 Juan Wang
Juan Wang Roger Stephan
Roger Stephan Katrin Zurfluh
Katrin Zurfluh Herbert Hächler
Herbert Hächler Séamus Fanning
Séamus Fanning- 1UCD Centre for Food Safety, School of Public Health, Physiotherapy and Population Science, University College Dublin, Dublin, Ireland
- 2Vetsuisse Faculty, Institute for Food Safety and Hygiene, University of Zurich, Zürich, Switzerland
- 3School of Biological Sciences, Institute for Global Food Security, Queen's University Belfast, Belfast, UK
Objectives: Previously 14 conjugative plasmids from multi-drug resistant (MDR) Escherichia coli from healthy humans and food-producing animals in Switzerland were sequenced. The aim of this study was to extend the genetic characterization of these plasmids with a focus on blaESBL genes including blaCTX-M-1 and blaTEM, class 1 integrons and toxin-antitoxin (TA) systems contained therein.
Methods: The nucleotide sequences and subsequent annotation therein of 14 conjugative plasmids were previously determined from their corresponding transconjugants. The TA loci were confirmed by RASTA-Bacteria.
Results: Eight of the conjugative plasmids identified were found to encode genes expressing ESBLs. Structural heterogeneity was noted in the regions flanking both the blaCTX-M-1 and blaTEM genes. The blaCTX-M-1 genes were associated with the common insertion sequences ISEcp1 and IS26, and uniquely with an IS5 element in one case; while blaTEM genes were found to be associated with IS26 and Tn2. A new blaTEM-210 gene was identified. Seven class 1 integrons were also identified and assigned into 3 groups, denoted as In54, In369 and In501. Sixteen TA loci belonging to 4 of the TA gene families (relBE, vapBC, ccd and mazEF) were identified on 11 of these plasmids.
Conclusions: Comparative sequence analysis of these plasmids provided data on the structures likely to contribute to sequence diversity associated with these accessory genes, including IS26, ISEcp1 and Tn2. All of them contribute to the dissemination of the corresponding resistance genes located on the different plasmids. There appears to be no association between β-lactam encoding genes and TA systems.
Introduction
Understanding the molecular epidemiology of antimicrobial drug resistance genes has been a complex task due to the plasmids' role in the spread of these elements (Carattoli et al., 2006; Hopkins et al., 2006). Moreover, the diversity and promiscuity of resistance genes has become a major global public health issue since antimicrobial resistance threatens the effective prevention and treatment of an ever-increasing number of infections in a clinical setting. Plasmid-located genes can be acquired from several sources and disseminated by horizontal gene transfer (HGT). The latter strategy contributes to the dissemination of many of the undesirable phenotypes associated with bacteria, including antibiotic resistance, virulence, and resistance to heavy metals (Davies and Davies, 2010; Baquero et al., 2013). Transmissible plasmids can be considered as particularly successful entities within the communal gene pool, and those that encode a full set of conjugation-encoding genes are referred to as conjugative, facilitating their dissemination over large taxonomic distances (Norman et al., 2009). Of note, there is a need to standardize surveillance methods with an emphasis on gene tracking by plasmid sequencing as an aid to reveal the transmission of resistance among bacteria from animals to humans vice versa.
Escherichia coli producing extended-spectrum β-lactamases (ESBLs) is a major problem in worldwide, since these ESBL genes can be spread by plasmid-mediated integrons, insertion sequence (IS) elements, and transposons among different bacteria species causing outbreaks as well as sporadic infections (Liebana et al., 2013). Plasmids expressing an ESBL phenotype frequently carry genes encoding resistance to other commonly used antimicrobial drug classes and genes for toxin-antitoxin (TA) systems, which contribute to the maintenance of plasmids in their host. Thus, understanding the roles of these genetic elements and the surrounding genetic structures are essential and relevant to the subsequent development of strategies to limit the dissemination and persistence of ESBL genes and others.
In a recent publication (Wang et al., 2014), we reported the complete nucleotide sequences of 14 large (>30 kb) conjugative plasmids identified in nine multi-drug resistant (MDR) E. coli isolates expressing an ESBL phenotype isolated from food-producing animals and healthy humans in Switzerland. A comparative analysis of these plasmid-backbone structures was carried out previously (Wang et al., 2014). The aim of the present study was to analyse, in detail, the accessory genes from these plasmids for (i) the genetic context in which blaCTX−M−1 and blaTEM genes were located, (ii) the structure of class 1 integrons and their gene cassettes, (iii) the identification of other related antimicrobial resistance genes and (iv) the identification of the TA systems.
Materials and Methods
Bacterial Isolates, Susceptibility Testing and Genetic Annotation
Nine ESBL-producing E. coli isolated from fecal samples of food-producing animals and healthy humans in Switzerland from 2009 to 2011 (Geser et al., 2012a,b), were identified previously. All of the associated technical protocols were described in earlier publications (Wang et al., 2013, 2014). The antimicrobial resistance profiles for these isolates are shown in a supplementary table (Table S1).
Bioinformatics Search for TA Pairs
Putative TA systems were identified in the 14 conjugative plasmids using the RASTA-Bacteria program version 2.12 (http://genoweb1.irisa.fr/duals/RASTA-Bacteria/). A summary of the TA features associated with the plasmids is shown in Table 1.
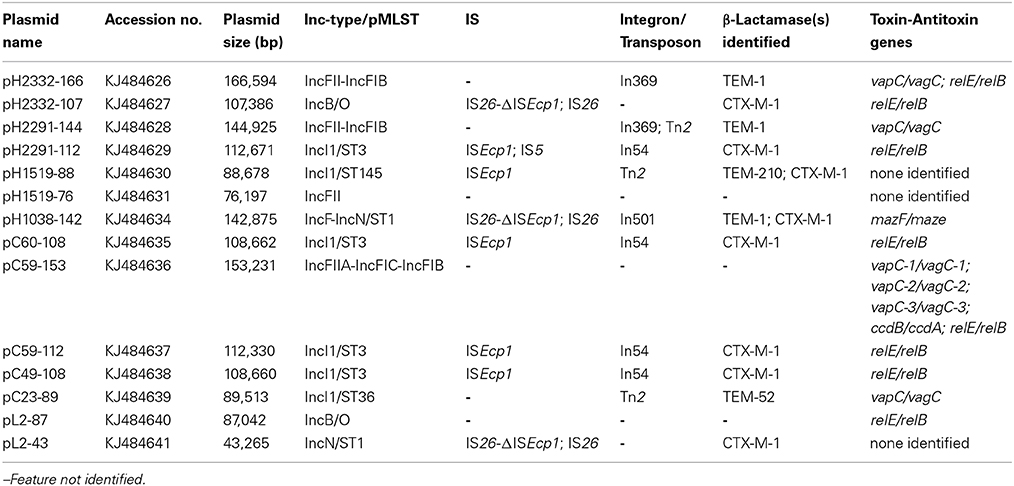
Table 1. Summary of the TA features associated with all 14 conjugative sequenced plasmids from healthy human and food-producing animal origin previously reported in Switzerland.
Reference Numbers for Class 1 Integrons
Standard methods were used to annotate the class 1 integrons by INTEGRALL platform version 1.2 (http://integrall.bio.ua.pt/), referred as In501, In369 or In54 (Figure 3).
Results and Discussion
Comparison of the Genetic Environment of blaCTX−M−1 Genes
Eight blaCTX−M−1 genes were found to be located within various genetic contexts in the present study. Either a complete or partial ISEcp1 gene was identified proximal to the resistance gene (Figure 1). A complete ISEcp1 gene was more often identified (Figure 1A), along with a partial ORF (denoted as Δorf477) and a Δmrx gene located distally in this arrangement. In three of the eight loci studied, a novel genetic arrangement was also noted wherein a truncated copy of ISEcp1 gene was identified (denoted as ΔISEcp1) and which proximally flanked the blaCTX−M−1 genes with an IS26 element being located further upstream (Figure 1B).
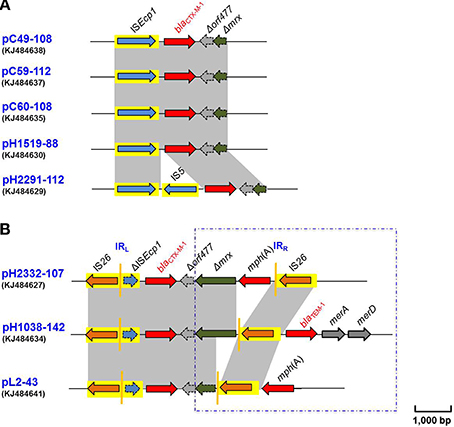
Figure 1. (A) Comparative schematic representation of the flanking regions of the blaCTX-M-1 genes in plasmids pC49-108, pC59-112, pC60-108, pH1519-88, and pH2291-112. Areas shades in gray indicate homologies in the corresponding genetic environment on each plasmid. The ORFs are shown as arrows, with the arrowhead indicating the direction of transcription. ISEcp1 elements are shown as arrowed boxes. (B) Comparative schematic representation of the flanking regions of the blaCTX-M-1 genes in plasmids pH2332-107, pH1038-142, and pL2-43. Areas shades in gray indicate homologies in the corresponding genetic environment on each plasmid. The ORFs are shown as arrows, with the arrowhead indicating the direction of transcription. IS elements are shown as arrowed boxes.
Further consideration of the eight blaCTX−M−1 containing plasmids suggested that they could be divided into two groups based on their different disrupted insertion sequence elements. The first group consisted of five of the eight plasmids, including pC49-108, pC59-112, pC60-108, pH1519-88, and pH2291-112 (Figure 1A and Table 1). All 5 plasmids were typed as IncI1 and showed a high degree of similarity (99% at the nucleotide level) with the reference IncI1 plasmid R64 (Table 1) (Wang et al., 2014). With the exception of pH2291-112, a tnpA gene of 1263 bp was found to be located upstream of the four blaCTX−M−1 genes (Figure 1A). This was typical of the genetic organization previously identified in blaCTX−M−1 containing plasmids, and the former DNA sequence was determined to be 100% identical to ISEcp1 (accession no.: AJ242809) in all cases. ISEcp1 was originally identified proximal to the plasmid-mediated cephalosporinase gene blaCMY−4 in an E. coli isolate identified in the United Kingdom (accession no.: AJ242809). This unusual IS element was also commonly found to be associated with several blaCTX−M genes (Figure S1) (Karim et al., 2001; Saladin et al., 2002; Poirel et al., 2003; Lartigue et al., 2004). ISEcp1 elements are known to play a dual role, acting both as a transposase and ensuring the expression of the downstream ORF via a strong putative promoter (Poirel et al., 2003, 2005; Hossain et al., 2004; Jacoby, 2009). In the case of pH2291-112, unusually the locus between the latter transposon and the β-lactamase resistance gene was interrupted by an IS5 element (Figure 1A).
Analysis of the DNA downstream of the blaCTX−M−1 genes showed a varied gene organization. In the CTX-M-1 cluster, a partial sequence of 342 bp denoted as Δorf477 was present in all structures studied and followed by a partially deleted Δmrx gene of 110 bp. This organization (ISEcp1- blaCTX−M−1-Δorf477-Δmrx) was determined to be identical to the corresponding region found in other IncI1 plasmid pCTX1261 (accession no.: HF549089) and also in an IncF plasmid pCTX2412 (accession no.: HF549091) originally purified from E. coli cultured from swine and horse samples in Germany (Schink et al., 2013). The data reported here further confirmed the importance of ISEcp1 that is thought to function in the mobilization of blaCTX−M genes among IncI1 type plasmids.
The insertion sequence element IS5 identified upstream of the blaCTX−M−1 gene in pH2291-112 (Figure 1A) has only been reported on one other plasmid previously. The ISEcp1-IS5 arrangement was originally identified on a Klebsiella pneumoniae plasmid p9701 (accession no.: FM246881) and this was associated with the mobilization of blaCMY−2 gene (Figure S1B) (Verdet et al., 2009). Moreover, this arrangement was recently reported in the genomic DNA of E. coli EC958, which was associated with one blaCMY−23 gene (Figure S1B) (Totsika et al., 2011).
The second group was unique in that a truncated copy of the ISEcp1 transposon (denoted as ΔISEcp1) was identified and found to be located between the blaCTX−M−1 and an IS26. This unusual arrangement was identified in the remaining three plasmids: pH2332-107 (IncB/O), pH1038-142 (IncF-IncN), and pL2-43 (IncN) (Figure 1B). In all cases, the truncated ΔISEcp1 was 214 bp in length. All of these sequences contained the putative promoter region involved in the transcription of blaCTX−M genes. When the region proximal to the blaCTX−M−1 gene was considered, it showed some unusual features. In the case of pH2332-107, the distal region of this gene consisted of the terminal 342 bp of orf477, followed by a partially deleted mrx gene (Δmrx), an entire mph(A) gene and a second copy of IS26 (Figure 1B). The gene module IS26-ΔISEcp1-blaCTX−M−1−Δmrx-mph(A)-IS26 was recently identified in plasmids purified from E. coli, including those from the successful clone ST131 and from clinical E. coli isolated from pigs in Germany (Cullik et al., 2010). In particular, this structure had been identified in several IncN and IncI1 plasmids (Schink et al., 2011; Dolejska et al., 2013). Since the identical structure of the blaCTX−M−1 region can be found in plasmids of different incompatibility groups, it has been hypothesized that the complete structure can be exchanged en-bloc between different plasmid backbones as a composite transposon mediated by an IS26 transposition event (Cullik et al., 2010; Dolejska et al., 2013).
In contrast to the genetic arrangement above, in pH1038-142, the locus containing the mph(A) gene was deleted and extended beyond the distal IS26 with a region containing another gene, blaTEM−1 in addition to the merA and merD genes, encoding resistance to mercury (Figure 1B). Based on our extensive searching of the current databases, this is the first description of such an arrangement. Further examination of the blaCTX−M−1 region in pL2-43 revealed other novel structural rearrangements. As shown in Figure 1B, similar to the situation described above for pH1038-142, the distal IS26 element, with intact left and right inverted repeats, was further extended with a copy of mph(A) followed by a hypothetical ORF (not shown in Figure 1B). These findings suggest that perhaps IS26 may play a hitherto unrecognized but important role in the mobilization of these ESBL genes.
Comparison of the Genetic Environment of blaTEM Regions
Five blaTEM genes were identified in this study. These were found to be located within three different genetic environments (Figure 2). In two of these (pH1519-88 and pH1038-142) the blaTEM gene was located on the same plasmid containing a blaCTX−M−1 gene. A complex transposon Tn2 was identified on three of these plasmids, pC23-89 (IncI1) containing blaTEM−52, pH1519-88 (IncI1) containing a new variant blaTEM−210 and pH2291-144 (IncFII-IncFIB) containing blaTEM−1 (Figure 2A). In this case, the β-lactamase-encoding resistance gene was located distal to the tnpR-encoding gene. In the remaining two cases, the blaTEM−1 gene was flanked by two copies of an IS26 element as shown for plasmid pH2332-166 (IncFII-IncFIB) and a proximally located IS26 element in pH1038-142 (IncF-IncN), followed by merA-merD genes (Figure 2B).
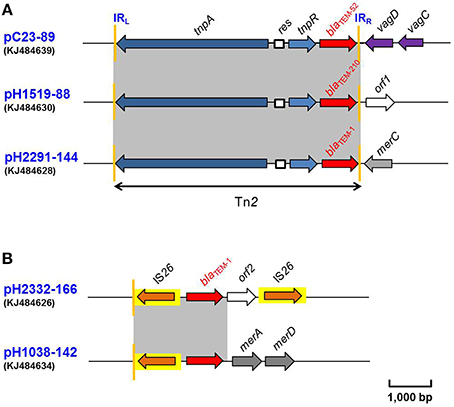
Figure 2. (A,B) Schematic presentation of the flanking regions of the blaTEM genes in plasmids pC23-89, pH1519-88, pH2291-144, pH2332-166, and pH1038-142. Areas shades in gray indicate homologies in the corresponding genetic region on each plasmid. The ORFs are shown as arrows, with the arrowhead indicating the direction of transcription. IS elements are shown as arrowed boxes.
The single blaTEM−52-carrying plasmid, pC23-89 was purified from an E. coli isolate of poultry origin and belonged to replicon type IncI1. Sequence analysis of the immediate flanking regions identified tnpA and tnpR genes of transposon Tn2 located upstream of the blaTEM−52 gene. The right inverted repeat (denoted as IRR in Figure 2A) of Tn2 was followed by the virulence-associated vagD and vagC genes (Figure 2A). These alleles showed 100% nucleotide identity with another two IncI1 plasmids, which also harbored blaTEM−52 genes. One (accession no.: EF141186) was identified in a Salmonella Infantis, isolated in Belgium in 2005 (Cloeckaert et al., 2007); and the other was located on plasmid pESBL-117 (accession no.: CP008734) from an ESBL-producing E. coli isolate recovered in Netherlands in 2014 (Brouwer et al., 2014). Recently, this blaTEM−52 gene had been reported within a partially-sequenced Tn2 transposon derivative on an IncI1 plasmid, pCTX909 (accession no.: HF549095) from a poultry source in Germany (Schink et al., 2013). [Bailey and coworkers (Bailey et al., 2011) reported that transposon Tn2 is annotated incorrectly as Tn3 transposon in the respective database entry].
Highly transmissible IncI1 plasmids carrying the blaTEM−52 are of particular interest as they are globally spread among E. coli populations from humans and animals (Carattoli, 2009). IncI1 plasmids carrying blaTEM−52 within transposon Tn2, similar to that observed for pC23-89 (Figure 2A), have been described in Salmonella enterica and E. coli isolates from poultry and humans in Belgium, Germany and the Netherlands, respectively, (Cloeckaert et al., 2007; Leverstein-Van Hall et al., 2011) and these appear to be widely distributed among different members of the Enterobacteriaceae family. Furthermore, a range of additional TEM variants (including TEM-20, −52, and −126) have similarly been detected in food-producing animals and from isolates cultured from foods in different European countries (De Champs et al., 2004; Sunde et al., 2009). Moreover, in relation to the ESBLs of the TEM class, the most frequently detected throughout the EU was reported to be TEM-52 [EFSA Panel on Biological Hazards (BIOHAZ), 2011].
Based on the sequencing data from pH1519-88, a novel TEM variant was identified and classified as blaTEM−210 (Figure 2A). Plasmid pH1519-88 also belonged to the IncI1 group. Sequence analysis of blaTEM−210 revealed two mutations which gave rise to two amino acid substitutions at positions corresponding to residues 49 and 69 when compared with a previously published sequence (accession no.: AF309824). The blaTEM−210 gene had also been identified within a Tn2 transposon (Figure 2A).
The Tn2-blaTEM−1 region of 4770 bp in length located on pH2291-144 exhibited 100% DNA sequence identity to that found in several plasmids in E. coli, including the conjugative plasmid pVQS1 (accession no.: JQ609357). The latter carries a qnrS1 gene associated with an IncN group and was originally identified in a Salmonella Virchow isolate, which was cultured from a patient in Switzerland returning from a foreign holiday (Karczmarczyk et al., 2012). It is interesting to note that, whereas pH2291-144 and pH2332-166 showed 99% nucleotide identity across their backbones, the blaTEM−1 genes contained on these plasmids were associated with different transposons (Figures 2A,B). The blaTEM−1 gene of pH2291-144 was located within Tn2 and inserted within a mer module (Figure 2A), whereas in pH2332-166, the blaTEM−1 was flanked by two copies of IS26 (Figure 2B).
Several blaTEM genes have been reported to be associated with different transposons, and with Tn2 in particular as the dominant blaTEM-containing transposon in commensal E. coli (Bailey et al., 2011). Although information on the genetic context of the blaTEM genes would be valuable in extending our understanding of how this important resistance gene is disseminated, for most TEM variants little or no sequence beyond the immediate flanking regions of the gene is available (Bailey et al., 2011).
Comparison of Class 1 Integron Regions
Several complete class 1 integron structures were noted among the 14 sequenced conjugative plasmids. Typically a class 1 integron is composed of three key features; a 5′-conserved structure (CS) containing an int1 site-specific recombinase; followed by a variable region containing one or more gene cassettes and then the 3′-CS containing the qacEΔ1 and a sul1-encoding genes. A complete class 1 integron was identified in plasmids pH2332-166 and pH2291-144 (Figure 3), wherein two gene cassettes dfrA1b-aadA1b were identified in the typical head-to-tail arrangement. Furthermore, on pH2332-166, three antimicrobial resistance genes, strB, strA and sul2, that encode resistance to streptomycin and sulphonamide, were identified and found to flank the 5′-CS region of a complete class 1 integron; these latter genes were lacking on pH2291-144 (Figure 3). Sequence analysis of the segment (sul2-strA-strB-intI1-dfrA1b-aadA1b-qacEΔ1-sul1) (Figure 3) on pH2332-166 exhibited 100% similarity to the corresponding sequence located on the plasmid pNRG857c (accession no.: CP001856), which was previously isolated from a clinical isolate of adherent and invasive E. coli in Germany (Nash et al., 2010).
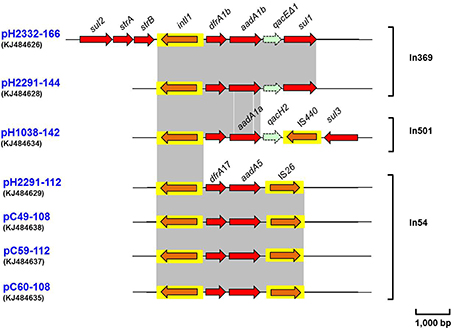
Figure 3. Schematic presentation showing the complete class I integrin along with the flanking regions identified on plasmids pH2332-166, pH2291-144, pH1038-142, pH2291-112, pC49-108, pC59-112, and pC60-108. Areas shaded in gray and the arrangements of the ORFs are as indicated in Figure 2 above. IS elements are shown as arrowed boxes.
Located on pH1038-142 was the gene cassette containing dfrA1b-aadA1a and this was located distally to a 5′-CS which in this case was devoid of a complete 3′-CS, being replaced instead by a single copy of qacH2 and an insertion sequence element, IS440 (Figure 3). Downstream of the latter element was a sul3 gene that encodes resistance to sulphonamides. The cluster of intl1-dfrA1b-aadA1a-qacH2-IS440-sul3 shared 100% identity to the corresponding DNA sequence with plasmid pEC54 (accession no.: FM244709), which was isolated from one sul3-encoding E. coli isolate of porcine origin in Germany.
A similarly structured class 1 integron was also identified from the R64-like IncI1 plasmids, including pH2291-112, pC49-108, pC59-112 and pC60-108; wherein the 3′-CS was replaced by IS26 (Figure 3). The gene cassette array of intl1-dfrA17-aadA5-IS26 (3,470 bp) exhibited 100% nucleotide sequence identity to that of the class 1 integron in Salmonella enteric serovar Indiana located on a non-conjugative plasmid pS414 from Shandong, China, 2008 (accession no.: KC237285) (Lai et al., 2013).
All seven integron sequences identified in this study were deposited in the INTEGRALL database (http://integrall.bio.ua.pt/) and were designated as In369, In54 or In501 (Figure 3).
Other Related Antimicrobial Resistance Genes
Three tetAR modules coding for a class A tetracycline efflux protein and a repressor, were identified on pH2332-166, pH2291-144 and pH1038-142 respectively (Figure 4 and Table 1). The loci (tnpA-pecM-tet(A)-tetR) showed a high degree of sequence similarity to each other, at 99% nucleotide identity. The tet(A) gene on each plasmid was flanked on the distal side by a pecM gene encoding a PecM-like protein, which has been reported previously to be associated with tetAR genes (Pasquali et al., 2005; Szczepanowski et al., 2011). On pH2332-166 and pH2291-144, the tetR genes were located on the distal side of a mercury resistance cassette (Wang et al., 2014). In plasmid pH1038-142 this module was located between a class 1 integron and a strAB module. It has been suggested previously that the tetAR and adjacent genes serve as a hot-spot for insertion of different mobile elements and can be modified by direct-repeat-mediated deletional events (Szczepanowski et al., 2011). It is conceivable that additional resistance modules could be integrated into the tet-platform.

Figure 4. Schematic presentation of the flanking gene regions of the tetAR-modules identified on plasmids pH2332-166, pH2291-144, and pH1038-142. Areas shaded in gray and the arrangements of the ORFs are as indicated in Figure 2 above. IS elements are shown as arrowed boxes.
A sul2-strA-strB gene cluster was noted on pH2332-166 (Figure 3), encoding resistance to sulphonamide and streptomycin, which showed a high degree of similarity with that of plasmid pCERC1 (accession no.: JN012467), at 99% nucleotide identity. The plasmid pCERC1 carrying the dfrA14 cassette in the strA gene of the sul2-strA-strB gene cluster, has been reported in many countries, indicating a global distribution and it appears to have been circulating in Gram-negative bacteria for more than 25 years (Anantham and Hall, 2012).
All of the transconjugant strains were re-screened for antimicrobial resistance genes as previously described (Karczmarczyk et al., 2011). The antimicrobial resistance profiles for the nine strains in this study are shown in a supplementary table (Table S1).
TA Systems Identified on Plasmids Isolated from E. coli in Healthy Food-Producing Animals and Humans
TA loci play a role in bacterial stress physiology and the stabilization of horizontally acquired elements. TA systems are composed of a toxin-encoding gene along with its cognate antitoxin and these features prevent post-segregational killing by the host bacterial cell. In addition, these genetic loci also act to eliminate compatible plasmids, therefore ensuring the maintenance of the plasmid in the bacterial cell during replication (Hayes, 2003; Unterholzner et al., 2013). In adapted E. coli, it has been proposed, previously that strain virulence and the association with IncF plasmids, could contribute to the success and spread of the blaCTX−M−15 genes (Clermont et al., 2008; Mnif et al., 2010). Furthermore, recent experiments reported that ESBL-plasmids carrying TA systems can be cured from field isolates of E. coli using a heat technique, and the cured ESBL-plasmids contained at least one complete TA system, whose loss would normally mean the death of bacterial cells (Schaufler et al., 2013). TA loci-encoded toxins themselves may represent a novel target that could be exploited for the development of future generation of antibacterial compounds (Gupta, 2009).
Using BLASTP and RASTA-Bacteria, we searched all 14 conjugative plasmids and identified four different TA families (Table S2). Six TA loci, denoted as VapC (toxin protein) and VagC (antitoxin/virulence-associated protein), were identified on plasmids pH2332-166, pH2291-144, pC59-153, and pC23-89. The addiction module toxin RelE and antitoxin RelB protein were detected from eight plasmids, pH2332-166, pH2332-107, pH2291-112, pC60-108, pC59-153, pC59-112, pC49-108, and pL2-87, respectively. A representative of the mazEF family was located on the multidrug resistant plasmid pH1038-142. The plasmid maintenance protein, toxin CcdB and antitoxin CcdA protein, were only detected on plasmid pC59-153. A summary of TA systems from the 14 conjugative plasmids and the corresponding protein sequences are shown in Table S2.
A phylogenetic tree containing the RelE/VapC/CcdB/MazF superfamily toxins identified on these plasmids in this study is shown in Figure 5. Four clusters containing these toxin sequences were noted. The RelE superfamily was located on plasmids with different Inc types in this study (Figure 5 and Table 1). In IncI1 plasmids, the RelE sequences were identical, with variations being evident from the clustering when located in plasmids of IncB/O or IncF types (Figure 5). It is also interesting to note that pC59-153 possessed three vapC toxin-encoding genes, which were located in different plasmid genomes. Additionally, the backbone of pC59-153 showed high levels of nucleotide similarity with several ColV plasmids that are known to be associated with an avian pathogenic E. coli strains (Wang et al., 2014). Thus, the presence of multiple vapC toxin-encoding genes and other TA systems in pC59-153 could support plasmid stability and virulence. Figure 5 shows that the six VapC sequences cluster into two different groups according to their amino acid sequence similarities, and all but one belonged to IncF types. While the relationship between the TA systems and the plasmid incompatibility types remains unclear, further study may help in the understanding of the nature of these TA systems in plasmids and their specific physiological role(s) in bacteria. Amino acid sequence similarity and clustering analysis of the antitoxin genes were carried out and these showed similar characteristics to their corresponding toxin genes (data not shown).
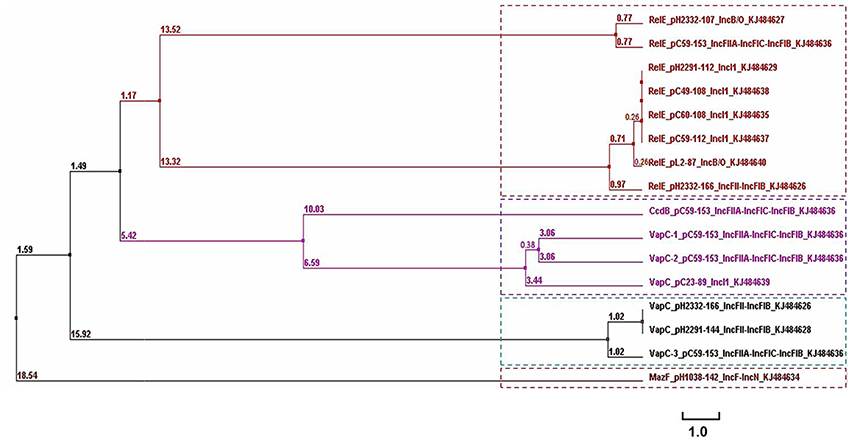
Figure 5. Phylogenetic tree (chladogram) for the amino acid sequences of plasmid-encoded toxin genes from 14 conjugative plasmids (see Table 1). The TA loci sequences were retrieved from Table S2 (raw sequences). The tree was calculated using Clustal Omega version 1.2.1. The lengths of the horizontal lines indicate relative evolutionary distances. A scale bar is also shown.
In many cases, TA loci were clustered or closely linked to mobile genetic elements (Pandey and Gerdes, 2005). In the most extreme of these cases, three vapC/vagC loci from plasmids pH2332-166, pH2291-144, and pC59-153, were found to be located in the traD-traI intergenic region. All relBE homologous loci identified in our study, were bona fide integron elements located between replication proteins and mobile element proteins. It is tempting to speculate that these features suggest that TA loci may represent a type of mobile cassette that can move between plasmids of different Inc types or different original bacterial sources and that undergo rapid evolution and horizontal transfer.
Concluding Remarks
In this study we extended our careful analysis of the accessory genes in 14 conjugative plasmids from nine unrelated human, poultry and lamb E. coli isolates. These plasmids have the capacity to transfer at high frequency to suitable recipient bacterial strains, providing a means by which the blaESBL genes harbored therein can be quickly disseminated. This feature may in turn compromise the use of this class of antibiotic for the treatment of infections in both animals and humans alike. Furthermore it suggests the existence of a reservoir of resistance genes that is highly mobile and contained in more than one genetic context. Our current study reports on some novel arrangements that were identified in the plasmids conferring resistance to β-lactam and other commonly-used antibiotics, and the structural heterogeneity associated with these regions flanking antibiotic resistance genes. Insertion sequences and transposons, such as ISEcp1, IS26, and Tn2, are likely to contribute to the dissemination of the blaCTX−M−1 and blaTEM gene containing regions between different ecosystems (human and animal). Seven class 1 integrons were identified and designated into three classes; and 16 TA loci from 11 of these plasmids were assigned into four groups according to their toxin sequence similarity. There appeared to be no link between the β-lactam resistance-encoding genes and the TA systems in this study. Moreover, the nature of specific addiction systems appears to differ with the type of replicon with no obvious epidemiological links. Nonetheless, these antimicrobial resistance genes and addiction modules are located on efficient conjugative plasmids (i.e., IncI1 and IncF-related), and this could contribute to the emergence and spread of these genes, in different bacterial hosts. Taken together these characteristics provide much of the motivation for extending our understanding of the molecular epidemiology and structural relationships of accessory genes that exist between plasmids from a variety of Inc types and bacterial sources. Further studies will be necessary to understand the driving forces associated with accessory gene cassettes transfer and plasmid spreading.
Funding
This study was supported by internal funding.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://www.frontiersin.org/journal/10.3389/fmicb.2014.00716/abstract
Figure S1 | (A) Comparison of the blaCTX−M−1 gene environment in plasmid pC49-108 with the blaCTX−M−15 gene in plasmid pEC_B24, and the blaCMY−6 gene in plasmid pA/C-LS6-SC29. Areas shades in gray indicate homologies in the corresponding genetic region on each plasmid. The ORFs are shown as arrows, with the arrowhead indicating the direction of transcription. ISEcp1 elements are shown as boxes with an arrow indicating the transcription orientation. (B) Comparison of the blaCTX−M−1 gene environment in plasmid pH2291-112 with the blaCMY−2 gene in plasmid p9701. Areas shades in gray indicate homologies in the corresponding genetic region on each plasmid. The ORFs are shown as arrows, with the arrowhead indicating the direction of transcription. ISEcp1 and IS5 elements are shown as boxes with an arrow indicating their transcription orientation.
References
Anantham, S., and Hall, R. M. (2012). pCERC1, a small, globally disseminated plasmid carrying the dfrA14 cassette in the strA gene of the sul2-strA-strB gene cluster. Microb. Drug Resist. 18, 364–371. doi: 10.1089/mdr.2012.0008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bailey, J. K., Pinyon, J. L., Anantham, S., and Hall, R. M. (2011). Distribution of the blaTEM gene and blaTEM-containing transposons in commensal Escherichia coli. J. Antimicrob. Chemother. 66, 745–751. doi: 10.1093/jac/dkq529
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Baquero, F., Tedim, A. P., and Coque, T. M. (2013). Antibiotic resistance shaping multi-level population biology of bacteria. Front. Microbiol. 4:15. doi: 10.3389/fmicb.2013.00015
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brouwer, M. S., Bossers, A., Harders, F., Van Essen-Zandbergen, A., Mevius, D. J., and Smith, H. E. (2014). Complete genome sequences of IncI1 plasmids carrying extended-spectrum beta-lactamase genes. Genome Announc. 2, e00859–e00814. doi: 10.1128/genomeA.00859-14
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Carattoli, A. (2009). Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 53, 2227–2238. doi: 10.1128/AAC.01707-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Carattoli, A., Miriagou, V., Bertini, A., Loli, A., Colinon, C., Villa, L., et al. (2006). Replicon typing of plasmids encoding resistance to newer beta-lactams. Emerg. Infect. Dis. 12, 1145–1148. doi: 10.3201/eid1207.051555
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Clermont, O., Lavollay, M., Vimont, S., Deschamps, C., Forestier, C., Branger, C., et al. (2008). The CTX-M-15-producing Escherichia coli diffusing clone belongs to a highly virulent B2 phylogenetic subgroup. J. Antimicrob. Chemother. 61, 1024–1028. doi: 10.1093/jac/dkn084
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cloeckaert, A., Praud, K., Doublet, B., Bertini, A., Carattoli, A., Butaye, P., et al. (2007). Dissemination of an extended-spectrum-beta-lactamase blaTEM-52 gene-carrying IncI1 plasmid in various Salmonella enterica serovars isolated from poultry and humans in Belgium and France between 2001 and 2005. Antimicrob. Agents Chemother. 51, 1872–1875. doi: 10.1128/AAC.01514-06
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cullik, A., Pfeifer, Y., Prager, R., Von Baum, H., and Witte, W. (2010). A novel IS26 structure surrounds blaCTX-M genes in different plasmids from German clinical Escherichia coli isolates. J. Med. Microbiol. 59, 580–587. doi: 10.1099/jmm.0.016188-0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Davies, J., and Davies, D. (2010). Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 74, 417–433. doi: 10.1128/MMBR.00016-10
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
De Champs, C., Chanal, C., Sirot, D., Baraduc, R., Romaszko, J. P., Bonnet, R., et al. (2004). Frequency and diversity of Class A extended-spectrum beta-lactamases in hospitals of the Auvergne, France: a 2 year prospective study. J. Antimicrob. Chemother. 54, 634–639. doi: 10.1093/jac/dkh395
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dolejska, M., Villa, L., Hasman, H., Hansen, L., and Carattoli, A. (2013). Characterization of IncN plasmids carrying blaCTX-M-1 and qnr genes in Escherichia coli and Salmonella from animals, the environment and humans. J. Antimicrob. Chemother. 68, 333–339. doi: 10.1093/jac/dks387
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
EFSA Panel on Biological Hazards (BIOHAZ). (2011). Scientific Opinion on the public health risks of bacterial strains producing extended-spectrum β-lactamases and/or AmpC β-lactamases in food and food-producing animals. EFSA J. 9:2322. doi: 10.2903/j.efsa.2011.2322
Geser, N., Stephan, R., and Hachler, H. (2012a). Occurrence and characteristics of extended-spectrum beta-lactamase (ESBL) producing Enterobacteriaceae in food producing animals, minced meat and raw milk. BMC Vet. Res. 8:21. doi: 10.1186/1746-6148-8-21
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Geser, N., Stephan, R., Korczak, B. M., Beutin, L., and Hachler, H. (2012b). Molecular identification of extended-spectrum-beta-lactamase genes from Enterobacteriaceae isolated from healthy human carriers in Switzerland. Antimicrob. Agents Chemother. 56, 1609–1612. doi: 10.1128/AAC.05539-11
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gupta, A. (2009). Killing activity and rescue function of genome-wide toxin-antitoxin loci of Mycobacterium tuberculosis. FEMS Microbiol. Lett. 290, 45–53. doi: 10.1111/j.1574-6968.2008.01400.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hayes, F. (2003). Toxins-antitoxins: plasmid maintenance, programmed cell death, and cell cycle arrest. Science 301, 1496–1499. doi: 10.1126/science.1088157
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hopkins, K. L., Liebana, E., Villa, L., Batchelor, M., Threlfall, E. J., and Carattoli, A. (2006). Replicon typing of plasmids carrying CTX-M or CMY beta-lactamases circulating among Salmonella and Escherichia coli isolates. Antimicrob. Agents Chemother. 50, 3203–3206. doi: 10.1128/AAC.00149-06
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hossain, A., Reisbig, M. D., and Hanson, N. D. (2004). Plasmid-encoded functions compensate for the biological cost of AmpC overexpression in a clinical isolate of Salmonella typhimurium. J. Antimicrob. Chemother. 53, 964–970. doi: 10.1093/jac/dkh240
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jacoby, G. A. (2009). AmpC beta-lactamases. Clin. Microbiol. Rev. 22, 161–182. doi: 10.1128/CMR.00036-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Karczmarczyk, M., Abbott, Y., Walsh, C., Leonard, N., and Fanning, S. (2011). Characterization of multidrug-resistant Escherichia coli Isolates from animals presenting at a university veterinary hospital. Appl. Environ. Microbiol. 77, 7104–7112. doi: 10.1128/Aem.00599-11
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Karczmarczyk, M., Stephan, R., Hachler, H., and Fanning, S. (2012). Complete nucleotide sequence of pVQS1 containing a quinolone resistance determinant from Salmonella enterica serovar Virchow associated with foreign travel. J. Antimicrob. Chemother. 67, 1861–1864. doi: 10.1093/jac/dks158
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Karim, A., Poirel, L., Nagarajan, S., and Nordmann, P. (2001). Plasmid-mediated extended-spectrum beta-lactamase (CTX-M-3 like) from India and gene association with insertion sequence ISEcp1. FEMS Microbiol. Lett. 201, 237–241. doi: 10.1016/S0378-1097(01)00276-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lai, J., Wang, Y., Shen, J., Li, R., Han, J., Foley, S. L., et al. (2013). Unique class 1 integron and multiple resistance genes co-located on IncHI2 plasmid is associated with the emerging multidrug resistance of Salmonella Indiana isolated from chicken in China. Foodborne. Pathog. Dis. 10, 581–588. doi: 10.1089/fpd.2012.1455
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lartigue, M. F., Poirel, L., and Nordmann, P. (2004). Diversity of genetic environment of bla(CTX-M) genes. FEMS Microbiol. Lett. 234, 201–207. doi: 10.1016/j.femsle.2004.01.051
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Leverstein-Van Hall, M. A., Dierikx, C. M., Cohen Stuart, J., Voets, G. M., Van Den Munckhof, M. P., Van Essen-Zandbergen, A., et al. (2011). Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 17, 873–880. doi: 10.1111/j.1469-0691.2011.03497.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Liebana, E., Carattoli, A., Coque, T. M., Hasman, H., Magiorakos, A. P., Mevius, D., et al. (2013). Public health risks of enterobacterial isolates producing extended-spectrum beta-lactamases or AmpC beta-lactamases in food and food-producing animals: an EU perspective of epidemiology, analytical methods, risk factors, and control options. Clin. Infect. Dis. 56, 1030–1037. doi: 10.1093/cid/cis1043
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mnif, B., Vimont, S., Boyd, A., Bourit, E., Picard, B., Branger, C., et al. (2010). Molecular characterization of addiction systems of plasmids encoding extended-spectrum beta-lactamases in Escherichia coli. J. Antimicrob. Chemother. 65, 1599–1603. doi: 10.1093/jac/dkq181
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nash, J. H., Villegas, A., Kropinski, A. M., Aguilar-Valenzuela, R., Konczy, P., Mascarenhas, M., et al. (2010). Genome sequence of adherent-invasive Escherichia coli and comparative genomic analysis with other E. coli pathotypes. BMC Genomics. 11:667. doi: 10.1186/1471-2164-11-667
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Norman, A., Hansen, L. H., and Sorensen, S. J. (2009). Conjugative plasmids: vessels of the communal gene pool. Philos. Trans. R. Soc. Lond. B Biol. Sci. 364, 2275–2289. doi: 10.1098/rstb.2009.0037
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pandey, D. P., and Gerdes, K. (2005). Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic. Acids. Res. 33, 966–976. doi: 10.1093/nar/gki201
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pasquali, F., Kehrenberg, C., Manfreda, G., and Schwarz, S. (2005). Physical linkage of Tn3 and part of Tn1721 in a tetracycline and ampicillin resistance plasmid from Salmonella Typhimurium. J. Antimicrob. Chemother. 55, 562–565. doi: 10.1093/jac/dkh553
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Poirel, L., Decousser, J. W., and Nordmann, P. (2003). Insertion sequence ISEcp1B is involved in expression and mobilization of a bla(CTX-M) beta-lactamase gene. Antimicrob. Agents Chemother. 47, 2938–2945. doi: 10.1128/AAC.47.9.2938-2945.2003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Poirel, L., Lartigue, M. F., Decousser, J. W., and Nordmann, P. (2005). ISEcp1B-mediated transposition of blaCTX-M in Escherichia coli. Antimicrob. Agents Chemother. 49, 447–450. doi: 10.1128/AAC.49.1.447-450.2005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Saladin, M., Cao, V. T. B., Lambert, T., Donay, J. L., Herrmann, J. L., Ould-Hocine, Z., et al. (2002). Diversity of CTX-M beta-lactamases and their promoter regions from Enterobacteriaceae isolated in three Parisian hospitals. FEMS Microbiol. Lett. 209, 161–168. doi: 10.1111/J.1574-6968.2002.tb11126.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schaufler, K., Wieler, L. H., Semmler, T., Ewers, C., and Guenther, S. (2013). ESBL-plasmids carrying toxin-antitoxin systems can be “cured” of wild-type Escherichia coli using a heat technique. Gut. Pathog. 5:34. doi: 10.1186/1757-4749-5-34
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schink, A. K., Kadlec, K., Kaspar, H., Mankertz, J., and Schwarz, S. (2013). Analysis of extended-spectrum-beta-lactamase-producing Escherichia coli isolates collected in the GERM-Vet monitoring programme. J. Antimicrob. Chemother. 68, 1741–1749. doi: 10.1093/jac/dkt123
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schink, A. K., Kadlec, K., and Schwarz, S. (2011). Analysis of bla(CTX-M)-carrying plasmids from Escherichia coli isolates collected in the BfT-GermVet study. Appl. Environ. Microbiol. 77, 7142–7146. doi: 10.1128/AEM.00559-11
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sunde, M., Tharaldsen, H., Slettemeas, J. S., Norstrom, M., Carattoli, A., and Bjorland, J. (2009). Escherichia coli of animal origin in Norway contains a blaTEM-20-carrying plasmid closely related to blaTEM-20 and blaTEM-52 plasmids from other European countries. J. Antimicrob. Chemother. 63, 215–216. doi: 10.1093/jac/dkn445
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Szczepanowski, R., Eikmeyer, F., Harfmann, J., Blom, J., Rogers, L. M., Top, E. M., et al. (2011). Sequencing and comparative analysis of IncP-1alpha antibiotic resistance plasmids reveal a highly conserved backbone and differences within accessory regions. J. Biotechnol. 155, 95–103. doi: 10.1016/j.jbiotec.2010.11.018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Totsika, M., Beatson, S. A., Sarkar, S., Phan, M. D., Petty, N. K., Bachmann, N., et al. (2011). Insights into a multidrug resistant Escherichia coli pathogen of the globally disseminated ST131 lineage: genome analysis and virulence mechanisms. PLoS ONE 6:e26578. doi: 10.1371/journal.pone.0026578
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Unterholzner, S. J., Poppenberger, B., and Rozhon, W. (2013). Toxin-antitoxin systems: biology, identification, and application. Mob. Genet. Elements 3:e26219. doi: 10.4161/mge.26219
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Verdet, C., Gautier, V., Chachaty, E., Ronco, E., Hidri, N., Decre, D., et al. (2009). Genetic context of plasmid-carried blaCMY-2-like genes in Enterobacteriaceae. Antimicrob. Agents Chemother. 53, 4002–4006. doi: 10.1128/AAC.00753-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wang, J., Stephan, R., Karczmarczyk, M., Yan, Q., Hachler, H., and Fanning, S. (2013). Molecular characterization of bla ESBL-harboring conjugative plasmids identified in multi-drug resistant Escherichia coli isolated from food-producing animals and healthy humans. Front. Microbiol. 4:188. doi: 10.3389/fmicb.2013.00188
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wang, J., Stephan, R., Power, K., Yan, Q., Hachler, H., and Fanning, S. (2014). Nucleotide sequences of 16 transmissible plasmids identified in nine multidrug-resistant Escherichia coli isolates expressing an ESBL phenotype isolated from food-producing animals and healthy humans. J. Antimicrob. Chemother. 69, 2658–2668. doi: 10.1093/jac/dku206
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: bla genes, plasmid sequencing, CTX-M, TEM, accessory genes
Citation: Wang J, Stephan R, Zurfluh K, Hächler H and Fanning S (2015) Characterization of the genetic environment of blaESBL genes, integrons and toxin-antitoxin systems identified on large transferrable plasmids in multi-drug resistant Escherichia coli. Front. Microbiol. 5:716. doi: 10.3389/fmicb.2014.00716
Received: 25 September 2014; Accepted: 01 December 2014;
Published online: 06 January 2015.
Edited by:
Fiona Walsh, National University of Ireland Maynooth, IrelandReviewed by:
Lei Dai, Iowa State University, USAFiona Fouhy, Teagasc - The Agriculture and Food Development Authority, Ireland
Copyright © 2015 Wang, Stephan, Zurfluh, Hächler and Fanning. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Séamus Fanning, UCD Centre for Food Safety, School of Public Health, Physiotherapy and Population Science, University College Dublin, Belfield, Dublin 4, Ireland e-mail:c2Zhbm5pbmdAdWNkLmll