 Philipp Schuster1,2
Philipp Schuster1,2- 1Institute of Medical Microbiology and Hygiene, University of Regensburg, Regensburg, Germany
- 2Institute of Clinical and Molecular Virology, Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany
- 3Nexigen, Cologne, Germany
Classical and plasmacytoid dendritic cells (DC) play important roles in the defense against murine and human infections with herpes simplex virus (HSV). So far, CD8α expression has only been reported for murine DC. CD8α+ DC have prominent cross-presenting activities, which are enhanced by murine CD8α+ PDC. The human orthologue of murine CD8α+ DC, the CD141 (BDCA3)+ DC, mainly cross-present after TLR3 ligation. We report here the serendipitous finding that a subset of human PDC upregulates CD8α upon HSV-1 stimulation, as shown by gene array and flow cytometry analyses. CD8α, not CD8ß, was expressed upon exposure. Markers of activation, migration, and costimulation were upregulated on CD8α-expressing human PDC. In these cells, increased cytokine and chemokine levels were detected that enhance development and function of T, B, and NK cells, and recruit immature DC, monocytes, and Th1 cells, respectively. Altogether, human CD8α+ PDC exhibit a highly activated phenotype and appear to recruit other immune cells to the site of inflammation. Further studies will show whether CD8α-expressing PDC contribute to antigen cross-presentation, which may be important for immune defenses against HSV infections in vitro and in vivo.
Introduction
Since Ralph Steinman first described a new subset of cells characterized by tree-like processes in 1973 (Steinman and Cohn, 1973), knowledge about dendritic cells (DC) in mice and humans has grown exponentially. These cells were originally identified as important players in the defense against “foreign” pathogens, but it turns out that they are similarly crucial in initiating immune responses against tumor-associated antigens (Vacchelli et al., 2013). Immature DC engulf extracellular antigens, but in the absence of appropriate danger signals, they induce peripheral tolerance. Only after appropriate activation, DC release cytokines and chemokines, undergo a maturation process, and migrate toward secondary lymphatic tissues to induce cytotoxic responses by other immune cells (Palucka and Banchereau, 2012; Merad et al., 2013).
In this perspective paper, we will focus on the role of CD8α-expressing DC. CD8 serves as useful subset marker for murine DC, which are highly efficient in cross-presenting foreign, self, and—most likely—tumor-associated antigens, although evidence is lacking that CD8 expression plays any role in the development and function of these cells (Shortman and Heath, 2010). So far, CD8 expression on human DC has not been reported (Naik, 2008). However, we report here a serendipitous finding of CD8α expression on human plasmacytoid dendritic cells (PDC) after stimulation with herpes simplex virus type 1 (HSV-1), which characterizes a highly activated subset of PDC. We will discuss how the knowledge about CD8α-expressing murine DC may translate into functions of CD8α-expressing human PDC. For the background of this topic, the reader is referred to excellent review articles by respected colleagues (Villadangos and Young, 2008; Shortman and Heath, 2010; Joffre et al., 2012; Nierkens et al., 2013; Bedoui and Greyer, 2014; Boltjes and van Wijk, 2014).
Murine CD8α+ DC: Prominent Cross-presentation
In the murine system, several different DC populations exist: lymphoid-organ resident CD8α+ or CD8α– DC; migratory dermal CD103+ or CD103– DC, which migrate to lymphatic tissue; Langerhans cells in the skin; inflammatory DC, which develop from monocytes; and PDC (Vremec et al., 1992; Bursch et al., 2007; Ginhoux et al., 2007, 2009; Leon et al., 2007; Poulin et al., 2010; Joffre et al., 2012). CD8α is a marker for lymphoid tissue-resident DC, which make up roughly 20% of spleen DC and 70% of thymic DC, whereas only 0.2% of peripheral blood mononuclear cells are CD8α+ DC (Crowley et al., 1989; Vremec et al., 2000; Donnenberg et al., 2001; Henri et al., 2001; Shortman and Heath, 2010). These cells express a CD8αα homodimer rather than the CD8αß heterodimer on T cells (Vremec et al., 1992, 2000). Precursors of CD8α+ DC may lack CD8 expression (Martinez del Hoyo et al., 2002). Apart from the classical CD8α+ DC population, this molecule is expressed by murine PDC in the spleen (O’Keeffe et al., 2002) and other migratory DC after activation (Anjuere et al., 1999, 2000; Merad et al., 2000; Henri et al., 2001). Mice with a knock-out for interferon regulatory factor (IRF) 8 neither develop CD8α+ DC nor PDC (Schiavoni et al., 2002; Aliberti et al., 2003; Tsujimura et al., 2003), whereas Batf3-deficient mice are only deficient in CD8α+ DC (Hildner et al., 2008; Edelson et al., 2010).
Amongst other receptors, the murine CD8α+ DC subset expresses CD11c, CD24, CD36, Necl2, MHC-II, the integrin CD103, the lectins CD205, CLEC9A, CLEC12A, and langerin (CD207) (Shortman and Heath, 2010). CLEC9A and CD36 are both involved in recognizing late apoptotic or necrotic cells (Albert et al., 1998; Caminschi et al., 2008; Huysamen et al., 2008; Sancho et al., 2009). Murine CD8α+ DC also express TLR3 and TLR9 (Edwards et al., 2003), and respond to TLR stimulation with proinflammatory IL-12 secretion and at least some type I interferon production (Hochrein et al., 2001). Upon stimulation, CD8α+ DC upregulate costimulatory markers CD40, CD80, and CD86 as well as CD25, CD62L, and MHC-II (Wilson et al., 2003).
CD8α+ DC are most efficient in antigen cross-presentation, a process in which extracellular antigen is not presented on MHC-II to CD4+ T cells, but instead shunted to MHC-I with subsequent induction of CD8+ T cells. Cross-presentation occurs through the cytosolic or vacuolar pathway (Joffre et al., 2012). The former involves proteasomal degradation with subsequent transport of peptides into the endoplasmic reticulum via transporter associated with antigen processing 1 (TAP), whereas the latter is based on lysosomal proteolysis with subsequent loading of peptides onto MHC-I molecules (Joffre et al., 2012).
In this respect, CD8α+ DC, but not CD8α– DC, were shown to cross-prime using a TAP-dependent pathway (den Haan et al., 2000; Pooley et al., 2001; Schnorrer et al., 2006; Lin et al., 2008). CD8α+ DC have been reported to selectively engulf dying cells in vitro and in vivo and present on MHC-I via a proteasome-dependent pathway (Iyoda et al., 2002; Schulz and Reis e Sousa, 2002). In these cells, endosomal acidification is limited (Savina et al., 2009), which fosters limited antigen degradation and efficient transport of the antigen to the cytosol (Delamarre et al., 2005). Overexpression of MHC-I loading complexes (Dudziak et al., 2007) by CD8α+ DC and expression of chemokine receptor XCR1, whose ligand XCL1 is secreted by activated CD8+ T cells, contribute to antigen cross-presentation and differentiation of cytotoxic T cells (Dorner et al., 2009).
In HSV infections, CD8α+ DC are able to present viral antigens and prime naïve CD4+ and CD8+ T cells, which appears to be mediated by cross-presentation (Allan et al., 2003; Smith et al., 2003; Belz et al., 2004a,b; Wilson et al., 2006; Bedoui et al., 2009; Lee et al., 2009). It is still a matter of debate how the viral antigen is transported from peripheral infected tissue to the lymphoid-resident CD8α+ DC. In this process, mainly other (migratory) DC are reported to be involved (Zhao et al., 2003; Carbone et al., 2004; Allan et al., 2006; Bedoui et al., 2009; Jirmo et al., 2009). These migratory DC either capture viral antigens or are infected within the peripheral tissue, although reduced migratory capacity has been reported for HSV-infected DC (Jones et al., 2003; Eidsmo et al., 2009; Puttur et al., 2010). The transfer of viral antigen can occur via exosomes, gap junctions, or uptake of apoptotic material following death of migratory DC (Thery et al., 2009; Mazzini et al., 2014). Another option is “crossdressing”, i.e., the transfer of preformed MHC-I complexes loaded with peptides from infected cells to murine DC via secreted membrane vesicles or transfer of membrane fragments (trogocytosis) (Thery et al., 2009; Wakim and Bevan, 2011; Joffre et al., 2012).
Murine CD8α+ PDC: Cross-presentation Help
Murine PDC were identified in the spleen of mice (Asselin-Paturel et al., 2001; O’Keeffe et al., 2003). Amongst other surface receptors, they express Ly6C, B220, and CD11c. Upon stimulation, type I interferons—and to a minor extent IL-12—are induced, and costimulatory markers CD40, CD69, CD80, and CD86 are upregulated (Asselin-Paturel et al., 2001; O’Keeffe et al., 2002; Lund et al., 2003). Unstimulated murine PDC express CD8α only to a minor extent, while exposure to CpG or viruses enhances expression of this molecule (Nakano et al., 2001; O’Keeffe et al., 2002, 2003). When CD8α+ and CD8α– PDC were separated and subsequently stimulated, they did not differ in cytokine production (O’Keeffe et al., 2002).
A few publications report TAP-dependent cross-presentation of soluble and particulate antigen by murine PDC after TLR ligation (Shinohara et al., 2006; Mouries et al., 2008; Kool et al., 2011). The majority of authors, however, deny cross-presentation by murine PDC (Chung et al., 2005; Janssen et al., 2006; Sapoznikov et al., 2007; GeurtsvanKessel et al., 2008; Reboulet et al., 2010; Hennies et al., 2011). In vitro stimulation of murine PDC with HSV-1 or influenza allowed priming of CD8+ T cells (Belz et al., 2004a). In in vivo HSV-1 infections, however, PDC do not participate in active cross-presentation (Allan et al., 2003; Lee et al., 2009; Swiecki et al., 2013). Still, murine PDC appear to be important in enhancing cross-presentation by other DC. An explanation of this phenomenon could be that type I interferons increase cross-presentation by decreasing antigen degradation in endocytic compartments and stimulating the survival of CD8α+ DC (Diamond et al., 2011; Fuertes et al., 2011; Lorenzi et al., 2011). In this respect, depletion of murine PDC was reported to impair CTL-mediated HSV-1 eradication in a CD2-, CD40L-, and type I interferon-dependent manner (Yoneyama et al., 2005). Also in the lymphocytic choriomeningitis model, virus-induced type I interferons were required for cross-priming of CD8+ T cells (Le Bon et al., 2003). When PDC were depleted in CLEC4C-DTR mice, PDC proved to be important for inducing CD8+ T cell responses in systemic HSV-1 and HSV-2 infections (Swiecki et al., 2013). Further functions of PDC in murine HSV-1 and HSV-2 infections are reviewed in (Schuster et al., 2011).
Human Orthologue of CD8α+ DC: Cross-presentation Following Activation
The conventional human blood DC population consist of three subsets specifically expressing CD1c (BDCA1), CD16, or CD141 (BDCA3) (Dzionek et al., 2000; MacDonald et al., 2002). Evidence is accumulating that the CD11c+ CD141+ DC subset represents the human orthologue of murine CD8α+ DC. These cells can be detected in lymphatic tissues such as lymph nodes, tonsils, bone marrow, spleen, and also liver (Galibert et al., 2005; Lindstedt et al., 2005; Velasquez-Lopera et al., 2008; Bamboat et al., 2009; Poulin et al., 2010). Genome-wide expression analyses revealed a similar transcriptomal signature between CD141+ human DC and murine CD8α+ DC (Robbins et al., 2008). Both subsets express Necl2 (Galibert et al., 2005), CLEC9A (Caminschi et al., 2008; Huysamen et al., 2008; Sancho et al., 2009; Jongbloed et al., 2010; Schreibelt et al., 2012), TLR3 (Edwards et al., 2003; Lindstedt et al., 2005; Jongbloed et al., 2010), as well as CD207, Batf3, and IRF8 (Poulin et al., 2010). BDCA3+ DC also express the chemokine receptor XCR1 and respond to respective ligands (Bachem et al., 2010; Crozat et al., 2010). Similar to murine CD8α+ DC, human BDCA3+ DC respond to TLR3 ligation with production of lambda interferons (Lauterbach et al., 2010). In contrast to murine CD8α+ DC, human BDCA3+ DC do not express TLR9 (Jongbloed et al., 2010).
Lymphoid tissue-derived human BDCA3+ DC were shown to be at least equivalent to other human DC subsets in cross-presenting soluble or cell-associated antigens, even in the absence of activation (Segura et al., 2012, 2013). This process can be enhanced by stimulation with TLR3 ligands, inducing superior cross-presenting activity by blood-derived BDCA3+ DC with induction of CD8+ T cell responses (Poulin et al., 2007; Bachem et al., 2010; Crozat et al., 2010; Jongbloed et al., 2010). There is evidence that cross-presentation by myeloid DC plays a role in human herpes virus infections (Bosnjak et al., 2005), but the importance of BDCA3+ DC needs to be further clarified.
Human CD8α+ PDC: Cross-presentation Help Following Viral Activation?
In 1999, two independent groups identified human PDC as major producers of type I interferons in the blood (Cella et al., 1999; Siegal et al., 1999). Amongst other receptors, PDC express BDCA2 and BDCA4, MHC-II, the lymph node-homing receptors CD62L and CCR7 (CD197), and costimulatory molecules (CD40, CD80, CD86, CD270, CD274, CD275) (Cella et al., 2000; Dzionek et al., 2000; Ito et al., 2007; Jaehn et al., 2008; Donaghy et al., 2009; Schuster et al., 2010, 2011; Cabezon et al., 2011). PDC recognize single-stranded RNA and CpG molecules via TLR7 and TLR9, respectively (Kadowaki et al., 2001).
Whether human PDC can cross-present soluble or particulate antigens is still a matter of debate. Viral antigen derived from influenza, recombinant vaccinia, tick-borne encephalitis or human immunodeficiency type I virus infection was taken up into recycling endosomes, loaded onto MHC-I molecules, and presented to CD8+ T cells (Fonteneau et al., 2003a,b; Hoeffel et al., 2007; Di Pucchio et al., 2008; Lui et al., 2009; Mittag et al., 2011; Tel et al., 2012). In addition, antigen loaded on synthetic microparticles or soluble tumor-associated antigen was presented to CD8+ T cells by exposed PDC (Tel et al., 2010; Guillerme et al., 2013; Segura et al., 2013). In contrast, other groups report no or only minor cross-presenting capacities of human PDC (Schnurr et al., 2005; Bachem et al., 2010; Crozat et al., 2010).
An early report by Fitzgerald-Bocarsly described the “interferon-producing cells” as being important for the lysis of HSV-infected fibroblasts (Feldman et al., 1992). PDC infiltrate herpetic lesions in the genital tract and tightly colocalize with NK and T cells (Donaghy et al., 2009). HSV-stimulated human PDC induce migration of activated T and NK cells via chemokine secretion (Megjugorac et al., 2004), and contribute to the activation of NK cells via IFNα- and TNFα-dependent mechanism (Vogel et al., 2014). In addition, HSV-exposed PDC were shown to prime IL-10 and IFN-γ production by cytotoxic regulatory CD4+ T cells (Kadowaki et al., 2000; Kawamura et al., 2006).
So far, expression of CD8 on human PDC has not been reported. Since the expression of this molecule on the surface of human PDC may define new and yet unknown capacities of these cells, we investigated whether PDC might upregulate these molecules upon viral stimulation. Recently, we analyzed the expression profile of human PDC, which were purified from PBMC of six donors. After exposure to IL-3 or IL-3 plus UV-inactivated HSV-1 (HSVUV), RNA was extracted from these cells and hybridized to a Human Genome U133 Plus 2_0 Array (Affymetrix, Santa Clara, CA, USA) (Schuster et al., 2010). In these analyses, we focused on the expression and regulation of surface receptors on PDC. Notably, the signal for CD8α expression increased from 57.2 to 100.1, which was slightly above the arbitrary threshold of 95, reflecting the expression signal of TLR9. In contrast, three probe sets for CD8β remained below this threshold. These data suggested a potential expression of CD8α on PDC upon stimulation with HSVUV.
To corroborate these data, we isolated PDC from a total of 15 different donors, and investigated CD8α expression on these cells in independent experiments after exposure to IL-3 (n = 16), IL-3 plus HSVUV (n = 16), or IL-3 plus infectious HSV-1 (HSVINF, n = 6) for 40 h. Flow cytometry confirmed a distinct expression of CD8α on a subset of HSVUV- and HSVINF-exposed PDC (Figure 1A). After stimulation with HSVUV or HSVINF, the percentage of CD8α-expressing PDC was significantly higher compared to PDC within freshly isolated PBMC (n = 9) (p < 0.001 for HSVUV and p < 0.05 for HSVINF, unpaired t-test) and purified PDC that were cultivated in the presence of IL-3 only (p < 0.001 for HSVUV and p < 0.05 for HSVINF, paired t-test) (Figure 1A). CD8α expression was not different between HSVUV- and HSVINF-exposed PDC (p = 0.27, n.s.). When we stained in parallel for CD8α and CD8β expression, we confirmed expression of CD8α by flow cytometry, while CD8β was neither detected on PDC exposed to HSVUV (n = 4) nor HSVINF (n = 3) (Figure 1B). These data indicated that PDC did not express a heterodimeric CD8αβ receptor upon stimulation. In further analyses, we investigated the kinetics of CD8α expression (n = 4). After exposure to HSVUV, the percentage of CD8α-expressing cells increased by day 1, but reached significance by day 2 post stimulation, compared to PDC cultivated with IL-3 alone (n < 0.05, paired t-test). Expression of CD8β was not detected at any of the time points analyzed (n = 3) (Figure 1C).
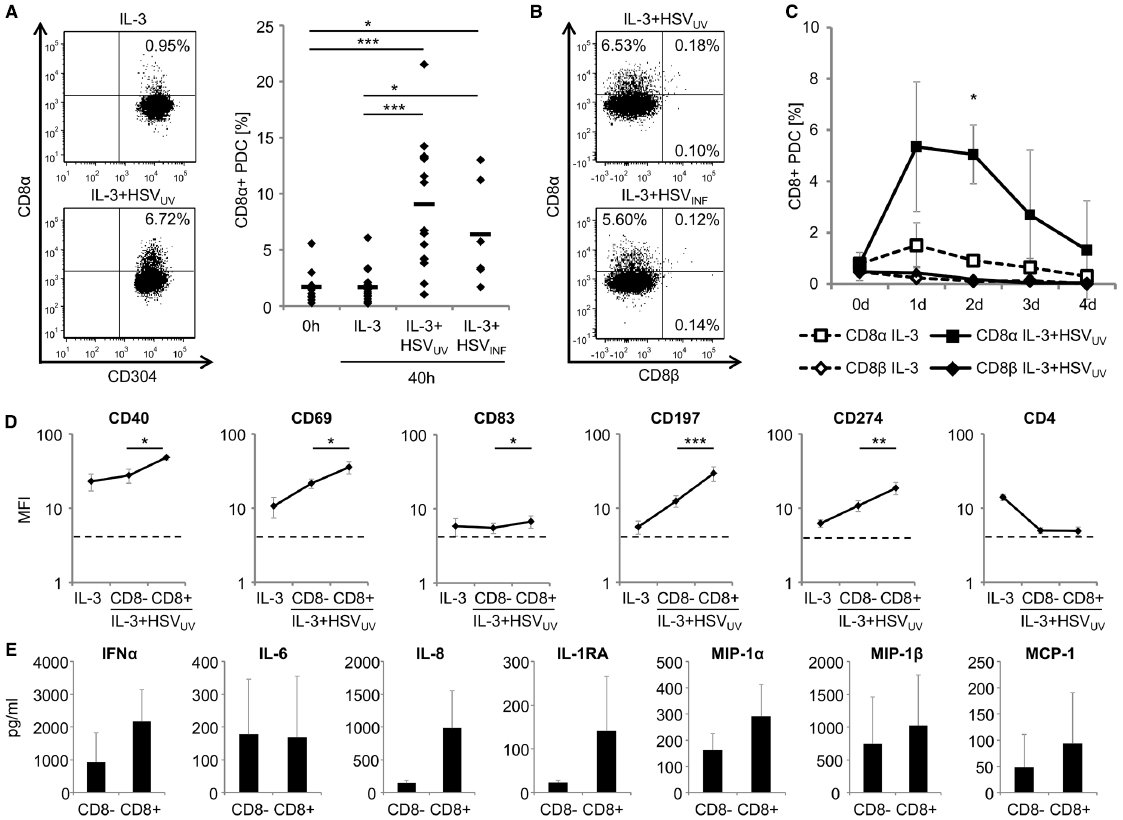
Figure 1. Characterization of CD8α-expressing human plasmacytoid dendritic cells (PDC). (A) Expression of CD8α on human PDC, as evaluated by flow cytometry within PBMC immediately after cell isolation (0 h) (n = 9) and after cultivation of purified PDC in the presence of IL-3 (10 ng/ml), IL-3 plus UV-inactivated herpes simplex virus type 1 (HSVUV) (n = 16) or infectious HSV-1 (HSVINF) (n = 6) (1 × 106 plaque-forming units/ml) for 40 h. One representative example of PDC exposed to IL-3 (upper left panel) and IL-3 plus HSVUV (lower left panel) and data of all donors including mean values (bars) are shown (right panel). (B) Representative expression of CD8α, not CD8β, on human PDC after exposure to IL-3 plus HSVUV (upper panel, n = 4) or HSVINF (lower panel, n = 3) for 40 h. (C) Kinetics of CD8α (n = 4) and CD8β (n = 3) expression after exposure of human PDC to IL-3 or IL-3 plus HSVUV for 4 days. Data are presented as mean and standard deviation. (D) Expression of markers for costimulation (CD40, CD274), activation (CD69), maturation (CD83), and migration (CD197) on CD8α+ and CD8α– human PDC after exposure to IL-3 plus HSVUV for 40 h. The dotted lines represent isotype controls. For statistical analysis, MFI values were transformed logarithmically to obtain normal distribution. Expression of CD4 as well as CD2, CD46, CD80, and CD123 (data not shown) was not significantly different between the two PDC subsets. Mean and standard deviation of four different donors (except for CD274, n = 3). (E) After stimulation of human PDC with IL-3 plus HSVUV for 40 h, cells were harvested and then separated using a CD8 cell isolation kit (Miltenyi Biotec, Bergisch-Gladbach, Germany). After stimulation with IL-3 or IL-3 plus HSVUV for another 20 h, different cytokines were analyzed in the cell culture supernatants using a multiplex cytokine and chemokine panel on a luminex platform (Invitrogen/Life Technologies, Darmstadt, Germany, and Affymetrix/ebioscience, Frankfurt, Germany). Mean and standard deviation of three different donors. *p < 0.05, **p < 0.01, ***p < 0.001.
To find out in how far CD8α+ and CD8α– PDC differed from each other, we analyzed the expression of cell surface markers for costimulation (CD40, CD274), activation (CD69), maturation (CD83), and migration (CD197) on these two subsets. All these markers were significantly upregulated on CD8α+ PDC compared to CD8α– PDC (p < 0.05, paired t-test) (Figure 1D), while five other surface molecules (CD2, CD4, CD46, CD80, and CD123) were not differently regulated after HSVUV stimulation. These data suggested that the subset of CD8α+ PDC was particularly activated. Eventually, we exposed PDC of three donors to HSVUV for 40 h, separated these cells using a CD8 cell isolation kit, and exposed the CD8α+ and CD8α– PDC to HSVUV for another 20 h. Subsequently, cell culture supernatants were analyzed using a multiplex cytokine bead assay. Of a total of 25 cytokines, we found IFN-α, IL-8, IL-1RA, MIP-1α, MIP-1β, and MCP-1 upregulated in CD8α-expressing PDC. In contrast, IL-6 secretion was not different between the two subsets, and other cytokines were either not induced (IL-1β, IL-17, IFN-γ, GM-CSF, MIG, RANTES) or expressed only at very low levels (IL-2, IL-4, IL-5, IL-7, IL-12p40, IL-13, IL-15, eotaxin) (Figure 1E). IFN-α and IL-6 enhance T cell, B cell, and NK cell development and function; IL-8 recruits T cells and induces their degranulation; IL-1RA inhibits IL-1 induced T cell activation, and the chemokines MIP-1α, MIP-1β, and MCP-1 recruit immature DC, monocytes, and Th1 cells. Altogether, these data indicate that a subset of PDC gradually upregulates a homodimeric CD8α receptor upon HSV-1 stimulation, exposes a highly activated phenotype, and appears to be particularly active in recruiting other immune cells to the site of inflammation.
Conclusion
This is—at least to our knowledge—the first report that a subset of human PDC is capable of expressing CD8α at the cell surface upon HSV-1 stimulation. This subset is phenotypically different from the CD8α– PDC in expressing increased levels of markers for activation, costimulation, and migration. In parallel, CD8α+ PDC secrete enhanced levels of proinflammatory cytokines and chemokines. Therefore, this subset may play an important role in innate and adaptive immune defenses in HSV-1 infections. So far, it is unclear whether CD8α+ PDC are just a more activated subset, which “does better” than CD8α– PDC, or whether they have additional or different functions, such as being actively involved in cross-presentation. Further studies are required to define the conditions under which PDC present antigen efficiently and which formulation of antigen fits best for PDC cross-presentation (Villadangos and Young, 2008). Notably, murine knockouts for IRF8 lead to deficiencies in PDC and lymphoid-resident CD8α+ DC (Schiavoni et al., 2002; Aliberti et al., 2003; Tsujimura et al., 2003). This phenomenon may point to a common link in development and possibly function of these two cell populations. Further analyses of human CD8α-expressing PDC will delineate their role in the defense against viral infections, and—if viral vectors are used—also in anti-tumor responses.
Author Contributions
PS, ST, and MW performed the experiments, JV contributed multiplex cytokine bead array data and performed proof-reading, and BS and PS wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Bernhard Fleckenstein, Erlangen, and André Gessner, Regensburg, for continuous support, and Ulrike Samulowitz, Pfizer Oligonucleotide Therapeutics Unit, Coley Pharmaceutical GmbH, Düsseldorf, Germany, for performing cytokine bead arrays. Part of this work was supported by the doctoral training programs GK1071 (“Viruses of the immune system”; to PS) and GRK1660 (“Key signals of adaptive immune response”; to ST), and the “Akademie der Wissenschaften und Literatur zu Mainz.”
Abbreviations
DC, dendritic cells; HSV, Herpes simplex virus; IFN, interferon; IL, interleukin; PBMC, peripheral blood mononuclear cells; PDC, plasmacytoid dendritic cells.
References
Albert, M. L., Pearce, S. F., Francisco, L. M., Sauter, B., Roy, P., Silverstein, R. L., et al. (1998). Immature dendritic cells phagocytose apoptotic cells via αvβ5 and CD36, and cross-present antigens to cytotoxic T lymphocytes. J. Exp. Med. 188, 1359–1368. doi: 10.1084/jem.188.7.1359
Aliberti, J., Schulz, O., Pennington, D. J., Tsujimura, H., Reis e Sousa, C., Ozato, K., et al. (2003). Essential role for ICSBP in the in vivo development of murine CD8α+ dendritic cells. Blood 101, 305–310. doi: 10.1182/blood-2002-04-1088
Allan, R. S., Smith, C. M., Belz, G. T., Van Lint, A. L., Wakim, L. M., Heath, W. R., et al. (2003). Epidermal viral immunity induced by CD8α+ dendritic cells but not by Langerhans cells. Science 301, 1925–1928. doi: 10.1126/science.1087576
Allan, R. S., Waithman, J., Bedoui, S., Jones, C. M., Villadangos, J. A., Zhan, Y., et al. (2006). Migratory dendritic cells transfer antigen to a lymph node-resident dendritic cell population for efficient CTL priming. Immunity 25, 153–162. doi: 10.1016/j.immuni.2006.04.017
Anjuere, F., Martin, P., Ferrero, I., Fraga, M. L., Del Hoyo, G. M., Wright, N., et al. (1999). Definition of dendritic cell subpopulations present in the spleen, Peyer’s patches, lymph nodes, and skin of the mouse. Blood 93, 590–598.
Anjuere, F., Martinez Del Hoyo, G., Martin, P., and Ardavin, C. (2000). Langerhans cells acquire a CD8+ dendritic cell phenotype on maturation by CD40 ligation. J. Leukoc. Biol. 67, 206–209.
Asselin-Paturel, C., Boonstra, A., Dalod, M., Durand, I., Yessaad, N., Dezutter-Dambuyant, C., et al. (2001). Mouse type I IFN-producing cells are immature APCs with plasmacytoid morphology. Nat. Immunol. 2, 1144–1150. doi: 10.1038/ni736
Bachem, A., Guttler, S., Hartung, E., Ebstein, F., Schaefer, M., Tannert, A., et al. (2010). Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells. J. Exp. Med. 207, 1273–1281. doi: 10.1084/jem.20100348
Bamboat, Z. M., Stableford, J. A., Plitas, G., Burt, B. M., Nguyen, H. M., Welles, A. P., et al. (2009). Human liver dendritic cells promote T cell hyporesponsiveness. J. Immunol. 182, 1901–1911. doi: 10.4049/jimmunol.0803404
Bedoui, S., and Greyer, M. (2014). The role of dendritic cells in immunity against primary herpes simplex virus infections. Front. Microbiol. 5:533. doi: 10.3389/fmicb.2014.00533
Bedoui, S., Whitney, P. G., Waithman, J., Eidsmo, L., Wakim, L., Caminschi, I., et al. (2009). Cross-presentation of viral and self antigens by skin-derived CD103+ dendritic cells. Nat. Immunol. 10, 488–495. doi: 10.1038/ni.1724
Belz, G. T., Smith, C. M., Eichner, D., Shortman, K., Karupiah, G., Carbone, F. R., et al. (2004a). Cutting edge: conventional CD8α+ dendritic cells are generally involved in priming CTL immunity to viruses. J. Immunol. 172, 1996–2000. doi: 10.4049/jimmunol.172.4.1996
Belz, G. T., Smith, C. M., Kleinert, L., Reading, P., Brooks, A., Shortman, K., et al. (2004b). Distinct migrating and nonmigrating dendritic cell populations are involved in MHC class I-restricted antigen presentation after lung infection with virus. Proc. Natl. Acad. Sci. U.S.A. 101, 8670–8675. doi: 10.1073/pnas.0402644101
Boltjes, A., and van Wijk, F. (2014). Human dendritic cell functional specialization in steady-state and inflammation. Front. Immunol. 5:131. doi: 10.3389/fimmu.2014.00131
Bosnjak, L., Miranda-Saksena, M., Koelle, D. M., Boadle, R. A., Jones, C. A., and Cunningham, A. L. (2005). Herpes simplex virus infection of human dendritic cells induces apoptosis and allows cross-presentation via uninfected dendritic cells. J. Immunol. 174, 2220–2227. doi: 10.4049/jimmunol.174.4.2220
Bursch, L. S., Wang, L., Igyarto, B., Kissenpfennig, A., Malissen, B., Kaplan, D. H., et al. (2007). Identification of a novel population of Langerin+ dendritic cells. J. Exp. Med. 204, 3147–3156. doi: 10.1084/jem.20071966
Cabezon, R., Sintes, J., Llinas, L., and Benitez-Ribas, D. (2011). Analysis of HLDA9 mAbs on plasmacytoid dendritic cells. Immunol. Lett. 134, 167–173. doi: 10.1016/j.imlet.2010.09.020
Caminschi, I., Proietto, A. I., Ahmet, F., Kitsoulis, S., Shin Teh, J., Lo, J. C., et al. (2008). The dendritic cell subtype-restricted C-type lectin Clec9A is a target for vaccine enhancement. Blood 112, 3264–3273. doi: 10.1182/blood-2008-05-155176
Carbone, F. R., Belz, G. T., and Heath, W. R. (2004). Transfer of antigen between migrating and lymph node-resident DCs in peripheral T-cell tolerance and immunity. Trends Immunol. 25, 655–658. doi: 10.1016/j.it.2004.09.013
Cella, M., Facchetti, F., Lanzavecchia, A., and Colonna, M. (2000). Plasmacytoid dendritic cells activated by influenza virus and CD40L drive a potent TH1 polarization. Nat. Immunol. 1, 305–310. doi: 10.1038/79747
Cella, M., Jarrossay, D., Facchetti, F., Alebardi, O., Nakajima, H., Lanzavecchia, A., et al. (1999). Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat. Med. 5, 919–923. doi: 10.1038/11360
Chung, Y., Chang, J. H., Kweon, M. N., Rennert, P. D., and Kang, C. Y. (2005). CD8alpha-11b+ dendritic cells but not CD8α+ dendritic cells mediate cross-tolerance toward intestinal antigens. Blood 106, 201–206. doi: 10.1182/blood-2004-11-4240
Crowley, M., Inaba, K., Witmer-Pack, M., and Steinman, R. M. (1989). The cell surface of mouse dendritic cells: FACS analyses of dendritic cells from different tissues including thymus. Cell. Immunol. 118, 108–125. doi: 10.1016/0008-8749(89)90361-4
Crozat, K., Guiton, R., Contreras, V., Feuillet, V., Dutertre, C. A., Ventre, E., et al. (2010). The XC chemokine receptor 1 is a conserved selective marker of mammalian cells homologous to mouse CD8α+ dendritic cells. J. Exp. Med. 207, 1283–1292. doi: 10.1084/jem.20100223
Delamarre, L., Pack, M., Chang, H., Mellman, I., and Trombetta, E. S. (2005). Differential lysosomal proteolysis in antigen-presenting cells determines antigen fate. Science 307, 1630–1634. doi: 10.1126/science.1108003
den Haan, J. M., Lehar, S. M., and Bevan, M. J. (2000). CD8+ but not CD8– dendritic cells cross-prime cytotoxic T cells in vivo. J. Exp. Med. 192, 1685–1696. doi: 10.1084/jem.192.12.1685
Di Pucchio, T., Chatterjee, B., Smed-Sorensen, A., Clayton, S., Palazzo, A., Montes, M., et al. (2008). Direct proteasome-independent cross-presentation of viral antigen by plasmacytoid dendritic cells on major histocompatibility complex class I. Nat. Immunol. 9, 551–557. doi: 10.1038/ni.1602
Diamond, M. S., Kinder, M., Matsushita, H., Mashayekhi, M., Dunn, G. P., Archambault, J. M., et al. (2011). Type I interferon is selectively required by dendritic cells for immune rejection of tumors. J. Exp. Med. 208, 1989–2003. doi: 10.1084/jem.20101158
Donaghy, H., Bosnjak, L., Harman, A. N., Marsden, V., Tyring, S. K., Meng, T. C., et al. (2009). Role for plasmacytoid dendritic cells in the immune control of recurrent human herpes simplex virus infection. J. Virol. 83, 1952–1961. doi: 10.1128/JVI.01578-08
Donnenberg, V. S., O’connell, P. J., Logar, A. J., Zeevi, A., Thomson, A. W., and Donnenberg, A. D. (2001). Rare-event analysis of circulating human dendritic cell subsets and their presumptive mouse counterparts. Transplantation 72, 1946–1951. doi: 10.1097/00007890-200112270-00014
Dorner, B. G., Dorner, M. B., Zhou, X., Opitz, C., Mora, A., Guttler, S., et al. (2009). Selective expression of the chemokine receptor XCR1 on cross-presenting dendritic cells determines cooperation with CD8+ T cells. Immunity 31, 823–833. doi: 10.1016/j.immuni.2009.08.027
Dudziak, D., Kamphorst, A. O., Heidkamp, G. F., Buchholz, V. R., Trumpfheller, C., Yamazaki, S., et al. (2007). Differential antigen processing by dendritic cell subsets in vivo. Science 315, 107–111. doi: 10.1126/science.1136080
Dzionek, A., Fuchs, A., Schmidt, P., Cremer, S., Zysk, M., Miltenyi, S., et al. (2000). BDCA-2, BDCA-3, and BDCA-4: three markers for distinct subsets of dendritic cells in human peripheral blood. J. Immunol. 165, 6037–6046. doi: 10.4049/jimmunol.165.11.6037
Edelson, B. T., Kc, W., Juang, R., Kohyama, M., Benoit, L. A., Klekotka, P. A., et al. (2010). Peripheral CD103+ dendritic cells form a unified subset developmentally related to CD8α+ conventional dendritic cells. J. Exp. Med. 207, 823–836. doi: 10.1084/jem.20091627
Edwards, A. D., Diebold, S. S., Slack, E. M., Tomizawa, H., Hemmi, H., Kaisho, T., et al. (2003). Toll-like receptor expression in murine DC subsets: lack of TLR7 expression by CD8α+ DC correlates with unresponsiveness to imidazoquinolines. Eur. J. Immunol. 33, 827–833. doi: 10.1002/eji.200323797
Eidsmo, L., Allan, R., Caminschi, I., Van Rooijen, N., Heath, W. R., and Carbone, F. R. (2009). Differential migration of epidermal and dermal dendritic cells during skin infection. J. Immunol. 182, 3165–3172. doi: 10.4049/jimmunol.0802950
Feldman, M., Howell, D., and Fitzgerald-Bocarsly, P. (1992). Interferon-α-dependent and -independent participation of accessory cells in natural killer cell-mediated lysis of HSV-1-infected fibroblasts. J. Leukoc. Biol. 52, 473–482.
Fonteneau, J. F., Gilliet, M., Larsson, M., Dasilva, I., Munz, C., Liu, Y. J., et al. (2003a). Activation of influenza virus-specific CD4+ and CD8+ T cells: a new role for plasmacytoid dendritic cells in adaptive immunity. Blood 101, 3520–3526. doi: 10.1182/blood-2002-10-3063
Fonteneau, J. F., Kavanagh, D. G., Lirvall, M., Sanders, C., Cover, T. L., Bhardwaj, N., et al. (2003b). Characterization of the MHC class I cross-presentation pathway for cell-associated antigens by human dendritic cells. Blood 102, 4448–4455. doi: 10.1182/blood-2003-06-1801
Fuertes, M. B., Kacha, A. K., Kline, J., Woo, S. R., Kranz, D. M., Murphy, K. M., et al. (2011). Host type I IFN signals are required for antitumor CD8+ T cell responses through CD8α+ dendritic cells. J. Exp. Med. 208, 2005–2016. doi: 10.1084/jem.20101159
Galibert, L., Diemer, G. S., Liu, Z., Johnson, R. S., Smith, J. L., Walzer, T., et al. (2005). Nectin-like protein 2 defines a subset of T-cell zone dendritic cells and is a ligand for class-I-restricted T-cell-associated molecule. J. Biol. Chem. 280, 21955–21964. doi: 10.1074/jbc.M502095200
GeurtsvanKessel, C. H., Willart, M. A., Van Rijt, L. S., Muskens, F., Kool, M., Baas, C., et al. (2008). Clearance of influenza virus from the lung depends on migratory langerin+CD11b– but not plasmacytoid dendritic cells. J. Exp. Med. 205, 1621–1634. doi: 10.1084/jem.20071365
Ginhoux, F., Collin, M. P., Bogunovic, M., Abel, M., Leboeuf, M., Helft, J., et al. (2007). Blood-derived dermal langerin+ dendritic cells survey the skin in the steady state. J. Exp. Med. 204, 3133–3146. doi: 10.1084/jem.20071733
Ginhoux, F., Liu, K., Helft, J., Bogunovic, M., Greter, M., Hashimoto, D., et al. (2009). The origin and development of nonlymphoid tissue CD103+ DCs. J. Exp. Med. 206, 3115–3130. doi: 10.1084/jem.20091756
Guillerme, J. B., Boisgerault, N., Roulois, D., Menager, J., Combredet, C., Tangy, F., et al. (2013). Measles virus vaccine-infected tumor cells induce tumor antigen cross-presentation by human plasmacytoid dendritic cells. Clin. Cancer Res. 19, 1147–1158. doi: 10.1158/1078-0432.CCR-12-2733
Hennies, C. M., Reboulet, R. A., Garcia, Z., Nierkens, S., Wolkers, M. C., and Janssen, E. M. (2011). Selective expansion of merocytic dendritic cells and CD8DCs confers anti-tumour effect of Fms-like tyrosine kinase 3-ligand treatment in vivo. Clin. Exp. Immunol. 163, 381–391. doi: 10.1111/j.1365-2249.2010.04305.x
Henri, S., Vremec, D., Kamath, A., Waithman, J., Williams, S., Benoist, C., et al. (2001). The dendritic cell populations of mouse lymph nodes. J. Immunol. 167, 741–748. doi: 10.4049/jimmunol.167.2.741
Hildner, K., Edelson, B. T., Purtha, W. E., Diamond, M., Matsushita, H., Kohyama, M., et al. (2008). Batf3 deficiency reveals a critical role for CD8α+ dendritic cells in cytotoxic T cell immunity. Science 322, 1097–1100. doi: 10.1126/science.1164206
Hochrein, H., Shortman, K., Vremec, D., Scott, B., Hertzog, P., and O’keeffe, M. (2001). Differential production of IL-12, IFN-α, and IFN-gamma by mouse dendritic cell subsets. J. Immunol. 166, 5448–5455. doi: 10.4049/jimmunol.166.9.5448
Hoeffel, G., Ripoche, A. C., Matheoud, D., Nascimbeni, M., Escriou, N., Lebon, P., et al. (2007). Antigen crosspresentation by human plasmacytoid dendritic cells. Immunity 27, 481–492. doi: 10.1016/j.immuni.2007.07.021
Huysamen, C., Willment, J. A., Dennehy, K. M., and Brown, G. D. (2008). CLEC9A is a novel activation C-type lectin-like receptor expressed on BDCA3+ dendritic cells and a subset of monocytes. J. Biol. Chem. 283, 16693–16701. doi: 10.1074/jbc.M709923200
Ito, T., Yang, M., Wang, Y. H., Lande, R., Gregorio, J., Perng, O. A., et al. (2007). Plasmacytoid dendritic cells prime IL-10-producing T regulatory cells by inducible costimulator ligand. J. Exp. Med. 204, 105–115. doi: 10.1084/jem.20061660
Iyoda, T., Shimoyama, S., Liu, K., Omatsu, Y., Akiyama, Y., Maeda, Y., et al. (2002). The CD8+ dendritic cell subset selectively endocytoses dying cells in culture and in vivo. J. Exp. Med. 195, 1289–1302. doi: 10.1084/jem.20020161
Jaehn, P. S., Zaenker, K. S., Schmitz, J., and Dzionek, A. (2008). Functional dichotomy of plasmacytoid dendritic cells: antigen-specific activation of T cells versus production of type I interferon. Eur. J. Immunol. 38, 1822–1832. doi: 10.1002/eji.200737552
Janssen, E., Tabeta, K., Barnes, M. J., Rutschmann, S., Mcbride, S., Bahjat, K. S., et al. (2006). Efficient T cell activation via a Toll-Interleukin 1 Receptor-independent pathway. Immunity 24, 787–799. doi: 10.1016/j.immuni.2006.03.024
Jirmo, A. C., Nagel, C. H., Bohnen, C., Sodeik, B., and Behrens, G. M. (2009). Contribution of direct and cross-presentation to CTL immunity against herpes simplex virus 1. J. Immunol. 182, 283–292. doi: 10.4049/jimmunol.182.1.283
Joffre, O. P., Segura, E., Savina, A., and Amigorena, S. (2012). Cross-presentation by dendritic cells. Nat. Rev. Immunol. 12, 557–569. doi: 10.1038/nri3254
Jones, C. A., Fernandez, M., Herc, K., Bosnjak, L., Miranda-Saksena, M., Boadle, R. A., et al. (2003). Herpes simplex virus type 2 induces rapid cell death and functional impairment of murine dendritic cells in vitro. J. Virol. 77, 11139–11149. doi: 10.1128/JVI.77.20.11139-11149.2003
Jongbloed, S. L., Kassianos, A. J., Mcdonald, K. J., Clark, G. J., Ju, X., Angel, C. E., et al. (2010). Human CD141+ BDCA-3+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J. Exp. Med. 207, 1247–1260. doi: 10.1084/jem.20092140
Kadowaki, N., Antonenko, S., Lau, J. Y., and Liu, Y. J. (2000). Natural interferon α/β-producing cells link innate and adaptive immunity. J. Exp. Med. 192, 219–226. doi: 10.1084/jem.192.2.219
Kadowaki, N., Ho, S., Antonenko, S., Malefyt, R. W., Kastelein, R. A., Bazan, F., et al. (2001). Subsets of human dendritic cell precursors express different toll-like receptors and respond to different microbial antigens. J. Exp. Med. 194, 863–869. doi: 10.1084/jem.194.6.863
Kawamura, K., Kadowaki, N., Kitawaki, T., and Uchiyama, T. (2006). Virus-stimulated plasmacytoid dendritic cells induce CD4+ cytotoxic regulatory T cells. Blood 107, 1031–1038. doi: 10.1182/blood-2005-04-1737
Kool, M., Geurtsvankessel, C., Muskens, F., Madeira, F. B., Van Nimwegen, M., Kuipers, H., et al. (2011). Facilitated antigen uptake and timed exposure to TLR ligands dictate the antigen-presenting potential of plasmacytoid DCs. J. Leukoc. Biol. 90, 1177–1190. doi: 10.1189/jlb.0610342
Lauterbach, H., Bathke, B., Gilles, S., Traidl-Hoffmann, C., Luber, C. A., Fejer, G., et al. (2010). Mouse CD8α+ DCs and human BDCA3+ DCs are major producers of IFN-lambda in response to poly IC. J. Exp. Med. 207, 2703–2717. doi: 10.1084/jem.20092720
Le Bon, A., Etchart, N., Rossmann, C., Ashton, M., Hou, S., Gewert, D., et al. (2003). Cross-priming of CD8+ T cells stimulated by virus-induced type I interferon. Nat. Immunol. 4, 1009–1015. doi: 10.1038/ni978
Lee, H. K., Zamora, M., Linehan, M. M., Iijima, N., Gonzalez, D., Haberman, A., et al. (2009). Differential roles of migratory and resident DCs in T cell priming after mucosal or skin HSV-1 infection. J. Exp. Med. 206, 359–370. doi: 10.1084/jem.20080601
Leon, B., Lopez-Bravo, M., and Ardavin, C. (2007). Monocyte-derived dendritic cells formed at the infection site control the induction of protective T helper 1 responses against Leishmania. Immunity 26, 519–531. doi: 10.1016/j.immuni.2007.01.017
Lin, M. L., Zhan, Y., Proietto, A. I., Prato, S., Wu, L., Heath, W. R., et al. (2008). Selective suicide of cross-presenting CD8+ dendritic cells by cytochrome c injection shows functional heterogeneity within this subset. Proc. Natl. Acad. Sci. U.S.A. 105, 3029–3034. doi: 10.1073/pnas.0712394105
Lindstedt, M., Lundberg, K., and Borrebaeck, C. A. (2005). Gene family clustering identifies functionally associated subsets of human in vivo blood and tonsillar dendritic cells. J. Immunol. 175, 4839–4846. doi: 10.4049/jimmunol.175.8.4839
Lorenzi, S., Mattei, F., Sistigu, A., Bracci, L., Spadaro, F., Sanchez, M., et al. (2011). Type I IFNs control antigen retention and survival of CD8α+ dendritic cells after uptake of tumor apoptotic cells leading to cross-priming. J. Immunol. 186, 5142–5150. doi: 10.4049/jimmunol.1004163
Lui, G., Manches, O., Angel, J., Molens, J. P., Chaperot, L., and Plumas, J. (2009). Plasmacytoid dendritic cells capture and cross-present viral antigens from influenza-virus exposed cells. PLoS ONE 4:e7111. doi: 10.1371/journal.pone.0007111
Lund, J., Sato, A., Akira, S., Medzhitov, R., and Iwasaki, A. (2003). Toll-like receptor 9-mediated recognition of Herpes simplex virus-2 by plasmacytoid dendritic cells. J. Exp. Med. 198, 513–520. doi: 10.1084/jem.20030162
MacDonald, K. P., Munster, D. J., Clark, G. J., Dzionek, A., Schmitz, J., and Hart, D. N. (2002). Characterization of human blood dendritic cell subsets. Blood 100, 4512–4520. doi: 10.1182/blood-2001-11-0097
Martinez del Hoyo, G., Martin, P., Arias, C. F., Marin, A. R., and Ardavin, C. (2002). CD8α+ dendritic cells originate from the CD8α– dendritic cell subset by a maturation process involving CD8α, DEC-205, and CD24 up-regulation. Blood 99, 999–1004. doi: 10.1182/blood.V99.3.999
Mazzini, E., Massimiliano, L., Penna, G., and Rescigno, M. (2014). Oral tolerance can be established via gap junction transfer of fed antigens from CX3CR1+ macrophages to CD103+ dendritic cells. Immunity 40, 248–261. doi: 10.1016/j.immuni.2013.12.012
Megjugorac, N. J., Young, H. A., Amrute, S. B., Olshalsky, S. L., and Fitzgerald-Bocarsly, P. (2004). Virally stimulated plasmacytoid dendritic cells produce chemokines and induce migration of T and NK cells. J. Leukoc. Biol. 75, 504–514. doi: 10.1189/jlb.0603291
Merad, M., Fong, L., Bogenberger, J., and Engleman, E. G. (2000). Differentiation of myeloid dendritic cells into CD8α-positive dendritic cells in vivo. Blood 96, 1865–1872.
Merad, M., Sathe, P., Helft, J., Miller, J., and Mortha, A. (2013). The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu. Rev. Immunol. 31, 563–604. doi: 10.1146/annurev-immunol-020711-074950
Mittag, D., Proietto, A. I., Loudovaris, T., Mannering, S. I., Vremec, D., Shortman, K., et al. (2011). Human dendritic cell subsets from spleen and blood are similar in phenotype and function but modified by donor health status. J. Immunol. 186, 6207–6217. doi: 10.4049/jimmunol.1002632
Mouries, J., Moron, G., Schlecht, G., Escriou, N., Dadaglio, G., and Leclerc, C. (2008). Plasmacytoid dendritic cells efficiently cross-prime naive T cells in vivo after TLR activation. Blood 112, 3713–3722. doi: 10.1182/blood-2008-03-146290
Naik, S. H. (2008). Demystifying the development of dendritic cell subtypes, a little. Immunol. Cell Biol. 86, 439–452. doi: 10.1038/icb.2008.28
Nakano, H., Yanagita, M., and Gunn, M. D. (2001). CD11c+B220+Gr-1+ cells in mouse lymph nodes and spleen display characteristics of plasmacytoid dendritic cells. J. Exp. Med. 194, 1171–1178. doi: 10.1084/jem.194.8.1171
Nierkens, S., Tel, J., Janssen, E., and Adema, G. J. (2013). Antigen cross-presentation by dendritic cell subsets: one general or all sergeants? Trends Immunol. 34, 361–370. doi: 10.1016/j.it.2013.02.007
O’Keeffe, M., Hochrein, H., Vremec, D., Caminschi, I., Miller, J. L., Anders, E. M., et al. (2002). Mouse plasmacytoid cells: long-lived cells, heterogeneous in surface phenotype and function, that differentiate into CD8+ dendritic cells only after microbial stimulus. J. Exp. Med. 196, 1307–1319. doi: 10.1084/jem.20021031
O’Keeffe, M., Hochrein, H., Vremec, D., Scott, B., Hertzog, P., Tatarczuch, L., et al. (2003). Dendritic cell precursor populations of mouse blood: identification of the murine homologues of human blood plasmacytoid pre-DC2 and CD11c+ DC1 precursors. Blood 101, 1453–1459. doi: 10.1182/blood-2002-03-0974
Palucka, K., and Banchereau, J. (2012). Cancer immunotherapy via dendritic cells. Nat. Rev. Cancer 12, 265–277. doi: 10.1038/nrc3258
Pooley, J. L., Heath, W. R., and Shortman, K. (2001). Cutting edge: intravenous soluble antigen is presented to CD4 T cells by CD8- dendritic cells, but cross-presented to CD8 T cells by CD8+ dendritic cells. J. Immunol. 166, 5327–5330. doi: 10.4049/jimmunol.166.9.5327
Poulin, L. F., Henri, S., De Bovis, B., Devilard, E., Kissenpfennig, A., and Malissen, B. (2007). The dermis contains langerin+ dendritic cells that develop and function independently of epidermal Langerhans cells. J. Exp. Med. 204, 3119–3131. doi: 10.1084/jem.20071724
Poulin, L. F., Salio, M., Griessinger, E., Anjos-Afonso, F., Craciun, L., Chen, J. L., et al. (2010). Characterization of human DNGR-1+ BDCA3+ leukocytes as putative equivalents of mouse CD8α+ dendritic cells. J. Exp. Med. 207, 1261–1271. doi: 10.1084/jem.20092618
Puttur, F. K., Fernandez, M. A., White, R., Roediger, B., Cunningham, A. L., Weninger, W., et al. (2010). Herpes simplex virus infects skin gamma delta T cells before Langerhans cells and impedes migration of infected Langerhans cells by inducing apoptosis and blocking E-cadherin downregulation. J. Immunol. 185, 477–487. doi: 10.4049/jimmunol.0904106
Reboulet, R. A., Hennies, C. M., Garcia, Z., Nierkens, S., and Janssen, E. M. (2010). Prolonged antigen storage endows merocytic dendritic cells with enhanced capacity to prime anti-tumor responses in tumor-bearing mice. J. Immunol. 185, 3337–3347. doi: 10.4049/jimmunol.1001619
Robbins, S. H., Walzer, T., Dembele, D., Thibault, C., Defays, A., Bessou, G., et al. (2008). Novel insights into the relationships between dendritic cell subsets in human and mouse revealed by genome-wide expression profiling. Genome Biol. 9, R17. doi: 10.1186/gb-2008-9-1-r17
Sancho, D., Joffre, O. P., Keller, A. M., Rogers, N. C., Martinez, D., Hernanz-Falcon, P., et al. (2009). Identification of a dendritic cell receptor that couples sensing of necrosis to immunity. Nature 458, 899–903. doi: 10.1038/nature07750
Sapoznikov, A., Fischer, J. A., Zaft, T., Krauthgamer, R., Dzionek, A., and Jung, S. (2007). Organ-dependent in vivo priming of naive CD4+, but not CD8+, T cells by plasmacytoid dendritic cells. J. Exp. Med. 204, 1923–1933. doi: 10.1084/jem.20062373
Savina, A., Peres, A., Cebrian, I., Carmo, N., Moita, C., Hacohen, N., et al. (2009). The small GTPase Rac2 controls phagosomal alkalinization and antigen crosspresentation selectively in CD8+ dendritic cells. Immunity 30, 544–555. doi: 10.1016/j.immuni.2009.01.013
Schiavoni, G., Mattei, F., Sestili, P., Borghi, P., Venditti, M., Morse, H. C., et al. (2002). ICSBP is essential for the development of mouse type I interferon-producing cells and for the generation and activation of CD8α+ dendritic cells. J. Exp. Med. 196, 1415–1425. doi: 10.1084/jem.20021263
Schnorrer, P., Behrens, G. M., Wilson, N. S., Pooley, J. L., Smith, C. M., El-Sukkari, D., et al. (2006). The dominant role of CD8+ dendritic cells in cross-presentation is not dictated by antigen capture. Proc. Natl. Acad. Sci. U.S.A. 103, 10729–10734. doi: 10.1073/pnas.0601956103
Schnurr, M., Chen, Q., Shin, A., Chen, W., Toy, T., Jenderek, C., et al. (2005). Tumor antigen processing and presentation depend critically on dendritic cell type and the mode of antigen delivery. Blood 105, 2465–2472. doi: 10.1182/blood-2004-08-3105
Schreibelt, G., Klinkenberg, L. J., Cruz, L. J., Tacken, P. J., Tel, J., Kreutz, M., et al. (2012). The C-type lectin receptor CLEC9A mediates antigen uptake and (cross-)presentation by human blood BDCA3+ myeloid dendritic cells. Blood 119, 2284–2292. doi: 10.1182/blood-2011-08-373944
Schulz, O., and Reis e Sousa, C. (2002). Cross-presentation of cell-associated antigens by CD8α+ dendritic cells is attributable to their ability to internalize dead cells. Immunology 107, 183–189. doi: 10.1046/j.1365-2567.2002.01513.x
Schuster, P., Boscheinen, J. B., Tennert, K., and Schmidt, B. (2011). the role of plasmacytoid dendritic cells in innate and adaptive immune responses against α herpes virus infections. Adv. Virol. 2011, 12. doi: 10.1155/2011/679271
Schuster, P., Donhauser, N., Pritschet, K., Ries, M., Haupt, S., Kittan, N. A., et al. (2010). Co-ordinated regulation of plasmacytoid dendritic cell surface receptors upon stimulation with herpes simplex virus type 1. Immunology 129, 234–247. doi: 10.1111/j.1365-2567.2009.03176.x
Segura, E., Durand, M., and Amigorena, S. (2013). Similar antigen cross-presentation capacity and phagocytic functions in all freshly isolated human lymphoid organ-resident dendritic cells. J. Exp. Med. 210, 1035–1047. doi: 10.1084/jem.20121103
Segura, E., Valladeau-Guilemond, J., Donnadieu, M. H., Sastre-Garau, X., Soumelis, V., and Amigorena, S. (2012). Characterization of resident and migratory dendritic cells in human lymph nodes. J. Exp. Med. 209, 653–660. doi: 10.1084/jem.20111457
Shinohara, M. L., Lu, L., Bu, J., Werneck, M. B., Kobayashi, K. S., Glimcher, L. H., et al. (2006). Osteopontin expression is essential for interferon-α production by plasmacytoid dendritic cells. Nat. Immunol. 7, 498–506. doi: 10.1038/ni1327
Shortman, K., and Heath, W. R. (2010). The CD8+ dendritic cell subset. Immunol. Rev. 234, 18–31. doi: 10.1111/j.0105-2896.2009.00870.x
Siegal, F. P., Kadowaki, N., Shodell, M., Fitzgerald-Bocarsly, P. A., Shah, K., Ho, S., et al. (1999). The nature of the principal type 1 interferon-producing cells in human blood. Science 284, 1835–1837. doi: 10.1126/science.284.5421.1835
Smith, C. M., Belz, G. T., Wilson, N. S., Villadangos, J. A., Shortman, K., Carbone, F. R., et al. (2003). Cutting edge: conventional CD8α+ dendritic cells are preferentially involved in CTL priming after footpad infection with herpes simplex virus-1. J. Immunol. 170, 4437–4440. doi: 10.4049/jimmunol.170.9.4437
Steinman, R. M., and Cohn, Z. A. (1973). Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J. Exp. Med. 137, 1142–1162. doi: 10.1084/jem.137.5.1142
Swiecki, M., Wang, Y., Gilfillan, S., and Colonna, M. (2013). Plasmacytoid dendritic cells contribute to systemic but not local antiviral responses to HSV infections. PLoS Pathog. 9:e1003728. doi: 10.1371/journal.ppat.1003728
Tel, J., Lambeck, A. J., Cruz, L. J., Tacken, P. J., De Vries, I. J., and Figdor, C. G. (2010). Human plasmacytoid dendritic cells phagocytose, process, and present exogenous particulate antigen. J. Immunol. 184, 4276–4283. doi: 10.4049/jimmunol.0903286
Tel, J., Smits, E. L., Anguille, S., Joshi, R. N., Figdor, C. G., and De Vries, I. J. (2012). Human plasmacytoid dendritic cells are equipped with antigen-presenting and tumoricidal capacities. Blood 120, 3936–3944. doi: 10.1182/blood-2012-06-435941
Thery, C., Ostrowski, M., and Segura, E. (2009). Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 9, 581–593. doi: 10.1038/nri2567
Tsujimura, H., Tamura, T., and Ozato, K. (2003). Cutting edge: IFN consensus sequence binding protein/IFN regulatory factor 8 drives the development of type I IFN-producing plasmacytoid dendritic cells. J. Immunol. 170, 1131–1135. doi: 10.4049/jimmunol.170.3.1131
Vacchelli, E., Vitale, I., Eggermont, A., Fridman, W. H., Fucikova, J., Cremer, I., et al. (2013). Trial watch: dendritic cell-based interventions for cancer therapy. Oncoimmunology 2, e25771. doi: 10.4161/onci.25771
Velasquez-Lopera, M. M., Correa, L. A., and Garcia, L. F. (2008). Human spleen contains different subsets of dendritic cells and regulatory T lymphocytes. Clin. Exp. Immunol. 154, 107–114. doi: 10.1111/j.1365-2249.2008.03734.x
Villadangos, J. A., and Young, L. (2008). Antigen-presentation properties of plasmacytoid dendritic cells. Immunity 29, 352–361. doi: 10.1016/j.immuni.2008.09.002
Vogel, K., Thomann, S., Vogel, B., Schuster, P., and Schmidt, B. (2014). Both plasmacytoid dendritic cells and monocytes stimulate natural killer cells early during human herpes simplex virus type 1 infections. Immunology 143, 588–600. doi: 10.1111/imm.12337
Vremec, D., Pooley, J., Hochrein, H., Wu, L., and Shortman, K. (2000). CD4 and CD8 expression by dendritic cell subtypes in mouse thymus and spleen. J. Immunol. 164, 2978–2986. doi: 10.4049/jimmunol.164.6.2978
Vremec, D., Zorbas, M., Scollay, R., Saunders, D. J., Ardavin, C. F., Wu, L., et al. (1992). The surface phenotype of dendritic cells purified from mouse thymus and spleen: investigation of the CD8 expression by a subpopulation of dendritic cells. J. Exp. Med. 176, 47–58. doi: 10.1084/jem.176.1.47
Wakim, L. M., and Bevan, M. J. (2011). Cross-dressed dendritic cells drive memory CD8+ T-cell activation after viral infection. Nature 471, 629–632. doi: 10.1038/nature09863
Wilson, N. S., Behrens, G. M., Lundie, R. J., Smith, C. M., Waithman, J., Young, L., et al. (2006). Systemic activation of dendritic cells by Toll-like receptor ligands or malaria infection impairs cross-presentation and antiviral immunity. Nat. Immunol. 7, 165–172. doi: 10.1038/ni1300
Wilson, N. S., El-Sukkari, D., Belz, G. T., Smith, C. M., Steptoe, R. J., Heath, W. R., et al. (2003). Most lymphoid organ dendritic cell types are phenotypically and functionally immature. Blood 102, 2187–2194. doi: 10.1182/blood-2003-02-0513
Yoneyama, H., Matsuno, K., Toda, E., Nishiwaki, T., Matsuo, N., Nakano, A., et al. (2005). Plasmacytoid DCs help lymph node DCs to induce anti-HSV CTLs. J. Exp. Med. 202, 425–435. doi: 10.1084/jem.20041961
Keywords: dendritic cells, plasmacytoid, virus, HSV, human, murine
Citation: Schuster P, Thomann S, Werner M, Vollmer J and Schmidt B (2015) A subset of human plasmacytoid dendritic cells expresses CD8α upon exposure to herpes simplex virus type 1. Front. Microbiol. 6:557. doi: 10.3389/fmicb.2015.00557
Received: 23 February 2015; Accepted: 20 May 2015;
Published: 02 June 2015.
Edited by:
Laura Hertel, Children’s Hospital Oakland Research Institute, USAReviewed by:
Giada Frascaroli, Ulm University Medical Center, GermanyChristiane S. Heilingloh, Universitätsklinikum Erlangen, Germany
Copyright © 2015 Schuster, Thomann, Werner, Vollmer and Schmidt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Barbara Schmidt, Institute of Medical Microbiology and Hygiene, University of Regensburg, Franz-Josef-Strauss-Allee 11, 93053 Regensburg, Germany,YmFyYmFyYS5zY2htaWR0QHVrci5kZQ==