 Bhabatosh Das
Bhabatosh Das- Centre for Human Microbial Ecology, Translational Health Science and Technology Institute, Gurgaon, India
Vibrio cholerae, the etiological agent of acute diarrhoeal disease cholera, harbors large numbers of lysogenic filamentous phages, contribute significantly to the host pathogenesis and provide fitness factors to the pathogen that help the bacterium to survive in natural environment. Most of the vibriophage genomes are not equipped with integrase and thus exploit two host-encoded tyrosine recombinases, XerC and XerD, for lysogenic conversion. Integration is site-specific and it occurs at dimer resolution site (dif) of either one or both chromosomes of V. cholerae. Each dif sequence contains two recombinase-binding sequences flanking a central region. The integration follows a sequential strand exchanges between dif and attP sites within a DNA-protein complex consisting of one pair of each recombinase and two DNA fragments. During entire process of recombination, both the DNA components and recombinases of the synaptic complex keep transiently interconnected. Within the context of synaptic complex, both of the actuated enzymes mediate cleavage of phosphodiester bonds. First cleavage generates a phosphotyrosyl-linked recombinase-DNA complex at the recombinase binding sequence and free 5′-hydroxyl end at the first base of the central region. Following the cleavage, the exposed bases with 5′-hydroxyl ends of the central region of dif and attP sites melt from their complementary strands and react with the recombinase-DNA phosphotyrosyl linkage of their recombining partner. Subsequent ligation between dif and attP strands requires complementary base pair interactions at the site of phosphodiester bond formation. Integration mechanism is mostly influenced by the compatibility of dif and attP sequences. dif sites are highly conserved across bacterial phyla. Different phage genomes have different attP sequences; therefore they rely on different mechanisms for integration. Here, I review our current understanding of integration mechanisms used by the vibriophages.
Introduction
Bacterial pathogens evolve continuously to adapt to the changing environment by adopting multiple mobile genetic elements into their compact, modularly organized mosaic genomes to help combat the environmental factors that are detrimental to their subsistence (Frost et al., 2005; Mercier et al., 2008; Hassan et al., 2010). Vibrio cholerae, the noxious enteric pathogen with extraordinary fitness competence resides in multiple niches across continents. A variety of the vibrio strains have acquired multiple genetic traits in their genomes (Heidelberg et al., 2000; Sack et al., 2004; Chun et al., 2009), with the purpose of contributing to the toxin production (Waldor and Mekalanos, 1996), intestinal colonization (Rhine and Taylor, 1994), disease development (Herrington et al., 1988), antimicrobial resistance (Mazel and Davies, 1998; Beaber et al., 2004), cell division (Hassan et al., 2010), and survival in aquatic as well as gut environments (Davies et al., 2012). In V. cholerae, most of the horizontally acquired genetic traits integrate site-specifically within a short region of sequence identity shared by the host chromosome and the integrative mobile genetic elements (IMGEs), using self- or host-encoded tyrosine recombinases (Huber and Waldor, 2002; Rajanna et al., 2003; Hazen et al., 2010; Das et al., 2013; Banerjee et al., 2014). Integration might be reversible or irreversible, depending upon the structures of pre- and post-integrative attachment sequences (Das et al., 2011a, 2013, 2014). Tyrosine recombinases can bind double stranded as well as folded single stranded DNA of the acquired exogenous genetic elements (Val et al., 2005; Mazel, 2006; Das et al., 2011b). Different acquired genetic traits reported in V. cholerae are heterogeneous, and they recognize different receptors for infection (Herrington et al., 1988; Campos et al., 2010; Das et al., 2013), follow different mechanisms to deliver DNA into the host cytoplasm (Heilpern and Waldor, 2000), have a wide range of genomic content (Heidelberg et al., 2000; Faruque and Mekalanos, 2003), and most importantly rely on catalytic activities of different recombinases for chromosomal integration (Rajanna et al., 2003; Das et al., 2010, 2011b; Midonet et al., in press). In a broader sense, these IMGEs can be classified into two different groups, depending upon the presence and absence of recombinases, the enzymes essential for their integration to the host genome. IMGEs, which encode and rely on their own recombinases for integration, are generally large in size and integrates specifically in front of tRNA or tmRNA operon (Karaolis et al., 1999; Heidelberg et al., 2000). On the other hand, IMGEs exploit host-encoded tyrosine recombinases (IMEX), are small in size and integrate at the dimer resolution sites (dif) of host chromosomes (Das et al., 2013, 2014).
The best-characterized IMEX, which exploits the host encoded tyrosine recombinases for its lysogenic conversion is CTXΦ, a temperate filamentous bacteriophage that encodes the cholera toxin in V. cholerae (Waldor and Mekalanos, 1996). The CTXΦ recognizes the toxin co-regulated pilus (TCP), a type IV pilus encoded by the vibrio pathogenic island–I (VPI-1), and subsequently introduces its ~ 7.0 × 103 nucleotides long circular single stranded genomic DNA (+ssDNA) into the V. cholerae cytoplasm. Once in the bacterial cytoplasm, ssDNA may convert to dsDNA, and start the rolling circle replication, or it can be recognized by the chromosomally-encoded tyrosine recombinases, that enable CTXΦ integration into the dif site of host chromosome. Tandemly integrated CTXΦ can also initiate the rolling circle replication from the chromosome and produce extrachromosomal ssDNA genome that may contribute in virion production (Davis and Waldor, 2000). Compared to other episomally replicative filamentous phages, the number of phage particles produced by the CTX-prophage is very low, even upon induction, resulting in one phage particle produced per 10–100 host cells (Davis et al., 2002). The other two well-characterized IMEXs that use the same recombinases for integration at the dif1 site of V. cholerae are VGJΦ and TLCΦ (Campos et al., 2003b; Hassan et al., 2010; Das et al., 2011b). Presence of single or multiple copies of VGJΦ and TLCΦ were reported in toxigenic V. cholerae isolates. Unlike CTXΦ, the virion production of VGJΦ, and TLCΦ mainly relies on the episomal replication. Although the genomes of the two latter elements do not encode any toxins, they are nevertheless implicated in the host fitness and play important role in acquisition of the cholera-toxin-encoding genes and CTXΦ dissemination (Hassan et al., 2010; Das et al., 2011b; Midonet et al., in press).
In this review, I will update the mechanistic insights into filamentous vibriophage integration and cooperative interactions amongst IMGEs, which support the emergence of new pathogenic strains by contributing efficient acquisition and rapid dissemination of the cholera toxin genes in closely or distantly related bacterial strains.
Xer Recombination System in Vibrio cholerae
The native function of Xer recombination system is to ensure the stable maintenance of monomeric circular replicon (Blakely et al., 1993). In V. cholerae, the principal components of the Xer recombination system are (i) two recombinases, XerC and XerD; (ii) a DNA motor protein, FtsK; (iii) a 28-bp DNA sequence, dif (Val et al., 2008) (Table 1). The Xer proteins are members of the tyrosine recombinase family (Esposito and Scocca, 1997). Members of this family share two conserved motifs, containing four highly conserved residues (R1-H-R2-Y), a catalytic tyrosine, which acts as the nucleophile during cleavage of phosphodiester bond, two arginines and a histidine, which have been implicated both the cleavage and rejoining of DNA strands (Esposito and Scocca, 1997). The two positively charged arginines are thought to stabilize the pentavalent phosphate transition state, whereas the histidine may act as a general base catalyst (Esposito and Scocca, 1997; Sherratt and Wigley, 1998).
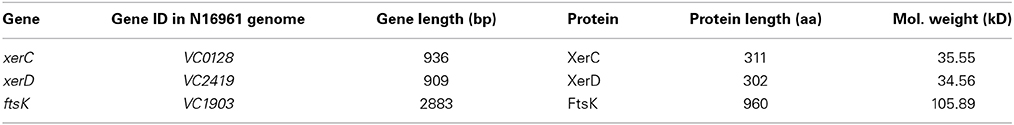
Table 1. Key components of the Xer recombination system in V. cholerae.
During chromosome dimer resolution at the end of DNA replication, tetrameric XerC and XerD recombinase complex catalyzes successive strand exchanges between the two dif sites of a dimeric chromosome. The resolution is completed by a two-step transesterification reaction in which the OH- group of catalytic tyrosine residue of each recombinase is directly involved in the phosphodiester bond formation (Figure 1). In the first step of dimer resolution reaction, cleavage of a phosphodiester bond in DNA is introduced by the active enzyme, generating a covalently linked enzyme-DNA complex at the recombinase binding site and a free 5′-hydroxyl group at the end of the central region. Following the cleavage, the exposed sequence at the 5′-hydroxyl ends of the central region melt from their complementary strands and react with their recombinase-DNA phosphotyrosyl linked recombining partner. Subsequent joining between dif and attP strands requires complementary base pair interactions at the site of the phosphodiester bond formation (MacDonald et al., 2006; Das et al., 2010).
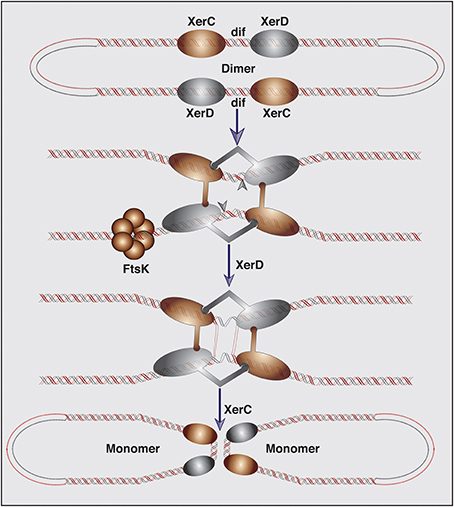
Figure 1. Schematic representation of the XerC-XerD mediated chromosome dimer resolution. Chromosome dimers are resolved by the XerC-XerD-mediated recombination at the dif site. DNA motor protein FtsK brings two dif sites together and induces the XerD catalytic activity required for the first pair of strand exchanges. Followed isomerization, XerC mediates the second pair of strand exchanges, and completes the recombination reaction.
FtsK, a bifunctional protein essential for cell division and chromosome partitioning, induces the XerD function during chromosome dimer resolution (Recchia et al., 1999; Aussel et al., 2002; Demarre et al., 2013). The N-terminal part of FtsK forms a transmembrane domain and is directly linked to the forming septum. The cytoplasmic C-terminal domain is involved in the inter- and intracellular DNA transfer and activation of XerD recombinases (Bigot et al., 2006). Recent reports demonstrated that several IMGEs exploit the conserved Xer recombination system of bacteria to mediate their integration in the dimer resolution site of host chromosomes (Das et al., 2013). The integration mechanisms of such elements were studied in details in V. cholerae and revealed that the IMEXs present in the dif region of V. cholerae make use of three distinct integration mechanisms for lysogenic conversion (Das et al., 2010, 2011a,b, 2013; Midonet et al., in press).
Lysogenic Filamentous Vibriophages Exploit Xer Recombinases
Most of the characterized filamentous vibriophages are lysogenic and integrate at the dif1 and/or dif2 sites of V. cholerae. All reported filamentous vibriophages are equipped with an autonomously replicating genetic module, with or without toxin-encoding genes (Figure 2). Some of the filamentous vibriophages are directly responsible for V. cholerae pathogenesis, others contribute to toxin acquisition and host fitness (Waldor and Mekalanos, 1996; Hassan et al., 2010). Their genomes may or may not be equipped with genes required for virion production and can exploit the virion structural and assembly genes of other filamentous phages for genome packaging (Hassan et al., 2010). Although most of the filamentous vibriophages use host-encoded Xer machinery for their integration, their attachment sites (attP), and their genomic organization are fairly distinct (Figure 2). The best-studied CTXΦ genome is organized into two structurally and functionally distinct modules called repeat sequence 2 (RS2) and core (Figure 2). The RS2 module carries genetic traits essential for CTXΦ replication, maintenance of the ssDNA genome, and transcriptional regulation from phage originated PRSTA promoter (Waldor et al., 1997). The replicating genome of CTXΦ is detrimental to V. cholerae growth, but is tolerated upon integration into the dif sites of either one or both chromosomes (Das et al., 2010; Faruque and Mekalanos, 2012). Compared to toxigenic strains, that contain a single or multiple integrated CTXΦ genomes, V. cholerae cells carrying a replicative form of CTXΦ grow slowly and rapidly lose the replicative phage both under the standard laboratory growth conditions and in rabbit gastrointestinal tract animal model (Faruque et al., 2001).
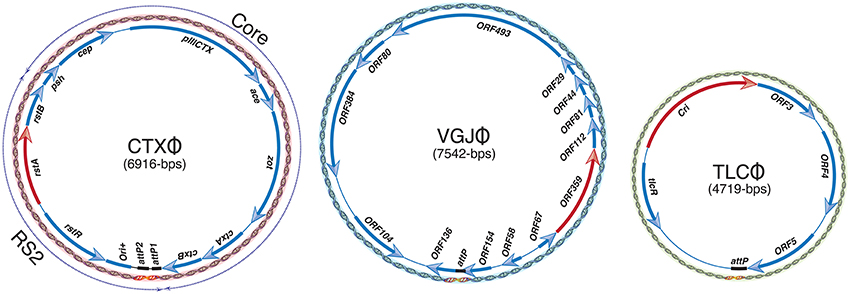
Figure 2. Genomic organization of CTXΦ, VGJΦ, and TLCΦ phages. A red arrow depicts gene essential for replication in each genome. A solid black line denotes attachment sequence, recognized by the Xer recombinases. Replicative form (RF) of each phage genome is presented as dsDNA. Arrows indicate the direction of transcription of each ORF. A thin solid line illustrates intergenic regions. Dashed arrows above the CTXΦ genome represent the core and RS2 modules.
VGJΦ, a filamentous (+)ssDNA lysogenic bacteriophage, infects both clinical and environmental isolates of cholera pathogen and exploits the host-encoded Xer recombinases to integrate at the dif1 site in the large chromosome of V. cholerae (Campos et al., 2003b, 2010; Das et al., 2011b). In contrast to complex attachment site of CTXΦ that contains two dif-like attP sites in an inverted orientation, the 7.5-kb genome of VGJΦ harbors a single dif-like 29-bp attachment sequence, called attPVGJ and integrates specifically into the dif1 site.
TLCΦ, a satellite temperate phage with an autonomous replication module but lacking the morphogenesis and structural virion-encoding genes, is often present in the dif1 region of toxigenic V. cholerae isolates (Rubin et al., 1998). Recent reports demonstrated that, like CTXΦ, and VGJΦ, TLCΦ also relies on the host-encoded Xer machinery for establishing lysogeny (Hassan et al., 2010; Midonet et al., in press). Interestingly, although both the XerC and XerD recombinases are essential for integration of IMEXs, none of the reported IMEXs rely on FtsK for recombination.
Different vibriophages have different attP structures and therefore rely on to a certain extent of different mechanisms of interaction with the Xer recombinases and integration (Das et al., 2013).
Mechanistic Insights into Lysogenic Conversion of Vibriophages
Most lysogenic filamentous vibriophages are IMEXs that depend on the host-encoded Xer recombinases for their lysogeny. The attachment site in the phage genome mimics the native chromosomal Xer binding sites and exploits Xer recombinases to catalyze site-specific recombination and enable phage integration. Based on the attP structure and integration mechanisms, vibriophages can be categorized into three classes (Table 2). Although the constituents of the integration reaction are very similar, integration is achieved by three distinct mechanisms, where catalytic activity of recombinases, order of strand exchanges and the resolution of reaction intermediates into end products are different (Figures 3–5). In this review, I will concentrate on CTXΦ, VGJΦ, and TLCΦ lysogenic conversions to describe the most updated integration mechanisms of these phages studied in V. cholerae and related bacterial cells.
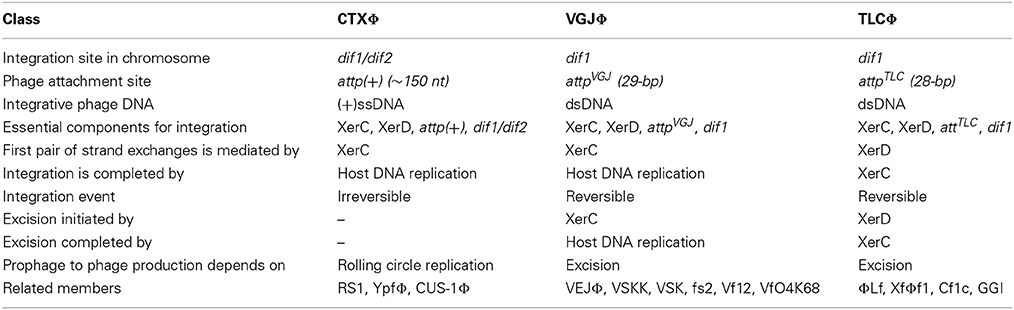
Table 2. Different components and strategies used by three different filamentous Vibriophages for their integration at dif sites of V. cholerae chromosome.
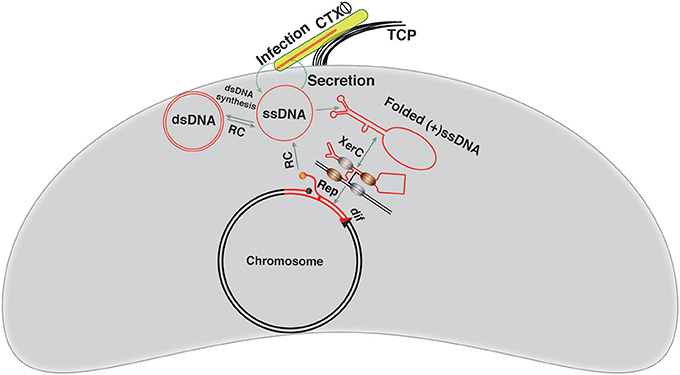
Figure 3. Visual depiction of the CTXΦ integration and replication. The CTXΦ virion recognizes the cognate V. cholerae cell surface receptor TCP and delivers its ssDNA genome into the V. cholerae cytoplasm. The ssDNA genome of CTXΦ may be converted into dsDNA or directly integrated into a chromosomal dif site using the host-encoded XerC-XerD recombinases. XerC mediates first pair of strand exchanges. The host DNA replication, possibly, resolves the resulting Holiday junction. The red line depicts the phage genome, while a black line shows bacterial genome. Newly generated dif site is represented by a triangle.
Mechanistic Insights into CTXΦ Integration
CTX-prophages are ubiquitously present in the chromosomes of all epidemics V. cholerae isolates (Mekalanos et al., 1983; Mekalanos, 1985; Vanden Broeck et al., 2007). Phage to prophage conversion is rapid and efficient process achieved through site-specific recombination between attP(+) of CTXΦ and dif sites of V. cholerae (Val et al., 2005) that results in a stable integration. The functional attP(+) structure is formed by intra-strand base pairing interactions between two inversely oriented attP sequences located between the ctxB and rstR genes in the circular CTXΦ genome (Figure 2). In a replicating form (RF) of the genome, attP1 consists of XerC and XerD binding sites separated by a 12 bp central region. The attP2 is consists of 11-bp XerC and XerD binding sites and a 5–7 bp central region. Neither attP1 nor attP2 are suitable for Xer-mediated integration due to extended central region or presence of incompatible bases in the central region, respectively (Das et al., 2010). The specificity and compatibility of CTXΦ integration is solely determined by the homology of bases next to the XerC-binding site between the overlapping (or corresponding) regions of attP and dif sequence (Das et al., 2010). A 90-bp DNA fragment separates the attP1 and attP2 of CTXΦ. The CTXΦ integration sites in V. cholerae chromosomes (dif1 and dif2) each consist of 28 bp DNA sequence. In dif1, a 6 bp central region, at the border of which the strand exchange occurs, separates the 11 bp binding sites of XerC and XerD. The chromosomally-encoded Xer recombinases can recognize both the double stranded chromosomal dif site and folded single stranded attP(+) of CTXΦ and catalyze recombination to facilitate the phage ssDNA integration.
The CTXΦ ssDNA integration takes place within a DNA-protein complex, where DNA strands are cleaved and rejoined sequentially by XerC recombinase through transient nucleoprotein covalent intermediates. The catalytic tyrosine residue of Xer recombinases functions as a nucleophile and mediates the cleavage of a phosphodiester bond between the last base of the recombinase-binding site and the first base of central region. The key components within the nucleoprotein complex are XerC, XerD, attP(+), and dif sequences. Although the presence of both recombinases are essential for recombination, successful integration of the CTXΦ ssDNA needs only the catalytic activity of XerC (Val et al., 2005). At the onset of recombination reaction, XerC introduces a cut of the phosphodiester bond between the last base of XerC binding site and the first base of the central region, of dif1/dif2, and attP(+) (Figure 3). On one side, a XerC-DNA complex is formed through a covalent phosphotyrosyl bond. On the other side, the DNA strand with a free 5′-OH end melts away from the central region of the reaction complex, migrates to the central region of the partner strand and forms complementary base pairing interactions to stabilize the exchanged strands and form of a phosphodiester bond between the deoxyriboses of two adjacent nucleotides. This step is the determining factor for phage integration specificity (Das et al., 2010). The resulting strand exchanges generate a Holiday Junction (HJ) intermediate. Host DNA replication, possibly, resolves the resulting, transient HJ intermediate and accomplishes the CTXΦ ssDNA integration into the V. cholerae chromosome (Figure 3). The absence of any homology between central region bases at the XerD-side of attP(+) and dif1/dif2 impedes any XerD mediated strand exchange. A recent report revealed that the host-encoded DNA repair protein, endonuclease III, a product of nth gene with N-glycosylase and AP-lyase functions, facilitates the CTXΦ genome integration, possibly by stabilizing the transient HJ intermediate (Bischerour et al., 2012). Each individual step in the CTXΦ genome integration pathway is in principle reversible, but the prophage excision has never been detected under the standard laboratory conditions. This is due to the loss of a functional folded attP(+) structure in the double-stranded form of prophage (Val et al., 2005), in which the intra- strand base pairing between attP1 and attP2 is excluded. In the prophage genome, both attP sequences retained the XerC and XerD binding sites, but the central region of both sites, where the strand exchange occurs during integration, are incompatible with the Xer reaction and make the CTXΦ integration irreversible (Das et al., 2010).
Mechanistic Insights into VGJΦ Integration
VGJΦ, a filamentous (+)ssDNA lysogenic bacteriophage, infects both clinical and environmental isolates of cholera pathogen and exploits the host-encoded Xer recombinases to integrate at the dif1site in the large chromosome of cholera pathogen (Campos et al., 2003b; Das et al., 2011b). In contrast to CTXΦ, the 7.5 kb genome of VGJΦ harbors a single dif like 29-bp DNA sequence, called attPVGJ (Figure 2). Presence of the single functional dif like site allows integration of VGJΦ genome into the V. cholerae chromosome as dsDNA. Like CTXΦ, integration of VGJΦ also relies on the catalytic activity of XerC (Figure 4). Due to an absence of the sequence homology in the central region adjacent to the XerD-binding site between the attPVGJ and dif1 site of V. cholerae, XerD catalytic activities are not been used either to generate HJ or to resolve the HJ during integration. The XerC generated HJ junction is resolved either by the host DNA replication or other DNA repair proteins and assist VGJΦ integration (Figure 4). After integration, two compatible Xer recombination sites (attL and attR) flank the VGJ-prophage. The 7.5-kb distance between the two Xer binding sites does not impede assembly of the recombination complex and excision of the VGJΦ prophage. Like integration, excision of VGJΦ also depends on the XerC-mediated strand exchanges (Figure 4). In vitro recombination reactions between synthetic attPVGJ and dif1 substrates confirmed that only XerC and XerD recombinases are sufficient for DNA rearrangement (Das et al., 2011b).
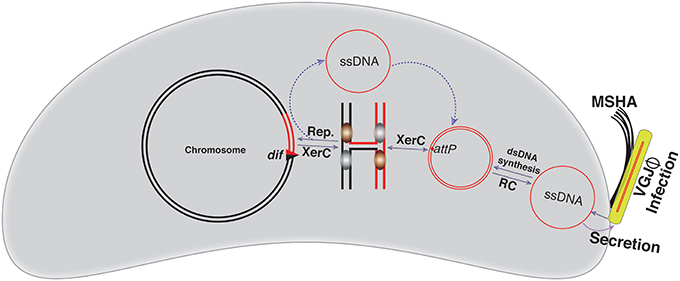
Figure 4. Schematic representation of the VGJΦ integration and replication. VGJΦ uses mannose-sensitive hemagglutinin A (MSHA) to as a primary host receptor and delivers its ssDNA genome into the host cytoplasm. The ssDNA genome of VGJΦ is first converted into dsDNA and for this it contains a dif like DNA sequence, attPVGJ. XerC mediates the first pair of strand exchanges and the host DNA replication resolves the resulting Holiday junction. The VGJΦ integration is reversible. Excision follows the same sequence of strand exchanges as described for integration. The red line depicts the phage genome while a black line depicts bacterial genome. Newly generated dif site is represented by a triangle.
Mechanistic Insights into TLCΦ Integration
TLCΦ, a satellite temperate phage with an autonomous replication module, is mostly present in the dif1 region of toxigenic V. cholerae isolates. Recent reports demonstrated that like CTXΦ, and VGJΦ, TLCΦ also relies on host encoded Xer machinery for its lysogeny (Hassan et al., 2010; Midonet et al., in press). In sharp contrast to the first two elements, the XerD binding site of TLCΦ is degenerated (Das et al., 2013; Midonet et al., in press). The XerC binding site and central region of attPTLC are almost identical to dif1 of V. cholerae, but XerD binding sequence has very little similarity (Midonet et al., in press). Despite the absence of bona fide XerD binding site in attPTLC, TLCΦ integration is strictly related to XerD catalytic activity (Midonet et al., in press). The integration mechanism of TLCΦ is unique compared to the integration strategy adopted by CTXΦ, and VGJΦ (Table 2).
Like CTXΦ and VGJΦ, TLCΦ integration also needs both XerC and XerD recombinases. During TLCΦ integration, XerC and XerD form a hetero-tetrameric complex with dif1 and attPTLC, within which XerD exchanges first pair of DNA strands and form the HJ. The resulting HJ proceed to DNA isomerization and is subsequently resolved by the XerC-mediated second strand exchanges (Figure 5). Tyrosine recombinases-mediated reactions are reversible, and therefore the TLCΦ integration is also reversible. Like integration excision is also depends on XerD catalytic activity (Figure 5). The action of both recombinases and the sequence of strand-exchanges stages in both integration and excision are very similar to chromosome dimer resolution, however the TLCΦ integration occurs without direct participation of DNA motor protein FtsK. Since the recombination reaction between dif1 and attPTLC is not reconstituted in defined in vitro reactions, it is yet not clear whether only the 28-bp attPTLC is sufficient for Xer reaction or whether the TLCΦ needs extended attPTLC region and support from additional host proteins for efficient integration. Similarly, the mechanistic insights into TLCΦ excision in defined reaction conditions are yet to be explored.
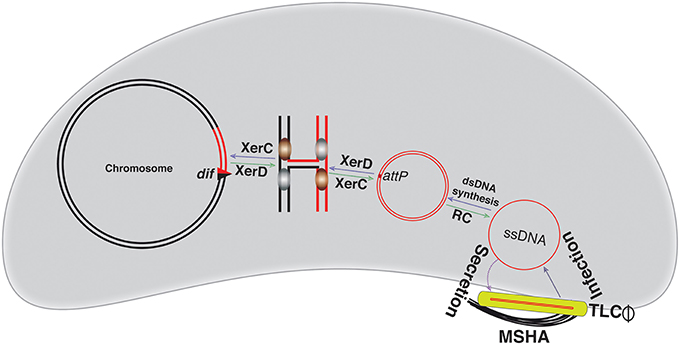
Figure 5. Illustration of the TLCΦ integration and replication. The TLCΦ satellite prophage uses morphogenesis proteins of other filamentous phages, like fs2 for assembly. It uses the mannose-sensitive hemagglutinin A (MSHA) as the primary host receptor and delivers its ssDNA genome into host cytoplasm. The ssDNA genome of the TLCΦ is first converted into dsDNA. The attPTLC in the dsDNA replicative genome of TLCΦ is recognized by the XerC-XerD recombinases. XerD mediates the first pair of strand exchanges and generates a Holiday junction. After isomerization, XerC mediates second pair of strand exchanges and enables the TLCΦ integration. TLCΦ integration, which is reversible. Excision follows the same sequence of strand exchanges as described for integration. The blue and green arrows indicate integration and excision pathways, respectively. The red line depicts phage genome, while a black line represents bacterial genome. Newly generated dif site is symbolized by a triangle.
Cooperative Interactions Among Filamentous Vibriophages
Metagenomic studies revealed several IMEXs integrated at one or both chromosomes of cholera pathogenic strains (Chun et al., 2009). The co-occurrence of multiple IMEXs in the genome of cholera pathogens is not simply a coincidence. Recent reports demonstrated that there are remarkable cooperative interactions between closely or distantly located IMEXs in cholera pathogen (Taylor et al., 1986; Hassan et al., 2010; Das et al., 2011b). Interactions between IMEXs happen at different levels, including the host recognition, receptor binding and DNA entry into the cytoplasm, chromosomal integration, and virion production (Waldor and Mekalanos, 1996; Hassan et al., 2010). The best example of this interaction is acquisition of cholera toxin. The first step of CTXΦ infection in V. cholerae is to recognize its host receptor. TCP, the cell surface receptor of V. cholerae for CTXΦ, is encoded by VPI-1, a ~41-kb mobile genomic island. The same genomic island also helps cholera toxin production by providing transcriptional inducer ToxT, which specifically binds to the “TATTA” repeat, invariably present upstream of the ctxAB operon.
Cooperative interactions between VGJΦ and CTXΦ have been reported very recently (Campos et al., 2003a; Das et al., 2011b). CTXΦ integration is irreversible and phage production depends on the presence of tandem copies of prophages or related RS1 elements (Waldor et al., 1997; Val et al., 2005). A recent report demonstrated that VGJΦ could help CTXΦ excision and its dissemination to the V. cholerae strains devoid of TCP island. When the CTXΦ prophage is inserted between the attachment site of VGJΦ and the dif1 of V. cholerae, a hybrid VGJΦ-CTXΦ is often detected (Das et al., 2011b). The hybrid phage genome is packed within the VGJΦ coat proteins and infects V. cholerae cells expressing mannose-sensitive hemagglutinin (MSHA) pilus, a type IV pilus present on the cell surface whose structural pilin subunit is encoded by the mshA gene. MSHA pilus is ubiquitous and constitutively expressed in all V. cholerae serotypes and thus, host range of the hybrid phage containing the CTXΦ genome encapsulated in the VGJΦ-encoded virion is not restricted to the cells that expressed TCP (Fullner and Mekalanos, 1999). Similarly, TLCΦ also helps the CTXΦ excision if both elements are present in tandem in the V. cholerae chromosome (Midonet et al., in press). Since the TLC prophage is present in most toxigenic strains where a single or multiple copies of the CTXΦ genome are integrated at the chromosomal dif1 site, the TLCΦ is more significant for the CTXΦ excision than is the VGJΦ-prophage. Other than TLCΦ and VGJΦ elements, several other IMGEs could help CTXΦ replication as well as its interaction with the V. cholerae in natural environment by providing cell surface receptor. They can also help increase the amount of cholera toxin production by up-regulating ctxAB operon, protein required for the hybrid virion production and dissemination in the environment.
Conclusions
In this review, several fundamental questions related to IMEXs integration have been addressed, such as contribution of these elements to continuous evolution of the cholera pathogenic strains. Although hypothesis for the adaptive advantage of lysogenic conversion of filamentous vibriophages was proposed, many important questions are still open and need to be addressed in order to improve understanding of the V. cholerae biology and cholera management (Box 1). Pangenomic studies revealed that IMEXs are not limited only to clinical isolates but are also widely distributed in the environmental V. cholerae isolates that are closely or distantly related to pathogenic strains. Much emphasis has been put on the biology of those IMEXs that are mostly present in the clinical isolates. Our knowledge on IMEXs present in the environmental isolates is limited. It is widely accepted that the environmental V. cholerae strains are ubiquitously distributed in aquatic environment and could serve as a reservoir of toxin-encoding genes as well as other fitness factors for clinical strains. Comprehensive studies of IMEXs present both in clinical and environmental isolates of V. cholerae and related pathogenic strains are required in order to understand the microevolution of the species pertinent to epidemiology. On the other hand, better mechanistic and structural knowledge of IMEXs will help to develop therapeutic agents and limit the emergence of the new cholera pathogenic strains and other strains that may pose a serious threat to human beings as well as animals.
Box 1. Outstanding questions.
❖ How is the CTXΦ integration efficiency modulated? Does TLCΦ modulate efficiency of CTXΦ integration into the V. cholerae chromosomes?
❖ How is the CTXΦ virion produced from toxigenic V. cholerae cells, in the case when it harbors only a single copy of the prophage in either chromosome?
❖ Does the TLCΦ contribute to the CTXΦ rolling circle replication?
❖ Is there any relation between the LexA regulon and the CTXΦ integration? Does ssDNA of CTXΦ induced the SOS response in the host cell?
❖ How does the TLCΦ genome integrate in the chromosome of classical V. cholerae strains, where both chromosomes contained an attPTLC incompatible dif2 sequence?
❖ Are there any accessory proteins other than Xer recombinases that may help the TLCΦ genome integration into the V. cholerae chromosome?
❖ What are the host factors implicated in the CTXΦ, VGJΦ, and TLCΦ replication?
Conflict of Interest Statement
The Guest Associate Editor, Jasna Rakonjac, declares that, despite having collaborated on the same research topic as author Bhabatosh Das, the review process was handled objectively and no conflict of interest exists.
Acknowledgments
I thank Dr. G.B. Nair and other members of CHME-THSTI for invaluable suggestions and help during the preparation of this review. The work is supported by the Department of Science and Technology (Grant No. SB/FT/LS-309/2012), Government of India (GOI) and the Department of Biotechnology (Grant No. BT/MB/THSTI/HMC-SFC/2011), GOI.
References
Aussel, L., Barre, F. X., Aroyo, M., Stasiak, A., Stasiak, A. Z., and Sherratt, D. (2002). FtsK Is a DNA motor protein that activates chromosome dimer resolution by switching the catalytic state of the XerC and XerD recombinases. Cell 108, 195–205. doi: 10.1016/S0092-8674(02)00624-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Banerjee, R., Das, B., Balakrish Nair, G., and Basak, S. (2014). Dynamics in genome evolution of Vibrio cholerae. Infect. Genet. Evol. 23C, 32–41. doi: 10.1016/j.meegid.2014.01.006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Beaber, J. W., Hochhut, B., and Waldor, M. K. (2004). SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature 427, 72–74. doi: 10.1038/nature02241
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bigot, S., Saleh, O. A., Cornet, F., Allemand, J. F., and Barre, F. X. (2006). Oriented loading of FtsK on KOPS. Nat. Struct. Mol. Biol. 13, 1026–1028. doi: 10.1038/nsmb1159
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bischerour, J., Spangenberg, C., and Barre, F. X. (2012). Holliday junction affinity of the base excision repair factor Endo III contributes to cholera toxin phage integration. EMBO J. 31, 3757–3767. doi: 10.1038/emboj.2012.219
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Blakely, G., May, G., McCulloch, R., Arciszewska, L. K., Burke, M., Lovett, S. T., et al. (1993). Two related recombinases are required for site-specific recombination at dif and cer in E. coli K12. Cell 75, 351–361. doi: 10.1016/0092-8674(93)80076-Q
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Campos, J., Martinez, E., Izquierdo, Y., and Fando, R. (2010). VEJ{phi}, a novel filamentous phage of Vibrio cholerae able to transduce the cholera toxin genes. Microbiology 156, 108–115. doi: 10.1099/mic.0.032235-0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Campos, J., Martinez, E., Marrero, K., Silva, Y., Rodriguez, B. L., Suzarte, E., et al. (2003a). Novel type of specialized transduction for CTX or its satellite phage RS1 mediated by filamentous phage VGJ in Vibrio cholerae. J. Bacteriol. 185, 7231–7240. doi: 10.1128/JB.185.24.7231-7240.2003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Campos, J., Martinez, E., Suzarte, E., Rodriguez, B. L., Marrero, K., Silva, Y., et al. (2003b). VGJ, a novel filamentous phage of Vibrio cholerae, integrates into the same chromosomal site as CTX. J. Bacteriol. 185, 5685–5696. doi: 10.1128/JB.185.19.5685-5696.2003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chun, J., Grim, C. J., Hasan, N. A., Lee, J. H., Choi, S. Y., Haley, B. J., et al. (2009). Comparative genomics reveals mechanism for short-term and long-term clonal transitions in pandemic Vibrio cholerae. Proc. Natl. Acad. Sci. U.S.A. 106, 15442–15447. doi: 10.1073/pnas.0907787106
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Das, B., Bischerour, J., and Barre, F. X. (2011a). Molecular mechanism of acquisition of the cholera toxin genes. Indian J. Med. Res. 133, 195–200.
Das, B., Bischerour, J., and Barre, F. X. (2011b). VGJphi integration and excision mechanisms contribute to the genetic diversity of Vibrio cholerae epidemic strains. Proc. Natl. Acad. Sci. U.S.A. 108, 2516–2521. doi: 10.1073/pnas.1017061108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Das, B., Bischerour, J., Val, M. E., and Barre, F. X. (2010). Molecular keys of the tropism of integration of the cholera toxin phage. Proc. Natl. Acad. Sci. U.S.A. 107, 4377–4382. doi: 10.1073/pnas.0910212107
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Das, B., Kumari, R., Pant, A., Sen Gupta, S., Saxena, S., Mehta, O., et al. (2014). A novel broad range CTXPhi derived stable integrative expression vector for functional studies. J. Bacteriol. 196, 4071–4080. doi: 10.1128/JB.01966-14
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Das, B., Martinez, E., Midonet, C., and Barre, F. X. (2013). Integrative mobile elements exploiting Xer recombination. Trends Microbiol. 21, 23–30. doi: 10.1016/j.tim.2012.10.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Davies, B. W., Bogard, R. W., Young, T. S., and Mekalanos, J. J. (2012). Coordinated regulation of accessory genetic elements produces cyclic di-nucleotides for V. cholerae virulence. Cell 149, 358–370. doi: 10.1016/j.cell.2012.01.053
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Davis, B. M., Kimsey, H. H., Kane, A. V., and Waldor, M. K. (2002). A satellite phage-encoded antirepressor induces repressor aggregation and cholera toxin gene transfer. EMBO J. 21, 4240–4249. doi: 10.1093/emboj/cdf427
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Davis, B. M., and Waldor, M. K. (2000). CTXphi contains a hybrid genome derived from tandemly integrated elements. Proc. Natl. Acad. Sci. U.S.A. 97, 8572–8577. doi: 10.1073/pnas.140109997
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Demarre, G., Galli, E., and Barre, F. X. (2013). The FtsK family of DNA pumps. Adv. Exp. Med. Biol. 767, 245–262. doi: 10.1007/978-1-4614-5037-5_12
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Esposito, D., and Scocca, J. J. (1997). The integrase family of tyrosine recombinases: evolution of a conserved active site domain. Nucleic Acids Res. 25, 3605–3614. doi: 10.1093/nar/25.18.3605
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Faruque, S. M., and Mekalanos, J. J. (2003). Pathogenicity islands and phages in Vibrio cholerae evolution. Trends Microbiol. 11, 505–510. doi: 10.1016/j.tim.2003.09.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Faruque, S. M., and Mekalanos, J. J. (2012). Phage-bacterial interactions in the evolution of toxigenic Vibrio cholerae. Virulence 3, 556–565. doi: 10.4161/viru.22351
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Faruque, S. M., Rahman, M. M., Hasan, A. K., Nair, G. B., Mekalanos, J. J., and Sack, D. A. (2001). Diminished diarrheal response to Vibrio cholerae strains carrying the replicative form of the CTX(Phi) genome instead of CTX(Phi) lysogens in adult rabbits. Infect. Immun. 69, 6084–6090. doi: 10.1128/IAI.69.10.6084-6090.2001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Frost, L. S., Leplae, R., Summers, A. O., and Toussaint, A. (2005). Mobile genetic elements: the agents of open source evolution. Nat. Rev. Microbiol. 3, 722–732. doi: 10.1038/nrmicro1235
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fullner, K. J., and Mekalanos, J. J. (1999). Genetic characterization of a new type IV-A pilus gene cluster found in both classical and El Tor biotypes of Vibrio cholerae. Infect. Immun. 67, 1393–1404.
Hassan, F., Kamruzzaman, M., Mekalanos, J. J., and Faruque, S. M. (2010). Satellite phage TLCphi enables toxigenic conversion by CTX phage through dif site alteration. Nature 467, 982–985. doi: 10.1038/nature09469
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hazen, T. H., Pan, L., Gu, J. D., and Sobecky, P. A. (2010). The contribution of mobile genetic elements to the evolution and ecology of Vibrios. FEMS Microbiol. Ecol. 74, 485–499. doi: 10.1111/j.1574-6941.2010.00937.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Heidelberg, J. F., Eisen, J. A., Nelson, W. C., Clayton, R. A., Gwinn, M. L., Dodson, R. J., et al. (2000). DNA sequence of both chromosomes of the cholera pathogen Vibrio cholerae. Nature 406, 477–483. doi: 10.1038/35020000
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Heilpern, A. J., and Waldor, M. K. (2000). CTXphi infection of Vibrio cholerae requires the tolQRA gene products. J. Bacteriol. 182, 1739–1747. doi: 10.1128/JB.182.6.1739-1747.2000
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Herrington, D. A., Hall, R. H., Losonsky, G., Mekalanos, J. J., Taylor, R. K., and Levine, M. M. (1988). Toxin, toxin-coregulated pili, and the toxR regulon are essential for Vibrio cholerae pathogenesis in humans. J. Exp. Med. 168, 1487–1492. doi: 10.1084/jem.168.4.1487
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Huber, K. E., and Waldor, M. K. (2002). Filamentous phage integration requires the host recombinases XerC and XerD. Nature 417, 656–659. doi: 10.1038/nature00782
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Karaolis, D. K., Somara, S., Maneval, D. R. Jr., Johnson, J. A., and Kaper, J. B. (1999). A bacteriophage encoding a pathogenicity island, a type-IV pilus and a phage receptor in cholera bacteria. Nature 399, 375–379. doi: 10.1038/20715
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
MacDonald, D., Demarre, G., Bouvier, M., Mazel, D., and Gopaul, D. N. (2006). Structural basis for broad DNA-specificity in integron recombination. Nature 440, 1157–1162. doi: 10.1038/nature04643
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mazel, D. (2006). Integrons: agents of bacterial evolution. Nat. Rev. Microbiol. 4, 608–620. doi: 10.1038/nrmicro1462
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mazel, D., and Davies, J. (1998). Antibiotic resistance. The big picture. Adv. Exp. Med. Biol. 456, 1–6. doi: 10.1007/978-1-4615-4897-3_1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mekalanos, J. J. (1985). Cholera toxin: genetic analysis, regulation, and role in pathogenesis. Curr. Top. Microbiol. Immunol. 118, 97–118. doi: 10.1007/978-3-642-70586-1_6
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mekalanos, J. J., Swartz, D. J., Pearson, G. D., Harford, N., Groyne, F., and de Wilde, M. (1983). Cholera toxin genes: nucleotide sequence, deletion analysis and vaccine development. Nature 306, 551–557. doi: 10.1038/306551a0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mercier, R., Petit, M. A., Schbath, S., Robin, S., El Karoui, M., Boccard, F., et al. (2008). The MatP/matS site-specific system organizes the terminus region of the E. coli chromosome into a macrodomain. Cell 135, 475–485. doi: 10.1016/j.cell.2008.08.031
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Midonet, C., Das, B., Paly, E., and Barre, F. X. (in press). XerD initiates the integration of TLCφ into the Vibrio cholerae genome independently of FtsK. Proc. Natl. Acad. Sci. U.S.A. doi: 10.1073/pnas.1404047111
Rajanna, C., Wang, J., Zhang, D., Xu, Z., Ali, A., Hou, Y. M., et al. (2003). The Vibrio pathogenicity island of epidemic Vibrio cholerae forms precise extrachromosomal circular excision products. J. Bacteriol. 185, 6893–6901. doi: 10.1128/JB.185.23.6893-6901.2003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Recchia, G. D., Aroyo, M., Wolf, D., Blakely, G., and Sherratt, D. J. (1999). FtsK-dependent and -independent pathways of Xer site-specific recombination. EMBO J. 18, 5724–5734. doi: 10.1093/emboj/18.20.5724
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rhine, J. A., and Taylor, R. K. (1994). TcpA pilin sequences and colonization requirements for O1 and O139 vibrio cholerae. Mol. Microbiol. 13, 1013–1020. doi: 10.1111/j.1365-2958.1994.tb00492.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rubin, E. J., Lin, W., Mekalanos, J. J., and Waldor, M. K. (1998). Replication and integration of a Vibrio cholerae cryptic plasmid linked to the CTX prophage. Mol. Microbiol. 28, 1247–1254. doi: 10.1046/j.1365-2958.1998.00889.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sack, D. A., Sack, R. B., Nair, G. B., and Siddique, A. K. (2004). Cholera. Lancet 363, 223–233. doi: 10.1016/S0140-6736(03)15328-7
Sherratt, D. J., and Wigley, D. B. (1998). Conserved themes but novel activities in recombinases and topoisomerases. Cell 93, 149–152. doi: 10.1016/S0092-8674(00)81566-4
Taylor, R. K., Miller, V. L., Furlong, D. B., and Mekalanos, J. J. (1986). Identification of a pilus colonization factor that is coordinately regulated with cholera toxin. Ann. Sclavo Collana Monogr. 3, 51–61.
Val, M. E., Bouvier, M., Campos, J., Sherratt, D., Cornet, F., Mazel, D., et al. (2005). The single-stranded genome of phage CTX is the form used for integration into the genome of Vibrio cholerae. Mol. Cell 19, 559–566. doi: 10.1016/j.molcel.2005.07.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Val, M. E., Kennedy, S. P., El Karoui, M., Bonne, L., Chevalier, F., and Barre, F. X. (2008). FtsK-dependent dimer resolution on multiple chromosomes in the pathogen Vibrio cholerae. PLoS Genet. 4:e1000201. doi: 10.1371/journal.pgen.1000201
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Vanden Broeck, D., Horvath, C., and De Wolf, M. J. (2007). Vibrio cholerae: cholera toxin. Int. J. Biochem. Cell Biol. 39, 1771–1775. doi: 10.1016/j.biocel.2007.07.005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Waldor, M. K., and Mekalanos, J. J. (1996). Lysogenic conversion by a filamentous phage encoding cholera toxin. Science 272, 1910–1914. doi: 10.1126/science.272.5270.1910
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Waldor, M. K., Rubin, E. J., Pearson, G. D., Kimsey, H., and Mekalanos, J. J. (1997). Regulation, replication, and integration functions of the Vibrio cholerae CTXphi are encoded by region RS2. Mol. Microbiol. 24, 917–926. doi: 10.1046/j.1365-2958.1997.3911758.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: CTXΦ, VGJΦ, TLCΦ, XerC, XerD, dif, attP
Citation: Das B (2014) Mechanistic insights into filamentous phage integration in Vibrio cholerae. Front. Microbiol. 5:650. doi: 10.3389/fmicb.2014.00650
Received: 02 August 2014; Accepted: 10 November 2014;
Published online: 28 November 2014.
Edited by:
Jasna Rakonjac, Massey University, New ZealandReviewed by:
Dahlene N. Fusco, Massachusetts General Hospital, USAChristophe Possoz, Centre National de la Recherche Scientifique, France
Copyright © 2014 Das. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bhabatosh Das, Centre for Human Microbial Ecology, Translational Health Science and Technology Institute, 496, Phase III, Udyog Vihar, Gurgaon 122 016, Haryana, India e-mail:YmhhYmF0b3NoQHRoc3RpLnJlcy5pbg==