 Nadja Bier
Nadja Bier Keike Schwartz
Keike Schwartz Beatriz Guerra
Beatriz Guerra Eckhard Strauch
Eckhard Strauch- Department of Biological Safety, Federal Institute for Risk Assessment, Berlin, Germany
An increase in the occurrence of potentially pathogenic Vibrio species is expected for waters in Northern Europe as a consequence of global warming. In this context, a higher incidence of Vibrio infections is predicted for the future and forecasts suggest that people visiting and living at the Baltic Sea are at particular risk. This study aimed to investigate antimicrobial resistance patterns among Vibrio vulnificus and Vibrio cholerae non-O1/non-O139 isolates that could pose a public health risk. Antimicrobial susceptibility of 141 V. vulnificus and 184 V. cholerae non-O1/non-O139 strains isolated from German coastal waters (Baltic Sea and North Sea) as well as from patients and retail seafood was assessed by broth microdilution and disk diffusion. Both species were susceptible to most of the agents tested (12 subclasses) and no multidrug-resistance was observed. Among V. vulnificus isolates, non-susceptibility was exclusively found toward aminoglycosides. In case of V. cholerae, a noticeable proportion of strains was non-susceptible to aminopenicillins and aminoglycosides. In addition, resistance toward carbapenems, quinolones, and folate pathway inhibitors was sporadically observed. Biochemical testing indicated the production of carbapenemases with unusual substrate specificity in four environmental V. cholerae strains. Most antimicrobial agents recommended for treatment of V. vulnificus and V. cholerae non-O1/non-O139 infections were found to be effective in vitro. However, the occurrence of putative carbapenemase producing V. cholerae in German coastal waters is of concern and highlights the need for systematic monitoring of antimicrobial susceptibility in potentially pathogenic Vibrio spp. in Europe.
Introduction
The family Vibrionaceae within the class of Gammaproteobacteria comprises eight genera of Gram-negative, facultative anaerobic, straight, or curved rods that are mostly oxidase-positive, halophilic, and motile (Farmer and Janda, 2004). Members of this family are ubiquitously distributed in aquatic ecosystems worldwide. They can be found as free-living bacteria and as commensals of aquatic organisms and play an important role in nutrient cycling of natural aquatic habitats. Due to their metabolic diversity and their adaptive abilities to changing environmental conditions, a seasonal, and geographical variability of total Vibrio populations is observed in response to climatic influences and seawater circulations (Mansergh and Zehr, 2014).
Among the Vibrionaceae, a number of important human pathogenic bacteria have been identified that can cause gastrointestinal infections, wound infections or septicemia. Vibrio cholerae, Vibrio vulnificus, and Vibrio parahaemolyticus are considered as the most clinically relevant human pathogens within the genus Vibrio (Daniels and Shafaie, 2000). The latter species is widely disseminated in estuarine, marine, and coastal so surroundings and the leading cause of human intestinal infections after consumption of raw and undercooked seafood (Letchumanan et al., 2014). V. cholerae and V. vulnificus are also part of the microbial community in coastal or estuarine aquatic ecosystems with moderate salinities (Thompson and Polz, 2006).
V. cholerae is a well-known human pathogen consisting of more than 200 serogroups (Kaper et al., 1995; Lutz et al., 2013). Toxigenic V. cholerae of the O1 or O139 serogroup are the causative agents of cholera, an endemic disease in many Asian and African countries with symptoms of severe watery diarrhea, vomiting, and dehydration. All other serogroups designated as V. cholerae non-O1/non-O139 have also been linked to sporadically occurring human infections ranging from extraintestinal wound or ear infections (Huhulescu et al., 2007) to relatively mild or sometimes severe gastroenteritis (Tobin-D'Angelo et al., 2008), whereby smaller diarrheal outbreaks were also reported (Luo et al., 2013). Additionally, rarely occurring bacteremia has been described with mortality rates up to 61.5% (Petsaris et al., 2010). Since 2000, around 40 cases of V. cholerae non-O1/non-O139 infections in the United States have been reported to the CDC annually1. V. cholerae infections contracted in Germany were mainly ear or wound infections caused by non-toxigenic non-O1/non-O139 strains that were acquired through contact to seawater (Huehn et al., 2014). Due to the rare occurrence of V. cholerae non-O1/non-O139 infections, there are no official recommendations on antibiotic therapy (Petsaris et al., 2010). However, in case of bacteremia an early administration of antibiotic therapy can prevent a fatal outcome. Several case studies on V. cholerae non-O1/non-O139 bacteremia and wound infection exist, where fluoroquinolones and third-generation cephalosporins have been used (Huhulescu et al., 2007; Petsaris et al., 2010). But also treatment with ampicillin or last-line carbapenems has been described (Feghali and Adib, 2011; Lu et al., 2014).
V. vulnificus is known as a highly virulent pathogen. Although infections occur only sporadically, they can rapidly progress to septicemia, especially in persons with predisposing risk factors (e.g., immunocompromising conditions or chronic liver diseases resulting in elevated serum iron levels; Oliver, 2006). Foodborne infections can either result in a relatively mild gastroenteritis or in primary septicemia with mortality rates of 61% (Shapiro et al., 1998; Oliver, 2006). A second infection route for V. vulnificus is through open wounds exposed to seawater. Due to the high multiplication rate of the pathogen, wound infections may quickly progress to necrotizing fasciitis, which often makes surgical debridement or amputation necessary (Daniels and Shafaie, 2000). Delayed treatment promotes progression to secondary septicemia with mortality rates about seventeen per cent (Shapiro et al., 1998; Daniels and Shafaie, 2000). Surgical interventions should be considered early to prevent a fatal outcome as poor blood perfusion in necrotic tissue can impede the achievement of effective concentrations of antimicrobial agents (Chen et al., 2012). However, to avoid septicemia and a further distribution of the pathogen additional antibiotic therapy is indispensable and should be administered as early as possible. Due to the fast progression of V. vulnificus infections, the presence of antimicrobial resistance preventing an effective therapy can be fatal for the patient. A combination of a tetracycline with a third-generation cephalosporin or single-agent therapy with fluoroquinolones is recommended by the CDC2, while trimethoprim-sulfamethoxazole in combination with an aminoglycoside is proposed for the treatment of pregnant women and children.
In the U.S., 95% of all seafood-related deaths can be attributed to V. vulnificus, whereas infections in Germany were almost exclusively wound infections occurring after contact to seawater (Oliver, 2006; Huehn et al., 2014). So far, Vibrio infections in Germany occur only sporadically but incidences peaked after extreme heatwaves (Huehn et al., 2014). Due to impacts of climate change, a rise in the occurrence of V. vulnificus and V. cholerae is predicted for European waters (Baker-Austin et al., 2012). Changing demography is expected to further contribute to higher incidences of Vibrio infections (Baker-Austin et al., 2012). In view of these forecasts and the potential severity of infections, an investigation on antimicrobial susceptibility of Vibrio spp. is demanded to provide guidance for medical intervention, but also for epidemiological purposes. For this reason, our study aimed to assess antimicrobial resistance prevalence among V. vulnificus and V. cholerae non-O1/non-O139 posing a public health risk for the population. Environmental isolates were obtained from German coastal and estuarine waters of the open North Sea and the intracontinental Baltic Sea. In addition, we also included isolates from clinical sources and retail seafood for comparison and to give a more comprehensive overview of antimicrobial resistance patterns of these two species in Germany. To our knowledge, this is the first study examining antimicrobial susceptibility of V. vulnificus and V. cholerae non-O1/non-O139 in Northern Europe on a large scale.
Materials and Methods
Bacterial Strains
The strains used in this study are summarized in Table 1 and listed in detail in Supplementary Tables S1, S2. Antimicrobial susceptibilities were determined for a total of 325 bacterial strains, including 141 isolates of V. vulnificus (19 clinical, 122 environmental) and 184 isolates of V. cholerae non-O1/non-O139 (18 clinical, 131 environmental, 35 retail). The majority of environmental strains were isolated by health authorities during the German research programs KLIWAS3 and VibrioNet4 between 2004 and 2014. Water and sediment samples were mostly collected at bathing sites along the Baltic Sea and North Sea coastline as well as within the estuaries of the rivers Ems and Weser (Böer et al., 2012). Environmental isolates from bivalve mollusks were obtained from coastal areas of the North Sea. Isolates from retail samples were collected by health authorities of Germany and sent to the National Reference Laboratory for Monitoring Bacteriological Contamination of Bivalve Mollusks of the Federal Institute for Risk Assessment (BfR), Germany. Clinical V. vulnificus and V. cholerae non-O1/non-O139 isolates were characterized in previous studies (Bier et al., 2013; Schirmeister et al., 2014).
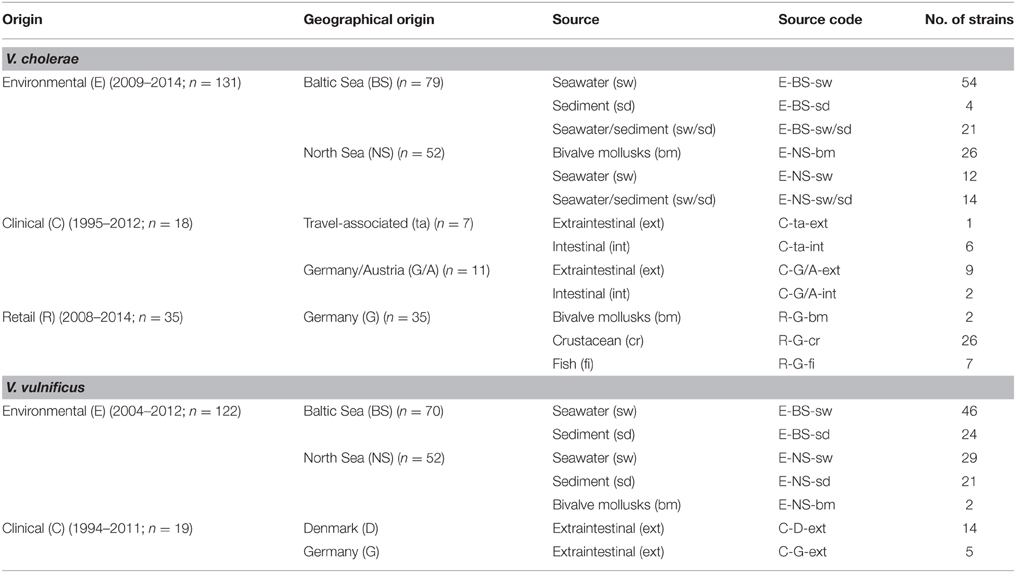
Table 1. Origin and source of V. cholerae non-O1/non-O139 (n = 184) and V. vulnificus (n = 141) strains included in the study.
DNA Extraction
DNA extraction was performed with two methods that are equally applicable for Vibrio species. Genomic DNA of V. vulnificus isolates was extracted from 1 ml of an overnight culture using the RTP Bacteria DNA Kit according to the manufacturer's protocol (STRATEC Biomedical AG, Birkenfeld, Germany).
Genomic DNA of V. cholerae strains was extracted using a boiling method: 1.5 ml of an overnight culture were centrifuged at 14,000 g for 4 min. The cell pellet was suspended in 300 μl TE buffer (10 mM Tris-HCl, 0.5 mM EDTA, pH 8), boiled for 10 min at 95°C, and subsequently cooled on ice. After centrifugation at 14,000 g for 2 min, a 200 μl aliquot of the supernatant was transferred to a new sterile tube. DNA preparations were stored at −20°C.
Species Confirmation
Species confirmation of all V. vulnificus and V. cholerae strains was carried out by species-specific toxR PCR amplification as previously described (Bauer and Røervik, 2007) and in parallel by MALDI-TOF MS analysis. MALDI-TOF MS analysis was performed using a Microflex LT system mass spectrometer (Bruker Daltonik, Bremen, Germany) following the manufacturer's settings. MALDI spectra were obtained by the direct transfer method according to the manufacturer's protocol as previously described (Schirmeister et al., 2014).
Characterization of V. cholerae Isolates
V. cholerae isolates were characterized and subtyped via multiplex PCR targeting rfb sequences specific for O1 and O139 serogroups, toxR, and ctxA. PCR amplification was performed in a final volume of 25 μl with 1x PCR buffer (3 mM MgCl2), 0.2 mM of each deoxynucleoside triphosphate, 0.5 μM of O1 rfb primers, 0.125 μM of O139 rfb, toxR, and ctxA primers, 1.5 U DreamTaq DNA polymerase (Thermo Fisher Scientific Biosciences GmbH, St. Leon-Rot, Germany), and 2 μl of genomic DNA. After an initial denaturation step at 94°C for 4 min, the cycling conditions were the following: 30 cycles at 94°C for 30 s, 59°C for 30 s, 72°C for 30 s, followed by a final extension step at 72°C for 5 min.
Antimicrobial Susceptibility Testing
Antimicrobial susceptibility to the following 13 antimicrobial agents was determined by broth microdilution according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2012a) using custom-defined microtiter plates (EUMVS2, Trek Diagnostic Systems, East Grinstead, United Kingdom): ampicillin, ceftazidime, cefotaxime, chloramphenicol, ciprofloxacin, colistin, florfenicol, gentamicin, kanamycin, nalidixic acid, streptomycin, tetracycline, and trimethoprim. Test ranges are shown in Supplementary Table S3. Additionally, all isolates were tested for their susceptibility to amoxicillin-clavulanic acid (20/10 μg), cefepime (30 μg), imipenem (10 μg), levofloxacin (5 μg), meropenem (10 μg), and sulfamethoxazole-trimethoprim (23.75/1.25 μg) by the disk diffusion method, according to the guidelines of the CLSI using commercially available disks (Oxoid GmbH, Wesel, Germany; CLSI, 2012b).
Strains showing non-susceptibility to imipenem (zone diameter ≤ 19) were tested against an additional panel of β-lactams by broth microdilution (imipenem, ertapenem, cefepime, cefoxitin, temocillin; EUVSEC2, Trek Diagnostic Systems) or disk diffusion (aztreonam, 30 μg).
Following the guidelines of the CLSI, tests were performed with Mueller-Hinton agar and cation-adjusted Mueller-Hinton broth without supplementation of additional sodium chloride (CLSI, 2010a). Escherichia coli ATCC 25922 was used for quality assurance. Minimal inhibitory concentration (MIC) values and inhibition zone diameters of all strains are listed in Supplementary Tables S1, S2. Results were interpreted using the criteria summarized in Supplementary Table S3. In general, results were interpreted according to CLSI clinical breakpoints specific for Vibrio spp. (CLSI, 2010a), which derived from breakpoints for Enterobacteriaceae (CLSI, 2010b). In cases where CLSI breakpoints for Vibrio spp. were obsolete or not available, the latest CLSI breakpoints for Enterobacteriaceae were used: document M100-S25 for aztreonam, cefepime, ertapenem, gentamicin, kanamycin, imipenem, meropenem, nalidixic acid, and trimethoprim (CLSI, 2015); document Vet01-S2 for florfenicol (CLSI, 2013). Other interpretive criteria were used for colistin (EUCAST clinical breakpoints for Enterobacteriaceae5; EUCAST, 2015), temocillin (BSAC interpretive criteria for systemic infections; Andrews, 2009), and streptomycin (based on different studies of Vibrio spp. and E. coli; National Food Institute, 2013; Shaw et al., 2014), as no CLSI breakpoints were available.
Molecular Investigation of Resistance Determinants
PCR amplification was conducted to detect specific antimicrobial resistance determinants depending on the phenotype found. Non-susceptible isolates were screened for genes mediating resistance to streptomycin (aadA1, aadA2, and strA/B) and β-lactams (blaPSE−1, blaOXA−1−like, blaTEM−1−like) that are widespread in Enterobacteriaceae and other Gram-negative bacteria. Specifically imipenem-resistant strains (zone diameter ≤ 19) were tested for the presence of several carbapenemase and AmpC β-lactamase encoding genes. Presence of class 1 integrons was investigated by PCR amplification of the corresponding integrase gene intI1 in all β-lactam and streptomycin non-susceptible strains.
Standard PCR reactions were performed using a Mastercycler EP gradient (Eppendorf, Hamburg, Germany) in a volume of 25 μl with 1x PCR buffer (2 mM MgCl2), 0.2 mM of each deoxynucleoside triphosphate (dNTP), 0.2 μM of each primer, 1.5 U DreamTaq DNA polymerase, and 2 μl of genomic DNA. After an initial denaturation step at 94°C for 4 min, the cycling conditions were the following: 30 cycles of denaturation at 94°C for 30 s, primer annealing for 30 s, and extension at 72 °C for 1 min per kb, followed by a final extension step at 72°C for 10 min.
All primer pairs, target genes, corresponding annealing temperatures, and amplicon sizes are listed in Supplementary Table S4. Enterobacterial strains carrying class 1 integrons and investigated resistance determinants (aadA1, aadA2, blaPSE−1, blaOXA−1, blaTEM−1, blaNDM−1, blaIMP−1, blaVIM−2, blaKPC−3, blaOXA−48, blaACC−1, blaCMY−1, blaCMY−2, blaDHA−1, blaACT−1, blaFOX−1, strA, and strB), as well as one V. cholerae non-O1/non-O139 isolate carrying a class 1 integron with the aadA1 gene were used as positive controls. Susceptible V. cholerae and V. vulnificus strains were included as negative controls.
All streptomycin resistant V. cholerae (VN-3469, VN-5095, VN-10191, and VN-10192) and V. vulnificus (VN-0098, VN-0100, VN-0125, VN-0129) isolates were further examined for mutations in the rpsL gene encoding ribosomal protein S12. Streptomycin susceptible (V. cholerae: VN-0298, VN-2997, VN-3955, VN-4226, and V. vulnificus: VN-0096, VN-0274, VN-3368) and intermediate resistant strains (V. cholerae: VN-3944, VN-4261, and V. vulnificus: VN-3418, VN-3981, VN-10121) were included as controls. For specific amplification and sequencing of the whole rpsL gene in V. cholerae and V. vulnificus, two primer pairs Vc-rpsL-F/Vc-rpsL-R and Vv-rpsL-F/Vv-rpsL-R were designed based on published genome sequences (V. cholerae strains NIH41, N16961, O395, and V. vulnificus strains CMCP6, YJ016, MO6-24/O). Purification of PCR products was performed using the MSB® Spin PCRapace Kit (STRATEC Biomedical AG, Berlin, Germany). Sequencing was conducted on both strands through sequencing service (Eurofins MWG GmbH, Ebersberg, Germany). Electropherograms were assembled and trimmed using SeqMan Pro (v12; DNASTAR Lasergene, Madison, Wisconsin). Sequences were analyzed and compared to the sequences of reference and control strains using Accelrys Gene (v2.5, Accelrys Inc., San Diego, California).
Test for Carbapenemase Activity: Carba NP Test II/Blue-CARBA Test
Strains non-susceptible to imipenem (zone diameter ≤ 19) were grown overnight at 37°C on chromID™ CARBA (bioMérieux, Nürtingen, Germany). Bacterial colonies were subsequently tested for carbapenemase activity with the improved Carba NP test II (Dortet et al., 2014) using 12 g/L imipenem/cilastatin (Zienam®, MSD SHARP, and DOHME GMBH, Haar, Germany) and two calibrated loops (10 μl) as bacterial inoculum to increase enzyme quantity. The Blue-CARBA test was performed as described (Pires et al., 2013) and in addition analogously to the Carba NP test II with two loops of bacterial colonies in 200 μl of the test solution in microcentrifuge tubes and supplementation of tazobactam or EDTA to inhibit class A or metallo-carbapenemases, respectively. Two E. coli strains positive for NDM-1 and KPC-2, respectively, as well as a KPC-3-positive Klebsiella pneumoniae strain served as positive controls.
Statistical Analyses
Descriptive statistics were used to analyze resistance prevalence to different antimicrobial agents (Table 2). Chi-square test for independence was applied with 2 × 2 contingency tables to test if observed differences displayed in Table 3 were statistically significant (P-values ≤ 0.05). MIC50 and MIC90 were defined as the concentration at which growth of 50 and 90% of the isolates was inhibited, respectively.
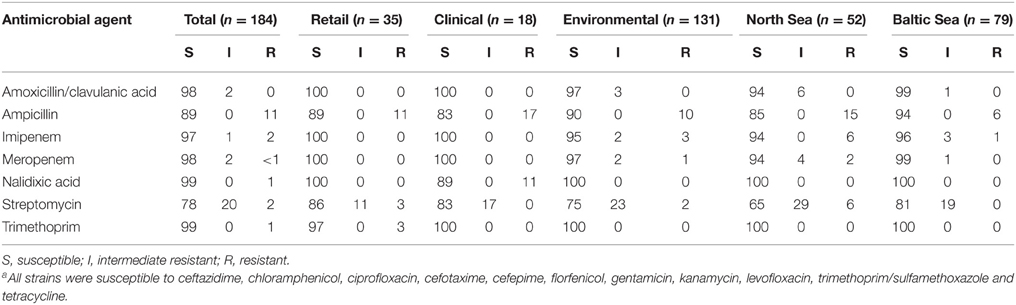
Table 2. Susceptibility vs. resistance occurrence (%) found among V. cholerae non-O1/non-O139 isolates from different originsa.
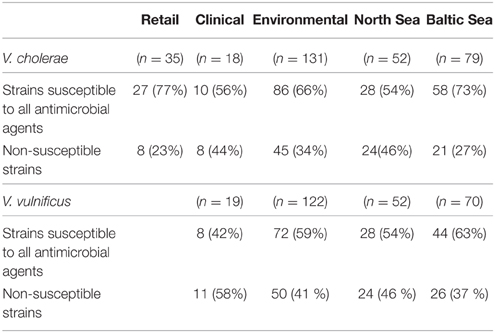
Table 3. Overall resistance occurrence in V. cholerae and V. vulnificus isolates with respect to different origins.
Results
Antimicrobial Susceptibility of Clinical and Environmental V. vulnificus Isolates
Fifty-seven per cent of all examined V. vulnificus isolates showed susceptibility to all antimicrobial agents tested (with the exception of colistin). All 141 V. vulnificus isolates, regardless of their origin were susceptible to quinolones, fluoroquinolones, phenicols, tetracyclines, folate pathway inhibitors, aminopenicillins with or without β-lactamase inhibitors, carbapenems, and third- and fourth- generation cephalosporins. The clinically relevant agents cefotaxime, ceftazidime, and tetracycline were among the most effective antimicrobial agents in vitro as they showed MIC90 values identical to the lowest concentration tested (Table 4). Non-susceptibility was exclusively observed toward aminoglycosides with 40 and 3% of all strains showing intermediate resistance (MIC 32 mg/L) and resistance (MIC 64 mg/L) to streptomycin, respectively. One clinical isolate was intermediate resistant to kanamycin (MIC 32 mg/L) while all strains were susceptible to gentamicin. The percentage of non-susceptible strains with respect to different origins is shown in Table 3. No significant difference was observed between clinical (n = 19) and environmental (n = 122) isolates, nor was there a significant difference between isolates from the Baltic Sea and the North Sea (p > 0.05, χ2). None of the examined gene determinants encoding streptomycin resistance (aadA1, aadA2, strA/B), nor class 1 integrons were detected in streptomycin non-susceptible V. vulnificus isolates. Sequence analysis of the rpsL gene revealed that all four streptomycin resistant V. vulnificus isolates (VN-0098, VN-0100, VN-0125, VN-0129) as well as two susceptible (VN-0274, VN-3368) and two intermediate resistant isolates (VN-3918, VN-10121) carried one silent point mutation A-291-T compared to the three reference strains (CMCP6, YJ016, and MO6-24/O). An additional silent mutation C-351-T within the rpsL gene was observed in strain VN-0100.
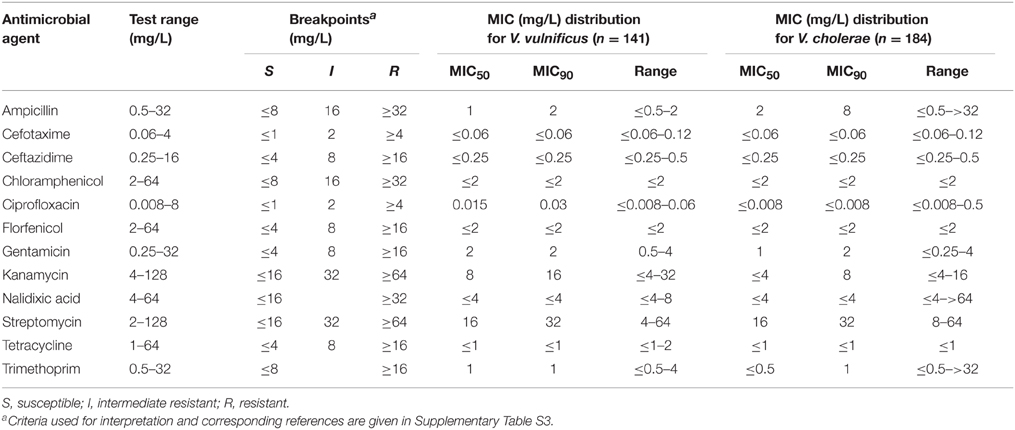
Table 4. Antimicrobial MIC distributions for the V. cholerae and V. vulnificus isolates tested.
Antimicrobial Susceptibility of V. cholerae non-O1/non-O139 Isolated from Clinical, Environmental and Seafood Samples
All 184 isolates investigated in this study were confirmed to be non-toxigenic V. cholerae non-O1/non-O139 isolates. The majority of isolates (67%) were susceptible to all antimicrobial agents tested (with the exception of colistin). Eighteen per cent of the strains showed intermediate resistance to one or two antimicrobial agents (mostly to streptomycin) and the remaining strains (15%) showed full resistance to at least one antimicrobial agent. None of the V. cholerae isolates showed multidrug-resistance, defined as resistance to three or more classes of antimicrobial agents (Chen et al., 2010). Resistance profiles are shown in Supplementary Table S2, while resistance occurrence is given in Table 2.
As observed among V. vulnificus, all V. cholerae strains were susceptible to ciprofloxacin, chloramphenicol, florfenicol, cefotaxime, sulfamethoxazole-trimethoprim, levofloxacin, ceftazidime, cefepime, gentamicin, kanamycin, and tetracycline. Additionally, 98% of the isolates were susceptible to amoxicillin/clavulanic acid. The most effective clinically relevant agents in vitro were ciprofloxacin, cefotaxime, ceftazidime, and tetracycline as they showed MIC90 values identical to the lowest concentration tested (Table 4).
Similar to the V. vulnificus isolates, a small proportion of V. cholerae strains showed resistance to streptomycin (2%), while 20% of the strains were intermediate resistant. In contrast to V. vulnificus, the most frequent antimicrobial resistance found among all V. cholerae isolates was resistance to ampicillin (11%). Resistance to nalidixic acid and trimethoprim was rarely observed in two clinical isolates and in one isolate from seafood. Non-susceptibility to the carbapenems imipenem and meropenem was observed in 5 and 3% of the environmental isolates, respectively.
Clinical strains showed the highest percentage of non-susceptible strains (44%), followed by environmental strains (34%) and by strains isolated from retail seafood (23%) (Table 3). However, statistical analysis revealed that the observed differences to environmental isolates are not significant (p > 0.05, χ2). Comparison between the geographical origin of environmental strains revealed that the percentages of strains non-susceptible to streptomycin, ampicillin, amoxicillin/clavulanic acid, meropenem, and imipenem were higher in the North Sea compared to the Baltic Sea (Table 2). The higher occurrence of non-susceptible V. cholerae strains in the North Sea (Table 3) was statistically significant (χ2 = 5.327, d.f. = 1, p < 0.05).
Analysis of MIC distributions revealed that susceptibilities of the two Vibrio species were rather similar (Table 4). Differences were exclusively observed for kanamycin and ciprofloxacin, where MIC90 values of V. cholerae were two and four times lower and in case of ampicillin four times higher than those for V. vulnificus.
Susceptibility to colistin was excluded from any statistical analysis and tables, since V. vulnificus and V. cholerae possess an intrinsic resistance to colistin, which is used for selective growth on cellobiose-polymyxin B-colistin agar (Massad and Oliver, 1987). However, five V. cholerae strains were highly susceptible to colistin (MICs ≤ 2 mg/L) and would therefore fail to grow on this selective agar.
Neither class 1 integrons, nor gene determinants encoding streptomycin resistance (aadA1, aadA2, strA/B) were detected among non-susceptible V. cholerae isolates. Sequence analysis of the rpsL gene revealed three silent point mutations C-198-T, A-251-T, and G-360-A in two streptomycin resistant V. cholerae isolates (VN-10191, VN-10192) as well as within a susceptible isolate (VN-4226). In addition, PCR amplifications to detect β-lactamase genes (blaPSE−1, blaOXA−1−like, blaTEM−1−like) were negative in all tested V. cholerae isolates.
Examination of Carbapenem Non-susceptible V. cholerae Isolates
Among the 131 environmental V. cholerae non-O1/non-O139 isolates analyzed, resistance to the carbapenem imipenem (zone diameter ≤ 19) was observed in four strains (VN-2808, VN-2825, VN-2923, and VN-2997). These strains additionally showed resistance to ampicillin, intermediate resistance to amoxicillin-clavulanic acid, as well as intermediate or full resistance to meropenem (Table 5). In contrast, they were susceptible to the third- and fourth-generation cephalosporins ceftazidime, cefotaxime, and cefepime. Further characterization revealed resistance to aztreonam and ertapenem, while the strains were susceptible to temocillin and intermediate resistant to cefoxitin. The four strains grew on chromID™ CARBA agar, while growth was inhibited on chromID™ OXA-48 (bioMérieux GmbH, Nürtingen, Germany), indicating the expression of carbapenem-hydrolyzing enzymes other than OXA-48 type carbapenemases. To further investigate the presence of carbapenemases, Blue-CARBA and Carba NP II tests were conducted on these strains. Imipenem-hydrolyzing activity was detected in intact cells and crude cell extracts of each of the four strains. This activity was inhibited by tazobactam but not by EDTA, suggesting the presence of Ambler class A carbapenemases rather than class B metallo-carbapenemases or class D OXA-carbapenemases (Dortet et al., 2012).
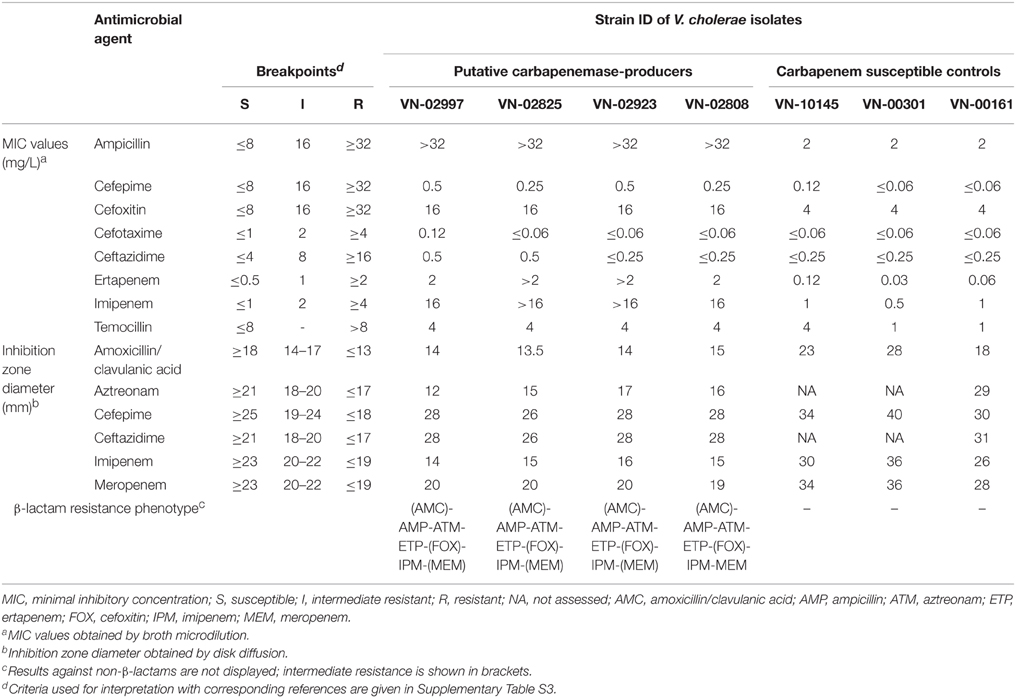
Table 5. β-lactam MIC values and inhibition zone diameters found in the putative carbapenemase-producers and in carbapenem susceptible isolates selected as negative controls.
PCRs to identify genes encoding Ambler class A carbapenemases were performed. However, no products with expected sizes were observed using primers for detection of NMC-A, SME 1-3, IMI 1-3, or KPC 1-5. In the case of NMC-A, SME, and IMI, a general failure of PCR amplification cannot be excluded as no positive control strains were available.
In addition, the strains were negative for PCR-amplification of specific genes encoding Ambler class B metallo-carbapenemases (VIM 1-2, IMP, NDM-1), class D carbapenemase (OXA-48), and AmpC-β-lactamases (MOX 1-2, CMY 1-11, LAT 1-4, BIL-1, DHA 1-2, ACC, MIR-1T, ACT-1, FOX 1-5b) as well as of class 1 integrons.
Discussion
Prevalence of Antimicrobial Resistance in V. vulnificus and V. cholerae non-O1/non-O139
In this study, V. vulnificus isolates from German coastal waters as well as of clinical origin were susceptible to quinolones, fluoroquinolones, phenicols, tetracyclines, folate pathway inhibitors, aminopenicillins with or without β-lactamase inhibitors, carbapenems, and third- and fourth-generation cephalosporins. Non-susceptibility was exclusively observed toward aminoglycosides; predominantly streptomycin and sporadically kanamycin. Similar observations were made by Han et al. (2007), who reported total susceptibility with comparable MIC90 values to chloramphenicol, ampicillin, ceftazidime, cefotaxime, ciprofloxacin, gentamicin, and tetracycline among V. vulnificus isolates from oysters of the Louisiana Gulf coast, USA (Han et al., 2007). In a recent study of V. vulnificus isolates from the Chesapeake Bay, USA (Shaw et al., 2014), the highest percentage of resistance was also observed against streptomycin. However, a large percentage of intermediate resistant strains to chloramphenicol (78%) and sporadically non-susceptibility to β-lactams was also reported in that study (Shaw et al., 2014). Compared to our study, higher percentages of non-susceptible strains to ampicillin, tetracycline, nalidixic acid, trimethoprim, and especially to the aminoglycosides streptomycin and gentamicin were observed in a study of 151 V. vulnificus isolates from South Carolina, USA, while resistance to chloramphenicol and meropenem was under one per cent (Baker-Austin et al., 2009).
Among V. cholerae non-O1/non-O139 isolated from clinical, environmental, and seafood samples in Germany, no multidrug-resistance was observed and the majority (67%) of isolates were susceptible to all antimicrobial agents tested. Full resistance was most frequently found toward ampicillin (11%) and streptomycin (2%). In addition, a considerable proportion of isolates showed intermediate resistance to streptomycin (20%). Resistance to nalidixic acid and trimethoprim was only sporadically found in isolates from clinical and seafood samples, respectively. While numerous studies on antimicrobial resistance of toxigenic V. cholerae O1, O139 strains have been published, data on V. cholerae non-O1/non-O139 are less frequent. A recent study showed similar antimicrobial resistance patterns among environmental V. cholerae non-O1/non-O139 isolates from the Chesapeake Bay, USA (Ceccarelli et al., 2015). No multidrug-resistant isolates were detected and resistance to β-lactams was found in some isolates. In one large-scale study from India on antimicrobial susceptibility of V. cholerae non-O1/non-O139 isolates, the highest percentage of resistant strains was also seen for ampicillin (88%) and streptomycin (85%) though with a considerably higher frequency (Kumar et al., 2009). In contrast to our study, Kumar et al. (2009) reported a high prevalence of multidrug resistance and only a small percentage of strains were susceptible to all ten antimicrobial agents tested (12%).
Several PCR analyses were performed to reveal the underlying molecular mechanisms responsible for the observed non-susceptibility to streptomycin, ampicillin, and imipenem.
Resistance to streptomycin is often mediated by enzymatic inactivation through adenylation by aminoglycoside (3″) adenyltransferases (aadA genes) or through phosphorylation by aminoglycoside phosphotransferases (strA/strB genes; Shaw et al., 1993; Tsai et al., 2014). However, none of the Vibrio isolates was positive for amplification of aadA1, aadA2, or strA/B genes that are commonly found in Enterobacteriaceae and that have already been identified in V. cholerae (Hochhut et al., 2001; Sá et al., 2010; Yu et al., 2012). Ribosomal alterations resulting from mutations in the rpsL gene encoding ribosomal protein S12 can be another cause of streptomycin resistance (Shaw et al., 1993; Tsai et al., 2014). However, amino acid sequences of ribosomal protein S12 were identical in all investigated strains, irrespective of the streptomycin resistance phenotype. Observed single nucleotide polymorphisms within the rpsL gene of some resistant as well as of some susceptible isolates were silent mutations. This indicates that other streptomycin inactivating enzymes or other resistance mechanisms, such as decreased permeability or other ribosomal alterations (e.g., mutations in the rrs gene encoding the 16S ribosomal RNA) may be responsible for the observed phenotype (Shaw et al., 1993; Tsai et al., 2014).
Likewise, all β-lactam non-susceptible Vibrio isolates were negative for amplification of blaPSE−1, blaOXA−1−like, and blaTEM−1−like genes encoding common β-lactamases in Enterobacteriaceae and other Gram-negative bacteria. This suggests that resistance may be encoded by other β-lactamase genes which are known to show a high diversity in Gram-negative bacteria. β-lactam resistance may also be mediated by other mechanisms such as reduced permeability, increased efflux, or target alterations (e.g., reduced affinity or increased amount of penicillin-binding protein (PBP); Foster, 1983).
Carbapenemase Producing V. cholerae
Antimicrobial susceptibility patterns as well as growth patterns on different selective media indicated the presence of a β-lactamase with carbapenem hydrolyzing activity in four environmental V. cholerae non-O1/non-O139 isolates from the Baltic Sea and the North Sea. The expression of carbapenemases was confirmed by positive Blue-CARBA and positive Carba NP II tests, which specifically detect imipenem hydrolyzing activity (Dortet et al., 2012; Pires et al., 2013). Inhibition of carbapenemase activity by tazobactam but not by EDTA suggested the presence of Ambler class A carbapenemases rather than class B metallo-carbapenemases or class D OXA-carbapenemases (Dortet et al., 2012). However, we found no evidence for the presence of specific genes encoding Ambler class A carbapenemases.
The four strains showed an exceptional resistance profile: They were non-susceptible to aminopenicillins, carbapenems, cefoxitin, and aztreonam and were only slightly inhibited by the β-lactamase inhibitor clavulanic acid. However, they were fully susceptible to third- and fourth-generation cephalosporins as well as to temocillin. The observed resistance to aminopenicillins coupled with susceptibility to extended spectrum cephalosporins may indicate the presence of an OXA-type carbapenemase (Ambler class D) reviewed by Walther-Rasmussen and Høiby (2006). However, with some exceptions, e.g., OXA-23, these enzymes are generally not inhibited by tazobactam, as seen for the four strains in the Carba NP II and Blue-CARBA tests and generally don't mediate resistance to aztreonam (Walther-Rasmussen and Høiby, 2006).
So far, the identity of the enzyme responsible for imipenem hydrolyzing activity in the four strains remains unclear, as none of the examined carbapenemase and AmpC-β-lactamase genes could be detected. It cannot be excluded that in addition to a carbapenem hydrolyzing enzyme other resistance mechanisms, such as reduced affinity of PBPs, porin alterations resulting in decreased membrane permeability or active efflux systems, either alone or in combination may also contribute to the observed phenotype (Walther-Rasmussen and Høiby, 2006; Queenan and Bush, 2007; Nordmann et al., 2012). Carbapenem-resistant V. cholerae have already been reported in other studies. NDM-1 carbapenemase was detected in a V. cholerae O1 El Tor Ogawa strain isolated from a 2-year old child (Mandal et al., 2012) as well as in V. cholerae isolated from seepage water in India (Walsh et al., 2011). Furthermore, increasing resistance to carbapenems was recently described among V. cholerae O1 or O139 strains isolated between 1986 and 2012 in southwest China (Gu et al., 2014).
Concluding remarks
In this study, antimicrobial agents recommended as first choice agents for the treatment of V. vulnificus and V. cholerae non-O1/non-O139 infections such as fluoroquinolones, tetracyclines, and extended spectrum cephalosporins were found to be effective in vitro against both species. However, the administration of aminopenicillins, carbapenems, or aminoglycosides for treatment of V. cholerae non-O1/non-O139 infections, which has been reported in few studies (Daniels and Shafaie, 2000; Feghali and Adib, 2011; Lu et al., 2014) should be considered carefully, as non-susceptibility was most frequently observed against ampicillin and streptomycin and sporadically to carbapenems. For V. vulnificus, non-susceptibility was exclusively observed to the aminoglycosides streptomycin and kanamycin. However, gentamicin was effective against both species and could be an aminoglycoside of choice for the treatment of children and pregnant woman, as was also suggested by others (Shaw et al., 2014).
We report the detection of carbapenemase producing V. cholerae from different locations of the German coast line (North Sea and Baltic Sea) representing an environmental reservoir of carbapenem resistance. An entry into the sea resulting from sanitary pollution or human recreational activities cannot be excluded, but seems not likely as vibrios are indigenous bacteria of the marine environment and not intestinal commensals of humans or terrestrial animals. The strains displaying carbapenemase activity showed resistance to an unusual pattern of β-lactams. Therefore, characterization of the underlying genetic background is necessary to identify the responsible genes e.g., using whole genome sequencing as the most promising approach. Further investigations on the mobility as well as on the location of encoding genes are also needed, since location on mobile genetic elements would imply a higher risk for interspecies spread. Carbapenems are last line antimicrobial agents for treatment of multidrug-resistant Gram-negative bacteria and are of high therapeutic value (Nordmann et al., 2012). The occurrence of putative carbapenemase producing V. cholerae in the North and Baltic Sea is therefore of great concern and highlights the need for systematic monitoring of antimicrobial susceptibility in potentially pathogenic Vibrio spp. in Europe.
Funding
This work was supported by the Federal Ministry of Education and Research (VibrioNet, BMBF grant 01KI1015A) and the Federal Institute for Risk Assessment (BfR 46-001). The German research program KLIWAS was funded by the Federal Ministry of Transport and Digital Infrastructure.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Dr. G. Hauk and Dr. O. Duty (Governmental Institute of Public Health and Social Affairs of Mecklenburg-Western Pomerania) as well as Dr. N. Brennholt and Dr. S. I. Böer (German Federal Institute of Hydrology), Dr. K. Luden and Dr. E.-A. Heinemeyer (Governmental Institute for Public Health of Lower Saxony) are greatly acknowledged for providing Vibrio isolates collected within the German research program KLIWAS. We thank Dr. A. Käsbohrer (Federal Institute for Risk Assessment, BfR) for providing laboratory facilities and A. Schabanowski, S. Schmoger, and T. Skladnikiewicz-Ziemer (BfR) for introduction into techniques of antimicrobial resistance testing. We further thank Dr. B.-A. Tenhagen (BfR) and Dr. B. Malorny (BfR) for advice.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2015.01179
Footnotes
1. ^http://www.cdc.gov/cholera/non-01-0139-infections.html
2. ^http://emergency.cdc.gov/disasters/vibriovulnificus.asp
3. ^http://www.kliwas.de/KLIWAS/EN/03_ResearchTasks/03_vh3/04_304/304_node.html
References
Andrews, J. M. (2009). BSAC standardized disc susceptibility testing method (version 8). J. Antimicrob. Chemother. 64, 454–489. doi: 10.1093/jac/dkp244
Baker-Austin, C., McArthur, J. V., Lindell, A. H., Wright, M. S., Tuckfield, R. C., Gooch, J., et al. (2009). Multi-site analysis reveals widespread antibiotic resistance in the marine pathogen Vibrio vulnificus. Microb. Ecol. 57, 151–159. doi: 10.1007/s00248-008-9413-8
Baker-Austin, C., Trinanes, J. A., Taylor, N. G. H., Hartnell, R., Siitonen, A., and Martinez-Urtaza, J. (2012). Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Change 3, 73–77. doi: 10.1038/nclimate1628
Bauer, A., and Røervik, L. M. (2007). A novel multiplex PCR for the identification of Vibrio parahaemolyticus, Vibrio cholerae and Vibrio vulnificus. Lett. Appl. Microbiol. 45, 371–375. doi: 10.1111/j.1472-765X.2007.02195.x
Bier, N., Bechlars, S., Diescher, S., Klein, F., Hauk, G., Duty, O., et al. (2013). Genotypic diversity and virulence characteristics of clinical and environmental Vibrio vulnificus isolates from the baltic sea region. Appl. Environ. Microbiol. 79, 3570–3581. doi: 10.1128/AEM.00477-13
Böer, S., Hauk, G., Duty, O., Luden, K., Heinemeyer, E.-A., and Brennholt, N. (2012). “Pathogenic Vibrio species in German coastal waters of the North Sea and the Baltic Sea – a comparison,” in Veranstaltungen International Symposium “Pathogenic Vibrio spp.in Northern European Waters” (Koblenz: German Federal Institute of Hydrology), 36–42. doi: 10.5675/BfG_Veranst_2012.4
Ceccarelli, D., Chen, A., Hasan, N. A., Rashed, S. M., Huq, A., and Colwell, R. R. (2015). Non-O1/non-O139 Vibrio cholerae carrying multiple virulence factors and V. cholerae O1 in the Chesapeake Bay, Maryland. Appl. Environ. Microbiol. 81, 1909–1918. doi: 10.1128/AEM.03540-14
Chen, S. C., Lee, Y. T., Tsai, S. J., Chan, K. S., Chao, W. N., Wang, P. H., et al. (2012). Antibiotic therapy for necrotizing fasciitis caused by Vibrio vulnificus: retrospective analysis of an 8 year period. J. Antimicrob. Chemother. 67, 488–493. doi: 10.1093/jac/dkr476
Chen, X., Naren, G. W., Wu, C. M., Wang, Y., Dai, L., Xia, L. N., et al. (2010). Prevalence and antimicrobial resistance of Campylobacter isolates in broilers from China. Vet. Microbiol. 144, 133–139. doi: 10.1016/j.vetmic.2009.12.035
CLSI (2010a). Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria; Approved Guideline —Second Edition M45-A2. Wayne, PA: Clinical and Laboratory Standards Institute, CLSI.
CLSI (2010b). Performance Standards for Antimicrobial Susceptibility Testing; Twentieth Informational Supplement M100-S20. Wayne, PA: Clinical and Laboratory Standards Institute, CLSI.
CLSI (2012a). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard—Ninth Edition M7-A9. Wayne, PA: Clinical and Laboratory Standards Institute, CLSI.
CLSI (2012b). Performance Standards for Antimicrobial Disk Susceptibility Tests; Approved Standard—Eleventh Edition M2-A11. Wayne, PA: Clinical and Laboratory Standards Institute, CLSI.
CLSI (2013). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals; Second Informational Supplement VET01-S2. Wayne, PA: Clinical and Laboratory Standards Institute, CLSI.
CLSI (2015). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-fifth Informational Supplement M100-S25. Wayne, PA: Clinical and Laboratory Standards Institute, CLSI.
Daniels, N. A., and Shafaie, A. (2000). A review of pathogenic Vibrio infections for clinicians. Infect. Med. 17, 665–685.
Dortet, L., Bréchard, L., Cuzon, G., Poirel, L., and Nordmann, P. (2014). Strategy for rapid detection of carbapenemase-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 58, 2441–2445. doi: 10.1128/AAC.01239-13
Dortet, L., Poirel, L., and Nordmann, P. (2012). Rapid identification of carbapenemase types in Enterobacteriaceae and Pseudomonas spp. By using a biochemical test. Antimicrob. Agents. Chemother. 56, 6437–6440. doi: 10.1128/AAC.01395-12
EUCAST (2015). The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 5.0. Available online at: http://www.eucast.org.
Farmer, J. J. III, and Janda, J. M. (2004). “Family I. Vibrionaceae,” in Bergey's Manual of Systematic Bacteriology, 2nd Edn., ed G. M. Garrity (New York, NY: Springer), 491–546.
Feghali, R., and Adib, S. M. (2011). Two cases of Vibrio cholerae non-O1/non-O139 septicaemia with favourable outcome in Lebanon. East. Mediterr. Health J. 17, 722–724.
Foster, T. J. (1983). Plasmid-determined resistance to antimicrobial drugs and toxic metal ions in bacteria. Microbiol. Rev. 47, 361–409.
Gu, W., Yin, J., Yang, J., Li, C., Chen, Y., Yin, J., et al. (2014). Characterization of Vibrio cholerae from 1986 to 2012 in Yunnan Province, southwest China bordering Myanmar. Infect. Genet. Evol. 21, 1–7. doi: 10.1016/j.meegid.2013.10.015
Han, F., Walker, R. D., Janes, M. E., Prinyawiwatkul, W., and Ge, B. (2007). Antimicrobial susceptibilities of Vibrio parahaemolyticus and Vibrio vulnificus isolates from Louisiana Gulf and retail raw oysters. Appl. Environ. Microbiol. 73, 7096–7098. doi: 10.1128/AEM.01116-07
Hochhut, B., Lotfi, Y., Mazel, D., Faruque, S. M., Woodgate, R., and Waldor, M. K. (2001). Molecular analysis of antibiotic resistance gene clusters in Vibrio cholerae O139 and O1 SXT constins. Antimicrob. Agents Chemother. 45, 2991–3000. doi: 10.1128/AAC.45.11.2991-3000.2001
Huehn, S., Eichhorn, C., Urmersbach, S., Breidenbach, J., Bechlars, S., Bier, N., et al. (2014). Pathogenic vibrios in environmental, seafood and clinical sources in Germany. Int. J. Med. Microbiol. 304, 843–850. doi: 10.1016/j.ijmm.2014.07.010
Huhulescu, S., Indra, A., Feierl, G., Stoeger, A., Ruppitsch, W., Sarkar, B., et al. (2007). Occurrence of Vibrio cholerae serogroups other than O1 and O139 in Austria. Wien. Klin. Wochenschr. 119, 235–241. doi: 10.1007/s00508-006-0747-2
Kumar, P. A., Patterson, J., and Karpagam, P. (2009). Multiple antibiotic resistance profiles of Vibrio cholerae non-O1 and non-O139. Jpn. J. Infect. Dis. 62, 230–232.
Letchumanan, V., Chan, K. G., and Lee, L. H. (2014). Vibrio parahaemolyticus: a review on the pathogenesis, prevalence, and advance molecular identification techniques. Front. Microbiol. 5:705. doi: 10.3389/fmicb.2014.00705
Lu, B., Zhou, H., Li, D., Li, F., Zhu, F., Cui, Y., et al. (2014). The first case of bacteraemia due to non-O1/non-O139 Vibrio cholerae in a type 2 diabetes mellitus patient in mainland China. Int. J. Infect. Dis. 25, 116–118. doi: 10.1016/j.ijid.2014.04.015
Luo, Y., Ye, J., Jin, D., Ding, G., Zhang, Z., Mei, L., et al. (2013). Molecular analysis of non-O1/non-O139 Vibrio cholerae isolated from hospitalised patients in China. BMC Microbiol. 13:52. doi: 10.1186/1471-2180-13-52
Lutz, C., Erken, M., Noorian, P., Sun, S., and McDougald, D. (2013). Environmental reservoirs and mechanisms of persistence of Vibrio cholerae. Front. Microbiol. 4:375. doi: 10.3389/fmicb.2013.00375
Mandal, J., Sangeetha, V., Ganesan, V., Parveen, M., Preethi, V., Harish, B. N., et al. (2012). Third-generation cephalosporin-resistant Vibrio cholerae, India. Emerg. Infect. Dis. 18, 1326–1328. doi: 10.3201/eid1808.111686
Mansergh, S., and Zehr, J. P. (2014). Vibrio diversity and dynamics in the Monterey Bay upwelling region. Front. Microbiol. 5:48. doi: 10.3389/fmicb.2014.00048
Massad, G., and Oliver, J. D. (1987). New selective and differential medium for Vibrio cholerae and Vibrio vulnificus. Appl. Environ. Microbiol. 53, 2262–2264.
National Food Institute, T. U. O. D. (2013). DANMAP 2013. Use of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Food Animals, Food and Humans in Denmark.
Nordmann, P., Dortet, L., and Poirel, L. (2012). Carbapenem resistance in Enterobacteriaceae: here is the storm! Trends Mol. Med. 18, 263–272. doi: 10.1016/j.molmed.2012.03.003
Oliver, J. D. (2006). “Vibrio vulnificus,” in The Biology of Vibrios, eds F. L. Thompson, B. Austin, and J. Swings, 349–366. doi: 10.1128/9781555815714.ch25
Petsaris, O., Nousbaum, J. B., Quilici, M. L., Le Coadou, G., Payan, C., and Abalain, M. L. (2010). Non-O1, non-O139 Vibrio cholerae bacteraemia in a cirrhotic patient. J. Med. Microbiol. 59, 1260–1262. doi: 10.1099/jmm.0.021014-0
Pires, J., Novais, A., and Peixe, L. (2013). Blue-carba, an easy biochemical test for detection of diverse carbapenemase producers directly from bacterial cultures. J. Clin. Microbiol. 51, 4281–4283. doi: 10.1128/JCM.01634-13
Queenan, A. M., and Bush, K. (2007). Carbapenemases: the versatile beta-lactamases. Clin. Microbiol. Rev. 20, 440–458. doi: 10.1128/CMR.00001-07
Sá, L. L., Fonseca, E. L., Pellegrini, M., Freitas, F., Loureiro, E. C., and Vicente, A. C. (2010). Occurrence and composition of class 1 and class 2 integrons in clinical and environmental O1 and non-O1/non-O139 Vibrio cholerae strains from the Brazilian Amazon. Mem. Inst. Oswaldo Cruz 105, 229–232. doi: 10.1590/S0074-02762010000200021
Schirmeister, F., Dieckmann, R., Bechlars, S., Bier, N., Faruque, S. M., and Strauch, E. (2014). Genetic and phenotypic analysis of Vibrio cholerae non-O1, non-O139 isolated from German and Austrian patients. Eur. J. Clin. Microbiol. Infect. Dis. 33, 767–778. doi: 10.1007/s10096-013-2011-9
Shapiro, R. L., Altekruse, S., Hutwagner, L., Bishop, R., Hammond, R., Wilson, S., et al. (1998). The role of Gulf Coast oysters harvested in warmer months in Vibrio vulnificus infections in the United States, 1988-1996. Vibrio Working Group. J. Infect. Dis. 178, 752–759. doi: 10.1086/515367
Shaw, K. J., Rather, P. N., Hare, R. S., and Miller, G. H. (1993). Molecular genetics of aminoglycoside resistance genes and familial relationships of the aminoglycoside-modifying enzymes. Microbiol. Rev. 57, 138–163.
Shaw, K. S., Rosenberg Goldstein, R. E., He, X., Jacobs, J. M., Crump, B. C., and Sapkota, A. R. (2014). Antimicrobial susceptibility of Vibrio vulnificus and Vibrio parahaemolyticus recovered from recreational and commercial areas of Chesapeake Bay and Maryland Coastal Bays. PloS ONE 9:e89616. doi: 10.1371/journal.pone.0089616
Thompson, J. R., and Polz, M. F. (2006). “Dynamics of vibrio populations and their role in environmental nutrient cycling,” in The Biology of Vibrios, eds F. L. Thompson, B. Austin, and J. Swings (Washington, DC: ASM Press), 190–203. doi: 10.1128/9781555815714.ch13
Tobin-D'Angelo, M., Smith, A. R., Bulens, S. N., Thomas, S., Hodel, M., Izumiya, H., et al. (2008). Severe diarrhea caused by cholera toxin-producing Vibrio cholerae serogroup O75 infections acquired in the southeastern United States. Clin. Infect. Dis. 47, 1035–1040. doi: 10.1086/591973
Tsai, Y. K., Liou, C. H., Lin, J. C., Ma, L., Fung, C. P., Chang, F. Y., et al. (2014). A suitable streptomycin-resistant mutant for constructing unmarked in-frame gene deletions using rpsL as a counter-selection marker. PLoS ONE 9:e109258. doi: 10.1371/journal.pone.0109258
Walsh, T. R., Weeks, J., Livermore, D. M., and Toleman, M. A. (2011). Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: an environmental point prevalence study. Lancet Infect. Dis. 11, 355–362. doi: 10.1016/S1473-3099(11)70059-7
Walther-Rasmussen, J., and Høiby, N. (2006). OXA-type carbapenemases. J. Antimicrob. Chemother. 57, 373–383. doi: 10.1093/jac/dki482
Keywords: antimicrobial resistance pattern, Baltic Sea, North Sea, carbapenemase, disk diffusion, broth microdilution
Citation: Bier N, Schwartz K, Guerra B and Strauch E (2015) Survey on antimicrobial resistance patterns in Vibrio vulnificus and Vibrio cholerae non-O1/non-O139 in Germany reveals carbapenemase-producing Vibrio cholerae in coastal waters. Front. Microbiol. 6:1179. doi: 10.3389/fmicb.2015.01179
Received: 05 August 2015; Accepted: 12 October 2015;
Published: 28 October 2015.
Edited by:
Learn-Han Lee, Monash University Malaysia, MalaysiaReviewed by:
Weili Liang, Chinese Center for Disease Control and Prevention, ChinaAdrian Canizalez-Roman, Autonomous University of Sinaloa, Mexico
Vengadesh Letchumanan, University of Malaya, Malaysia
Copyright © 2015 Bier, Schwartz, Guerra and Strauch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eckhard Strauch, ZWNraGFyZC5zdHJhdWNoQGJmci5idW5kLmRl