 Leyla Slamti
Leyla Slamti Christelle Lemy
Christelle Lemy Céline Henry
Céline Henry Eugénie Huillet
Eugénie Huillet Didier Lereclus
Didier Lereclus- Micalis Institute, INRA, AgroParisTech, Université Paris-Saclay, Jouy-en-Josas, France
In Gram-positive bacteria, cell–cell communication mainly relies on cytoplasmic sensors of the RNPP family. Activity of these regulators depends on their binding to secreted signaling peptides that are imported into the cell. These quorum sensing regulators control important biological functions in bacteria of the Bacillus cereus group, such as virulence and necrotrophism. The RNPP quorum sensor PlcR, in complex with its cognate signaling peptide PapR, is the main regulator of virulence in B. cereus and Bacillus thuringiensis (Bt). Recent reports have shown that the global stationary phase regulator CodY, involved in adaptation to nutritional limitation, is required for the expression of virulence genes belonging to the PlcR regulon. However, the mechanism underlying this regulation was not described. Using genetics and proteomics approaches, we showed that CodY regulates the expression of the virulence genes through the import of PapR. We report that CodY positively controls the production of the proteins that compose the oligopeptide permease OppABCDF, and of several other Opp-like proteins. It was previously shown that the pore components of this oligopeptide permease, OppBCDF, were required for the import of PapR. However, the role of OppA, the substrate-binding protein (SBP), was not investigated. Here, we demonstrated that OppA is not the only SBP involved in the recognition of PapR, and that several other OppA-like proteins can allow the import of this peptide. Altogether, these data complete our model of quorum sensing during the lifecycle of Bt and indicate that RNPPs integrate environmental conditions, as well as cell density, to coordinate the behavior of the bacteria throughout growth.
Introduction
Quorum-sensing (QS) is a mode of cell–cell communication that controls important adaptive processes in bacteria such as conjugation, virulence, sporulation, or competence (Dunny and Leonard, 1997; Perego, 2013; Cook and Federle, 2014; Slamti et al., 2014). This type of communication depends on the secretion of small molecules that trigger regulation mechanisms. In Gram-positive species QS mainly relies on the secretion of auto-inducing oligopeptides. These autoinducers can either act on the outside of the bacterium, by interacting with a sensor in the membrane, or in the cytoplasm of the cell. In the latter case, once internalized, the peptides bind an effector protein whose activity is subsequently modified (Perego and Hoch, 1996; Dunny and Leonard, 1997; Lazazzera et al., 1997; Miller and Bassler, 2001). Examples of these interactions are provided by the protein-peptide pairs of the RNPP family (for Rap-NprR-PrgX-PlcR, the defining members of this family; Declerck et al., 2007). The Rap phosphatases-Phr peptides are involved in competence and sporulation in Bacillus subtilis (Lazazzera et al., 1997), the PrgX transcriptional regulator-cCf10 peptide controls conjugation in Enterococcus faecalis (Suzuki et al., 1984) and NprR-NprX and PlcR-PapR, two transcriptional regulator/peptide pairs of the Bacillus cereus (Bc) group, are required for necrotrophism and virulence, respectively (Perchat et al., 2011; Dubois et al., 2012; Slamti et al., 2014).
The quorum-sensor PlcR is one of the defining proteins of the RNPP family and has been a model for understanding the mode of action of the protein-peptide complex at the molecular level (Grenha et al., 2013). PlcR is a transcription factor that regulates the expression of about 45 genes in the sporulating Gram-positive bacteria of the Bc group (Agaisse et al., 1999; Gohar et al., 2008). In addition to Bc sensu stricto, responsible for foodborne and opportunistic infections, this group comprises six other species including the causative agent of anthrax, Bacillus anthracis (Ba) and the insect pathogen Bacillus thuringiensis (Bt). PlcR is truncated and thus inactive in Ba (Agaisse et al., 1999; Mignot et al., 2001). However, the opportunistic properties of Bc and Bt (i.e., the ability to cause non-foodborne infections) are PlcR-dependent (Raymond et al., 2010). Most of the PlcR-controlled genes in Bc and Bt encode proteins whose functions are related to food supply and virulence (phospholipases, proteases, hemolysins, and toxins), cell protection and environment-sensing. Deletion of plcR results in a drastic reduction of Bt and Bc virulence in insect and mouse models of infection (Salamitou et al., 2000).
Activity of the regulator depends on the binding of the signaling peptide PapR (Slamti and Lereclus, 2002), which was shown to be imported by the oligopeptide permease Opp (Gominet et al., 2001). Oligopeptide permeases are ATP-binding cassette transporters composed of five proteins: two integral membrane proteins that form the actual pore (OppB and OppC), two ATPases bound to the membrane proteins that provide the energy required for translocation (OppD and OppF) and a membrane-anchored substrate-binding protein (SBP) facing the outside of the cell (OppA; Hiles et al., 1987; Higgins, 1992; Hiron, 2007). While it was reported that OppB-C-D-F that form the pore of the permease were required for the import of PapR (Gominet et al., 2001), the involvement of OppA, was not described. Regulation of the PlcR-PapR system is quite complex as PlcR activates its own transcription and that of the genes under its control, which include papR, at the end of the exponential growth phase (Lereclus et al., 1996; Agaisse et al., 1999). In addition, the master regulator of sporulation, Spo0A, represses the transcription of plcR during stationary phase by binding and presumably obstructing the plcR promoter region (Lereclus et al., 2000; Gominet et al., 2001). It has also been recently reported that the global regulator CodY was required for expression of plcR and PlcR-dependent genes in Bc (Frenzel et al., 2012; Lindback et al., 2012). However, the influence of CodY on the expression of the virulence factors does not appear to be via a direct binding of CodY to the promoter regions of plcR or of the PlcR-dependent genes.
CodY is conserved in low G+C Gram-positive bacteria and is a transcriptional regulator of metabolism (Sonenshein, 2005, 2007). Activity of CodY depends on cofactors such as branched-chain amino acids and GTP, depending on the bacterial species. CodY mainly directly or indirectly represses metabolism-related genes during exponential growth. When the nutrient availability decreases, it induces a drop in the intracellular concentration of GTP and amino acids available for CodY activation. This leads to the derepression of CodY-dependent genes when cells enter stationary phase. Aside from controlling the direct response to starvation, CodY is also involved in the regulation of virulence genes in several pathogens (Sonenshein, 2005; Stenz et al., 2011; Richardson et al., 2015). CodY is required for toxin expression in Ba, via the control of AtxA levels in the cell. A Ba ΔcodY deletion mutant is severely attenuated in its virulence against mice (van Schaik et al., 2009; Chateau et al., 2011). In Clostridium difficile, CodY represses toxin genes expression by direct binding to the promoter of tcdR, the sigma factor required for toxin gene transcription (Dineen et al., 2007), resulting in a hypervirulent phenotype for a ΔcodY mutant (Richardson et al., 2015). In Staphylococcus aureus, the codY deletion leads to an increase of the hemolytic activity and increased virulence in a mouse model of infection (Montgomery et al., 2012; Rivera et al., 2012). This is due to the derepression of the QS Agr-dependent genes in exponential phase (Majerczyk et al., 2008) and a recent report showed that this CodY-mediated regulation was indirect (Roux et al., 2014). However, as for Bt and Bc, the authors did not report the mechanism responsible for the effect of CodY on the virulence regulators.
Here, we investigated the mechanism underlying this regulation. We report that CodY is involved in the expression of Bt virulence factors through its role in the import of the signaling peptide PapR. Using proteomics and genetics approaches, we show that a ΔcodY mutant is impaired in its ability to import PapR. We also show that OppA is not the only SBP involved in the recognition of PapR, and that several other OppA-like proteins can allow the import of this peptide. This report brings new insights into the regulation of virulence gene expression and its link to the metabolic state of the bacteria.
Materials and Methods
Bacterial Strains and Growth Conditions
The acrystalliferous B. thuringiensis 407 Cry- strain (Bt 407-; Lereclus et al., 1989) was used as the parental strain to create all the strains used in this study. Escherichia coli strain DH5α (Taylor et al., 1993) was used as the host strain for plasmid construction. E. coli strain ET12567 (MacNeil et al., 1992) was used to prepare DNA prior to electroporation in B. thuringiensis. Unless otherwise noted, cells were grown in LB medium (1% tryptone, 0.5% yeast extract, 1% NaCl) at 37°C and stored at -80°C in LB containing 15% glycerol.
For B. thuringiensis cultures, t0 corresponds to the beginning of the transition between the exponential and stationary growth phases. It is defined as the point where the slope starts to decrease at the end of the exponential phase.
Columbia agar plates (BioMérieux) containing 5% sheep blood were used to evaluate the hemolytic activity of the B. thuringiensis strains. BHI (Beckton-Dickinson) agar plates containing 5% egg yolk were used to evaluate the lecithinase activity of the B. thuringiensis strains.
The antibiotic concentrations used for selection of B. thuringiensis and E. coli were as follows: erythromycin, 10 μg/mL; kanamycin, 200 μg/mL; ampicillin, 100 μg/mL.
When required, xylose was used at a concentration of 20 mM.
Strain Construction
The plasmids used for strain construction are listed in Table 1. A list of the strains used in this study is presented Table 2. Strain Bt ΔcodY was constructed as follows. pRN-ΔcodY was integrated in the chromosome of Bt 407- following a single recombination event as described previously (Lereclus et al., 1992) and verified by PCR using primer pairs codY7/RN3 and RN2/codYVerifRev (Supplementary Table S1). The resulting Bt codY::pRN-ΔcodY cells were then transformed with p1618K-Pxyl or p1618K-Pxyl’-codY. The second recombination event was allowed to proceed in these strains in parallel with the parental Bt codY::pRN-ΔcodY cells. Occurrence of this event was screened by PCR using primer pair codY7/codYDnBam (Supplementary Table S1) on the chromosome of ErmS (and KanR for the strains harboring the replicative plasmid) cells. Supplementary Table S2 shows the number of clones tested to screen for codY deletion. We could only obtain a deletion of the codY gene in cells harboring a copy of codY in trans. We then cultured Bt ΔcodY (p1618K-Pxyl’-codY) cells in parallel with the Bt (p1618K-Pxyl’-codY) strain and compared the percentage of cells that lost the plasmid in these two strains (Supplementary Materials and Methods). Supplementary Table S3 shows the number of clones tested. Only 10% of the Bt ΔcodY cells had lost the plasmid at the end of the experiment. In contrast, there was a marked loss of the plasmid in wild-type Bt cells. We chose one of the Bt ΔcodY mutants to pursue our experiments. In order to complement the mutation this clone was transformed with (p1618K-Pxyl’-codY) resulting in strain Bt ΔcodY (p1618K-Pxyl’-codY)B.

TABLE 1. Plasmids used this study.
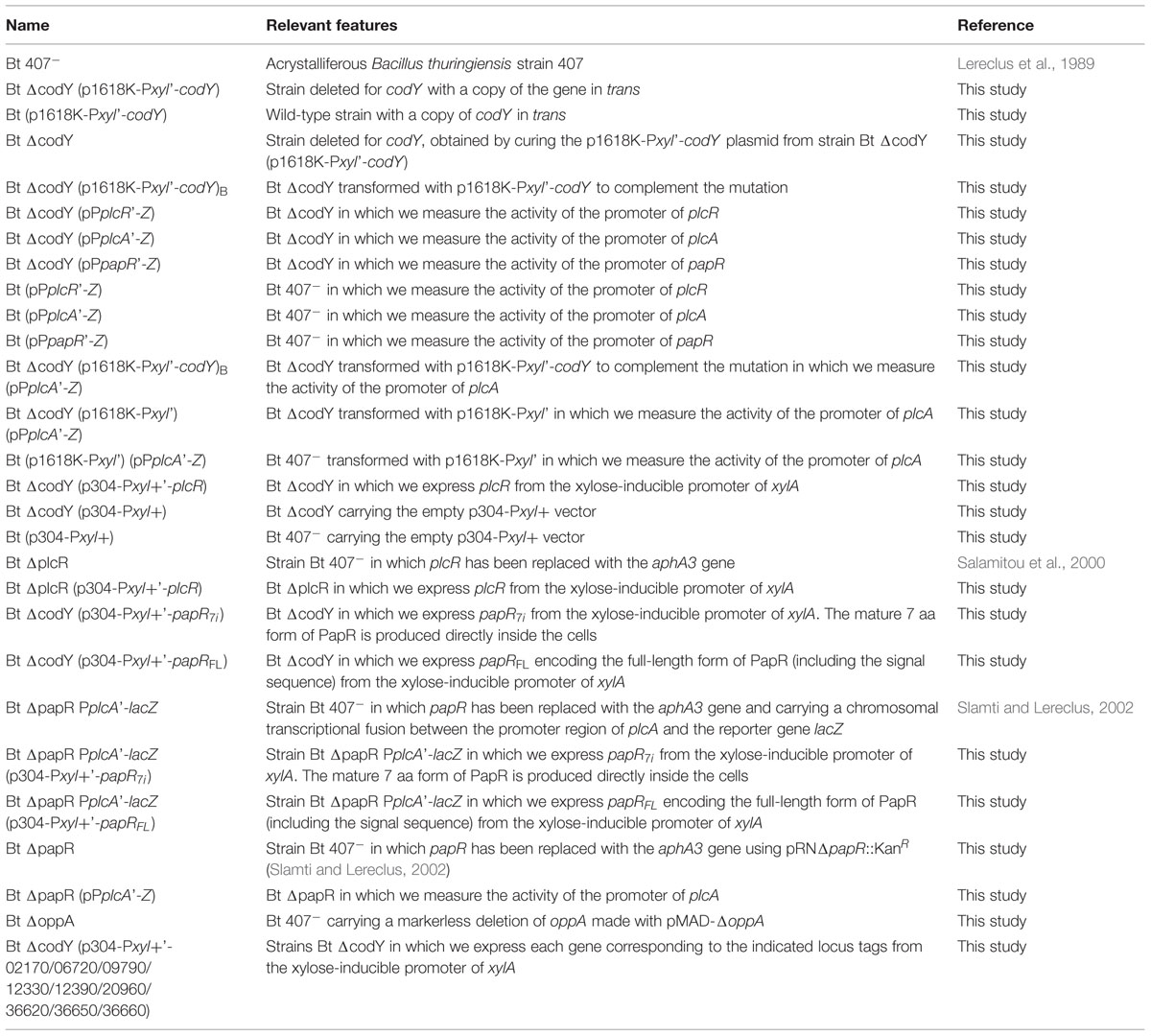
TABLE 2. Strains used this study.
β-galactosidase Assay
β-galactosidase activities were measured as described previously with the exception of the incubation temperature which was set to 30°C instead of room temperature (Perchat et al., 2011). Each assay was repeated at least twice.
Lecithinase Assay
Two mL samples were harvested from cell cultures by centrifugation at the indicated times for each experiment. The supernatant was then filter-sterilized and kept at 4°C overnight. 180 μL of a 0.5% egg yolk saline solution were distributed in a 96-well plate and 20 μL of each filtered supernatant was added to two wells. This was considered time zero of the assay. The plate was then incubated at 25°C in a Tecan Infinite F200 Pro reader for up to 20 h and the OD600 was recorded every 10 min after a short vigorous shaking. OD600 obtained from wells containing the supernatant of Bt ΔpapR PplcA’-lacZ (a lecithinase minus strain) were substracted from the OD600 corresponding to the strains of interest. The resulting OD600 were then plotted as a function of time and the slope of each curve was determined from the linear part of the curve. A specific activity was determined using the following formula: slope/[OD600 of the culture at the time of sampling × volume of supernatant used in the assay (L)]. This formula was adapted from (Taniguchi et al., 2009). Each assay was repeated at least twice.
Analysis of the Wild-Type and ΔcodY Cell Culture Supernatants Using Mass-spectrometry
To facilitate detection of PapR in a peptide-poor medium, Bt 407-, Bt ΔcodY and Bt ΔpapR PplcA’-lacZ cells were grown at 37°C in S medium (1.7% synthetic broth AOAC, 0.2% NaCl) supplemented with 0.3% glucose instead of LB. In this medium, the expression profile of the PlcR-dependent gene plcA was similar to that in LB in the strains used (data not shown). 1 mL samples were harvested from cell cultures by centrifugation 30 min after entry into stationary phase. The supernatant was then filter-sterilized and kept at -20°C until analysis.
Peptides produced in S medium + glucose were isolated from 1 ml of supernatant by solid-phase extraction on StataX cartridges (Phenomenex) using a 50% acetonitrile solution for the elution step. Peptides were then concentrated by evaporation. Dried fractions were resuspended in 40 μl of 0.1% TFA, 2% acetonitrile and peptides abundance were measured by LC–MS/MS on an LTQ-Orbitrap mass spectrometer at the PAPPSO platform (http://pappso.inra.fr; See Supplementary Material and Methods for details).
Analysis of the Opp Content in Membrane-Enriched Fractions of Wild-Type and ΔcodY Cells Using Mass-spectrometry
Membrane-Enriched Fraction Preparation
Bt 407- and Bt ΔcodY cells were grown at 37°C in LB and samples were collected by centrifugation 1 h before (t-1), at (t0), and 1 h after (t1) the transition from exponential to stationary phase (100, 50, and 50 mL, respectively). The pellets were washed once with PBS and stored at -20°C until further processing. The pellets were thawed, resuspended in lysis buffer [KPO4 pH6.6 100 mM, lysozyme 100 μg/mL, anti-protease cocktail (Roche) 1x] at an OD600 = 35 and incubated at 37°C for 30 min. The suspensions were sonicated on ice and sonication efficiency was verified by microscopic observation. A nuclease cocktail (DNAseI 10 μg/mL, RNaseA 10 μg/mL, MgCl2 1 mM, CaCl2 1 mM was added to the samples that were then incubated on ice for 1 h. The suspensions were centrifuged at 5000 g for 10 min at 4°C and the supernatant was then further centrifuged at 100000 g for 1 h at 4°C. The pellets were then resuspended in Laemmli sample buffer (Sigma) and kept at 4°C overnight until completely solubilized before protein concentration was measured using the 2D Quant kit (GE Healthcare). This experiment was repeated three times and 10 μg of proteins from each sample were then loaded on 12% SDS-polyacrylamide gels poured together form the same master mix. A short migration was then performed.
Protein In-gel Digestion
Each lane of short migration was cut and washed for 15 min with an acetonitrile/100 mM ammonium bicarbonate mixture (1:1). Digestion was performed in 50 mM ammonium bicarbonate pH 8.0 and the quantity of modified trypsin (Promega, sequencing grade) was 0.1 μg per sample. Digestion was achieved for 6 h at 37°C. The supernatant was conserved. Peptides were extracted by 5% formic acid in water/acetonitrile (v/v). Supernatant and extracted tryptic peptides were dried and resuspended in 50 μl of 0.1% (v/v) formic acid and 2% (v/v) acetonitrile. Proteins were detected and quantified by spectral counting approach by LC-MS/MS on a Q-Exactive mass spectrometer at the PAPPSO platform (http://pappso.inra.fr; See Supplementary Material and Methods for details). The raw data were submitted to the Pride database (Vizcaino et al., 2013) as a Proteom Xchange dataset (http://www.ebi.ac.uk/pride/archive/projects/PXD003311).
For ease of reading we removed “BTB_c” or “BTB_RS” from the locus tags and only kept the number associated.
The threshold for being referenced in Supplementary Table S5 was a mean number of spectra of at least 2 in at least 1 condition for the wild-type and the ΔcodY samples.
Peptide Synthesis
Synthetic PapR7 (ADLPFEF) and PapR27 (DTAFEKSQIISHNDQEVQVAADLPFEF) were ordered from Genscript at a purity >95% and were resuspended in water.
Results
PlcR-Dependent Genes are Down-regulated in a Bt ΔcodY Mutant
We decided to construct a ΔcodY mutant in strain Bt 407- in order to examine the effect of this regulator in a genetic background in which the PlcR-PapR quorum-sensing system has been extensively characterized (for a review see Slamti et al., 2014). This strain is also capable of completing a full infectious cycle in our insect model Galleria mellonella. After unsuccessful attempts at disrupting codY by homologous recombination, it appeared that we could only delete codY in the presence of a copy of the gene in trans. We obtained the Bt ΔcodY strain after curing the cells of the plasmid carrying the ectopic copy of codY (see Materials and Methods). We cannot exclude that generation of this mutant might have resulted in the selection of one or more suppressor mutations elsewhere in the genome. ΔcodY mutant cultures presented a slower growth rate in exponential phase and reached lower OD600 during stationary phase than wild-type cell cultures (Supplementary Figure S1). The mutant cells look also more elongated and thinner than wild-type cells and seem to present a chaining phenotype, in particular during exponential phase (Supplementary Figure S2). Disruption of the codY gene resulted in a drastic loss of hemolytic and lecithinase activities on sheep blood and egg yolk agar plates, respectively (Supplementary Figure S3). These activities are due to the production of PlcR-dependent gene products (Salamitou et al., 2000; Slamti et al., 2004). Introducing p1618K-Pxyl’-codY, a plasmid carrying the codY gene under the control of the xylose-inducible promoter Pxyl, in the ΔcodY mutant restored the hemolytic and lecithinase activities of this strain (Supplementary Figure S3), confirming that the loss of these activities was due to the deletion of codY. These phenotypes are in agreement with what has been previously reported for B. cereus (Frenzel et al., 2012; Lindback et al., 2012).
We also measured the transcriptional activity of three PlcR-dependent promoters, fused to the reporter gene lacZ, in wild-type and ΔcodY cells (Figure 1). The activity of the plcR and papR promoters is reduced by 60 and 70% in the ΔcodY mutant compared to wild-type cells, respectively. In contrast, the activity of plcA, encoding the PI-phospholipase PlcA, is almost abolished in the mutant compared to the wild-type. Transcription of plcA was restored in ΔcodY cells complemented with a copy of codY in trans (Supplementary Figure S4). In aggregate, these results show that CodY positively regulates the transcription of PlcR-dependent genes in Bt 407-, either indirectly or by direct interaction with the promoter of plcR or papR.
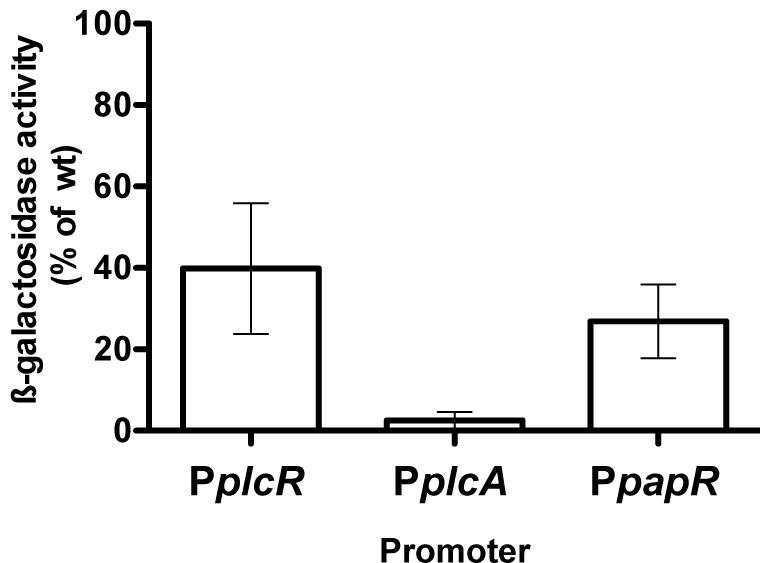
FIGURE 1. Expression of PlcR-dependent genes in Bt ΔcodY compared to the wild-type strain. ΔcodY and wild-type cells harbor a transcriptional fusion between the promoter of interest and the lacZ gene. Cells were grown in LB at 37°C and harvested 2 h after entry into stationary phase. The β-galactosidase specific activity was then assayed. The results are the mean values of at least two independent experiments, and the error bars represent standard deviation.
CodY Controls the Activity of PlcR
As plcR is autoregulated, we needed to determine if CodY was involved in the transcriptional activation of plcR or in the activity of the protein itself. We constructed p304-Pxyl+’-plcR, a plasmid carrying the plcR gene under the control of a modified version of the xylose-inducible promoter Pxyl (see Materials and Methods). This allowed us to uncouple plcR transcription from the activity of its product. p304-Pxyl+’-plcR was introduced in Bt ΔplcR and Bt ΔcodY. Lecithinase production was then assayed in these strains and compared to that of the wild-type strain (Figure 2). As plcB, the gene encoding the lecithinase PC-PLC, is under the control of PlcR and is absent in the supernatant of a ΔplcR mutant (Agaisse et al., 1999; Gohar et al., 2002, 2008), the activity of its product reflects the activity of the regulator. No lecithinase activity could be detected at t1 (1 h after the onset of stationary phase) in the ΔcodY and ΔplcR mutants. Lecithinase activity was restored to almost wild-type levels in Bt ΔplcR (p304-Pxyl+’-plcR) showing that our assay was functional, i.e., that plcR is transcribed and active in this strain. However, we could not detect any lecithinase activity in Bt ΔcodY (p304-Pxyl+’-plcR) indicating that PlcR is not active in this strain and therefore that CodY controls PlcR activity.
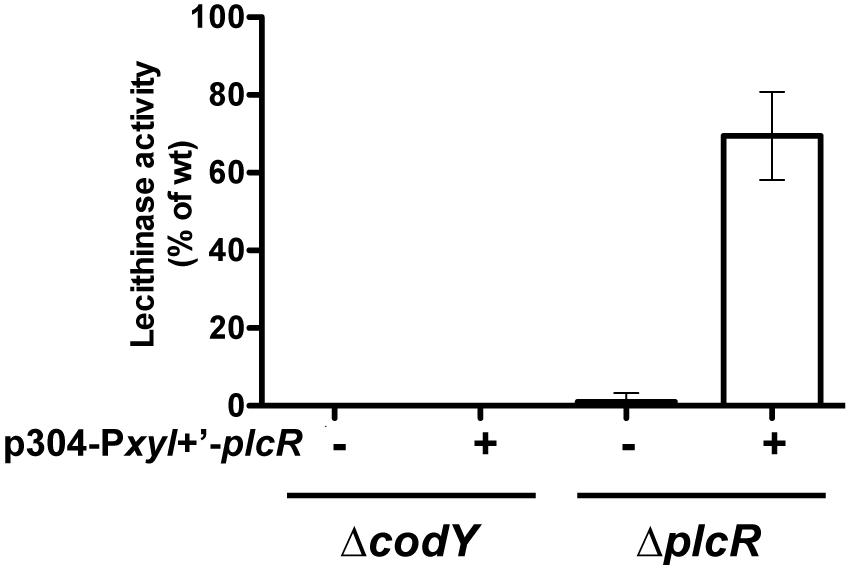
FIGURE 2. Lecithinase production in Bt ΔcodY compared to the wild-type strain when plcR is overexpressed. ΔcodY and ΔplcR cells harbor (+) or not (-) the p304-Pxyl+’-plcR vector carrying a transcriptional fusion between the xylose-inducible promoter PxylA and the plcR gene. Filter-sterilized supernatants from samples harvested 1 h after entry into stationary phase were assayed for lecithinase production. The results are the mean values of two independent experiments, and the error bars represent standard deviation.
Activity of PlcR is Restored by Expressing the Mature form of PapR Inside ΔcodY Cells
It has been shown that the activity of PlcR directly depends on the binding of the PapR peptide (Slamti and Lereclus, 2002). We investigated whether loss of the PlcR-dependent phenotypes in the codY mutant was due to an effect of CodY on either step of PapR production (export, maturation, or import). We have already shown that papR transcription was strongly reduced, but not abolished, in ΔcodY cells (Figure 1). We introduced p304-Pxyl+’-papR7i or p304-Pxyl+’-papRFL in the ΔcodY and ΔpapR mutant cells. The former plasmid allows the production of the mature 7 aa form of PapR directly inside the cells. The latter encodes the full-length version of PapR, including the signal sequence. We then assayed the lecithinase activity of these strains and compared it to that of the wild-type strain (Figure 3). As above, no lecithinase activity could be detected at t1 in the ΔcodY mutant. The lecithinase activity for ΔpapR cells was very low and the measurements obtained for this mutant were used to normalize the results (see Materials and methods). Therefore the data corresponding to this strain could not be plotted on this graph. Figure 3 shows that lecithinase activity was above wild-type levels in Bt ΔpapR PplcA’-lacZ (p304-Pxyl+’-papR7i) and Bt ΔpapR PplcA’-lacZ (p304-Pxyl+’- papRFL) showing that our assay was functional, i.e., that papR7i and papRFL are transcribed and active in these strains. The same result was observed for Bt ΔcodY (p304-Pxyl+’-papR7i) indicating that expressing the mature form of PapR inside ΔcodY cells is enough to restore the production of the PlcR-dependent protein PC-PLC. In contrast, lecithinase production was only 16% of that of the wild-type in Bt ΔcodY (p304-Pxyl+’-papRFL) at t1. Since PapRFL is not able to complement the codY mutation, these data suggest that export and/or maturation and/or import of PapR is affected in ΔcodY mutant cells.
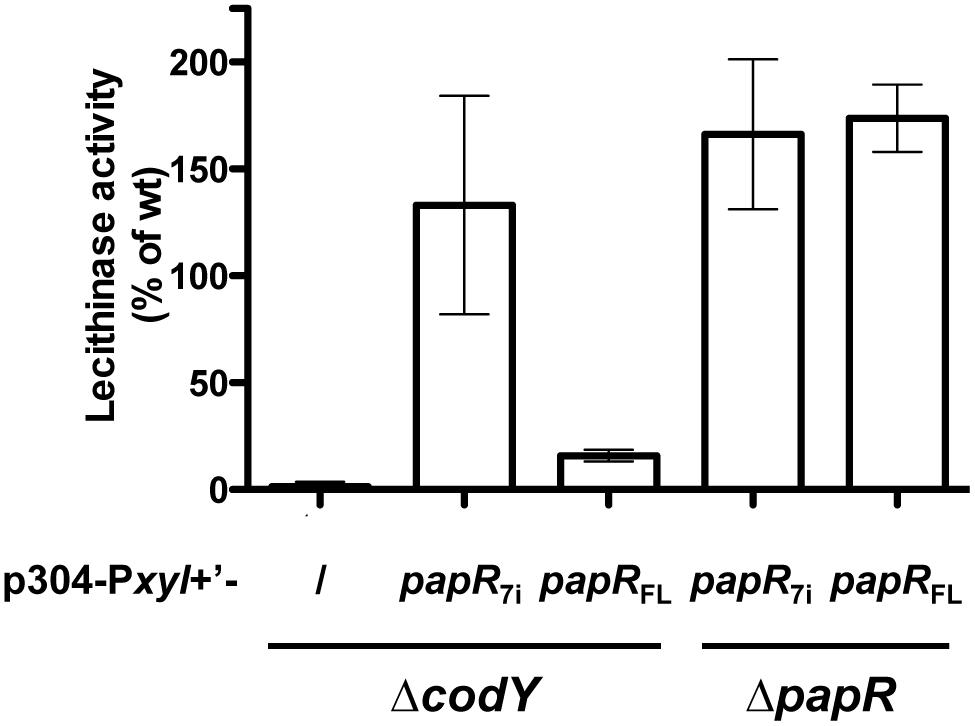
FIGURE 3. Lecithinase production in Bt ΔcodY compared to the wild-type strain when the mature form of PapR is produced inside the cells. ΔcodY and ΔpapR cells harbor the p304-Pxyl+’ vector carrying a transcriptional fusion between the xylose-inducible promoter PxylA and either papR7i, coding for the mature form of PapR (7 aa), or papRFL, coding for full length PapR (including the signal sequence). / indicates that the cells are devoid of these plasmids. Filter-sterilized supernatants from samples harvested 1 h after entry into stationary phase were assayed for lecithinase production. The results are the mean values of two independent experiments, and the error bars represent standard deviation.
PapR7 is Less Abundant in the Culture Supernatant of a ΔcodY Mutant than in the Culture Supernatant of Wild-Type Cells
To determine which step of PapR production was controlled by CodY, we examined if the signaling peptide could be detected in the supernatant of Bt ΔcodY. The supernatant of wild-type and ΔcodY cells was harvested at t0.5 and filter-sterilized before being subjected to LC-MS/MS analysis. The arrows on the PapR peptide sequence in Figure 4 indicate the main cleavage sites deduced from the peptides found in the supernatant of both strains. Table 3 and Figure 4 show the sequence and abundance of each peptide found either in the wild-type or the ΔcodY culture supernatant or in both. The most commonly found N-terminal end is the alanine in position 15 in both cases. However, the similitude does not much extend further as the only other common cleavage site is after the alanine in position 41 (green arrows in Figure 4). The PapR-derived peptidic sequences found in the two supernatants are otherwise different, suggesting a different processing between the strains (Table 3 and compare the purple and orange arrows in Figure 4). It is also important to note that the abundance of PapR7 (ADLPFEF) was lower in the supernatant of the ΔcodY cells than in that of the wild-type strain, as this could result in the inability to reach the concentration required for the activation of PlcR.
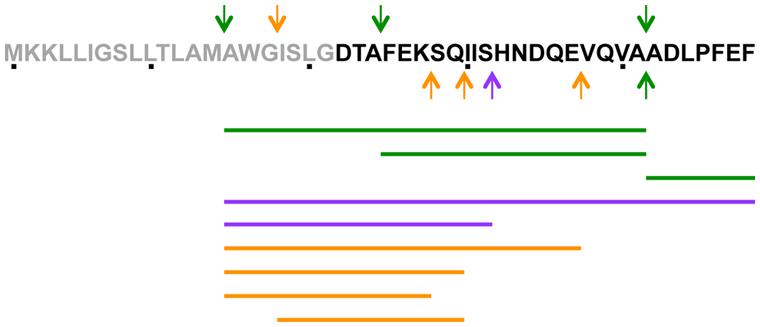
FIGURE 4. Schematic representation of the presence of PapR-derived peptides in the supernatant of Bt ΔcodY compared to the wild-type strain. The culture supernatant of ΔcodY and wild-type cells was filter-sterilized before concentration and injection in an LTQ-Orbitrap for LCMS/MS analysis. Cells were grown in synthetic broth + glucose at 37°C. Samples were collected 0.5 h after entry into stationary phase. The arrows on the PapR peptide sequence indicate the main cleavage sites (in N-terminus, above the sequence and in C-terminus, below the sequence) deduced from the peptides found in the supernatant of both strains. The lines below the peptidic sequence schematize the peptides found in the supernatants. Green refers to cleavage sites and peptides common to ΔcodY and wild-type, purple are specific to wild-type and orange to ΔcodY.
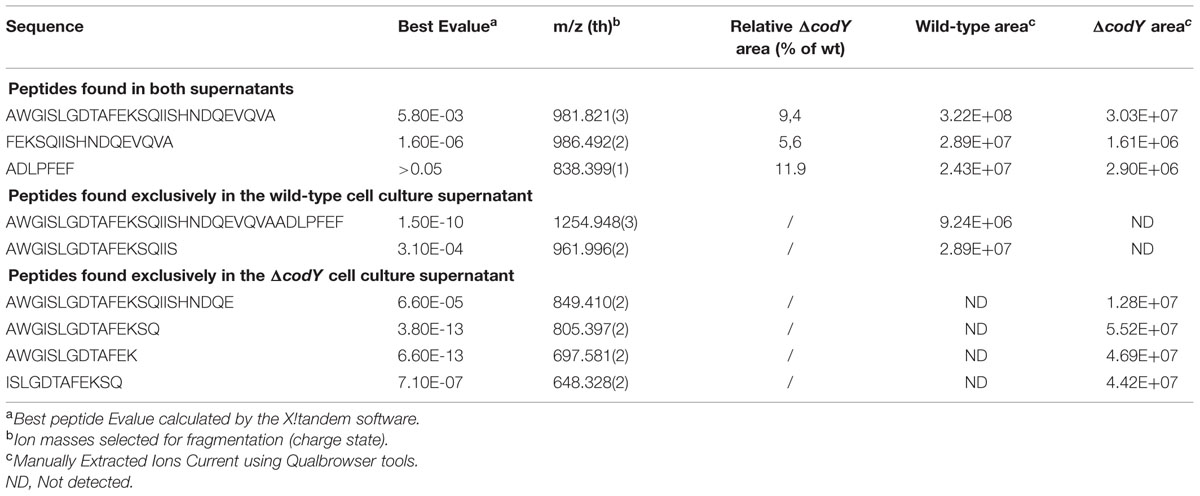
TABLE 3. PapR-derived peptides detected in wild-type and ΔcodY cell culture supernatants.
The Culture Supernatant of the Bt ΔcodY Strain Contains Enough PapR to Complement ΔpapR Cells
We then examined if the amount of PapR present in the ΔcodY culture supernatant was able to complement a ΔpapR mutant for the expression of a PlcR-dependent gene. β-galactosidase production was assayed in Bt ΔpapR PplcA’-lacZ cells grown in the presence of supernatant harvested from wild-type, ΔcodY or ΔpapR cell cultures at t0.5. Figure 5 shows that the activity of the plcA promoter in Bt ΔpapR PplcA’-lacZ cells grown in the presence of supernatant from ΔpapR cells is about 10% of its activity compared to when they are grown in conditioned medium from wild-type cells. This is consistent with what has been previously reported (Slamti and Lereclus, 2002). In contrast, supernatant from ΔcodY cell cultures was able to partially complement Bt ΔpapR PplcA’-lacZ cells and to induce transcription at the plcA promoter to 70% of its activity compared to when cells are cultured in supernatant from wild-type cells. This indicates that Bt ΔcodY cells secrete enough PapR to activate the PlcR regulator.
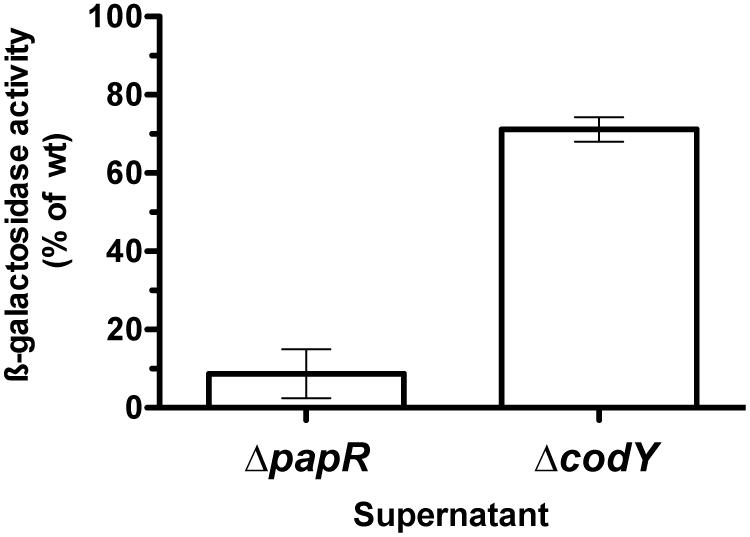
FIGURE 5. Extracellular complementation assay of ΔpapR cells with conditioned medium from ΔcodY compared to wild-type cells. The culture supernatant of ΔcodY, ΔpapR and wild-type cells was filter-sterilized before being used to resuspend ΔpapR cell pellets. The latter harbor a chromosomal transcriptional fusion between the promoter of plcA and lacZ. Cells were grown in LB at 37°C. All samples were collected 0.5 h after entry into stationary phase. β-galactosidase production was assayed after resuming growth for 1.5 h. The results are the mean values of two independent experiments, and the error bars represent standard deviation.
Import of PapR is Impaired in ΔcodY Cells
To directly assess if PapR could be reimported in the ΔcodY mutant, we measured β-galactosidase production in Bt ΔcodY (pPplcA’-Z) cells grown in the presence of synthetic PapR7 peptide (Figure 6). The results show that the ß-galactosidase activity in these cells is 27% of that of Bt (pPplcA’-Z), whereas it reaches 137% in Bt ΔpapR (pPplcA’-Z) cells cultured in the same conditions. This indicates that deletion of codY impairs the import of the mature form of PapR in Bt cells.
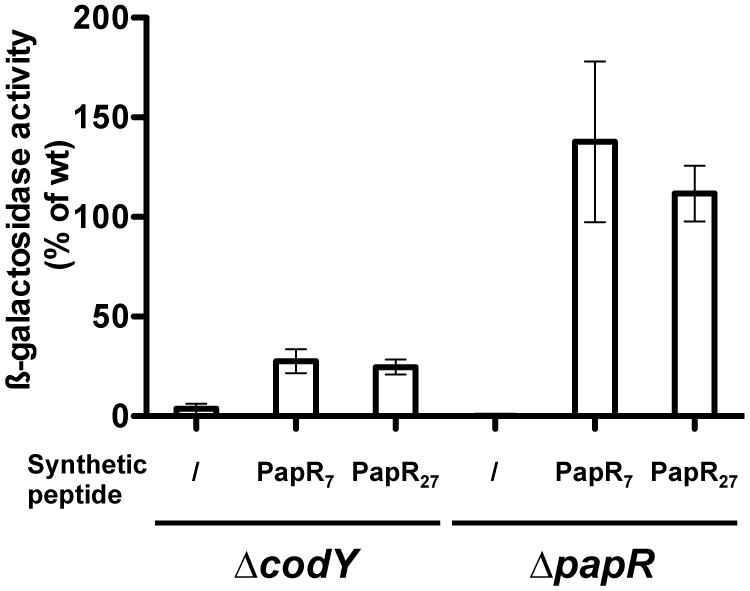
FIGURE 6. Extracellular complementation assay of ΔcodY cells with synthetic PapR peptides. ΔcodY and ΔpapR cell cultures were split at the entry of stationary phase and synthetic peptides were added when indicated at a final concentration of 5 μM. The cells harbor the pPplcA’-Z plasmid, carrying a transcriptional fusion between the promoter of plcA and lacZ. Cells were grown in LB at 37°C. Samples were collected for β-galactosidase assay 2.5 h after addition of the peptides. The sequence of the PapR peptides is indicated in Section “Materials and Methods.” The results are the mean values of two independent experiments, and the error bars represent standard deviation.
Furthermore, synthetic PapR27, which corresponds to full length PapR (excluding the predicted signal sequence) also induced the same activation of PplcA’-Z in ΔcodY as did PapR7, suggesting that maturation of PapR can occur in this strain. PapR7 and PapR27 also induced the same activation of PplcA’-Z in ΔpapR cells.
OppA is Involved in -but is Dispensable for- the Import of PapR
The pore components required for proper activation of the expression of the PlcR regulon were shown to be OppB-C-D-F (corresponding to proteins 12340, 12350, 12360, and 12370, respectively; Gominet et al., 2001). However, no information was reported about the SBP. We examined if OppA (protein 12330) was responsible for the import of PapR. We assayed lecithinase production of ΔcodY cells in which we overexpressed oppA. We showed that expression of oppA induces an increase in lecithinase activity in the ΔcodY mutant in the presence of PapR7, compared to that of the strain in which the gene is not expressed (about 60 and 10% of the activity of wild-type samples, respectively; Figure 7). This strongly supports the direct involvement of OppA in the import of the peptide. We then deleted the oppA gene and assessed the hemolytic activity of the Bt ΔoppA strain on sheep blood agar plates as a reporter of PlcR activity. Contrary to what was reported previously for a ΔoppB mutant (Gominet et al., 2001), there was no obvious difference between the ΔoppA mutant and the wt strain (Supplementary Figure S5), indicating that, although OppA is involved in the import of PapR, other SBPs are able to fulfill the same function.
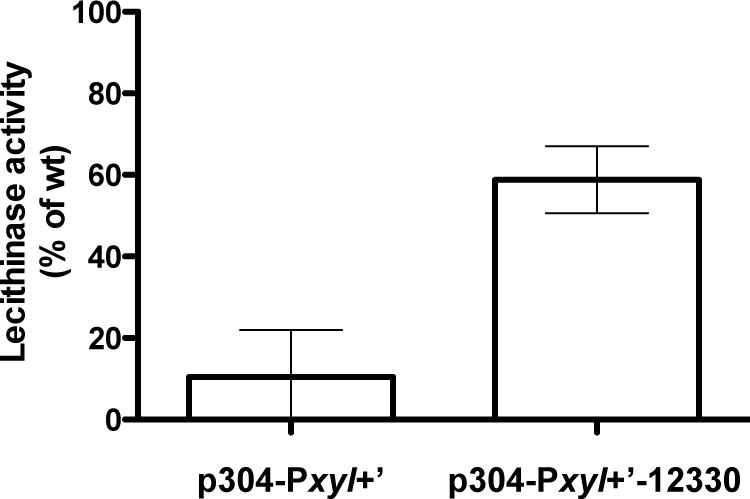
FIGURE 7. Lecithinase activity of ΔcodY cells overexpressing oppA in the presence of synthetic PapR peptide. ΔcodY cells harbor the empty p304-Pxyl+’ vector or express the oppA gene under the control of the Pxyl+ promoter (p304-Pxyl+’-12330). The cultures were split 0.5 h before the transition into stationary phase (t-0.5) and the synthetic peptide PapR7 was added at a final concentration of 5 μM. Cells were grown in LB supplemented with xylose at 37°C. Samples were collected for lecithinase assay 2.5 h after addition of the peptide. The sequence of the peptide is indicated in Section “Materials and Methods.” The results are the mean values of two independent experiments, and the error bars represent standard deviation.
Several Opp-Like Proteins are Differentially Detected in ΔcodY Membrane-Enriched Fractions Compared to Wild-Type Fractions
In order to assess which permease component involved in the import of PapR was affected by the codY mutation, and because Bt harbors 40 genes that are predicted to encode a component of potential oligopeptide permease systems (Supplementary Table S4), we chose to use a general approach. Membrane enriched fractions of wild-type and ΔcodY cells harvested 1 h before (t-1), at (t0), and 1 h after (t1) the transition from exponential to stationary phase were prepared and subjected to mass spectrometry analysis. Label free quantification of proteins was achieved using spectral counting which allows the relative quantification of proteins based on the number of spectra obtained with tryptic peptides in mass spectrometry. This quantification relies on the fact that the more of a particular protein is present in a sample, the more MS spectra are detected for peptides from that protein. We identified a total of 1366 proteins in all the samples (the complete results are available on the PRIDE database (Vizcaino et al., 2013) as a Proteom Xchange dataset under the submission number PXD003311). 22% of these proteins were predicted to be membrane-associated using the PSORTb v3.0 software (Yu et al., 2010). Nineteen Opp-like proteins passed our detection threshold. They are listed in Supplementary Table S5. We could identify all the components of the permease required for the import of PapR (i.e., OppB-C-D-F). The results show that there was no difference in the detection of the pore components at t-1 in wt and ΔcodY cells (Figure 8A and Supplementary Table S5). However, there was less OppB-C-D-F in the codY mutant than in the wt strain at t0, and, at t1, almost none of these proteins were detected in the ΔcodY membrane-enriched fraction (Figures 8B,C and Supplementary Table S5). In contrast, OppA (protein 12330) seems to follow a different pattern. Figure 8A shows that more OppA was detected in the ΔcodY sample than in the wt preparation. At t0 there was no significant difference between the samples, and, at t1, no OppA was detected in ΔcodY membrane-enriched preparation (Figures 8B,C and Supplementary Table S5).
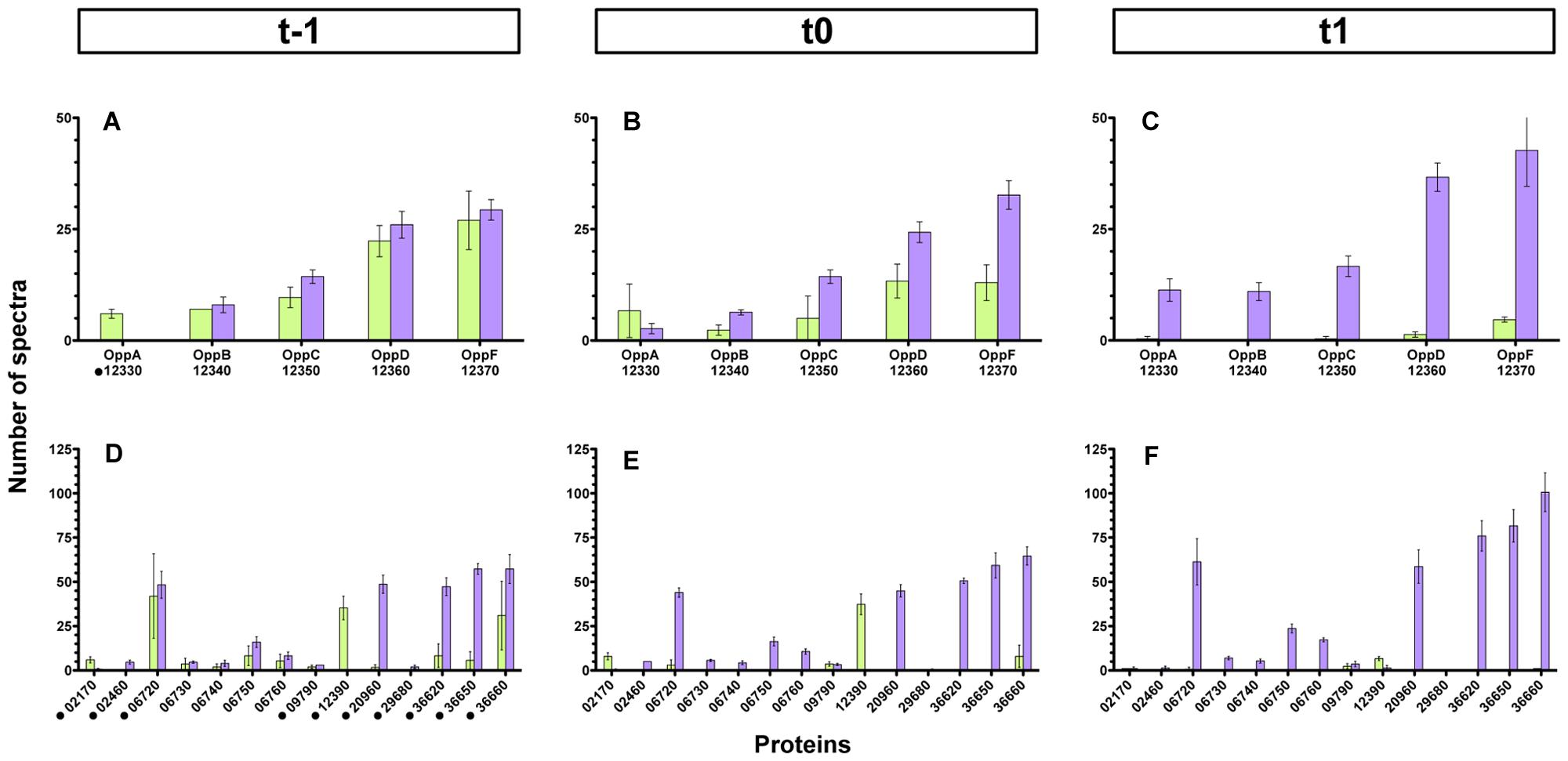
FIGURE 8. Detection of Opp-like proteins in membrane-enriched fractions from ΔcodY and wild-type cells. Membrane-enriched fractions obtained from ΔcodY and wild-type cells harvested 1 h before (t-1, A,D), at (t0, B,E), or 1 h after (t1, C,F) the transition into stationary phase were analyzed using mass spectrometry. The histograms represent the number of spectra detected in the samples for each protein indicated on the x-axis. The upper panels report the data for the components of the oligopeptide permease Opp of which the pore is required for PapR import. The lower panels report the data for the other Opp-like proteins detected. The results are the mean values of three independent experiments, and the error bars represent standard deviations. The black dots in (A,D) indicate an OppA-like protein. Purple bars, wild-type samples; green bars, ΔcodY samples.
Interestingly, other Opp-like proteins were differentially detected in the wild-type and ΔcodY samples. Panels D, E and F of Figure 8 (as well as Supplementary Table S5) recapitulate all these proteins. Protein 12390 was significantly more represented in the ΔcodY samples than in the wt preparations at all times. However, its number of spectra was strongly reduced between t0 and t1. To a lesser extent, protein 02170 was also significantly more represented in the ΔcodY samples than in the wt preparations, except at t1 where it was no longer detected. In contrast, proteins 20960, 36620, 36650, and 36660 saw their level drastically reduced in the ΔcodY membrane-enriched fraction compared to their wt level, at all times. The level of protein 06720 was also drastically reduced in the ΔcodY background compared to its wt level, but only at t0 and t1. Proteins 06730, 06740, 06750, and 06760 follow the same pattern but with less amplitude. The other proteins show little -or no significant- difference between the samples.
Ectopic Over-expression of Selected OppA-Like-Encoding Genes Partially Restores the Hemolytic Phenotype of ΔcodY Cells
In order to assess if we could restore the import of PapR in ΔcodY cells, we over expressed seven genes encoding for OppA-like proteins that were less abundant in Bt ΔcodY than in the wt strain [06720, 12330 (OppA), 20960, 36620, 36650, 36660], according to the results presented above. We also overexpressed two genes encoding OppA-like proteins that were more abundant in Bt ΔcodY than in the wt strain (02170 and 12390) as a control. We then assayed the hemolytic activity of these strains. The results presented in Figure 9 show that over-expression of some OppA-like-encoding genes, but not all, partially restored the hemolytic activity of ΔcodY cells. We observed the same results when monitoring the lecithinase activity of these cells (Supplementary Figure S6). Over-expression of the 02170 and 12390 genes did not induce hemolysis of the red blood cells. This is in agreement with the fact that their corresponding proteins were more abundant in ΔcodY than in wt preparations (Figure 8, lower panels). Interestingly, over-expression of gene 06720 does not have any effect on the hemolytic phenotype of the ΔcodY cells, despite the fact that it was more abundant in the wild-type than in the ΔcodY samples (Figure 8, lower panels). This result suggests that this protein is probably not involved in the uptake of PapR. In contrast, over-expression of gene 12330, which encodes OppA, and of genes 20960, 36620, 36650 and 36660, which are more abundant in the wt samples than in the ΔcodY samples (Figure 8, lower panels), induce an increase in the hemolytic activity of the ΔcodY cells, suggesting that they are all able to bind PapR and allow its import.

FIGURE 9. Hemolytic activity of ΔcodY cells over-expressing oppA and oppA-like-encoding genes. Cells were patched onto a Columbia sheep blood agar plate supplemented with xylose and grown at 37°C for 16 h. The cells either harbor the empty vector p304-Pxyl+ or overexpress our gene of interest under the control of a xylose-inducible promoter. wt, Bt 407- (p304-Pxyl+); ΔcodY, Bt ΔcodY (p304-Pxyl+); (02170), Bt ΔcodY (p304-Pxyl+-02170); (06720), Bt ΔcodY (p304-Pxyl+-06720); (09790), Bt ΔcodY (p304-Pxyl+-09790); (12330), Bt ΔcodY (p304-Pxyl+-12330); (12390), Bt ΔcodY (p304-Pxyl+-12390); (20960), Bt ΔcodY (p304-Pxyl+-20960); (36620), Bt ΔcodY (p304-Pxyl+-36620); (36650), Bt ΔcodY (p304-Pxyl+-36650); (36660), Bt ΔcodY (p304-Pxyl+-36660). This picture is representative of at least three independent experiments.
Discussion
We assessed the mechanism underlying the control of the global regulator CodY on the expression of the PlcR-dependent virulence genes in Bt, an entomopathogen of the Bc group. Involvement of CodY in the expression of these genes had been demonstrated in two Bc strains, but no mechanistic explanation has been reported (Frenzel et al., 2012; Lindback et al., 2012). Several unsuccessful attempts at deleting the codY gene in Bt 407-, our reference strain, had been made in our laboratory and the difficulties we encountered to generate our mutant strongly suggest that this gene is essential for the survival of this strain in our laboratory conditions. A codY deletion mutant in Bt 407- could only be generated in a strain harboring an ectopic copy of the gene. The loss of the plasmid carrying this copy resulted in obtaining a marker-less ΔcodY mutant. It is possible that the mutant we obtained contains mutations allowing for the growth of Bt in the absence of codY. It has been reported that CodY was essential in two Streptococcus pneumoniae strains (Caymaris et al., 2010). The authors speculated that codY inactivation might lead to an increase in iron uptake and that iron toxicity might explain the lethality of this mutation. To survive in the presence of the codY knock-out, the cells contained additional mutations in the putative iron transporters. In Bt, we showed that the hemolytic and lecithinase activities linked to PlcR-regulated genes were abolished in the ΔcodY mutant and were restored when the codY mutation was complemented in trans. This indicates that the regulatory link between PlcR and CodY was not affected by these hypothetical suppressor mutations. There is no mention of the putative essential nature of the codY gene in the previous reports using ΔcodY mutants in 3 Bc strains of differing origins (Hsueh et al., 2008; Frenzel et al., 2012; Lindback et al., 2012). This might reflect a different requirement for CodY in these genetic backgrounds.
In agreement with results reported in Bc (Frenzel et al., 2012; Lindback et al., 2012), we showed that three PlcR-dependent promoters were downregulated in Bt ΔcodY compared to the wild-type strain. Our study shows that, although the papR gene is less transcribed in Bt ΔcodY than in its wild-type parent, PapR-derived peptides were found in the supernatant of the mutant cells. However, in addition to the lower concentration in PapR derivatives, the sequences of the peptides detected were different between the mutant and the wild-type strain. It appeared that PapR was cleaved in Bt ΔcodY at sites that were not used in the wild-type strain. It has been reported that the PlcR-controlled NprB peptidase was, at least in part, responsible for the maturation of PapR (Pomerantsev et al., 2009). However, PapR27 was able to activate PlcR in ΔpapR cells, suggesting that other peptidases might be involved in the maturation process, at least before the positive autoregulatory loop is triggered and NprB is produced. nprB is among the most downregulated genes in the Bc ΔcodY mutant compared to the wild-type cells in the transcriptomic analysis reported by Lindback et al. (2012). It is then unlikely that it could fulfill its function in ΔcodY cells in a similar manner as in wild-type cells. We hypothesize that, in ΔcodY cells, NprB is rare and that this could facilitate the access of other peptidases to PapR, including peptidases that would usually be less abundant. We also noted that there was no concordance between the predicted cleavage site after the putative signal sequence and the peptides found in the supernatant of both strains. We did not detect peptides with D22 in the N-terminal position, as expected from the prediction. Aside from the ADLPFEF peptide, we only found peptides with A15 or F25 at their N-terminal end in the wild-type cell culture supernatant. This suggests that the signal peptide might have been improperly predicted and might be defined as the first 14 amino acids of the PapR sequence.
Expressing the mature form of PapR inside ΔcodY cells was enough to restore the production of PlcR-dependent proteins. We also showed that the concentration of PapR in a ΔcodY cell culture supernatant is sufficient to activate PlcR in ΔpapR cells. These results indicate an import deficiency in Bt ΔcodY compared to wild-type. Import of PapR has been previously investigated and it is known that it requires the pore components of the Opp permease, OppB-C-D-F (Gominet et al., 2001). However, no data was reported concerning the involvement of the OppA SBP. We showed that an oppA deletion mutant has a hemolytic phenotype similar to that of its wild-type parent on sheep blood agar plates, indicating that, even though we demonstrated that OppA was involved in the import of PapR, the cell produces other SBP with the same function. Bt carries 40 genes that encode proteins potentially involved in peptide import. Five loci encoding all the components of oligopeptide permeases were identified, including the one encoding OppA-B-C-D-F mentioned above. Aside from the latter, no function has been described for the other putative transporters. In addition to transporting peptides, members of the Opp family are also able to import nickel. Without functional assays, it is difficult to assign them a role. Interestingly, eight OppA-like proteins seem orphan, which is not common among bacteria. In order to understand the effect of the codY mutation on the production of the Opp permease and the other Opp-like proteins we undertook a proteomics approach and compared the protein contents of membrane-enriched fractions of ΔcodY cells to that of wild-type cells. Among the 1366 proteins detected in all the samples 19 are putative components of oligopeptide permeases. It was previously shown that CodY impacts the expression of genes encoding oligopeptide permeases in Lactococcus lactis (Guédon et al., 2001) and B. subtilis (Slack et al., 1995; Molle et al., 2003). In a Bc ΔcodY strain however, several Opp-like-encoding genes were downregulated whereas others were upregulated (Lindback et al., 2012).
Mass spectrometry analysis of membrane-enriched fractions allowed us to detect 19 out of the 40 Opp-like proteins predicted to be encoded in the genome of Bt. Some of these proteins were either upregulated in ΔcodY cells compared to wild-type or were not affected by the mutation. Ten of these proteins were downregulated in Bt ΔcodY compared to the wild-type strain, including the components of the Opp permease, except OppA in late exponential phase. Results of the overexpression of the genes encoding the OppA-like proteins with this profile in the Bt ΔcodY background showed that some proteins [09790, 12330 (OppA), 20960, 36620, 36650, and 36660] were able to partially restore PlcR-dependent phenotypes in the mutant. It is worth noting that several OppA-like proteins are potentially able to bind PapR and to interact with its dedicated permease in order to allow its import in the cell. Proteins 02170 and 12390, that were less detected in ΔcodY than in wt samples did not have any influence on PlcR activity, indicating that hemolysis restoration was not an artifact linked to the overexpression of any OppA-encoding gene. Remarkably, overexpression of gene 06720, whose abundance was higher in ΔcodY than in wt preparations, did not affect the hemolytic activity of ΔcodY cells.
Interestingly, and although the opp locus was reported as being transcribed as an operon (Gominet et al., 2001), there is a difference in the detection profile of OppA and the other components of the Opp permease. We are not able to explain this observation. We can, however, hypothesize that OppA is not present during exponential phase in wild-type cells to avoid the untimely entry of other peptides, such as Phrs, involved, for example, in triggering sporulation, and that the other OppA-like proteins that allow the import of PapR, might have a better affinity for the latter than for the Phrs. This would be in agreement with the need of functional SBPs during late vegetative growth to import and accumulate PapR in the bacterial cytoplasm. OppA is eventually produced and is able to import PapR, in order to reinforce the feedback loop.
In our study, we show that the positive effect that CodY exerts on the expression of PlcR-regulated genes depends on its effect on OppA-like proteins. However, we do not know if this effect is direct or not. It was recently shown that CodY indirectly activated the expression of dtpT (a putative oligopeptide permease in B. subtilis) via the direct repression of the gene encoding the repressor ScoC (Belitsky et al., 2015), also present in Bt. We cannot exclude that CodY influences the expression of opp genes indirectly in the same manner, or by activating another activator.
Involvement of CodY in the control of the import of PapR allows the PlcR-PapR QS system to integrate two types of external signals: the composition of the medium and the accumulation of the peptide. This mechanism adds another layer of complexity to the regulatory circuits governing QS in Bt. Indeed, it has been shown that CodY directly represses the transcription of nprR, a quorum sensor responsible for the ability of Bt to survive on insect cadavers after the bacteria have killed their host (Dubois et al., 2013). nprR is also under the positive control of PlcR. During stationary phase, Spo0A, the major regulator of sporulation, represses plcR transcription (Lereclus et al., 2000; Gominet et al., 2001). Our laboratory has shown that, in vivo, in the insect model of infection Galleria mellonella, a PlcR-dependent gene was activated during the virulent stage of the bacterium, then its expression decreased as the expression of an NprR-dependent gene increased, after the insect death (Dubois et al., 2012). The influence of CodY on the virulence of pathogens has been previously reported, as mentioned in the introduction, linking the virulent behavior of the bacterium to its metabolic state. Altogether, our data are in line with these reports and describe one of the mechanisms underlying these finely tuned regulatory circuits that control interconnected physiological stages to coordinate the behavior of the bacteria throughout growth.
Author Contributions
Conceived and designed the study: LS, DL; designed the experiments: LS, DL; performed the experiments: LS, CL, CH, AG; constructed a mutant strain: EH; analyzed the data: LS, DL, CH, AG; wrote the paper: LS.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Ludovic Bridoux for technical assistance. This work was supported by the INRA, the French National Institute for Agronomical research. The proteomics data were obtained at the PAPPSO facility (Micalis Institute).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2015.01501
References
Agaisse, H., Gominet, M., Okstad, O. A., Kolsto, A. B., and Lereclus, D. (1999). PlcR is a pleiotropic regulator of extracellular virulence factor gene expression in Bacillus thuringiensis. Mol. Microbiol. 32, 1043–1053. doi: 10.1046/j.1365-2958.1999.01419.x
Agaisse, H., and Lereclus, D. (1994). Structural and functional analysis of the promoter region involved in full expression of the cryIIIA toxin gene of Bacillus thuringiensis. Mol. Microbiol. 13, 97–107. doi: 10.1111/j.1365-2958.1994.tb00405.x
Arantes, O., and Lereclus, D. (1991). Construction of cloning vectors for Bacillus thuringiensis. Gene 108, 115–119. doi: 10.1016/0378-1119(91)90495-W
Arnaud, M., Chastanet, A., and Debarbouille, M. (2004). New vector for efficient allelic replacement in naturally nontransformable, low-GC-content, gram-positive bacteria. Appl. Environ. Microbiol. 70, 6887–6891. doi: 10.1128/AEM.70.11.6887-6891.2004
Belitsky, B. R., Barbieri, G., Albertini, A. M., Ferrari, E., Strauch, M. A., and Sonenshein, A. L. (2015). Interactive regulation by the Bacillus subtilis global regulators CodY and ScoC. Mol. Microbiol. 97, 698–716. doi: 10.1111/mmi.13056
Caymaris, S., Bootsma, H. J., Martin, B., Hermans, P. W., Prudhomme, M., and Claverys, J. P. (2010). The global nutritional regulator CodY is an essential protein in the human pathogen Streptococcus pneumoniae. Mol. Microbiol. 78, 344–360. doi: 10.1111/j.1365-2958.2010.07339.x
Chateau, A., Van Schaik, W., Six, A., Aucher, W., and Fouet, A. (2011). CodY regulation is required for full virulence and heme iron acquisition in Bacillus anthracis. FASEB J. 25, 4445–4456. doi: 10.1096/fj.11-188912
Cook, L. C., and Federle, M. J. (2014). Peptide pheromone signaling in Streptococcus and Enterococcus. FEMS Microbiol. Rev. 38, 473–492. doi: 10.1111/1574-6976.12046
Declerck, N., Bouillaut, L., Chaix, D., Rugani, N., Slamti, L., Hoh, F., et al. (2007). Structure of PlcR: insights into virulence regulation and evolution of quorum sensing in Gram-positive bacteria. Proc. Natl. Acad. Sci. U.S.A 104, 18490–18495. doi: 10.1073/pnas.0704501104
Dineen, S. S., Villapakkam, A. C., Nordman, J. T., and Sonenshein, A. L. (2007). Repression of Clostridium difficile toxin gene expression by CodY. Mol. Microbiol. 66, 206–219. doi: 10.1111/j.1365-2958.2007.05906.x
Dubois, T., Faegri, K., Perchat, S., Lemy, C., Buisson, C., Nielsen-Leroux, C., et al. (2012). Necrotrophism is a quorum-sensing-regulated lifestyle in Bacillus thuringiensis. PLoS Pathog 8:e1002629. doi: 10.1371/journal.ppat.1002629
Dubois, T., Perchat, S., Verplaetse, E., Gominet, M., Lemy, C., Aumont-Nicaise, M., et al. (2013). Activity of the Bacillus thuringiensis NprR-NprX cell-cell communication system is co-ordinated to the physiological stage through a complex transcriptional regulation. Mol. Microbiol. 88, 48–63. doi: 10.1111/mmi.12168
Dunny, G. M., and Leonard, B. A. (1997). Cell-cell communication in gram-positive bacteria. Annu. Rev. Microbiol. 51, 527–564. doi: 10.1146/annurev.micro.51.1.527
Frenzel, E., Doll, V., Pauthner, M., Lucking, G., Scherer, S., and Ehling-Schulz, M. (2012). CodY orchestrates the expression of virulence determinants in emetic Bacillus cereus by impacting key regulatory circuits. Mol. Microbiol. 85, 67–88. doi: 10.1111/j.1365-2958.2012.08090.x
Gohar, M., Faegri, K., Perchat, S., Ravnum, S., Okstad, O. A., Gominet, M., et al. (2008). The PlcR virulence regulon of Bacillus cereus. PLoS ONE 3:e2793. doi: 10.1371/journal.pone.0002793
Gohar, M., Okstad, O. A., Gilois, N., Sanchis, V., Kolsto, A. B., and Lereclus, D. (2002). Two-dimensional electrophoresis analysis of the extracellular proteome of Bacillus cereus reveals the importance of the PlcR regulon. Proteomics 2, 784–791. doi: 10.1002/1615-9861(200206)2:6
Gominet, M., Slamti, L., Gilois, N., Rose, M., and Lereclus, D. (2001). Oligopeptide permease is required for expression of the Bacillus thuringiensis plcR regulon and for virulence. Mol. Microbiol. 40, 963–975. doi: 10.1046/j.1365-2958.2001.02440.x
Grenha, R., Slamti, L., Nicaise, M., Refes, Y., Lereclus, D., and Nessler, S. (2013). Structural basis for the activation mechanism of the PlcR virulence regulator by the quorum-sensing signal peptide PapR. Proc. Natl. Acad. Sci. U.S.A. 110, 1047–1052. doi: 10.1073/pnas.1213770110
Guédon, E., Serror, P., Ehrlich, S. D., Renault, P., and Delorme, C. (2001). Pleiotropic transcriptional repressor CodY senses the intracellular pool of branched-chain amino acids in Lactococcus lactis. Mol. Microbiol. 40, 1227–1239. doi: 10.1046/j.1365-2958.2001.02470.x
Higgins, C. F. (1992). ABC transporters: from microorganisms to man. Annu. Rev. Cell Biol. 8, 67–113. doi: 10.1146/annurev.cb.08.110192.000435
Hiles, I. D., Gallagher, M. P., Jamieson, D. J., and Higgins, C. F. (1987). Molecular characterization of the oligopeptide permease of Salmonella typhimurium. J. Mol. Biol. 195, 125–142. doi: 10.1016/0022-2836(87)90332-9
Hiron, A. (2007). Les Transporteurs de Peptides de Staphylococcus aureus, Ph.D. thesis, Paris: AgroParisTech.
Hsueh, Y. H., Somers, E. B., and Wong, A. C. (2008). Characterization of the codY gene and its influence on biofilm formation in Bacillus cereus. Arch. Microbiol. 189, 557–568. doi: 10.1007/s00203-008-0348-8
Lazazzera, B. A., Solomon, J. M., and Grossman, A. D. (1997). An exported peptide functions intracellularly to contribute to cell density signaling in B. subtilis. Cell 89, 917–925. doi: 10.1016/S0092-8674(00)80277-9
Lereclus, D., Agaisse, H., Gominet, M., Salamitou, S., and Sanchis, V. (1996). Identification of a Bacillus thuringiensis gene that positively regulates transcription of the phosphatidylinositol-specific phospholipase C gene at the onset of the stationary phase. J. Bacteriol. 178, 2749–2756.
Lereclus, D., Agaisse, H., Grandvalet, C., Salamitou, S., and Gominet, M. (2000). Regulation of toxin and virulence gene transcription in Bacillus thuringiensis. Int. J. Med. Microbiol. 290, 295–299. doi: 10.1016/S1438-4221(00)80024-7
Lereclus, D., Arantes, O., Chaufaux, J., and Lecadet, M. (1989). Transformation and expression of a cloned delta-endotoxin gene in Bacillus thuringiensis. FEMS Microbiol. Lett. 51, 211–217. doi: 10.1111/j.1574-6968.1989.tb03448.x
Lereclus, D., Vallade, M., Chaufaux, J., Arantes, O., and Rambaud, S. (1992). Expansion of insecticidal host range of Bacillus thuringiensis by in vivo genetic recombination. Biotechnology 10, 418–421. doi: 10.1038/nbt0492-418
Lindback, T., Mols, M., Basset, C., Granum, P. E., Kuipers, O. P., and Kovacs, A. T. (2012). CodY, a pleiotropic regulator, influences multicellular behaviour and efficient production of virulence factors in Bacillus cereus. Environ. Microbiol. 14, 2233–2246. doi: 10.1111/j.1462-2920.2012.02766.x
MacNeil, D. J., Gewain, K. M., Ruby, C. L., Dezeny, G., Gibbons, P. H., and Macneil, T. (1992). Analysis of Streptomyces avermitilis genes required for avermectin biosynthesis utilizing a novel integration vector. Gene 111, 61–68. doi: 10.1016/0378-1119(92)90603-M
Majerczyk, C. D., Sadykov, M. R., Luong, T. T., Lee, C., Somerville, G. A., and Sonenshein, A. L. (2008). Staphylococcus aureus CodY negatively regulates virulence gene expression. J. Bacteriol. 190, 2257–2265. doi: 10.1128/JB.01545-07
Mignot, T., Mock, M., Robichon, D., Landier, A., Lereclus, D., and Fouet, A. (2001). The incompatibility between the PlcR- and AtxA-controlled regulons may have selected a nonsense mutation in Bacillus anthracis. Mol. Microbiol. 42, 1189–1198. doi: 10.1046/j.1365-2958.2001.02692.x
Miller, M. B., and Bassler, B. L. (2001). Quorum sensing in bacteria. Annu. Rev. Microbiol. 55, 165–199. doi: 10.1146/annurev.micro.55.1.165
Molle, V., Nakaura, Y., Shivers, R. P., Yamaguchi, H., Losick, R., Fujita, Y., et al. (2003). Additional targets of the Bacillus subtilis global regulator CodY identified by chromatin immunoprecipitation and genome-wide transcript analysis. J. Bacteriol. 185, 1911–1922. doi: 10.1128/JB.185.6.1911-1922.2003
Montgomery, C. P., Boyle-Vavra, S., Roux, A., Ebine, K., Sonenshein, A. L., and Daum, R. S. (2012). CodY deletion enhances in vivo virulence of community-associated methicillin-resistant Staphylococcus aureus clone USA300. Infect. Immun. 80, 2382–2389. doi: 10.1128/IAI.06172-11
Perchat, S., Dubois, T., Zouhir, S., Gominet, M., Poncet, S., Lemy, C., et al. (2011). A cell-cell communication system regulates protease production during sporulation in bacteria of the Bacillus cereus group. Mol. Microbiol. 82, 619–633. doi: 10.1111/j.1365-2958.2011.07839.x
Perego, M. (2013). Forty years in the making: understanding the molecular mechanism of peptide regulation in bacterial development. PLoS Biol. 11:e1001516. doi: 10.1371/journal.pbio.1001516
Perego, M., and Hoch, J. A. (1996). Cell-cell communication regulates the effects of protein aspartate phosphatases on the phosphorelay controlling development in Bacillus subtilis. Proc. Natl. Acad. Sci. U.S.A. 93, 1549–1553. doi: 10.1073/pnas.93.4.1549
Pomerantsev, A. P., Pomerantseva, O. M., Camp, A. S., Mukkamala, R., Goldman, S., and Leppla, S. H. (2009). PapR peptide maturation: role of the NprB protease in Bacillus cereus 569 PlcR/PapR global gene regulation. FEMS Immunol. Med. Microbiol. 55, 361–377. doi: 10.1111/j.1574-695X.2008.00521.x
Raymond, B., Johnston, P. R., Nielsen-Leroux, C., Lereclus, D., and Crickmore, N. (2010). Bacillus thuringiensis: an impotent pathogenΔ Trends Microbiol. 18, 189–194. doi: 10.1016/j.tim.2010.02.006
Richardson, A. R., Somerville, G. A., and Sonenshein, A. L. (2015). Regulating the intersection of metabolism and pathogenesis in gram-positive bacteria. Microbiol. Spectr. 3:MBP-0004-2014. doi: 10.1128/microbiolspec.MBP-0004-2014
Rivera, F. E., Miller, H. K., Kolar, S. L., Stevens, S. M. Jr., and Shaw, L. N. (2012). The impact of CodY on virulence determinant production in community-associated methicillin-resistant Staphylococcus aureus. Proteomics 12, 263–268. doi: 10.1002/pmic.201100298
Roux, A., Todd, D. A., Velazquez, J. V., Cech, N. B., and Sonenshein, A. L. (2014). CodY-mediated regulation of the Staphylococcus aureus Agr system integrates nutritional and population density signals. J. Bacteriol. 196, 1184–1196. doi: 10.1128/JB.00128-13
Salamitou, S., Ramisse, F., Brehelin, M., Bourguet, D., Gilois, N., Gominet, M., et al. (2000). The PlcR regulon is involved in the opportunistic properties of Bacillus thuringiensis and Bacillus cereus in mice and insects. Microbiology 146(Pt 11), 2825–2832. doi: 10.1099/00221287-146-11-2825
Slack, F. J., Serror, P., Joyce, E., and Sonenshein, A. L. (1995). A gene required for nutritional repression of the Bacillus subtilis dipeptide permease operon. Mol. Microbiol. 15, 689–702. doi: 10.1111/j.1365-2958.1995.tb02378.x
Slamti, L., and Lereclus, D. (2002). A cell-cell signaling peptide activates the PlcR virulence regulon in bacteria of the Bacillus cereus group. EMBO J. 21, 4550–4559. doi: 10.1093/emboj/cdf450
Slamti, L., Perchat, S., Gominet, M., Vilas-Boas, G., Fouet, A., Mock, M., et al. (2004). Distinct mutations in PlcR explain why some strains of the Bacillus cereus group are nonhemolytic. J. Bacteriol. 186, 3531–3538. doi: 10.1128/JB.186.11.3531-3538.2004
Slamti, L., Perchat, S., Huillet, E., and Lereclus, D. (2014). Quorum Sensing in Bacillus thuringiensis is required for completion of a full infectious cycle in the insect. Toxins (Basel) 6, 2239–2255. doi: 10.3390/toxins6082239
Sonenshein, A. L. (2005). CodY, a global regulator of stationary phase and virulence in Gram-positive bacteria. Curr. Opin. Microbiol. 8, 203–207. doi: 10.1016/j.mib.2005.01.001
Sonenshein, A. L. (2007). Control of key metabolic intersections in Bacillus subtilis. Nat. Rev. Microbiol. 5, 917–927. doi: 10.1038/nrmicro1772
Stammen, S., Muller, B. K., Korneli, C., Biedendieck, R., Gamer, M., Franco-Lara, E., et al. (2010). High-yield intra- and extracellular protein production using Bacillus megaterium. Appl. Environ. Microbiol. 76, 4037–4046. doi: 10.1128/AEM.00431-10
Stenz, L., Francois, P., Whiteson, K., Wolz, C., Linder, P., and Schrenzel, J. (2011). The CodY pleiotropic repressor controls virulence in gram-positive pathogens. FEMS Immunol. Med. Microbiol. 62, 123–139. doi: 10.1111/j.1574-695X.2011.00812.x
Suzuki, A., Mori, M., Sakagami, Y., Isogai, A., Fujino, M., Kitada, C., et al. (1984). Isolation and structure of bacterial sex pheromone, cPD1. Science 226, 849–850. doi: 10.1126/science.6436978
Taniguchi, L., De Fatima Faria, B., Rosa, R. T., De Paula, E. C. A., Gursky, L. C., Elifio-Esposito, S. L., et al. (2009). Proposal of a low-cost protocol for colorimetric semi-quantification of secretory phospholipase by Candida albicans grown in planktonic and biofilm phases. J. Microbiol. Methods 78, 171–174. doi: 10.1016/j.mimet.2009.05.012
Taylor, R. G., Walker, D. C., and Mcinnes, R. R. (1993). E. coli host strains significantly affect the quality of small scale plasmid DNA preparations used for sequencing. Nucleic Acids Res. 21, 1677–1678. doi: 10.1093/nar/21.7.1677
van Schaik, W., Chateau, A., Dillies, M. A., Coppee, J. Y., Sonenshein, A. L., and Fouet, A. (2009). The global regulator CodY regulates toxin gene expression in Bacillus anthracis and is required for full virulence. Infect. Immun. 77, 4437–4445. doi: 10.1128/IAI.00716-09
Villafane, R., Bechhofer, D. H., Narayanan, C. S., and Dubnau, D. (1987). Replication control genes of plasmid pE194. J. Bacteriol. 169, 4822–4829.
Vizcaíno, J. A, Côté, R. G., Csordas, A., Dianes, J. A., Fabregat, A., Foster, J. M., et al. (2013). The PRoteomics IDEntifications (PRIDE) database and associated tools: status in 2013. Nucleic Acids Res. 41, D1063–D1069. doi: 10.1093/nar/gks1262
Yu, N. Y., Wagner, J. R., Laird, M. R., Melli, G., Rey, S., Lo, R., et al. (2010). PSORTb 3.0: improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 26, 1608–1615. doi: 10.1093/bioinformatics/btq249
Keywords: quorum-sensing, regulation, signaling peptide, Opp, virulence regulon, mass spectrometry, shot gun
Citation: Slamti L, Lemy C, Henry C, Guillot A, Huillet E and Lereclus D (2016) CodY Regulates the Activity of the Virulence Quorum Sensor PlcR by Controlling the Import of the Signaling Peptide PapR in Bacillus thuringiensis. Front. Microbiol. 6:1501. doi: 10.3389/fmicb.2015.01501
Received: 27 August 2015; Accepted: 14 December 2015;
Published: 06 January 2016.
Edited by:
Peter Mullany, University College London, UKReviewed by:
Chien-Yi Chang, Newcastle University, UKAbraham L. Sonenshein, Tufts University, USA
Beth Lazazzera, University of California, Los Angeles, USA
Copyright © 2016 Slamti, Lemy, Henry, Guillot, Huillet and Lereclus. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Leyla Slamti, bGV5bGEuc2xhbXRpQGpvdXkuaW5yYS5mcg==