 Rani Soni
Rani Soni Drista Sharma
Drista Sharma Tarun K. Bhatt
Tarun K. Bhatt- Department of Biotechnology, School of Life Sciences, Central University of Rajasthan, Rajasthan, India
Plasmodium falciparum is the causative agent of deadly malaria disease. It is an intracellular eukaryote and completes its multi-stage life cycle spanning the two hosts viz, mosquito and human. In order to habituate within host environment, parasite conform several strategies to evade host immune responses such as surface antigen polymorphism or modulation of host immune system and it is mediated by secretion of proteins from parasite to the host erythrocyte and beyond, collectively known as, malaria secretome. In this review, we will discuss about the deployment of parasitic secretory protein in mechanism implicated for immune evasion, protein trafficking, providing virulence, changing permeability and cyto-adherence of infected erythrocyte. We will be covering the possibilities of developing malaria secretome as a drug/vaccine target. This gathered information will be worthwhile in depicting a well-organized picture for host-pathogen interplay during the malaria infection and may also provide some clues for the development of novel anti-malarial therapies.
Introduction
World Health Organization report summarized that about 198 million cases and 0.58 million deaths occurred in year 2013 (World Health Organization [WHO], 2014). Amongst different species of Plasmodium, P. falciparum is the most dangerous and responsible for severe complexities during infection like multi-organ failure, cerebral malaria, coma, and death (Miller et al., 1994; Mendis and Carter, 1995). P. falciparum completes its life cycle spanning two alternate host, human, and mosquito. Within the human host, parasite undergoes series of developmental stages in the liver and erythrocytes (RBCs). The intra-erythrocytic cycle is found to be important as it is responsible for patho-physiology of the disease (Miller et al., 2002). Within the erythrocyte, the parasite proceeds through the different morphological stages such as ring, trophozoite, and schizont (Bannister et al., 2000; Florens et al., 2002). After completion of infection cycle, erythrocyte gets ruptured, and merozoites are released into the host bloodstream. The released merozoites initiate next round of erythrocytic cycle by infecting fresh erythrocytes (Gilson and Crabb, 2009). The survival of parasite inside the host cell is difficult, ascribed to which Plasmodium adapts various strategies to avoid the host immune response (Miller et al., 1994; Hisaeda et al., 2005). The strategies encompass the secretion of hitherto of proteins against infected RBC (iRBC) surface and beyond it into the host plasma (Singh et al., 2009). Infected erythrocyte surface remodeling is an example of such phenomena, involving the insertion of secreted proteins into the iRBC membrane (Deitsch and Wellems, 1996; Parker et al., 2004). Remodeling assists in cyto-adherence of iRBCs to the endothelial lining of blood vessels and thus averting them from splenic clearance (Newbold et al., 1999; Cooke et al., 2001; Rowe et al., 2009). Host immune modulation is another phenomenon involving the release of secretory proteins before or along with rupture of iRBCs. The released proteins interact with the components of the host immune system to provide conducive environment for merozoites before they invade new erythrocytes (Singh et al., 2009). The entire set of secreted proteins is known as ‘Secretome.’ These proteins are implicated in the processes essential for parasite survival such as host–parasite interactions and immune modulation (Ranganathan and Garg, 2009). Hence, in this review we will be focusing on parasite secretome and its significance in the malaria biology.
Identification of Malaria Parasite’s Secretome
The export of secretory proteins from various species including P. falciparum has been thoroughly studied (van Ooij et al., 2008). Transport of secretory proteins follows a complicated route due to the presence of three membranes of parasite and vacuole inside the host cell (Martin et al., 2009). There must be a defined mechanism or recognizable sequence motifs behind such complex transport. Advent of bioinformatics tools facilitates the prediction of such conserved signal sequences responsible for the export and localization of the secretory proteins (Hiller et al., 2004; Marti et al., 2004, 2005).
The identification of sequence motifs necessary for export of parasite proteins is required for unearthing the complete secretome of the parasite. The first report proposed the presence of host targeting signal (HT motif) (Hiller et al., 2004) or Plasmodium export element (PEXEL motif) (Marti et al., 2004) in the sequence that is a requisite for the export of secretory proteins from parasitophorous vacuole (PV). The HT/PEXEL motif, present in more than 400 parasitic proteins, comprises short amino-terminal sequence, ‘R/KxLxE/Q’. The role of motif in the export of both soluble and surface-associated protein is determined by green fluorescent protein (GFP) and yellow fluorescent protein (YFP) assays involving the fusion of secretory proteins such as Knob associated histidine rich protein (KAHRP), PfEMP-1 (with PEXEL like motif), Glycophorin binding protein (GBP130), and members of repetitive interspersed family (rifin). Mutation or truncation of the PEXEL motif interrupted the transport of proteins and rendered their accumulation in PV itself (Marti et al., 2004; van Ooij et al., 2008). Prior to the secretion of PEXEL containing proteins to destination, N-terminus is processed in endoplasmic reticulum (ER) by protease enzyme plasmepsin V (PMV), followed by N-acetylation of the cleaved product (Chang et al., 2008; Boddey et al., 2010; Russo et al., 2010). The decisive role of PMV in export of proteins is also demonstrated by identification of transition state (TS) inhibitor, WEHI-916 (Walter and Eliza Hall Institute of Medical Research). The WEHI-916 inhibitor is found to compete with the PEXEL containing substrate resulting in blocking of activity of PMV and ultimately lead to the cessation of parasite growth at the trophozoite stage. Knockdown studies of PMV further supported the inhibitory role on PMV. Indirect hindrance of export of PfEMP-1 and the loss of virulence and cytoadherence of iRBC has been observed due to inhibitory activity of WEHI-916 (Sleebs et al., 2014a,b). Recently, another inhibitor of PMV, WEHI-842 has been identified. The inhibitory role of WEHI-842 is assessed through the immunoblotting of GFP tagged PEXEL containing PfEMP-3. It is found to be more effective in comparison to WEHI-916 (Hodder et al., 2015). The presence of PEXEL motif leads to identification of first set of parasite secretome. However, HT/PEXEL is found missing in various secretory proteins (Lingelbach and Przyborski, 2006). Analysis of such sequences showed the presence of a hydrophobic stretch in the internal region of trans-membrane proteins which helps in crossing the Parasitophorous Vacuolar Membrane (PVM). Immune localization experiments and GFP-tagged approach investigated that first 20 amino acids of N-terminus along with hydrophobic residues in trans-membrane domain are common features of all PEXEL negative export proteins (PNEPs). Thus PNEPs have further expanded the secretome repertoire of the parasite (Spielmann et al., 2006; Spielmann and Gilberger, 2010; Heiber et al., 2013).
Apart from sequence-motifs based approach, other approaches are used to predict secretory proteins of Plasmodium. A Position-Specific Scoring Matrix (PSSM) profile based method is adapted that employs phylogenetic relationship derived through PSI–Blast against the non-redundant database. Based on these data, web server called ‘Plasmodium Secretory and Infected erythrocyte Associated Protein prediction’ (PSEApred) is developed to predict the secretory nature of plasmodium proteins (Verma et al., 2008). Similarly, the presence of N-myristoylation site, a cysteine S-palmitoylation site and some basic residues at N-terminus of parasite proteins are found to be responsible for the targeting to PVM and beyond (Gunaratne et al., 2000; Ma et al., 2012; Thavayogarajah et al., 2015; Wetzel et al., 2015). Together, both classical and non-classical path of secretion of proteins from parasite to the host cell have enhanced the secretome of the parasite. However, there is a possibility of appending more proteins to the growing secretome of parasite by identifying new signatures and patterns of secretion.
Trafficking Pathway of Secretome
The protein containing PEXEL motif moves from ER after cleavage by PMV to the PV either in the form of vesicular cargo (Barnwell, 1989) or through the secretory apparatus (Hinterberg et al., 1994; Taraschi et al., 2003). Proteins that are not cleaved by PMV have been shown to bind with phosphatidylinositol 3- phosphate (PI3-P) in the ER and proceed for further trafficking pathway (Bhattacharjee et al., 2012). The PNEP proteins require transmembrane domain for their transport (Heiber et al., 2013). All the proteins in the PV are found to be exported through Plasmodium Translocon of Exported protein (PTEX) complex (de Koning-Ward et al., 2009; Beck et al., 2014; Elsworth et al., 2014). These proteins undergo unfolding for their translocation across PV (Charpian and Przyborski, 2008; Gehde et al., 2009; Gruring et al., 2012). P. falciparum has developed a membranous structure in the cytoplasm of the host cell called ‘Maurer’s cleft’. It is a secondary organelle and required for the export of the proteins involved in virulence, modification of host cell environment (Trager et al., 1966; Rudzinska and Trager, 1968) and for trafficking of membrane localized proteins (Przyborski et al., 2003; Lanzer et al., 2006; Mundwiler-Pachlatko and Beck, 2013). The exported proteins from PV are found to reside in the Maurer’s cleft (Haldar et al., 2002). Knock-down studies of Maurer’s cleft residing proteins like Membrane-Associated Histidine-Rich Protein (MAHRP1) and Skeletal binding protein-1 (SBP-1) proved its vitality in protein sorting (Epp and Deitsch, 2006; Maier et al., 2007; Spycher et al., 2008). In addition, most of the known secretory proteins including three antigenic families of parasite proteins (Stevor, Rifin, and Var) are localized in the Maurer’s cleft (Cheng et al., 1998) via PTEX export system (de Koning-Ward et al., 2009). PTEX export system, found exclusively in the genus Plasmodium, is responsible for the translocation of proteins targeted beyond the vacuolar membrane of the parasite (de Koning-Ward et al., 2009; Desai and Miller, 2014). It is a complex of five proteins including PTEX150, Heat shock protein 101 (HSP101), exported protein 2 (EXP2), PTEX88, and thioredoxin 2 (TRX2). The passage for directing proteins toward the cytosol of host erythrocyte is formed by EXP2 (de Koning-Ward et al., 2009). TRX 2 is found to be involved in unfolding of proteins destined to pass through the PTEX. Inhibition of HSP101 leads to the obstruction in protein export and eventually the accumulation of proteins such as Ring Infected Erythrocyte Surface Antigen (RESA), Ring Exported Protein 3 (REX3), Histidine Rich Protein-1 (HRP1), and KAHRP in parasitic compartment (Beck et al., 2014). It is observed that deletion of PTEX components prevent proteins from crossing PVM, resulting in interference of parasitic growth at the ring and trophozoite stage (Elsworth et al., 2014). However, mode of recognition between proteins to be exported and those to be retained by the PTEX complex still remains unclear. The mechanism of unfolding during protein export is also not defined and therefore it opens a new window of opportunity for scientists to explore and explain the facts related to PTEX system. In addition, the presence of this export system exclusively in Plasmodium genus makes it a captivating drug target (de Koning-Ward et al., 2009).
Host Cell Remodeling
In order to make opportune environment within host, parasite makes substantial modifications in the host erythrocytes (Haldar and Mohandas, 2007). The modifications are predominantly mediated by secretion of parasite proteins across the PVM (Charpian and Przyborski, 2008; Maier et al., 2009; Goldberg and Cowman, 2010; Marti and Spielmann, 2013; Elsworth et al., 2014). The process of erythrocyte remodeling includes.
Cytoadherence
To circumvent immune clearance in spleen, infected erythrocytes get adhered to endothelial wall, which is mediated through various cell adhesion molecules like ICAMs, CD36 on blood vessels (Gardner et al., 1996; Ho and White, 1999; Bhalla et al., 2015). Some events during adhesion process such as rosette formation with fresh erythrocyte (Udomsangpetch et al., 1989), auto-agglutination due to clumping of iRBCs and platelets (Pain et al., 2001) leads to severe disease pathologies (Rowe et al., 1995, 2002; Newbold et al., 1999). P. falciparum erythrocytic membrane protein-1 (PfEMP-1) is major virulent factor present on surface of erythrocyte (Magowan et al., 1988; Chen et al., 1998). A study regarding transgenic lines of P. falciparum with altered PfEMP-1 expression shows strong immune response targeted against PfEMP-1 (Chan et al., 2012). Alteration in functioning of B-cells during parasite infection comprehends the interaction between cysteine-rich inter-domain region 1α (CIDRα) of PfEMP-1 and B-cells. The complex formed causes the activation of NF-kB pathway resulting in functional impairment of B-cells (Simone et al., 2011). Multiple PfEMP-1 proteins of P. falciparum bind to Fc portion of IgM (Jeppesen et al., 2015; Stevenson et al., 2015a) and found to be involved in rosette formation (Stevenson et al., 2015a,b). In addition to PfEMP-1, sub-telomeric variant open reading frame (STEVOR) and RIFIN members also play decisive role in rosette formation (Cheng et al., 1998; Kyes et al., 1999; Niang et al., 2014). The antigenic variation of proteins allows the parasite to escape host immune response (Bull et al., 1998). A protruding structure on the surface of erythrocyte namely ‘knob’ is found to be essential in adhesion process of iRBCs (Crabb et al., 1997). Some proteins localized to knob interact with surface proteins of erythrocytes. Interactions include binding of KAHRP with ankyrin R and pro-coagulant glucosaminoglycans (Waller et al., 1999; Wickham et al., 2001; Rug et al., 2006; Weng et al., 2014) and binding of Plasmodium helical interspersed sub-telomeric domain (PHIST) to PfEMP-1 (Oberli et al., 2014). In case of cerebral malaria, Pf14_075, member of PHIST family is found to be highly up-regulated and binds to human brain endothelial cell line (HBEC-5i). The study indicates its mantle in cyto-adherence (Claessens et al., 2012). Proteins such as erythrocyte membrane protein 3 (PfEMP3), Mature parasite-infected Erythrocyte Surface Antigen (MESA; Lustigman et al., 1990), RESA, PfEMP-1 (Sharma, 1997; Horrocks et al., 2005), KAHRP (Rug et al., 2006) and PfEMP3 (Knuepfer et al., 2005) are involved in knob formation. Merozoites Surface Protein-1 (MSP-1), another knob protein, shows interaction with RBC surface proteins like Band 3 and Glycophorin A (GPA). A study of mouse model deficient in GPA-Band3 complex described the role of knob formation in cyto-adherence. (Goel et al., 2003; Baldwin et al., 2015).
Membrane Permeability
Secretory proteins make astonishing alterations in the permeability of iRBCs membrane for ions and nutrient exchange. (Homewood and Neame, 1974; Ginsburg et al., 1983; Kutner et al., 1983). The presence of ion channels such as Plasmodium Surface Anion Channel (PSAC) is responsible for induction of drug resistance. The identification of structural composition of this complex would contribute to better understanding of pathogenic interaction and drug resistance mechanism and therefore suggested for therapeutic intervention (Lisk et al., 2008; Desai, 2012). Cytoadherence-linked antigen3 (Clag3) protein, found on the host membrane is appraised to be associated with PSAC in ion and nutrient transport through channels (Nguitragool et al., 2011; Pillai et al., 2012; Sharma et al., 2015). Secretory proteins involved in regulation of net flux of Na+, K+, and other ions are on the focus (Kirk, 2015). For instance, P-type ATP4 (PfATP4), regulating the transport of Na + ions is contemplated as a potential drug target (Spillman et al., 2013). Membrane permeabilization is found to be a necessary event for egress of parasites from iRBCs. Cysteine proteases have been shown to play cardinal role in rupture of erythrocyte membrane for the release of parasite (Hadley et al., 1983; Dluzewski et al., 1986; McKerrow et al., 1993; Raphael et al., 2000; Lee and Fidock, 2008). One of the members of this class, falcipian 2 is responsible for the cleavage of ankyrin and protein 4.1 of erythrocytic cytoskeleton (Dua et al., 2001). It has been evident through the gene disruption studies that expression of Plasmodium perforin like protein 2 (PPLP2) is paramount for membrane permeabilization during the gametocyte release from infected erythrocytes. It had been illustrated that gametocytes are unable to release from PPLP2 (-) lines of parasite, thereby reducing the transfer of gametocyte to vector (Wirth et al., 2014). MSP-1 has also been demonstrated to interact with host cytoskeleton spectrin causing the membrane destabilization and thereby enabling the release of merozoites from iRBC (Das et al., 2015).
Membrane Rigidity
Apart from cytoadherence, membrane rigidness or loss of deformability is also responsible for the sequestration of iRBCs (Bull et al., 1998). It has been clarified that knobs are liable for causing stiffness and hardening of the iRBCs (Zhang et al., 2015). Deformability of parasitized RBC is reduced due to association of RESA with spectrin (Mills et al., 2007). Pf332 exported on the membrane is directly involved in membrane rigidity and adhesion (Glenister et al., 2009). The KAHRP along with the membrane skeleton imparts rigidity to infected cell and will eventually obstruct blood flow (Waller et al., 1999; Pei et al., 2005). PHIST protein increases membrane rigidity by binding to membrane skeleton (Parish et al., 2013). Thus, it can be surmised that proteins responsible for rigidity are directly linked to virulence, providing an evidence for secretome in establishment of infection.
Secretory Protein Exported Beyond the Erythrocyte
Most of the data reported with respect to secretome is related to proteins secreted into the erythrocytes cytosol or membrane. Interestingly, some proteins, which are not restricted to iRBCs membrane rather squeeze out from iRBCs membrane and get secreted out. First experimental evidence (Singh et al., 2009) identified secretion of 27 novel proteins beyond the erythrocyte membrane before it gets ruptured. Immune localization and immune electron microscopic studies confirmed the secretion of proteins beyond iRBC (Singh et al., 2009). Some of them are functionally characterized. The protein containing Sel-1 functional domain is found to be involved in regulating ‘Notch signaling pathway’ which in turn has been hypothesized to influence the T cell differentiation (Grant and Greenwald, 1996; Singh et al., 2009). In most protozoan parasites, to evade host immune response, common mechanism includes altered T-helper cell differentiation (Zambrano-Villa et al., 2002; Rodrigues et al., 2014). Some proteins, closely associated with highly polymorphic genes, contribute to antigenic determinants of parasite (Singh et al., 2009). Secretory protein with LCCL (Limulus clotting factor C) domain, conserved across apicomplexan parasite, assumed to have role in immune evasion mechanism, (Claudianos et al., 2002; Dessens et al., 2004), defense mechanism and shows binding with lipid A of lipopolysaccharides. CRISPLD2 (Cysteine-Rich Secretory Protein LCCL Domain containing 2), an example of LCCL domain containing protein, has an anti-inflammatory function and is related to disease pathology (Vásárhelyi et al., 2014).
Sequence similarity studies suggested that some proteins viz, virulent immuno-reactive protein (specific to bacteria and virus), PFB0765w (uncharacterized protein), rhoptry neck protein (RON4), moving junction protein and MAL13P1.39 (uncharacterized protein) are involved in modulation of host immune response (Singh et al., 2009). Domain analysis demonstrated the presence of extracellular domain responsible for the interaction with other proteins, speculative of being involved in host–parasite interactions. [Table 1: Domains identified by CDD (Conserved Domain Database), NCBI]. It is depicted through flowcytometery and confocal microscopy that translationally controlled tumor protein (TCTP) analog released by Plasmodium in host serum is responsible for release of histamine and IL-8 from basophils and eosinophils, respectively, (MacDonald et al., 2001) and reduction in B-cell immune response. In another study, a canonical tyrosyl-tRNA synthetase (PfTyrRS) from Plasmodium is evidenced to be secreted out from the iRBCs and involved in non-canonical function of immune cell binding and modulation (Bhatt et al., 2011). Likewise in other intracellular pathogens such as Mycobacterium tuberculosis immune modulation ability is found in secretory proteins (Giacomini et al., 2001). During infection, secretome is also charged for causing alteration in functioning of antigen-presenting cells and dendritic cells (Sacks and Sher, 2002; Langhorne et al., 2004; Millington et al., 2006; Sponaas et al., 2006; Teirlinck et al., 2015). The presence of proteins on the surface or in secretion implicate their role in host–parasite interactions and probably in immune modulation for better survival of parasite and it would be fascinating to have information related to the ‘Interactome’ of the secretory proteins. In-silico knock-out studies and graphical analysis of protein–protein interaction network (PPIN) explored newer approach in order to identify the interacting partners vital to parasite during host–parasite interaction (Bhattacharyya and Chakrabarti, 2015). Nevertheless, some more studies are required to understand the role of secretory proteins in regulating host pathways. The role of secretory proteins of the parasite could have larger impact on malaria biology. Besides available knowledge, there is a need to identify signature motif or pattern responsible for secretion of proteins outside iRBCs. The identification of marker responsible for the localization of proteins to infected erythrocyte membrane and their export will be highly beneficial in interaction studies. There is a requirement of classifying secretome in terms of cellular localization and expression during developmental stages of parasite in order to understand its role in better way. It would be interesting to explore trafficking pathway of protein exported beyond the iRBCs membrane.
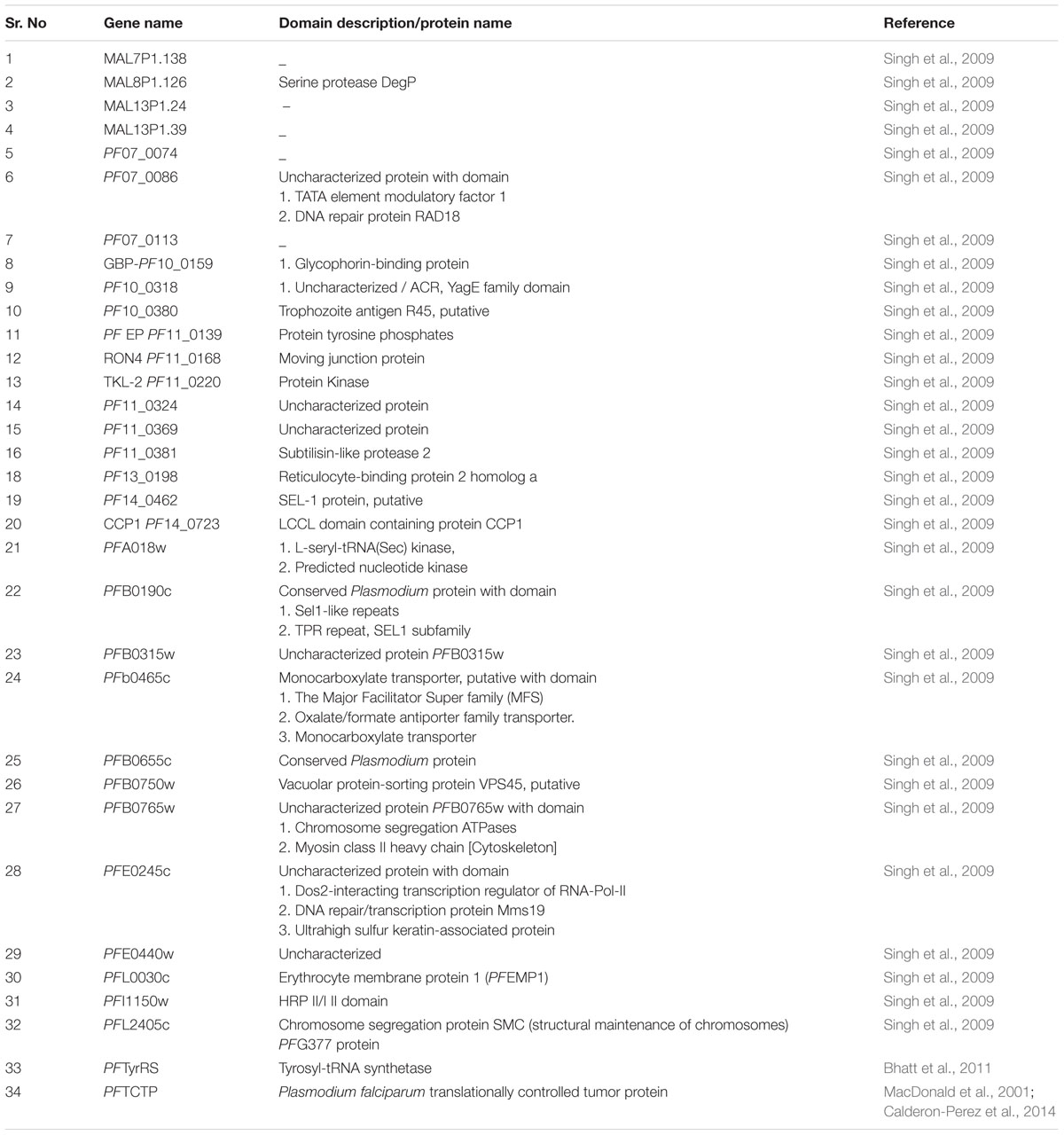
TABLE 1. Some secretory protein exported out from the iRBC.
Secretome as Potential Drug/Vaccine Target
The intracellular parasite adapts different strategies for protein export in order to survive in host environment. As the secretome is intimately associated with disease pathology and parasite survival is reliant on them, any interference in the secretory pathway or inhibition of secretory proteins itself would jeopardize the parasite. In addition, utilization of information of secretome available shall provide clues to certain strategies involved in host–parasite interaction at molecular level (Ranganathan and Garg, 2009). The function of these proteins can be annotated by comparing with homologous protein of known function in other organisms. Homology modeling of secretory proteins could also provide a starting point for the lead identification in the process of drug development. Vaccine and drug development against the secretory protein is in progress in various other pathogens like H. pylori (Lower et al., 2008), Helminths parasite (Hewitson et al., 2009) etc. Till date, various parasite proteins involved in secretory pathway have been characterized and may be critical in anti-malarial drug targeting such as inhibition of PTEX complex. Another important drug target capturing the interest in context of drug development area is PMV (plasmepsin V). Indispensability of PMV in virulence, cytoadherence, and parasitic growth makes PMV an attractive anti-malarial drug target (Sleebs et al., 2014a,b). Structural determination of PMV–WEHI 842 inhibitor complex provides an insight for interaction between active site residue and inhibitor. This study paves the way for developing potent anti-malarial by blocking export machinery of parasite (Hodder et al., 2015).
Taken together, the functional characterization of secretory proteins and proteins involved in their export, implicated in knob formation, involved in trafficking pathway, or those involved in host pathogenic interaction and invasion of host immune system, are all indispensible for parasite survival or pathogenicity. Therefore, complete investigation and characterization of secretome may provide us better understanding to get effective therapies for malaria disease. Allelic replacement and GFP tagging revealed the importance of PMV in protein export and parasite survival thus making it an attractive target for anti malarial drugs.
Conclusion
Export of secretory parasite proteins into host cytoplasm will lead to apprehension of host cell functions required for parasite growth and survival by modulating crucial phenomena of malaria biology such as immune evasion and virulence. Deep understanding and investigation of role played by malaria secretome will be not only beneficial in deciphering host–pathogen interactions but it may also lead to better therapeutic intervention for malaria disease.
Author Contributions
RS and DS provided data and TB wrote the manuscript.
Funding
Authors thank Department of Science and Technology, SERB, Government of India for providing financial assistance (SB/FT/LS-191/2012) to the lab. RS is CSIR-UGC Junior Research Fellow.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Baldwin, M. R., Li, X., Hanada, T., Liu, S.-C., and Chishti, A. H. (2015). Merozoite surface protein 1 recognition of host glycophorin A mediates malaria parasite invasion of red blood cells. Blood 125, 2704–2711. doi: 10.1182/blood-2014-11-611707
Bannister, L. H., Hopkins, J. M., Fowler, R. E., Krishna, S., and Mitchell, G. H. (2000). A Brief illustrated guide to the ultrastructure of Plasmodium falciparum asexual blood stages. Parasitol. Today 16, 427–433. doi: 10.1016/S0169-4758(00)01755-5
Barnwell, J. (1989). Vesicle-mediated transport of membrane and proteins in malaria-infected erythrocytes. Blood Cells 16, 379–395.
Beck, J. R., Muralidharan, V., Oksman, A., and Goldberg, D. E. (2014). PTEX component HSP101 mediates export of diverse malaria effectors into host erythrocytes. Nature 511, 592–595. doi: 10.1038/nature13574
Bhalla, K., Chugh, M., Mehrotra, S., Rathore, S., Tousif, S., Prakash Dwivedi, V., et al. (2015). Host ICAMs play a role in cell invasion by Mycobacterium tuberculosis and Plasmodium falciparum. Nat. Commun. 6:6049. doi: 10.1038/ncomms7049
Bhatt, T. K., Khan, S., Dwivedi, V. P., Banday, M. M., Sharma, A., Chandele, A., et al. (2011). Malaria parasite tyrosyl-tRNA synthetase secretion triggers pro-inflammatory responses. Nat. Commun. 2:530. doi: 10.1038/ncomms1522
Bhattacharjee, S., Stahelin, R. V., Speicher, K. D., Speicher, D. W., and Haldar, K. (2012). Endoplasmic reticulum PI(3)P lipid binding targets malaria proteins to the host cell. Cell 148, 201–212. doi: 10.1016/j.cell.2011.10.051
Bhattacharyya, M., and Chakrabarti, S. (2015). Identification of important interacting proteins (IIPs) in Plasmodium falciparum using large-scale interaction network analysis and in-silico knock-out studies. Malar. J. 14:70. doi: 10.1186/s12936-015-0562-1
Boddey, J. A., Hodder, A. N., Gunther, S., Gilson, P. R., Patsiouras, H., Kapp, E. A., et al. (2010). An aspartyl protease directs malaria effector proteins to the host cell. Nature 463, 627–631. doi: 10.1038/nature08728
Bull, P. C., Lowe, B. S., Kortok, M., Molyneux, C. S., Newbold, C. I., and Marsh, K. (1998). Parasite antigens on the infected red cell surface are targets for naturally acquired immunity to malaria. Nat. Med. 4, 358–360. doi: 10.1038/nm0398-358
Calderon-Perez, B., Xoconostle-Cazares, B., Lira-Carmona, R., Hernandez-Rivas, R., Ortega-Lopez, J., and Ruiz-Medrano, R. (2014). The Plasmodium falciparum translationally controlled tumor protein (TCTP) is incorporated more efficiently into B cells than its human homologue. PLoS ONE 9:e85514. doi: 10.1371/journal.pone.0085514
Chan, J.-A., Howell, K. B., Reiling, L., Ataide, R., Mackintosh, C. L., Fowkes, F. J. I., et al. (2012). Targets of antibodies against Plasmodium falciparum–infected erythrocytes in malaria immunity. J. Clin. Invest. 122, 3227–3238. doi: 10.1172/JCI62182
Chang, H. H., Falick, A. M., Carlton, P. M., Sedat, J. W., DeRisi, J. L., and Marletta, M. A. (2008). N-terminal processing of proteins exported by malaria parasites. Mol. Biochem. Parasitol. 160, 107–115. doi: 10.1016/j.molbiopara.2008.04.011
Charpian, S., and Przyborski, J. M. (2008). Protein transport across the parasitophorous vacuole of Plasmodium falciparum: into the great wide open. Traffic 9, 157–165. doi: 10.1111/j.1600-0854.2007.00648.x
Chen, Q., Barragan, A., Fernandez, V., Sundström, A., Schlichtherle, M., Sahlén, A., et al. (1998). Identification of Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) as the rosetting ligand of the malaria parasite P. falciparum. J. Exp. Med. 187, 15–23. doi: 10.1084/jem.187.1.15
Cheng, Q., Cloonan, N., Fischer, K., Thompson, J., Waine, G., Lanzer, M., et al. (1998). stevor and rif are Plasmodium falciparum multicopy gene families which potentially encode variant antigens. Mol. Biochem. Parasitol. 97, 161–176. doi: 10.1016/S0166-6851(98)00144-3
Claessens, A., Adams, Y., Ghumra, A., Lindergard, G., Buchan, C. C., Andisi, C., et al. (2012). A subset of group A-like var genes encodes the malaria parasite ligands for binding to human brain endothelial cells. Proc. Natl. Acad. Sci. U.S.A. 109, E1772–E1781. doi: 10.1073/pnas.1120461109
Claudianos, C., Dessens, J. T., Trueman, H. E., Arai, M., Mendoza, J., Butcher, G. A., et al. (2002). A malaria scavenger receptor-like protein essential for parasite development. Mol. Microbiol. 45, 1473–1484. doi: 10.1046/j.1365-2958.2002.03118.x
Cooke, B. M., Mohandas, N., and Coppel, R. L. (2001). The malaria-infected red blood cell: structural and functional changes. Adv. Parasitol. 50, 1–86. doi: 10.1016/S0065-308X(01)50029-9
Crabb, B. S., Cooke, B. M., Reeder, J. C., Waller, R. F., Caruana, S. R., Davern, K. M., et al. (1997). Targeted gene disruption shows that knobs enable malaria-infected red cells to cytoadhere under physiological shear stress. Cell 89, 287–296. doi: 10.1016/S0092-8674(00)80207-X
Das, S., Hertrich, N., Perrin A. J., Withers-Martinez, C., Collins, C. R., Jones, M. L., et al. (2015). Processing of Plasmodium falciparum merozoite surface protein msp1 activates a spectrin-binding function enabling parasite egress from RBCs. Cell Host Microbe 18, 433–444. doi: 10.1016/j.chom.2015.09.007
Deitsch, K. W., and Wellems, T. E. (1996). Membrane modifications in erythrocytes parasitized by Plasmodium falciparum. Mol. Biochem. Parasitol. 76, 1–10. doi: 10.1016/0166-6851(95)02575-8
de Koning-Ward, T. F., Gilson, P. R., Boddey, J. A., Rug, M., Smith, B. J., Papenfuss, A. T., et al. (2009). A novel protein export machine in malaria parasites. Nature 459, 945–949. doi: 10.1038/nature08104
Desai, S. A. (2012). Ion and nutrient uptake by malaria parasite-infected erythrocytes. Cell Microbiol. 14, 1003–1009. doi: 10.1111/j.1462-5822.2012.01790.x
Desai, S. A., and Miller, L. H. (2014). Malaria: protein-export pathway illuminated. Nature 511, 541–542. doi: 10.1038/nature13646
Dessens, J. T., Sinden, R. E., and Claudianos, C. (2004). LCCL proteins of apicomplexan parasites. Trends Parasitol. 20, 102–108. doi: 10.1016/j.pt.2004.01.002
Dluzewski, A. R., Rangachari, K., Wilson, R. J. M., and Gratzer, W. B. (1986). Plasmodium falciparum: protease inhibitors and inhibition of erythrocyte invasion. Exp. Parasitol. 62, 416–422. doi: 10.1016/0014-4894(86)90050-0
Dua, M., Raphael, P., Sijwali, P. S., Rosenthal, P. J., and Hanspal, M. (2001). Recombinant falcipain-2 cleaves erythrocyte membrane ankyrin and protein 4.1. Mol. Biochem. Parasitol. 116, 95–99. doi: 10.1016/S0166-6851(01)00306-1
Elsworth, B., Matthews, K., Nie, C. Q., Kalanon, M., Charnaud, S. C., Sanders, P. R., et al. (2014). PTEX is an essential nexus for protein export in malaria parasites. Nature 511, 587–591. doi: 10.1038/nature13555
Epp, C., and Deitsch, K. (2006). Deciphering the export pathway of malaria surface proteins. Trends Parasitol. 22, 401–404. doi: 10.1016/j.pt.2006.07.002
Florens, L., Washburn, M. P., Raine, J. D., Anthony, R. M., Grainger, M., Haynes, J. D., et al. (2002). A proteomic view of the Plasmodium falciparum life cycle. Nature 419, 520–526. doi: 10.1038/nature01107
Gardner, J. P., Pinches, R. A., Roberts, D. J., and Newbold, C. I. (1996). Variant antigens and endothelial receptor adhesion in Plasmodium falciparum. Proc. Natl. Acad. Sci. U.S.A. 93, 3503–3508. doi: 10.1073/pnas.93.8.3503
Gehde, N., Hinrichs, C., Montilla, I., Charpian, S., Lingelbach, K., and Przyborski, J. M. (2009). Protein unfolding is an essential requirement for transport across the parasitophorous vacuolar membrane of Plasmodium falciparum. Mol. Microbiol. 71, 613–628. doi: 10.1111/j.1365-2958.2008.06552.x
Giacomini, E., Iona, E., Ferroni, L., Miettinen, M., Fattorini, L., Orefici, G., et al. (2001). Infection of human macrophages and dendritic cells with Mycobacterium tuberculosis induces a differential cytokine gene expression that modulates T cell response. J. Immunol. 166, 7033–7041. doi: 10.4049/jimmunol.166.12.7033
Gilson, P. R., and Crabb, B. S. (2009). Morphology and kinetics of the three distinct phases of red blood cell invasion by Plasmodium falciparum merozoites. Int. J. Parasitol. 39, 91–96. doi: 10.1016/j.ijpara.2008.09.007
Ginsburg, H., Krugliak, M., Eidelman, O., and Ioav Cabantchik, Z. (1983). New permeability pathways induced in membranes of Plasmodium falciparum infected erythrocytes. Mol. Biochem. Parasitol. 8, 177–190. doi: 10.1016/0166-6851(83)90008-7
Glenister, F. K., Fernandez, K. M., Kats, L. M., Hanssen, E., Mohandas, N., Coppel, R. L., et al. (2009). Functional alteration of red blood cells by a megadalton protein of Plasmodium falciparum. Blood 113, 919–928. doi: 10.1182/blood-2008-05-157735
Goel, V. K., Li, X., Chen, H., Liu, S.-C., Chishti, A. H., and Oh, S. S. (2003). Band 3 is a host receptor binding merozoite surface protein 1 during the Plasmodium falciparum invasion of erythrocytes. Proc. Natl. Acad. Sci. U.S.A. 100, 5164–5169. doi: 10.1073/pnas.0834959100
Goldberg, D. E., and Cowman, A. F. (2010). Moving in and renovating: exporting proteins from Plasmodium into host erythrocytes. Nat. Rev. Microbiol. 8, 617–621. doi: 10.1038/nrmicro2420
Grant, B., and Greenwald, I. (1996). The Caenorhabditis elegans sel-1 gene, a negative regulator of lin-12 and glp-1, encodes a predicted extracellular protein. Genetics 143, 237–247.
Gruring, C., Heiber, A., Kruse, F., Flemming, S., Franci, G., Colombo, S. F., et al. (2012). Uncovering common principles in protein export of malaria parasites. Cell Host Microbe 12, 717–729. doi: 10.1016/j.chom.2012.09.010
Gunaratne, R. S., Sajid, M., Ling, I. T., Tripathi, R., Pachebat, J. A., and Holder, A. A. (2000). Characterization of N-myristoyltransferase from Plasmodium falciparum. Biochem. J. 348(Pt 2), 459–463. doi: 10.1042/bj3480459
Hadley, T., Aikawa, M., and Miller, L. H. (1983). Plasmodium knowlesi: studies on invasion of rhesus erythrocytes by merozoites in the presence of protease inhibitors. Exp. Parasitol. 55, 306–311. doi: 10.1016/0014-4894(83)90027-9
Haldar, K., and Mohandas, N. (2007). Erythrocyte remodeling by malaria parasites. Curr. Opin. Hematol. 14, 203–209. doi: 10.1097/MOH.0b013e3280f31b2d
Haldar, K., Mohandas, N., Samuel, B. U., Harrison, T., Hiller, N. L., Akompong, T., et al. (2002). Protein and lipid trafficking induced in erythrocytes infected by malaria parasites. Cell Microbiol. 4, 383–395. doi: 10.1046/j.1462-5822.2002.00204.x
Heiber, A., Kruse, F., Pick, C., Gruring, C., Flemming, S., Oberli, A., et al. (2013). Identification of new PNEPs indicates a substantial non-PEXEL exportome and underpins common features in Plasmodium falciparum protein export. PLoS Pathog. 9:e1003546. doi: 10.1371/journal.ppat.1003546
Hewitson, J. P., Grainger, J. R., and Maizels, R. M. (2009). Helminth immunoregulation: the role of parasite secreted proteins in modulating host immunity. Mol. Biochem. Parasitol. 167, 1–11. doi: 10.1016/j.molbiopara.2009.04.008
Hiller, N. L., Bhattacharjee, S., van Ooij, C., Liolios, K., Harrison, T., Lopez-Estrano, C., et al. (2004). A host-targeting signal in virulence proteins reveals a secretome in malarial infection. Science 306, 1934–1937. doi: 10.1126/science.1102737
Hinterberg, K., Scherf, A., Gysin, J., Toyoshima, T., Aikawa, M., Mazie, J. C., et al. (1994). Plasmodium falciparum: the Pf332 antigen is secreted from the parasite by a brefeldin A-dependent pathway and is translocated to the erythrocyte membrane via the Maurer’s clefts. Exp. Parasitol. 79, 279–291. doi: 10.1006/expr.1994.1091
Hisaeda, H., Yasutomo, K., and Himeno, K. (2005). Malaria: immune evasion by parasites. Int. J. Biochem. Cell Biol. 37, 700–706. doi: 10.1016/j.biocel.2004.10.009
Ho, M., and White, N. J. (1999). Molecular mechanisms of cytoadherence in malaria. Am. J. Physiol. Cell Physiol. 276, C1231–C1242.
Hodder, A. N., Sleebs, B. E., Czabotar, P. E., Gazdik, M., Xu, Y., O’Neill, M. T., et al. (2015). Structural basis for plasmepsin V inhibition that blocks export of malaria proteins to human erythrocytes. Nat. Struct. Mol. Biol. 22, 590–596. doi: 10.1038/nsmb.3061
Homewood, C. A., and Neame, K. D. (1974). Malaria and the permeability of the host erythrocyte. Nature 252, 718–719. doi: 10.1038/252718a0
Horrocks, P., Pinches, R. A., Chakravorty, S. J., Papakrivos, J., Christodoulou, Z., Kyes, S. A., et al. (2005). PfEMP1 expression is reduced on the surface of knobless Plasmodium falciparum infected erythrocytes. J. Cell Sci. 118, 2507–2518. doi: 10.1242/jcs.02381
Jeppesen, A., Ditlev, S. B., Soroka, V., Stevenson, L., Turner, L., Dzikowski, R., et al. (2015). Multiple Plasmodium falciparum erythrocyte membrane protein 1 variants per genome can bind IgM via Its Fc fragment fcmu. Infect. Immun. 83, 3972–3981.
Kirk, K. (2015). Ion regulation in the malaria parasite. Annu. Rev. Microbiol. 69, 341–359. doi: 10.1146/annurev-micro-091014-104506
Knuepfer, E., Rug, M., Klonis, N., Tilley, L., and Cowman, A. F. (2005). Trafficking determinants for PfEMP3 export and assembly under the Plasmodium falciparum-infected red blood cell membrane. Mol. Microbiol. 58, 1039–1053. doi: 10.1111/j.1365-2958.2005.04895.x
Kutner, S., Ginsburg, H., and Cabantchik, Z. I. (1983). Permselectivity changes in malaria (Plasmodium falciparum) infected human red blood cell membranes. J. Cell. Physiol. 114, 245–251. doi: 10.1002/jcp.1041140215
Kyes, S. A., Rowe, J. A., Kriek, N., and Newbold, C. I. (1999). Rifins: a second family of clonally variant proteins expressed on the surface of red cells infected with Plasmodium falciparum. Proc. Natl. Acad. Sci. U.S.A. 96, 9333–9338. doi: 10.1073/pnas.96.16.9333
Langhorne, J. R., Albano, F., Hensmann, M., Sanni, L., Cadman, E., Voisine, C., et al. (2004). Dendritic cells, pro-inflammatory responses, and antigen presentation in a rodent malaria infection. Immunol. Rev. 201, 35–47.
Lanzer, M., Wickert, H., Krohne, G., Vincensini, L., and Breton, C. B. (2006). Maurer’s clefts: a novel multi-functional organelle in the cytoplasm of Plasmodium falciparum-infected erythrocytes. Int. J. Parasitol. 36, 23–36. doi: 10.1016/j.ijpara.2005.10.001
Lee, M. C. S., and Fidock, D. A. (2008). Arresting malaria parasite egress from infected red blood cells. Nat. Chem. Biol. 4, 161–162. doi: 10.1038/nchembio0308-161
Lingelbach, K., and Przyborski, J. M. (2006). The long and winding road: protein trafficking mechanisms in the Plasmodium falciparum infected erythrocyte. Mol. Biochem. Parasitol. 147, 1–8. doi: 10.1016/j.molbiopara.2006.01.014
Lisk, G., Pain, M., Gluzman, I. Y., Kambhampati, S., Furuya, T., Su, X.-Z., et al. (2008). Changes in the plasmodial surface anion channel reduce leupeptin uptake and can confer drug resistance in Plasmodium falciparum-infected erythrocytes. Antimicrob. Agents Chemother. 52, 2346–2354.
Lower, M., Weydig, C., Metzler, D., Reuter, A., Starzinski-Powitz, A., Wessler, S., et al. (2008). Prediction of extracellular proteases of the human pathogen Helicobacter pylori reveals proteolytic activity of the Hp1018/19 protein HtrA. PLoS ONE 3:e3510. doi: 10.1371/journal.pone.0003510
Lustigman, S., Anders, R. F., Brown, G. V., and Coppel, R. L. (1990). The mature-parasite-infected erythrocyte surface antigen (MESA) of Plasmodium falciparum associates with the erythrocyte membrane skeletal protein, band 4.1. Mol. Biochem. Parasitol. 38, 261–270. doi: 10.1016/0166-6851(90)90029-L
Ma, J., Rahlfs, S., Jortzik, E., Schirmer, R. H., Przyborski, J. M., and Becker, K. (2012). Subcellular localization of adenylate kinases in Plasmodium falciparum. FEBS Lett. 586, 3037–3043. doi: 10.1016/j.febslet.2012.07.013
MacDonald, S. M., Bhisutthibhan, J., Shapiro, T. A., Rogerson, S. J., Taylor, T. E., Tembo, M., et al. (2001). Immune mimicry in malaria: Plasmodium falciparum secretes a functional histamine-releasing factor homolog in vitro and in vivo. Proc. Natl. Acad. Sci. U.S.A. 98, 10829–10832. doi: 10.1073/pnas.201191498
Magowan, C., Wollish, W., Anderson, L., and Leech, J. (1988). Cytoadherence by Plasmodium falciparum-infected erythrocytes is correlated with the expression of a family of variable proteins on infected erythrocytes. J. Exp. Med. 168, 1307–1320. doi: 10.1084/jem.168.4.1307
Maier, A. G., Cooke, B. M., Cowman, A. F., and Tilley, L. (2009). Malaria parasite proteins that remodel the host erythrocyte. Nat. Rev. Microbiol. 7, 341–354. doi: 10.1038/nrmicro2110
Maier, A. G., Rug, M., O’Neill, M. T., Beeson, J. G., Marti, M., Reeder, J., et al. (2007). Skeleton-binding protein 1 functions at the parasitophorous vacuole membrane to traffic PfEMP1 to the Plasmodium falciparum–infected erythrocyte surface. Blood 109, 1289–1297. doi: 10.1182/blood-2006-08-043364
Marti, M., Baum, J., Rug, M., Tilley, L., and Cowman, A. F. (2005). Signal-mediated export of proteins from the malaria parasite to the host erythrocyte. J. Cell Biol. 171, 587–592. doi: 10.1083/jcb.200508051
Marti, M., Good, R. T., Rug, M., Knuepfer, E., and Cowman, A. F. (2004). Targeting malaria virulence and remodeling proteins to the host erythrocyte. Science 306, 1930–1933. doi: 10.1126/science.1102452
Marti, M., and Spielmann, T. (2013). Protein export in malaria parasites: many membranes to cross. Curr. Opin. Microbiol. 16, 445–451. doi: 10.1016/j.mib.2013.04.010
Martin, R. E., Ginsburg, H., and Kirk, K. (2009). Membrane transport proteins of the malaria parasite. Mol. Microbiol. 74, 519–528. doi: 10.1111/j.1365-2958.2009.06863.x
McKerrow, J. H., Sun, E., Rosenthal, P. J., and Bouvier, J. (1993). The proteases and pathogenicity of parasitic protozoa. Annu. Rev. Microbiol. 47, 821–853. doi: 10.1146/annurev.mi.47.100193.004133
Mendis, K. N., and Carter, R. (1995). Clinical disease and pathogenesis in malaria. Parasitol. Today 11, TI1–TI16. doi: 10.1016/0169-4758(95)80143-X
Miller, L. H., Baruch, D. I., Marsh, K., and Doumbo, O. K. (2002). The pathogenic basis of malaria. Nature 415, 673–679. doi: 10.1038/415673a
Miller, L. H., Good, M. F., and Milon, G. (1994). Malaria pathogenesis. Science 264, 1878–1883. doi: 10.1126/science.8009217
Millington, O. R., Di Lorenzo, C., Phillips, R. S., Garside, P., and Brewer, J. M. (2006). Suppression of adaptive immunity to heterologous antigens during Plasmodium infection through hemozoin-induced failure of dendritic cell function. J. Biol. 5:5. doi: 10.1186/jbiol34
Mills, J., Diez-Silva, M., Quinn, D., Dao, M., Lang, M., Tan, K., et al. (2007). Effect of plasmodial RESA protein on deformability of human red blood cells harboring Plasmodium falciparum. Proc. Natl. Acad. Sci. U.S.A. 104, 9213–9217. doi: 10.1073/pnas.0703433104
Mundwiler-Pachlatko, E., and Beck, H.-P. (2013). Maurer’s clefts, the enigma of Plasmodium falciparum. Proc. Natl. Acad. Sci. U.S.A. 110, 19987–19994. doi: 10.1073/pnas.1309247110
Newbold, C., Craig, A., Kyes, S., Rowe, A., Fernandez-Reyes, D., and Fagan, T. (1999). Cytoadherence, pathogenesis and the infected red cell surface in Plasmodium falciparum. Int. J. Parasitol. 29, 927–937. doi: 10.1016/S0020-7519(99)00049-1
Nguitragool, W., Bokhari, A. A. B., Pillai, A. D., Rayavara, K., Sharma, P., Turpin, B., et al. (2011). Malaria parasite clag genes determine nutrient uptake channel activity on infected red blood cells. Cell 145, 665–677. doi: 10.1016/j.cell.2011.05.002
Niang, M., Bei A. K., Madnani, K. G., Pelly, S., Dankwa, S., Kanjee, U., et al. (2014). STEVOR is a Plasmodium falciparum erythrocyte binding protein that mediates merozoite invasion and rosetting. Cell Host Microbe 16, 81–93. doi: 10.1016/j.chom.2014.06.004
Oberli, A., Slater, L. M., Cutts, E., Brand, F., Mundwiler-Pachlatko, E., Rusch, S., et al. (2014). A Plasmodium falciparum PHIST protein binds the virulence factor PfEMP1 and comigrates to knobs on the host cell surface. FASEB J. 28, 4420–4433. doi: 10.1096/fj.14–256057
Pain, A., Ferguson, D. J., Kai, O., Urban, B. C., Lowe, B., Marsh, K., et al. (2001). Platelet-mediated clumping of Plasmodium falciparum-infected erythrocytes is a common adhesive phenotype and is associated with severe malaria. Proc. Natl. Acad. Sci. U.S.A. 98, 1805–1810. doi: 10.1073/pnas.98.4.1805
Parish, L. A., Mai, D. W., Jones, M. L., Kitson, E. L., and Rayner, J. C. (2013). A member of the Plasmodium falciparum PHIST family binds to the erythrocyte cytoskeleton component band 4.1. Malar. J. 12, 160–160. doi: 10.1186/1475-2875-12–160
Parker, P. D., Tilley, L., and Klonis, N. (2004). Plasmodium falciparum induces reorganization of host membrane proteins during intraerythrocytic growth. Blood 103, 2404–2406. doi: 10.1182/blood-2003-08-2692
Pei, X., An, X., Guo, X., Tarnawski, M., Coppel, R., and Mohandas, N. (2005). Structural and functional studies of interaction between Plasmodium falciparum knob-associated histidine-rich protein (KAHRP) and erythrocyte spectrin. J. Biol. Chem. 280, 31166–31171. doi: 10.1074/jbc.M505298200
Pillai, A. D., Nguitragool, W., Lyko, B., Dolinta, K., Butler, M. M., Nguyen, S. T., et al. (2012). Solute restriction reveals an essential role for clag3-associated channels in malaria parasite nutrient acquisition. Mol. Pharmacol. 82, 1104–1114. doi: 10.1124/mol.112.081224
Przyborski, J. M., Wickert, H., Krohne, G., and Lanzer, M. (2003). Maurer’s clefts—a novel secretory organelle? Mol. Biochem. Parasitol. 132, 17–26. doi: 10.1016/S0166-6851(03)00212-3
Ranganathan, S., and Garg, G. (2009). Secretome: clues into pathogen infection and clinical applications. Genome Med. 1:113. doi: 10.1186/gm113
Raphael, P., Takakuwa, Y., Manno, S., Liu, S. C., Chishti, A. H., and Hanspal, M. (2000). A cysteine protease activity from Plasmodium falciparum cleaves human erythrocyte ankyrin. Mol. Biochem. Parasitol. 110, 259–272. doi: 10.1016/S0166-6851(00)00283-8
Rodrigues, V., Cordeiro-da-Silva, A., Laforge, M., Ouaissi, A., Akharid, K., Silvestre, R., et al. (2014). Impairment of T cell function in parasitic infections. PLoS Negl. Trop. Dis. 8:e2567. doi: 10.1371/journal.pntd.0002567
Rowe, A., Obeiro, J., Newbold, C. I., and Marsh, K. (1995). Plasmodium falciparum rosetting is associated with malaria severity in Kenya. Infect. Immun. 63, 2323–2326.
Rowe, J. A., Claessens, A., Corrigan, R. A., and Arman, M. (2009). Adhesion of Plasmodium falciparum-infected erythrocytes to human cells: molecular mechanisms and therapeutic implications. Expert Rev. Mol. Med. 11:e16. doi: 10.1017/S1462399409001082
Rowe, J. A., Obiero, J., Marsh, K., and Raza, A. (2002). Short report: positive correlation between rosetting and parasitemia in Plasmodium falciparum clinical isolates. Am. J. Trop. Med. Hyg. 66, 458–460.
Rudzinska, M. A., and Trager, W. (1968). The fine structure of trophozoites and gametocytes in Plasmodium coatneyi∗. J. Protozool. 15, 73–88. doi: 10.1111/j.1550-7408.1968.tb02091.x
Rug, M., Prescott, S. W., Fernandez, K. M., Cooke, B. M., and Cowman, A. F. (2006). The role of KAHRP domains in knob formation and cytoadherence of P. falciparum-infected human erythrocytes. Blood 108, 370–378. doi: 10.1182/blood-2005-11-4624
Russo, I., Babbitt, S., Muralidharan, V., Butler, T., Oksman, A., and Goldberg, D. E. (2010). Plasmepsin V licenses Plasmodium proteins for export into the host erythrocyte. Nature 463, 632–636. doi: 10.1038/nature08726
Sacks, D., and Sher, A. (2002). Evasion of innate immunity by parasitic protozoa. Nat. Immunol. 3, 1041–1047. doi: 10.1038/ni1102-1041
Sharma, P., Rayavara, K., Ito, D., Basore, K., and Desai, S. A. (2015). A CLAG3 mutation in an amphipathic transmembrane domain alters malaria parasite nutrient channels and confers leupeptin resistance. Infect. Immun. 83, 2566–2574. doi: 10.1128/IAI.02966-14
Simone, O., Bejarano, M. T., Pierce, S. K., Antonaci, S., Wahlgren, M., Troye-Blomberg, M., et al. (2011). TLRs innate immunereceptors and Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) CIDR1alpha-driven human polyclonal B-cell activation. Acta Trop. 119, 144–150. doi: 10.1016/j.actatropica.2011.05.005
Singh, M., Mukherjee, P., Narayanasamy, K., Arora, R., Sen, S. D., Gupta, S., et al. (2009). Proteome analysis of Plasmodium falciparum extracellular secretory antigens at asexual blood stages reveals a cohort of proteins with possible roles in immune modulation and signaling. Mol. Cell. Proteom. 8, 2102–2118. doi: 10.1074/mcp.M900029-MCP200
Sleebs, B. E., Gazdik, M., O’Neill, M. T., Rajasekaran, P., Lopaticki, S., Lackovic, K., et al. (2014a). Transition state mimetics of the Plasmodium export element are potent inhibitors of plasmepsin V from P. falciparum and P. vivax. J. Med. Chem. 57, 7644–7662. doi: 10.1021/jm500797g
Sleebs, B. E., Lopaticki, S., Marapana, D. S., O’Neill, M. T., Rajasekaran, P., Gazdik, M., et al. (2014b). Inhibition of Plasmepsin V activity demonstrates its essential role in protein export, PfEMP1 display, and survival of malaria parasites. PLoS Biol 12:e1001897. doi: 10.1371/journal.pbio.1001897
Spielmann, T., and Gilberger, T.-W. (2010). Protein export in malaria parasites: do multiple export motifs add up to multiple export pathways? Trends Parasitol. 26, 6–10. doi: 10.1016/j.pt.2009.10.001
Spielmann, T., Hawthorne, P. L., Dixon, M. W. A., Hannemann, M., Klotz, K., Kemp, D. J., et al. (2006). A cluster of ring stage–specific genes linked to a locus implicated in cytoadherence in Plasmodium falciparum codes for PEXEL-negative and PEXEL-positive proteins exported into the host cell. Mol. Biol. Cell 17, 3613–3624. doi: 10.1091/mbc.E06-04-0291
Spillman, N. J., Allen, R. J., McNamara, C. W., Yeung, B. K., Winzeler, E. A., Diagana, T. T., et al. (2013). Na+ regulation in the malaria parasite Plasmodium falciparum involves the cation ATPase PfATP4 and is a target of the spiroindolone antimalarials. Cell Host Microbe 13, 227–237. doi: 10.1016/j.chom.2012.12.006
Sponaas, A.-M., Cadman, E. T., Voisine, C., Harrison, V., Boonstra, A., O’Garra, A., et al. (2006). Malaria infection changes the ability of splenic dendritic cell populations to stimulate antigen-specific T cells. J. Exp. Med. 203, 1427–1433. doi: 10.1084/jem.20052450
Spycher, C., Rug, M., Pachlatko, E., Hanssen, E., Ferguson, D., Cowman, A. F., et al. (2008). The Maurer’s cleft protein MAHRP1 is essential for trafficking of PfEMP1 to the surface of Plasmodium falciparum-infected erythrocytes. Mol. Microbiol. 68, 1300–1314. doi: 10.1111/j.1365-2958.2008.06235.x
Stevenson, L., Huda, P., Jeppesen, A., Laursen, E., Rowe, J. A., Craig, A., et al. (2015a). Investigating the function of Fc-specific binding of IgM to Plasmodium falciparum erythrocyte membrane protein 1 mediating erythrocyte rosetting. Cell Microbiol. 17, 819–831. doi: 10.1111/cmi.12403
Stevenson, L., Laursen, E., Cowan, G. J., Bandoh, B., Barfod, L., Cavanagh, D. R., et al. (2015b). α 2-Macroglobulin can crosslink multiple Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) molecules and may facilitate adhesion of parasitized erythrocytes. PLoS Pathog. 11:e1005022. doi: 10.1371/journal.ppat.1005022
Taraschi, T. F., O’Donnell, M., Martinez, S., Schneider, T., Trelka, D., Fowler, V. M., et al. (2003). Generation of an erythrocyte vesicle transport system by Plasmodium falciparum malaria parasites. Blood 102, 3420–3426. doi: 10.1182/blood-2003-05–1448
Teirlinck, A. C., Roestenberg, M., Bijker, E. M., Hoffman, S. L., Sauerwein, R. W., and Scholzen, A. (2015). Plasmodium falciparum infection of human volunteers activates monocytes and CD16+ dendritic cells and induces upregulation of CD16 and CD1c expression. Infect. Immun. 83, 3732–3739. doi: 10.1128/IAI.00473-15
Thavayogarajah, T., Gangopadhyay, P., Rahlfs, S., Becker, K., Lingelbach, K., Przyborski, J. M., et al. (2015). Alternative protein secretion in the malaria parasite Plasmodium falciparum. PLoS ONE 10:e0125191. doi: 10.1371/journal.pone.0125191
Trager, W., Rudzinska, M. A., and Bradbury, P. C. (1966). The fine structure of Plasmodium falciparum and its host erythrocytes in natural malarial infections in man. Bull. World Health Organ. 35, 883–885.
Udomsangpetch, R., Wahlin, B., Carlson, J., Berzins, K., Torii, M., Aikawa, M., et al. (1989). Plasmodium falciparum-infected erythrocytes form spontaneous erythrocyte rosettes. J. Exp. Med. 169, 1835–1840. doi: 10.1084/jem.169.5.1835
van Ooij, C., Tamez, P., Bhattacharjee, S., Hiller, N. L., Harrison, T., Liolios, K., et al. (2008). The malaria secretome: from algorithms to essential function in blood stage infection. PLoS Pathog. 4:e1000084. doi: 10.1371/journal.ppat.1000084
Vásárhelyi, V., Trexler, M., and Patthy, L. (2014). Both LCCL-domains of human CRISPLD2 have high affinity for lipid A. Biochimie 97, 66–71. doi: 10.1016/j.biochi.2013.09.021
Verma, R., Tiwari, A., Kaur, S., Varshney, G. C., and Raghava, G. P. (2008). Identification of proteins secreted by malaria parasite into erythrocyte using SVM and PSSM profiles. BMC Bioinform. 9:201. doi: 10.1186/1471-2105-9-201
Waller, K. L., Cooke, B. M., Nunomura, W., Mohandas, N., and Coppel, R. L. (1999). Mapping the binding domains involved in the interaction between the Plasmodium falciparum knob-associated histidine-rich protein (kahrp) and the cytoadherence ligand P. falciparum erythrocyte membrane protein 1 (PfEMP1). J. Biol. Chem. 274, 23808–23813. doi: 10.1074/jbc.274.34.23808
Weng, H., Guo, X., Papoin, J., Wang, J., Coppel, R., Mohandas, N., et al. (2014). Interaction of Plasmodium falciparum knob-associated histidine-rich protein (KAHRP) with erythrocyte ankyrin R is required for its attachment to the erythrocyte membrane. Biochim. Biophys. Acta 1838, 185–192. doi: 10.1016/j.bbamem.2013.09.014
Wetzel, J., Herrmann, S., Swapna, L. S., Prusty, D., John Peter, A. T., Kono, M., et al. (2015). The role of palmitoylation for protein recruitment to the inner membrane complex of the malaria parasite. J. Biol. Chem. 290, 1712–1728. doi: 10.1074/jbc.M114.598094
World Health Organization [WHO] (2014). World Malaria Report 2013. Geneva: World Health Organization.
Wickham, M. E., Rug, M., Ralph, S. A., Klonis, N., McFadden, G. I., Tilley, L., et al. (2001). Trafficking and assembly of the cytoadherence complex in Plasmodium falciparum-infected human erythrocytes. EMBO J. 20, 5636–5649. doi: 10.1093/emboj/20.20.5636
Wirth, C. C., Glushakova, S., Scheuermayer, M., Repnik, U., Garg, S., Schaack, D., et al. (2014). Perforin-like protein PPLP2 permeabilizes the red blood cell membrane during egress of Plasmodium falciparum gametocytes. Cell Microbiol. 16, 709–733. doi: 10.1111/cmi.12288
Zambrano-Villa, S., Rosales-Borjas, D., Carrero, J. C., and Ortiz-Ortiz, L. (2002). How protozoan parasites evade the immune response. Trends Parasitol. 18, 272–278. doi: 10.1016/S1471-4922(02)02289-4
Keywords: plasmodium, secretome, cytoadherence, immune modulation, anti-malarial therapy
Citation: Soni R, Sharma D and Bhatt TK (2016) Plasmodium falciparum Secretome in Erythrocyte and Beyond. Front. Microbiol. 7:194. doi: 10.3389/fmicb.2016.00194
Received: 02 November 2015; Accepted: 05 February 2016;
Published: 19 February 2016.
Edited by:
Alexandre Morrot, Federal University of Rio de Janeiro, BrazilReviewed by:
Anne-Marie Zeeman, Biomedical Primate Research Centre, NetherlandsWai-Hong Tham, The Walter and Eliza Hall Institute, Australia
Copyright © 2016 Soni, Sharma and Bhatt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tarun K. Bhatt, dGFydW5AY3VyYWouYWMuaW4=