 Ruth Gómez Expósito
Ruth Gómez Expósito Joeke Postma
Joeke Postma Jos M. Raaijmakers
Jos M. Raaijmakers Irene De Bruijn
Irene De Bruijn- 1Department of Microbial Ecology, Netherlands Institute of Ecology (NIOO-KNAW), Wageningen, Netherlands
- 2Laboratory of Phytopathology, Wageningen University and Research Centre, Wageningen, Netherlands
- 3Plant Research International, Wageningen University and Research Centre, Wageningen, Netherlands
The genus Lysobacter includes several species that produce a range of extracellular enzymes and other metabolites with activity against bacteria, fungi, oomycetes, and nematodes. Lysobacter species were found to be more abundant in soil suppressive against the fungal root pathogen Rhizoctonia solani, but their actual role in disease suppression is still unclear. Here, the antifungal and plant growth-promoting activities of 18 Lysobacter strains, including 11 strains from Rhizoctonia-suppressive soils, were studied both in vitro and in vivo. Based on 16S rRNA sequencing, the Lysobacter strains from the Rhizoctonia-suppressive soil belonged to the four species Lysobacter antibioticus, Lysobacter capsici, Lysobacter enzymogenes, and Lysobacter gummosus. Most strains showed strong in vitro activity against R. solani and several other pathogens, including Pythium ultimum, Aspergillus niger, Fusarium oxysporum, and Xanthomonas campestris. When the Lysobacter strains were introduced into soil, however, no significant and consistent suppression of R. solani damping-off disease of sugar beet and cauliflower was observed. Subsequent bioassays further revealed that none of the Lysobacter strains was able to promote growth of sugar beet, cauliflower, onion, and Arabidopsis thaliana, either directly or via volatile compounds. The lack of in vivo activity is most likely attributed to poor colonization of the rhizosphere by the introduced Lysobacter strains. In conclusion, our results demonstrated that Lysobacter species have strong antagonistic activities against a range of pathogens, making them an important source for putative new enzymes and antimicrobial compounds. However, their potential role in R. solani disease suppressive soil could not be confirmed. In-depth omics'–based analyses will be needed to shed more light on the potential contribution of Lysobacter species to the collective activities of microbial consortia in disease suppressive soils.
Introduction
Lysobacter are Gram-negative bacteria widely distributed in diverse ecosystems, including soil, rhizosphere, and freshwater habitats (Reichenbach, 2006). The genus Lysobacter was first described in 1978 by Christensen and Cook and included four species. Lysobacter spp. are closely related to members of the genus Xanthomonas and were initially misclassified as Cytophaga, Sorangium, or Myxobacter (Christensen and Cook, 1978). Currently, 30 Lysobacter species have been taxonomically accepted (for updates see http://www.bacterio.net/lysobacter.html) and new Lysobacter species have been recently identified (Du et al., 2015; Lin et al., 2015; Singh et al., 2015) but are not yet included in the database. Various members of this bacterial genus have activity against a range of other (micro)organisms, including Gram-negative and Gram-positive bacteria, fungi, oomycetes, and nematodes (Reichenbach, 2006). They are well-known for the production of a variety of extracellular enzymes and antimicrobial compounds. Enzymes identified for Lysobacter include chitinases (Zhang and Yuen, 2000; Zhang et al., 2001), glucanases (Palumbo et al., 2005), proteases (Stepnaya et al., 2008; Gökçen et al., 2014; Vasilyeva et al., 2014), lipases (Folman et al., 2003; Ko et al., 2009) as well as elastases, keratinases, phosphatases, endonucleases, endoamylases, and esterases (Reichenbach, 2006). Antimicrobial compounds described for Lysobacter include lysobactin, tripopeptin, xanthobaccin, maltophilin, dihydromaltophilin, phenazine, lactivicin (Xie et al., 2012), HSAF (Li et al., 2008), and WAP-8294A2 (Zhang et al., 2011). Currently, WAP-8294A2 is in phase I/II clinical trials for controlling methicillin-resistant Staphylococcus aureus (anti-MRSA) (Zhang et al., 2011; Wang et al., 2013).
In terms of ecosystem services, Postma et al. (2010a) showed a correlation between the abundance of three Lysobacter species (Lysobacter antibioticus, Lysobacter capsici, and Lysobacter gummosus) in soil and the level of suppressiveness against Rhizoctonia solani, a devastating fungal pathogen of numerous economically important crops such as sugar beet, potato, and rice. Also in the study by Mendes et al. (2011), the Xanthomonadaceae family, to which Lysobacter belongs, was found more abundant in a soil suppressive against R. solani on sugar beet. Several studies have shown that application of Lysobacter spp. reduced diseases caused by different plant pathogens in several crops such as cucumber (Folman et al., 2004; Postma et al., 2009), bean (Yuen et al., 2001), rice (Ji et al., 2008), pepper (Ko et al., 2009), grapevine (Puopolo et al., 2014), sugar beet, spinach (Islam et al., 2005), and tomato (Puopolo et al., 2010). To date, however, few data are available on the frequency and diversity of Lysobacter species in natural habitats and little is known about the ecology and the determinative role of Lysobacter species in plant growth promotion and disease suppressive soils.
The work described here focused on elucidating the role of Lysobacter spp. in protecting plants against soil-borne diseases and in stimulating plant growth. To that end, we determined (i) the genetic and phenotypic diversity of 18 different Lysobacter strains obtained from soil and plant-associated environments, (ii) their activity against a range of pathogens, (iii) if these Lysobacter strains alone can suppress damping-off disease of sugar beet and cauliflower caused by R. solani, and (iv) if Lysobacter can promote plant growth via direct contact and/or via production of volatile compounds.
Materials and Methods
Strains, Culture, and Storage Conditions
The Lysobacter strains used in this study (Table 1) were isolated from different Dutch soils suppressive to R. solani. Reference strains (Table 1) were obtained from the DSM strain collection (Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures, Braunschweig, Germany). For the activity and plant growth promotion assays, Lysobacter strains were pre-cultured in tryptone soya broth (TSB, Oxoid) for 2–3 days at 25°C on a rotary shaker at 200 rpm and cells were washed 3 times with 0.9% NaCl unless mentioned otherwise. The fungal pathogens used in this study were mostly provided by the Institute of Sugar Beet Research (IRS). Fusarium oxysporum Forl1 was provided by the University of Turin, Italy (Clematis et al., 2009), Verticillium dahliae JR2 by B. Thomma [Wageningen University (WUR)], Phytophthora infestans by F. Govers (WUR), and Aspergillus niger was provided by L. de Graaf (WUR) (Table S1). The bacterial strains were kept in 40% (v/v) glycerol at −80°C; the fungi and oomycetes were kept in mineral oil at 10°C.
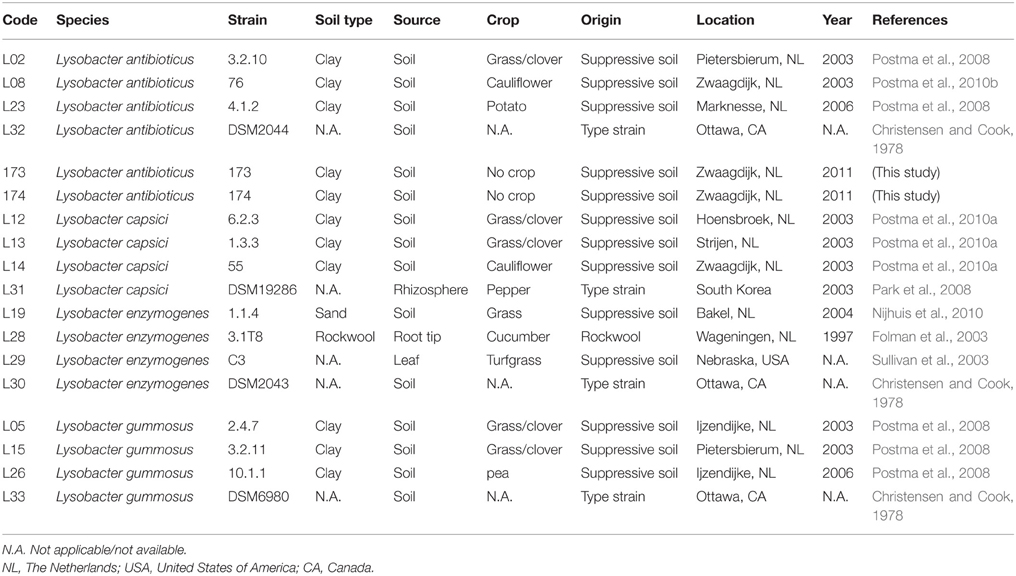
Table 1. Isolation details of the Lysobacter strains used in this study.
Soil Collection and Storage
The non-suppressive (conducive) soil to R. solani was collected from a pear orchard located in Zwaagdijk, The Netherlands (52°41′53.549″ N, 5°6′58.643″ E) in June 2012 at a depth of 10–40 cm. The soil, classified as clay soil with loam texture (29.9% of the particles are >50 μm, 26.4% of the particles are < 2 μm), was air-dried, sieved (0.5 cm mesh) to remove plant/root material and stored at 8°C until use for the in vivo activity test of Lysobacter spp. against R. solani on cauliflower.
Genetic and Phenotypic Characterization of the Lysobacter Strains
BOX-PCR
To determine the genetic variation among Lysobacter strains, the repetitive elements in their genome were analyzed by BOX-PCR according to Rademaker et al. (2004). Amplification reactions were conducted in 25 μl volume composed of 1 μl BOX-A1R primer (10 μM), 1.25 μl dNTPs (25 mM each), 0.4 μl BSA (10 mg/ml), 2.5 μl 100% DMSO, 5 μl 5x Gitschier buffer, 0.4 μl Taq polymerase (5U/μl SuperTaq), and 14.45 μl miliQ water. DNA was added by a toothpick inoculation of bacterial cells in the reaction mix. The reaction volume was heated to 95°C for 2 min, followed by 30 cycles of 3 s at 94°C, 92°C for 30 s, 50°C for 1 min, and 65°C for 8 min. The PCR reaction was finished with an 8 min incubation at 65°C for and then kept at 8°C. Five microliter of the PCR product was loaded on an 1.5% (w/v) agarose gel and ran overnight at 40V.
Phylogenetic Analyses
For each Lysobacter strain, the sequences of the 16S ribosomal RNA gene, the gene encoding a recombination/repair protein (recN) and the gene encoding the subunit C of the excinuclease ABC (uvrC) were amplified using primers described in Table 2. The markers recN and uvrC were chosen based on Zeigler (2003) who showed that these candidate genes will provide high fidelity for species prediction, and the 16S rRNA gene was included because of its broad use in taxonomic studies. Amplification reactions were conducted in 25 μl volume composed of 1 μl each of forward and reverse primer (10 μM), 1 μl dNTPs (5 mM each), 1.5 μl MgCl2 (25 mM), 5 μl 5x GoTaq Flexibuffer, 0.125 μl GoTaq polymerase (5 U/μl), and 15.375 μl miliQ water. DNA was added by a toothpick inoculation of bacterial cells in the reaction mix. The reaction volume was heated to 95°C for 3 min, followed by 35 cycles of: 1 min at 95°C, 58°C for 1 min, 72°C for 1.4 min (for 16S rRNA), 1 min at 95°C, 57.2°C for 1 min, 72°C for 1.2 min (for recN), and 1 min at 95°C, 58°C for 1 min, 72°C for 2 min (for uvrC). The PCR reaction were finished with an 5 min incubation at 72°C for and then kept at 12°C. Five microliter of the PCR product were visualized on an 1.5% (w/v) agarose and PCR products were sequenced by Macrogen Inc. (Amsterdam, The Netherlands). Phylogenetic trees were constructed with the three markers independently or concatenated using ClustalW alignments (Thompson et al., 1994) and neighbor joining tree constructions using the Tamura 3 parameter model and discrete Gamma distribution in MEGA6 (Tamura et al., 2013).
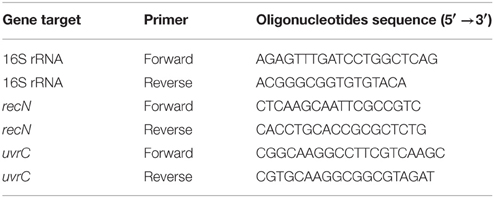
Table 2. Primer sets used for phylogenetic analysis.
The sequences obtained during this study are deposited in NCBI GenBank under accession numbers KT851449 to KT851466 for uvrC, KT851467 to KT851484 for 16S rRNA and KT851485 to KT851502 for recN.
Swarming Ability
Motility of the Lysobacter strains was assessed on soft standard succinate medium (SSM) as described in De Bruijn and Raaijmakers (2009). In brief, 5 μl of Lysobacter suspensions was spot-inoculated in the center of SSM agar Petri dishes [(32.8 mM K2HPO4, 22 mM KH2PO4, 7.6 mM (NH4)2SO4, 0.8 mM MgSO4, 34 mM succinic acid (w/v)), adjusted pH to 7 and 0.6% agar (w/v)]. Petri dishes were incubated for 2–12 days at 25°C.
Enzymatic Activity
Chitinase, glucanase, and protease activity of the Lysobacter strains were tested as described in De Bruijn et al. (in press). In brief, 2–5 μl of Lysobacter suspensions (of stationary phase of growth) was spot-inoculated in the center of different media containing 1.5–2% agar. For chitinase activity, R2A (Oxoid) and 1/10th strength TSB agar Petri dishes were used containing 0.2% colloidal chitin prepared from crab shell chitin (Sigma) and Petri dishes were incubated for 3–7 days at 25°C. For glucanase activity, R2A medium containing 0.5% laminarin was used and Petri dishes were incubated for 3 days at 25°C. The colonies were removed by washing with water and the medium was stained with 1% congo red. After destaining, coloration of the medium was determined. For protease activity, bacteria were inoculated on 15 g/l skimmed milk powder, 4 g/l blood agar base and 0.5 g/l yeast extract and Petri dishes were incubated for 3–7 days at 25°C.
In Vitro Antagonistic Activity
Lysobacter strains (Table 1) were grown in 5 ml TSB for 2 days at 25°C on a rotary shaker at 200 rpm. Suspensions were washed once by centrifugation at 3800 × g for 5 min and 10x concentrated in 0.9% NaCl.
To test activity against bacterial pathogens, R2A, 1/5th potato dextrose agar (PDA, Oxoid) and Luria-Bertani (LB, Difco) agar Petri dishes were prepared with an overlay of 1% water agar cooled down to 50°C to which washed cells of a culture of the bacterial pathogens (Table S1) were added. Subsequently, 2–5 μl of the Lysobacter cell suspensions (of stationary phase of growth) was spot-inoculated on the medium. Petri dishes were incubated for 3–7 days at 25°C and clearing zones surrounding the colonies were monitored.
To test inhibition of mycelial growth, oomycetes, and fungal strains (Table S1) were grown on PDA at 25°C. Four 5 μl of the Lysobacter suspensions were spot-inoculated at the edges of Petri dishes containing 20 ml of R2A, 1/5th PDA or PDA and a fresh 5 mm agar plug with actively grown mycelium was placed in the middle of the Petri dish.
To test antagonism against fungal spores, fungi (Table S1) were grown on PDA until sporulation. To enhance spore production, Cercospora and Stemphylium were grown on vegetable juice agar Petri dishes [(vegetable juice (V8) solified with 1.5% agar)] (Beckman and Payne, 1983; Rossi et al., 2005).
Under 16 h photoperiod, and to enhance spore collection from Verticillium and Aspergillus, the spores of those two fungi were scratched from the mycelium and streaked on fresh PDA Petri dishes. Fungal spores were collected as described in Trifonova et al. (2008) with slight modifications. In brief, spores were released from the mycelium by adding 10 ml of 0.9% NaCl and scratching the surface with a sterile spatula, collected, 10-fold diluted and added to the culture media (PDA, 1/5th PDA and R2A) of 48–55°C to a final concentration of 5% (v/v). Four 5 μl of the Lysobacter suspensions were spot-inoculated at the edges of Petri dishes containing 20 ml of medium with spores.
For each assay, three replicates per media were used. Petri dishes without Lysobacter were used as controls. All Petri dishes were incubated at 25°C for 1 week and subsequent inhibitory halo formation was monitored.
In Vivo Activity of Lysobacter spp. Against Rhizoctonia solani
Spontaneous rifampicin-resistant mutants of the Lysobacter strains were verified by BOX-PCR. These mutants exhibited chitinase activity to the same extent as their parental strains. The rifampicin-resistant mutants were grown in 10 ml of TSB supplemented with 50 μg/ml rifampicin for 2 days at 25°C on a rotary shaker at 200 rpm. Cultures were centrifuged, washed 3 times and resuspended in 0.9% NaCl. Cell suspensions were mixed in a potting soil:river sand (1:9, w/w) mixture at an initial density of 107 cells/g soil and approximately 20% hydration. Rectangle shape trays (19.5 × 6 × 3.5 cm) were filled with 250 g of the potting soil:sand mixture (eight replicates per treatment) and 16 sugar beet seeds coated with thiram, hymexazol, and poncho-beta were sown in a row, 1 cm apart. Non-inoculated soil was used as a control. Trays were placed in boxes with transparent lids in a growth chamber at 24°C with a 16 h photoperiod. After 5 days, seeds germinated and a single fresh 1/5th PDA agar plug (5 mm) grown with R. solani AG2-2 IIIB was placed touching the first seedling, with the mycelial side toward the plant. Spread of R. solani was scored at regular intervals during 2 weeks by scoring the number of diseased plants as well as the distance between the inoculum and the most distal plant suffering from damping-off. In addition, the area under the disease progress curve (AUDPC) was calculated to determine the disease dispersal over time as:
where ti are the time points in a sequence (days) and yi are measures of the disease dispersal (cm). Therefore, y(0) is defined as the initial infection at t = 0 and A(tx) is the AUDPC (total accumulated diseased dispersal until t = tx).
From each tray, the rhizospheres of two healthy sugar beet plants that were the closest to the last infected one were collected. Two replicates were pooled together in 4 ml 0.9% NaCl, vortexed for 1 min, sonicated for 1 min and vortexed for 15 s. Fifty microliter of a 10-, 100-, and 1000-fold dilution were plated on selective medium, R2A supplemented with 50 μg/ml rifampicin, 200 μg/ml ampicillin, 25 μg/ml kanamycin, and 100 μg/ml delvocid. Petri dishes were incubated at 25°C for 1 week. Colony forming units (CFU) were counted and CFU/g rhizosphere was calculated. The in vivo assay and the rhizosphere colonization test were done twice.
A similar experiment was performed in cauliflower using the same set up as described above with slight differences. Bacterial strains were grown in 10 ml of LB broth supplemented with 50 μg/ml rifampicin at 25°C for 3 days. The selected Lysobacter strains for this assay were L08, L14, L15, L19 and L29. Bacterial strains were inoculated in Zwaagdijk conducive soil at an initial density of 105 and 107 cells/g soil. Sowing, R. solani AG2-1/21 inoculation, growth of the plants, disease scoring, and AUDPC calculation was done as described above. The experiment was repeated twice, once with rifampicin resistant Lysobacter and once with non-rifampicin resistant Lysobacter. Statistically significant differences were determined by One-way ANOVA and post-hoc Dunnet's analysis (P < 0.05) performed in SPSS 22.0.
In Vitro Plant Growth Promotion Assay
Seed Preparation
Prior to surface sterilization, naked sugar beet (Beta vulgaris) seeds were soaked in 0.03 N HCl for 6 h under rotation, washed with sterile milliQ water and air-dried to enhance seed germination (Habib, 2010). Surface sterilization of sugar beet, cabbage (Brassica oleracea), and onion (Allium cepa) seeds was performed by washing the seeds in 2% sodium hypochlorite for 5 min and rinsing them with sterile milliQ water. Seeds were placed on Whatman filter paper moistened with 3 ml sterile milliQ water and pre-germinated at 25°C for 2–3 days. Arabidopsis thaliana (Columbia 0) seeds were sterilized in an exicator with 50 ml of commercial bleach (10% v/v) + 3% of concentrated HCl for 4 h, placed in wet Whatman filter paper and incubated at 4°C in darkness for 3 days.
Seed Inoculation
Two day-old pre-germinated sugar beet seeds were soaked in 3 ml of Lysobacter suspensions of 109 cells/ml for 30 min. Subsequently, sugar beet seeds (six seeds per container) were placed in cylinder shaped plastic containers (9 cm diameter, 8 cm height) with transparent lids containing 150 ml of 0.5 × Murashige and Skoog (MS) medium (including vitamins), and incubated in a growth chamber at 24°C with a 16 h photoperiod. Fresh and dry weight of shoots and roots were determined after 2 weeks. The experiment was done twice, with three replicates per treatment.
Root Tip Inoculation
Two days-old pre-germinated sugar beet seeds were placed in square Petri dishes (10 × 10 × 2 cm) containing 50 ml of 0.5 × MS medium (four seeds/Petri dish). Petri dishes were incubated in vertical position in a growth chamber at 24°C with a 16 h photoperiod until the roots were approximately 1 cm long and 2 μl of the Lysobacter suspensions of 109 cells/ml were, spotted onto each root tip and incubated for 1 week. Fresh and dry weight of shoots and roots was determined. The experiment was done once, with three replicates per treatment.
Volatile Assay
Two days-old pre-germinated seeds of sugar beet, cauliflower and onion were placed in containers as described above containing either 150 ml of 0.5 × MS medium or 150 g of a sterile mixture of potting soil:sand (1:9) with 20% humidity. A small Petri dish (35 mm diameter), containing 4 ml of R2A medium was placed in the middle of the container, and the Lysobacter strains were inoculated into the small Petri dishes at a density of 107 cells/Petri dish. Containers were incubated in a growth chamber at 24°C with a 16 h photoperiod for 2 weeks and fresh and dry weight of shoots and roots as well as leaf area were determined. The experiment was performed three times for sugar beet, once for cauliflower and once for onion, with five replicates per treatment. For the volatile assay in A. thaliana, L. antibioticus L08, L. capsici L14, L. gummosus L15, and Pseudomonas fluorescens SBW25 [known by its ability in promoting plant growth in A. thaliana when growing on King's B (KB) agar medium and used as a positive control (J. M. Raaijmakers, personal communication)] were used. Each bacterial strain was pre-cultured in LB broth for 2 days at 25°C, and then washed three times with 10 mM MgSO4. A 10 μl drop of a bacterial suspension of 109 cells/ml was spotted in the small Petri dish (35 mm diameter) containing 4 ml of R2A, LB or KB agar medium and Petri dishes were incubated for 1 day at 25°C. Small Petri dishes were placed into big Petri dishes (150 mm diameter) containing 50 ml of 0.5 × MS medium and five 3-days-old pre-germinated seeds were sown per Petri dish. Petri dishes with medium but without bacteria were included as controls. Petri dishes were incubated in vertical position in a growth chamber at 21°C with a 16 h photoperiod for 21 days. After that period, fresh and dry weight of shoots and roots were determined. The experiment was repeated once with five replicates/treatment.
Seed Colonization Ability
Naked sugar beet seeds were surface sterilized as described above and soaked in 3 ml of bacterial suspensions containing 109 cells/ml for 30 min as described above for the seed inoculation assay (22 seeds/bacterial treatment). Six seeds from each bacterial suspension were placed in 4 ml 0.9% NaCl, vortexed 1 min, sonicated 1 min, and vortexed 15 s. Fifty microliter of both undiluted suspensions and 10, 100, 1000, and 10000x time dilutions were plated on R2A agar dishes and incubated at 25°C for 1 week. The remaining seeds were sown in squared Petri dishes containing 50 ml of 0.5 MS (four seeds/Petri dish, four replicates per treatment) and incubated as described above for the root tip inoculation assay. After 1 week, the roots of the seedlings from each Petri dish were excised and placed in 4 ml of 0.9% NaCl, vortexed 1 min, sonicated 1 min, and vortexed 15 s. Fifty microliter of both undiluted suspensions and 10, 100, 1000, and 10000x fold dilution were plated on R2A agar dishes, incubated at 25°C for 1 week and the amount of colony forming units (CFU) per seed and per root were determined by colony counting.
Results
Genetic and Phenotypic Characterization of the Lysobacter Strains
BOX-PCR profiling of the 18 Lysobacter strains revealed a high genetic diversity among the different Lysobacter species and between strains of a given species (Figure 1A). L. gummosus strains showed the lowest intraspecific diversity whereas L. enzymogenes strains showed the highest diversity. Based on 16S rRNA sequences, the most phylogenetically distant species was L. enzymogenes (Figure S1A). When using either recN or uvrC or the three molecular markers together, however, L. antibioticus was the most distant of the four species (Figure 1B and Figures S1B,C).
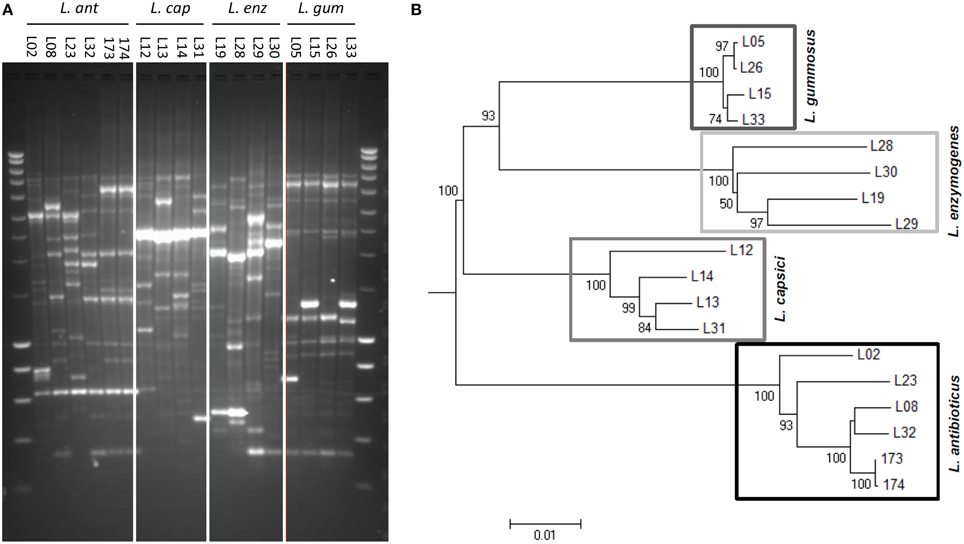
Figure 1. Genetic diversity of 18 selected Lysobacter strains belonging to four different species. (A) Genetic profiling by BOX-PCR. Lanes on complete left and right shows Smartladder (Eurogentec) marker. (B) Phylogenetic tree of the Lysobacter strains based on the concatenated sequences of the 16S ribosomal RNA gene (16S rRNA), a gene encoding a recombination/repair protein (recN) and a gene encoding the subunit C of the excinuclease ABC (uvrC). The evolutionary relationship of the Lysobacter strains was inferred by alignment with ClustalW and neighbor-joining tree construction. The numbers at the nodes indicate the level of bootstrap support of 50 or higher, based on neighbor-joining analysis of 1000 resampled data sets. The bar indicates the relative number of substitutions per site.
The Lysobacter strains did not show any motility after 4 days of incubation on soft SSM agar medium. After 12 days of incubation, however, L. capsici (L12, L13, L14, and L31) and L. enzymogenes (L19, L28, L29, L30) did spread from the point of inoculation, most likely due to gliding motility (Figure 2). All Lysobacter strains used in this study showed extracellular chitinase and glucanase activities (Figure 2). Most strains presented proteolytic activity except for two L. gummosus and four L. antibioticus strains (Figure 2). Variation in these three enzymatic activities among strains belonging to the same species was observed, especially for the L. antibioticus strains.
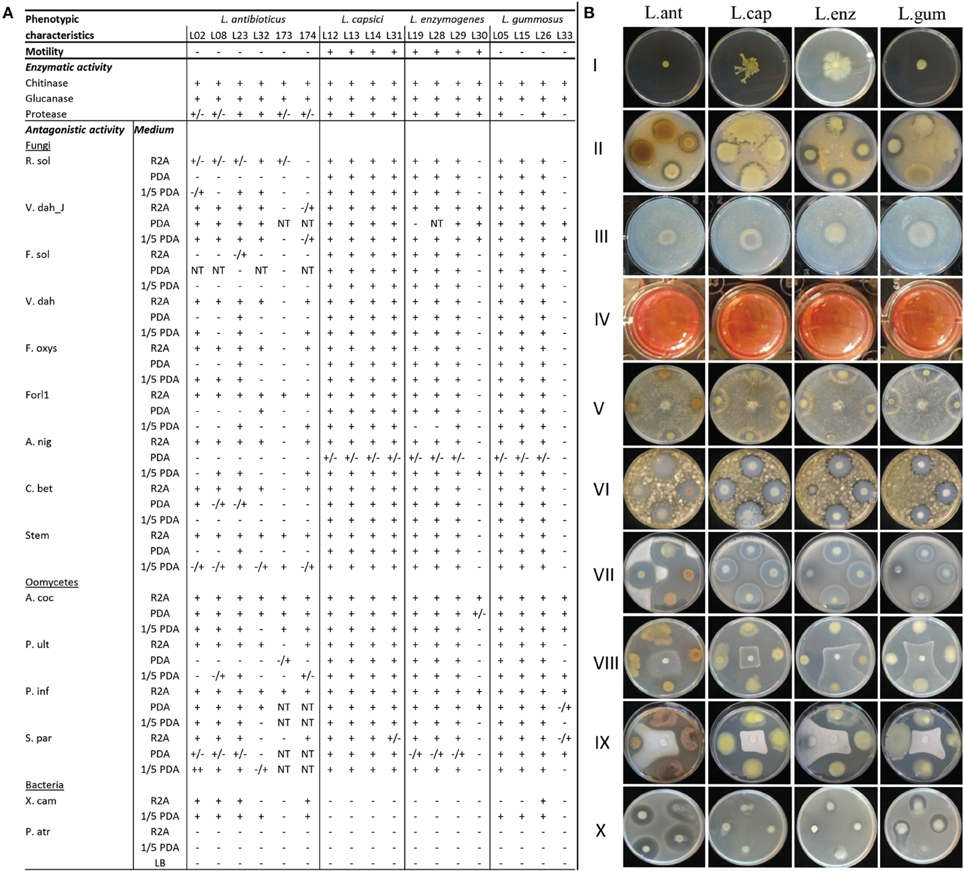
Figure 2. Phenotypic characterization of the Lysobacter strains, including (A) motility, protease, chitinase and glucanase activities, and antagonistic activity against pathogenic fungi, oomycetes and bacteria. + indicates activity; − indicates no activity; ± indicates antagonistic activity observed after 2–3 days of incubation, but the activity disappeared upon longer incubation. For the enzymatic activity, the ± indicates weak activity; NT indicates not tested. (B) Pictures of phenotypic characterization of L. antibioticus (L. ant), L. capsici (L. cap), L. enzymogenes (L. enz), and L. gummosus (L. gum) for I: motility on SSM medium; II: chitinase activity; III: glucanase activity, positive glucanase activity is given by the change from red to orange color (not shown); IV: protease activity; and in vitro antagonistic activity on R2A (except when otherwise indicated) against V: R. solani; VI: Cercospora beticola; VII: Verticillium dahliae; VIII: Pythium ultimum; IX: Aphanomyces cochlioides on PDA and X: Xanthomonas campestris pv. campestris on 1/5th PDA.
The antimicrobial activity of the Lysobacter strains (Table 1) was tested on different media. Almost all Lysobacter strains showed a strong antagonistic activity against all pathogens tested (Table S1), except against the plant pathogenic bacterium Pectobacterium atrosepticum. The magnitude of the antagonistic activity of Lysobacter was media-dependent, with the strongest activity on R2A medium and the weakest activity on PDA medium (Figure 2). L. capsici was the most consistent species in terms of antagonistic activity, with all L. capsici strains showing activity on R2A against all pathogens tested except for X. campestris and L. capsici strain L31 against S. parasitica (Figure 2). On R2A, all L. enzymogenes and L. gummosus strains, with the exception of the type strains, showed activity against all pathogens tested. The type strain of L. enzymogenes did show activity against V. dahliae JR2, A. cochlioides and P. infestans, whereas the L. gummosus type strain had activity against all oomycetes tested except P. ultimum (Figure 2). L. antibioticus strains showed the highest variation in activity, with strain L23 having the broadest antimicrobial activity (Figure 2).
In Vivo Activity of Lysobacter spp. against Rhizoctonia solani
The efficacy of the Lysobacter strains, several of which originate from Rhizoctonia suppressive soil, to control Rhizoctonia damping-off disease of sugar beet seedlings was tested in a sterilized (by autoclaving twice) sand-potting soil mixture and in a non-sterilized agricultural soil. Seed germination was not affected by the Lysobacter strains. In two bioassays, none of the strains was able to consistently suppress damping-off disease caused by R. solani after 2 weeks of plant growth (Figure 3A). For example, strains L19 and L05 significantly reduced damping-off disease of sugar beet in bioassay 2 but not in bioassay 1 (Figure 3A).
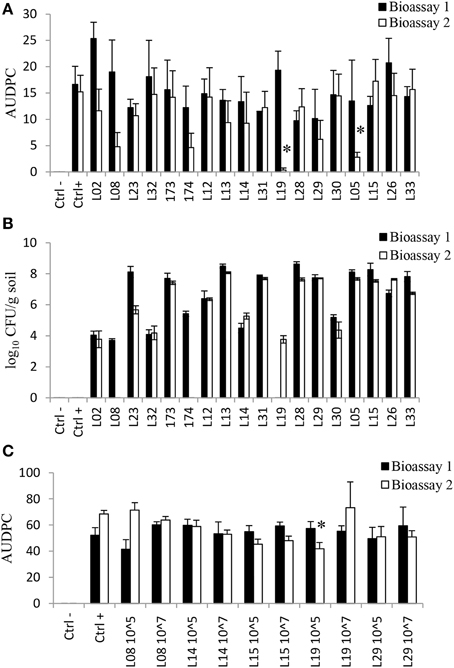
Figure 3. In vivo Rhizoctonia disease suppression and rhizosphere colonization ability by Lysobacter strains. (A) Area under disease progress curve (AUDPC) of disease spread for sugar beet when Lysobacter strains were applied at an initial density of 107 CFU/g into a mixture potting soil:sand (1:9); (B) Colonization of the rhizosphere of sugar beet by the Lysobacter strains when applied at an initial density of 107 CFU/g into a mixture potting soil:sand (1:9). (C) AUDPC of disease spread for cauliflower when Lysobacter strains were applied into a conducive soil. 10^7 and 10^5 means an initial density of the inoculum at 107 and 105 cells/g soil, respectively; L. antibioticus: L02, L08, L23, L32, 173, 174; L. capsici: L12, L13, L14, L31; L. enzymogenes: L19, L28, L29, L30, and L. gummosus: L05, L15, L26, L33. For each of the two bioassays, an asterisk indicates a significant difference (p < 0.05) with the control treatment calculated by analysis of variance and Dunnet's post-hoc analysis.
The results further showed that after an initial application of 107 CFU/g soil, Lysobacter strains established densities in the rhizosphere of sugar beet ranging from 103 to 108 CFU/g (Figure 3B), with substantial variation between strains and between the two bioassays. In general, L. gummosus strains were better rhizosphere colonizers whereas L. antibioticus showed the highest variation among strains. L. antibioticus strains L08 and 174 were only detected in the sugar beet rhizosphere in bioassay 1. L. antibioticus L23 was detected at high densities (108 CFU/g) in bioassay 1, but at 1000-fold lower densities in bioassay 2. L. enzymogenes L19 was only detected in bioassay 2 (Figure 3B).
The ability of Lysobacter to suppress Rhizoctonia damping-off disease of another host plant (cauliflower) was assessed for Lysobacter strains L08, L14, L15, L19, and L29 at two initial densities of 105 and 107 CFU/g of soil. Also for this crop, germination was not affected by the introduced bacterial strains and again no significant and consistent reduction in disease incidence was observed. When applied at 105 CFU/g of soil, strain L19 significantly reduced disease incidence but only in bioassay 2 (Figure 3C). For bioassay 2, colonization of cauliflower rhizosphere by the Lysobacter strains was determined. The results showed that the densities recovered were lower (101 to 103) than initially applied except for L. enzymogenes L29 and L. gummosus L15 when applied at 107 CFU/g soil (Figure S2). After an initial application of 105 cells/g soil, only L. gummosus L15 and L enzymogenes L19 and L29 were detected in the rhizosphere of cauliflower.
Plant Growth Promotion
The ability of the Lysobacter strains to promote plant growth in vitro was tested for sugar beet, cauliflower, onion, and A. thaliana. For sugar beet, the 18 Lysobacter strains were applied to the seeds as well as to the root tips. For the first seed inoculation assay, almost all L. antibioticus strains negatively affected plant growth, decreasing plant biomass with 15–38% compared to the untreated control (Figure 4). One L. capsici and two L. enzymogenes strains negatively affected shoot biomass. In the second bioassay, no negative or positive effects on plant growth were observed for any of the strains (Figure 4), except for L. gummosus L26 which promoted root growth.
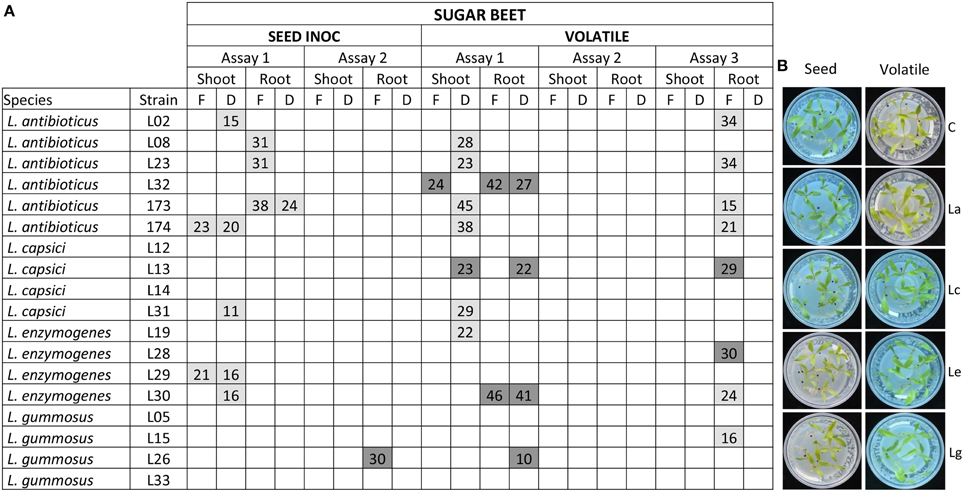
Figure 4. Sugar beet plant growth promotion by Lysobacter strains. (A) Sugar beet seeds were grown on 0.5 MS medium and plant growth promotion was determined when Lysobacter strains were inoculated on seeds or by volatiles. Each assay was performed with three to five replicates. F indicates fresh weight; D indicates dry weight. Light gray boxes indicate a statistical significant negative effect in plant growth when compared to the control and dark gray boxes indicate a statistical significant positive effect. Values within the boxes, indicates the % of increase/decrease of plant weight compared to the control. (B) Pictures of the plant growth promotion assays. C, control; La: L. antibioticus; Lc: L. capsici; Le: L. enzymogenes; Lg: L. gummosus. Significant differences (p < 0.05) with the uninoculated control were calculated using analysis of variance and Dunnet's post-hoc analysis.
The ability of Lysobacter to colonize the surface of the seeds and the roots was determined for bioassay 2. Whilst bacteria were applied at an initial density of 108 cells/seed, bacterial recovery from the seed after 30 min of incubation ranged from approximately 103–104 cells/seed, with even lower numbers for L. antibioticus L32 (102 cells/seed; Table S2). After 1 week of plant growth, bacteria could not be detected on sugar beet roots (Table S2). Hence, Lysobacter appears to be a poor root colonizer under these experimental conditions.
In the root tip inoculation assay, positive effects (ranging from 17 to 28% biomass increase) were observed for dry weight of shoots by two L. antibioticus, two L. capsici, and one L. enzymogenes strains (Figure S3). One L. antibioticus and one L. gummosus strain increased fresh (33%) and dry (38%) root biomass respectively (Figure S3).
To determine if Lysobacter emits volatile compounds that promote plant growth, assays were conducted in a split Petri dish where Lysobacter was physically separated from sugar beet seedlings. A high variation in plant phenotypes was observed between assays. For example, L. antibioticus L32 increased shoot biomass with 24% and root biomass with 42% only in the first assay. L. enzymogenes L30 increased root biomass in the first assay whereas in the third assay it showed a negative effect on plant growth (Figure 4). The volatile assays were repeated in sterile potting soil:sand mixture with sugar beet, cauliflower, and onion. Also in these assays, no significant and consistent results were obtained for the Lysobacter strains tested (data not shown). In addition, plant growth promotion was also determined by measurement of the leaf surface and no positive or negative effects of the Lysobacter strains were observed (data not shown).
L. antibioticus strain L08, L. capsici L14, L. gummosus L15 were also tested for volatile-mediated growth promotion of A. thaliana on different media. The positive control P. fluorescens SBW25 significantly increased shoot and root biomass (Figure S4). However, none of the Lysobacter strains tested showed a plant growth promoting effect on A. thaliana. Furthermore, when growing on LB medium, all the three Lysobacter as well as P. fluorescens SBW25 showed a notable adverse effect on plant growth (Figure S4).
Discussion
The genus Lysobacter is receiving substantial ecological and biotechnological interest as producers of different exoenzymes and antibiotics (Pidot et al., 2014). During the last years, several Lysobacter species have been isolated from Dutch soils suppressive to the fungal root pathogen R. solani (Postma et al., 2008, 2010b). Here, we showed that 18 Lysobacter strains from Rhizoctonia suppressive soils showed a high genetic diversity. In a recent study, comparative genomics of seven Lysobacter strains (five of which are included in this study) belonging to four Lysobacter species showed only 55% overlap in genome content (De Bruijn et al., in press). A high genetic diversity can confer an advantage under adverse environmental conditions as some members may exhibit phenotypes that allow them to survive and proliferate (Foster, 2005). Genome analysis also revealed the lack of flagellar genes (De Bruijn et al., in press), which supports our findings that none of the Lysobacter strains tested were motile on soft agar. Nonetheless, some dispersal was observed for L. capsici and L. enzymogenes after 12 days of incubation, most likely due to gliding motility as described previously for other Lysobacter species (Sullivan et al., 2003; Hayward et al., 2010).
Lysobacter is known to produce a variety of bioactive compounds, including enzymes and antimicrobial compounds. Hence, they were pointed out as an untapped source of new bioactive products (Xie et al., 2012; Pidot et al., 2014). Our results showed that the Lysobacter strains possess chitinase and glucanase activity, confirming and extending previous research (Zhang and Yuen, 2000; Zhang et al., 2001; Palumbo et al., 2005; De Bruijn et al., in press). Protease activity was observed for all strains belonging to L. capsici and L. enzymogenes, whereas only two out of four strains from L. gummosus and two out of six from L. antibioticus showed this activity. Chitinase, glucanase and protease activities may contribute to antimicrobial activity, since chitin, α- and β-glucans and glycoproteins are the major components of the cell walls of fungi (Barreto-Bergter and Figueiredo, 2014).
Most of the Lysobacter strains effectively inhibited the growth of oomycetes and fungi; only L. antibioticus and L. gummosus strains showed antibacterial activity. Differences in activity were observed between Lysobacter species and between strains of a given species, suggesting that the genus Lysobacter indeed may have a large reservoir of putative novel bioactive compounds. The in vitro antagonistic activity was media-dependent, showing stronger activity on poor medium, confirming and extending results obtained previously for the activity of L. enzymogenes 3.1T8 against Pythium aphanidermatum (Folman et al., 2004).
Due to their broad spectrum activity, Lysobacter members have been proposed as promising candidates for biological control of plant diseases (Hayward et al., 2010). However, none of the Lysobacter strains used in this study were able to consistently reduce R. solani infection on sugar beet and cauliflower. These results differ from those in previous studies where several Lysobacter strains significantly controlled plant pathogens, including P. aphanidermatum on cucumber (Folman et al., 2004; Postma et al., 2009), Bipolaris sorokiniana on tall fescue (Kilic-Ekici and Yuen, 2003), Uromyces appendiculatus on bean (Yuen et al., 2001), Xanthomonas oryzae pv. oryzae on rice (Ji et al., 2008), Phytophthora capsici on pepper (Ko et al., 2009), Plasmopara viticola on grapevine (Puopolo et al., 2014), Aphanomyces cochlioides in sugar beet and spinach (Islam et al., 2005) and F. oxysporum f. sp. radicis-lycopersici on tomato (Puopolo et al., 2010). Furthermore, L. capsici YS1215 was reported to have nematicidal activity, reducing root-knot caused by Meloidogyne incognita by inhibiting egg hatching (Lee et al., 2014).
Most of the Lysobacter strains tested here poorly colonized the rhizosphere of sugar beet and cauliflower. Given the importance of root colonization for biocontrol (Bull et al., 1991; Johnson, 1994; Raaijmakers et al., 1995), this suggests that the inconsistency in disease control by the Lysobacter strains may be due to their lack of competitiveness in the rhizosphere of sugar beet and cauliflower. The rhizosphere differs from the bulk soil by the presence of plant root exudates that create an environment rich in nutrients. Chemotaxis and active motility toward root exudates represent the first steps in rhizosphere colonization (Benizri et al., 2001; De Weert and Bloemberg, 2006). This motility may be active, through flagellar movements, or passive, through percolating water or vectors. None of the 18 Lysobacter strains possess flagella, what limits the capacity of the strains to effectively compete against flagellated soil bacteria for a niche in the rhizosphere. The adherence to root tissues through biofilm formation is the next step in rhizosphere colonization (Benizri et al., 2001; Ramey et al., 2004; Danhorn and Fuqua, 2007). Several traits are involved in biofilm formation including cell wall structures and extracellular polysaccharide production (Lugtenberg et al., 2001). Biofilm production in vitro has been described for L. capsici AZ78 and appeared medium specific, (Puopolo et al., 2014). Biofilm formation was observed for Lysobacter sp. strain SB-K88 on roots of sugar beet (Islam et al., 2005). Biofilm formation in situ was not tested for our 18 Lysobacter strains and will be subject of future studies. The root exudate composition is plant specific (Mandimba et al., 1986) and the ability to assimilate specific amino acids, vitamin B1, carbohydrates, organic acids as well as pH tolerance and competition for limiting resources also determine the rhizosphere competence (Dekkers et al., 1999; Benizri et al., 2001; Lugtenberg and Kamilova, 2009; Ghirardi et al., 2012). In the rhizosphere there is often a limitation for soluble iron, commonly used as a cofactor in enzymes that are involved in pathways that are essential for microbial growth. Therefore, the ability to produce siderophores (small high-affinity iron chelating compounds) confers a competitive advantage. The role of competition for iron by siderophore production of Lysobacter sp. seems species or strain specific and not all strains, including several strains used in this study, possess iron-chelating capacity (Puopolo et al., 2010; Ko et al., 2011; De Bruijn et al., in press).
The soil type may also influence rhizosphere colonization and biocontrol activity. For example, the colonization of Pseudomonas sp. strain ITRI53 and Pantoea sp. strain BTRH79 of Italian ryegrass was higher in loamy soils compared with sandy soils (Afzal et al., 2011). The agricultural soil used in this study is a clay soil with loam texture. Several of our Lysobacter strains were isolated from this agricultural soil and we expected that those conditions would provide a “home-field advantage” for rhizosphere colonization of sugar beet and cauliflower. In a potting soil:sand mixture, we observed higher rhizosphere population densities on sugar beet seedlings as compared to the agricultural soil, with densities higher than the minimal dose of 105 CFU/g soil reported for other biocontrol strains (Xu and Gross, 1986; Leeman et al., 1995; Raaijmakers et al., 1995). Despite these densities, no significant and/or consistent biocontrol activity was observed for any of the Lysobacter strains tested.
Several biocontrol agents not only suppress disease but also promote plant growth (Johansson et al., 2003). None of the Lysobacter strains tested in this study, however, were able to significantly and consistently promote growth of 4 different crops when applied to seeds or root tips or when applied physically separated from the crop. Furthermore, volatiles produced by the Lysobacter strains when grown on LB medium even showed a negative effect on growth of A. thaliana. This may be due to the accumulation of toxic volatiles that are produced by Lysobacter spp. when growing in rich media. Weise et al. (2013) showed that Serratia odorifera inhibited the growth of A. thaliana plants due to the production of ammonia when grown on peptone-rich nutrient media. Iwata et al. (2010) reported that Lysobacter sp. E4 was able to fix nitrogen under free-living conditions and accumulated ammonia in the culture broth. Also hydrogen cyanide (HCN) produced by Chromobacterium, Pseudomonas, and Serratia have been shown to inhibit the growth of A. thaliana (Blom et al., 2011). More research needs to be conducted to determine if HCN or other toxic volatiles are produced by Lysobacter.
Overall, our results indicate that none of the 18 Lysobacter strains have the potential to control Rhizoctonia or promote plant growth of sugar beet and cauliflower, probably due to insufficient rhizosphere competence. However, the Lysobacter strains showed a high diversity in in vitro activity against 14 different pathogenic fungi, oomycetes and bacteria, suggesting that the genus Lysobacter constitutes an extensive source of (new) enzymes and antimicrobial compounds. Possibly Lysobacter needs to interact with a specific microbial community to become antagonistic to Rhizoctonia or to promote plant growth in natural environments. To better understand the potential contribution of Lysobacter species to the overall activities of the microbial communities responsible for soil suppressiveness against R. solani, in-depth metagenomic and metatranscriptomic analyses of the bacterial community compositions and functions will be needed to unravel the role of this genus in disease suppressiveness. Future work will include testing Lysobacter mixtures or mixtures with other bacterial genera abundant in soils suppressive to R. solani. Interactions of Lysobacter with other bacteria may stimulate the production of antimicrobial compounds as was shown recently for other bacterial genera (Tyc et al., 2014).
Author Contributions
All authors were involved in the design of the experiments. RG and IB performed in vitro and in vivo activity bioassays, BOX-PCR and phylogenetic analyses. RG performed plant growth promotion assays. All authors contributed to the writing of the manuscript and approved submission.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Bram Hanse (IRS) for providing sugar beet seeds and most of the pathogenic strains, Liesbeth van der Heijden (Bejo Zaden B.V.) for providing cauliflower and onion seeds and Bart Thomma, Francine Govers, and Leo de Graaf (WUR) for providing the pathogenic strains. We also thank Reinier van Velzen for his help in part of the in vivo bioassays. This manuscript is publication number 5947 of Netherlands Institute of Ecology (NIOO-KNAW). This research was funded by the Dutch Technology Foundation (STW), project number 11755.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2015.01243
References
Afzal, M., Yousaf, S., Reichenauer, T. G., Kuffner, M., and Sessitsch, A. (2011). Soil type affects plant colonization, activity and catabolic gene expression of inoculated bacterial strains during phytoremediation of diesel. J. Hazard. Mater. 186, 1568–1575. doi: 10.1016/j.jhazmat.2010.12.040
Barreto-Bergter, E., and Figueiredo, R. T. (2014). Fungal glycans and the innate immune recognition. Front. Cell. Infect. Microbiol. 4:145. doi: 10.3389/fcimb.2014.00145
Beckman, P. M., and Payne, G. A. (1983). Cultural techniques and conditions influencing growth and sporulation of Cercospora zeae-maydis and lesion development in corn. Phytopathology 73, 286–289. doi: 10.1094/Phyto-73-286
Benizri, E., Baudoin, E., and Guckert, A. (2001). Root colonization by inoculated plant growth-promoting rhizobacteria. Biocontrol Sci. Technol. 11, 557–574. doi: 10.1080/09583150120076120
Blom, D., Fabbri, C., Eberl, L., and Weisskopf, L. (2011). Volatile-mediated killing of Arabidopsis thaliana by bacteria is mainly due to hydrogen cyanide. Appl. Environ. Microbiol. 77, 1000–1008. doi: 10.1128/AEM.01968-10
Bull, C. T., Weller, D., and Thomashow, L. S. (1991). Relationship between root colonization and suppression of Gaeumannomyces graminis var. tritici by Pseudomonas fluorescens strain 2-79. Phytopathology 81, 954–959. doi: 10.1094/Phyto-81-954
Christensen, P., and Cook, F. D. (1978). Lysobacter, a new genus of nonfruiting, gliding bacteria with a high base ratio. Int. J. Syst. Evol. Micr. 28, 367–393. doi: 10.1099/00207713-28-3-367
Clematis, F., Minuto, A., Gullino, M. L., and Garibaldi, A. (2009). Suppressiveness to Fusarium oxysporum f. sp. radicis lycopersici in re-used perlite and perlite–peat substrates in soilless tomatoes. Biol. Control 48, 108–114. doi: 10.1016/j.biocontrol.2008.10.001
Danhorn, T., and Fuqua, C. (2007). Biofilm formation by plant-associated bacteria. Annu. Rev. Microbiol. 61, 401–422. doi: 10.1146/annurev.micro.61.080706.093316
De Bruijn, I., Cheng, X., de Jager, V., Gómez Expósito, R., Watrous, J., Patel, N., et al. (in press). Comparative genomics and metabolic profiling of the genus Lysobacter. BMC Genomics.
De Bruijn, I., and Raaijmakers, J. M. (2009). Regulation of Cyclic Lipopeptide Biosynthesis in Pseudomonas fluorescens by the ClpP Protease. J. Bacteriol. 191, 1910–1923. doi: 10.1128/JB.01558-08
Dekkers, L., Phoelich, C., and Lugtenberg, B. (1999). Bacterial Traits and Genes Involved in Rhizosphere Colonization. Halifax, NS: Atlanta Canada Society for Microbial Ecology.
De Weert, S., and Bloemberg, G. (2006). “Rhizosphere competence and the role of root colonization in biocontrol,” in Plant-Associated Bacteria, ed S. Gnanamanickam (Netherlands: Springer), 317–333.
Du, J., Singh, H., Ngo, H. T., Won, K., Kim, K. Y., and Yi, T. H. (2015). Lysobacter tyrosinelyticus sp. nov. isolated from gyeryongsan national park soil. J. Microbiol. 53, 365–370. doi: 10.1007/s12275-015-4729-9
Folman, L. B., De Klein, M. J. E. M., Postma, J., and Van Veen, J. A. (2004). Production of antifungal compounds by Lysobacter enzymogenes isolate 3.1T8 under different conditions in relation to its efficacy as a biocontrol agent of Pythium aphanidermatum in cucumber. Biol. Control 31, 145–154. doi: 10.1016/j.biocontrol.2004.03.008
Folman, L. B., Postma, J., and Van Veen, J. A. (2003). Characterisation of Lysobacter enzymogenes (christensen and cook 1978) strain 3.1T8, a powerful antagonist of fungal diseases of cucumber. Microbiol. Res. 158, 107–115. doi: 10.1078/0944-5013-00185
Foster, P. L. (2005). Stress responses and genetic variation in bacteria. Mutat. Res. 569, 3–11. doi: 10.1016/j.mrfmmm.2004.07.017
Ghirardi, S., Dessaint, F., Mazurier, S., Corberand, T., Raaijmakers, J. M., Meyer, J.-M., et al. (2012). Identification of traits shared by rhizosphere-competent strains of fluorescent pseudomonads. Microb. Ecol. 64, 725–737. doi: 10.1007/s00248-012-0065-3
Gökçen, A., Vilcinskas, A., and Wiesner, J. (2014). Biofilm-degrading enzymes from Lysobacter gummosus. Virulence 5, 378–387. doi: 10.4161/viru.27919
Habib, M. (2010). Sugarbeet (Beta vulgaris L.) seed pre-treatment with water and HCl to improve germination. Afr. J. Biotechnol. 9, 1338–1342. doi: 10.5897/AJB10.1460
Hayward, A. C., Fegan, N., Fegan, M., and Stirling, G. R. (2010). Stenotrophomonas and Lysobacter: ubiquitous plant-associated gamma-proteobacteria of developing significance in applied microbiology. J. Appl. Microbiol. 108, 756–770. doi: 10.1111/j.1365-2672.2009.04471.x
Islam, M. T., Hashidoko, Y., Deora, A., Ito, T., and Tahara, S. (2005). Suppression of damping-off disease in host plants by the rhizoplane bacterium Lysobacter sp. strain SB-K88 is linked to plant colonization and antibiosis against soilborne Peronosporomycetes. Appl. Environ. Microbiol. 71, 3786–3796. doi: 10.1128/AEM.71.7.3786-3796.2005
Iwata, K., Azlan, A., Yamakawa, H., and Omori, T. (2010). Ammonia accumulation in culture broth by the novel nitrogen-fixing bacterium, Lysobacter sp. E4. J. Biosci. Bioeng. 110, 415–418. doi: 10.1016/j.jbiosc.2010.05.006
Ji, G.-H., Wei, L.-F., He, Y.-Q., Wu, Y.-P., and Bai, X.-H. (2008). Biological control of rice bacterial blight by Lysobacter antibioticus strain 13-1. Biol. Control 45, 288–296. doi: 10.1016/j.biocontrol.2008.01.004
Johansson, P. M., Johnsson, L., and Gerhardson, B. (2003). Suppression of wheat-seedling diseases caused by Fusarium culmorum and Microdochium nivale using bacterial seed treatment. Plant Pathol. 52, 219–227. doi: 10.1046/j.1365-3059.2003.00815.x
Johnson, K. B. (1994). Dose-response relationships and inundative biological control. Phytopathology 84, 780–784. doi: 10.1094/phyto-84-780
Kilic-Ekici, O., and Yuen, G. Y. (2003). Induced Resistance as a mechanism of biological control by Lysobacter enzymogenes Strain C3. Phytopathology 93, 1103–1110. doi: 10.1094/PHYTO.2003.93.9.1103
Ko, H. S., Jin, R. D., Krishnan, H. B., Lee, S. B., and Kim, K. Y. (2009). Biocontrol ability of Lysobacter antibioticus hs124 against phytophthora blight is mediated by the production of 4-hydroxyphenylacetic acid and several lytic enzymes. Curr. Microbiol. 59, 608–615. doi: 10.1007/s00284-009-9481-0
Ko, H. S., Tindwa, H., De Jin, R., Lee, Y. S., Hong, S. H., Hyun, H. N., et al. (2011). Investigation of siderophore production and antifungal activity against Phytophthora capsici as related to iron (iii) nutrition by Lysobacter antibioticus HS124. Korean J. Soil. Sci. Fert. 44, 650–656. doi: 10.7745/KJSSF.2011.44.4.650
Lee, Y. S., Naning, K. W., Nguyen, X. H., Kim, S. B., Moon, J. H., and Kim, K. Y. (2014). Ovicidal activity of lactic acid produced by Lysobacter capsici ys1215 on eggs of root-knot nematode, Meloidogyne incognita. J. Microbiol. Biotechnol. 24, 1510–1515. doi: 10.4014/jmb.1405.05014
Leeman, M., Van Pelt, J., Den Ouden, F., Heinsbroek, M., Bakker, P., and Schippers, B. (1995). Induction of systemic resistance against Fusarium wilt of radish by lipopolysaccharides of Pseudomonas fluorescens. Phytopathology 85, 1021–1027. doi: 10.1094/Phyto-85-1021
Li, S., Jochum, C. C., Yu, F., Zaleta-Rivera, K., Du, L., Harris, S. D., et al. (2008). An antibiotic complex from Lysobacter enzymogenes strain c3: antimicrobial activity and role in plant disease control. Phytopathology 98, 695–701. doi: 10.1094/PHYTO-98-6-0695
Lin, S. Y., Hameed, A., Wen, C. Z., Liu, Y. C., Hsu, Y. H., Lai, W. A., et al. (2015). Lysobacter lycopersici sp. nov., isolated from tomato plant Solanum lycopersicum. Antonie Van Leeuwenhoek 107, 1261–1270. doi: 10.1007/s10482-015-0419-1
Lugtenberg, B. J., Dekkers, L., and Bloemberg, G. V. (2001). Molecular determinants of rhizosphere colonization by Pseudomonas. Annu. Rev. Phytopathol. 39, 461–490. doi: 10.1146/annurev.phyto.39.1.461
Lugtenberg, B., and Kamilova, F. (2009). Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 63, 541–556. doi: 10.1146/annurev.micro.62.081307.162918
Mandimba, G., Heulin, T., Bally, R., Guckert, A., and Balandreau, J. (1986). Chemotaxis of free-living nitrogen-fixing bacteria towards maize mucilage. Plant Soil 90, 129–139. doi: 10.1007/BF02277392
Mendes, R., Kruijt, M., de Bruijn, I., Dekkers, E., Van Der Voort, M., Schneider, J. H., et al. (2011). Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 332, 1097–1100. doi: 10.1126/science.1203980
Nijhuis, E. H., Pastoor, R., and Postma, J. (2010). Specific detection of Lysobacter enzymogenes (christensen and cook 1978) strain 3.1T8 with taqman PCR. J. Appl. Microbiol. 108, 1155–1166. doi: 10.1111/j.1365-2672.2009.04519.x
Palumbo, J. D., Yuen, G. Y., Jochum, C. C., Tatum, K., and Kobayashi, D. Y. (2005). Mutagenesis of beta-1,3-glucanase genes in Lysobacter enzymogenes strain c3 results in reduced biological control activity toward bipolaris leaf spot of tall fescue and pythium damping-off of sugar beet. Phytopathology 95, 701–707. doi: 10.1094/PHYTO-95-0701
Park, J. H., Kim, R., Aslam, Z., Jeon, C. O., and Chung, Y. R. (2008). Lysobacter capsici sp. nov., with antimicrobial activity, isolated from the rhizosphere of pepper, and emended description of the genus Lysobacter. Int. J. Syst. Evol. Microbiol. 58, 387–392. doi: 10.1099/ijs.0.65290-0
Pidot, S. J., Coyne, S., Kloss, F., and Hertweck, C. (2014). Antibiotics from neglected bacterial sources. Int. J. Med. Microbiol. 304, 14–22. doi: 10.1016/j.ijmm.2013.08.011
Postma, J., Nijhuis, E. H., and Yassin, A. F. (2010a). Genotypic and phenotypic variation among Lysobacter capsici strains isolated from rhizoctonia suppressive soils. Syst. Appl. Microbiol. 33, 232–235. doi: 10.1016/j.syapm.2010.03.002
Postma, J., Scheper, R. W. A., and Schilder, M. T. (2010b). Effect of successive cauliflower plantings and Rhizoctonia solani AG 2-1 inoculations on disease suppressiveness of a suppressive and a conducive soil. Soil Biol. Biochem. 42, 804–812. doi: 10.1016/j.soilbio.2010.01.017
Postma, J., Schilder, M. T., Bloem, J., and Van Leeuwen-Haagsma, W. K. (2008). Soil suppressiveness and functional diversity of the soil microflora in organic farming systems. Soil Biol. Biochem. 40, 2394–2406. doi: 10.1016/j.soilbio.2008.05.023
Postma, J., Stevens, L. H., Wiegers, G. L., Davelaar, E., and Nijhuis, E. H. (2009). Biological control of Pythium aphanidermatum in cucumber with a combined application of Lysobacter enzymogenes strain 3.1T8 and chitosan. Biol. Control 48, 301–309. doi: 10.1016/j.biocontrol.2008.11.006
Puopolo, G., Giovannini, O., and Pertot, I. (2014). Lysobacter capsici AZ78 can be combined with copper to effectively control Plasmopara viticola on grapevine. Microbiol. Res. 169, 633–642. doi: 10.1016/j.micres.2013.09.013
Puopolo, G., Raio, A., and Zoina, A. (2010). Identification and characterization of Lysobacter capsici strain PG4: a new plant health-promoting rhizobacterium. J. Plant Pathol. 92, 157–164. doi: 10.4454/jpp.v92i1.25
Raaijmakers, J. M., Leeman, M., Van Oorschot, M. M., Van Der Sluis, I., Schippers, B., and Bakker, P. (1995). Dose-response relationships in biological control of fusarium wilt of radish by Pseudomonas spp. Phytopathology 85, 1075–1080. doi: 10.1094/Phyto-85-1075
Rademaker, J. L. W., Louws, F. J., Versalovic, J., and Bruijn, F. J. (2004). “Characterization of the diversity of ecological important microbes by rep-PCR genomic fingerprinting,” in Molecular Microbial Ecology Manual II (Dordrecht: Kluwer), 611–643.
Ramey, B. E., Koutsoudis, M., Von Bodman, S. B., and Fuqua, C. (2004). Biofilm formation in plant-microbe associations. Curr. Opin. Microbiol. 7, 602–609. doi: 10.1016/j.mib.2004.10.014
Reichenbach, H. (2006). “The Genus Lysobacter,” in The Prokaryotes, eds M. Dworkin, S. Falkow, E. Rosenberg, K.-H. Schleifer and E. Stackebrandt (New York, NY: Springer), 939–957.
Rossi, V., Pattori, E., Giosué, S., and Bugiani, R. (2005). Growth and sporulation of Stemphylium vesicarium, the causal agent of brown spot of pear, on herb plants of orchard lawns. Eur. J. Plant Pathol. 111, 361–370. doi: 10.1007/s10658-004-5273-3
Singh, H., Du, J., Ngo, H. T., Won, K., Yang, J. E., Kim, K. Y., et al. (2015). Lysobacter fragariae sp. nov. and Lysobacter rhizosphaerae sp. nov. isolated from rhizosphere of strawberry plant. Antonie Van Leeuwenhoek 107, 1437–1444. doi: 10.1007/s10482-015-0439-x
Stepnaya, O. A., Tsfasman, I. M., Chaika, I. A., Muranova, T. A., and Kulaev, I. S. (2008). Extracellular yeast-lytic enzyme of the bacterium lysobacter sp. XL 1. Biochemistry 73, 310–314. doi: 10.1134/s0006297908030115
Sullivan, R. F., Holtman, M. A., Zylstra, G. J., White, J. F., and Kobayashi, D. Y. (2003). Taxonomic positioning of two biological control agents for plant diseases as Lysobacter enzymogenes based on phylogenetic analysis of 16s rdna, fatty acid composition and phenotypic characteristics. J. Appl. Microbiol. 94, 1079–1086. doi: 10.1046/j.1365-2672.2003.01932.x
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680. doi: 10.1093/nar/22.22.4673
Trifonova, R., Postma, J., Verstappen, F. W., Bouwmeester, H. J., Ketelaars, J. J., and Van Elsas, J. D. (2008). Removal of phytotoxic compounds from torrefied grass fibres by plant-beneficial microorganisms. FEMS Microbiol. Ecol. 66, 158–166. doi: 10.1111/j.1574-6941.2008.00508.x
Tyc, O., Van Den Berg, M., Gerards, S., Van Veen, J. A., Raaijmakers, J. M., De Boer, W., et al. (2014). Impact of interspecific interactions on antimicrobial activity among soil bacteria. Front. Microbiol. 5:567. doi: 10.3389/fmicb.2014.00567
Vasilyeva, N. V., Shishkova, N. A., Marinin, L. I., Ledova, L. A., Tsfasman, I. M., Muranova, T. A., et al. (2014). Lytic peptidase L5 of lysobacter sp. XL1 with broad antimicrobial spectrum. J. Mol. Microbiol. Biotechnol. 24, 59–66. doi: 10.1159/000356838
Wang, Y., Qian, G., Liu, F., Li, Y. Z., Shen, Y., and Du, L. (2013). Facile method for site-specific gene integration in Lysobacter enzymogenes for yield improvement of the anti-MRSA antibiotics WAP-8294A and the antifungal antibiotic HSAF. ACS Synth. Biol. 2, 670–678. doi: 10.1021/sb4000806
Weise, T., Kai, M., and Piechulla, B. (2013). Bacterial ammonia causes significant plant growth inhibition. PLoS ONE 8:e63538. doi: 10.1371/journal.pone.0063538
Xie, Y., Wright, S., Shen, Y., and Du, L. (2012). Bioactive natural products from lysobacter. Nat. Prod. Rep. 29, 1277–1287. doi: 10.1039/c2np20064c
Xu, G., and Gross, D. (1986). Selection of fluorescent pseudomonads antagonistic to Erwinia carotovora and suppressive of potato seed piece decay. Phytopathology 76, 414–422. doi: 10.1094/Phyto-76-414
Yuen, G. Y., Steadman, J. R., Lindgren, D. T., Schaff, D., and Jochum, C. (2001). Bean rust biological control using bacterial agents1. Crop Prot. 20, 395–402. doi: 10.1016/S0261-2194(00)00154-X
Zeigler, D. R. (2003). Gene sequences useful for predicting relatedness of whole genomes in bacteria. Int. J. Syst. Evol. Microbiol. 53, 1893–1900. doi: 10.1099/ijs.0.02713-0
Zhang, W., Li, Y., Qian, G., Wang, Y., Chen, H., Li, Y. Z., et al. (2011). Identification and characterization of the anti-methicillin-resistant Staphylococcus aureus WAP-8294A2 biosynthetic gene cluster from Lysobacter enzymogenes OH11. Antimicrob. Agents Chemother. 55, 5581–5589. doi: 10.1128/AAC.05370-11
Zhang, Z., and Yuen, G. Y. (2000). The role of chitinase production by Stenotrophomonas maltophilia strain c3 in biological control of Bipolaris sorokiniana. Phytopathology 90, 384–389. doi: 10.1094/PHYTO.2000.90.4.384
Keywords: Lysobacter, Rhizoctonia solani, Beta vulgaris, disease suppression, plant growth promotion
Citation: Gómez Expósito R, Postma J, Raaijmakers JM and De Bruijn I (2015) Diversity and Activity of Lysobacter Species from Disease Suppressive Soils. Front. Microbiol. 6:1243. doi: 10.3389/fmicb.2015.01243
Received: 24 August 2015; Accepted: 26 October 2015;
Published: 16 November 2015.
Edited by:
Jesús Mercado-Blanco, Consejo Superior de Investigaciones Científicas, SpainReviewed by:
Gerardo Puopolo, Edmund Mach Foundation, ItalyJane Debode, Institute for Agricultural and Fisheries Research, Belgium
Copyright © 2015 Gómez Expósito, Postma, Raaijmakers and De Bruijn. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Irene De Bruijn, aS5kZWJydWlqbkBuaW9vLmtuYXcubmw=