 Danièle Sohier
Danièle Sohier Sonia Pavan
Sonia Pavan Armelle Riou
Armelle Riou Jérôme Combrisson
Jérôme Combrisson Florence Postollec
Florence Postollec- 1Food Safety and Quality Unit, ADRIA Développement, Agri-Food Technical Institute, Quimper, France
- 2Bretagne Biotechnologie Alimentaire dairy association member, Analytical Sciences, Danone Research, Palaiseau, France
Traditionally, culture-based methods have been used to enumerate microbial populations in dairy products. Recent developments in molecular methods now enable faster and more sensitive analyses than classical microbiology procedures. These molecular tools allow a detailed characterization of cell physiological states and bacterial fitness and thus, offer new perspectives to integration of microbial physiology monitoring to improve industrial processes. This review summarizes the methods described to enumerate and characterize physiological states of technological microbiota in dairy products, and discusses the current deficiencies in relation to the industry’s needs. Recent studies show that Polymerase chain reaction-based methods can successfully be applied to quantify fermenting microbes and probiotics in dairy products. Flow cytometry and omics technologies also show interesting analytical potentialities. However, they still suffer from a lack of validation and standardization for quality control analyses, as reflected by the absence of performance studies and official international standards.
Introduction
Fermenting microorganisms play a pivotal role in the development of physicochemical and sensory properties of food products. They also contribute to product safety by limiting the growth of pathogenic and spoilage microorganisms (Caplice and Fitzgerald, 1999). Therefore, evaluation of cell viability is of great importance for the fermented food industry in general, and more specifically for the dairy sector. As fermenting microbes are responsible for organoleptic properties, it is essential to be able to characterize not only cells able to divide but also metabolic activities and bacterial fitness, in order to improve quality controls and end products. For probiotic products, it is important to ensure the presence of sufficient numbers of viable cells that will bring about beneficial health effects (Karimi et al., 2012).
The number of food products that involve microbial activities during at least one step of their production is substantial. As a consequence, numerous methods have been described to characterize microbial populations participating in fermentation processes. Traditionally, descriptive culture methods have been used, and remain the most employed to determine the presence/absence of colonies (i.e., cultivable cells) and their numbers. However, these straightforward methods provide a very simplistic, often biased, view of the physiological state of microbial populations in which several subpopulations characterized by various levels of “viability” and metabolic activity may coexist (Davey, 2011). The emergence of molecular techniques has opened new opportunities to characterize the numerous intermediate states of microbial cells, so much so that the well-being, fitness, and metabolic activities are now being targeted through the quantification of biomarkers, rather than just growth/no growth quantifications (Sieuwerts et al., 2008; de Vos, 2011). A biomarker is defined by the National Institutes of Health as “a characteristic that is objectively measured and evaluated as an indicator of normal biological processes, pathogenic processes, or pharmacologic responses to a therapeutic intervention” (Atkinson et al., 2001). We propose an adaptation of this definition to food processes as “a characteristic that is objectively measured and evaluated as an indicator of normal biological processes, pathogenic processes, or cellular responses to food processes.” Molecular methods encompass fluorescent in situ hybridization (FISH), flow cytometry (FC), “omics,” and Polymerase chain reaction (PCR)-based technologies. Some of these, such as FC, have been described for a few decades and were successfully used for very diverse research purposes (Díaz et al., 2010), but their routine application to dairy industry analyses is just being seriously considered (Figure 1). More recently developed, omics technologies are very promising to better understand microbial communities and to identify biomarkers, but until now they have not been applied to quality control purposes. PCR-based techniques are now being routinely used for the analysis of pathogens and the characterization of technological microbiota in fermented products (Postollec et al., 2011).
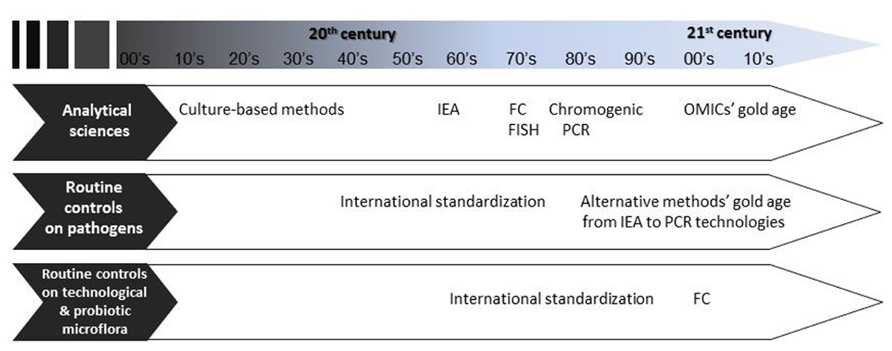
FIGURE 1. Evolution of analytical methods, and the gaps between routine controls depending on the targeted microbiota. IEA, immuno-enzymatic assay; FC, flow cytometry; FISH, fluorescent in situ hybridization; PCR, polymerase chain reaction.
What about their standard application to quantify fitness biomarkers? What is the gap between all the described methods and the specific needs of the dairy industry for fast, efficient, reliable, and standardized methods? This article will review the existing methods described to enumerate technological microbiota and characterize their physiological status that contribute to quality control of processes and end-products.
In addition to global Internet search, a total of four international databases were screened for journal articles, books, patents, conferences, and symposia proceedings in the field of food science, food industry, life science, and biomedical information (FSTA®, BIOSIS® Preview, Medline®, Foodline®). As this article aims at providing basis for discussion about the real use of these methods by dairy industry laboratories, current lacks, and future possibilities to facilitate the transfer from research knowledge to food industry applications, only references published between 2000 and 2013 were considered. Only representative publications for each technique and type of application were retained, and priority was given to reviews discussing about specific techniques and their applications. Methods for strain identification or characterization were excluded.
Culture Methods, Outdated or Essential?
The growth of colonies on nutrient agar is routinely used in all microbiology laboratories and is the simplest way to detect and quantify viable microbes. Protocols based on culture methods and colony counting are validated as reference or alternative methods according to the European and International Standard Organization (EN ISO) standards, to detect and enumerate food-borne pathogens, total microbiota and hygiene indicator microbes. A few ISO methods are also available to quantify probiotics and fermenting microbes employed in the dairy industry (Boyer and Combrisson, 2013).
The main limitations of these enumeration methods are the lack of discrimination between the targeted microbes and the endogenous microbiota, the time-to-result, false positive counts, matrix-dependent efficiency, and last but not least, the impossibility to recover viable but non-cultivable (VBNC) cells, which are seen as dead. The concept of viability of microorganisms was for long considered as the ability to multiply on an optimal medium (Postgate et al., 1961). Then, in the 80s, the concept of VBNC was developed (Roszak et al., 1984; Roszak and Colwell, 1987), with the application of additional methods such as Live/Dead detection staining kits or reverse transcription (RT)-PCR, to differentiate cells able to divide, VBNC and dead ones. It is now universally recognized that many intermediate states exist between live and dead bacteria (Kell et al., 1998; Nebe-von-Caron et al., 2000; Díaz et al., 2010) and that colonies recovered by application of a culture method correspond to the cells (or groups of cells) that are able to replicate under the provided growth conditions.
In spite of the evolution of the viability concept, of the increased knowledge relating to the physiological states of microbial cells and of the development of molecular methods that do not display some of the above mentioned limitations, there is still a great interest in culture methods as reflected by the regularly published studies. Various agar plate media were described for selective enumeration of probiotic and non-probiotic lactic acid bacteria (LAB) and Bifidobacterium species in yogurt, cheese, or in pure cultures (Tharmaraj and Shah, 2003; Van de Casteele et al., 2006; De Carvalho Lima et al., 2009; Ashraf and Shah, 2011; Saccaro et al., 2011; Karimi et al., 2012). Probiotic survival in commercial products including powders, frozen material, micro-encapsulated cultures, capsules, foods, and drinks was evaluated using various agar plate media (Corcoran et al., 2004; Phillips et al., 2006; Antunes et al., 2007; Magariños et al., 2007; Champagne et al., 2010, 2011; do Espírito Santo et al., 2011; Forssten et al., 2011; Oberg et al., 2011). Probiotic shelf-life during which the minimal content in viable cells has to be guaranteed was determined. The parameters influencing probiotic survival in yogurt and cheese were also studied (Donkor et al., 2007; Karimi et al., 2011). To improve the efficiency of LAB selective media, many variations of the basic agar formulas and culture conditions were proposed, including the use of antibiotics, different incubation temperatures, NaCl concentrations, pH or carbon sources (Van de Casteele et al., 2006; Antunes et al., 2007; Saccaro et al., 2011). However, the limitations already present with older media remain. For microbiologically complex products, the autochthonous microbiota is often undistinguishable from the technological microbiota (Phillips et al., 2006; Ashraf and Shah, 2011). In addition, most culture methods allow discriminating bacteria at the genus level only, and at best at the species level (Tharmaraj and Shah, 2003; Ashraf and Shah, 2011; Saccaro et al., 2011). Matrix effect is another important aspect to consider when applying a culture method. Indeed, protocols developed to enumerate microbes in a given food product may not be reliable with another food product, as exemplified in many studies where variable bacterial counts were found depending on the matrix used (Van de Casteele et al., 2006; do Espírito Santo et al., 2011; Forssten et al., 2011). This underlines the need to develop specific methods, but also the lack of quality controls. Indeed, a majority of the developed methods focus on feasibility aspects but do not demonstrate to what extent the method is accurate, reliable, biased, and do not establish the detection limits. Similarly to the matrix effect, a strain effect may also occur (Van de Casteele et al., 2006), and again, the lack of quality controls to evaluate whether a given method can be accurately extrapolated to other strains is blatant. Considering the currently available cultural tools, it is generally recommended to use an additional identification method (preferably molecular), to ensure the accuracy and reliability of the results. This is especially important with probiotic bacteria enumeration, because minimal viable numbers have to be present in the product to guarantee health effects. Therefore, the need for clear recommendations is obvious. The set-up and validation of methods according to official standards would highly benefit the dairy industry. Recommendations for each analytical step of the viability assessment of probiotics in food products, beverages, dry or frozen preparations have been recently proposed (Champagne et al., 2011). Although specific for probiotics, most of these recommendations can be applied to the analysis of fermented products.
Strikingly, the use of chromogenic media has been developed for the analysis of pathogenic and spoilage bacteria and validated and normalized methods are available, but nothing of the kind was set up for technological microbiota. Indeed, colonies of Listeria, Salmonella, Bacillus cereus, Escherichia coli, and other enterobacteria can now be easily distinguished by a large range of colors. Chromogenic media are user-friendly culture methods, and can be used without knowledge in molecular biology. Such media would undoubtedly be very useful for microbiological controls in the dairy industry, and their development would contribute to fill in the lack of specific methods.
Microscopy, Flow Cytometry, and Other Fluorescent Labeling-Based Methods: Applicability for the Food Industry?
Fluorescent in situ hybridization was a popular technique in research laboratories in the 90s. It is based on the identification of cells containing specific nucleic acid sequences. Oligomer probes conjugated to fluorescent molecules hybridize to their target DNA or RNA, and fluorescence is detected by microscopic observation (Bottari et al., 2006). In the last years, a few publications have described the use of FISH to enumerate dairy microbes. Propionibacteria were detected and enumerated in cheese with good correlations with bacterial counts (Babot et al., 2011). García-Hernández et al. (2012) have combined a direct viable count method with FISH to enumerate viable yogurt strains inoculated in feces. FISH allows highly specific bacterial detection and, unlike other molecular techniques, provides information about spacial distribution of bacteria in their environment due to the use of imaging. In spite of these advantages, FISH remains difficult to set up, has low repeatability and artifacts and interferences with the food matrix often occur. Therefore its application for routine analyses is difficult to implement. The use of other fluorescent labeling techniques, sometimes combined with FISH, is also described (Moreno et al., 2006; Zotta et al., 2012). Fluorescent labeling is always associated with microscopy technologies. While the performances of methods based on fluorescent labeling and microscopic detection are usually very good (Auty et al., 2001; Bernardeau et al., 2001), a routine use of epifluorescence and/or confocal microscopy for food industry quality controls is hardly conceivable.
Another technology that has greatly evolved together with the development of fluorescent dyes is FC. It allows characterizing individual cells within a cell population, and to discriminate between various VBNC and physiological states (Sheehan et al., 2005; Lahtinen et al., 2006; Papadimitriou et al., 2006, 2007; Quiros et al., 2007; Sunny-Roberts and Knorr, 2008; El Arbi et al., 2011; Zotta et al., 2012), which is a major advantage over culture-based methods. The excellent review from Díaz et al. (2010) describes the principle of FC, a range of fluorescent probes associated with various cellular functions and applications to industrial microbial bioprocesses including dairy industry applications. Nowadays, FC appears as the most promising labeling technology (Tracy et al., 2010; Davey, 2011) and many publications describe its use in food microbiology. A large choice of dyes is available to target cell components (nucleic acids, proteins, lipids), intracellular pH, membrane integrity, intracellular ions, viability markers, and membrane energization (Maecker et al., 2004; Sheehan et al., 2005; Papadimitriou et al., 2006; Berney et al., 2007; Quiros et al., 2007; Comas-Riu and Rius, 2009; Díaz et al., 2010; Doherty et al., 2010; El Arbi et al., 2011; Zotta et al., 2012). A fine characterization of the cellular status is possible by combining several dyes (Chen et al., 2011). The effects of acid, oxidative, osmotic, or cold stresses encountered during food production or storage on the cellular status were studied (Ananta et al., 2005; Lahtinen et al., 2006; Moreno et al., 2006; Papadimitriou et al., 2007; Sunny-Roberts and Knorr, 2008; Kramer et al., 2009; El Arbi et al., 2011; Zotta et al., 2012). Population dynamics in batch cultures can also be characterized (Quiros et al., 2007; Comas-Riu and Rius, 2009; Lopes Da Silva et al., 2009), as well as early detection of bacteriophage infection (Michelsen et al., 2007), membrane changes at various cheese cooking temperatures (Sheehan et al., 2005), and antibacterial effects of bacteriocins (Budde and Rasch, 2001). FC has also been proposed as a means to enumerate viable probiotic populations in commercial products (Maukonen et al., 2006). Correlations between FC enumerations and plate counts are often very good during exponential growth and when cells are not submitted to stress (Bunthof and Abee, 2002; El Arbi et al., 2011). In other situations FC counts generally outnumber the plate counts by 0.3–1 log of CFU/g (Gunasekera et al., 2000; Quiros et al., 2007; Sunny-Roberts and Knorr, 2008). Among the other advantages of FC over traditional culture methods, one should mention the shorter analysis time and possible automation (Maukonen et al., 2006; Díaz et al., 2010; Davey, 2011). However, while a whole population cellular activity can be characterized, distinction between near genera or species remains difficult because specific nucleic acid and antigen probes have not yet been extensively used. In addition, the quantification – and sometimes detection – limit is relatively high, for instance 103–104 cells/ml in milk (Gunasekera et al., 2000; Maecker et al., 2004). Protocol development and data analyses also require some experience (Davey, 2011). Indeed, depending on their composition some samples need additional preparation, such as a reduction of protein amounts (Gunasekera et al., 2000; Doherty et al., 2010). False-positives or data dispersion may be observed with some dyes (Maecker et al., 2004).
In spite of the few limitations, FC appears as a very promising tool for the food industry (Díaz et al., 2010; Tracy et al., 2010; Davey, 2011). Beyond the simple enumeration of cells, FC can provide higher knowledge about microbial fitness and metabolic activities during bioprocesses. Therefore, this will improve optimization of technological processes involving dairy bacteria, prediction of microbial performances along the whole process and the presence/absence of activity during storage. Such application would, for instance, benefit the quality control of probiotic products during their shelf life, and inclusion of FC as a validated technology for this purpose is supported by some authors (Lahtinen et al., 2006). Today, existing kits and automated systems are rather dedicated to the detection of industrial and environmental contaminants (Díaz et al., 2010). But FC is currently used for milk quality control, and an ISO standardized method is being set-up under coordination of the International Dairy Federation, for enumeration of LAB in starter cultures and their applications (International Dairy Federation, 2012). An automated system is being commercialized by AES-Chemunex to enumerate viable bacteria in industrial products (http://www.aeschemunex.com). An important step is still to overcome for FC routine analysis, and developed applications require thorough calibration and validation of their performances and limitations.
Omic Methods: Research Tools Only?
The number of sequenced lactobacilli and lactococci genomes has greatly increased during the last decade (Liu et al., 2005; Makarova et al., 2006; de Vos, 2011). High throughput functional genomics and comparative metagenomic studies have also known a strong expansion and many recent articles describe research applications of omics technologies in the field of food fermentations and associated microbes. Transcriptomics and proteomics studies have enabled analyzing the response of LAB to various conditions of culture, stresses and industrial processes (de Vos, 2011). For instance, several articles reported the use of omics to characterize gene expression and protein synthesis during growth in milk or cheese and bacterial adaptation during commercial preparation (Gitton et al., 2005; Raynaud et al., 2005; Herve-Jimenez et al., 2008; Azcarate-Peril et al., 2009; Cretenet et al., 2011; Taibi et al., 2011; Thevenard et al., 2011). Transcriptomic, proteomic, and metabolic responses of LAB and bifidobacteria to temperature, acidic or oxidative conditions, bile, osmotic stress, limited carbon sources, or basic pH were described (De Dea Lindner et al., 2007; Cretenet et al., 2011; Lee et al., 2011; Li et al., 2011). The behavior of LAB in mixed cultures in milk, yogurt, cheese or probiotic products was also evaluated (Herve-Jimenez et al., 2008; Sieuwerts et al., 2008, 2010; Ruiz et al., 2009; Thevenard et al., 2011). Moreover, Omics technologies were applied to study genome evolution and biodiversity in various ecological niches (Siezen et al., 2004, 2008; Makarova et al., 2006; Rasmussen et al., 2008; O’Sullivan et al., 2009; Pastink et al., 2009; Taibi et al., 2011). One of the goals of these omics studies is to identify key biomarkers that could be used for the screening of new probiotic or technologically interesting strains and for the evaluation of their physiological states in order to improve functionality during industrial processes (Reid et al., 2002; Ventura et al., 2007; Rasmussen et al., 2008; Izquierdo et al., 2009; Sieuwerts et al., 2010; Thierry et al., 2011). The wealth of data generated by omics approaches also necessitates powerful tools to analyze and interpret them. It is interesting to mention the method developed by Segata et al. (2011) available as an online interface, to quickly select for candidate biomarkers by screening metagenomic data.
Usefulness of omics technologies to understand microbial behaviors and cellular pathways with the aim to optimize industrial processes is obvious. By contrast, applications of these high throughput tools to improve analytical methods have been neglected. However, identification of biomarkers that could help developing new simple and fast analytical methods for quality controls deserves attention.
The Popular PCR and PCR-Based Methods: Ready for Routine Analyses?
Since its development in the 80s, PCR has become fundamental to biological and medical research laboratories (Bartlett and Stirling, 2003). Widely employed for descriptive purposes such as the detection of microbes and analyses of ecosystems composition in combination with other technologies, it is now routinely used for the detection of pathogenic and spoilage microbes in food products (Postollec et al., 2011). Detection of food-borne pathogens by PCR is recognized by the ISO and standardized through several guidelines (ISO, 2005a,b, 2006a,b, 2011a,b). PCR is also used to confirm characteristic colonies from agar plates, as specified by ISO (ISO, 2007). In the last decade, this amplification technique has strongly evolved toward quantitative PCR (qPCR; Masco et al., 2007; Malorny et al., 2008; Le Dréan et al., 2010), and ISO guidelines describing the use of qPCR for the detection of food-borne pathogens in foodstuff have also been developed (ISO, 2012, 2013). The last 10 years have witnessed a large number of research articles and reviews describing PCR-based applications for probiotics and microbes involved in fermentation processes. Arisen from comparative genomics analyses, these methods aim at specifically quantifying microbial populations (Friedrich and Lenke, 2006; Masco et al., 2007; Randazzo et al., 2009; Sheu et al., 2009, 2010; Abdulamir et al., 2010; Reimann et al., 2010; Sohier et al., 2012), at assessing the role of microbes in dairy products processes (Collado et al., 2006; Ben Amor et al., 2007; Randazzo et al., 2009; Masoud et al., 2011), or at allowing simultaneous identification of dairy and probiotic bacteria containing multiple strains (Wong and Medrano, 2005; Sul et al., 2007; Senan et al., 2008; Smith and Osborn, 2009).
Polymerase chain reaction techniques were also used to evaluate physiological states and viability of microorganisms during products processes (Lahtinen et al., 2008; Marco and Kleerebezem, 2008; García-Cayuela et al., 2009; Randazzo et al., 2009; Matijašić et al., 2010; Meng et al., 2010; Bove et al., 2011). One way to distinguish between viable and dead bacteria is to use RT-qPCR which targets RNA instead of DNA (Matsuda et al., 2009; Falentin et al., 2010; Reimann et al., 2010). However, due to the short and variable half-life of RNA molecules and difficulties to extract high quality RNA from complex matrices, the reliable use of RT-qPCR remains delicate and necessitates thorough quality controls and standardizations (Postollec et al., 2011). Other alternatives have been proposed, among which is the viability PCR approach (Fittipaldi et al., 2012). DNA intercalating agents such as propidium monoazide (PMA) and ethidium monoazide (EMA) are used. They penetrate only into dead cells with compromised membranes and subsequently prevent amplification of DNA by PCR. Quantification of viable cells in probiotic products or viable LAB in fermented milk was proposed using EMA-PCR and PMA-PCR, and showed good correlations with plate counts (García-Cayuela et al., 2009; Matijašić et al., 2010; Meng et al., 2010). RT-qPCR is a method of choice to study physiological states. However, in spite of the increasing availability of omics data, and due to the above mentioned difficulties to work with RNA molecules, validated methods to quantify bacterial fitness biomarkers still lag behind.
Compared to standard plate counts, the reduced time-to-results and higher specificity of PCR-based methods make them very appropriate for dairy industry needs (Boyer and Combrisson, 2013). However, a big step leading to routine use for dairy quality controls is still present because many studies do not take into account these specific needs. Indeed, robustness, accuracy and limits of the developed methods are rarely characterized, and validations on final products are not always performed.
A drawback of the high sensitivity of PCR and RT-qPCR is that these methods are also very sensitive to small variations in sample preparation, amplification and mode of data expression, which may have a major impact on the results. The lack of consensus on how best to perform experiments and interpret PCR data is regularly pointed out (Bustin, 2009; Boyer and Combrisson, 2013). MIQE guidelines were published in order to ensure reproducibility and comparability of data (Bustin et al., 2009). Recently, a dMIQE checklist proposing the minimum information for publication of digital PCR (dPCR; see below) requirements was set up (Huggett et al., 2013). Based on the current lack and knowledge we previously proposed a list of recommendations for better use of qPCR in food industry analyses (Postollec et al., 2011). An ISO standard defines the minimal requirements for the detection of food-borne pathogens in foodstuffs by qPCR (ISO, 2011b) but no similar standard is available for fermenting or probiotic microbes. All these recommendations should help designing qPCR-based methods as alternative methods for quantification of dairy microorganisms that would be further validated according to EN ISO 16140 (ISO, 2003a).
Digital PCR or droplet digital PCR (ddPCR) that uses the well-known principle of the most probable number (MPN) employed in microbiology appears as an interesting alternative to qPCR (Baker, 2012). It is based on amplification of single target DNA molecules, and thus provides absolute quantification without the need to set up a standard curve, which would be more accurate than qPCR and should facilitate comparability of data (White et al., 2009). As for standard qPCR, dPCR can also be multiplexed, thus reducing the bias coming from the competition between targets (Ottesen et al., 2006; Hua et al., 2010; Zhong et al., 2011). Applications of dPCR for the detection or quantification of microbes in food have not yet been described. Clinical microbiology studies have reported good performances of the method and good correlations with qPCR (Hayden et al., 2013). Other interesting clinical and environmental applications [for instance, Devonshire et al., 2013; Heredia et al., 2013; Kelley et al., 2013; Song et al., 2013] suggest a promising future for dPCR that could easily be extended to food microbiology.
Existing Patents and Standards
Very few patents targeting nucleic acid sequences of LAB were deposited. They relate to nucleic acid sequences of L. acidophilus encoding cell-surface protein homologues and stress-related proteins and their uses (Klaenhammer et al., 2004, 2005), L. rhamnosus polynucleotides, probes, primers, genetic constructs, polypeptides, and methods for using them (Dekker et al., 2002, 2004; Glenn et al., 2004), Lactobacillus nucleic acids and proteins involved in stress response and recombinant expression vectors and host cells (Klaenhammer et al., 2006, 2010). A patent describing a method to detect live microbiological contaminants in food, by detecting a mRNA coding for an elongation factor synthesized by the contaminants, is also available (Gendre and Brignon, 1998). The nucleic acid molecules and metabolic products described in these patents could be used as biomarkers to characterize bacterial activity and fitness.
Only a few ISO standards relating to fermenting LAB or bifidobacteria in dairy products are available. Some of these standards specify plate-count methods for the enumeration of targeted microbiota in milk or yogurt (ISO, 1998, 2003c, 2006c, 2010b). Some others do not refer to bacterial counts but specify the tests for the identification of the characteristic microorganisms in yogurt or the composition of starters (ISO, 2003b, 2010a).
Conclusion
Table 1 summarizes the current uses and lacks of the main analytical methods in dairy microbiology. Indeed, many databases are now available and provide a deeper understanding of the physiology and metabolic characteristics of strains. Still underexploited, they could help developing routine methods to evaluate LAB and probiotics physiological states. For instance, the current biological and physiological knowledge about fermenting bacteria should help developing chromogenic media based on biochemical reactions specific for intrinsic strain properties. These chromogenic tools are very straightforward, more specific than classical agar media, and very relevant for use in microbiological controls performed by the dairy industry.
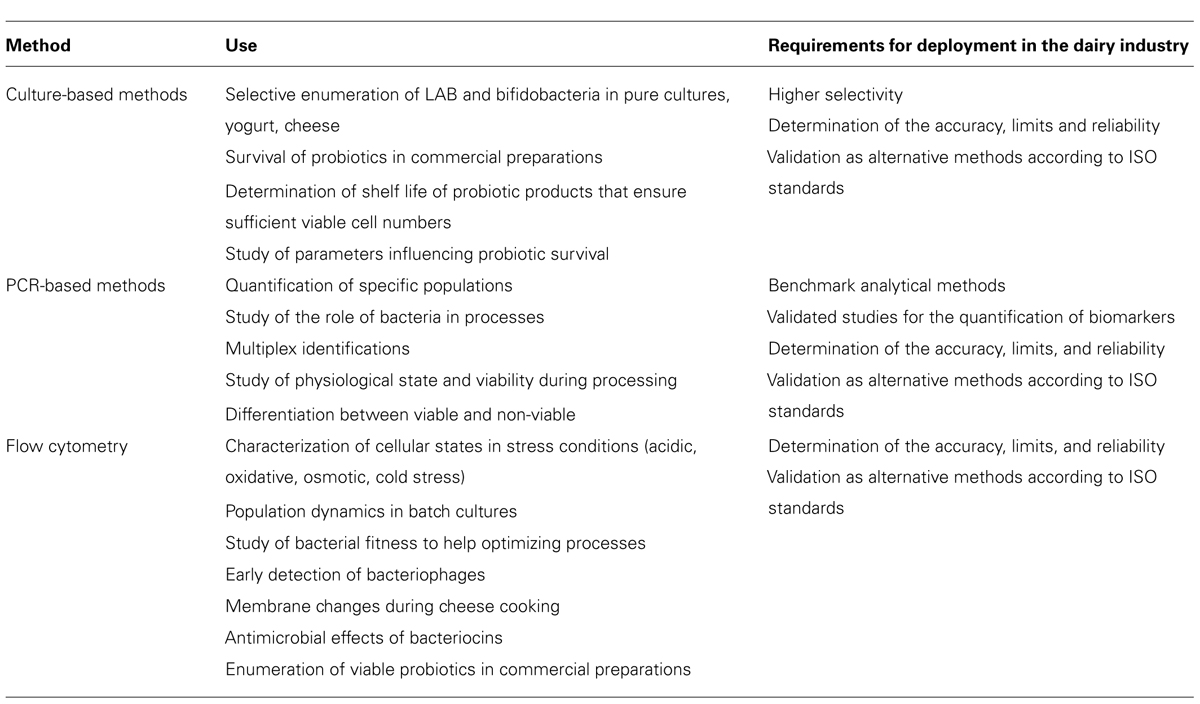
TABLE 1. Current uses and lacks of the most employed analytical methods.
Promising FC methods were described for analyses of mixed cultures in milk, yogurt, and other fermented milk products. They could be applied for quality controls during processes or during product shelf life. Nevertheless, they would require careful optimization to prevent artifacts due to the presence of milk proteins. One must also keep in mind that the quantification limits remain high and that current probes are not specific for a species or genus but for a target cell component. Therefore FC would rather be used to characterize physiological states than for specific counts.
OMICS data have allowed identifying many key biomarkers, which could be quantified in order to quickly characterize physiological states. PCR is already routinely used to detect food-borne pathogens and spoilage microbes. This could easily be extended to fermenting and probiotic bacteria, providing standardization of nucleic acid extraction, PCR procedures, and data interpretation. PCR (and PCR-based methods) would be a useful tool to quickly select interesting fermenting strains, or to ensure that optimal conditions are present during processes. Today, standards, patents, or commercial kits that would allow to quickly transferring methods to study fitness and physiological states of LAB and probiotics are not available.
Common to all the techniques reviewed in this study, two major gaps were identified: evaluation of the methods in agreement with the specific needs of the dairy industry (characterization of the robustness, accuracy, and limits) and validation according to official standards (Table 1). Indeed, while standard methods have been developed and validated for food-borne pathogens, LAB and probiotics seem to be the poor relations of the diagnosis industry and method standardization (Figure 1). This is surprising when considering the amount of quality controls performed by the dairyindustry.
Author Contributions
Danièle Sohier has designed the study, analyzed the data, and reviewed the manuscript. Sonia Pavan has written the manuscript. Armelle Riou has collected the data. Jérôme Combrisson and Florence Postollec have reviewed the manuscript and participated in the discussion.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was carried out according to the request of BBA dairy association (Rennes, France) and was supported by VALORIAL competitiveness pole (Rennes, France).
References
Abdulamir, A., Yoke, T., Nordin, N., and Abu Bakar, F. (2010). Detection and quantification of probiotic bacteria using optimized DNA extraction, traditional and real-time PCR methods in complex microbial communities. Afr. J. Biotechnol. 9, 1481–1492.
Ananta, E., Volkert, M., and Knorr, D. (2005). Cellular injuries and storage stability of spray-dried Lactobacillus rhamnosus GG. Int. Dairy J. 15, 399–409. doi: 10.1016/j.idairyj.2004.08.004
Antunes, A. E. C., Grael, E. T., Moreno, I., Rodrigues, L. G., Dourado, F. M., Saccaro, D. M., et al. (2007). Selective enumeration and viability of Bifidobacterium animalis subsp. lactis in a new fermented milk product. Braz. J. Microbiol. 38, 173–177. doi: 10.1590/S1517-83822007000100035
Ashraf, R., and Shah, N. P. (2011). Selective and differential enumerations of Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus, Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium spp. in yoghurt – a review. Int. J. Food Microbiol. 149, 194–208. doi: 10.1016/j.ijfoodmicro.2011.07.008
Atkinson, A., Colburn, W. A., Degruttola, V. G., Demets, D. L., Downing, G. J., Hoth, D. F., et al. (2001). Biomarkers and surrogate endpoints: preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 69, 89–95. doi: 10.1067/mcp.2001.113989
Auty, M. A. E., Gardiner, G. E., McBrearty, S. J., O’Sullivan, E. O., Mulvihill, D. M., Collins, J. K., et al. (2001). Direct in situ viability assessment of bacteria in probiotic dairy products using viability staining in conjunction with confocal scanning laser microscopy. Appl. Environ. Microbiol. 67, 420–425. doi: 10.1128/AEM.67.1.420-425.2001
Azcarate-Peril, M. A., Tallon, R., and Klaenhammer, T. R. (2009). Temporal gene expression and probiotic attributes of Lactobacillus acidophilus during growth in milk. J. Dairy Sci. 92, 870–886. doi: 10.3168/jds.2008–1457
Babot, J. D., Hidalgo, M., Arganaraz-Martinez, E., Apella, M. C., and Perez Chaia, A. (2011). Fluorescence in situ hybridization for detection of classical propionibacteria with specific 16S rRNA-targeted probes and its application to enumeration in Gruyere cheese. Int. J. Food Microbiol. 145, 221–228. doi: 10.1016/j.ijfoodmicro.2010.12.024
Bartlett, J. M., and Stirling, D. (2003). A short history of the polymerase chain reaction. Methods Mol. Biol. 226, 3–6. doi: 10.1385/1-59259-384-4-3
Ben Amor, K., Vaughan, E. E., and De Vos, W. M. (2007). Advanced molecular tools for the identification of lactic acid bacteria. J. Nutr. 137, 741S–747S.
Bernardeau, M., Vernoux, J. P., and Gueguen, M. (2001). Usefulness of epifluorescence for quantitative analysis of lactobacilli in probiotic feed. J. Appl. Microbiol. 91, 1103–1109. doi: 10.1046/j.1365-2672.2001.01481.x
Berney, M., Hammes, F., Bosshard, F., Weilenmann, H. U., and Egli, T. (2007). Assessment and interpretation of bacterial viability by using the LIVE/DEAD BacLight Kit in combination with flow cytometry. Appl. Environ. Microbiol. 73, 3283–3290. doi: 10.1128/aem.02750-6
Bottari, B., Ercolini, D., Gatti, M., and Neviani, E. (2006). Application of FISH technology for microbiological analysis: current state and prospects. Appl. Microbiol. Biotechnol. 73, 485–494. doi: 10.1007/s00253-006-0615-z
Bove, C. G., Lazzi, C., Bernini, V., Bottari, B., Neviani, E., and Gatti, M. (2011). cDNA-amplified fragment length polymorphism to study the transcriptional responses of Lactobacillus rhamnosus growing in cheese-like medium. J. Appl. Microbiol. 111, 855–864. doi: 10.1111/j.1365-2672.2011.05101.x
Boyer, M., and Combrisson, J. (2013). Analytical opportunities of quantitative polymerase chain reaction in dairy microbiology. Int. Dairy J. 30, 45–52. doi: 10.1016/j.idairyj.2012.11.008
Budde, B. B., and Rasch, M. (2001). A comparative study on the use of flow cytometry and colony forming units for assessment of the antibacterial effect of bacteriocins. Int. J. Food Microbiol. 63, 65–72. doi: 10.1016/S0168-1605(00)00399–398
Bunthof, C. J., and Abee, T. (2002). Development of a flow cytometric method to analyze subpopulations of bacteria in probiotic products and dairy starters. Appl. Environ. Microbiol. 68, 2934–2942. doi: 10.1128/AEM.68.6.2934-2942.2002
Bustin, S. A. (2009). Why the need for qPCR publication guidelines? The case for MIQE. Methods 50, 217–226. doi: 10.1016/j.ymeth.2009.12.006
Bustin, S. A., Benes, V., Garson, J. A., Hellemans, J., Huggett, J., Kubista, M., et al. (2009). The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622. doi: 10.1373/clinchem.2008.112797
Caplice, E., and Fitzgerald, G. F. (1999). Food fermentations: role of microorganisms in food production and preservation. Int. J. Food Microbiol. 15, 131–149. doi: 10.1016/S0168-1605(99)00082–83
Champagne, C. P., Raymond, Y., and Tompkins, T. A. (2010). The determination of viable counts in probiotic cultures microencapsulated by spray-coating. Food Microbiol. 27, 1104–1111. doi: 10.1016/j.fm.2010.07.017
Champagne, C. P., Ross, R. P., Saarela, M., Hansen, K. F., and Charalampopoulos, D. (2011). Recommendations for the viability assessment of probiotics as concentrated cultures and in food matrices. Int. J. Food Microbiol. 149, 185–193. doi: 10.1016/j.ijfoodmicro.2011.07.005
Chen, S., Ferguson, L. R., Shu, Q., and Garg, S. (2011). The application of flow cytometry to the characterisation of a probiotic strain Lactobacillus reuteri DPC16 and the evaluation of sugar preservatives for its lyophilization. Food Sci. Technol. 44, 1873–1879. doi: 10.1016/j.lwt.2011.05.006
Collado, M. C., Moreno, Y., Cobo, J., and Hernández, M. (2006). Microbiological evaluation and molecular characterization of bifidobacteria strains in commercial fermented milks. Eur. Food Res. Technol. 222, 112–117. doi: 10.1007/s00217-005-0093-z
Comas-Riu, J., and Rius, N. (2009). Flow cytometry applications in the food industry. J. Ind. Microbiol. Biotechnol. 36, 999–1011. doi: 10.1007/s10295-009-0608-x
Corcoran, B. M., Ross, R. P., Fitzgerald, G. F., and Stanton, C. (2004). Comparative survival of probiotic lactobacilli spray-dried in the presence of prebiotic substances. J. Appl. Microbiol. 96, 1024–1039. doi: 10.1111/j.1365-2672.2004.02219.x
Cretenet, M., Laroute, V., Ulve, V., Jeanson, S., Nouaille, S., Even, S., et al. (2011). Dynamic analysis of the Lactococcus lactis transcriptome in cheeses made from milk concentrated by ultrafiltration reveals multiple strategies of adaptation to stresses. Appl. Environ. Microbiol. 77, 247–257. doi: 10.1128/aem.01174–1110
Davey, H. M. (2011). Life, death, and in-between: meanings and methods in microbiology. Appl. Environ. Microbiol. 77, 5571–5576. doi: 10.1128/aem.00744–711
De Carvalho Lima, K. G., Kruger, M. F., Behrens, J., Destro, M. T., Landgraf, M., and De Melo Franco, B. D. G. (2009). Evaluation of culture media for enumeration of Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium animalis in the presence of Lactobacillus delbrueckii subsp bulgaricus and Streptococcus thermophilus. Food Sci. Technol. 42, 491–495. doi: 10.1016/j.lwt.2008.08.011
De Dea Lindner, J., Canchaya, C., Zhang, Z., Neviani, E., Fitzgerald, G. F., Van Sinderen, D., et al. (2007). Exploiting Bifidobacterium genomes: the molecular basis of stress response. Int. J. Food Microbiol. 120, 13–24. doi: 10.1016/j.ijfoodmicro.2007.06.016
Dekker, J., Glenn, M., Havukkala, I. J., and Lubbers, M. W. (2002). Lactobacillus rhamnosus Polynucleotides, Polypeptides and Methods for using them. WO2002044383 A1. NZ patent application.
Dekker, J., Glenn, M., Havukkala, I. J., and Lubbers, M. W. (2004). Polynucleotides, Materials Incorporating them, and Methods for using them. US 20040009490 A1.
Devonshire, A. S., Sanders, R., Wilkes, T. M., Taylor, M. S., Foy, C. A., and Huggett, J. F. (2013). Application of next generation qPCR and sequencing platforms to mRNA biomarker analysis. Methods 59, 89–100. doi: 10.1016/j.ymeth.2012.07.021
de Vos, W. M. (2011). Systems solutions by lactic acid bacteria: from paradigms to practice. Microb. Cell Fact. 10(Suppl. 1), S2. doi: 10.1186/1475-2859-10-s1-s2
Díaz, M., Herrero, M., García, L. A., and Quirós, C. (2010). Application of flow cytometry to industrial microbial bioprocesses. Biochem. Eng. J. 48, 385–407. doi: 10.1016/j.bej.2009.07.013
do Espírito Santo, A. P., Perego, P., Converti, A., and Oliveira, M. N. (2011). Influence of food matrices on probiotic viability – a review focusing on the fruity bases. Trends Food Sci. Technol. 22, 377–385. doi: 10.1016/j.tifs.2011.04.008
Doherty, S. B., Wang, L., Ross, R. P., Stanton, C., Fitzgerald, G. F., and Brodkorb, A. (2010). Use of viability staining in combination with flow cytometry for rapid viability assessment of Lactobacillus rhamnosus GG in complex protein matrices. J. Microbiol. Methods 82, 301–310. doi: 10.1016/j.mimet.2010.07.003
Donkor, O. N., Tsangalis, D., and Shah, N. P. (2007). Viability of probiotic bacteria and concentrations of organic acids in commercial yoghurts during refrigerated storage. Food Aust. 59, 121–126.
El Arbi, A., Ghorbal, S., Delacroix-Buchet, A., and Bouix, M. (2011). Assessment of the dynamics of the physiological states of Lactococcus lactis ssp. cremoris SK11 during growth by flow cytometry. J. Appl. Microbiol. 111, 1205–1211. doi: 10.1111/j.1365-2672.2011.05114.x
Falentin, H., Postollec, F., Parayre, S., Henaff, N., Le Bivic, P., Richoux, R., et al. (2010). Specific metabolic activity of ripening bacteria quantified by real-time reverse transcription PCR throughout Emmental cheese manufacture. Int. J. Food Microbiol. 144, 10–19. doi: 10.1016/j.ijfoodmicro.2010.06.003
Fittipaldi, M., Nocker, A., and Codony, F. (2012). Progress in understanding preferential detection of live cells using viability dyes in combination with DNA amplification. J. Microbiol. Methods 91, 276–289. doi: 10.1016/j.mimet.2012.08.007
Forssten, S. D., Sindelar, C. W., and Ouwehand, A. C. (2011). Probiotics from an industrial perspective. Anaerobe 17, 410–413. doi: 10.1016/j.anaerobe.2011.04.014
Friedrich, U., and Lenke, J. (2006). Improved enumeration of lactic acid bacteria in mesophilic dairy starter cultures by using multiplex quantitative real-time PCR and flow cytometry-fluorescence in situ hybridization. Appl. Environ. Microbiol. 72, 4163–4171. doi: 10.1128/aem.02283–2285
García-Cayuela, T., Tabasco, R., Peláez, C., and Requena, T. (2009). Simultaneous detection and enumeration of viable lactic acid bacteria and bifidobacteria in fermented milk by using propidium monoazide and real-time PCR. Int. Dairy J. 19, 405–409. doi: 10.1016/j.idairyj.2009.02.001
Garcia-Hernandez, J., Moreno, Y., Amorocho, C. M., and Hernandez, M. (2012). A combination of direct viable count and fluorescence in situ hybridization for specific enumeration of viable Lactobacillus delbrueckii subsp.bulgaricus and Streptococcus thermophilus. Lett. Appl. Microbiol. 54, 247–254. doi: 10.1111/j.1472-765X.2011.03201.x
Gendre, F., and Brignon, P. (1998). Method for Detecting Live Microbiological Contaminants in a Food Product Sample. Patent FR2755145 (A1)–1998-04-30.
Gitton, C., Meyrand, M., Wang, J., Caron, C., Trubuil, A., Guillot, A., et al. (2005). Proteomic signature of Lactococcus lactis NCDO763 cultivated in milk. Appl. Environ. Microbiol. 71, 7152–7163. doi: 10.1128/aem.71.11.7152-7163.2005
Glenn, M., Havukkala, I. J., Lubbers, M. W., and Dekker, J. (2004). Polynucleotides and Polypeptides Derived from Lactobacillus rhamnosus HN001. WO2004031389. NZ patent application.
Gunasekera, T. S., Attfield, P. V., and Veal, D. A. (2000). A flow cytometry method for rapid detection and enumeration of total bacteria in milk. Appl. Environ. Microbiol. 66, 1228–1232. doi: 10.1128/AEM.66.3.1228-1232.2000
Hayden, R. T., Gu, Z., Ingersoll, J., Abdul-Ali, D., Shi, L., Pounds, S., et al. (2013). Comparison of droplet digital PCR to real-time PCR for quantitative detection of cytomegalovirus. J. Clin. Microbiol. 51, 540–546. doi: 10.1128/jcm.02620–2612
Heredia, N. J., Belgrader, P., Wang, S., Koehler, R., Regan, J., Cosman, A. M., et al. (2013). Droplet Digital PCR quantitation of HER2 expression in FFPE breast cancer samples. Methods 59, S20–S23. doi: 10.1016/j.ymeth.2012.09.012
Herve-Jimenez, L., Guillouard, I., Guedon, E., Gautier, C., Boudebbouze, S., Hols, P., et al. (2008). Physiology of Streptococcus thermophilus during the late stage of milk fermentation with special regard to sulfur amino-acid metabolism. Proteomics 8, 4273–4286. doi: 10.1002/pmic.200700489
Hua, Z., Rouse, J. L., Eckhardt, A. E., Srinivasan, V., Pamula, V. K., Schell, W. A., et al. (2010). Multiplexed real-time polymerase chain reaction on a digital microfluidic platform. Anal. Chem. 82, 2310–2316. doi: 10.1021/ac902510u
Huggett, J. F., Foy, C. A., Benes, V., Emslie, K., Garson, J. A., Haynes, R., et al. (2013). The digital MIQE guidelines: minimum information for publication of quantitative digital PCR experiments. Clin. Chem. 59, 892–902. doi: 10.1373/clinchem.2013.206375
International Dairy Federation. (2012). IDF Programme of Work. Available at: http://www.ukidf.org/documents/pow_Sep12.pdf (accessed October 10, 2013).
ISO. (1998). Microbiology of Food and Animal Feeding Stuffs – Horizontal Method for the Enumeration of Mesophilic Lactic Acid Bacteria – Colony-Count Technique at 30 Degrees. Geneva: International Standardisation Organisation
ISO. (2003a). Microbiology of Food and Animal Feeding Stuffs – Protocol for the Validation of Alternative Methods. Geneva: International Standardisation Organisation.
ISO. (2003b). Yogurt – Identification of Characteristic Microorganisms (Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus). Geneva International Standardisation Organisation.
ISO. (2003c). Yogurt – Enumeration of Characteristic Microorganisms –- Colony-Count Technique at 37 Degrees. Geneva: International Standardisation Organisation.
ISO. (2005a). Microbiology of Food and Animal Feeding Stuffs – Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens – Performance Testing for Thermal Cyclers. Geneva: International Standardisation Organisation.
ISO. (2005b). Microbiology of Food and Animal Feeding Stuffs. Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens. General Requirements and Definitions. Geneva: International Standardisation Organisation.
ISO. (2006a). Microbiology of Food and Animal Feeding Stuffs – Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens – Requirements for Amplification and Detection for Qualitative Methods. Geneva: International Standardisation Organisation.
ISO. (2006b). Microbiology of Food and Animal Feeding Stuffs – Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens – Requirements for Sample Preparation for Qualitative Detection. Geneva: International Standardisation Organisation.
ISO. (2006c). Milk, Milk Products and Mesophilic Starter Cultures – Enumeration of Citrate-Fermenting Lactic Acid Bacteria – Colony-Count Technique at 25 Degrees. Geneva: International Standardisation Organisation.
ISO. (2007). Microbiology of Food and Animal Feeding Stuffs – General Requirements and Guidance for Microbiological Examinations. Geneva: International Standardisation Organisation.
ISO. (2010a). Fermented Milk Products – Bacterial Starter Cultures – Standard of Identity. Geneva: International Standardisation Organisation.
ISO. (2010b). Milk Products – Enumeration of Presumptive Bifidobacteria – Colony Count Technique at 37 Degrees. Geneva: International Standardisation Organisation.
ISO. (2011a). Microbiology of Food and Animal Feeding Stuffs – Polymerase Chain Reaction (PCR) for the Detection and Quantification of Food-Borne Pathogens – Performance Characteristics. Geneva: International Standardisation Organisation.
ISO. (2011b). Microbiology of Food and Animal Feeding Stuffs – Real-Time Polymerase Chain Reaction (PCR) for the Detection of Food-Borne Pathogens – General Requirements and Definitions. Geneva: International Standardisation Organisation.
ISO. (2012). Microbiology of Food and Animal Feed – Real-Time Polymerase Chain Reaction (PCR)-Based Method for the Detection of Food-Borne Pathogens – Horizontal Method for the Detection of Shiga Toxin-Producing Escherichia coli (stec) and the Determination of o157, o111, o26, o103 and o145 Serogroups. Geneva: International Standardisation Organisation.
ISO. (2013). Microbiology of Food and Animal Feed – Horizontal Method for Determination of Hepatitis A Virus and Norovirus in Food using Real-Time RT-PCR – Part 1: Method for Quantification. Geneva: International Standardisation Organisation.
Izquierdo, E., Horvatovich, P., Marchioni, E., Aoude-Werner, D., Sanz, Y., and Ennahar, S. (2009). Two-DE and MS analysis of key proteins in the adhesion of Lactobacillus plantarum, a first step toward early selection of probiotics based on bacterial biomarkers. Electrophoresis 30, 949–956. doi: 10.1002/elps.200800399
Karimi, R., Mortazavian, A., and Cruz, A. (2011). Viability of probiotic microorganisms in cheese during production and storage: a review. Dairy Sci. Technol. 91, 283–308. doi: 10.1007/s13594-011-0005-x
Karimi, R., Mortazavian, A. M., and Amiri-Rigi, A. (2012). Selective enumeration of probiotic microorganisms in cheese. Food Microbiol. 29, 1–9. doi: 10.1016/j.fm.2011.08.008
Kell, D. B., Kaprelyants, A. S., Weichart, D. H., Harwood, C. R., and Barer, M. R. (1998). Viability and activity in readily culturable bacteria: a review and discussion of the practical issues. Antonie Van Leeuwenhoek 73, 169–187. doi: 10.1023/A:1000664013047
Kelley, K., Cosman, A., Belgrader, P., Chapman, B., and Sullivan, D. C. (2013). Detection of methicillin-resistant Staphylococcus aureus by a duplex droplet digital polymerase chain reaction. J. Clin. Microbiol. 51, 2033–2039. doi: 10.1128/jcm.00196-13
Klaenhammer, T., Russell, W. M., and Altermann, E. (2006). Nucleic Acid Sequences Encoding Two-Component Sensing and Regulatory Proteins, Antimicrobial Proteins and Uses Therefore. US 20060134745 A1.
Klaenhammer, T. R., Altermann, E., Azcarate-Peril, A., McAuliffe, O., and Russell, W. M. (2005). Lactobacillus acidophilus Nucleic Acid Sequences Encoding Stress-Related Proteins and Uses Therefore. WO2005086794. USA patent application.
Klaenhammer, T. R., Russell, W. M., Altermann, E., and Azcarate-Peril, A. (2010). Nucleic Acids Encoding Two-Component Sensing and Regulatory Proteins, Antimicrobial Proteins And Uses Therefore. US 20100279349 A1.
Klaenhammer, T. R., Russell, W. M., Altermann, E., Cano, R. J., and Hamrick, A. (2004). Lactobacillus acidophilus Nucleic Acid Sequences Encoding Cell-Surface Protein Homologues and Uses Therefore. WO2004096992. USA patent application.
Kramer, M., Obermajer, N., Bogovic Matijasic, B., Rogelj, I., and Kmetec, V. (2009). Quantification of live and dead probiotic bacteria in lyophilised product by real-time PCR and by flow cytometry. Appl. Microbiol. Biotechnol. 84, 1137–1147. doi: 10.1007/s00253-009-2068-7
Lahtinen, S. J., Ahokoski, H., Reinikainen, J. P., Gueimonde, M., Nurmi, J., Ouwehand, A. C., et al. (2008). Degradation of 16S rRNA and attributes of viability of viable but nonculturable probiotic bacteria. Lett. Appl. Microbiol. 46, 693–698. doi: 10.1111/j.1472-765X.2008.02374.x
Lahtinen, S. J., Ouwehand, A. C., Reinikainen, J. P., Korpela, J. M., Sandholm, J., and Salminen, S. J. (2006). Intrinsic properties of so-called dormant probiotic bacteria, determined by flow cytometric viability assays. Appl. Environ. Microbiol. 72, 5132–5134. doi: 10.1128/aem.02897-05
Le Dréan, G., Mounier, J., Vasseur, V., Arzur, D., Habrylo, O., and Barbier, G. (2010). Quantification of Penicillium camemberti and P. roqueforti mycelium by real-time PCR to assess their growth dynamics during ripening cheese. Int. J. Food Microbiol. 138, 100–107. doi: 10.1016/j.ijfoodmicro.2009.12.013
Lee, K., Rho, B. S., Pi, K., Kim, H. J., and Choi, Y. J. (2011). Proteomic analysis of protein expression in Lactobacillus plantarum in response to alkaline stress. J. Biotechnol. 153, 1–7. doi: 10.1016/j.jbiotec.2011.02.008
Li, J. S., Bi, Y. T., Dong, C., Yang, J. F., and Liang, W. D. (2011). Transcriptome analysis of adaptive heat shock response of Streptococcus thermophilus. PLoS ONE 6:e25777. doi: 10.1371/journal.pone.0025777
Liu, M., Van Enckevort, F. H., and Siezen, R. J. (2005). Genome update: lactic acid bacteria genome sequencing is booming. Microbiology 151, 3811–3814. doi: 10.1099/mic.0.28557-0
Lopes Da Silva, T., Piekova, L., Mileu, J., and Roseiro, J. (2009). A comparative study using the dual staining flow cytometric protocol applied to Lactobacillus rhamnosus and Bacillus licheniformis batch cultures. Enzyme Microb. Technol. 45, 134–138. doi: 10.1016/j.enzmictec.2009.03.001
Maecker, H. T., Frey, T., Nomura, L. E., and Trotter, J. (2004). Selecting fluorochrome conjugates for maximum sensitivity. Cytometry A 62, 169–173. doi: 10.1002/cyto.a.20092
Magariños, H., Selaive, S., Costa, M., Flores, M., and Pizarro, O. (2007). Viability of probiotic micro-organisms (Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp. lactis Bb-12) in ice cream. Int. J. Dairy Technol. 60, 128–134. doi: 10.1111/j.1471-0307.2007.00307.x
Makarova, K., Slesarev, A., Wolf, Y., Sorokin, A., Mirkin, B., Koonin, E., et al. (2006). Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. U.S.A. 103, 15611–15616. doi: 10.1073/pnas.0607117103
Malorny, B., Lofstrom, C., Wagner, M., Kramer, N., and Hoorfar, J. (2008). Enumeration of Salmonella bacteria in food and feed samples by real-time PCR for quantitative microbial risk assessment. Appl. Environ. Microbiol. 74, 1299–1304. doi: 10.1128/AEM.02489-07
Marco, M. L., and Kleerebezem, M. (2008). Assessment of real-time RT-PCR for quantification of Lactobacillus plantarum gene expression during stationary phase and nutrient starvation. J. Appl. Microbiol. 104, 587–594. doi: 10.1111/j.1365-2672.2007.03578.x
Masco, L., Vanhoutte, T., Temmerman, R., Swings, J., and Huys, G. (2007). Evaluation of real-time PCR targeting the 16S rRNA and recA genes for the enumeration of bifidobacteria in probiotic products. Int. J. Food Microbiol. 113, 351–357. doi: 10.1016/j.ijfoodmicro.2006.07.021
Masoud, W., Takamiya, M., Vogensen, F. K., Lillevang, S., Al-Soud, W. A., Sørensen, S. J., et al. (2011). Characterization of bacterial populations in Danish raw milk cheeses made with different starter cultures by denaturating gradient gel electrophoresis and pyrosequencing. Int. Dairy J. 21, 142–148. doi: 10.1016/j.idairyj.2010.10.007
Matijašić, B. B., Obermajer, T., and Rogelj, I. (2010). Quantification of Lactobacillus gasseri, Enterococcus faecium and Bifidobacterium infantis in a probiotic OTC drug by real-time PCR. Food Control 21, 419–425. doi: 10.1016/j.foodcont.2009.07.001
Matsuda, K., Tsuji, H., Asahara, T., Matsumoto, K., Takada, T., and Nomoto, K. (2009). Establishment of an analytical system for the human fecal microbiota, based on reverse transcription-quantitative PCR targeting of multicopy rRNA molecules. Appl. Environ. Microbiol.75, 1961–1969. doi: 10.1128/aem.01843-08
Maukonen, J., Alakomi, H. -L., Nohynek, L., Hallamaa, K., Leppämäki, S., Mättö, J., et al. (2006). Suitability of the fluorescent techniques for the enumeration of probiotic bacteria in commercial non-dairy drinks and in pharmaceutical products. Food Res. Int. 39, 22–32. doi: 10.1016/j.foodres.2005.05.006
Meng, X., Pang, R., Wang, C., and Wang, L. (2010). Rapid and direct quantitative detection of viable bifidobacteria in probiotic yogurt by combination of ethidium monoazide and real-time PCR using a molecular beacon approach. J. Dairy Res. 77, 498–504. doi: 10.1017/S0022029910000658
Michelsen, O., Cuesta-Dominguez, A., Albrechtsen, B., and Jensen, P. R. (2007). Detection of bacteriophage-infected cells of Lactococcus lactis by using flow cytometry. Appl. Environ. Microbiol. 73, 7575–7581. doi: 10.1128/aem.01219-07
Moreno, Y., Collado, M. C., Ferrús, M. A., Cobo, J. M., Hernández, E., and Hernández, M. (2006). Viability assessment of lactic acid bacteria in commercial dairy products stored at 4 °C using LIVE/DEAD® BacLightTM staining and conventional plate counts. Int. J. Food Sci. Technol. 41, 275–280. doi: 10.1111/j.1365-2621.2005.01060.x
Nebe-von-Caron, G., Stephens, P. J., Hewitt, C. J., Powell, J. R., and Badley, R. A. (2000). Analysis of bacterial function by multi-colour fluorescence flow cytometry and single cell sorting. J. Microbiol. Methods 42, 97–114. doi: 10.1016/S0167-7012(00)00181-0
O’Sullivan, O., O’Callaghan, J., Sangrador-Vegas, A., McAuliffe, O., Slattery, L., Kaleta, P., et al. (2009). Comparative genomics of lactic acid bacteria reveals a niche-specific gene set. BMC Microbiol. 9:50. doi: 10.1186/1471-2180-9-50
Oberg, C. J., Moyes, L. V., Domek, M. J., Brothersen, C., and McMahon, D. J. (2011). Survival of probiotic adjunct cultures in cheese and challenges in their enumeration using selective media. J. Dairy Sci. 94, 2220–2230. doi: 10.3168/jds.2010–3934
Ottesen, E. A., Hong, J. W., Quake, S. R., and Leadbetter, J. R. (2006). Microfluidic digital PCR enables multigene analysis of individual environmental bacteria. Science 314, 1464–1467. doi: 10.1126/science.1131370
Papadimitriou, K., Pratsinis, H., Nebe-Von-Caron, G., Kletsas, D., and Tsakalidou, E. (2006). Rapid assessment of the physiological status of Streptococcus macedonicus by flow cytometry and fluorescence probes. Int. J. Food Microbiol. 111, 197–205. doi: 10.1016/j.ijfoodmicro.2006.04.042
Papadimitriou, K., Pratsinis, H., Nebe-Von-Caron, G., Kletsas, D., and Tsakalidou, E. (2007). Acid tolerance of Streptococcus macedonicus as assessed by flow cytometry and single-cell sorting. Appl. Environ. Microbiol. 73, 465–476. doi: 10.1128/aem.01244–1246
Pastink, M. I., Teusink, B., Hols, P., Visser, S., De Vos, W. M., and Hugenholtz, J. (2009). Genome-scale model of Streptococcus thermophilus LMG18311 for metabolic comparison of lactic acid bacteria. Appl. Environ. Microbiol. 75, 3627–3633. doi: 10.1128/aem.00138–139
Phillips, M., Kailasapathy, K., and Tran, L. (2006). Viability of commercial probiotic cultures (L. acidophilus, Bifidobacterium sp., L. casei, L. paracasei and L. rhamnosus) in cheddar cheese. Int. J. Food Microbiol. 108, 276–280. doi: 10.1016/j.ijfoodmicro.2005.12.009
Postgate, J. R., Crumpton, J. E., and Hunter, J. R. (1961). The measurement of bacterial viabilities by slide culture. J. Gen. Microbiol. 24, 15–24. doi: 10.1099/00221287-24-1-15
Postollec, F., Falentin, H., Pavan, S., Combrisson, J., and Sohier, D. (2011). Recent advances in quantitative PCR (qPCR) applications in food microbiology. Food Microbiol. 28, 848–861. doi: 10.1016/j.fm.2011.02.008
Quiros, C., Herrero, M., Garcia, L. A., and Diaz, M. (2007). Application of flow cytometry to segregated kinetic modeling based on the physiological states of microorganisms. Appl. Environ. Microbiol. 73, 3993–4000. doi: 10.1128/aem.00171–177
Randazzo, C. L., Caggia, C., and Neviani, E. (2009). Application of molecular approaches to study lactic acid bacteria in artisanal cheeses. J. Microbiol. Methods 78, 1–9. doi: 10.1016/j.mimet.2009.04.001
Rasmussen, T. B., Danielsen, M., Valina, O., Garrigues, C., Johansen, E., and Pedersen, M. B. (2008). Streptococcus thermophilus core genome: comparative genome hybridization study of 47 strains. Appl. Environ. Microbiol. 74, 4703–4710. doi: 10.1128/aem.00132–138
Raynaud, S., Perrin, R., Cocaign-Bousquet, M., and Loubiere, P. (2005). Metabolic and transcriptomic adaptation of Lactococcus lactis subsp. lactis biovar diacetylactis in response to autoacidification and temperature downshift in skim milk. Appl. Environ. Microbiol. 71, 8016–8023. doi: 10.1128/aem.71.12.8016-8023.2005
Reid, G., Gan, B. S., She, Y. M., Ens, W., Weinberger, S., and Howard, J. C. (2002). Rapid identification of probiotic Lactobacillus biosurfactant proteins by ProteinChip tandem mass spectrometry tryptic peptide sequencing. Appl. Environ. Microbiol. 68, 977–980. doi: 10.1128/AEM.68.2.977-980.2002
Reimann, S., Grattepanche, F., Rezzonico, E., and Lacroix, C. (2010). Development of a real-time RT-PCR method for enumeration of viable Bifidobacterium longum cells in different morphologies. Food Microbiol. 27, 236–242. doi: 10.1016/j.fm.2009.10.010
Roszak, D. B., and Colwell, R. R. (1987). Survival strategies of bacteria in the natural environment. Microbiol. Rev. 51, 365–379.
Roszak, D. B., Grimes, D. J., and Colwell, R. R. (1984). Viable but nonrecoverable stage of Salmonella enteritidis in aquatic systems. Can. J. Microbiol. 30, 334–338. doi: 10.1139/m84-049
Ruiz, L., Sanchez, B., De Los Reyes-Gavilan, C. G., Gueimonde, M., and Margolles, A. (2009). Coculture of Bifidobacterium longum and Bifidobacterium breve alters their protein expression profiles and enzymatic activities. Int. J. Food Microbiol. 133, 148–153. doi: 10.1016/j.ijfoodmicro.2009.05.014
Saccaro, D. M., Hirota, C. Y., Tamime, A. Y., and De Oliveira, M. N. (2011). Evaluation of different selective media for enumeration of probiotic micro-organisms in combination with yogurt starter cultures in fermented milk. African J. Microbiol. Res. 6, 2239–2245.
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W. S., et al. (2011). Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60. doi: 10.1186/gb-2011-12-6-r60
Senan, S., Grover, S., and Batish, V. K. (2008). Comparison of specificity of different primer pairs for the development of multiplex PCR assays for rapid identification of dairy lactobacilli. Int. J. Sci.Technol. 3, 123–137.
Sheehan, A., O’Loughlin, C., O’Cuinn, G., Fitzgerald, R. J., and Wilkinson, M. G. (2005). Cheddar cheese cooking temperature induces differential lactococcal cell permeabilization and autolytic responses as detected by flow cytometry: implications for intracellular enzyme accessibility. J. Appl. Microbiol. 99, 1007–1018. doi: 10.1111/j.1365-2672.2005.02718.x
Sheu, S. J., Hwang, W. Z., Chen, H. C., Chiang, Y. C., and Tsen, H. Y. (2009). Development and use of tuf gene-based primers for the multiplex PCR detection of Lactobacillus acidophilus, Lactobacillus casei group, Lactobacillus delbrueckii, and Bifidobacterium longum in commercial dairy products. J. Food Prot. 72, 93–100.
Sheu, S. J., Hwang, W. Z., Chiang, Y. C., Lin, W. H., Chen, H. C., and Tsen, H. Y. (2010). Use of tuf gene-based primers for the PCR detection of probiotic Bifidobacterium species and enumeration of bifidobacteria in fermented milk by cultural and quantitative real-time PCR methods. J. Food Sci. 75, M521–M527. doi: 10.1111/j.1750-3841.2010.01816.x
Sieuwerts, S., De Bok, F. A., Hugenholtz, J., and Van Hylckama Vlieg, J. E. (2008). Unraveling microbial interactions in food fermentations: from classical to genomics approaches. Appl. Environ. Microbiol. 74, 4997–5007. doi: 10.1128/aem.00113–118
Sieuwerts, S., Molenaar, D., Van Hijum, S. A., Beerthuyzen, M., Stevens, M. J., Janssen, P. W., et al. (2010). Mixed-culture transcriptome analysis reveals the molecular basis of mixed-culture growth in Streptococcus thermophilus and Lactobacillus bulgaricus. Appl. Environ. Microbiol. 76, 7775–7784. doi: 10.1128/aem.01122–1110
Siezen, R. J., Starrenburg, M. J., Boekhorst, J., Renckens, B., Molenaar, D., and Van Hylckama Vlieg, J. E. (2008). Genome-scale genotype-phenotype matching of two Lactococcus lactis isolates from plants identifies mechanisms of adaptation to the plant niche. Appl. Environ. Microbiol. 74, 424–436. doi: 10.1128/aem.01850–1857
Siezen, R. J., Van Enckevort, F. H., Kleerebezem, M., and Teusink, B. (2004). Genome data mining of lactic acid bacteria: the impact of bioinformatics. Curr. Opin. Biotechnol. 15, 105–115. doi: 10.1016/j.copbio.2004.02.002
Smith, C. J., and Osborn, A. M. (2009). Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS Microbiol. Ecol. 67, 6–20. doi: 10.1111/j.1574-6941.2008.00629.x
Sohier, D., Jamet, E., Le Dizes, A-S., Dizin, M., Pavan, S., Postollec, F., et al. (2012). Polyphasic approach for quantitative analysis of obligately heterofermentative Lactobacillus species in cheese. Food Microbiol. 31, 271–277. doi: 10.1016/j.fm.2012.01.009
Song, L., Shan, D., Zhao, M., Pink, B. A., Minnehan, K. A., York, L., et al. (2013). Direct detection of bacterial genomic DNA at sub-femtomolar concentrations using single molecule arrays. Anal. Chem. 85, 1932–1939. doi: 10.1021/ac303426b
Sul, S. Y., Kim, H. J., Kim, T. W., and Kim, H. Y. (2007). Rapid identification of Lactobacillus and Bifidobacterium in probiotic products using multiplex PCR. J. Microbiol. Biotechnol. 17, 490–495.
Sunny-Roberts, E. O., and Knorr, D. (2008). Evaluation of the response of Lactobacillus rhamnosus VTT E-97800 to sucrose-induced osmotic stress. Food Microbiol. 25, 183–189. doi: 10.1016/j.fm.2007.05.003
Taibi, A., Dabour, N., Lamoureux, M., Roy, D., and Lapointe, G. (2011). Comparative transcriptome analysis of Lactococcus lactis subsp. cremoris strains under conditions simulating Cheddar cheese manufacture. Int. J. Food Microbiol. 146, 263–275. doi: 10.1016/j.ijfoodmicro.2011.02.034
Tharmaraj, N., and Shah, N. P. (2003). Selective enumeration of Lactobacillus delbrueckii ssp. bulgaricus, Streptococcus thermophilus, Lactobacillus acidophilus, bifidobacteria, Lactobacillus casei, Lactobacillus rhamnosus, and propionibacteria. J. Dairy Sci. 86, 2288–2296. doi: 10.3168/jds.S0022-0302(03)73821–73821
Thevenard, B., Rasoava, N., Fourcassie, P., Monnet, V., Boyaval, P., and Rul, F. (2011). Characterization of Streptococcus thermophilus two-component systems in silico analysis, functional analysis and expression of response regulator genes in pure or mixed culture with its yogurt partner, Lactobacillus delbrueckii subsp. bulgaricus. Int. J. Food Microbiol. 151, 171–181. doi: 10.1016/j.ijfoodmicro.2011.08.019
Thierry, A., Deutsch, S. M., Falentin, H., Dalmasso, M., Cousin, F. J., and Jan, G. (2011). New insights into physiology and metabolism of Propionibacterium freudenreichii. Int. J. Food Microbiol. 149, 19–27. doi: 10.1016/j.ijfoodmicro.2011.04.026
Tracy, B. P., Gaida, S. M., and Papoutsakis, E. T. (2010). Flow cytometry for bacteria: enabling metabolic engineering, synthetic biology and the elucidation of complex phenotypes. Curr. Opin. Biotechnol. 21, 85–99. doi: 10.1016/j.copbio.2010.02.006
Van de Casteele, S., Vanheuverzwijn, T., Ruyssen, T., Van Assche, P., Swings, J., and Huys, G. (2006). Evaluation of culture media for selective enumeration of probiotic strains of lactobacilli and bifidobacteria in combination with yoghurt or cheese starters. Int. Dairy J. 16, 1470–1476. doi: 10.1016/j.idairyj.2005.12.002
Ventura, M., O’Connell-Motherway, M., Leahy, S., Moreno-Munoz, J. A., Fitzgerald, G. F., and Van Sinderen, D. (2007). From bacterial genome to functionality; case bifidobacteria. Int. J. Food Microbiol. 120, 2–12. doi: 10.1016/j.ijfoodmicro.2007.06.011
White, R. A. III, Blainey, P. C., Fan, H. C., and Quake, S. R. (2009). Digital PCR provides sensitive and absolute calibration for high throughput sequencing. BMC Genomics 10:116. doi: 10.1186/1471-2164-10–116
Wong, M. L., and Medrano, J. F. (2005). Real-time PCR for mRNA quantitation. Biotechniques 39, 75–85. doi: 10.2144/05391RV01
Zhong, Q., Bhattacharya, S., Kotsopoulos, S., Olson, J., Taly, V., Griffiths, A. D., et al. (2011). Multiplex digital PCR: breaking the one target per color barrier of quantitative PCR. Lab Chip 11, 2167–2174. doi: 10.1039/c1lc20126c
Keywords: dairy, food microbiology, probiotic, analytical method, bacterial fitness, PCR, flow cytometry, quality control
Citation: Sohier D, Pavan S, Riou A, Combrisson J and Postollec F (2014) Evolution of microbiological analytical methods for dairy industry needs. Front. Microbiol. 5:16. doi: 10.3389/fmicb.2014.00016
Received: 15 October 2013; Accepted: 10 January 2014;
Published online: 07 February 2014.
Edited by:
Danilo Ercolini, Università degli Studi di Napoli Federico II, ItalyReviewed by:
Paul David Cotter, Teagasc, IrelandAntonia Picon, Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, Spain
Copyright © 2014 Sohier, Pavan, Riou, Combrisson and Postollec. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Danièle Sohier, Food Safety and Quality Unit, ADRIA Développement, Agri-Food Technical Institute, UMT 08.3, Z.A. Creac’h Gwen, 29196 Quimper, France e-mail:ZGFuaWVsZS5zb2hpZXJAYWRyaWEudG0uZnI=
†Present address: Jérôme Combrisson, Sample preparation Competence Centre, bioMerieux, Grenoble, France