ducks, 34 geese, 9 pigeons, 12 partridges, and 37 cows, were collected from six different provinces in China from 2004 to 2008. We found that 12.6% (112/892) of them carried rmtB gene, while the armA gene was detected in only one isolate. This result demonstrates the widespread dissemination of the rmtB gene among multiple animal sources in China, as reported in other studies (Chen et al., 2007; Liu et al., 2008; Du et al., 2009; Deng et al., 2011a; Li et al., 2012). The prevalence rate of rmtB among E. coli isolated from different animals was much higher than those reported to be of patient origin, which was less than 5% (Fritsche et al., 2008; Kang et al., 2008). Nonetheless, the incidence of armA-carrying Enterobacteriaceae isolates of patient origin is higher in comparison with the incidences determined from data obtained regarding isolates derived from animal sources (Lee et al., 2006; Kang et al., 2008).
QepA is a plasmid-mediated efflux pump first discovered in an E. coli strain isolated from the urine specimen of an inpatient in Japan in 2002 (Yamane et al., 2007). The plasmid bearing the qepA gene increased the MICs of nalidixic acid, ciprofloxacin and norfloxacin by 2-, 32-, and 64-fold, respectively. Since its initial discovery, a variant of qepA possessing two amino acid substitutions was identified and named QepA2 (Cattoir et al., 2008); this variant conferred a resistance phenotype similar to that of QepA, which has now been renamed QepA1. Interestingly, qepA1-positive isolates from Japan, Belgium, China, and South Korea were found to be associated with the rmtB gene on the same Tn3 transposon (Périchon et al., 2007; Yamane et al., 2007; Périchon et al., 2008; Kim et al., 2009; Deng et al., 2011b), whereas qepA2 was flanked by a novel element, ISCR3C, with no rmtB associated with it (Cattoir et al., 2008). In this study, the qepA gene was found in 36 of 892 isolates (4.0%), and 32.1% of rmtB-positive strains harbored qepA. qepA was not detected in rmtB-negative strains, implying that qepA gene was associated with rmtB gene on a same mobile genetic element as reported recently (Liu et al., 2008; Périchon et al., 2008; Yamane et al., 2008; Park et al., 2009; Yao et al., 2011), except a recent report by Baudry et al. (2009). The high prevalence of rmtB and qepA coexisting in E. coli samples of diseased food-producing animals is worrisome, because they may be rapidly spreading among animals, humans and even in the environment; this may be occurring by direct or indirect contact and co-selection, with various antimicrobials possibly contributing to its dissemination and further limiting therapeutic options.
Molecular Typing of the 16S rRNA Methylases-Positive Strains
Among the 112 rmtB-positive strains, 79 distinct PFGE patterns were found, and 21 isolates concomitantly harbored qepA gene. Only small numbers of isolates with rmtB were clonally related. PFGE analysis indicated that a diversity of PFGE patterns was present in strains of different origins. However, identical patterns were found in the strains derived from the same farm. These data suggested that the high prevalence of rmtB-positive isolates was not mainly caused by clonal dissemination. Phylogenetic background of the 113 16S rRNA methylases-positive strains were also conducted in this study. Studies have shown that phylogenetic groups B2 and D usually carry virulence factors (Clermont et al., 2000). Compared with the other phylogenetic groups, a greater number of isolates in phylogenetic group B2 from human patients has been reported (Baudry et al., 2009; Song et al., 2009). However, we only obtained one rmtB-positive strain from chicken belonged to B2. The strains collected from diseased animals belong mainly to group A, including armA-positive, rmtB-positive and qepA-positive strains, indicating that most strains were not involved in pathogenicity.
Conjugation Experiment of 16S rRNA Methylases-Positive Strains
To investigate the transferability of rmtB to the recipient Ecoli J53, conjugation experiment was conducted. Forty three rmtB-positive conjugative plasmids were obtained, 17 of which carried both rmtB and qepA. E. coli isolates harbored transferable aminoglycoside resistance determinants and the increasing prevalence of transferable quinolone resistance determinants may have been an important driving force for selection and dissemination of aminoglycoside- and quinolone-resistant isolates. Our previous work showed that rmtB and qepA were found located on a very similar F2:A-:B- plasmids, which have disseminated among pigs, human and environment (Deng et al., 2011b). Another reports showed that 24 out of 35 transconjugants bearing rmtB and qepA originating from companion animals were also associated with the F2:A-:B- plasmid, 21 of which were sharing the identical plasmid restriction patterns (Deng et al., 2011a). It suggested that co-existence of rmtB and qepA on the same plasmid may contribute to dissemination of both aminoglycoside and quinolone resistance in different animal sources.
In conclusion, the present screen revealed a high prevalence of 16S rRNA methylase genes among E. coli isolated from various diseased food-producing animals in six provinces of China from 2004 to 2008. The dissemination of rmtB and qepA genes in the E. coli of food-producing animals was mainly mediated by a conjugated plasmid. The coexistence of these resistance determinants on a single plasmid increases the selection by one or more of the antimicrobials used in clinical practice. Prudent use of antimicrobial agents in veterinary clinics, especially those treating food-producing animals, should be reinforced.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by grants from National Natural Science Foundation of China (No. 30972218 and No. U1031004) and Guangdong Natural Science Foundation (No. S2012030006590).
References
Baudry, P. J., Nichol, K., Decorby, M., Lagacé-Wiens, P., Olivier, E., Boyd, D., et al. (2009). Mechanisms of resistance and mobility among multidrug-resistant CTX-M-producing Escherichia coli from Canadian intensive care units: the 1st report of QepA in North America. Diagn. Microbiol. Infect. Dis. 63, 319–326. doi: 10.1016/j.diagmicrobio.2008.12.001
Berçot, B., Poirel, L., and Nordmann, P. (2008). Plasmid-mediated 16S rRNA methylases among extended-spectrum beta-lactamase-producing Enterobacteriaceae isolates. Antimicrob. Agents Chemother. 52, 4526–4527. doi: 10.1128/AAC.00882-08
Bogaerts, P., Galimand, M., Bauraing, C., Deplano, A., Vanhoof, R., De Mendonca, R., et al. (2007). Emergence of ArmA and RmtB aminoglycoside resistance 16S rRNA methylases in Belgium. J. Antimicrob. Chemother. 59, 459–464. doi: 10.1093/jac/dkl527
Bueno, M. F., Francisco, G. R., O'Hara, J. A., de Oliveira Garcia, D., and Doi, Y. (2013). Co-production of 16S ribosomal RNA methyltransferase RmtD and RmtG with KPC-2 and CTX-M-group ESBLs in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 53, 2397–2400. doi: 10.1128/AAC.02108-12
Cattoir, V., Poirel, L., and Nordmann, P. (2008). Plasmid-mediated quinolone resistance pump QepA2 in an Escherichia coli isolate from France. Antimicrob. Agents Chemother. 52, 3801–3804. doi: 10.1128/AAC.00638-08
Chen, L., Chen, Z. L., Liu, J. H., Zeng, Z. L., Ma, J. Y., and Jiang, H. X. (2007). Emergence of RmtB methylase-producing Escherichia coli and Enterobacter cloacae isolates from pigs in China. J. Antimicrob. Chemother. 59, 880–885. doi: 10.1093/jac/dkm065
Clermont, O., Bonacorsi, S., and Bingen, E. (2000). Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 66, 4555–4558. doi: 10.1128/AEM.66.10.4555-4558.2000
Clinical Laboratory Standard Institute (CLSI). (2008). “Performance standards for antimicrobial disk suseptibility tests for bacteria isolated from animals; approved standard,” in Third Edition Document M31-A3 (Wayne, PA: CLSI).
Clinical Laboratory Standard Institute. (CLSI). (2010). “Performance standards for antimicrobial suseptibility testing,” in Twentieth Informational Supplement M100-S20 (Wayne, PA: CLSI).
Davis, M. A., Baker, K. N., Orfe, L. H., Shah, D. H., Besser, T. E., and Call, D. R. (2010). Discovery of a gene conferring multiple-aminoglycoside resistance in Escherichia coli. Antimicrob. Agents Chemother. 54, 2666–2669. doi: 10.1128/AAC.01743-09
Deng, Y., He, L., Chen, S., Zheng, H., Zeng, Z., Liu, Y., et al. (2011a). F33:A-:B- and F2:A-:B- plasmids mediate dissemination of rmtB-blaCTX–M–9 group genes and rmtB-qepA in Enterobacteriaceae isolates from pets in China. Antimicrob. Agents Chemother. 55, 4926–4929. doi: 10.1128/AAC.00133-11
Deng, Y., Zeng, Z., Chen, S., He, L., Liu, Y., Wu, C., et al. (2011b). Dissemination of IncFII plasmids carrying rmtB and qepA in Escherichia coli from pigs, farm workers and the environment. Clin. Microbiol. Infect. 17, 1740–1745. doi: 10.1111/j.1469-0691.2011.03472.x
Doi, Y., and Arakawa, Y. (2007). 16S ribosomal RNA methylation: emerging resistance mechanism against aminoglycosides. Clin. Infect. Dis. 45, 88–94. doi: 10.1086/518605
Du, X. D., Wu, C. M., Liu, H. B., Li, X. S., Beier, R. C., Xiao, F., et al. (2009). Plasmid-mediated ArmA and RmtB 16S rRNA methylases in Escherichia coli isolated from chickens. J. Antimicrob. Chemother. 64, 1328–1330. doi: 10.1093/jac/dkp354
Fritsche, T. R., Castanheira, M., Miller, G. H., Jones, R. N., and Armstrong, E. S. (2008). Detection of methyltransferases conferring high-level resistance to aminoglycosides in Enterobacteriaceae from Europe, North America, and Latin America. Antimicrob. Agents Chemother. 52, 1843–1845. doi: 10.1128/AAC.01477-07
Galimand, M., Courvalin, P., and Lambert, T. (2003). Plasmid-mediated high-level resistance to aminoglycosides in Enterobacteriaceae due to 16S rRNA methylation. Antimicrob. Agents Chemother. 47, 2565–2571. doi: 10.1128/AAC.47.8.2565-2571.2003
Galimand, M., Courvalin, P., and Lambert, T. (2012). RmtF, a new member of the aminoglycoside resistance 16S rRNA N7 G1405 methyltransferase family. Antimicrob. Agents Chemother. 56, 3960–3962. doi: 10.1128/AAC.00660-12
González-Zorn, B., Teshager, T., Casas, M., Porrero, M. C., Moreno, M. A., Courvalin, P., et al. (2005). armA and aminoglycoside resistance in Escherichia coli. Emerg. Infect. Dis. 11, 954–956. doi: 10.3201/eid1106.040553
Hopkins, K. L., Escudero, J. A., Hidalgo, L., and Gonzalez-Zorn, B. (2010). 16S rRNA methyltransferase RmtC in Salmonella enterica serovar Virchow. Emerg. Infect. Dis. 16, 712–715. doi: 10.3201/eid1604.090736
Kang, H. Y., Kim, J., Seol, S. Y., Lee, Y. C., Lee, J. C., and Cho, D. T. (2009). Characterization of conjugative plasmids carrying antibiotic resistance genes encoding 16S rRNA methylase, extended-spectrum beta-lactamase, and/or plasmid-mediated AmpC beta-lactamase. J. Microbiol. 47, 68–75. doi: 10.1007/s12275-008-0158-3
Kang, H. Y., Kim, K. Y., Kim, J., Lee, J. C., Lee, Y. C., Cho, D. T., et al. (2008). Distribution of conjugative-plasmid-mediated 16S rRNA methylase genes among amikacin-resistant Enterobacteriaceae isolates collected in 1995 to 1998 and 2001 to 2006 at a university hospital in South Korea and identification of conjugative plasmids mediating dissemination of 16S rRNA methylase. J. Clin. Microbiol. 46, 700–706. doi: 10.1128/JCM.01677-07
Kim, E. S., Jeong, J. Y., Choi, S. H., Lee, S. O., Kim, S. H., Kim, M. N., et al. (2009). Plasmid-mediated fluoroquinolone efflux pump gene, qepA, in Escherichia coli clinical isolates in Korea. Diagn. Microbiol. Infect. Dis. 65, 335–338. doi: 10.1016/j.diagmicrobio.2009.07.006
Kim, J., Bae, I. K., Jeong, S. H., Chang, C. L., Lee, C. H., and Lee, K. (2011). Characterization of IncF plasmids carrying the blaCTX–M–14 gene in clinical isolates of Escherichia coli from Korea. J. Antimicrob. Chemother. 66, 1263–1268. doi: 10.1093/jac/dkr106
Lee, H., Yong, D., Yum, J. H., Roh, K. H., Lee, K., Yamane, K., et al. (2006). Dissemination of 16S rRNA methylase-mediated highly amikacin-resistant isolates of Klebsiella pneumoniae and Acinetobacter baumannii in Korea. Diagn. Microbiol. Infect. Dis. 56, 305–312. doi: 10.1016/j.diagmicrobio.2006.05.002
Li, D. X., Zhang, S. M., Hu, G. Z., Wang, Y., Liu, H. B., Wu, C. M., et al. (2012). Tn3-associated rmtB together with qnrS1, aac(6')-Ib-cr and blaCTX–M–15 are co-located on an F49:A-:B- plasmid in an Escherichia coli ST10 strain in China. J. Antimicrob. Chemother. 67, 236–238. doi: 10.1093/jac/dkr428
Liu, J. H., Deng, Y. T., Zeng, Z. L., Gao, J. H., Chen, L., Arakawa, Y., et al. (2008). Coprevalence of plasmid-mediated quinolone resistance determinants QepA, Qnr, and AAC(6')-Ib-cr among 16S rRNA methylase RmtB-producing Escherichia coli isolates from pigs. Antimicrob. Agents Chemother. 52, 2992–2993. doi: 10.1128/AAC.01686-07
O'Hara, J. A., McGann, P., Snesrud, E. C., Clifford, R. J., Waterman, P. E., Lesho, E. P., et al. (2013). Novel 16S ribosomal RNA methyltransferase RmtH produced by Klebsiella pneumoniae associated with war-related trauma. Antimicrob. Agents Chemother. 57, 2413–2416. doi: 10.1128/AAC.00266-13
Park, Y. J., Yu, J. K., Kim, S. I., Lee, K., and Arakawa, Y. (2009). Accumulation of plasmid-mediated fluoroquinolone resistance genes, qepA and qnrS1, in Enterobacter aerogenes co-producing RmtB and class A beta-lactamase LAP-1. Ann. Clin. Lab. Sci. 39, 55–59.
Périchon, B., Bogaerts, P., Lambert, T., Frangeul, L., Courvalin, P., and Galimand, M. (2008). Sequence of conjugative plasmid pIP1206 mediating resistance to aminoglycosides by 16S rRNA methylation and to hydrophilic fluoroquinolones by efflux. Antimicrob. Agents Chemother. 52, 2581–2592. doi: 10.1128/AAC.01540-07
Périchon, B., Courvalin, P., and Galimand, M. (2007). Transferable resistance to aminoglycosides by methylation of G1405 in 16S rRNA and to hydrophilic fluoroquinolones by QepA-mediated efflux in Escherichia coli. Antimicrob. Agents Chemother. 51, 2464–2469. doi: 10.1128/AAC.00143-07
Song, S., Lee, E. Y., Koh, E. M., Ha, H. S., Jeong, H. J., Bae, I. K., et al. (2009). Antibiotic resistance mechanisms of Escherichia coli isolates from urinary specimens. Korean J. Lab. Med. 29, 17–24. doi: 10.3343/kjlm.2009.29.1.17
Tenover, F. C., Arbeit, R. D., Goering, R. V., Mickelsen, P. A., Murray, B. E., Persing, D. H., et al. (1995). Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol. 33, 2233–2239.
Wachino, J., Shibayama, K., Kurokawa, H., Kimura, K., Yamane, K., Suzuki, S., et al. (2007). Novel plasmid-mediated 16S rRNA m1A1408 methyltransferase, NpmA, found in a clinically isolated Escherichia coli strain resistant to structurally diverse aminoglycosides. Antimicrob. Agents Chemother. 51, 4401–4409. doi: 10.1128/AAC.00926-07
Yamane, K., Wachino, J., Suzuki, S., and Arakawa, Y. (2008). Plasmid-mediated qepA gene among Escherichia coli clinical isolates from Japan. Antimicrob. Agents Chemother. 52, 1564–1566. doi: 10.1128/AAC.01137-07
Yamane, K., Wachino, J., Suzuki, S., Kimura, K., Shibata, N., Kato, H., et al. (2007). New plasmid-mediated fluoroquinolone efflux pump, QepA, found in an Escherichia coli clinical isolate. Antimicrob. Agents Chemother. 51, 3354–3360. doi: 10.1128/AAC.00339-07
Yan, J. J., Wu, J. J., Ko, W. C., Tsai, S. H., Chuang, C. L., Wu, H. M., et al. (2004). Plasmid-mediated 16S rRNA methylases conferring high-level aminoglycoside resistance in Escherichia coli and Klebsiella pneumoniae isolates from two Taiwanese hospitals. J. Antimicrob. Chemother. 54, 1007–1012. doi: 10.1093/jac/dkh455
Yao, Q., Zeng, Z., Hou, J., Deng, Y., He, L., Tian, W., et al. (2011). Dissemination of the rmtB gene carried on IncF and IncN plasmids among Enterobacteriaceae in a pig farm and its environment. J. Antimicrob. Chemother. 66, 2475–2479. doi: 10.1093/jac/dkr328
Yokoyama, K., Doi, Y., Yamane, K., Kurokawa, H., Shibata, N., Shibayama, K., et al. (2003). Acquisition of 16S rRNA methylase gene in Pseudomonas aeruginosa. Lancet 362, 1888–1893. doi: 10.1016/S0140-6736(03)14959-8
Appendix
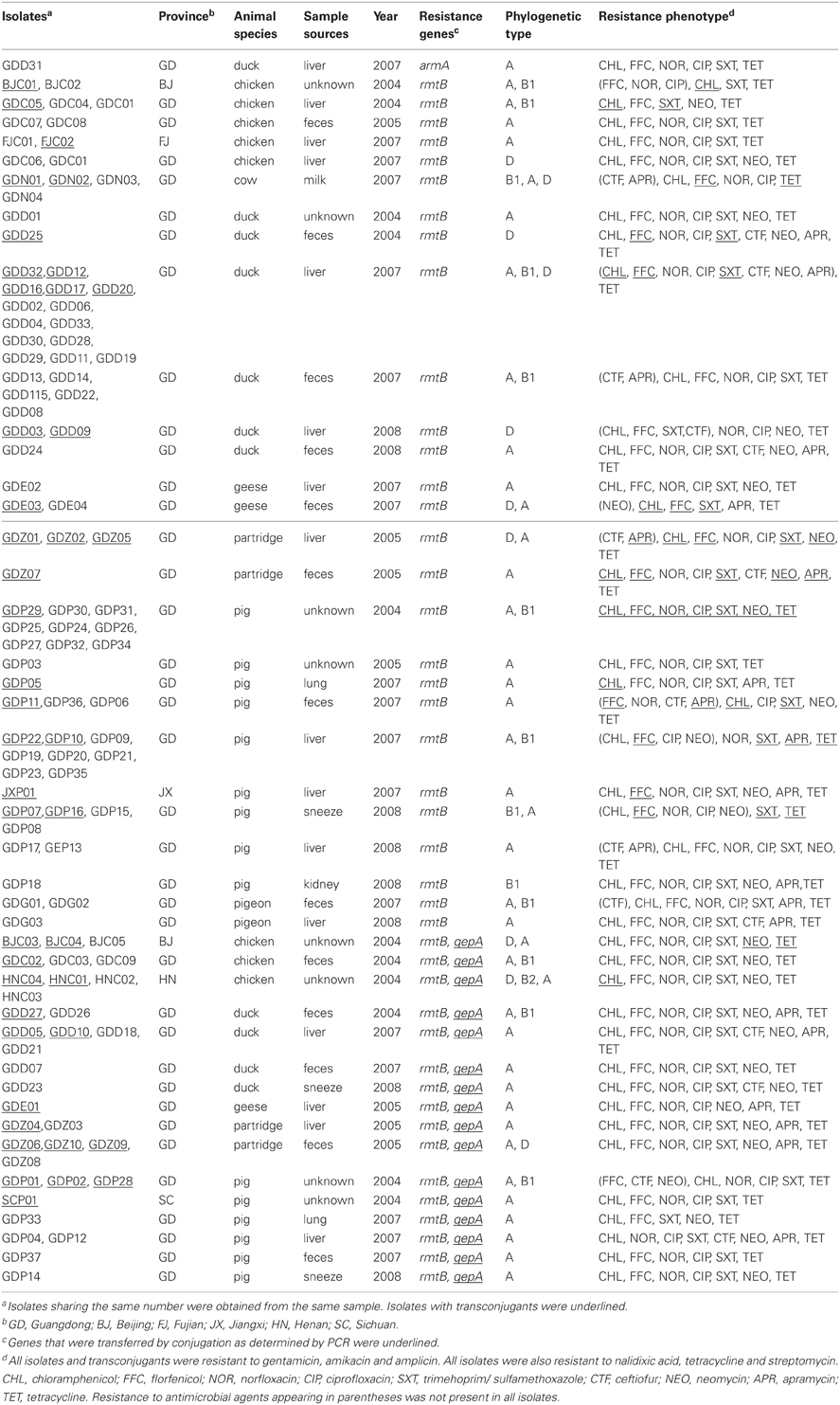
Table A1. Details of armA and rmtB-positive isolates.
Keywords: 16S rRNA methylases, qepA, E. coli, animal, molecular typing
Citation: Deng Y-T, Zeng Z-L, Tian W, Yang T and Liu J-H (2013) Prevalence and characteristics of rmtB and qepA in Escherichia coli isolated from diseased animals in China. Front. Microbiol. 4:198. doi: 10.3389/fmicb.2013.00198
Received: 17 March 2013; Paper pending published: 26 April 2013;
Accepted: 25 June 2013; Published online: 15 July 2013.
Edited by:
Michel S. Zygmunt, Institut National de la Recherche Agronomique, FranceReviewed by:
Patrick R. Butaye, Ghent University, BelgiumAtte V. Wright, University of Eastern Finland, Finland
Copyright © 2013 Deng, Zeng, Tian, Yang and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Jian-Hua Liu, National Reference Laboratory of Veterinary Drug Residues, College of Veterinary Medicine, South China Agricultural University, Wushan Road 483, Guangzhou 510642, China e-mail:amhsaXVAc2NhdS5lZHUuY24=