 Amlan K. Patra
Amlan K. Patra Zhongtang Yu
Zhongtang Yu- 1Department of Animal Sciences, The Ohio State University, Columbus, OH, USA
- 2Department of Animal Nutrition, West Bengal University of Animal and Fishery Sciences, Kolkata, India
In a previous study origanum oil (ORO), garlic oil (GAO), and peppermint oil (PEO) were shown to effectively lower methane production, decrease abundance of methanogens, and change abundances of several bacterial populations important to feed digestion in vitro. In this study, the impact of these essential oils (EOs, at 0.50 g/L) on the rumen bacterial community composition and population was further examined using the recently developed RumenBactArray. Species richness (expressed as number of operational taxonomic units, OTUs) in the phylum Firmicutes, especially those in the class Clostridia, was decreased by ORO and GAO, but increased by PEO, while that in the phylum Bacteroidetes was increased by ORO and PEO. Species richness in the genus Butyrivibrio was lowered by all the EOs. Increases of Bacteroidetes OTUs mainly resulted from increases of Prevotella OTUs. Overall, 67 individual OTUs showed significant differences (P ≤ 0.05) in relative abundance across the EO treatments. The predominant OTUs affected by EOs were diverse, including those related to Syntrophococcus sucromutans, Succiniclasticum ruminis, and Lachnobacterium bovis, and those classified to Prevotella, Clostridium, Roseburia, Pseudobutyrivibrio, Lachnospiraceae, Ruminococcaceae, Prevotellaceae, Bacteroidales, and Clostridiales. In total, 60 OTUs were found significantly (P ≤ 0.05) correlated with feed degradability, ammonia concentration, and molar percentage of volatile fatty acids. Taken together, this study demonstrated extensive impact of EOs on rumen bacterial communities in an EO type-dependent manner, especially those in the predominant families Prevotellaceae, Lachnospiraceae, and Ruminococcaceae. The information from this study may aid in understanding the effect of EOs on feed digestion and fermentation by rumen bacteria.
Introduction
In recent years, a variety of plant bioactive compounds, including saponins, essential oils (EOs), tannins, and flavonoids have been evaluated for their ability to modulate rumen microbial fermentation processes to improve feed utilization efficiency while decreasing methane emission and nitrogen excretion (Patra and Saxena, 2009, 2010). EOs have received more research interest than other types of plant bioactive compounds because they can effectively improve several important aspects of microbial metabolism in the rumen. For example, EOs can slow down degradation of starch and protein degradation, thereby reducing the risk of rumen acidosis in cattle fed high concentrate diets and decreasing intra-ruminal nitrogen turnover and nitrogen excretion, respectively, while inhibiting methanogenesis (McIntosh et al., 2003; Calsamiglia et al., 2007; Patra, 2011). Several studies also showed that supplementation of EOs to dairy cows resulted in increased milk yield and feed efficiency (Kung et al., 2008; Tassoul and Shaver, 2009; Giannenas et al., 2011). Although EOs have shown promise to inhibit the methanogenic archaea and methane production in the rumen (Patra and Saxena, 2010), adverse effects on fiber digestion and fermentation have also been reported, with the magnitude of these adverse effects varying considerably depending upon the type and dose of EO and diet composition (Calsamiglia et al., 2007; Macheboeuf et al., 2008). Determination of the effect of EOs on rumen bacterial communities is essential to understanding how EOs influence feed digestion and fermentation.
Phylogenetic microarrays enable simultaneous detection and semi-quantitation of thousands of different members of a microbiome (Rajilic-Stojanovic et al., 2009; Schatz et al., 2010). They have been used in investigations of bacteria in various environments, such as soil, human gut, human feces, sludges, and lakes (Small et al., 2001; Adamczyk et al., 2003; Castiglioni et al., 2004; Palmer et al., 2006; Rajilic-Stojanovic et al., 2009; Kang et al., 2010). A phylochip specifically for comprehensive analysis of rumen bacterial communities was recently developed based on 16S rRNA gene sequence with operational taxonomic units (OTUs) calculated at 97% sequence similarity (Kim et al., 2014). This phylochip, referred to as RumenBactArray, has more than 1600 OTU-specific probes that allow detection and semi-quantification of rumen bacteria. The RumenBactArray detects as few as 106 copies of a target and has a linear detection range of >4 orders of magnitude. The objectives of the present study were to assess the effect of three different EOs, which were shown to inhibit methanogenesis and modify rumen fermentation characteristics and select microbial populations and community (Patra and Yu, 2012), on the bacterial communities using the new RumenBactArray, and to identify associations between microbial populations, and digestion and fermentation variables. Broad effects were revealed and different sets of bacterial groups were affected differently by origanum oil (ORO), garlic oil (GAO), and peppermint oil (PEO). The results may help better understand the effect of these EOs on feed digestion and fermentation in the rumen.
Materials and Methods
Source of DNA Samples
The DNA samples analyzed in the present study had been analyzed previously using DGGE and qPCR (Patra and Yu, 2012). In that study, clove oil (CLO), eucalyptus oil (EUO), GAO, ORO, and PEO were evaluated in vitro at different doses (0.25, 0.50, and 1.0 g/L) for their effects on methane production, feed digestion, and fermentation. Their effects on communities of bacteria and of archaea were examined using DGGE, while changes in abundances of total bacteria, total archaea, total protozoa, and select cellulolytic bacteria (including Fibrobacter succinogenes, Ruminococcus flavefaciens, and Ruminococcus albus) were determined using specific qPCR. Different EOs were found to have different effect on most of the measurements in a dose dependent manner (Patra and Yu, 2012). In the present study, the effect of three of the EOs was further evaluated using RumenBactArray. These three EOs included GAO, ORO, and PEO. These three EOs were chosen because their principal bioactive components represent different chemical structures and stereochemistry: GAO contains alliin and allicin (organosulphur compounds); ORO contains thymol (monoterpinoid monoclyclic phenol); and PEO contains menthol (monoterpinoid monoclyclic non-phenol). Only the in vitro cultures (three replicates) that received 0.50 g/L each EO were used in the present study. This dose generally resulted in mild negative effects on digestion and rumen fermentation compared with high concentration (1 g/L). The control culture (three replicates) that did not receive EO was included in parallel. The detailed procedures of the in vitro experiment, sampling, and DNA extraction are available in the recent paper by Patra and Yu (2012).
Sample Preparation, Labeling, and Microarray Hybridization
Samples were prepared, labeled, and then subjected to microarray hybridization as described previously (Kim et al., 2014). Briefly, nearly full-length 16S rRNA genes were amplified from each metagenomic DNA sample using the universal primer set 27F (5′-AGA GTT TGA TCM TGG CTC AG-3′) and T7/1492R (5′-TCT AAT ACG ACT CAC TAT AGG GGG YTA CCT TGT TAC GAC TT-3′) as described previously. The amplicons were purified using a PCR purification Kit (Qiagen, Valencia, CA, USA) and then used in preparation of complementary RNA (cRNA) using a MEGAScript T7 in vitro transcription kit (Ambion, Austin, TX, USA). Following purification of the cRNA using a MEGAclear kit (Ambion, Austin, TX, USA), the cRNA was labeled with Cy5 at 37°C for 1 h using a Label IT®μ ArrayCy3/Cy5 Labeling kit (Mirus, Madison, WI, USA). The labeled cRNA was again purified to remove the free Cy5 dye using a MEGAclear kit. The labeled cRNA was quantified using the NanoDropt™ 1000 spectrophotometer, and then stored at −80°C until microarray hybridization.
Microarray hybridization was performed using Agilent Technologies' Hybridization gasket slides as described previously (Kim et al., 2014). Briefly, the hybridization solution containing 6× SSPE, 0.01% Tween-20, 0.01 mg/ml acetylated bovine serum albumin (BSA), 10% formamide, and 150 ng labeled cRNA was incubated at 65°C for 5 min and then placed on ice for 5 min. The Agilent hybridization cassette, the Agilent gasket slide and the microarray slide were preheated at 65°C while the hybridization solution was prepared. The hybridization solution was added to the center of the Agilent gasket slide, and then the RumenBactArray slide was placed over the gasket slide. The assembled cassette was placed in a HB-1000 hybridization oven (UVP, LLC) preset at 45°C to allow hybridization for 18 h with rotation set at 10 rpm. After hybridization, the microarray slides were bathed in 1× SSPE buffer (45°C) for 3 min twice and then in 0.25× SSPE for 30 s once prior to drying by centrifugation for 2 min at 400 g at room temperature.
Signal Detection and Data Analysis
The microarray slides were scanned and the fluorescent images of the hybridized microarrays were analyzed as previously described (Kim et al., 2014). Briefly, the hybridization images were captured with a GenePix 4000B Scanner (Axon Instruments, Union City, CA). The Cy5 fluorescence signal at each probe spot was measured using the GenePix®Pro 6.0 program (Axon Instruments). Probe spots on the scanned images were recognized by manually superimposing the gene allocation list (GAL) file provided by the manufacturer, which carries the annotation information of each spot on the microarray, over scanned images. Poor-quality probe spots that had a signal intensity lower than the background threshold, irregular size, or overlap with an adjacent spot were excluded from further analysis. Images were also inspected manually, and probe spots in low-quality areas of the microarray were also flagged and excluded from further analysis. The local median background signal intensity was subtracted from the median hybridization signal intensity of each separate probe spot. After background subtraction, normalization was performed based on the signal intensity of internal control probes targeting the bovine mitochondrial rRNA gene. Relative abundance of each OTU was calculated as its probe signal intensity percentage of total bacterial probe signal intensity. To assess if relative abundance of each bacterial OTU differed significantly (P ≤ 0.05) or tended (0.05 <P ≤ 0.10) to differ between control and the EO treatments, One-Way ANOVA was performed. When P ≤ 0.05, Tukey's test was employed to determine significant differences among the treatments. Both raw and normalized data are available in NCBI GEO under accession number GSE62624. Principal component analysis (PCA) using the MeV program (Saeed et al., 2006) was performed to compare the bacterial communities among the samples. The PCA scores on the first three principal components were further analyzed by multivariate analysis of variance (MANOVA) to test for differences in community composition among the treatments using SAS (2001).
Pearson correlation coefficients were calculated using SAS (2001) to examine correlation between relative abundances of each bacterial OTU and each of the fermentation data, including dry matter degradability (DMD), concentrations of ammonia, and molar percentages of acetate, propionate and butyrate, which were obtained in the previous study (Patra and Yu, 2012). Significant correlation was considered at P ≤ 0.05.
Results
Effects of EOs on Richness and Distribution of Ruminal Bacteria
The species richness, expressed as numbers of OTUs detected, was affected by all the EO treatments (Table 1). Overall, 228 OTUs with relative abundances greater than 0.5% were identified within different phyla among all the treatments. Of the OTUs with a relative abundance ≥0.5% (of total bacterial probe signal intensity), the number of OTUs in the phylum Firmicutes was considerably lower, especially in the class Clostridia, in the cultures that received ORO or GAO than in the control (Figure 1). However, the number of OTUs with a relative abundance ≥1.0% was increased by PEO compared with the control, while the number of OTUs with a relative abundance ≥0.5% was similar between the control and the PEO cultures. Compared to the control, all the EO treatments decreased the number of OTUs in the genus Butyrivibrio. The numbers of OTUs with a relative abundance ≥0.5% and unclassified within Ruminococcaceae were decreased by GAO and ORO. The numbers of OTUs with a relative abundance ≥1% in the phylum Bacteroidetes were increased by ORO and PEO, but were not affected by GAO; whereas the numbers of OTUs with a relative abundance ≥0.5% in this phylum was greater for PEO, but not for ORO or GAO, than for the control. The increases of OTU richness in Bacteroidetes mainly resulted from increases of Prevotella OTUs. No significant changes were observed in the number of OTUs classified to other phyla, families, or genera for any of the EO treatments.
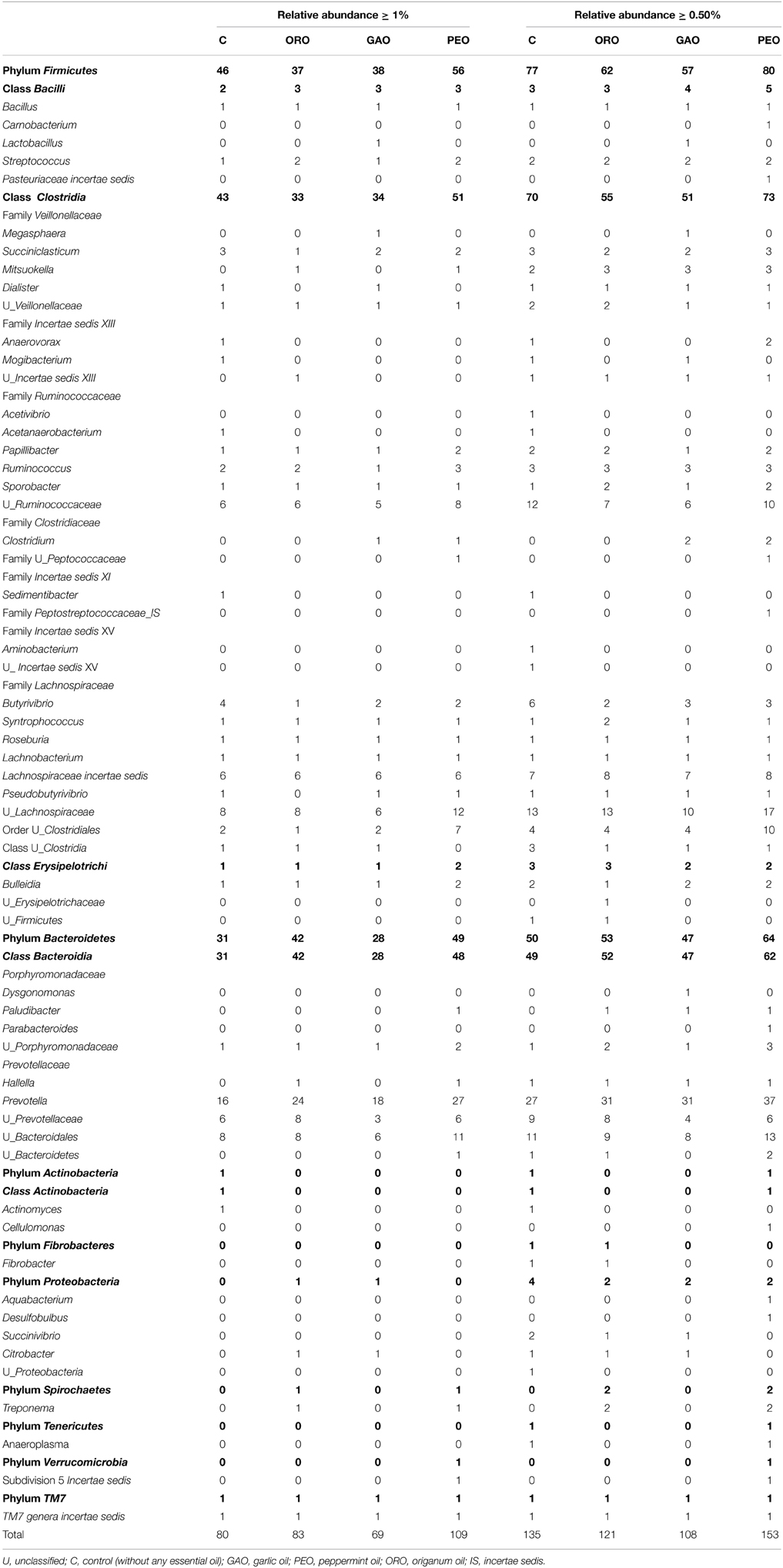
Table 1. Effects of essential oils on richness and distribution of major ruminal bacterial taxa.
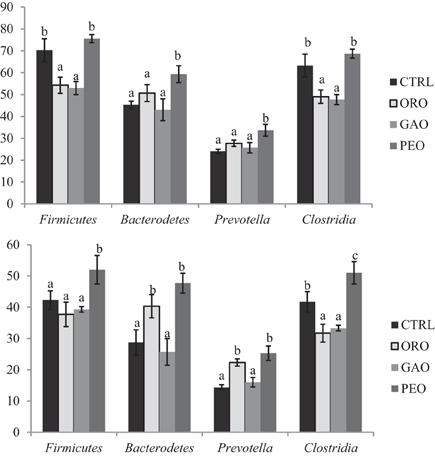
Figure 1. Average number of major bacterial OTUs (relative abundance of ≥0.5 or ≥1%) identified in Firmicutes, Bacteroidetes, Clostridia, and Prevotella. CTRL, control; ORO, origanum oil; GAO, garlic oil; PEO, peppermint oil; Different letters (a–c) above the bars indicate significant (P ≤ 0.05) differences among the treatments (three replicates per treatment).
Effects of EOs on Populations of Ruminal Bacteria
The effect of the tested EOs was assessed on individual bacterial populations as reflected by changes in their relative abundance. Overall, 67 individual OTUs showed significant differences (P ≤ 0.05) and 44 OTUs tended (0.05 <P ≤ 0.10) to change in relative abundance in response to the EOs treatments. The populations in the following phyla were examined closely as they represent most of the OTUs detected:
Phylum Firmicutes
In the phylum Firmicutes, 42 individual OTUs showed significant (P ≤ 0.05) differences (Table 2) and 27 OTUs tended (0.05 < P ≤ 0.10) to differ (Table S1) in relative abundance among the EO treatments (Table 2 and Table S1). Compared with the control, the following OTUs were decreased significantly by ORO (Table 2): Succiniclasticum_9 (a taxon name followed by “_” and a number represents a specific species-equivalent OTU within that taxon), unclassified Ruminococcaceae_20, unclassified Ruminococcaceae_49, Syntrophococcus_1, Roseburia_1, Lachnospiraceae incertae sedis_69, and unclassified Lachnospiraceae_137. On the other hand, other OTUs in this phylum, including unclassified Ruminococcaceae_132, and unclassified Clostridiales_73 were increased in response to the ORO addition. The relative abundance of Lachnospiraceae incertae sedis_69 was lower for the GAO treatment than for the control. However, GAO increased the relative abundance of unclassified Ruminococcaceae_132, unclassified Ruminococcaceae_75, Clostridium_2, Clostridium_9, and unclassified Lachnospiraceae_49. For PEO, some of the OTUs related to Syntrophococcus sucromutans, Succiniclasticum ruminis, together with some OTUs unclassified within the families Ruminococcaceae_49 and Lachnospiraceae_69 were decreased, whereas other OTUs classified to Roseburia, Prevotella, Pseudobutyrivibrio, and unclassified within Ruminococcaceae, Lachnospiraceae, Clostridiales, Bacteroidales and Ad-C-8H were increased notably.
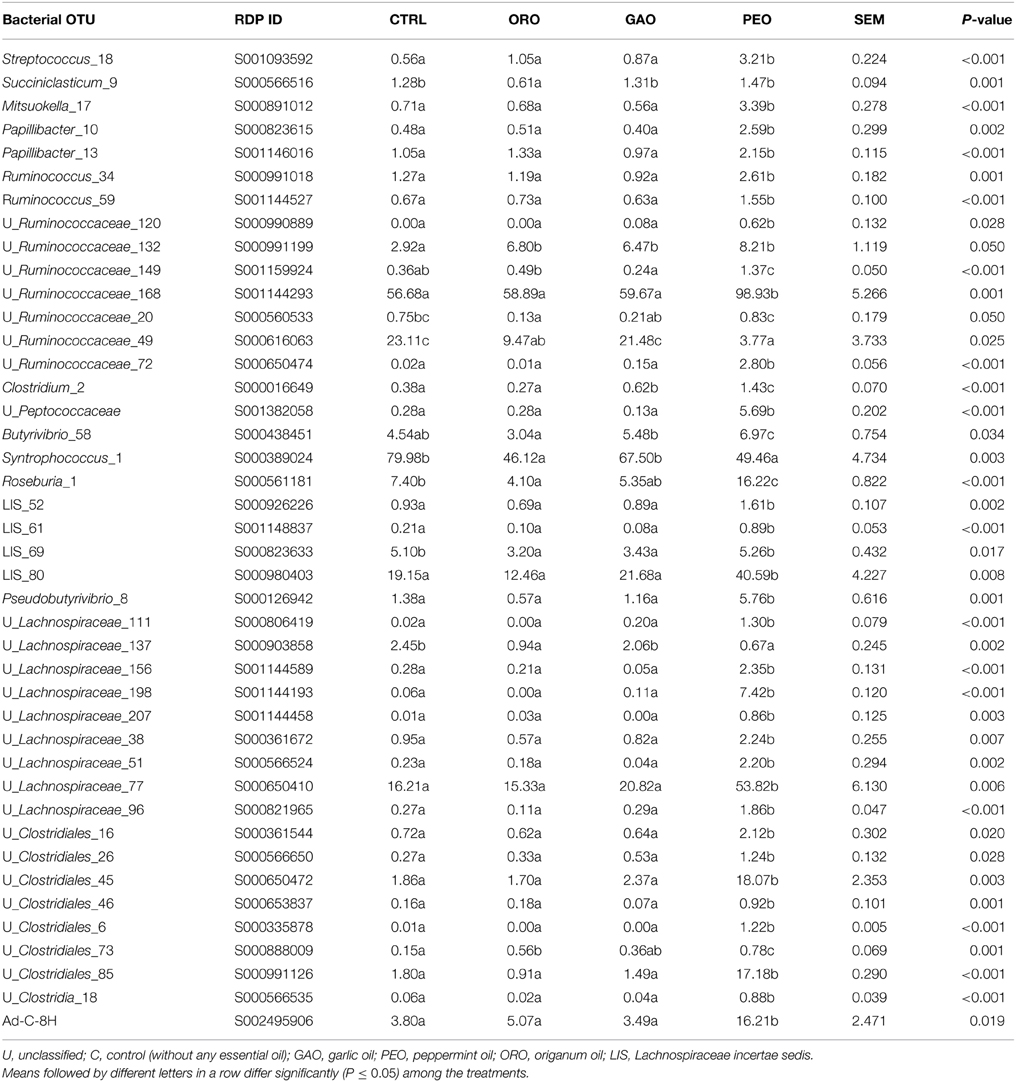
Table 2. Effects of essential oils on populations of the ruminal bacteria in the phylum Firmicutes [only the OTUs with significant (P ≤ 0.05) changes in relative abundance are shown].
Phylum Bacteroidetes
Twenty-four OTUs in the phylum Bacteroidetes showed a significant difference (Table 3) and 15 OTUs tended (0.05 < P ≤ 0.10) to differ (Table S2) in relative abundance between the EO treatments and the control, with 25 of them being members of the family Prevotellaceae (Table 3). All the OTUs affected by ORO showed increases in relative abundances compared with the control. These OTUs included Prevotella_18, _142, and_143 unclassified Prevotellaceae_34, and unclassified Bacteroidales_55, and _63. The GAO treatment did not significantly increase the relative abundance of any OTUs. Numerous OTUs had increased relative abundance in response to the addition of PEO, including OTUs in the families Porphyromonadaceae (Parabacteroides_1, unclassified Porphyromonadaceae_16, and _33) and Prevotellaceae (Hallella_14, Prevotella_2, _26, _68, _74, _115, _178, and _195, and unclassified Prevotellaceae_31), and OTUs remain unclassified in the order Bacteroidales (unclassified_Bacteroidales_18, _25, _55, _61, _75, _76, _170 and _217).
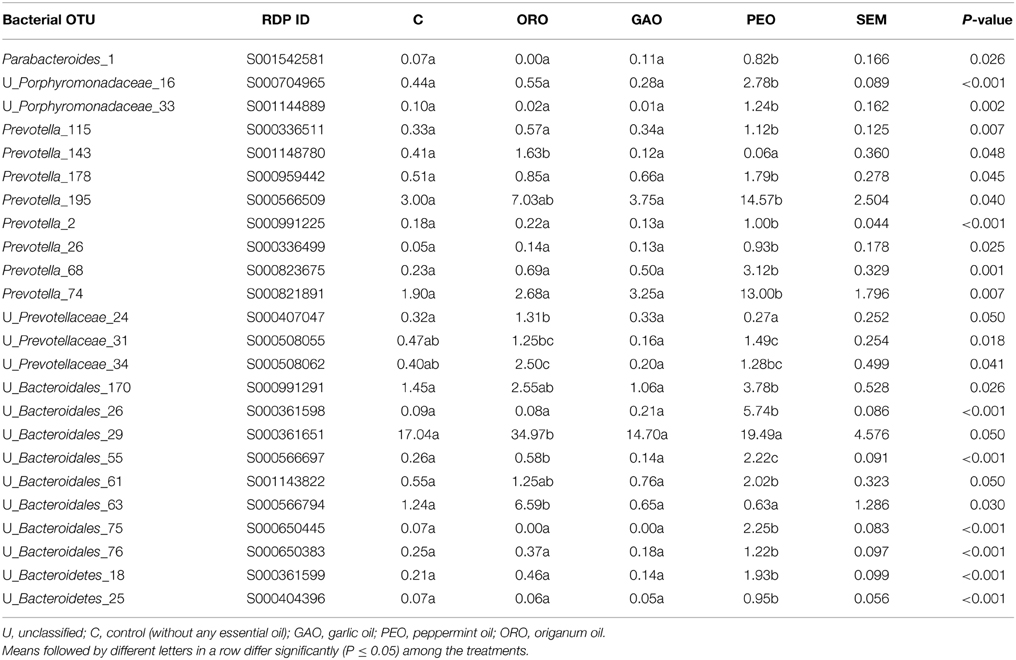
Table 3. Effects of essential oils on populations of ruminal bacteria in the phylum Bacteroidetes and Spirochaetes [only the OTUs with significant (P ≤ 0.05) changes in relative abundance are shown].
Phylum Spirochaetes
Only three OTUs in the phylum Spirochaetes showed a tendency to change in relative abundance in response to the EO treatments (Table S2).
Overall Effect on Bacterial Communities
Much of the variance (83%) was explained by the first three principal components (Figure 2). As shown along the PC2 axis, the PEO treatment resulted in a more distinct ruminal bacterial community than the control and the other EO treatments. The other two EOs, however, did not result in distinct bacterial communities compared to the control. Variations among the three replicates of each treatment were primarily displayed along the PC1 axis. The MANOVA also showed that the bacterial community was not significantly affected by ORO (P = 0.17) or GAO (P = 0.15) when compared to that of the control, but was significantly affected by PEO (P = 0.001). Among the three EO treatments, the ORO and the GAO treatments did not have significantly different (P = 0.089) bacterial community, but the PEO treatment had different bacterial community than the ORO (P = 0.036) and the GAO (P < 0.001) treatments (data not shown).
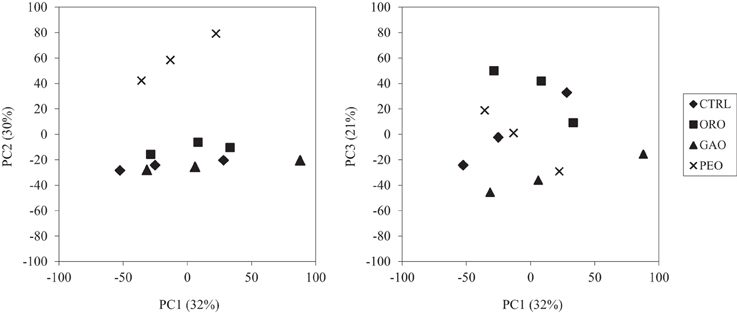
Figure 2. PCA plots of the bacterial OTUs detected by the RumenBactArray among the in vitro ruminal cultures. CTRL, control; ORO, origanum oil; GAO, garlic oil; PEO, peppermint oil. All the treatments were in triplicates.
Correlation between OTUs and Rumen Fermentation Characteristics
As determined by Pearson correlation coefficients (Table 4), 36 OTUs were negatively and 9 OTUs were positively correlated with DMD; 17 OTUs were negatively and 6 OTUs were positively correlated with ammonia concentrations; 3 OTUs were negatively and 29 OTUs were positively correlated with acetate concentrations; 39 OTUs were negatively and 5 were positively correlated with propionate concentration; and 5 OTUs were positively and 37 were positively correlated with butyrate concentrations. Methane production correlated positively with 43 OTUs and negatively with only one OTU, i.e., Syntrophococcus_9 (Figure 3). Only four OTUs (i.e., Syntrophococcus_1, Syntrophococcus_9, Succiniclasticum_9, and Lachnospiraceae incertae sedis_69) that correlated with methane production were associated with VFA profile and DM digestion.
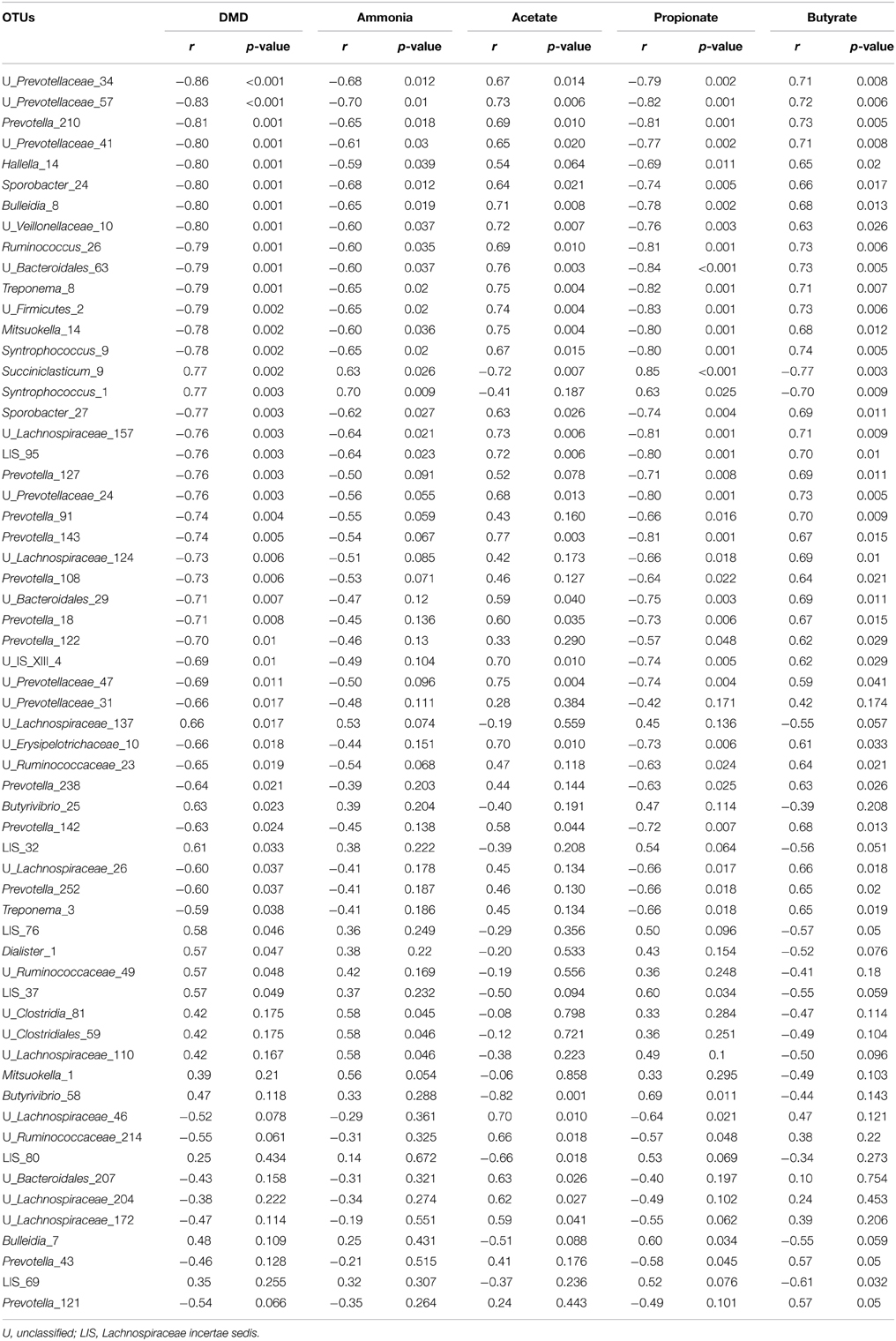
Table 4. Significant (P ≤ 0.05) Pearson correlation coefficients (r) between relative abundances of operational taxonomic units (OTUs) and some of the rumen fermentation characteristics.

Figure 3. Significant (P ≤ 0.05) Pearson correlation coefficients (r) between methane production and the relative abundances of OTUs.
Discussion
The rumen bacterial community is extremely diverse, especially at the species and genus levels, collectively containing over 5200 OTUs at species level and 3500 OTUs at genus levels (Kim et al., 2011; Creevey et al., 2014). In most studies that examined the effects of dietary interventions, including mitigation of methane emission by anti-methanogenic compounds or substances, only a few small groups of bacteria were analyzed (Patra and Yu, 2012; Fiorentini et al., 2013; Martínez-Fernández et al., 2014). The narrow scopes of these studies prevent full revelation of the actual impact of the feed additives on the rumen bacterial communities and the effect on the interactions among different bacteria. As a result, it is often difficult to understand the mode(s) of action or explain the observed efficacy of the feed additives. The rumen bacterial populations are highly dynamic in how they respond to changes in diet, feed additives, feeding regiments, and physiological status of the ruminants (Wang et al., 2009; Li et al., 2012; Wu et al., 2012). Most of these effectors typically cause fluctuations of bacterial populations, rather than completely eliminate bacterial populations or bring about the emergence of new bacterial populations. In the present study, RumenBactArray was used to comparatively examine the impact of EOs on rumen bacteria in a semi-quantitative manner.
The EOs evaluated affected the rumen bacterial community composition differently at species level depending upon the EO types. A number of mechanisms of action have been proposed to explain the antimicrobial properties of EOs, with chemical structures and physical properties being most important to determine their antimicrobial potency (Dorman and Deans, 2000; Burt, 2004). The presence of phenolic structure and the position of a hydroxyl group in the phenolic structure of EOs (e.g., EOs containing thymol or eugenol) can influence the antimicrobial potency of EOs (Dorman and Deans, 2000; Ultee et al., 2002). The greater antibacterial potency of ORO (containing a phenol) than PEO (containing a cyclohexane) shown in this study (Table 2) corroborates the importance of the phenolic ring to the antimicrobial activities of EOs (Ultee et al., 2002). Gram-negative bacteria are usually thought to be less susceptible to EOs than Gram-positive bacteria due to the presence of a protecting outer membrane (Dorman and Deans, 2000; Burt, 2004). Members of Prevotella, a Gram-negative genus, increased their populations in response to the addition of ORO and PEO, while GAO did not affect the populations of Prevotella. On the other hand, members of the Firmicutes, a largely Gram-positive phylum, were decreased by all the three EOs, and bacterial groups in the class Clostridia, which contains most of the Gram-positive rumen bacteria, were decreased by ORO and GAO, but not by PEO. Evidently, effects of EOs on rumen bacteria are both species and EO type dependent.
Addition of a few EOs to diets fed to ruminants has been shown to decrease degradation of starch and protein, improve rumen fermentation, and inhibit methanogenesis and biohydrogenation of polyunsaturated fatty acids in the rumen (McIntosh et al., 2003; Calsamiglia et al., 2007; Patra, 2011), which was considered to be a consequence of modification of microbial populations in the rumen. The three EOs evaluated in this study decreased the number of members in the genus Butyrivibrio. Butyrivibrio fibrisolvens, a major cultured species of butyrate-producing Gram-positive bacteria ubiquitous in the rumen, was found to be very sensitive to a blend of EOs (McIntosh et al., 2003). The population of this species was inhibited by ORO, but not by PEO at the similar dose level (Patra and Yu, 2014). Zhu et al. (2014) also reported decreased 16S rRNA gene clones related to Butyrivibrio proteoclasticus and Pseudobutyribrio ruminis in the rumen of goats fed 0.8 g/d of GAO. In the present study, the ORO supplementation decreased the relative abundances of many OTUs of S. sucromutans, Lachnospiraceae incertae sedis, and unclassified Ruminococcaceae, but increased that of S. ruminis, and some members of Prevotella, and unclassified Bacteroidales, Lachnospiraceae, and Prevotellaceae. Butyrivibrio, Anaerovoax (member of the Lachnospiraceae incertae sedis family), and unclassified Clostridiales and Ruminococcaceae have been suggested to have a key role in rumen biohydrogenation (Huws et al., 2011). Thus, modification of these microbial compositions by EOs may be associated with changes in the rumen biohydrogenation process (Lourenço et al., 2008; Ramos-Morales et al., 2013) and the increased concentrations of conjugated linoleic acid (CLA) and other poly unsaturated fatty acids in milk and tissues of ruminants fed EOs (Morsy et al., 2012; Mandal et al., 2014).
Supplementation of ORO and PEO increased while GAO did not affect the predominant representatives of Prevotella in the present study. The relative abundances of several Prevotella OTUs increased in response to EOs probably due to reduced competition from other bacteria that were inhibited by EOs. The genus Prevotella genus is present in the rumen across a variety of diets and exhibits substantial metabolic diversity (Petri et al., 2013). This genus comprises species that appear to be involved in protein degradation in the rumen (Wallace et al., 1997). It was suggested that EOs can decrease protein degradation and ammonia concentrations in the rumen (McIntosh et al., 2003; Patra, 2011). Indeed, addition of ORO significantly lowered ammonia concentrations, while PEO numerically reduced ammonia concentration at the dose level of 0.50 g/L in the mixed in vitro rumen cultures (Patra and Yu, 2014). The reduced ammonia production was also associated with reduced abundances of the major protein-degrading and amino acid-fermenting bacteria (Patra and Yu, 2014). Thus, it appears that few members of Prevotella or other OTUs which were not detected in this study may play a major role in protein metabolism in the rumen.
Inclusion of PEO increased relative abundances of several unclassified bacterial species of Ruminococcaceae and Lachnospiraceae. Zhu et al. (2014) also noted that some bacterial 16S rRNA genes classified to the genus Ruminococcus and other genera within the family Ruminococcaceae were specifically found in the GAO group. The number of OTUs of the unclassified Ruminococcaceae was lower in all the EO treatments than in the control. In our earlier study (Patra and Yu, 2012), ORO and PEO decreased the abundance of R. flavefaciens and R. albus. The members of Ruminococcaceae contribute significantly to fiber metabolism (Koike and Kobayashi, 2009), and thus EOs at high doses may decrease fiber digestion (Patra and Yu, 2012). In a previous study, DGGE-based analysis has shown that PEO and ORO resulted in bacterial communities that were distinctly different from that of control at the dose selected in this study (Patra and Yu, 2012). However, in the present study, the bacterial communities were only different between the PEO treatment and the control. This discrepancy was likely due to techniques used for community analysis. Much fewer bacterial groups can be detected by DGGE-based analysis than by microarray.
Addition of EOs changed the abundances of a few less-known bacteria. The relative abundance of Syntrophococcus was considerably decreased by ORO and PEO. The role of S. sucromutans in the rumen metabolism is not well-understood, but it produces acetate only from pyruvate and various carbohydrates (Krumholtz and Bryant, 1986). All three EOs also lowered the abundances of the genus Succinivibrio. The representatives of this genus increased when ruminants were fed high levels of grains or rapidly degradable carbohydrate such as diets rich in starch (Patterson and Hespell, 1985; O'Herrin and Kenealy, 1993). Succinivibrio dextrinosolvens may be one of the major rumen bacteria fermenting dextrin and levans in starch-based diets (Patterson and Hespell, 1985). Because all the three EOs were inhibitory to Succinivibrio, the effect of EOs on starch metabolism in the rumen may be attributed to their effect on this group of bacteria. The relative abundance of Acetanaerobacterium, a genus of Gram-positive bacteria, was generally lower in the EO treatments than in the control. The role of this bacterial genus in rumen metabolism is poorly understood except fermenting sugars to acetate from sugars (Chen and Dong, 2004). One OTU of the genus Actinomyces was detected in the control, while it was not detected in any of the EO treatments. Actinomyces ruminicola hydrolyzes xylan and starch and ferment several kinds of mono-, di-, and oligosaccharides (An et al., 2006). Much of the basic understandings on the rumen microbiome and the specific roles of microorganisms in rumen metabolism were obtained from studies on individual bacteria isolated in pure cultures (Petri et al., 2013). However, the majority of the ruminal microorganisms remain uncultured (Kim et al., 2011; Creevey et al., 2014), and thus their specific roles in the overall rumen fermentation remain to be elucidated. A large amount of the rumen bacteria will remain to be uncultured in the foreseeable future. As shown in the present study, the RumenBactArray can help in establishing associations between bacterial abundance and fermentation variables in the rumen in repeated studies.
Feed degradability and VFA profiles in the rumen are some of the most important parameters indicative of bacterial metabolism. Thus, correlation coefficients were determined to investigate association between OTUs and major rumen fermentation characteristics. Overall 60 OTUs were associated with the changes in DMD, ammonia concentration, and VFA profiles, which suggest that the dynamic changes of these OTUs in response to the EO treatments are probably responsible for or caused by the shifts in these fermentation characteristics. Correlation between specific bacteria and VFA profiles in the rumen has been scarcely reported, with a positive correlation being noted between abundance of genus Butyrivibrio and proportion of butyrate (Mohammed et al., 2014), but significant correlations were noted between VFA concentrations and several bacterial community in the feces of cattle (Mao et al., 2012). Future studies using quantitative tools, such as the RumenBactArray, may provide opportunities to determine the bacteria that are associated with some of the important fermentation characteristics in the rumen. In this study, 17 OTUs were found to have negative correlation with DMD, concentration of ammonia, and molar percentage of propionate, but positive correlation with molar percentages of acetate and butyrate, irrespective of EO treatments. Only four OTUs that were associated with VFA characteristics correlated with methane production. As noted by Mao et al. (2012), changes in bacterial populations and VFA profile depend on many other variables such as competition among bacteria for substrates, synthesis of antimicrobial agents, and bacterial metabolism. Future studies are needed to confirm these correlations and to determine their causality. Although not confirmatory, the results of the present study also suggest that a relatively small group of rumen bacteria may be responsible for and/or indicative of each of the important characteristics of feed digestion and fermentation. The members of these groups can be identified in future studies (both in vitro and in vivo) in which each of these feed digestion and fermentation characteristics is changed intentionally through dietary interventions or by addition of those particular bacterial groups in vitro or in vivo. Conceivably, once confirmed, these responsible or indicator groups of bacteria, rather than the entire rumen microbiome, can be specifically analyzed more effectively to support nutritional studies of ruminant animals.
The in vitro rumen fermentation technique is useful to assess digestibility, rumen fermentation characteristics, and microbial community structure influenced by feeds and feed additives, which is easy to conduct and inexpensive compared with in vivo measurements. However, the extrapolation of the results of in vitro studies to in vivo conditions may be sometimes unrepresentative due to continuous absorption of VFA, neutralization by saliva, changes in the microbial ecosystem such as a decrease in total microbial biomass and shifts in bacterial community composition, and low number of fungi and protozoa in vitro (Soto et al., 2012). In addition, different in vitro conditions can have different effects on certain rumen microbial populations (Weimer et al., 2011). Besides, the number of replicate samples for statistical analyses was low, especially for a microbial ecology study. Thus, the results obtained in this in vitro study might have limitations, and should be interpreted accordingly.
In conclusion, this study demonstrated for the first time using microarray analysis that EOs can affect the population dynamics of a number of bacteria, especially those in the families Prevotellaceae, Butyrivibrio, Lachnospiraceae, and Ruminococcaceae, in an EO type-dependent manner. Many bacterial OTUs were found to be associated with changes in feed digestibility and rumen fermentation characteristics, which may explain the modulation of rumen fermentation due to feed additives.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported in part by an OARDC grant (2010-007). AP's tenure at The Ohio State University was supported by a BOYSCAST fellowship from the Department of Science and Technology, India.
Supplementary Material
The Supplementary Material for this article can be found online at: http://www.frontiersin.org/journal/10.3389/fmicb.2015.00297/abstract
References
Adamczyk, J., Hesselsoe, M., Iversen, N., Horn, M., Lehner, A., Nielsen, P. H., et al. (2003). The isotope array, a new tool that employs substrate-mediated labeling of rRNA for determination of microbial community structure and function. Appl. Environ. Microbiol. 69, 6875–6887. doi: 10.1128/AEM.69.11.6875-6887.2003
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
An, D., Cai, S., and Dong, X. (2006). Actinomyces ruminicola sp. nov., isolated from cattle rumen. Int. J. Syst. Evol. Microbiol. 56, 2043–2048. doi: 10.1099/ijs.0.64059-0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Burt, S. (2004). Essential oils: their antibacterial properties and potential applications in foods-a review. Int. J. Food Microbiol. 94, 223–253. doi: 10.1016/j.ijfoodmicro.2004.03.022
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Calsamiglia, S., Busquet, M., Cardozo, P. W., Castillejos, L., and Ferret, A. (2007). Invited review: essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 90, 2580–2595. doi: 10.3168/jds.2006-644
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Castiglioni, B., Rizzi, E., Frosini, A., Sivonen, K., Rajaniemi, P., Rantala, A., et al. (2004). Development of a universal microarray based on the ligation detection reaction and 16S rRNA gene polymorphism to target diversity of cyanobacteria. Appl. Environ. Microbiol. 70, 7161–7172. doi: 10.1128/AEM.70.12.7161-7172.2004
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chen, S., and Dong, X. (2004). Acetanaerobacterium elongatum gen. nov., sp. nov., from paper mill waste water. Int. J. Syst. Evol. Microbiol. 54, 2257–2262. doi: 10.1099/ijs.0.63212-0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Creevey, C. J., Kelly, W. J., Henderson, G., and Leahy, S. C. (2014). Determining the culturability of the rumen bacterial microbiome. Microb. Biotechnol. 7, 467–479. doi: 10.1111/1751-7915.12141
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dorman, H. J. D., and Deans, S. G. (2000). Antimicrobial agents from plants: antibacterial activity of plant volatile oils. J. Appl. Microbiol. 88, 308–316. doi: 10.1046/j.1365-2672.2000.00969.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fiorentini, G., Messana, J. D., Dian, P. H. M., Reis, R. A., Canesin, R. C., Pires, A. V., et al. (2013). Digestibility, fermentation and rumen microbiota of crossbred heifers fed diets with different soybean oil availabilities in the rumen. Anim. Feed Sci. Technol. 181, 26–34. doi: 10.1016/j.anifeedsci.2013.01.011
Giannenas, I., Skoufos, J., Giannakopoulos, C., Wiemann, M., Gortzi, O., Lalas, S., et al. (2011). Effects of essential oils on milk production, milk composition, and rumen microbiota in Chios dairy ewes. J. Dairy Sci. 94, 5569–5577. doi: 10.3168/jds.2010-4096
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Huws, S. A., Kim, E. J., Lee, M. R., Scott, M. B., Tweed, J. K., Pinloche, E., et al. (2011). As yet uncultured bacteria phylogenetically classified as Prevotella, Lachnospiraceae incertae sedis and unclassified Bacteroidales, Clostridiales and Ruminococcaceae may play a predominant role in ruminal biohydrogenation. Environ. Microbiol. 13, 1500–1512. doi: 10.1111/j.1462-2920.2011.02452.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kang, S., Denman, S. E., Morrison, M., Yu, Z., Dore, J., Leclerc, M., et al. (2010). Dysbiosis of fecal microbiota in Crohn's disease patients as revealed by a custom phylogenetic microarray. Inflamm. Bowel Dis. 16, 2034–2042. doi: 10.1002/ibd.21319
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kim, M., Morrison, M., and Yu, Z. (2011). Status of the phylogenetic diversity census of ruminal microbiomes. FEMS Microbiol. Ecol. 76, 49–63. doi: 10.1111/j.1574-6941.2010.01029.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kim, M., Wang, L., Morrison, M., and Yu, Z. (2014). Development of a phylogenetic microarray for comprehensive analysis of ruminal bacterial communities. J. Appl. Microbiol. 117, 949–960. doi: 10.1111/jam.12598
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Koike, S., and Kobayashi, Y. (2009). Fibrolytic rumen bacteria: their ecology and functions. Asian Aust. J. Anim. Sci. 22, 131–138. doi: 10.5713/ajas.2009.r.01
Krumholtz, L. R., and Bryant, M. P. (1986). Syntrophococcus sucromutans sp. nov.gen. nov. uses carbohydrates as electron donors and formate, methoxymonobenzenoids or methanobrevibacter as electron acceptor systems. Archiv. Microbiol. 143, 313–318. doi: 10.1007/BF00412795
Kung, L. Jr., Williams, P., Schmidt, R. J., and Hu, W. (2008). A blend of essential plant oils used as an additive to alter silage fermentation or used as a feed additive for lactating dairy cows. J. Dairy Sci. 91, 4793–4800. doi: 10.3168/jds.2008-1402
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, R. W., Connor, E. E., Li, C., Baldwin Vi, R. L., and Sparks, M. E. (2012). Characterization of the rumen microbiota of pre-ruminant calves using metagenomic tools. Environ. Microbiol. 14, 129–139. doi: 10.1111/j.1462-2920.2011.02543.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lourenço, M., Cardozo, P. W., Calsamiglia, S., and Fievez, V. (2008). Effects of saponins, quercetin, eugenol, and cinnamaldehyde on fatty acid biohydrogenation of forage polyunsaturated fatty acids in dual-flow continuous culture fermenters. J. Anim. Sci. 86, 3045–3053. doi: 10.2527/jas.2007-0708
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Macheboeuf, D., Morgavi, D. P., Papon, Y., Mousset, J. L., and Arturo-Schaan, M. (2008). Dose-response effects of essential oils on in vitro fermentation activity of the rumen microbial population. Anim. Feed Sci. Technol. 145, 335–350. doi: 10.1016/j.anifeedsci.2007.05.044
Mandal, G. P., Roy, A., and Patra, A. K. (2014). Effects of feeding plant additives rich in saponins and essential oils on the performance, carcass traits and conjugated linoleic acid concentrations in muscle and adipose tissues of Black Bengal goats. Anim. Feed Sci. Technol. 197, 76–84. doi: 10.1016/j.anifeedsci.2014.08.008
Mao, S., Zhang, R., Wand, D., and Zhu, W. (2012). The diversity of the fecal bacterial community and its relationship with the concentration of volatile fatty acids in the feces during subacute rumen acidosis in dairy cows. BMC Vet. Res. 8:237. doi: 10.1186/1746-6148-8-237
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Martínez-Fernández, G., Abecia, L., Ramos-Morales, E., Martin-García, A. I., Molina-Alcaide, E., and Yáñez-Ruiz, D. R. (2014). Effects of propyl propane thiosulfinate on nutrient utilization, ruminal fermentation, microbial population and methane emissions in goats. Anim. Feed Sci. Technol. 191, 16–25. doi: 10.1016/j.anifeedsci.2014.01.008
McIntosh, F. M., Williams, P., Losa, R., Wallace, R. J., Beever, D. A., and Newbold, C. J. (2003). Effects of essential oils on ruminal microorganisms and their protein metabolism. Appl. Environ. Microbiol. 69, 5011–5014. doi: 10.1128/AEM.69.8.5011-5014.2003
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mohammed, R., Brink, G. E., Stevenson, D. M., Neumann, A. P., Beauchemin, K. A., Suen, G., et al. (2014). Bacterial communities in the rumen of Holstein heifers differ when fed orchardgrass as pasture vs. hay. Front. Microbiol. 5:689. doi: 10.3389/fmicb.2014.00689
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Morsy, T. A., Kholif, S. M., Matloup, O. H., Abdo, M. M., and El-Shafie, M. H. (2012). Impact of anise, clove and juniper oils as feed additives on the productive performance of lactating goats. Int. J. Dairy Sci. 7, 20–28. doi: 10.3923/ijds.2012.20.28
O'Herrin, S. M., and Kenealy, W. R. (1993). Glucose and carbon dioxide metabolism by Succinivibrio dextrinosolvens. Appl. Environ. Microbiol. 59, 748–755.
Palmer, C., Bik, E. M., Eisen, M. B., Eckburg, P. B., Sana, T. R., Wolber, P. K., et al. (2006). Rapid quantitative profiling of complex microbial populations. Nucleic Acids Res. 34:e5. doi: 10.1093/nar/gnj007
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Patra, A. K. (2011). Effects of essential oils on rumen fermentation, microbial ecology and ruminant production. Asian J. Anim. Vet. Adv. 6, 416–428. doi: 10.3923/ajava.2011.416.428
Patra, A. K., and Saxena, J. (2009). Dietary phytochemicals as rumen modifiers: a review of the effects on microbial populations. Anton van Leeuwenhoek 96, 363–375. doi: 10.1007/s10482-009-9364-1
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Patra, A. K., and Saxena, J. (2010). A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. Phytochemistry 71, 1198–1222. doi: 10.1016/j.phytochem.2010.05.010
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Patra, A. K., and Yu, Z. (2012). Effect of essential oil on methane production and fermentation by, and abundance and diversity of, rumen microbial populations. Appl. Environ. Microbiol. 78, 4271–4280. doi: 10.1128/AEM.00309-12
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Patra, A. K., and Yu, Z. (2014). Effects of vanillin, quillaja saponin, and essential oils on in vitro fermentation and protein-degrading microorganisms of the rumen. Appl. Microbiol. Biotechnol. 98, 897–905. doi: 10.1007/s00253-013-4930-x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Patterson, J. A., and Hespell, R. B. (1985). Glutamine synthetase activity in the ruminal bacterium Succinivibrio dextrinosolvens. Appl. Environ. Microbiol. 50, 1014–1020.
Petri, R. M., Schwaiger, T., Penner, G. B., Beauchemin, K. A., Forster, R. J., McKinnon, J. J., et al. (2013). Characterization of the core rumen microbiome in cattle during transition from forage to concentrate as well as during and after an acidotic challenge. PLoS ONE 8:e83424. doi: 10.1371/journal.pone.0083424
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rajilic-Stojanovic, M., Heilig, H. G., Molenaar, D., Kajander, K., Surakka, A., Smidt, H., et al. (2009). Development and application of the human intestinal tract chip, a phylogenetic microarray: analysis of universally conserved phylotypes in the abundant microbiota of young and elderly adults. Environ. Microbiol. 11, 1736–1751. doi: 10.1111/j.1462-2920.2009.01900.x
Ramos-Morales, E., Martínez-Fernández, G., Abecia, L., Martin-García, A. I., Molina-Alcaide, E., and Yáñez-Ruiz, D. R. (2013). Garlic derived compounds modify ruminal fatty acid biohydrogenation and induce shifts in the Butyrivibrio community in continuous-culture fermenters. Anim. Feed Sci. Technol. 184, 38–48. doi: 10.1016/j.anifeedsci.2013.05.017
Saeed, A. I., Bhagabati, N. K., Braisted, J. C., Liang, W., Sharov, V., Howe, E. A., et al. (2006). TM4 microarray software suite. Methods Enzymol. 411, 134–193. doi: 10.1016/S0076-6879(06)11009-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schatz, M. C., Phillippy, A. M., Gajer, P., Desantis, T. Z., Andersen, G. L., and Ravel, J. (2010). Integrated microbial survey analysis of prokaryotic communities for the phylochip microarray. Appl. Environ. Microbiol. 76, 5636–5638. doi: 10.1128/AEM.00303-10
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Small, J., Call, D. R., Brockman, F. J., Straub, T. M., and Chandler, D. P. (2001). Direct detection of 16S rRNA in soil extracts by using oligonucleotide microarrays. Appl. Environ. Microbiol. 67, 4708–4716. doi: 10.1128/AEM.67.10.4708-4716.2001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Soto, E., Yáñez-Ruiz, D., Cantalapiedra-Hijar, G., Vivas, A., and Molina-Alcaide, E. (2012). Changes in ruminal microbiota due to rumen content processing and incubation in single-flow continuous-culture fermenters. Anim. Prod. Sci. 52, 813–822. doi: 10.1071/AN11312
Tassoul, M. D., and Shaver, R. D. (2009). Effect of a mixture of supplemental dietary plant essential oils on performance of periparturient and early lactation dairy cows. J. Dairy Sci. 92, 1734–1740. doi: 10.3168/jds.2008-1760
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ultee, A., Bennink, M. H. J., and Moezelaar, R. (2002). The phenolic hydroxyl group of carvacrol is essential for action against the food-borne pathogen Bacillus cereus. Appl. Environ. Microbiol. 68, 1561–1568. doi: 10.1128/AEM.68.4.1561-1568.2002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wallace, R. J., Onodera, R., and Cotta, M. A. (1997). “Metabolism of nitrogen-containing compounds,” in The Rumen Microbial Ecosystem, eds P. N. Hobson and C. S. Stewart (London: Blackie Academic & Professional), 246–282. doi: 10.1007/978-94-009-1453-7_7
Wang, C. J., Wang, S. P., and Zhou, H. (2009). Influences of flavomycin, ropadiar, and saponin on nutrient digestibility, rumen fermentation, and methane emission from sheep. Anim. Feed Sci. Technol. 148, 157–166. doi: 10.1016/j.anifeedsci.2008.03.008
Weimer, P. J., Stevenson, D. M., Mertens, D. R., and Hall, M. B. (2011). Fiber digestion, VFA production, and microbial population changes during in vitro ruminal fermentations of mixed rations by monensin-adapted and unadapted microbes. Anim. Feed Sci. Technol. 169, 68–78. doi: 10.1016/j.anifeedsci.2011.06.002
Wu, S., Baldwin, R. L. VI, Li, W., Li, C., Connor, E. E., and Li, R. W. (2012). The bacterial community composition of the bovine rumen detected using pyrosequencing of 16S rRNAgenes. Metagenomics 1:235571. doi: 10.4303/mg/235571
Zhu, Z., Hang, S., Mao, S., and Zhu, W. (2014). Diversity of Butyrivibrio group bacteria in the rumen of goats and its response to the supplementation of garlic oil. Asian-Aust. J. Anim. Sci. 27, 179–186. doi: 10.5713/ajas.2013.13373
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: essential oil, bacterial composition, microarray, rumen, RumenBactArray
Citation: Patra AK and Yu Z (2015) Essential oils affect populations of some rumen bacteria in vitro as revealed by microarray (RumenBactArray) analysis. Front. Microbiol. 6:297. doi: 10.3389/fmicb.2015.00297
Received: 08 December 2014; Accepted: 26 March 2015;
Published: 10 April 2015.
Edited by:
Emilio M. Ungerfeld, Instituto de Investigaciones Agropecuarias - Carillanca, ChileReviewed by:
Carl James Yeoman, Montana State University, USARobin Anderson, United States Department of Agriculture/Agricultural Research Service, USA
David R. Yanez-Ruiz, Consejo Superior de Investigaciones Científicas, Spain
Copyright © 2015 Patra and Yu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amlan K. Patra, Department of Animal Nutrition, West Bengal University of Animal and Fishery Sciences, 37 K. B. Sarani, Belgachia, Kolkata 700037, IndiacGF0cmFfYW1sYW5AeWFob28uY29t;
Zhongtang Yu, Department of Animal Sciences, The Ohio State University, 2029 Fyffe Road, Columbus, OH 43210, USAeXUuMjI2QG9zdS5lZHU=