 Li Fu
Li Fu Tianze Song2,3
Tianze Song2,3 Yahai Lu
Yahai Lu- 1College of Resources and Environmental Sciences, China Agricultural University, Beijing, China
- 2School of Life Science, Fudan University, Shanghai, China
- 3College of Urban and Environmental Sciences, Peking University, Beijing, China
Zoige wetland in Tibetan plateau represents a cold environment at high altitude where significant methane emission has been observed. However, it remains unknown how the production and emission of CH4 from Zoige wetland will respond to a warming climate. Here we investigated the temperature sensitivity of methanogen community in a Zoige wetland soil under the laboratory incubation conditions. One soil sample was collected and the temperature sensitivity of the methanogenic activity, the structure of methanogen community and the methanogenic pathways were determined. We found that the response of methanogenesis to temperature could be separated into two phases, a high sensitivity in the low temperature range and a modest sensitivity under mesophilic conditions, respectively. The aceticlastic methanogens Methanosarcinaceae were the main methanogens at low temperatures, while hydrogenotrophic Methanobacteriales, Methanomicrobiales, and Methanocellales were more abundant at higher temperatures. The total abundance of mcrA genes increased with temperature indicating that the growth of methanogens was stimulated. The growth of hydrogenotrophic methanogens, however, was faster than aceticlastic ones resulting in the shift of methanogen community. Determination of carbon isotopic signatures indicated that methanogenic pathway was also shifted from mainly aceticlastic methanogenesis to a mixture of hydrogenotrophic and aceticlastic methanogenesis with the increase of temperature. Collectively, the shift of temperature responses of methanogenesis was in accordance with the changes in methanogen composition and methanogenic pathway in this wetland sample. It appears that the aceticlastic methanogenesis dominating at low temperatures is more sensitive than the hydrogenotrophic one at higher temperatures.
Introduction
The temperature sensitivity of soil microbial activity has gained increasing attention in recent decades (Davidson and Janssens, 2006). The global surface temperature is expected to increase by 3.7—4.8°C by the year 2100 (IPCC: Climate Change, 2014). Understanding the response of soil microbial activity is imperative to predict the feedback of global climate change. Low temperature has been a major constraint to the degradation of organic matter in high latitude and high altitude regions, where large amounts of readily degradable organic carbon have been stored (Davidson and Janssens, 2006; Liu et al., 2011). These regions, however, are warming rapidly. Temperature sensitivity of soil microbial activity is critical to the vulnerability of carbon stocks in these areas.
Temperature sensitivity of soil respiration has been estimated recently using meta-analysis and modeling. Based on the published data, Bond-Lamberty and Thomson (2010) obtained the mean temperature dependence of terrestrial respiration (Q10) of 1.5. Temperature sensitivity might be influenced by environmental and biological factors such as microbial community structure and substrate availability. Mahecha et al. (2010) derived the so-called intrinsic temperature dependences, and showed an almost universal Q10 value (1.4 ± 0.1) for ecosystems ranging from croplands to mixed forests and woody savanna. Similarly, Yvon-Durocher et al. (2012) reported that the sensitivity of ecosystem respiration to seasonal changes in temperature was statistically indiscernible across environments from aquatic to terrestrial ecosystems. The reconciled temperature sensitivity was also revealed for methanogenic systems from pure cultures to natural ecosystems (Yvon-Durocher et al., 2014). Thus, the meta-analysis and modeling indicated a remarkable consistency in temperature sensitivity of either terrestrial respiration or methanogenesis. This contrasts strikingly with the huge diversity of microbiota and their metabolisms contained in terrestrial ecosystems (Karhu et al., 2014).
Cold wetlands including boreal and alpine fens are among the most important biogenic sources of atmospheric methane (Conrad, 2009; Kirschke et al., 2013). To understand and predict the production and emission of CH4, numerous studies have been conducted in these areas to determine the spatiotemporal patterns of CH4 fluxes and the structure and function of methanogenic archaeal communities (e.g., Yrjala et al., 2011; Godin et al., 2012; Yang et al., 2014). The key factors controlling methanogenic activity have been identified that include temperature, water level, vegetation, surface topography, substrate availability, soil pH and depth (Galand et al., 2002, 2003, 2005; Høj et al., 2008; Juottonen et al., 2008; Tian et al., 2012a,b). Temperature stands out among the most important factors (Høj et al., 2008; Juottonen et al., 2008). Quantitative characterization of temperature sensitivity of methanogenic activity, however, is rare. Zoige wetland is an open fen located in Tibetan plateau with the average altitude of 3500 m, mean annual temperature of around 1°C and mean annual precipitation of 650 mm; it covers a total area of 6180 km2 (Chen et al., 2009a,b). The vegetation was dominated by Carex muliensis and Eleocharis valleculosa (Cai et al., 1965). Despite the cold climate, it has been estimated that the mean annual emission of CH4 amounts to 0.65–1.0 Tg, accounting for about one third of total emissions from natural wetlands in China (Chen et al., 2013a,b). Similar as in boreal fens and high arctic peats (Høj et al., 2008; Juottonen et al., 2008), temperature has been identified as the major factor controlling the seasonal and annual CH4 emissions from Zoige wetland (Chen et al., 2009b, 2013a). Investigation of methanogen community revealed the prevalence of psychrophilic aceticlastic and methylotrophic methanogens in Zoige wetland (Zhang et al., 2008b). For instance, a methanol-utilizing pure culture isolated from this wetland, Methanolobus psychrophilus, showed the optimum growth at 18°C, being active down to 0°C but stopped growth at 25°C (Zhang et al., 2008a). Apparently, the significant emission of CH4 is related to the existence of cold-adapted methanogens in this wetland. It, however, remains unknown how the methanogen community and methanogenesis in Zoige wetland will respond to a warming climate.
The investigations in rice field soils have shown that the methanogen community shifted from a mixture of aceticlastic and hydrogenotrophic methanogens under mesophilic conditions to the dominance of hydrogenotrophic Methanocellales at above 40°C (Fey and Conrad, 2000; Peng et al., 2008; Rui et al., 2011) or to the dominance of aceticlastic Methanosaetaceae and Methanosarcinaceae when temperature decreases to 15°C (Chin et al., 1999; Conrad et al., 2009). A preliminary study in Zoige wetland also showed the change in composition of methanogens and the pathway of methanogenesis between 15 and 30°C (Zhang et al., 2008a). We hypothesized that a shift in methanogenic community could induce change in temperature sensitivity of CH4 production in Zoige wetland. The purpose of the present study, therefore, was to determine the effects of temperature on methanogen composition and methanogenic activity and to evaluate if the temperature sensitivity of CH4 production in Zoige wetland was related with changes in methanogen community and methanogenic pathway. A laboratory incubation experiment was conducted with a soil sample collected from Zoige wetland. The incubations under different temperatures allowed us to explicitly link the sensitivity of methanogenic activity to the structure of methanogen community and the methanogenic pathways.
Experimental Methods
Soil Sampling and Anaerobic Incubation
The soil sample was collected from an open fen close to the Wetland National Nature Reserve of Zoige located in Qinghai-Tibetan Plateau (33°47′ N, 102°57′ E). The sampling site is covered dominantly by Carex muliensis. The soil is flooded periodically depending on precipitation and meltwater runoff. The microtopography consists of hummocks and hollows. The water table level was about 5 cm below the hollow surface at the time of sampling. Soil samples were collected on 25 July 2012 in about a square meter area at the depth of 5–20 cm below the hollow surface. Vegetation and organic debris was removed by hands during the sampling. About 10 kilograms of wet soil samples were placed in an ice box and transported to the laboratory within 24 h for immediate processing. The soil sample had the following characteristics: pH 7.5, organic C of 152.6 g kg−1, total N of 10.6 g kg−1, and C:N of 14.4. Soil slurries were prepared by mixing soil samples with autoclaved and degassed water. The slurries were passed through 2-mm sieves to homogenize and remove the coarse materials. Thirty grams of soil slurry was filled into 50-ml glass bottles with the final soil (d.w.) to water ratio of 1:3.5. The bottles were closed with butyl stoppers and flushed with N2. Soil slurries were incubated for 81 days at 10°C, 15°C, 20°C, 25°C, 30°C, and 35°C, respectively. Each temperature treatment was carried out in triplicate.
Measurement of Gases and Volatile Fatty Acids
Gas samples (0.1 ml) were taken from headspace with a pressure-lock precision analytical syringe (Baton Rouge, LA, USA). The concentrations of CH4, CO2, and H2 were analyzed using gas chromatographs GC-7890 (Agilent Technologies, USA) equipped with a thermal conductivity detector. The 13C abundance (δ13C) of CH4 and CO2 was analyzed by a gas chromatography-isotope ratio mass spectrometry system (Yuan and Lu, 2009). Liquid samples (0.5 ml) were taken with sterile syringes and centrifuged 15 min at 17,949 × g at 4°C. The supernatant was collected, passed through 0.25-μm-pore-size filters, and stored at −20°C. Acetate and propionate were analyzed with an HPLC-1200 using a Zorbax SB-AQ C18 column (Agilent Technologies, USA).
Temperature sensitivity was calculated according to the Arrhenius equation:
where P is the rate of CH4 production, E is the activation energy (eV), T is the absolute temperature (K), R is the Boltzmann constant (8.623 × 10−5 eV K−1), and M is the theoretical rate of CH4 production in the absence of activation energy. The maximal rate of CH4 production was obtained from the cumulating curve of CH4 partial pressure in the headspace of incubation bottles. To incorporate enzyme concentration into the calculation, the maximal rates of CH4 production were normalized against the total abundances of mcrA that were quantified by real time PCR as described below.
Nucleic Acid Extraction and Purification
The total DNA of soil samples was extracted using the protocol by Ma et al. (2012). Briefly, 2 g of soil slurry was extracted sequentially with TPMS buffer (50 mM Tris-HCl [pH 7.0], 1.7% [wt/vol] polyvinylpyrrolidone K25, 20 mM MgCl2, 1%[wt/vol] sodium dodecyl sulfate) and phenol-based lysis buffer (5 mMTris-HCl [pH 7.0], 5 mM Na2 EDTA, 1% [wt/vol] sodium dodecyl sulfate, 6% [vol/vol] water-saturated phenol). Beads-beating was performed in FastPrep-24 (MP Biomedicals, USA). The supernatants were further extracted with water-saturated phenol, phenol-chloroform-isoamyl alcohol (25:24:1 [vol/vol/vol]), and chloroform-isoamyl alcohol (24:1 [vol/vol]). The extracts were purified by cold ethanol and sodium acetate. The quality and purity of DNA were checked by 1% agarose gel electrophoresis and NanoDrop1000 spectrophotometry (NanoDrop Technologies, Wilmington, DE).
Terminal Restriction Fragment Length Polymorphism (T-RFLP) Analysis
PCR amplification and terminal restriction fragment polymorphism (T-RFLP) analyses of archaeal 16S rRNA gene fragments followed the protocols described in Peng et al. (2008). PCR was carried out using the primer set Ar109f and Ar915r (Lueders et al., 2004). The 5′ end of the Ar915r primer was labeled with 6-carboxyfluorescein. The 50-μl reaction mixture contained 1 μl of DNA template (in 1:100 dilution of original extracts), 5 μl of 10 × buffer, 3 μl of 25 mM MgCl2, 1 μl of a 10 mM concentration of the deoxynucleoside triphosphates, 0.5 μl of each primer (50 μM), and 2.5 U of Tag DNA polymerase (TaKaRa). The thermal profile was as follows: 3 min at 94°C; 32 cycles of 60 s at 94°C, 45 s at 52°C, and 90 s at 72°C; and finally 5 min at 72°C. The PCR product was purified using an agarose gel DNA extraction kit (TaKaRa) and digested at 65°C for 3.5 h by TaqI (Fermentas, Canada). The digestion products were purified with SigmaSpin Post-Reaction Clean-Up Columns (Sigma), and a portion was mixed with deionized formamide and the internal standard GeneScan-1000 LIZ (Applied Biosystems). The mixtures were denatured for 3 min at 95°C, and the DNA fragments were size separated using a 3730 × l Genetic Analyzer (Applied Biosystems). The percent abundance (Ap) of individual terminal restriction fragments (T-RFs) were calculated as the percentage of each peak height in the sum of all peak heights in a given T-RFLP profile and only those T-RFs with A p > 1% were considered in further analyses (Noll et al., 2005).
Cloning, Sequencing, and Phylogenic Analysis
Three clone libraries of the archaeal 16S rRNA genes were constructed from soil slurries incubated 49 days at 15°C, 25°C, and 35°C, respectively. The PCR amplification used the same primers as indicated above without FAM labeling. PCR products were purified and ligated into the pMD19-T vector (TaKaRa) according to the manufacturer's instructions. Plasmids were transformed into E. coli cells, and more than 100 clones were randomly selected from each clone library and sequenced with an ABI 3730 × l sequencer using BigDye Terminator cycle sequencing chemistry (Applied Biosystems) (Peng et al., 2008; Rui et al., 2009). Rarefaction curves (Figure S1) were calculated for three clone libraries according to the method described by Schloss and Handelsman (2005). Coverage of each clone library was calculated according to methods described by Good (1953) using the formula 1 − (n1/N)] × 100 (n1 and N denote the number of sole OTUs and total number of clones in a library, respectively). The coverage of analyzed clones by phylotype-richness estimates were 89, 92, and 81% for 15°C, 25°C, and 35°C treatment respectively, indicating that the clone libraries were adequately examined. Phylogenetic trees were constructed using the neighbor-joining algorithm according to the protocol of Lueders and Friedrich (2000), and bootstrap analysis implemented 1000 replicates.
Quantitative (Real-Time) PCR Analysis
Quantitative PCR of mcrA genes were carried out in a 7500 real-time PCR system (Applied Biosystems) using the primer pair mlas and mcrA-rev (Steinberg and Regan, 2008). Quantitative PCR was performed in a total volume of 25 μl containing 12.5 μl of Power SYBRsGreen PCR Master mix kit (Applied Biosystems), 0.75 μl (10 μM) primers, 200 ng bovine serum albumin and 2 μl of a 100:1 dilution of DNA sample. The thermal cycles and fluorescence signal acquisition followed the protocols described in (Steinberg and Regan, 2008). The DNA standards were prepared from the cloned sequence of mcrA gene, which were further amplified with vector-specific primers. The PCR products were purified with a UNIQ-10 column kit (Sangon Biotech, Shanghai, China). The obtained PCR products were quantified using the PicoGreen dsDNA quantification kit (Invitrogen, Eugene, OR) and then converted into the copy number of DNA molecules per unit volume ranging from 1.0 × 103 to 1.0 × 108 copies μl−1. Three replicates of each measurement were done.
Nucleotide Sequence Accession Numbers
The sequences of the 16S rRNA clones obtained in this study have been deposited in the EMBL nucleotide sequence database under the following accession numbers: KJ644784-KJ645071.
Results
Temperature Sensitivity of Methanogenesis
Production of CH4 showed a lag phase that increased with the decrease of temperature (Figure S2). Emission of CO2 into the headspace, however, occurred immediately in all incubations. Accumulation of CH4 in the headspace increased markedly with the increase of temperature. We calculated the rate of CH4 production at each incubation temperature (Figure S3). The rate reached to maxima around 20 days at 30°C and 35°C, but delayed substantially at lower temperatures. The maximal rates corresponded roughly to substrate availability inferred from the dynamics of acetate and propionate in incubations (Figure S4). Based on maximal rates of CH4 production, we calculated the activation energy of methanogenesis according to Arrhenius equation (Figure 1). It revealed that the temperature sensitivity could be separated into two phases with the first activation energy of 1.07 eV between 20°C and 35°C and the second 3.91 eV at lower temperatures (Figure 1A). Similar calculation produced only a single value (0.60 eV) for production of CO2 across the temperature range tested (Figure 1B). Since temperature dependence could be influenced by enzyme concentration, we quantified mcrA (Table 1), the gene encoding the subunit A of methyl coenzyme-M reductase that metabolized the last step of CH4 production. The activation energy was recalculated using the maximal rates of CH4 production normalized against the maximal abundance of mcrA. The pattern of temperature dependences did not change, i.e., showing two phases of temperature sensitivity (Figure 1C). The values of activation energy, however, decreased to 0.52 eV in upper temperature range (15–35°C) and 2.67 eV in lower temperature range (<15°C).
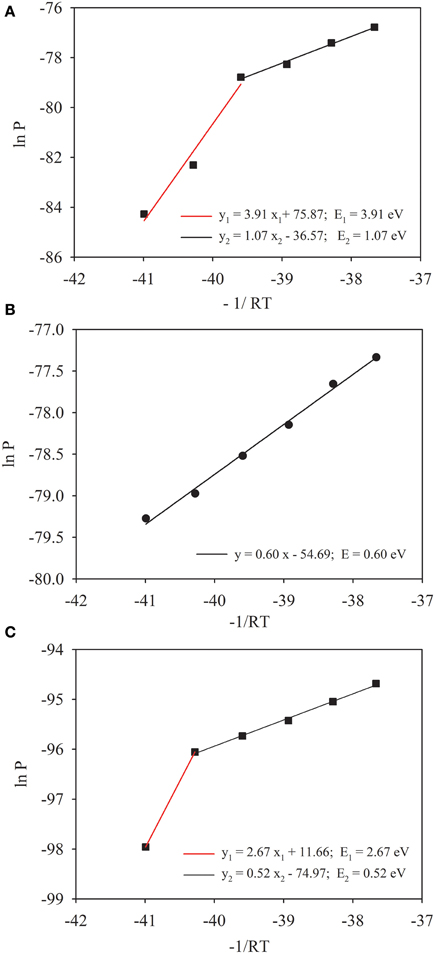
Figure 1. Temperature dependences of CH4 (A) and CO2 (B) production in Zoige wetland. Temperature dependence was characterized through plotting Arrhenius equation with “lnP” as a function of “−1/RT,” where P is the maximal rate of CH4 production, T is the absolute temperature and R is the Boltzmann constant (8.623 NU × 10−5 eV K−1). To take into account the effect of enzyme abundance, the maximal rate of CH4 production was normalized against the abundance of mcrA (C). Solid lines [except the red line in (C)] are the regression fits, where the slope indicates the apparent activation energy E (eV).
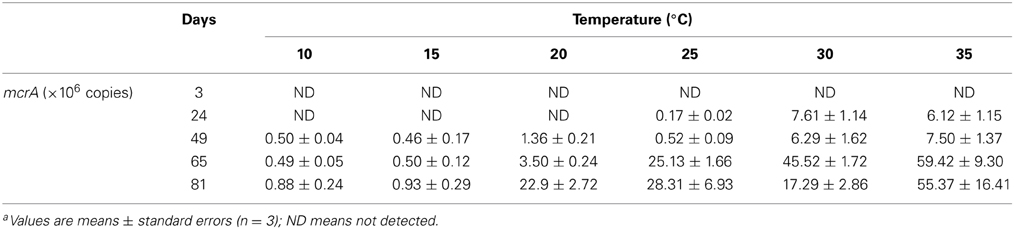
Table 1. Temperature and time dependent total mcrA gene abundancesa.
Community Structure and Methanogenic Pathway
Methanogen community was analyzed by using cloning, sequencing and T-RFLP analysis of the archaeal 16S rRNA genes. Three hundreds of 16S rRNA clones were retrieved from slurries incubated 49 days at 15°C, 25°C, and 35°C, respectively. The phylogenetic analysis of clone sequences showed that the archaeal community consisted of Methanosarcinaceae, Methanosaetaceae, Methanocellales, Methanomicrobiaceae, Methanobacteriaceae, and the uncultured euryarchaeotal RC-III, RC-V, LDS cluster and the crenarchaeotal group 1.1b and group 1.3 (Figure 2). The crenarchaeotal group 1.1b dominated accounting for over 40% of total archaeal clones (Figure S5). The relative abundances of methanogens and LDS cluster, however, increased relatively at 25°C and 35°C, while that of crenarchaeotal group 1.1b decreased.
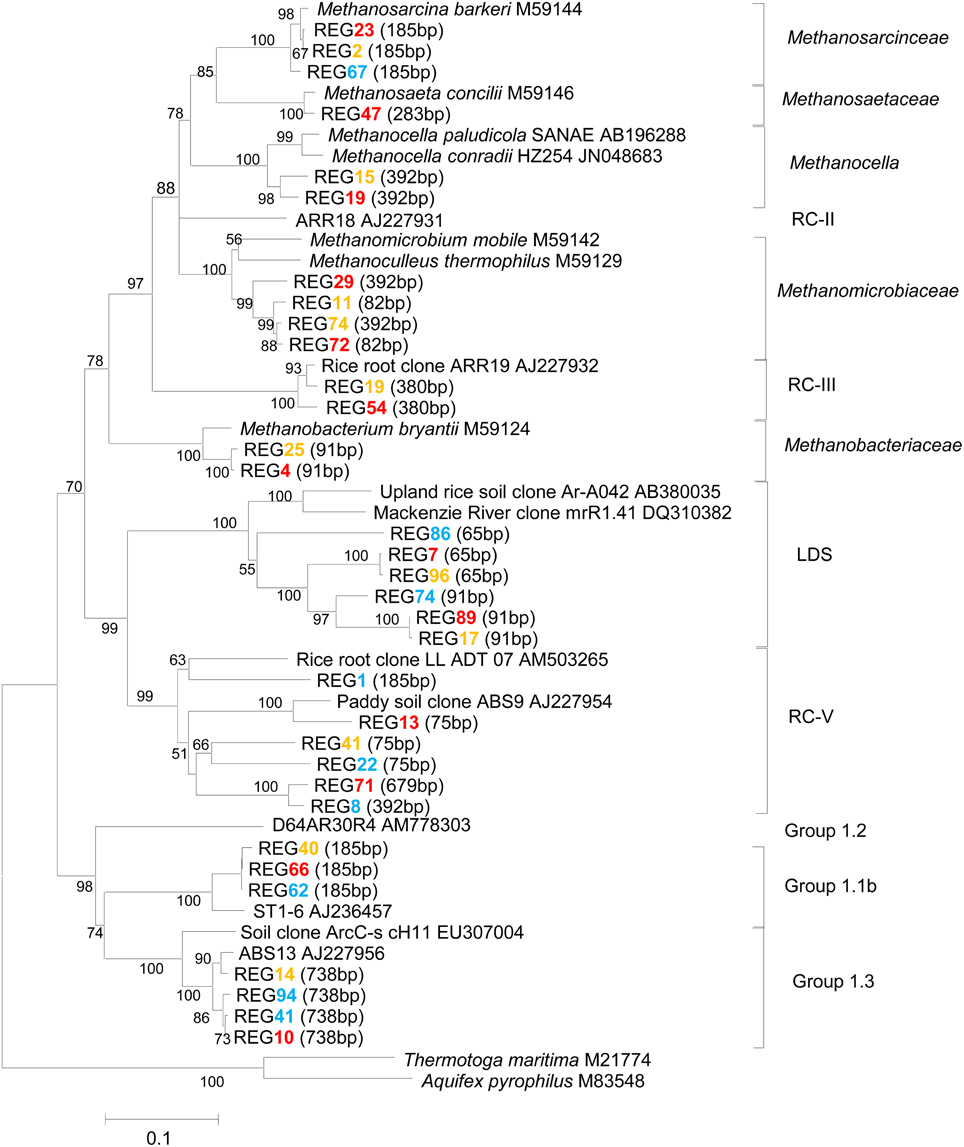
Figure 2. Phylogenetic relationship of representative archaeal 16S rRNA gene clone sequences generated from our Zoige wetland samples incubated 49 days at 15°C (blue), 25°C (orange) and 35°C (red), respectively. Clone libraries were constructed using primer set Ar109f and Ar915r, which produced about 800 bp size fragments. Sequences of this size were used for constructing the phylogenetic tree. The scale bar represents 10% sequence divergence. GenBank accession numbers of the reference sequences are indicated and in silico T-RF sizes are given in parentheses. Bootstrap values (%) were generated from 1000 replications and indicated at individual nodes.
T-RFLP profiles of archaeal 16S rRNA genes revealed that the 185-bp T-RF was predominant at the beginning and it remained dominant throughout the incubation at 10°C and 15°C (Figure 3). The relative abundance of this T-RF, however, decreased with incubation at higher temperatures. In contrast, the relative abundance of the 91-bp and 392-bp T-RFs increased over time in the incubations at 20°C and above. In silico analysis of clone sequences indicated that the 185-bp T-RF was related to Methanosarcinaceae and crenarchaeotal group 1.1b; the 91-bp T-RF to Methanobacteriaceae and LDS cluster, and the 392-bp T-RF to Methanomicrobiaceae and Methanocellales (Figure 2). Thus, the analyses of T-RFLP and clone sequences indicated that Methanosarcinaceae (185 bp) dominated the methanogen community at 10°C and 15°C, while the hydrogenotrophic methanogens Methanobacteriales (91 bp), Methanomicrobiales and Methanocellales (392 bp) increased when temperature increased to 20°C and above. The total abundance of mcrA increased with the increase of temperature (Table 1), indicating that the growth of methanogens was stimulated. Apparently, the growth of hydrogenotrophic methanogens was faster than aceticlastic ones, resulting in the shift of methanogen community.
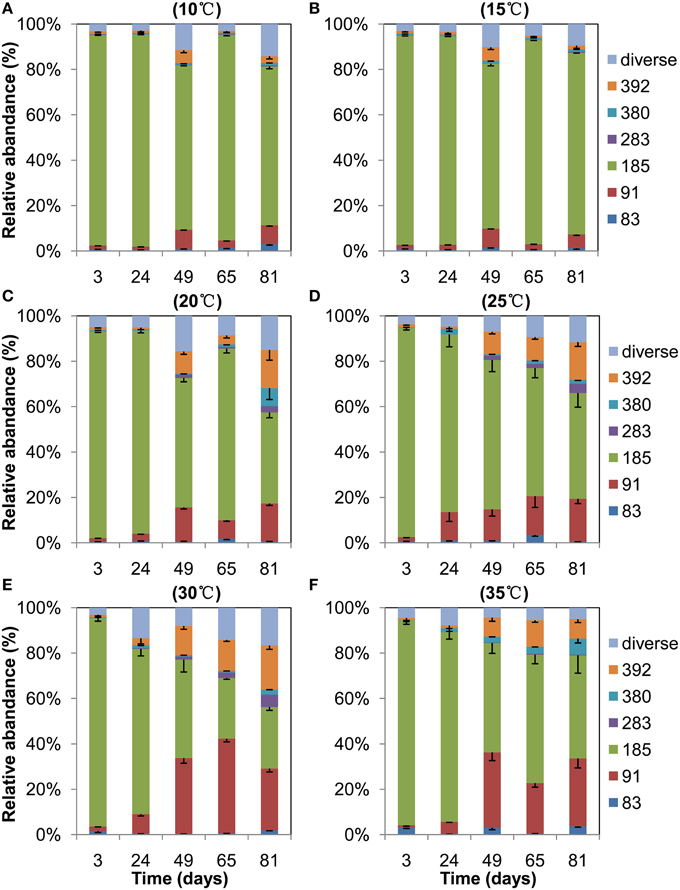
Figure 3. T-RFLP profiles of the archaeal community in anaerobically incubated Zoige wetland soil at 10°C (A), 15°C (B), 20°C (C), 25°C (D), 30°C (E), and 35°C (F), respectively. Data are means minus standard error (n = 3). Only major T-RFs are shown, the minor T-RFs are combined as Diverse.
To determine if the pathway of CH4 production was altered by temperature, we analyzed the δ13C abundances of CH4 and CO2 in incubations at 15°C, 25°C and 35°C, respectively (Figure 4). It is known that CH4 produced from CO2 reduction is more depleted in δ13C compared with CH4 produced from acetate cleavage (Conrad et al., 2009). The relative contribution of hydrogenotrophic versus aceticlastic methanogenesis, thus, can be inferred from isotopic signatures. We found that the δ13C values of CH4 were between −60 and −70‰ at 25°C and 35°C after day 20 when CH4 production was most active, while the values at 15°C were much higher (Figure 4A). In correspondence, the δ13C value of CO2 increased slightly in incubations at 25°C and 35°C, but showed decreasing tendency at 15°C. The apparent isotopic fractionation factor [αapp = 13CO2 + 103)/(δ 13CH4 + 103)] showed a gradual increase from 1.04 to 1.06 in incubations at 25°C and 35°C, but remaining lower than 1.03 over the incubation at 15°C (Figure 4B). The αapp value of 1.04 is characteristic typically for CH4 production from both CO2 and acetate (Conrad et al., 2009). The decrease of δ13C values of CH4 and the increase of αapp at high temperatures indicate that more of CH4 was produced from CO2 reduction. The δ13C data, therefore, indicated that methanogenic pathway shifted from the aceticlastic methanogenesis at 15°C to a mixture of both hydrogenotrophic and aceticlastic methanogenesis at 25°C and 35°C. This shift was in coincidence with the change in methanogen community as described above.
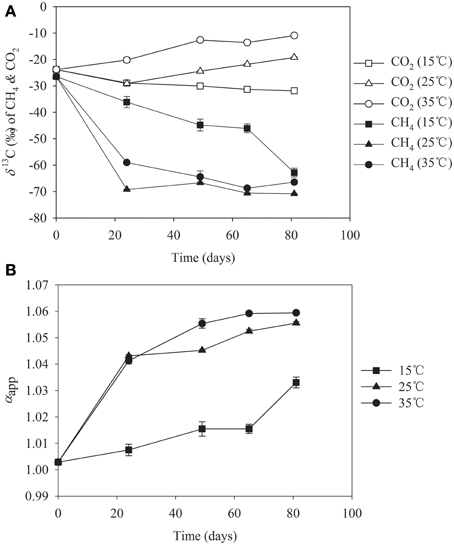
Figure 4. δ13C values of CO2 and CH4 (A) and the apparent fractionation factor (αapp) (B) in slurries of Zoige wetland incubated at 15°C (■), 25°C (▼), and 35°C (●), respectively.
Discussion
We showed here that temperature sensitivity of CH4 production in a Tibetan wetland soil sample changed with temperature. Specifically, two phases of temperature dependence can be distinguished, a high sensitivity in the low temperature range (<15°C) and a modest sensitivity under mesophilic conditions. This change of temperature sensitivity was in accordance with the shifts of methanogen composition and methanogenic pathway. It appeared that the sensitivity of aceticlastic methanogenesis was greater than that of hydrogenotrophic methanogenesis. The temperature sensitivity of methanogenesis revealed in the present experiment was in contrast to the meta-analysis and modeling that showed a universal temperature dependence of methane production across different systems (Yvon-Durocher et al., 2014). It, however, was in line with the finding that the temperature sensitivity differed markedly between photosynthesis, respiration and methanogenesis (Macdonald et al., 1998; Van Hulzen et al., 1999; Hartley et al., 2006; Zou and Gao, 2013), which indicated the dependence of temperature sensitivity on metabolic mechanisms.
Temperature sensitivity was known to be influenced by substrate availability and enzyme concentration (Davidson and Janssens, 2006). We used the maximal rate of CH4 production to reduce the influence of substrate availability. The obtained sensitivity (1.07 eV) at the upper temperature range (20–35°C) was close to that derived from the meta-analysis (Yvon-Durocher et al., 2014). But when the enzyme factor (i.e., methanogen biomass) was incorporated, a much lower sensitivity (0.52 eV) was obtained. This difference was apparently due to the growth of methanogen populations with increasing temperature. The seasonal change (growth) in methanogen populations has been often observed in various ecosystems (He et al., 2014; Kanta Gaihre et al., 2014; Sabrekov et al., 2014; Wei et al., 2014). To obtain the intrinsic temperature dependence, the growth of methanogens should be taken into account.
In contrast to CH4 production, CO2 production showed a single sensitivity factor over the temperature range tested. This pattern and the activation energy (0.60 eV) were in agreement with previous predictions from the meta-analysis (Yvon-Durocher et al., 2012). One possibility might be that the metabolic mechanisms for CO2 production did not change with temperature. Cautions, however, have to be taken with this explanation, because the mechanisms for CO2 production in anoxic slurries are complicated and remain unclear, and in addition CO2 in the headspace is in equilibrium with liquid that is controlled by slurry pH. We did not measure slurry pH and hence the effect of chemical equilibrium was not counted.
We found a very high sensitivity of CH4 production in low temperature range. The methanogen community was dominated by Methanosarcinaceae at these temperatures. Members of Methanosarcina are substrate-versatile, using acetate, hydrogen and methylated C1 compounds for methanogenesis. The 13C signatures of CH4 and CO2, however, indicated that it was the aceticlastic pathway that dominated methanogenesis at low temperatures. At higher temperatures, the relative abundance of hydrogenotrophic methanogens, consisting of Methanobacteriales, Methanomicrobiales and Methanocellales, increased. We did not detect Methanomicrobiales-like Fen Cluster as observed often in acidic boreal fens (Galand et al., 2005; Juottonen et al., 2008). This was probably due to the neutral condition (pH 7.5) in our wetland soil sample. Despite different compositions, the shifting pattern of methanogen community and methanogenic pathway upon temperature changes is consistent with previous studies on sediment (Conrad, 1999; Glissmann et al., 2004), rice soil (Fey and Conrad, 2000; Peng et al., 2008) and high arctic peat (Høj et al., 2008). Thus, it appears that the shift of methanogen community and methanogenic pathway in response to temperature change is ubiquitous across ecosystems.
In seasonality studies under field conditions, other factors like vegetation growth, substrate availability and water level could also be important driving forces for the shift of methanogen community and methanogenesis (Juottonen et al., 2008). Temperature sensitivity factor of methanogenic activity has not been determined in previous studies. But in the study on boreal fen, it was shown that the relative abundance of Methanosarcinaceae increased while the hydrogenotrophic Methanomicrobiales-associated fen cluster (FC) was decreased in winter compared with summer (Juottonen et al., 2008). This shift of community composition was accompanied with a markedly enhanced temperature response for potential CH4 production in winter soil sample relative to summer sample. If similar pattern of temperature sensitivity as revealed in this study and that in boreal fen exists ubiquitously in cold environments, large increase of global CH4 emissions may eventually occur in a warming climate.
In summary, we showed that temperature responses of CH4 production in a Zoige wetland soil sample displayed a high sensitivity in the low temperature range and a modest sensitivity under mesophilic conditions. This change in sensitivity was correlated with shifts of methanogen community and methanogenic pathway. We have to indicate that only one soil sample was tested in the present study, hence representing only a snapshot of temperature sensitivity of methanogens in Zoige wetland. Further researches shall be necessary to elucidate the mechanisms of this pathway-dependent temperature sensitivity across different ecosystems and take it into account in the future modeling and prediction of climate change impacts and feedbacks.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was partly supported by the National Basic Research Program of China (2011CB100505) and the National Natural Science Foundation of China (41130527).
Supplementary Material
The Supplementary Material for this article can be found online at: http://www.frontiersin.org/journal/10.3389/fmicb.2015.00131/abstract
References
Bond-Lamberty, B., and Thomson, A. (2010). Temperature-associated increases in the global soil respiration record. Nature 464, 579–582. doi: 10.1038/nature08930
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chen, H., Wu, N., Gao, Y., Wang, Y., Luo, P., and Tian, J. (2009a). Spatial variations on methane emissions from Zoige alpine wetlands of southwest China. Sci. Total Environ. 407, 1097–1104. doi: 10.1016/j.scitotenv.2008.10.038
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chen, H., Wu, N., Wang, Y., Zhu, D., Yang, G., Gao, Y., et al. (2013a). Inter-annual variations of methane emission from an open fen on the Qinghai-Tibetan Plateau: a three-year study. PLoS ONE 8:e53878. doi: 10.1371/journal.pone.0053878
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chen, H., Wu, N., Yao, S., Gao, Y., Wang, Y., Tian, J., et al. (2009b). Diurnal variation of methane emissions from an alpine wetland on the eastern edge of Qinghai-Tibetan Plateau. Environ. Monit. Assess. 164, 21–28. doi: 10.1007/s10661-009-0871-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chen, H., Zhu, Q. A., Peng, C., Wu, N., Wang, Y., Fang, X., et al. (2013b). Methane emissions from rice paddies natural wetlands, and lakes in China: synthesis and new estimate. Glob. Chang. Biol. 19, 19–32. doi: 10.1111/gcb.12034
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chin, K. J., Lukow, T., and Conrad, R. (1999). Effect of temperature on structure and function of the methanogenic archaeal community in an anoxic rice field soil. Appl. Environ. Microbiol. 65, 2341–2349.
Conrad, R. (1999). Contribution of hydrogen to methane production and control of hydrogen concentrations in methanogenic soils and sediments. FEMS Microbiol. Ecol. 28, 193–202. doi: 10.1111/j.1574-6941.1999.tb00575.x
Conrad, R. (2009). The global methane cycle: recent advances in understanding the microbial processes involved. Environ. Microbiol. Rep. 1, 285–292. doi: 10.1111/j.1758-2229.2009.00038.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Conrad, R., Klose, M., and Noll, M. (2009). Functional and structural response of the methanogenic microbial community in rice field soil to temperature change. Environ. Microbiol. 11, 1844–1853. doi: 10.1111/j.1462-2920.2009.01909.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Davidson, E. A., and Janssens, I. A. (2006). Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440, 165–173. doi: 10.1038/nature04514
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fey, A., and Conrad, R. (2000). Effect of temperature on carbon and electron flow and on the archaeal community in methanogenic rice field soil. Appl. Environ. Microbiol. 66, 4790–4797. doi: 10.1128/AEM.66.11.4790-4797.2000
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Galand, P. E., Fritze, H., Conrad, R., and Yrjala, K. (2005). Pathways for methanogenesis and diversity of methanogenic archaea in three boreal peatland ecosystems. Appl. Environ. Microbiol. 71, 2195–2198. doi: 10.1128/AEM.71.4.2195-2198.2005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Galand, P. E., Fritze, H., and Yrjala, K. (2003). Microsite-dependent changes in methanogenic populations in a boreal oligotrophic fen. Environ. Microbiol. 5, 1133–1143. doi: 10.1046/j.1462-2920.2003.00520.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Galand, P. E., Saarnio, S., Fritze, H., and Yrjala, K. (2002). Depth related diversity of methanogen archaea in Finnish oligotrophic fen. FEMS Microbiol. Ecol. 42, 441–449. doi: 10.1111/j.1574-6941.2002.tb01033.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Glissmann, K., Chin, K. J., Casper, P., and Conrad, R. (2004). Methanogenic pathway and archaeal community structure in the sediment of eutrophic Lake Dagow: effect of temperature. Microb. Ecol. 48, 389–399. doi: 10.1007/s00248-003-2027-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Godin, A., Mclaughlin, J. W., Webster, K. L., Packalen, M., and Basiliko, N. (2012). Methane and methanogen community dynamics across a boreal peatland nutrient gradient. Soil Biol. Biochem. 48, 96–105. doi: 10.1016/j.soilbio.2012.01.018
Good, I. J. (1953). The population frequencies of species and the estimation of population parameters. Biometrika 40, 237–264. doi: 10.1093/biomet/40.3-4.237
Hartley, I. P., Armstrong, A. F., Murthyw, R., Barron-Gafford, G., Ineson, P., and Atkin, O. K. (2006). The dependence of respiration on photosynthetic substrate supply and temperature: integrating leaf, soil and ecosystem measurements. Glob. Chang. Biol. 12, 1954–1968. doi: 10.1111/j.1365-2486.2006.01214.x
He, G., Li, K., Liu, X., Gong, Y., and Hu, Y. (2014). Fluxes of methane, carbon dioxide and nitrous oxide in an alpine wetland and an alpine grassland of the Tianshan Mountains, China. J. Arid Land 6, 717–724. doi: 10.1007/s,40333-014-0070-0
Høj, L., Olsen, R. A., and Torsvik, V. L. (2008). Effects of temperature on the diversity and community structure of known methanogenic groups and other archaea in high Arctic peat. ISME J. 2, 37–48. doi: 10.1038/ismej.2007.84
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
IPCC: Climate Change. (2014). Mitigation of Climate Change. Working Group III Contribution to the IPCC Fifth Assessment Report. Avaliable online at: http://www.mitigation2014.org
Juottonen, H., Tuittila, E.-S., Juutinen, S., Fritze, H., and Yrjala, K. (2008). Seasonality of rDNA- and rRNA-derived archaeal communities and methanogenic potential in a boreal mire. ISME J. 2, 1157–1168. doi: 10.1038/ismej.2008.66
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kanta Gaihre, Y., Wassmann, R., Tirol-Padre, A., Villegas-Pangga, G., Aquino, E., and Kimball, B. A. (2014). Seasonal assessment of greenhouse gas emissions from irrigated lowland rice fields under infrared warming. Agric. Ecosyst. Environ. 184, 88–100. doi: 10.1016/j.agee.2013.11.024
Karhu, K., Auffret, M. D., Dungait, J. A. J., Hopkins, D. W., Prosser, J. I., Singh, B. K., et al. (2014). Temperature sensitivity of soil respiration rates enhanced by microbial community response. Nature 513, 81–84. doi: 10.1038/nature13604
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kirschke, S., Bousquet, P., Ciais, P., Saunois, M., Canadell, J. G., Dlugokencky, E. J., et al. (2013). Three decades of global methane sources and sinks. Nat. Geosci. 6, 813–823. doi: 10.1038/ngeo1955
Liu, D. Y., Ding, W. X., Jia, Z. J., and Cai, Z. C. (2011). Relation between methanogenic archaea and methane production potential in selected natural wetland ecosystems across China. Biogeosciences 8, 329–338. doi: 10.5194/bg-8-329-2011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lueders, T., and Friedrich, M. (2000). Archaeal population dynamics during sequential reduction processes in rice field soil. Appl. Environ. Microbiol. 66, 2732–2742. doi: 10.1128/AEM.66.7.2732-2742.2000
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lueders, T., Manefield, M., and Friedrich, M. W. (2004). Enhanced sensitivity of DNA-and rRNA-based stable isotope probing by fractionation and quantitative analysis of isopycnic centrifugation gradients. Environ. Microbiol. 6, 73–78. doi: 10.1046/j.1462-2920.2003.00536.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ma, K., Conrad, R., and Lu, Y. H. (2012). Responses of methanogen mcrA genes and their transcripts to an alternate dry/wet cycle of paddy field soil. Appl. Environ. Microbiol. 78, 445–454. doi: 10.1128/AEM.06934-11
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Macdonald, J. A., Fowler, D., Hargreaves, K. J., Skiba, U., Leith, I. D., and Murray, M. B. (1998). Methane emission rates from a northern wetland; response to temperature, water table and transport. Atmos. Environ. 32, 3219–3227. doi: 10.1016/S1352-2310(97)00464-0
Mahecha, M. D., Reichstein, M., Carvalhais, N., Lasslop, G., Lange, H., Seneviratne, S. I., et al. (2010). Global convergence in the temperature sensitivity of respiration at ecosystem level. Science 329, 838–840. doi: 10.1126/science.1189587
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Noll, M., Matthies, D., Frenzel, P., Derakshani, M., and Liesack, W. (2005). Succession of bacterial community structure and diversity in a paddy soil oxygen gradient. Environ. Microbiol. 7, 382–395. doi: 10.1111/j.1462-2920.2005.00700.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Peng, J., Lu, Z., Rui, J., and Lu, Y. (2008). Dynamics of the methanogenic archaeal community during plant residue decomposition in an anoxic rice field soil. Appl. Environ. Microbiol. 74, 2894–2901. doi: 10.1128/AEM.00070-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rui, J., Peng, J., and Lu, Y. (2009). Succession of bacterial populations during plant residue decomposition in rice field soil. Appl. Environ. Microbiol. 75, 4879–4886. doi: 10.1128/AEM.00702-09
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rui, J., Qiu, Q., and Lu, Y. (2011). Syntrophic acetate oxidation under thermophilic methanogenic condition in Chinese paddy field soil. FEMS Microbiol. Ecol. 77, 264–273. doi: 10.1111/j.1574-6941.2011.01104.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sabrekov, A. F., Runkle, B. R. K., Glagolev, M. V., Kleptsova, I. E., and Maksyutov, S. S. (2014). Seasonal variability as a source of uncertainty in the West Siberian regional CH4 flux upscaling. Environ. Res. Lette. 9. doi: 10.1088/1748-9326/9/4/045008
Schloss, P. D., and Handelsman, J. (2005). Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 71, 1501–1506. doi: 10.1128/AEM.71.3.1501-1506.2005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Steinberg, L. M., and Regan, J. M. (2008). Phylogenetic comparison of the methanogenic communities from an acidic, oligotrophic fen and an anaerobic digester treating municipal wastewater sludge. Appl. Environ. Microbiol. 74, 6663–6671. doi: 10.1128/AEM.00553-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tian, J., Chen, H., Dong, X., and Wang, Y. (2012a). Relationship between archaeal community structure and vegetation type in a fen on the Qinghai-Tibetan Plateau. Biol. Fertil. Soils 48, 349–356. doi: 10.1007/s00374-011-0597-6
Tian, J., Zhu, Y., Kang, X., Dong, X., Li, W., Chen, H., et al. (2012b). Effects of drought on the archaeal community in soil of the Zoige wetlands of the Qinghai-Tibetan plateau. Eur. J. Soil Biol. 52, 84–90. doi: 10.1016/j.ejsobi.2012.07.003
Van Hulzen, J. B., Segers, R., Van Bodegom, P. M., and Leffelaar, P. A. (1999). Temperature effects on soil methane production: an explanation for observed variability. Soil Biol. Biochem. 31, 1919–1929. doi: 10.1016/S0038-0717(99)00109-1
Wei, D., Xu, R., Liu, Y., Wang, Y., and Wang, Y. (2014). Three-year study of CO2 efflux and CH4/N2O fluxes at an alpine steppe site on the central Tibetan Plateau and their responses to simulated N deposition. Geoderma 232, 88–96. doi: 10.1016/j.geoderma.2014.05.002
Yang, G., Chen, H., Wu, N., Tian, J., Peng, C., Zhu, Q., et al. (2014). Effects of soil warming, rainfall reduction and water table level on CH4 emissions from the Zoige peatland in China. Soil Biol. Biochem. 78, 83–89. doi: 10.1016/j.soilbio.2014.07.013
Yrjala, K., Tuomivirta, T., Juottonen, H., Putkinen, A., Lappi, K., Tuittila, E.-S., et al. (2011). CH4 production and oxidation processes in a boreal fen ecosystem after long-term water table drawdown. Glob. Chang. Biol. 17, 1311–1320. doi: 10.1111/j.1365-2486.2010.02290.x
Yuan, Q., and Lu, Y. (2009). Response of methanogenic archaeal community to nitrate addition in rice field soil. Environ. Microbiol. Rep. 1, 362–369. doi: 10.1111/j.1758-2229.2009.00065.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yvon-Durocher, G., Allen, A. P., Bastviken, D., Conrad, R., Gudasz, C., St-Pierre, A., et al. (2014). Methane fluxes show consistent temperature dependence across microbial to ecosystem scales. Nature 507, 488–491. doi: 10.1038/nature13164
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yvon-Durocher, G., Caffrey, J. M., Cescatti, A., Dossena, M., Del Giorgio, P., Gasol, J. M., et al. (2012). Reconciling the temperature dependence of respiration across timescales and ecosystem types. Nature 487, 472–476. doi: 10.1038/nature11205
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhang, G., Jiang, N., Liu, X., and Dong, X. (2008a). Methanogenesis from methanol at low temperatures by a novel psychrophilic methanogen, “Methanolobus psychrophilus” sp nov., prevalent in Zoige wetland of the Tibetan plateau. Appl. Environ. Microbiol. 74, 6114–6120. doi: 10.1128/AEM.01146-08
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhang, G., Tian, J., Jiang, N., Guo, X., Wang, Y., and Dong, X. (2008b). Methanogen community in Zoige wetland of Tibetan plateau and phenotypic characterization of a dominant uncultured methanogen cluster ZC-I. Environ. Microbiol. 10, 1850–1860. doi: 10.1111/j.1462-2920.2008.01606.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: methanogenesis, methanogens, temperature sensitivity, Zoige wetland, Tibetan plateau
Citation: Fu L, Song T and Lu Y (2015) Snapshot of methanogen sensitivity to temperature in Zoige wetland from Tibetan plateau. Front. Microbiol. 6:131. doi: 10.3389/fmicb.2015.00131
Received: 05 November 2014; Accepted: 04 February 2015;
Published online: 19 February 2015.
Edited by:
Paul Bodelier, Netherlands Institute of Ecology, NetherlandsReviewed by:
Xiuzhu Dong, Chinese Academy of Sciences, ChinaKim Yrjälä, University of Helsinki, Finland
Copyright © 2015 Fu, Song and Lu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yahai Lu, College of Urban and Environmental Sciences, Peking University, No. 5 Yiheyuan Road, Beijing 100871, China e-mail:bHV5aEBwa3UuZWR1LmNu