 Reshmi Upreti
Reshmi Upreti Pious Thomas
Pious Thomas- Endophytic and Molecular Microbiology Laboratory, Division of Biotechnology, ICAR – Indian Institute of Horticultural Research, Bangalore, India
This study was undertaken to assess if the root-associated native bacterial endophytes in tomato have any bearing in governing the host resistance to the wilt pathogen Ralstonia solanacearum. Internal colonization of roots by bacterial endophytes was confirmed through confocal imaging after SYTO-9 staining. Endophytes were isolated from surface-sterilized roots of 4-weeks-old seedlings of known wilt resistant (R) tomato cultivar Arka Abha and susceptible (S) cv. Arka Vikas on nutrient agar after plating the tissue homogenate. Arka Abha displayed more diversity with nine distinct organisms while Arka Vikas showed five species with two common organisms (Pseudomonas oleovorans and Agrobacterium tumefaciens). Screening for general indicators of biocontrol potential showed more isolates from Arka Abha positive for siderophore, HCN and antibiotic biosynthesis than from Arka Vikas. Direct challenge against the pathogen indicated strong antagonism by three Arka Abha isolates (P. oleovorans, Pantoea ananatis, and Enterobacter cloacae) and moderate activity by three others, while just one isolate from Arka Vikas (P. oleovorans) showed strong antagonism. Validation for the presence of bacterial endophytes on three R cultivars (Arka Alok, Arka Ananya, Arka Samrat) showed 8–9 antagonistic bacteria in them in comparison with four species in the three S cultivars (Arka Ashish, Arka Meghali, Arka Saurabhav). Altogether 34 isolates belonging to five classes, 16 genera and 27 species with 23 of them exhibiting pathogen antagonism were isolated from the four R cultivars against 17 isolates under three classes, seven genera and 13 species from the four S cultivars with eight isolates displaying antagonistic effects. The prevalence of higher endophytic bacterial diversity and more antagonistic organisms associated with the seedling roots of resistant cultivars over susceptible genotypes suggest a possible role by the root-associated endophytes in natural defense against the pathogen.
Introduction
Endophytic microorganisms colonize plants internally without any apparent adverse effects on the host (Hallmann et al., 1997; Gaiero et al., 2013). There is a growing interest in endophytic bacteria on account of their potential use in plant growth promotion, antagonistic effect on pests and pathogens, alleviation of abiotic stress and in phytoremediation (Compant et al., 2005; Ryan et al., 2008; Mercado-Blanco and Lugtenberg, 2014). Bacterial endophytes are generally known to enter the host from the surrounding soil through wounds in the roots (Hallmann et al., 1997; Compant et al., 2010) or through root hairs (Prieto et al., 2011; Mercado-Blanco and Prieto, 2012). They traverse the root cortex and reach various plant organs through the vascular system (Hallmann et al., 1997; Compant et al., 2010, 2011) while some use the apoplastic route (Sattelmacher, 2001; Reinhold-Hurek et al., 2007). Bacterial endophytes were earlier considered to be primarily colonizers in the inter-cellular or apoplastic spaces in the roots being present in relatively fewer numbers (Hallmann et al., 1997; Hallmann, 2001). Molecular studies have shown that there is considerable species diversity of bacterial endophytes albeit being present largely in a non-cultivable form (Lundberg et al., 2012; Sessitsch et al., 2012; Podolich et al., 2015). Intracellular colonization has also been documented in some plant systems (Pirttilä et al., 2000; de Almeida et al., 2009). A recent study employing banana shoot tissue has shown abundant endophytic bacteria in the two intracellular niches, namely in the cytoplasm and in the perispace between the cell wall and plasma membrane, and the terms ‘Cytobacts’ and ‘Peribacts’ have been coined to recognize the microorganisms in the respective intracellular niches (Thomas and Reddy, 2013; Thomas and Sekhar, 2014).
Bacterial wilt caused by the vascular pathogen, Ralstonia solanacearum (syn. Pseudomonas solanacearum) is a major constraint for tomato cultivation world over (Hayward, 1991; Genin and Denny, 2012). The wide host range covering major food and other economically important crops, broad geographic distribution, adaptation to survive in soil and water for long periods and the huge economic loss incited make the pathogen a very significant one worldwide (Genin and Denny, 2012; Mansfield et al., 2012). R. solanacearum invades the host through root injuries. The pathogen crosses the root cortex and overruns the xylem vessels leading to sudden wilting and plant death (Hayward, 1991; Genin and Denny, 2012). The similarities between bacterial endophytes and R. solanacearum in xylem colonization render the former as potential antagonistic and biocontrol agents against such vascular pathogens (Achari and Ramesh, 2014; Ting, 2014). Use of antagonistic bacteria for the biocontrol of bacterial wilt in tomato has been documented either as rhizospheric organisms (Vanitha et al., 2009) or as endophytes isolated from the same crop (Feng et al., 2013) or unrelated crops (Thomas and Upreti, 2014a).
Endophytic bacteria share an intimate symbiotic association with the host which makes them more valuable biocontrol agents (Compant et al., 2005; Bakker et al., 2013). Endophytes get an edge over their rhizospheric antagonist-counterparts on account of their ability to enter the host system without stimulating pathogen induced vulnerability responses but triggering host defense pathways (Conn et al., 2008; Gómez-Lama Cabanás et al., 2014; Podolich et al., 2015). Being internal colonizers, they could provide a barrier against the invading pathogens directly or through the production of bio-active compounds (Thomas and Upreti, 2014a; Podolich et al., 2015). Endophytes are better protected against abiotic stress and competing microbes compared with the rhizospheric counterparts (Hallmann et al., 1997; Ryan et al., 2008; Turner et al., 2013). While a vast majority of bacterial endophytes are known to be non-amenable for cultivation on common media (Lundberg et al., 2012; Sessitsch et al., 2012; Thomas and Sekhar, 2014), it entails that the organisms are easily cultivated to allow their agricultural exploitations. The present study was undertaken with a view to explore the extent of cultivable endophytic bacteria in transplantable-stage seedling roots of tomato cultivars that are either resistant or susceptible to R. solanacearum. Further, it was envisaged to evaluate the antagonistic and biocontrol features of the isolates to determine if the native endophytes played any role in governing the resilient property of the resistant cultivars.
Materials and Methods
Plant Material
Ralstonia solanacearum resistant (R) tomato (Solanum lycopersicum L.) cultivar Arka Abha and susceptible (S) cv. Arka Vikas (Thomas et al., 2015) were taken up as the primary test material in this study. In order to validate the findings, additional resistant (Arka Alok, Arka Ananya)/moderately resistant (Arka Samrat) and susceptible (Arka Ashish, Arka Meghali, and Arka Saurabhav) cultivars were employed. The names of genotypes are prefixed with R, MR, or S for easy recognition as resistant, moderately resistant or susceptible, respectively. Seedlings were raised in pasteurized organic cocopeat in protrays (Thomas et al., 2015) and used for the isolation of endophytes after 3½–4 weeks which corresponded to the stage of transplanting to the field when seedlings normally get exposed to the field pathogen inoculum (Thomas and Upreti, 2014b).
Confocal Imaging of Seedling Roots
Seedling roots were examined for bacterial colonization through confocal laser scanning microscopy (CLSM) after SYTO-9 staining. For this, tender roots from 3 to 4 weeks-old cocopeat – grown seedlings were washed, cut to ∼1 cm segments and were treated with 1× SYTO-9 (12 μM) from the LIVE/DEAD BacLight® bacterial viability kit L13152 (Molecular Probes, Invitrogen) as per the kit instructions. After 10–15 min staining, the lateral roots and root hairs were examined using a LSM 5 LIVE confocal microscope and the images were processed as described elsewhere (Thomas and Reddy, 2013). Root tissues were also examined after surface sterilization which involved a quick dip in 90% ethanol, a rinse in sterile distilled water (SDW) and 1 min sodium hypochlorite (2% available chlorine) treatment followed by six SDW rinses.
Isolation of Endophytes from Seedling Roots
Twenty randomly picked seedlings from RArka Abha and SArka Vikas 4 weeks after sowing were lifted with the plug of cocopeat and washed under running water taking care to minimize root injury. Seedlings were excised below the cotyledonary node and surface-sterilized essentially as per Zinniel et al. (2002). This involved a quick dip in 90% ethanol, a rinse in SDW and 1 min NaOCl (2% chlorine) treatment as above. After three rinses in SDW, 2% Na2S2O3 (10 min) was used to remove chloramine residues before finally rinsing the roots in SDW thrice. Root part was excised, blotted dry, weighed aseptically and macerated in a mortar employing 12.5 mM potassium phosphate buffer (Zinniel et al., 2002). After adjusting the volume to 10 ml g-1 tissue weight (100 stock), serial dilutions (101–105) were applied on NA through spotting- and tilt-spreading (SATS) approach (Thomas et al., 2012) with three replications per dilution. The plates were incubated at 30°C and the colony forming units (cfu) g-1 root tissue was determined on the third day. The NA plates used in this study were pre-monitored for absolute microbial sterility.
Identification of Organisms
Distinct bacterial colony types that emerged on NA from the root homogenate of RArka Abha (Tm-Ab01 to Tm-Ab09) and sArka Vikas (Tm-Av01 to Av05), serially numbered in the order of their relative abundance, were further purified through three rounds of streaking on NA. They were identified through partial 16S rRNA sequence homology analysis. For this polymerase chain reaction (PCR) was carried out with the primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R-Y (5′-GGYTACCTTGTTACGACTT-3′; Y = C/T) with the thermocyling conditions as described elsewhere (Thomas et al., 2008). The identity of these organisms was established and validated through megablast analysis to the cultured organisms at the National Centre for Biotechnological Information (NCBI) and the Seqmatch analysis with the Type Strains at the Ribosomal Database Project (RDP), Michigan State University. Wherever the identification was inconclusive based on NCBI homologies in the case of less common organisms, the highest species homology from NCBI or the similarity score from RDP was adopted to suggest the identity at sequence data submission to NCBI. The final identity was fixed as per the genus/species assigned by the GenBank at the acceptance of sequence data.
Screening of Organisms for the Indicators of Biocontrol Property
The endophytic organisms were tested for siderophore production through chrome azurol S method (Schwyn and Neilands, 1987) and for HCN production as per Ahmad et al. (2008). The isolates were screened through PCR for functional genes involved in the biosynthesis of bacterial non-ribosomal peptide synthetase (NRPS) and polyketide synthase (PKS) as markers for antibiotic production as per Miller et al. (2012). The primers MTF2 (5′-GCNGGYGGYGCNTAYGTNCC-3′) and MTR2 (5′-CCNCGDAYTTNACYTG-3′) were employed for NRPS giving a PCR product of ∼1000 bp, and the primers DKF (5′-GTGCCGGTNCCRTGNGYYTC-3′) and DKR (5′-GCGATGGAYCCNCARCARMG-3′) for PKS yielding ∼650–700 bp PCR product.
Pathogen and Culture Media
Ralstonia solanacearum ‘NH-Av01’ strain (NCBI acc. no. KJ412034; biovar 3) isolated from the bacterial ooze of a wilted ‘Arka Vikas’ plant as described elsewhere (Thomas and Upreti, 2014b,c) was used in antagonistic assays. The culture was stored as glycerol stocks at –80°C and revived on Kelman (1954) medium containing 1.0 g l-1 casein hydrolysate (C), 10 g l-1 bacteriological peptone (P), 5 g l-1 glucose (G), and 15 g l-1 bacteriological agar (A) and was fortified with 0.005% 2,3,5-Triphenyltetrazolium chloride (KM-TTC). The media constitutes were sourced from Hi Media Biosciences, Mumbai, except for TTC (Sigma, St. Louis, MO, USA) employing P14 lot of Type-1 peptone as per Thomas and Upreti (2014c). This was based on the observation that the colony characteristics, lawn formation and inhibition zone development were significantly influenced by the type and batch of peptone. Other media employed included casein-peptone-glucose-agar (CPGA) or CPG broth. Three additional Ralstonia isolates, namely, NH-Av05, NH-Av07, and KAU-Av01 were also used in the antagonistic assays.
Antagonistic Assay
Antagonistic assays were set up essentially as described earlier (Thomas and Upreti, 2014a). Briefly, 200 μl of 2-days-old CPGA or KM-TTC culture of 0.1 OD at 600 nm (approximately cfu of 108 ml-1) in peptone – salt (1 g l-1 each peptone and NaCl; Thomas et al., 2012) was spread over KM-TTC medium in 12 cm × 12 cm plates (Hi Media Biosciences, Mumbai) and wells of 6–7 mm diameter were prepared. After allowing R. solanacearum to establish at 30°C for 4 h, 50 μl of 0.2 OD endophytic bacterial inoculums in peptone – salt (approximately cfu in the range of 107–108 ml-1 for 0.1 OD culture depending on the organism) was applied in marked wells. After 20–25 min of surface drying, the plates were incubated inverted at 30°C. The antagonistic potential was rated based on the extent of clear zone formation, namely, strong (>20 mm; +++), medium (15–20 mm; ++), low (10–15 mm; +), or none.
Validation with Additional Tomato Cultivars
This included three additional resistant cultivars/F1 hybrids (RArka Alok, RArka Ananya F1, MRArka Samrat F1) and three susceptible cultivars (SArka Ashish, SArka Meghali, SArka Saurabhav; Thomas et al., 2015). Seedlings were grown in cocopeat in protrays and 5–10 surface-sterilized seedlings at 3½–4 weeks stage were employed for isolating the root endophytes. Tissue processing, culture purification, identification and assay for the antagonistic potential against the pathogen were undertaken as described earlier.
Nucleotide Sequences
The partial 16SrRNA gene sequences of the organisms have been deposited with the NCBI GenBank. The accession numbers are indicated in the Tables describing their identification.
Results
Confocal Imaging of Seedling Roots
The tender roots from 3 to 4 weeks-old RArka Abha and SArka Vikas seedlings showed green fluorescing bacterial cells on the root surface, inside the roots and in the surrounding film of water after SYTO-9 staining (Figures 1A1,B1). Root hairs showed abundant bacteria internally both along the cell periphery and in the cytoplasm (Figures 1A2,B2) confirming the endophytic colonization. Following surface sterilization, confocal imaging was impaired due to rapid signal bleaching (data not shown). However, it was possible to track the bacterial cells in both tender roots and root hairs with a notable reduction in the counts.
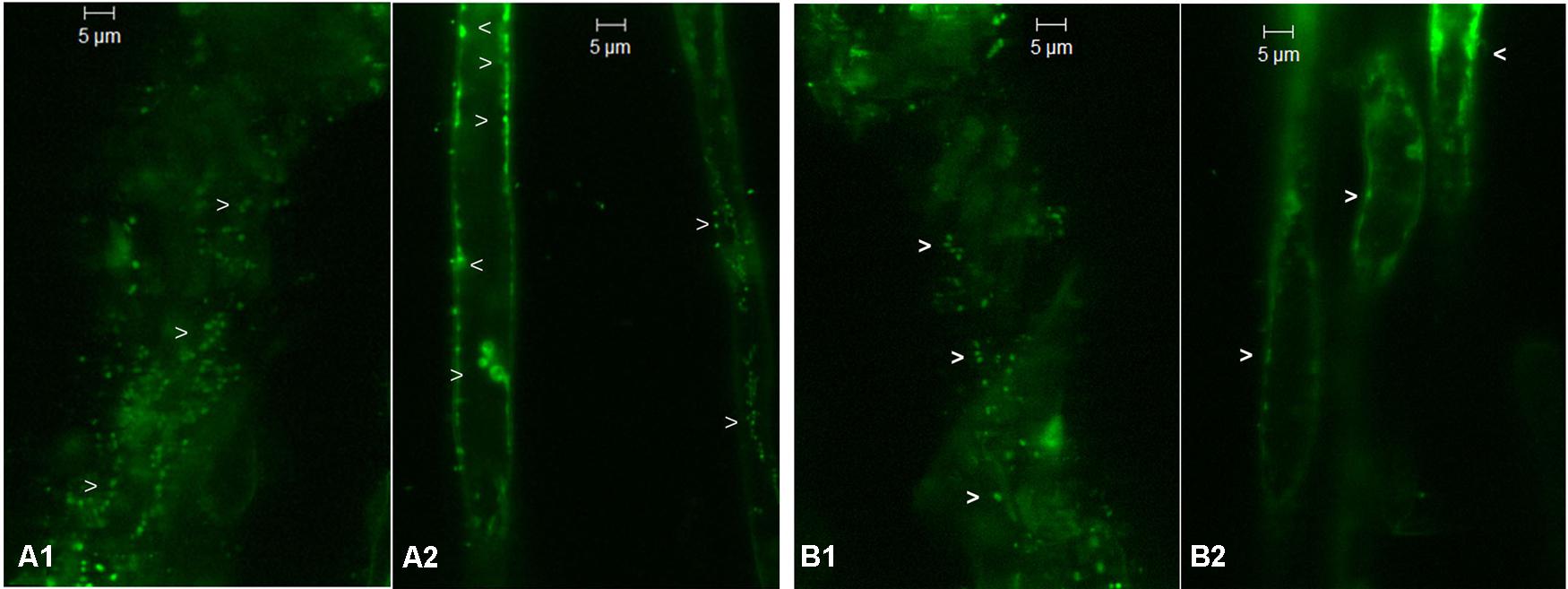
FIGURE 1. Confocal laser scanning microscopy images from SYTO-9 treated non-surface sterilized roots of tomato SArka Vikas and RArka Abha showing green fluorescing bacteria (indicated by arrow heads) on the surface (A1,B1, respectively) and internally along the cell periphery and inside root hairs (A2,B2, respectively).
Isolation and Identification of Endophytes from RArka Abha and SArka Vikas
Root growth in RArka Abha seedlings at endophyte isolation stage was relatively low compared with SArka Vikas. However, both the genotypes showed similar cfu estimates per unit fresh tissue weight (3.9 × 104 and 4.3 × 104, respectively). A number of distinct colonies were picked up which were finally assigned to nine distinct species in RArka Abha and five species in SArka Vikas (Table 1). The organisms from RArka Abha as per 16S rRNA gene sequence data accepted at NCBI GenBank included Pseudomonas oleovorans, Pseudomonas plecoglossicida, Pantoea ananatis, Citrobacter freundii, Staphylococcus hominis, Sphingobacterium multivorum, Enterobacter cloacae, Arthrobacter globiformis, and Agrobacterium tumefaciens. The isolates from SArka Vikas constituted P. oleovorans, Stenotrophomonas maltophilia, Bacillus pumilus, A. tumefaciens, and Microbacterium pumilum. The resistant cultivar apparently displayed more endophytic bacterial diversity with two organisms (P. oleovorans and A. tumefaciens) common to both the cultivars. Both RArka Abha and SArka Vikas showed more of Gram-negative bacteria (78 and 60%, respectively) and γ-subclass of Proteobacterium formed the commonest single phylogenetic group in both the cultivars (56 and 40%, respectively).
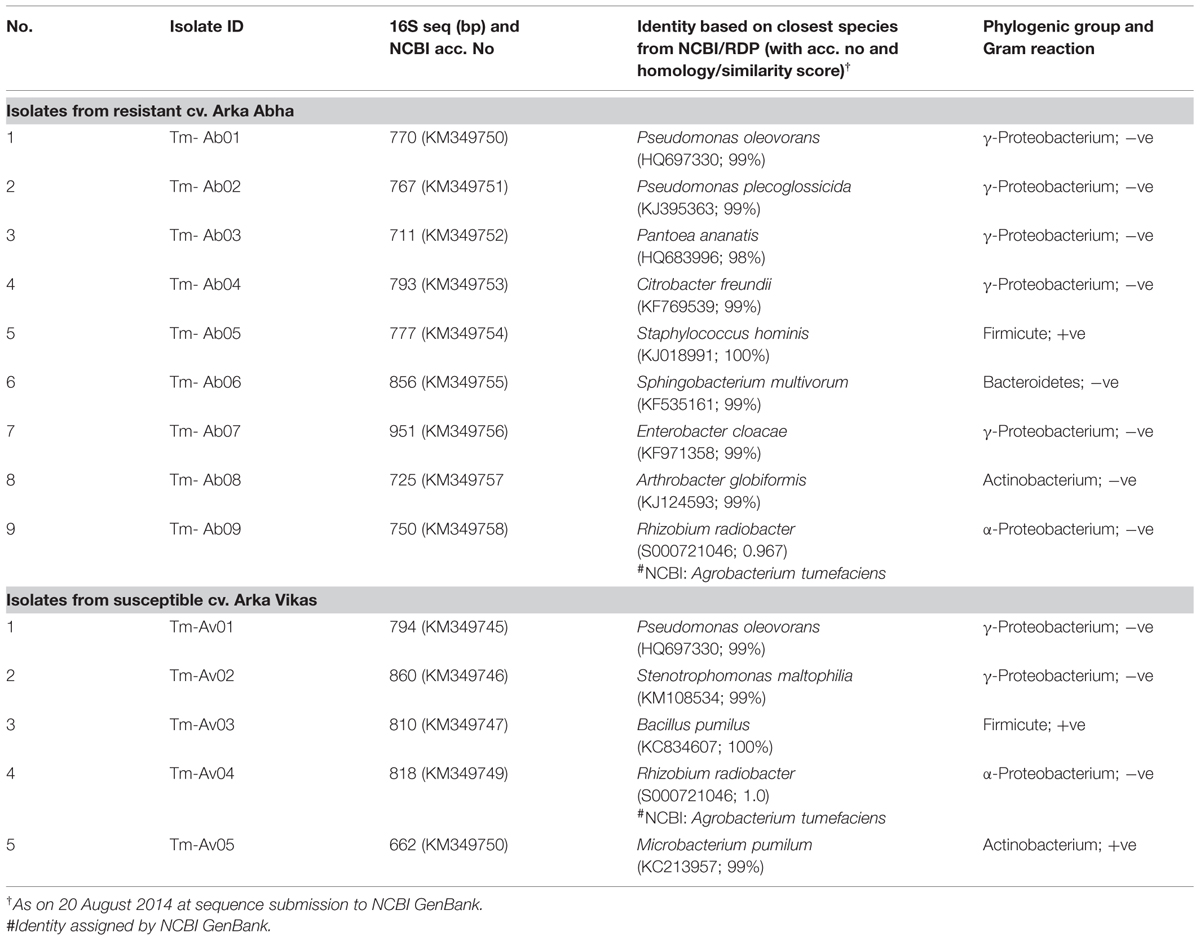
TABLE 1. Identification of bacterial endophytes isolated from the seedling root tissue of tomato cvs. Arka Abha and Arka Vikas.
Assessing the Endophytes for the Indicators of Biocontrol Property
Two of the RArka Abha isolates (Tm-Ab01, Tm-Ab03) showed siderophore production, two isolates (Tm-Ab03, Tm-Ab07) HCN production and three isolates (Tm-Ab02, Tm-Ab06, Tm-Ab08) proved positive for NRPS/ PKS (Table 2). The respective numbers for SArka Vikas were one, zero and one. Thus, the resistant cultivar harbored more organisms with biocontrol properties than the susceptible cultivar.
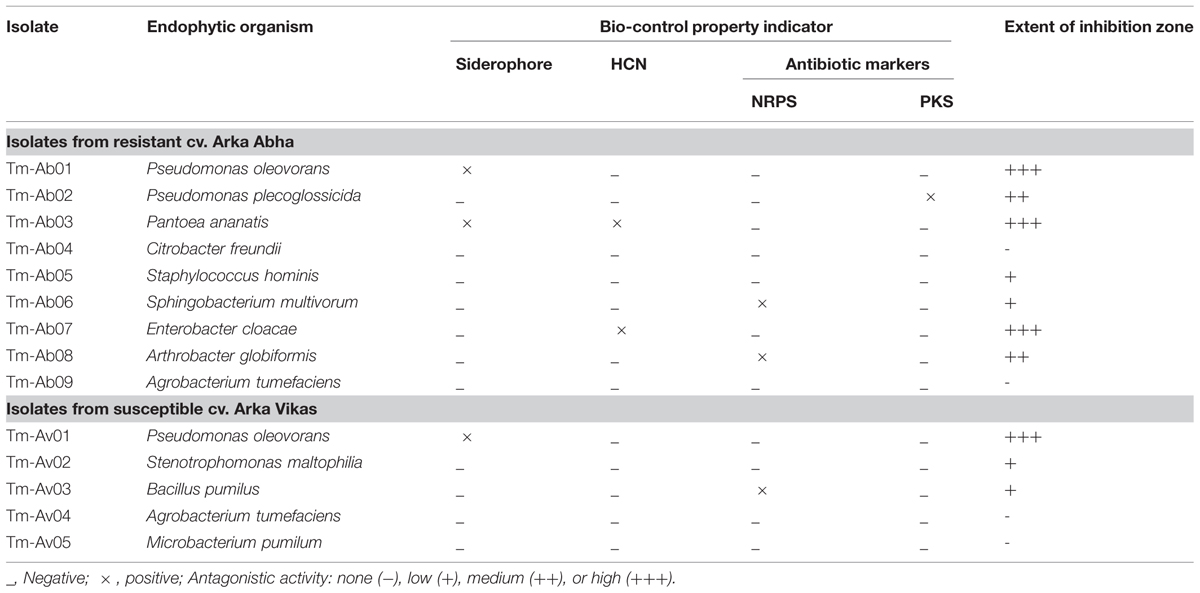
TABLE 2. Screening of bacterial endophytes from Ralstonia resistant Arka Abha and susceptible Arka Vikas tomato cultivars for the indicators of bio-control property.
Screening of Endophytes for Ralstonia Antagonistic Activity
Seven isolates from RArka Abha showed varying extents of antagonistic activity against R. solanacearum with Tm-Ab01 (P. oleovorans), Tm-Ab03 (P. ananatis), and Tm-Ab07 (E. cloacae) displaying significant effects, two isolates (Tm-Ab02, Tm-Ab08) offering medium activity and two others (Tm-Ab05, Tm-Ab06) showing low activity (Table 2). Among the SArka Vikas isolates, Tm-Av01 (P. oleovorans) showed strong antagonism while Tm-Av02 and Tm-Av03 displayed low activity. This was found true in a repeat assay and with three other isolates of R. solanacearum, namely, NH-Av05, NH-Av07 and KAU-Av01 (Figure 2).
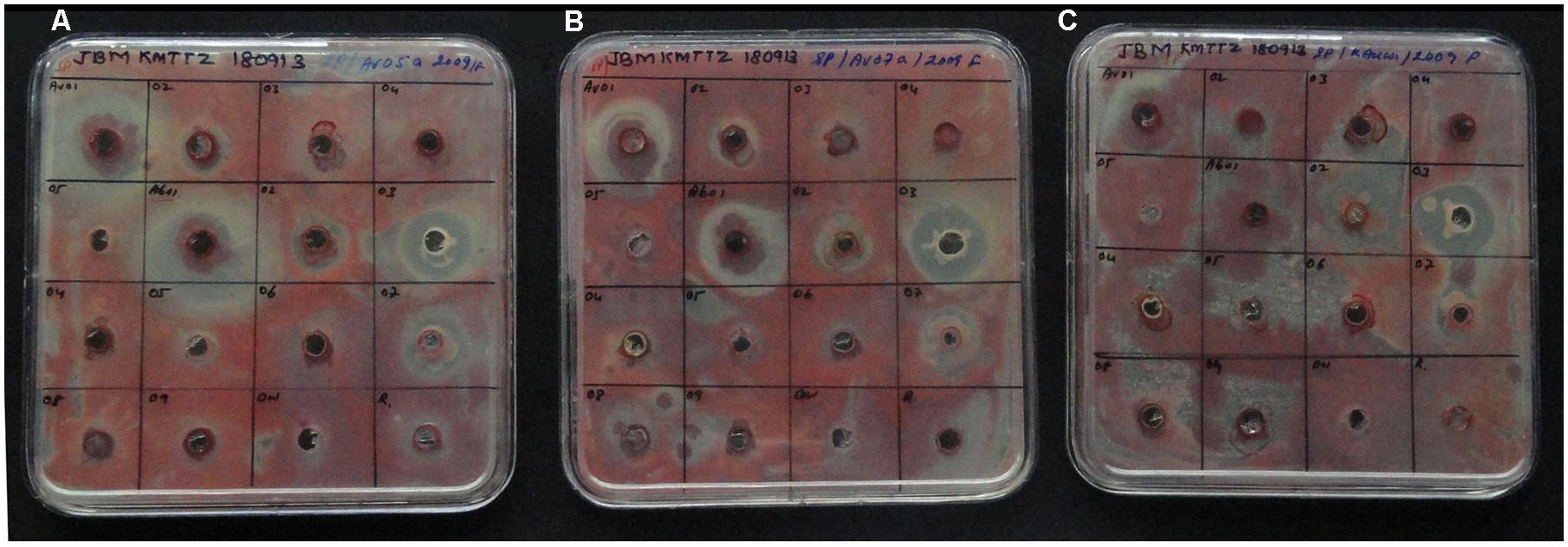
FIGURE 2. Screening of bacterial endophytes from susceptible cv. Arka Vikas and resistant cv. Arka Abha for the antagonistic activity against Ralstonia solanacearum isolates NH-Av05 (A), NH-Av07 (B), and KAU-Av01 (C). Treatment order: Row 1: Tm-Av01 to Av04; Row 2: Tm-Av05, Tm-Ab01 to Ab03; Row 3: Tm-Ab04 to Ab07; Row 4: Tm-Ab08, Ab09, distilled water control, Ralstonia inoculum, respectively.
Validation with Additional Resistant and Susceptible Cultivars
RArka Alok, RArka Ananya, and MRArka Samrat yielded 8–9 distinct organisms each while SArka Ashish, SArka Meghali, and SArka Saurabhav gave rise to four species each constituting a total of 37 isolates (Table 3). In general, there was a predominance of Gram negative bacteria in four cultivars (78, 62.5, 75, and 75%, respectively in RArka Alok, RArka Ananya, SArka Ashish, and SArka Saurabhav). However, MRArka Samrat and SArka Meghali showed 88 and 50% Gram positive organisms, respectively. The resistant cultivars showed more organisms with antagonistic potential in comparison with susceptible cultivars (Table 3) as discussed below.
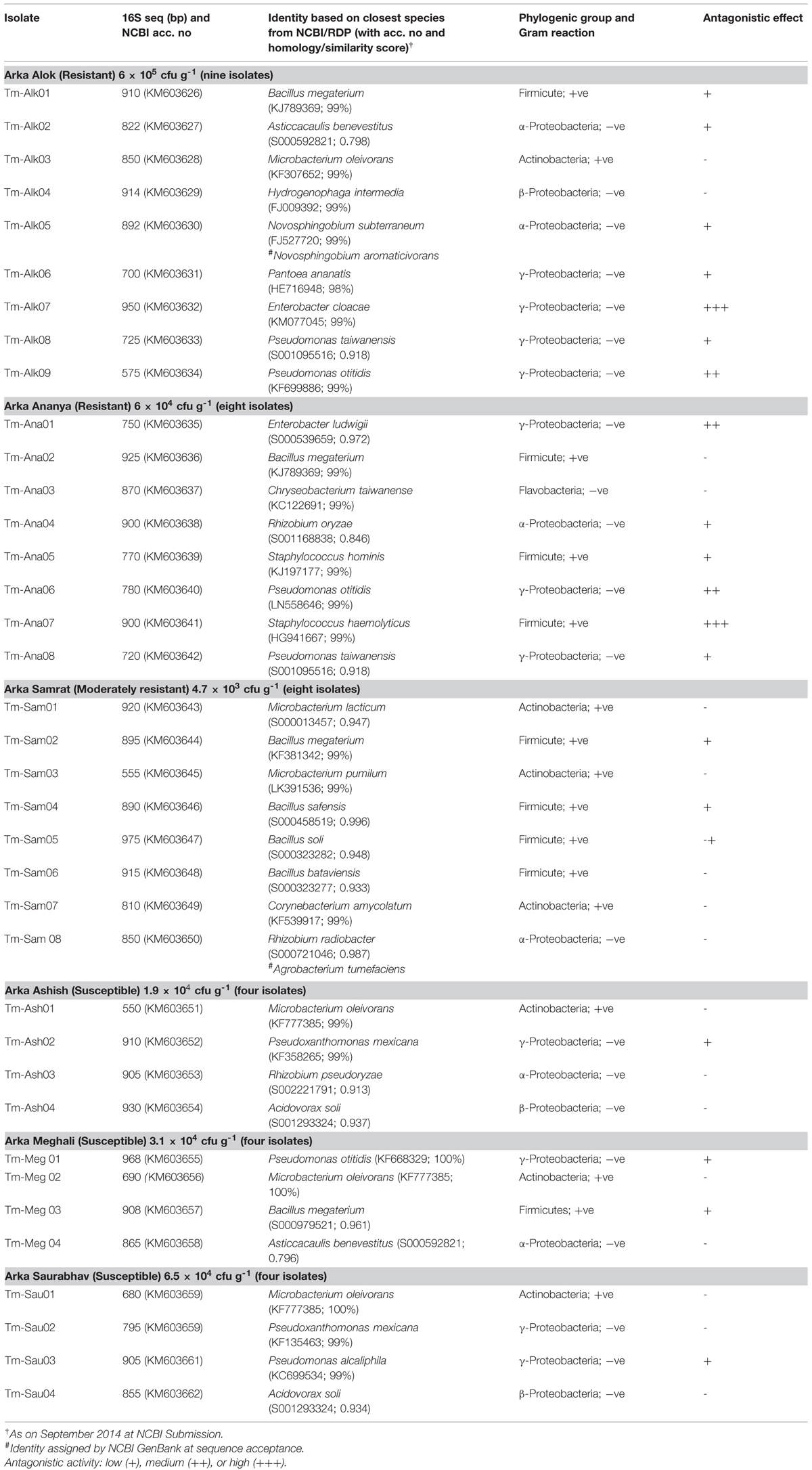
TABLE 3. Identification of bacterial endophytes from additional resistant and susceptible cultivars and their antagonistic activity against Ralstonia solanacearum NH-Av01 determined through agar-well diffusion assay.
Endophytes in Resistant and Susceptible Cultivars in Relation to Pathogen Antagonism
When the whole spectrum of root-associated bacterial endophytes in the four resistant and four susceptible cultivars of this investigation is considered, γ-Proteobacteria formed the commonest group followed by Actinobacteria, α-Proteobacteria and spore-forming Firmicutes (Figure 3A). The four resistant cultivars together yielded 34 endophytic bacteria which belonged to five classes (Proteobacteria, Actinobacteria, Firmicutes, Bacteroidetes, and Flavobacteria), 16 genera and 27 species while the isolates from susceptible cultivars represented three classes (Proteobacteria, Actinobacteria, and Firmicutes) including seven genera and 13 species (Table 4). The number of organisms displaying antagonistic activity during agar-well diffusion assay ranged from 4 to 7 in the former group while it was only one or two in the latter. Thus, among the R-cultivar isolates, 23 of them displayed varying levels of antagonistic effects while just seven from the S- category displayed such responses. Further, the extent of antagonistic activity as indicated by the diameter of clear zone was more with the isolates from R sources which included P. oleovorans, P. ananatis, and E. cloacae from RArka Abha, E. cloacae and P. otitidis from RArka Alok, and E. ludwigii, P. otitidis, and Staphylococcus haemolyticus from RArka Ananya. Maximum organisms with the antagonistic activity was observed with the γ-Proteobacteria group constituted by the genera Enterobacter, Pseudomonas, and Pantoea spp. with 15 out of 17 isolates showing antagonistic effects (Figure 3B). The next most promising group included non-spore forming Firmicutes, namely S. haemolyticus and S. hominis with all three isolates displaying good antagonistic potential.
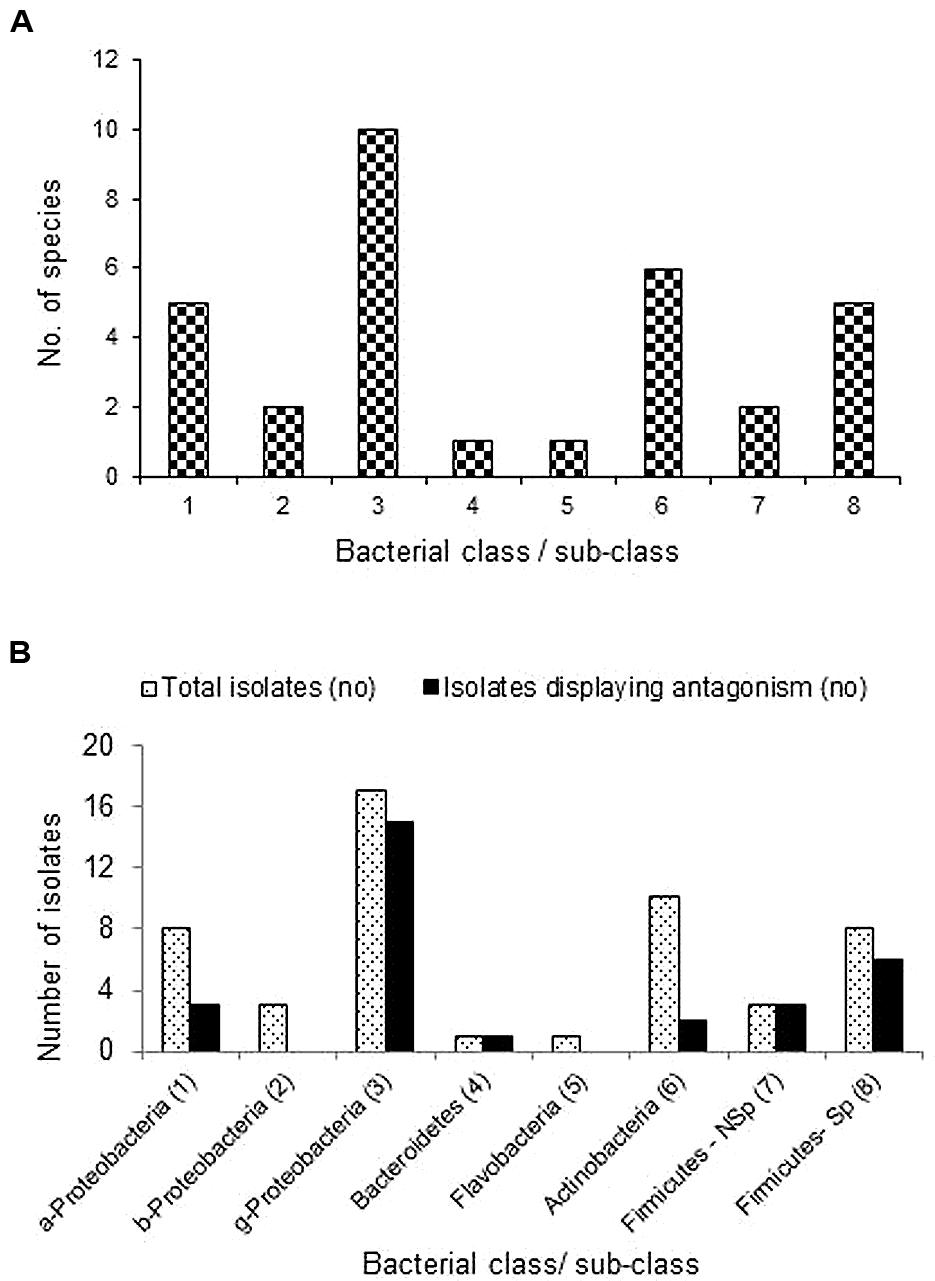
FIGURE 3. Extent of cultivable bacterial endophytes belonging to different bacterial classes and sub-classes across four Ralstonia resistant and four susceptible tomato cultivars (A), and the extent of organisms belonging to different phylogenetic groups displaying antagonistic activity against the pathogen (B); a-, b-, g- Proteobacteria represents α, β, and γ sub-classes; Firmicutes-Nsp and Sp represent non-sporulating and spore-forming Firmicutes.

TABLE 4. Extent of diversity of endophytic bacteria in Ralstonia resistant and susceptible cultivars of tomato in relation to pathogen antagonistic effect.
Discussion
Bacterial endophytes are known to confer protection against pathogens in a number of diseases (Compant et al., 2005; Mercado-Blanco and Lugtenberg, 2014) including Ralstonia wilt in tomato (Tan et al., 2011; Feng et al., 2013) and in related solanaceous crops (Ramesh and Phadke, 2012; Achari and Ramesh, 2014). Not many studies have addressed the diversity of endophytes or their possible involvement in offering a natural protection against this pathogen. The present study covering a number of tomato cultivars belonging to the resistant and susceptible categories enunciated the presence of greater cultivable endophytic bacterial diversity and more organisms with pathogen antagonistic potential in resistant cultivars. The isolates with antagonistic potential from resistant cultivars often showed accentuated pathogen inhibitory activity with one exception of Arka Samrat, which belonged to the moderately resistant category (Thomas et al., 2015). These observations suggested the possibility of an active role played by the endophytes in providing a natural protection against the pathogen in resistant cultivars. A recent study in tomato involving just one cultivar each from Ralstonia resistant and susceptible categories showed higher endophytic colonization, greater diversity and more pathogen antagonistic organisms in the former (Feng et al., 2013). Studies with other plant systems have also suggested the prevalence of a similar relationship (Araújo et al., 2002; Reiter et al., 2002). The endophytic communities perhaps are not random guests but essential associates interacting with the hosts (Gaiero et al., 2013; Podolich et al., 2015). It is postulated that the endophytic bacteria, which are largely in non-cultivable form, perhaps play an active role in crop protection through their revival to active form in response to pathogen attack or environmental stress (Podolich et al., 2015).
It was significant to note that several of the endophytes from RArka Abha were positive for biocontrol properties compared to SArka Vikas. The promising antagonistic organisms P. oleovorans and P. ananatis were siderophore producers while E. cloacae and P. ananatis showed HCN production indicating a relationship between antagonistic ability and siderophore/HCN production. On the other hand, no clear relationship between antibiotic (NRPS/PKS) biosynthesis capability and antagonistic property was observed. Therefore, it was imperative to undertake direct pathogen challenge assays to determine the antagonistic potential of the organisms.
Past investigations that reported elucidation of wilt-disease resistance mechanisms against R. solanacearum often laid emphasis on tissue-structural (Rahman and Abdullah, 1997; Rahman et al., 1999), genetic (Wang et al., 2000; Yang and Francis, 2006), or molecular attributes (Jacobs et al., 2012; Coll and Valls, 2013). It is generally concluded that the resistance trait of different cultivars is under genetic control. A perusal of reports on genetic basis of Ralstonia wilt resistance in tomato, however, showed considerable variations in the inheritance of this trait depending on the test hybrid combinations or the pathogen-isolate employed. This varied from monogenic to digenic dominant or recessive, or polygenic inheritance (Grimault et al., 1995; Mohamed et al., 1997; Hanson et al., 1998). The resistant cultivars have shown considerably low internal colonization by this pathogen than susceptible genotypes (Grimault et al., 1994; Rahman and Abdullah, 1997). The observations documented in this study raise a query whether the bacterial endophytes play either a direct active part or a supportive role in governing the resistance feature of a cultivar synergistic with the current concept of genetic inheritance of resistance.
Generally it is believed that the endophytes are recruited from the soil environment by the host influenced by the soil type where the host genotype is also known to have a significant influence (Compant et al., 2010; Lundberg et al., 2012; Mueller et al., 2015). It is difficult to visualize selective acquisition/recruitment of endophytes to take place from the soil in a resistant cultivar. The present study in which the seedlings were grown in pasteurized cocopeat ensured to be devoid of pathogenic Ralstonia leaves no room for such selective recruitment. The host genotype is known to play a significant role in governing the plant associated microorganisms, particularly endophytes (Hartmann et al., 2009; Lundberg et al., 2012; Bakker et al., 2013; Podolich et al., 2015). There are also reports on transmission of endophytes through seeds (Hardoim et al., 2012; Truyens et al., 2014) which would explain their possible integral association with a particular host cultivar. This study, supported by the recent reports on intracellular colonization by bacterial endophytes (Thomas and Reddy, 2013; Thomas and Sekhar, 2014), suggests the possibility of maternal inheritance of endophytes as seed colonizers. This hypothesis necessitates the isolation of same organisms from different batches of a cultivar. A subsequent trial with SArka Vikas showed three of the five isolates same as the earlier set (P. oleovorans, A. tumefaciens, and Microbacterium sp.) while two isolates constituted different organisms (Mitsuaria chitosanitabida and Kocuria palustris) indicating vertical transmission as well as lateral recruitment of bacterial endophytes. Three repeat trials with RArka Abha showed antagonistic P. oleovorans as a common associate. The current opinion on seed-transmission of endophytes appears divided with some in favor while others remaining inconclusive (Hallmann, 2001; Hardoim et al., 2012; Truyens et al., 2014). It now calls for more detailed investigations on seed colonization and vertical transmission of endophytes vis-à-vis genetic control of disease resistance. Observations with aseptically grown watermelon (Thomas and Aswath, 2014) and preliminary observations with papaya in vitro systems (Thomas, unpublished data) endorsed this possibility.
In this study, our main objective was to understand if the native endophytes in different tomato genotypes had any bearing on the inherent resistance characteristic of a cultivar. This study was confined to the natural endophytes without any external fortifications. It needs further investigations to elucidate how the organisms protect the crop in natural conditions; whether they act singly or synergistically, and their interactive action with other rhizospheric organisms. For instance, P. oleovorans constituted the most common endophyte in Arka Vikas, but this cultivar was susceptible to the pathogen (Thomas et al., 2015). It is possible that the population level of this antagonist in SArka Vikas was low to offer any formidable protection against the pathogenic intruder. It is feasible to increase the population levels of this endophyte through seed/seedling fortification which perhaps may impart some pathogen resistance in this cultivar. There is a general criticism that the in vitro antagonism activity by the endophytes may not be translated into effective biocontrol strategies. Our preliminary trials also suggested that exploiting antagonistic agents as potential biocontrol agents has uncertain results. The biocontrol effects are influenced by various other factors. The significance of microbe–microbe interactions in antimicrobial activity among soil bacteria is being increasingly recognized now (Tyc et al., 2014). Therefore, additional trials are needed to work out the biocontrol strategy which forms the next action plan.
In this study, the identification of the organisms was determined based on 16S rRNA sequence homology to the sequences available at the NCBI GenBank and RDP databases, and the final identity was fixed as per the genus/species assigned by the GenBank. The identification of some of the organisms based on such single gene data may not be conclusive as demonstrated with Pseudomonas spp. (Hilario et al., 2004). Classification based on additional genes is envisaged as we proceed with the biocontrol studies in the case of promising organisms.
The isolates from RArka Abha (P. oleovorans, P. ananatis, and E. cloacae) which showed strong antagonistic activity and that from SArka Vikas (P. oleovorans) are now short listed for further biocontrol investigations. The two isolates of P. oleovorans (Tm-Av01 and Tm-Ab01) and one A. tumefaciens isolate (Tm-Ab09) also showed higher seedling vigor index over uninoculated control in both tomato cultivars offering scope for their exploitation in organic vegetable growing (Thomas and Upreti, 2015). The hallmark of this study has been the elucidation that the native endophytic bacterial floras associated with the seedlings in resistant cultivars perhaps play a role in natural defense against the pathogen which hypothesis goes synergistic with the current concept of genetic inheritance of disease resistance. The present findings contribute to a better understanding of the basic aspects related to host - pathogen - endophyte interactions and open the scope for further explorations into the biological control of this pathogen.
Author Contributions
The experiments were planned together by the two authors. Bacterial isolation, PCR, and antagonistic assays were undertaken by RU. Bacterial identification, data interpretation, and manuscript preparation were done by PT. This work forms a part of the doctoral thesis of RU. The publication bears the Institute Contribution No. IIHR 92/2014.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Dr. T. P. Rajendran, Director, National Institute of Biotic Stress Management, Raipur, India for the critical reading of the manuscript and the suggestions. The study was funded by the ICAR – AMAAS Network project on ‘Endophytic microorganisms: Exploration of the endophytic microbial diversity in horticultural crops through metagenomics and cultivation’ through the National Bureau of Agriculturally Important Microorganisms (NBAIM), Mau Nath Bhanjan, Uttar Pradesh, India. The access to the confocal microscope facility at the Centre for Cellular and Molecular Platforms (C-CAMP), NCBS Campus, Bangalore and the help by Dr. Krishnamurthy, Navya and Shreyas in confocal microscopy work are acknowledged.
References
Achari, G. A., and Ramesh, R. (2014). Diversity, biocontrol, and plant growth promoting abilities of xylem residing bacteria from solanaceous crops. Int. J. Microbiol. 2014:14. doi: org/10.1155/2014/296521
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ahmad, F., Ahmad, I., and Khan, M. S. (2008). Screening of free living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 163, 173–181. doi: 10.1016/j.micres.2006.04.001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Araújo, W. L., Marcon, J., Maccheroni, W., van Elsas, J. D., van Vuurde, J. W., and Azevedo, J. L. (2002). Diversity of endophytic bacterial populations and their interaction with Xylella fastidiosa in citrus plants. Appl. Environ. Microbiol. 68, 4906–4914. doi: 10.1128/AEM.68.10.4906-4914.2002
Bakker, P. A. H. M., Berendsen, R. L., Doornbos, R. F., Wintermans, P. C. A., and Pieterse, C. M. J. (2013). The rhizosphere revisited: root microbiomics. Front. Plant Sci. 4:165. doi: 10.3389/fpls.2013.00165
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Coll, N. S., and Valls, M. (2013). Current knowledge on the Ralstonia solanacearum type III secretion system. Microb. Biotechnol. 6, 614–620.
Compant, S., Clément, C., and Sessitsch, A. (2010). Plant growth-promoting bacteria in the rhizo- and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 42, 669–678. doi: 10.1016/j.soilbio.2009.11.024
Compant, S., Duffy, B., Nowak, J., Clément, C., and Barka, E. A. (2005). Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 71, 4951–4959. doi: 10.1128/AEM.71.9.4951-4959.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Compant, S., Mitter, B., Colli-Mull, J. G., Gangl, H., and Sessitsch, A. (2011). Endophytes of grapevine flowers, berries and seeds: identification of cultivable bacteria, comparison with other plant parts, and visualization of niches of colonization. Microb. Ecol. 62, 188–197. doi: 10.1007/s00248-011-9883-y
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Conn, V. M., Walker, A. R., and Franco, C. M. M. (2008). Endophytic actinobacteria induce defense pathways in Arabidopsis thaliana. Mol. Plant Microb. Interact. 21, 208–218. doi: 10.1094/MPMI-21-2-0208
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
de Almeida, C. V., Andreote, F. D., Yara, R., Tanaka, F. A. O., Azevedo, J. L., and de Almeida, M. (2009). Bacteriosomes in axenic plants: endophytes as stable endosymbionts. World J. Microbiol. Biotech. 25, 1757–1764. doi: 10.1007/s11274-009-0073-8
Feng, H., Li, Y., and Liu, Q. (2013). Endophytic bacterial communities in tomato plants with differential resistance to Ralstonia solanacearum. Afr. J. Microbiol. Res. 7, 1311–1318.
Gaiero, J. R., McCall, C. A., Thompson, K. A., Day, N. J., Best, A. S., and Dunfield, K. E. (2013). Inside the root microbiome: bacterial root endophytes and plant growth promotion. Am. J. Bot. 100, 1738–1750. doi: 10.3732/ajb.1200572
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Genin, S., and Denny, T. P. (2012). Pathogenomics of the Ralstonia solanacearum species complex. Annu. Rev. Phytopathol. 50, 67–89. doi: 10.1146/annurev-phyto-081211-173000
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gómez-Lama Cabanás, C., Schilirò, E., Valverde-Corredor, A., and Mercado-Blanco, J. (2014). The biocontrol endophytic bacterium Pseudomonas fluorescens PICF7 induces systemic defense responses in aerial tissues upon colonization of olive roots. Front. Microbiol. 5:427. doi: 10.3389/fmicb.2014.00427
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Grimault, V., Anais, G., and Prior, P. (1994). Distribution of Pseudomonas solanacearum in the stem tissues of tomato plants with different levels of resistance to bacterial wilt. Plant Pathol. 43, 663–668. doi: 10.1111/j.1365-3059.1994.tb01604.x
Grimault, V., Prior, P., and Anais, G. (1995). A monogenic dominant resistance of tomato to bacterial wilt in Hawaii 7996 is associated with plant colonization by Pseudomonas solanacearum. J. Phytopathol. 143, 349–352. doi: 10.1111/j.1439-0434.1995.tb00274.x
Hallmann, J. (2001). “Plant interactions with endophytic bacteria,” in Biotic Interactions in Plant-Pathogen Associations, eds M. J. Jeger and N. J. Spence (Wallingford, Oxon: CABI Publishing), 87–119. doi: 10.1079/9780851995120.0087
Hallmann, J., Quadt-Hallmann, A., Mahaffe, W. F., and Kloepper, J. W. (1997). Bacterial endophytes in agricultural crops. Can. J. Microbiol. 43, 895–914. doi: 10.1139/m97-131
Hanson, P. M., Licardo, O., Wang, J. F., and Chen, J. T. (1998). Diallel analysis of bacterial wilt resistance in tomato derived from different sources. Plant Dis. 82, 74–78. doi: 10.1094/PDIS.1998.82.1.74
Hardoim, P. R., Hardoim, C. C. P., van Overbeek, L. S., and van Elsas, J. D. (2012). Dynamics of seed-borne rice endophytes on early plant growth stages. PLoS ONE 7:e30438. doi: 10.1371/journal.pone.0030438
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hartmann, A., Schmid, M., van Tuinen, D., and Berg, G. (2009). Plant-driven selection of microbes. Plant Soil 321, 235–257. doi: 10.1007/s11104-008-9814-y
Hayward, A. C. (1991). Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol. 29, 65–87. doi: 10.1146/annurev.py.29.090191.000433
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hilario, E., Buckley, T. R., and Young, J. M. (2004). Improved resolution on the phylogenetic relationships among Pseudomonas by the combined analysis of atpD, carA, recA and 16S rDNA. Antonie Van Leeuwenhoek 86, 51–64. doi: 10.1023/B:ANTO.0000024910.57117.16
Jacobs, J. M., Babujee, L., Meng, F., Milling, A., and Allen, C. (2012). The in planta transcriptome of Ralstonia solanacearum: conserved physiological and virulence strategies during bacterial wilt of tomato. MBio 3:e00114-12 doi: 10.1128/mBio.00114-12
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kelman, A. (1954). The relationship of pathogenicity in Pseudomonas solanacearum to colony appearance on a tetrazolium medium. Phytopathology 44, 693–696.
Lundberg, D. S., Lebeis, S. L., Paredes, S. H., Yourstone, S., Gehring, J., Malfatti, S., et al. (2012). Defining the core Arabidopsis thaliana root microbiome. Nature 488, 86–90. doi: 10.1038/nature11237
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mansfield, J., Genin, S., Magori, S., Citovsky, V., Sriariyanum, M., Ronald, P., et al. (2012). Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 13, 614–629. doi: 10.1111/j.1364-3703.2012.00804.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mercado-Blanco, J., and Lugtenberg, B. J. J. (2014). Biotechnological applications of bacterial endophytes. Curr. Biotechnol. 3, 60–75. doi: 10.2174/22115501113026660038
Mercado-Blanco, J., and Prieto, P. (2012). Bacterial endophytes and root hairs. Plant Soil 361, 301–306. doi: 10.1007/s11104-012-1212-9
Miller, K. I., Qing, C., Sze, D. M. Y., and Neilan, B. A. (2012). Investigations of the biosynthetic potential of endophytes in traditional Chinese anticancer herbs. PLoS ONE 7:e35953. doi: 10.1371/journal.pone.0035953
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mohamed, M. E. S., Umaharan, P., and Phelps, R. H. (1997). Genetic nature of bacterial wilt resistance in tomato (Lycopersicon esculentum Mill.) accession La 1421. Euphytica 96, 323–326. doi: 10.1023/A:1003075427304
Mueller, H., Berg, C., Landa, B. B., Auerbach, A., Moissl-Eichinger, C., and Berg, G. (2015). Plant genotype-specific archaeal and bacterial endophytes but similar Bacillus antagonists colonize Mediterranean olive trees. Front. Microbiol. 6:138. doi: 10.3389/fmicb.2015.00138
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pirttilä, A. M., Laukkanen, H., Pospiech, H., Myllyla, R., and Hohtola, A. (2000). Detection of intracellular bacteria in the buds of Scotch pine (Pinus sylvestris L.) by in situ hybridization. Appl. Environ. Microbiol. 66, 3073–3077. doi: 10.1128/AEM.66.7.3073-3077.2000
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Podolich, O., Ardanov, P., Zaets, I., Pirttilä, A. M., and Kozyrovska, N. (2015). Reviving of the endophytic bacterial community as a putative mechanism of plant resistance. Plant Soil 388, 367–377. doi: 10.1007/s11104-014-2235-1
Prieto, P., Schilirò, E., Maldonado-González, M. M., Valderrama, R., Barroso-Albarracín, J. B., and Mercado-Blanco, J. (2011). Root hairs play a key role in the endophytic colonization of olive roots by Pseudomonas spp. with biocontrol activity. Microb. Ecol. 62, 435–445. doi: 10.1007/s00248-011-9827-6
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rahman, M. A., and Abdullah, H. (1997). Susceptibility of Capsicum species and cultivars to Ralstonia solanacearum: anatomical differences and bacterial multiplication in resistant and susceptible cultivars. Pertanika J. Trop. Agric. Sci. 20, 1–11.
Rahman, M. A., Abdullah, H., and Vanhaecke, M. (1999). Histopathology of susceptible and resistant Capsicum annuum cultivars infected with Ralstonia solanacearum. J. Phytopathol. 147, 129–140. doi: 10.1111/j.1439-0434.1999.tb03819.x
Ramesh, R., and Phadke, G. S. (2012). Rhizosphere and endophytic bacteria for the suppression of eggplant wilt caused by Ralstonia solanacearum. Crop Prot. 37, 35–41. doi: 10.1016/j.cropro.2012.02.008
Reinhold-Hurek, B., Krause, A., Leyser, B., Miché, L., and Hurek, T. (2007). “The rice apoplast as a habitat for endophytic N2-fixing bacteria,” in The Apoplast of Higher Plants: Compartment of Storage, Transport and Reactions, eds B. Sattelmacher and W. J. Horst (Berlin: Springer), 427–443. doi: 10.1007/978-1-4020-5843-1_30
Reiter, B., Pfeifer, U., Schwab, H., and Sessitsch, A. (2002). Response of endophytic bacterial communities in potato plants to infection with Erwinia carotovora subsp. atroseptica. Appl. Environ. Microbiol. 68, 2261–2268. doi: 10.1128/AEM.68.5.2261-2268.2002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ryan, R. P., Germaine, K., Franks, A., Ryan, D. J., and Dowling, D. N. (2008). Bacterial endophytes: recent development and applications. FEMS Microbiol. Lett. 278, 1–9. doi: 10.1111/j.1574-6968.2007.00918.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sattelmacher, B. (2001). The apoplast and its significance for plant mineral nutrition. New Phytol. 149, 167–192. doi: 10.1046/j.1469-8137.2001.00034.x
Schwyn, B., and Neilands, J. B. (1987). Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160, 47–56. doi: 10.1016/0003-2697(87)90612-9
Sessitsch, A., Hardoim, P., Döring, J., Weilharter, A., Krause, A., Woyke, T., et al. (2012). Functional characteristics of an endophyte community colonizing rice roots as revealed by metagenomic analysis. Mol. Plant Microb. Interact. 25, 28–36. doi: 10.1094/MPMI-08-11-0204
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tan, H., Zhou, S., Deng, Z., He, M., and Cao, L. (2011). Ribosomal-sequence-directed selection for endophytic streptomycete strains antagonistic to Ralstonia solanacearum to control tomato bacterial wilt. Biol. Control 59, 245–254. doi: 10.1016/j.biocontrol.2011.07.018
Ting, A. S. Y. (2014). “Biosourcing endophytes as biocontrol agents of wilt diseases,” in Advances in Endophytic Research, eds V. C. Verma and A. C. Gange (New Delhi: Springer), 283–300.
Thomas, P., and Aswath, C. (2014). In vitro introduction of hardy alcohol resistant Bacillus spp. through aseptically grown watermelon seedlings. Adv. Microbiol. 4, 504–510. doi: 10.4236/aim.2014.49056
Thomas, P., and Reddy, M. K. (2013). Microscopic elucidation of abundant endophytic bacteria colonizing the cell wall – plasma membrane peri-space in the shoot-tip tissue of banana. AoB Plants 5:plt011. doi: 10.1093/aobpla/plt011
Thomas, P., Sadashiva, A. T., Upreti, R., and Mujawar, M. M. (2015). Direct delivery of inoculum to shoot tissue interferes with genotypic resistance to Ralstonia solanacearum in tomato seedlings. J. Phytopathol. 163, 320–323. doi: org/10.1111/jph.12281
Thomas, P., and Sekhar, A. C. (2014). Live cell imaging reveals extensive intracellular cytoplasmic colonization of banana genotypes by normally non-cultivable endophytic bacteria. AoB Plants 6:plu002. doi: 10.1093/aobpla/plu002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Thomas, P., Sekhar, A. C., and Mujawar, M. M. (2012). Non-recovery of varying proportions of viable bacteria during spread-plating governed by the extent of spreader usage and proposal for an alternate spotting-spreading approach to maximize the CFU. J. Appl. Microbiol. 113, 339–350. doi: 10.1111/j.1365-2672.2012.05327.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Thomas, P., Swarna, G. K., Roy, P. K., and Patil, P. (2008). Identification of cultivable and originally non-culturable endophytic bacteria isolated from shoot tip cultures of banana cv. Grand Naine. Plant Cell Tiss. Org. Cult. 93, 55–63. doi: 10.1007/s11240-008-9341-9
Thomas, P., and Upreti, R. (2014a). Testing of bacterial endophytes from non-host sources as potential antagonistic agents against tomato wilt pathogen Ralstonia solanacearum. Adv. Microbiol. 4:656–666. doi: 10.4236/aim.2014.410071
Thomas, P., and Upreti, R. (2014b). Influence of seedling age on the susceptibility of tomato plants to Ralstonia solanacearum during protray screening and at transplanting. Am. J. Plant Sci. 5, 1755–1762. doi: 10.4236/ajps.2014.512190
Thomas, P., and Upreti, R. (2014c). Significant effects due to peptone in Kelman medium on colony characteristics and virulence of Ralstonia solanacearum in tomato. Open J. Microbiol. 8, 87–105. doi: 10.2174/1874285801408010095
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Thomas, P., and Upreti, R. (2015). Evaluation of tomato seedling-root associated bacterial endophytes towards organic seedling production Org. Agri. doi: 10.1007/s13165-015-0111-9
Truyens, S., Weyens, N., Cuypers, A., and Vangronsveld, J. (2014). Bacterial seed endophytes: genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 7, 40–50. doi: 10.1111/1758-2229.12181
Turner, T. R., James, E. K., and Poole, P. S. (2013). The plant microbiome. Genome Biol. 14:209. doi: 10.1186/gb-2013-14-6-209
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tyc, O., van den Berg, M., Gerards, S., van Veen, J. A., Raaijmakers, J. M. de Boer, W., and Garbeva, P. (2014). Impact of interspecific interactions on antimicrobial activity among soil bacteria. Front. Microbiol. 5:567. doi: 10.3389/fmicb.2014.00567
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Vanitha, S. C., Niranjana, S. R., Mortensen, C. N., and Umesha, S. (2009). Bacterial wilt of tomato in Karnataka and its management by Pseudomonas fluorescens. Biocontrol 54, 685–695. doi: 10.1007/s10526-009-9217-x
Wang, J. F., Olivier, J., Thoquet, P., Mangin, B., Sauviac, L., and Grimsley, N. H. (2000). Resistance of tomato line Hawaii7996 to Ralstonia solanacearum Pss4 in Taiwan is controlled mainly by a major strain-specific locus. Mol. Plant Microb. Interact. 13, 6–13. doi: 10.1094/MPMI.2000.13.1.6
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yang, W., and Francis, D. M. (2006). “Genetics and breeding for bacterial diseases in tomato: prospects for marker-assisted selection,” in Genetic Improvement of Solanaceous Crops, Vol. 2, Tomato, eds M. J. Razdan and A. K. Mattoo (Boca Raton, FL: CRC Press), 381–414.
Zinniel, D. K., Lambrecht, P., and Harris, N. B. (2002). Isolation and characterization of endophytic colonizing bacteria from agronomic crops and prairie plants. Appl. Environ. Microbiol. 68, 2198–2208. doi: 10.1128/AEM.68.5.2198-2208.2002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: 16S rRNA homology, bacterial wilt resistance, biological control, confocal microscopy, endophytic bacteria, Ralstonia solanacearum, Solanum lycopersicum, tomato
Citation: Upreti R and Thomas P (2015) Root-associated bacterial endophytes from Ralstonia solanacearum resistant and susceptible tomato cultivars and their pathogen antagonistic effects. Front. Microbiol. 6:255. doi: 10.3389/fmicb.2015.00255
Received: 09 December 2014; Accepted: 15 March 2015;
Published online: 14 April 2015.
Edited by:
Jesús Mercado-Blanco, Consejo Superior de Investigaciones Científicas, SpainReviewed by:
Stephane Compant, Austrian Institute of Technology GmbH, AustriaDavid Ruano–Rosa, Instituto de Agricultura Sostenible–Consejo Superior de Investigaciones Científicas, Spain
Longxian Ran, Agricultural University of Hebei, China
Copyright © 2015 Upreti and Thomas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pious Thomas, Endophytic and Molecular Microbiology Laboratory, Division of Biotechnology, ICAR –Indian Institute of Horticultural Research, Hessaraghatta Lake, Bangalore–560089, Karnataka, IndiacGlvdXN0QGdtYWlsLmNvbQ==;cGlvdXN0QGlpaHIuZXJuZXQuaW4=