 Sophi Marmen
Sophi Marmen Dikla Aharonovich
Dikla Aharonovich Michal Grossowicz
Michal Grossowicz Lior Blank
Lior Blank Yosef Z. Yacobi
Yosef Z. Yacobi Daniel J. Sher
Daniel J. Sher- 1Department of Marine Biology, Charney School of Marine Sciences, University of Haifa, Haifa, Israel
- 2Department of Plant Pathology and Weed Research, ARO, Volcani Center, Bet Dagan, Israel
- 3Yigal Allon Kinneret Limnological Laboratory, Israel Oceanographic and Limnological Research, Migdal, Israel
Toxic cyanobacterial blooms are a growing threat to freshwater bodies worldwide. In order for a toxic bloom to occur, a population of cells with the genetic capacity to produce toxins must be present together with the appropriate environmental conditions. In this study, we investigated the distribution patterns and phylogeny of potentially-toxic Microcystis (indicated by the presence and/or phylogeny of the mcyD and mcyA genes). Samples were collected from the water column of almost 60 water bodies across widely differing gradients of environmental conditions and land use in Israel. Potentially, toxic populations were common but not ubiquitous, detected in ~65% of the studied sites. Local environmental factors, including phosphorus and ammonia concentrations and pH, as well as regional conditions such as the distance from built areas and nature reserves, were correlated with the distribution of the mcyD gene. A specific phylogenetic clade of Microcystis, defined using the sequence of the mcyA gene, was preferentially associated with aquaculture facilities but not irrigation reservoirs. Our results reveal important environmental, geospatial, and land use parameters affecting the geographic distribution of toxinogenic Microcystis, suggesting non-random dispersal of these globally abundant toxic cyanobacteria.
Introduction
Cyanobacteria, as part of a larger algal community, form the base of the food web in many aquatic environments (both marine and freshwater; Paerl and Paul, 2012). However, under appropriate conditions, many cyanobacterial species can grow at a rapid rate and form massive “blooms” which negatively impact water quality, especially when the blooming species produces toxins (Schinder and Valentyne, 2008; Paerl and Otten, 2013). Such blooms, termed “Harmful Cyanobacterial Blooms” (cyanoHABs), are one of the major threats to water quality worldwide, affecting many ecologically and economically important water bodies, and occasionally causing severe health problems and/or mortality in livestock and humans (Pouria et al., 1998; Carmichael, 2001; Brianda et al., 2003; Falconer and Humpage, 2005; Paerl and Huisman, 2009). In the United States alone, cyanoHABs have been estimated to inflict as much as $2 billion a year in losses from water made unfit for recreation, drinking or agriculture (Stone, 2011). Over the last several decades the frequency and severity of reported cyanoHABs has increased significantly, and toxic blooms have been observed in water bodies where they have not previously been recorded (Sukenik et al., 2012). To date, it is unknown whether these newly-observed blooms occur due to cyanobacteria that were always part of the microbial population (and potentially had bloomed but this was not reported) or whether these organisms have recently colonized the water body in question. If the latter is true, it is unclear whether the newly-introduced cyanobacteria migrated naturally (e.g., carried by the wind, animals or birds, Sukenik et al., 2012; van Leeuwen et al., 2012), or were introduced by man, for example through fish stocking or ballast water (Padilla and Williams, 2004).
The toxins that are most commonly found in freshwater environments worldwide, and potentially have the highest impact on water use, are microcystins (Carmichael, 2001; Valério, 2010). Microcystins are cyclic peptides (Valério, 2010), which are synthesized by a non-ribosomal peptide/polyketide synthase (NRPS/PKS) enzyme complex encoded in the highly conserved mcy gene cluster which comprises two operons (Dittmann and Börner, 2005; Vasconcelos et al., 2010). The large microcystin synthetase complex consists of peptide synthetases (McyA–C), a polyketide synthase (McyD) and hybrid enzymes (McyE-G) (Pearson et al., 2004; Campos and Vasconcelos, 2010). In mammals, the organ most affected by microcystins is the liver, and specifically the hepatocytes that contain the target protein phosphatases type 1 and 2 (Dittmann and Wiegand, 2006; Valério, 2010). High concentrations of microcystins in water have been directly linked to cases of intoxication and death in animals and humans, and they are also potential carcinogens (Carmichael et al., 2001; Hernández et al., 2009). Microcystins are most commonly produced by cyanobacteria of the genus Microcystis, although other genera (e.g., Planktothrix, Oscillatoria, Anabaena, Anabaenopsis, Nostoc, Hapalosiphon, Snowella, and Woronichinia) are also known to produce the toxin (Allender et al., 2009; Campos and Vasconcelos, 2010; Valério, 2010; Kurmayer et al., 2014).
The abundance and severity of cyanoHABs caused by Microcystis are rising worldwide, affecting millions of people (Carmichael, 2001; O'Neil et al., 2012). It is known that increases in nutrient load, temperature, salinity, and UV light may all contribute to the emergence of microcystin—producing cyanoHABs (Davis et al., 2009; Dziallas and Grossart, 2011; Paerl et al., 2011a; O'Neil et al., 2012). However, it is currently unclear whether all water sources contain toxic cyanobacteria, (Kurmayer et al., 2011; van Gremberghe et al., 2011), or whether the distribution is patchy, with some locations harboring toxinogenic populations and others not. It is also unclear whether the presence of such potentially-toxic populations is related to the conditions within the water body or the region surrounding it. Importantly, since cyanoHAB development requires the presence of cells capable of toxin biosynthesis either in the water body or in the sediment (Green et al., 2008; Tanabe et al., 2009), it is likely that the patterns of local distribution determine, at short time scales, where and when these blooms will occur.
To start answering these questions, we studied the distribution of Microcystis with the genetic capacity to produce microcystins in the water column of almost 60 different freshwater bodies across Israel. Despite its small geographic size, Israel is rich in different climatic and geographic regions: from Mediterranean climate (cool, wet winters and hot, dry summers) to desert (with an average annual precipitation of less than 25 mm), from highly urban to almost unsettled and from highly industrial to mainly agricultural or natural areas. Many small water sources, such as springs, irrigation reservoirs and aquaculture facilities are found within this tapestry of different local and regional environmental conditions (Supplementary Figure 1). Most of these water sources are relatively isolated, i.e., they are not directly connected to each other (e.g., through channels or streams). Similar conditions are found in many Mediterranean and semi-arid regions. The small size of the country and its conspicuous physical variation provide a unique natural laboratory for analyzing the effects of local and regional climate and land use on aquatic microbial communities. The goals of the study were: (1) to map the distribution of potentially-toxic Microcystis strains during the period of the year when blooms are most common (and thus cells most likely to be found in the water column), using a highly conserved fragment of the mcyD gene; (2) to characterize the environmental (local and regional) factors associated with the presence of toxin-producing strains in the water column, and, (3) to determine, using the phylogenetically-informative mcyA gene, whether toxinogenic strains in Israel belong to a single or multiple populations, each potentially associated with a specific aquatic niche.
Materials and Methods
Collection of Samples for Molecular and Meta-Data Analyses
A total of 58 water bodies were sampled across Israel (Figure 1, Table 1, Supplementary Figure 1). Most of the samples (51) were collected between July and the beginning of November of 2011, a period that was characterized by stable, hot and dry weather. During this period Microcystis blooms are often observed in small reservoirs around Israel, maximizing the possibility of detecting cells in the water column. Another, seven locations at the desert south of Israel were sampled during the following winter for technical reasons (rows 54–60 in Table 1, sampled during January and March, 2012). Each location was sampled once from the edge of the water body during the late morning to early afternoon. During sampling, dissolved oxygen, temperature and pH were measured using field probes (Eutech instruments, Singapore). At each sampling location, 5 l of surface water were collected. The collected water was filtered on GF/F filters (nominal pore size 0.7 μm, Whatman, UK) for DNA and particulate nutrients and on GF/C filters (1.2 μm) for chlorophyll extraction. DNA sample were overlaid by lysis buffer (50 mM Tris pH = 8.3, 0.75 M Sucrose, 40 mM EDTA). The filtrate from the GF/F filters was collected for dissolved nutrients analysis. Filtration was performed until the filters were blocked, and the volume of filtered water was recorded (Table 1). All filtration steps were performed within 1 h at the sampling site, using a hand-held vacuum pump (Mityvac, USA). All samples were placed in a cooler with dry ice, and transferred to a −80°C freezer (DNA and chlorophyll) and −20°C (dissolved nutrients) within 10 h. The filtration equipment was washed with ethyl alcohol (70%) and distilled water between sampling sites, to prevent any cross contamination.
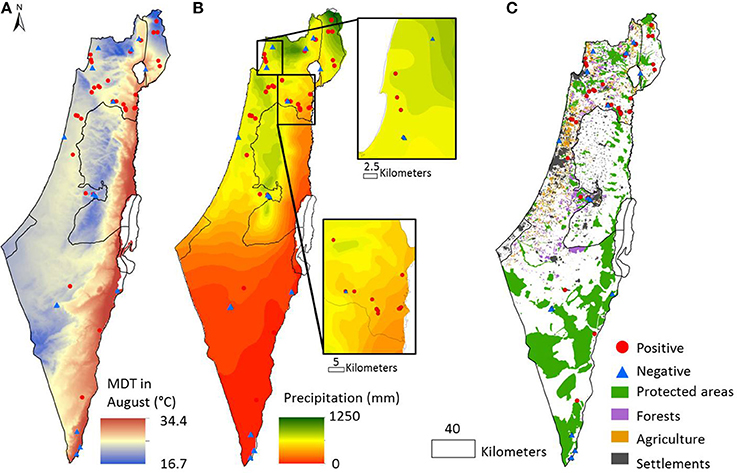
Figure 1. Environmental gradients and the distribution of toxinogenic Microcystis across Israel. Sampling locations where mcyD gene was detected are represented by a red circle, those where mcyD was not detected by a blue triangle. (A) Average day temperature range in August (mdt8); (B) Average annual precipitation; (C) Land use. Exact GPS coordinates of the sampling locations, as well as all of the parameters measured, are presented in Table 1.
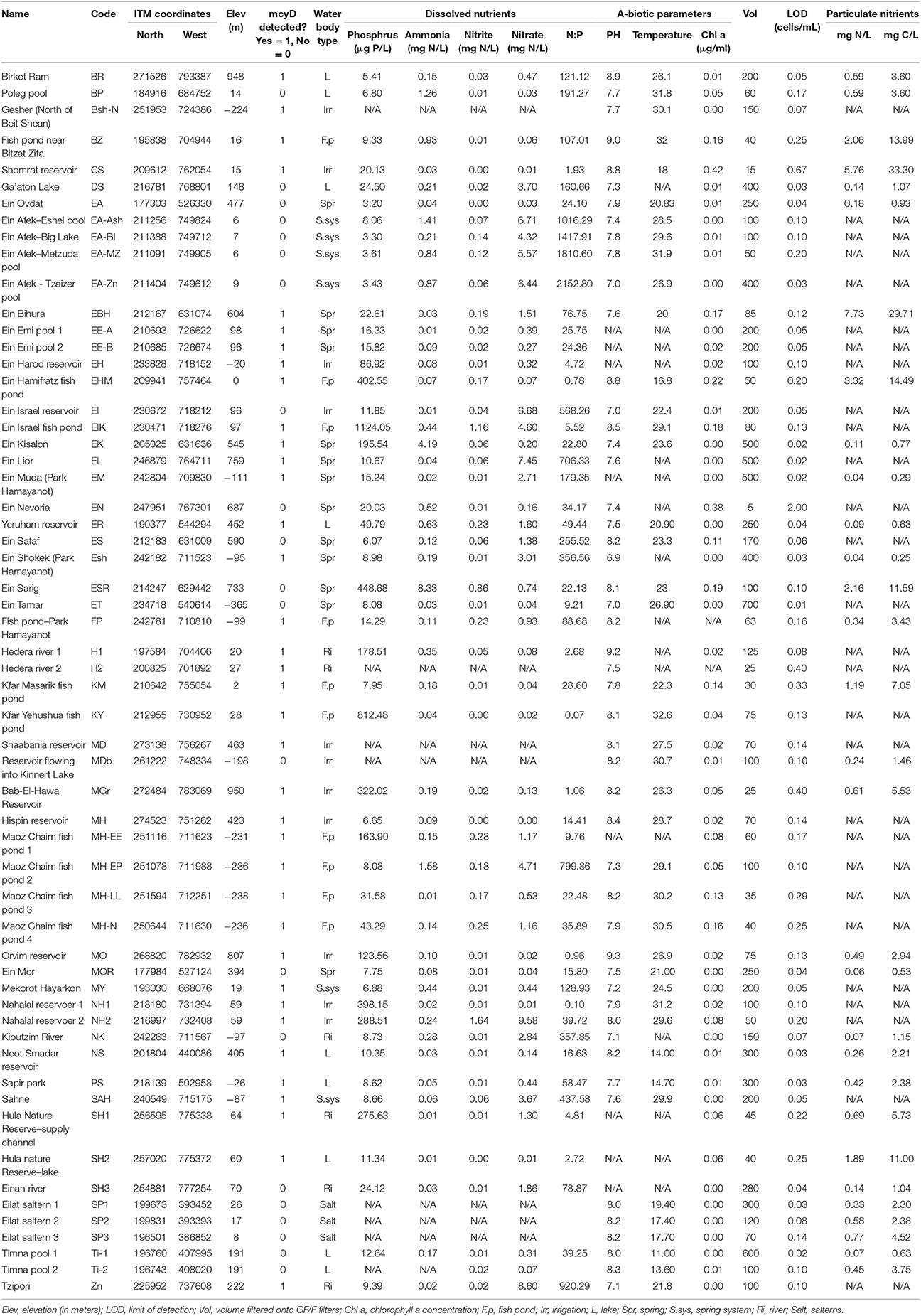
Table 1. Summarization of the sampled locations, ITM coordinates and all measured a-biotic parameters.
Nutrients and Chlorophyll Analyses
The concentration of dissolved nutrients was determined using a colorimetric standard method (APHA, 2005) in a flow injection automated ion analyzer (Quikchem, 8000 LACHAT instruments). The concentration of phosphorus was determined by the MAGIC method (Karl and Tien, 1992). Filters for particulate nutrient analysis were dried overnight at 60°C, weighed and analyzed using a C/H/N analyzer (Perkin Elmer). All of the nutrient measurements were performed at the Kinneret Limnological Laboratory, Israel.
Extraction of Chlorophyll was performed in absolute methanol for 12 h at room temperature in the dark and the extract filtered through a 0.2 μm filter. Chlorophyll a (Chl a) concentration was determined spectrophotometrically (Ritchie, 2008).
Environmental DNA Extraction, PCR Assays, and Cloning Procedure
Genomic DNA extraction was performed using a previously published protocol (Massana et al., 1997) with several modifications (Tzahor et al., 2009).
To determine whether there are toxic Microcystis species in the sampled waters, a sensitive end-point Polymerase Chain Reaction (PCR) was performed using MSF/R (Tillett et al., 2001) and mcyDF2/R2 (Kaebernick et al., 2000) primers sets for mcyA and mcyD genes respectively (Supplementary Table 1). The mcyD primers amplify the relevant genes from Microcystis but not from other cyanobacterial genera, as determined by two approaches: (1) BLAST against cyanobacterial genomes in the Integrated Microbial Genomes (IMG) server (Markowitz et al., 2014); (2) cloning and sequencing eight PCR fragments from four locations, all of which produced sequences which clustered together with mcyD from Microcystis aureginosa in a phylogenetic tree (Supplementary Figure 2). The mcyD primers were more sensitive than the mcyA primers, with a limit of detection of 10 toxinogenic cells/filter for mcyD and 100 toxinogenic cells/filter for mcyA, (Supplementary Figure 3), but were not phylogenetically informative (compare Supplementary Figures 2, 5). The mcyA primers were also Microcystis-specific, as shown in Figure 5 (see also Tillett et al., 2001). The end-point PCR for mcyD was at least as sensitive as quantitative PCR, with an average limit of detection of ~160 cells/L compared to ~260–400 cells/L in other studies (Rinta-Kanto et al., 2005; Baxa et al., 2010). For these reasons, and because our study focuses on identifying where and when potentially toxic cells are found in the water body rather than on the relationship between the quantity of toxinogenic strains and environmental conditions, we used end-point PCR with the mcyD primers for the identification of toxinogenic populations, and the mcyA gene for phylogenetic analyses. In all locations from which mcyA was amplified, mcyD amplification was also observed. As a positive control (e.g., to rule out PCR inhibitors), PCR was performed also using primers for general 16S rRNA (Frank et al., 2008) and cyanobacterial 16S rRNA genes (Nübel et al., 1997). Full details of the PCR program, primer sequences and enzymes used are detailed in the Supplementary Experimental Procedures. The mcyA and mcyD sequences were deposited in GenBank under accession numbers KU867658 - KU867777.
Phylogenetic Analysis
One hundred and twenty mcyA sequences, obtained from 17 different water reservoirs, were aligned using ClustalW in MEGA5 (Tamura et al., 2011) and a maximum likelihood tree with 1000 bootstraps was constructed. The tree was visualized with ITOL (http://itol.embl.de/; Letunic and Bork, 2011), and two datasets of explanatory variables were added: the location of the sample and the water body type.
Geographic Analyses
To determine whether there are regional environmental factors that may be used as predictive variables for the presence of toxinogenic Microcystis population, we analyzed seven environmental parameters: elevation, mean daily temperature of the hottest month (August—mdt8), mean annual rainfall, distance from built areas (including cities, villages, army bases, and industrial zones), distance from forests, distance from natural protected areas and distance from agricultural fields. Elevation was derived from the digital elevation model (DEM) at 33-m resolution using ArcGIS (ESRI, Redlands, CA). To test whether the sampled locations that were found positive to the presence of mcyD gene differed from the negative locations in these parameters, we used Mann–Whitney U-test with Bonferroni correction. SPSS was used for all statistical analysis (version 21.0).
Statistical Analyses
All multivariate analyses were performed with R i386 2.15. The abiotic characterization of 35 sampling locations, for which the full suite of metadata was available (Table 1), was analyzed here with 2D non-metric multidimensional scaling (nMDS) ordination using measurements of phosphorus, ammonia, nitrite, nitrate, pH, temperature, and chlorophyll-A concentrations. The data were first standardized, and then log10-transformed prior to nMDS analysis. We used Bray-Curtis for the dissimilarity distances matrix. The nMDS has a stress value which represents the divergence of the real value from the ordination output (Warwick and Clarke, 1993). Stress values lower than 0.2 mean that the ordination is useful, but higher values (>0.1) should be analyzed with caution. With this analysis, grouping was performed according to the geography, water-body type (category), and presence or absence of the mcyD gene. One-way ANOSIM (Analysis of Similarities) was performed to statistically differentiate the abiotic characterization of each grouping. This test is a-parametric and does not assume normality of the data. The calculated test statistic R has a value between −1 and 1 and practically rarely goes below 0 (Warwick and Clarke, 1993). R = 1 means that all the repeats within a group are similar to each other rather than to repeats in other groups. When R = 0, the similarity within and among all groups is averagely the same. SIMPER (Similarity Percentages) identify the “important” component from all the abiotic factors; i.e., what is the relative contribution of each abiotic factor to the dissimilarity between all inter-group pairs of samples.
To find whether there are associations between different nutrient concentrations or abiotic conditions and the presence of mcyD gene, we used chi square (χ2) test (α < 0.05). In order to determine whether specific water body types are over- or under-represented in different clades of the phylogenetic tree, we performed a two-sided Fisher's Exact Test with Bonferroni correction using the free online GraphPad software (http://graphpad.com/quickcalcs/contingency1.cfm). To determine whether clades were associated with differences in the concentration of nutrients or chlorophyll, a Kruskall-Wallis test was performed in SPSS.
To relate the environmental variables to the binary dependent variable (presence/absence of the mcyD genes), we used logistic regression in the framework of Generalized Linear Models (GLMs). Full details of the GLM procedure are found in the Supplementary Methods.
Results
Characterization of Sampling Sites
To map the distribution of potentially toxic Microcystis across different environmental conditions in Israel, we sampled 58 different water bodies belonging to seven functional categories: fish ponds, irrigation reservoirs, lakes, natural springs, spring systems (e.g., systems of several natural springs which flow from one into another), rivers and salterns (e.g., ponds for salt production). The sampling locations were selected in order to represent a wide diversity of environmental conditions (some of which are shown in Figure 1), and, for most locations, without prior knowledge of past cyanoHABs. The abiotic factors (e.g., pH values, phosphorus, ammonia, nitrate, and nitrite concentrations), which are expected to have a direct effect on the microbial community, differed widely between the sampled water bodies: pH values ranged from 6.9 to 9.3, phosphorus concentrations ranged from 3 to 1124 μg L−1, and total nitrogen concentration spanned from 0.03 to 11.5 mg L−1 (Table 1). Non-metric multidimensional scaling (nMDS) analysis of 35 locations for which a full suite of data were available (Table 1) revealed some grouping of the water bodies by both category (i.e., aquaculture, irrigation reservoirs or nature conservation), and geography (Figures 2A,B) with stress value of 0.149. Two of the categories of natural waters, namely spring systems and lakes, were each different from the agricultural waters (irrigation and fish-ponds, ANOSIM, Global R = 0.32, p < 0.001, pair-wise ANOSIM, R = 0.39–0.56, p < 0.01) mainly by the criteria of Chl a, phosphorus and nitrate values (Simper analysis). In terms of geography, the samples from the Negev Desert were different from the Jizreel Valley (Global R = 0.21, p = 0.003, pair-wise ANOSIM, R = 0.56, p < 0.01) and, with lower statistical support (R = 0.29–0.46, p < 0.05), from the other regions as well (Golan heights, Galilee, and Central region, Figure 2B). The main parameters for these variances were Chl a, phosphorus, nitrate and temperature values, as determined by Simper analysis. This may be due to the different sampling season (winter in the desert, summer in all other locations) or to inherent differences between the water bodies in the two climatically-different regions.
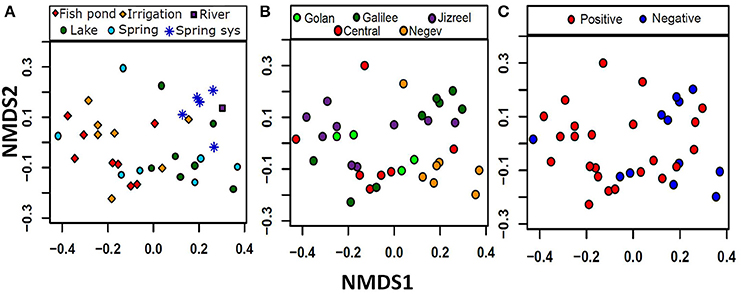
Figure 2. nMDS multivariate analysis indicates grouping of water bodies by geography, water use and mcyD gene presence. The samples ordinate to some extent by both the category or intended use of the water body (A) and the geographic location (B). A statisticaly significante (P = 0.011) separation could also be observed between samples in which mcyD gene presence was observed and those in which is the gene was not detected (C). The nMDS had a stress value of 0.149.
Presence of Potentially-Toxic Microcystis Cells in the Water Column of Most Sampled Water Bodies
We next determined whether potentially-toxic Microcystis were present in the water column of the locations described above. Using the Microcystis-specific PCR assay we found that the mcyD gene (amplified using Microcystis-specific mcyD primers, see Materials and Methods) was detected in 64% (37/58) of the sampling locations (Figure 1, Table 1). Consistent amplification of mcyD genes was obtained from 10 cells/filter or more, representing an approximate limit of detection of one toxinogenic cell ml−1 depending upon the volume filtered at each sampling location (Table 1, Supplementary Figure 3A). Importantly, we re-visited nine sampling locations from which mcyD genes could not be amplified 4 years after the initial samples were taken (June 2015), collecting samples from the water column as well as the sediment. In six of these nine locations we still could not amplify the mcyD gene in the water column, with the other three locations all being part of an interconnected set of springs (Ein Afek, Table 1). Nevertheless, the mcyD gene could be amplified from all sediment samples. This suggests that, despite the presence of potentially toxic cells in the sediment, in most of these locations low densities of potentially toxinogenic strains in the water column are the norm during summer. It is tempting to speculate that these locations are inherently less hospitable to the potentially-toxic Microcystis strains.
Presence of mcyD Genes and the Relationships with Environmental Factors
Many environmental factors are known to be associated with the presence or toxicity of Microcystis blooms, chief among them the concentrations of dissolved phosphorus and inorganic nitrogen (Vézie et al., 2002; Davis et al., 2009; Xu et al., 2010; Paerl et al., 2011b; Paerl and Paul, 2012). We therefore asked whether these factors are associated not only with the blooms or toxicity but also with the presence of potentially-toxic cells in general. As shown in Figure 3A, the probability of detecting the mcyD gene was significantly higher as the concentration of dissolved phosphorous increases. Furthermore, a negative trend was observed between the probability of mcyD detection and the N:P ratio, suggesting that the availability of phosphorus, rather than nitrogen, is correlated with, and potentially drives, the distribution of the mcyD containing strains (Supplementary Figure 4A). Moreover, when the presence/absence patterns of the mcyD genes were plotted on the nMDS analysis described above (Figure 2C), the mcyD-positive water bodies tended to cluster in the same area on the nMDS plot as the agricultural waters. All of the aquaculture water and most of the irrigation reservoirs contained potentially-toxic Microcystis strains, whereas only 50–60% of the natural water bodies (e.g., lakes and springs, the latter often collected into man-made pools, Supplementary Figure 1) contained potentially toxinogenic cells in sufficient concentrations for detection by the PCR assay (Figure 3B). In order to determine whether the correlation with high phosphate concentrations is in fact due to a cross-correlation between phosphate and aquaculture (implying that something else in the aquaculture-related water is in fact responsible for the presence of potentially-toxic populations), we performed the same statistical analysis without including values from fish ponds. The results exhibited a similar trend, with mcyD still associated with high phosphorus levels (Supplementary Figure 4C).
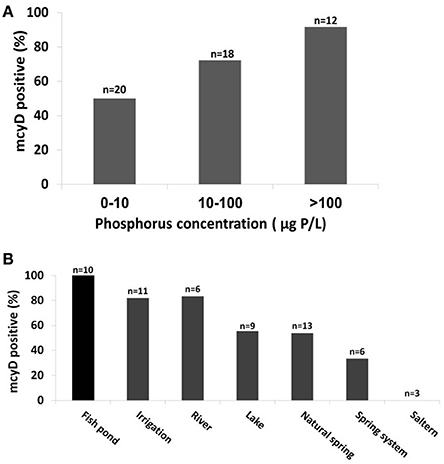
Figure 3. The probability of mcyD being detected in the water bodies increases with phosphorus concentrations and in aquaculture facilities. (A) A higher percent of mcyD gene presence was found in water with high P concentrations (a, p < 0.05). (B) Significant differences in the distribution of mcyD positive locations was observed among the different water body types (Kruskal-Wallis ANOVA analysis with p < 0.05). Note that all fish ponds contain toxinogenic Microcystis populations.
The Effect of Local and Regional Conditions on the Probability of mcyD Presence
We next asked, are there additional factors besides nutrient concentrations that are correlated with a high probability of mcyD detection? To answer this question, taking into account not only the conditions within the water body but also those prevalent in the region surrounding it (up to several tens of kilometers), we superimposed our data onto geographically resolved maps of the mean summer day temperature during August (mdt8, Figure 1A), precipitation (Figure 1B) and elevation (Supplementary Figure 1). In addition, we used a composite map describing the boundaries of four categories of land use: forests, agricultural land, built areas and nature reserves (Figure 1C, Materials and Methods). Due to the limitations of the publically-available data, the latter map contains data for only ~37% of the total analyzed areas, and we therefore used the distance from the closest representative of each of these specific land-use categories for subsequent analyses. We used logistic regression in the framework of Generalized Linear Models (GLMs) to relate both local (e.g., abiotic parameters of the water body) and regional environmental variables to the presence of mcyD genes and determine the relative importance of each variable in the final distribution model (Supplementary Tables 2–4). Despite challenges associated with relationships between the explanatory variables and the differences between the desert south of Israel and the rest of the locations (see Supplemental Methods, Supplementary Figure 5), robust inferences can be drawn from this analysis: first, three local parameters, namely pH and the concentrations of ammonia and phosphorus, were designated as important model terms (Figure 4). The pH of the water bodies and phosphorus were positively related with mcyD presence (as shown by the coefficient of these parameters in the models, Supplementary Table 3) whereas ammonia revealed a negative relationship. Second, several regional parameters were also important in these models, specifically, the distance from built areas which was always inversely related to mcyD presence, suggesting that urban runoff may be a factor contributing to the presence of potentially-toxic cells in the water (Figure 4, Supplementary Table 3). Conversely, the distance from nature reserves was positively related to mcyD presence, suggesting that water bodies found in or around nature reserves have less of a chance to contain potentially-toxic populations. The regional inferences were weaker when the samples from the desert south of Israel were not included in the analysis, potentially due to the low density of built areas and forests in this region (Supplementary Figure 5).
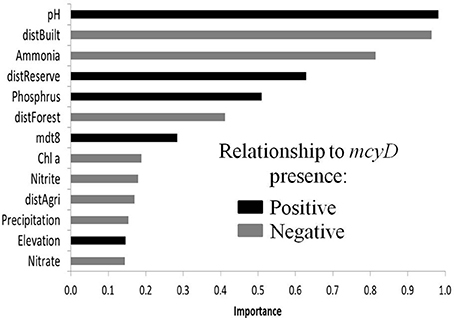
Figure 4. The relative importance of different local and regional parameters using GLM. The results of the same analysis performed with cross correlated in a distribution model of mcyD genes produced parameters removed and for locations found in the north of Israel are shown in Supplementary Table 3, and the coefficients relating each parameter to the presence/absence of mcyD are shown in Supplementary Table 4.
Phylogenetic Distribution of Potentially-Toxic Microcystis Populations among the Sampling Locations
Given the significant differences in environmental conditions between the sampling locations, we next asked whether any genetic differences could be identified between the potentially-toxic populations inhabiting each water body, using the mcyA gene as a phylogenetic marker. No clear distance-decay pattern could be observed in the similarity of the mcyA gene assemblages (e.g., as shown using automatic ribosomal intergenic spacer analysis for stream bacteria, Lear et al., 2013), suggesting that the geographic distances between the sampled sites did not strongly affect the population structure of potentially-toxic Microcystis. This could also be due to the relatively small number of sampling locations and mcyA sequences (120 sequences from 17 locations). Nevertheless, as shown in Figure 5, some mcyA clades were preferentially associated with specific water body types. Specifically, one clade of mcyA was significantly associated with aquaculture facilities and negatively related to irrigation reservoirs (Figure 5, clade 2C, p < 0.01, Fisher's exact test with Bonferroni correction). No association was observed between specific clades and the concentrations of inorganic nitrogen phosphorus or Chl a (Krusal-Wallis test, p>0.05). This suggests that the potentially-toxic strains are not randomly distributed, and that the type of the water body or its intended use, rather than trophic state or the concentrations of specific nutrients, affect this distribution.
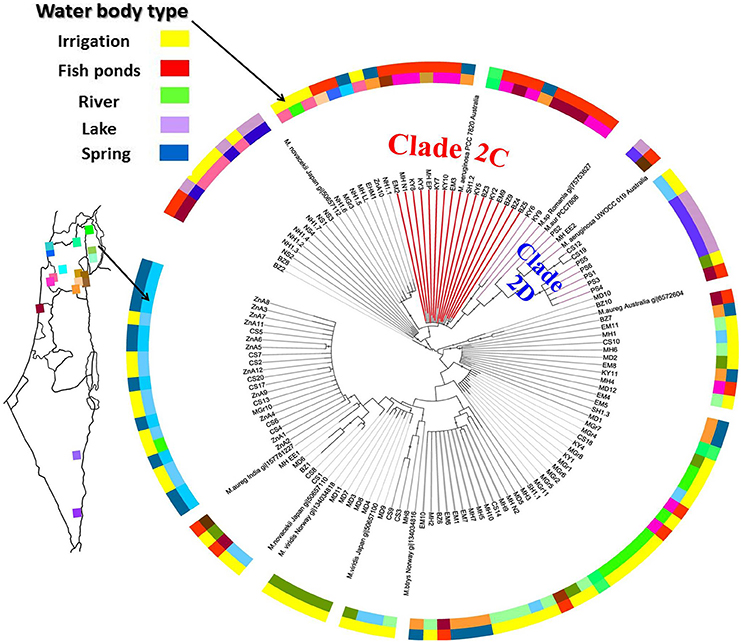
Figure 5. A specific mcyA gene lineage is enriched in fish ponds and under-represented in irrigation reservoirs. A maximum likelihood phylogenetic tree of the cloned mcyA genes, including also sequences from Microcystis isolates, is shown. Circles indicate branches with >50% bootstrap support. The tree is rooted at mid-point, sequences from other cyanobacteria (Anabaena, Nostoc, and Planktothrix) all clustered together as an outgroup (not shown). Color datasets on the rings surrounding the tree are the specific location of the sample (inner ring, with similar shades indicating geogrphically close locations) and the water body type (outer ring). Fish ponds are over-represented and irrigation reservoirs under-represented in the clade 2C (red, p < 0.01, fisher's exact test with Bonferroni correction). Lakes are over-represented in clade 2D (p < 0.05), however, most of the sequences from clade 2D came from the same lake and therefore this association requires further validation.
Discussion
What May Influence the Distribution of Potentially Toxinogenic Microcystis?
Harmful algal blooms in general, and those produced by Microcystis in particular, are on the increase. Elevated water temperature, increased salinity, stronger light intensity (especially UV), reduced mixing and higher nutrient loads may all provide a competitive advantage to Microcystis over other phytoplankton (Paerl and Huisman, 2008; Davis et al., 2009; Dziallas and Grossart, 2011; Van de Waal et al., 2011; Paerl and Paul, 2012). Many of the same environmental changes may also provide an advantage to toxic Microcystis strains (or other microcystin-producing bacteria) over non-toxic ones, although the mechanism (or the biological role of microcystins) is yet unclear (Davis et al., 2009; Dziallas and Grossart, 2011; Van de Waal et al., 2011; Kurmayer et al., 2014; Meissner et al., 2014). However, in order for a bloom to occur the water body in question must contain a viable “seed population” of toxinogenic strains, either in the water column or as resting stages in the sediment, that can respond to the favorable growth conditions. Microcystis, as a genus, has a global distribution, with strains isolated from all continents except Antarctica (van Gremberghe et al., 2011). Additionally, no clear phylogeographic patterns have been observed, suggesting few barriers to global dispersal (van Gremberghe et al., 2011; Moreira et al., 2013). Our results suggest that, at local to regional scales, potentially-toxic strains, identified through PCR of the mcyD gene, are common in the water column but not ubiquitous. Typically, planktonic Microcystis blooms are defined as >7 × 104 cells L−1 (Baxa et al., 2010), and develop over a period of several weeks, from a starting planktonic population of at least >1000 cells L−1 (Davis et al., 2009). Given the sensitivity of the end-point PCR method we used (Supplementary Figure 3) and the typical rates of bloom formation and decline in nature (e.g., Davis et al., 2009; Baxa et al., 2010), it is likely that location in which mcyD genes were not detected by the PCR method, did not experience high Microcystis densities over the period of 2–3 months prior to our sampling, nor did such blooms occur over the following 2–3 months. However, we caution that such extrapolations are fraught with uncertainty, and cases have been recorded where blooms emerged quite rapidly (e.g., at the Antioch sampling site of the San Franscisco Estuary, toxic cell equivalents increased from just below our detection limit to ~2 × 107 cells/L within less than a month and a half, Baxa et al., 2010).
The reasons for the observed “patchiness” in potentially-toxic Microcystis seems to be a complex interaction between many factors, including the intended use of the water (e.g., for aquaculture, irrigation etc.) and the location of the water body relative to urban centers and nature reserves. We suggest that these land- and water use parameters determine the concentration of inorganic nutrients and the pH of the water, which are the “proximal” drivers of potentially-toxic Microcystis distributions (Mattikalli and Richards, 1996; Crosbie and Chow-Fraser, 1999; Caccia and Boyer, 2005). As shown in Figure 4, Supplementary Figure 4B, and Supplementary Table 2, pH values and phosphorus concentrations were positively related to mcyD presence, whereas ammonia concentrations revealed a negative relationship. Inorganic phosphorus has previously been shown to be an important limiting factor for Microcystis growth (Nalewajko and Murphy, 2001; Xie et al., 2003; Paerl and Otten, 2013), and reduction of inorganic phosphorus in freshwaters has been suggested as a strategy to mitigate toxic blooms (Vézie et al., 2002). Our results extend this observation, suggesting that inorganic phosphorus also determines, to some extent, whether potentially-toxic populations inhabit a water body irrespective of whether or not they bloom. Notably, urban runoff often has very high inorganic phosphorus concentrations (Smil, 2000; Bartley et al., 2012), perhaps in part explaining why water bodies close to urban areas tended to have a higher probability of mcyD presence. In contrast, a negative correlation was observed between ammonia concentrations and mcyD presence in our dataset. High ammonia concentrations combined with strong sunlight (the latter being quite common in Israel and other semi-arid lands) have been shown to negatively affect Microcystis (Dai et al., 2012). Alternatively, the inverse correlation between mcyD presence and ammonia (as well as the inverse correlation with the dissolved ratio of nitrogen to phosphorus, Supplementary Figure 4A) may be due to nitrogen uptake by Microcystis and other phytoplankton. Unraveling the effect of different nutrients on Microcystis presence, bloom dynamics and toxicity is clearly required in order to ascertain which are the best methods to address toxic blooms (Paerl et al., 2014).
What causes some water bodies to have a higher pH, and how this relates to mcyD presence, is less clear. The measured high pH values may have been caused by CO2 depletion due to photosynthesis and thus may partly be indicative of high phytoplankton biomass (the samples were mostly collected around mid-day during the summer months). Indeed, pH and Chl a concentrations are somewhat positively correlated in our dataset (Supplementary Table 2). However, the pH of freshwater may also be affected by other parameters, such as the bedrock or sediment type and the concentrations and uptake kinetics of inorganic nitrogen compounds. The pH may also be affected by many types of organic and inorganic compounds, including some widely used by industry. Regardless of what causes the high pH, such conditions may select for Microcystis over other phytoplankton species, as they often have higher pH tolerance, and may also select for toxic over non-toxic Microcystis strains (Van de Waal et al., 2011).
An intriguing result of our analysis is that the distance from agricultural land does not emerge as an important predictor of mcyD presence in our dataset. Several studies have suggested that freshwater bodies with catchment areas comprising a high percentage of agricultural land tend to have higher cyanobacterial biomass (Katsiapi et al., 2012) as well as potentially higher microcystin levels (Beaver et al., 2014). Such a link between regional land use and algal or cyanobacterial biomass, however, may depend on the connectivity of the water bodies (i.e., to other water bodies through streams or rivers, Catherine et al., 2008; Lear et al., 2013) and on the ratio of the water body volume and the catchment area (Katsiapi et al., 2012). In Israel, most of the water bodies are isolated (not connected to a network). Additionally, many of the sampled water bodies receive multiple inputs, including precipitation runoff, groundwater, waste-water after different levels of treatment and spring water. These aspects may explain why agricultural land use seems to be not important for mcyD presence in our dataset. While our sampling was limited to Israel, a combination of similar hydrology and the presence of toxic Microcystis blooms is observed in many other Mediterranean and semi-arid regions, including much of the Middle East, Greece (e.g., Gkelis et al., 2015), Sicily (Naselli-Flores et al., 2007), Spain (Asencio, 2013), and parts of Australia (Sinang et al., 2013). Further, research is needed to determine to what extent our results can be generalized to these climatically- and hydrologically- similar regions.
Importantly, in this study we considered only the planktonic cells found in the surface water next to the shore, however, spatial heterogeneity in the distribution of phytoplankton may occur within a single water body, especially with floating organisms such as Microcystis which are strongly affected by wind conditions. Moreover, Microcystis may also survive extended periods of adverse conditions as dormant resting stages in the sediment, rapidly reviving and contributing to blooms (Ståhl-Delbanco et al., 2003; Cirés et al., 2013). Nevertheless, most of the locations that were mcyD-negative remained so 4 years later, suggesting that some locations are inherently less hospitable to the potentially-toxic Microcystis strains. Additional studies with better resolved land-use maps, sampling regimes incorporating also the sediment and following the same locations over time are required in order to determine how stable the observed patterns are.
Do Niche Separation, Non-random Dispersal or Biotic Interactions Underlie the Preference of a Toxinogenic Microcystis Clade to Fish Ponds?
Microcystis is one of several globally abundant freshwater cyanobacteria that are able to colonize and flourish in a wide range of habitats (Wilson et al., 2005; Fan and Wu, 2012). In the model marine cyanobacterium Prochlorococcus, genetically and physiologically different ecotypes have evolved to live under different conditions, for example under high- and low light conditions (Biller et al., 2014). In contrast, previous studies have suggested that Microcystis populations are not organized in ecotypes (van Gremberghe et al., 2011; Humbert et al., 2013; Moreira et al., 2013), and that their success may be attributed to a large and highly plastic genomes. Such genomes encode many regulatory and metabolic genes, enabling rapid acclimation and adaptation to fluctuating conditions in “unstable” freshwater environments (Humbert et al., 2013). Nevertheless, we have observed that at least one clade of Microcystis strains, defined using the mcyA gene sequence, preferentially inhabits fish ponds and is under-represented in irrigation reservoirs (Figure 5). It is tempting to speculate that this clade is specifically adapted to fish ponds, representing the first example of niche specialization in Microcystis. However, other explanations may be given, for example, that the association of this clade with fish ponds is due to non-random dispersal. Specifically, most of the Israeli aquaculture system relies on hatching and growth of juvenile fish in a small number of dedicated facilities, with the fish then distributed as fingerlings to fish ponds. This could enable concurrent transfer of the hatcheries microbiota, including Microcystis, to the open fish ponds. Another potential vector for non-random Microcystis dispersal is with migratory birds, many of which visit multiple fish ponds on their annual return route from Africa to Europe (van Leeuwen et al., 2012; Moreira et al., 2014). The observed patterns could also represent historical distribution patterns, because established populations often have a competitive advantage over newly introduced migrants, a phenomenon termed the “priority effect” (Van Gremberghe et al., 2009). Finally, other biotic factors associated with fish ponds may serve to control the distribution of Microcystis. For example, some species of the dinoflagellate Peridinium were documented inhibiting Microcystis growth and reproduction (Wu et al., 1998; Vardi et al., 2002), and heterotrophic bacterial communities may also affect the structure of Microcystis populations (Choi et al., 2005; Shen et al., 2011; Zhang et al., 2011). Long term analysis of the presence and abundance of toxinogenic Microcystis species, combined with whole-population 16S and 18S analyses, may reveal co-occurrence patterns between Microcystis and other micro-organisms that will help test these hypotheses. In parallel, isolation and characterization of fish-pond associated strains may help determine whether such strains have growth advantages under conditions commonly found in fish ponds, or whether the changes in mcyA gene sequence are associated with differences in the microcystin molecule, as previously shown for the mcyA gene (Allender et al., 2009). Our observations of a clade of Microcystis associated with fish ponds could not be expanded using mcyA sequences found in public databases, since the vast majority of these sequences are not associated with any metadata describing the location from which they were isolated. This highlights the need for better association between sequence and environmental data in order to facilitate our understanding of the factors controlling the distribution and abundance of microbes in nature.
As the Earth's climate, population, water-, and land-use rapidly change, predicting when and where aquatic microorganisms, including Microcystis and other toxic phytoplankton, will live and bloom is critical in order to facilitate the design of cost-effective monitoring systems and management approaches to reduce human and environmental exposure. Integrating our understanding of the biology and ecology of Microcystis and other aquatic microbes into a framework that encompasses geography, land, and water-use (Catherine et al., 2008; Lear et al., 2013) is critical as we inch forward to understand how water resources and their biotic communities are shaped under anthropogenic pressures.
Author Contributions
SM and DS designed the research, SM, DA, MG, YY, and DS performed field sampling, SM, DA, and YY analyzed samples, LB formulated and performed the GLM analyses and MG performed the multivariate analyses. All authors analyzed the data and wrote the manuscript.
Funding
This study was supported in part by grant 3-10342 from the Israel Ministry of Science and Technology and by Marie Curie Career Integration Grant MICROBES-2-MODEL.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Hans-Peter Grossart and Assaf Sukenik for providing toxinogenic and non-toxinogenic strains and for helpful discussions. We also thank Tamar Rachamim, Eliezra Glasser, Elad Rachmilovich, Omri Shabbat, and Sophia Barinova for help with sampling, and many water managers for sampling access and helpful insights. Sampling was performed in accordance with permit number 40258 from the Israel National Parks Authority.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.00271
References
Allender, C. J., LeCleir, G. R., Rinta-Kanto, J. M., Small, R. L., Satchwell, M. F., Boyer, G. L., et al. (2009). Identifying the source of unknown microcystin gens and predicting microcystin variants by linking multiple genes within uncultured cyanobacterial cells. Appl. Environ. Microbiol. 75, 3598–3604. doi: 10.1128/AEM.02448-08
APHA (2005). Standard Methods for Examination of Water and Waste Water, 21st Edn. Washington, DC: American Public Health Association.
Asencio, A. D. (2013). Determination of microcystins in reservoirs of different basins in a semiarid area. Appl. Phycol. 25, 1753–1762. doi: 10.1007/s10811-013-0025-4
Bartley, R., Speirs, W. J., Ellis, T. W., and Waters, D. K. (2012). A review of sediment and nutrient concentration data from Australia for use in catchment water quality models. Mar. Pollut. Bull. 65, 101–116. doi: 10.1016/j.marpolbul.2011.08.009
Baxa, D. V., Kurobe, T., Ger, K. A., Lehman, P. W., and Teh, S. J. (2010). Estimating the abundance of toxic Microcystis in the San Francisco Estuary using quantitative real-time PCR. Harmful Algae 9, 342–349. doi: 10.1016/j.hal.2010.01.001
Beaver, J. R., Manis, E. E., Loftin, K. A., Graham, J. L., Pollard, A. I., and Mitchell, R. M. (2014). Land use patterns, ecoregion, and microcystin relationships in U.S. lakes and reservoirs: a preliminary evaluation. Harmful Algae 36, 57–62. doi: 10.1016/j.hal.2014.03.005
Biller, S. J., Berube, P. M., Lindell, D., and Chisholm, S. W. (2014). Prochlorococcus: the structure and function of collective diversity. Nat. Rev. Micro. 13, 13–27. doi: 10.1038/nrmicro3378
Brianda, J. F., Jacqueta, S., Bernardb, C., and Humberta, J. F. (2003). Health hazards for terrestrial vertebrates from toxic cyanobacteria in surface water ecosystems. Vet. Res. 34, 361–377. doi: 10.1051/vetres:2003019
Caccia, V. G., and Boyer, J. N. (2005). Spatial patterning of water quality in Biscayne Bay, Florida as a function of land use and water management. Mar. Pollut. Bull. 50, 1416–1429. doi: 10.1016/j.marpolbul.2005.08.002
Campos, A., and Vasconcelos, V. (2010). Molecular mechanisms of microcystin toxicity in animal cells. Int. J. Mol. Sci. 11, 268–287. doi: 10.3390/ijms11010268
Carmichael, W. W. (2001). Health effects of toxin-producing cyanobacteria: “The CyanoHABs”. Hum. Ecol. Risk Assess. 7, 1393–1407. doi: 10.1080/20018091095087
Carmichael, W. W., Azevedo, S. M., An, J. S., Molica, R. J., Jochimsen, E. M., Lau, S., et al. (2001). Human fatalities from cyanobacteria: chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 109, 663–668. doi: 10.1289/ehp.01109663
Catherine, A., Troussellier, M., and Bernard, C. (2008). Design and application of a stratified sampling strategy to study the regional distribution of cyanobacteria (Ile-de-France, France). Water Res. 42, 4989–5001. doi: 10.1016/j.watres.2008.09.028
Choi, H., Kim, B., Kim, J., and Han, M. (2005). Streptomyces neyagawaensis as a control for the hazardous biomass of Microcystis aeruginosa (Cyanobacteria) in eutrophic freshwaters. Biol. Control 33, 335–343. doi: 10.1016/j.biocontrol.2005.03.007
Cirés, S., Wörmer, L., Agha, R., and Quesada, A. (2013). Overwintering populations of Anabaena, Aphanizomenon and Microcystis as potential inocula for summer blooms. J. Plankton Res. 35, 1254–1266. doi: 10.1093/plankt/fbt081
Crosbie, B., and Chow-Fraser, P. (1999). Percentage land use in the watershed determines the water and sediment quality of 22 marshes in the Great Lakes basin. Can. J. Fish. Aquat. Sci. 56, 1781–1791. doi: 10.1139/f99-109
Dai, G. Z., Shang, J. L., and Qiu, B. S. (2012). Ammonia may play an important role in the succession of cyanobacterial blooms and the distribution of common algal species in shallow freshwater lakes. Global Change Biol. 18, 1571–1581. doi: 10.1111/j.1365-2486.2012.02638.x
Davis, T. W., Berry, D. L., Boyer, G. L., and Gobler, C. J. (2009). The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 8, 715–725. doi: 10.1016/j.hal.2009.02.004
Dittmann, E., and Börner, T. (2005). Genetic contributions to the risk assessment of microcystin in the environment. Toxicol. Appl. Pharmacol. 203, 192–200. doi: 10.1016/j.taap.2004.06.008
Dittmann, E., and Wiegand, C. (2006). Cyanobacterial toxins – occurrence, biosynthesis and impact on human affairs. Mol. Nutr. Food Res. 50, 7–17. doi: 10.1002/mnfr.200500162
Dziallas, C., and Grossart, H. P. (2011). Increasing oxygen radicals and water temperature select for toxic Microcystis sp. PLoS ONE 6:e25569. doi: 10.1371/journal.pone.0025569
Falconer, I. R., and Humpage, A. R. (2005). Health risk assessment of cyanobacterial (Blue-green Algal) toxins in drinking water. Int. J. Environ. Res. Public Health 2, 43–50. doi: 10.3390/ijerph2005010043
Fan, X., and Wu, Q. L. (2012). Intra-habitat differences in the composition of the methanogenic archaeal community between the Microcystis-dominated and the macrophyte-dominated bays in Taihu Lake. Geomicrobiol. J. 31, 907–916. doi: 10.1080/01490451.2012.737091
Frank, J. A., Reich, C. I., Sharma, S., Weisbaum, J. S., Wilson, B. A., and Olsen, G. J. (2008). Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 74, 2461–2470. doi: 10.1128/AEM.02272-07
Gkelis, S., Lanaras, T., and Sivonen, K. (2015). Cyanobacterial toxic and bioactive peptides in freshwater bodies of Greece: concentrations, occurrence patterns, and implications for human health. Mar. Drugs 13, 6319–6335. doi: 10.3390/md13106319
Green, J. L., Bohannan, B. J. M., and Whitaker, R. J. (2008). Microbial biogeography: from taxonomy to traits. Science 320, 1039–1043. doi: 10.1126/science.1153475
Hernández, J. M., López-Rodas, V., and Costas, E. (2009). Microcystins from tap water could be a risk factor for liver and colorectal cancer: a risk intensified by global change. Med. Hypotheses 72, 539–540. doi: 10.1016/j.mehy.2008.11.041
Humbert, J. F., Barbe, V., Latifi, A., Gugger, M., Calteau, A., Coursin, T., et al. (2013). A tribute to disorder in the genome of the bloom-forming freshwater cyanobacterium Microcystis aeruginosa. PLoS ONE 8:e70747. doi: 10.1371/journal.pone.0070747
Kaebernick, M., Neilan, B. A., Börner, T., and Dittmann, E. (2000). Light and the transcriptional response of the microcystin biosynthesis gene cluster. Appl. Environ. Microbiol. 66, 3387–3392. doi: 10.1128/AEM.66.8.3387-3392.2000
Karl, D., and Tien, G. (1992). MAGIC: a sensitive and precise method for measuring dissolved phosphorus in aquatic environments. Limnol. Oceanogr. 37, 105–116. doi: 10.4319/lo.1992.37.1.0105
Katsiapi, M., Mazaris, A. D., Charalampous, E., and Moustaka-Gouni, M. (2012). Watershed land use types as drivers of freshwater phytoplankton structure. Hydrobiologia 698, 121–131. doi: 10.1007/s10750-012-1095-z
Kurmayer, R., Blom, J. F., Deng, L., and Pernthaler, J. (2014). Integrating phylogeny, geographic niche partitioning and secondary metabolite synthesis in bloom-forming Planktothrix. ISME J. 9, 909–921. doi: 10.1038/ismej.2014.189
Kurmayer, R., Schober, E., Tonk, L., Visser, P. M., and Christiansen, G. (2011). Spatial divergence in the proportions of genes encoding toxic peptide synthesis among populations of the cyanobacterium Planktothrix in European lakes. FEMS Microbiol. Lett. 317, 127–137. doi: 10.1111/j.1574-6968.2011.02222.x
Lear, G., Washington, V., Neale, M., Case, B., Buckley, H., and Lewis, G. (2013). The biogeography of stream bacteria. Global Ecol. Biogeogr. 22, 544–554. doi: 10.1111/geb.12046
Letunic, I., and Bork, P. (2011). Interactive tree of life v2: online annotation and display of phylogenetic trees made easy. Nucleic Acids Res. 39, 475–478. doi: 10.1093/nar/gkr201
Markowitz, V. M., Chen, I. M. A., Palaniappan, K., Chu, K., Szeto, E., Pillay, M., et al. (2014). IMG 4 version of the integrated microbial genomes comparative analysis system. Nucleic Acids Res. 42, 560–567. doi: 10.1093/nar/gkt963
Massana, R., Murray, A. E., Preston, C. M., and DeLong, E. F. (1997). Vertical distribution and phylogenetic characterization of marine planktonic Archaea in the Santa Barbara Channel. Appl. Environ. Microbiol. 63, 50–56.
Mattikalli, N. M., and Richards, K. S. (1996). Estimation of surface water quality changes in response to land use change: application of the export coefficient model using remote sensing and Geographical Information System. J. Environ. Manage. 48, 263–282. doi: 10.1006/jema.1996.0077
Meissner, S., Steinhauser, D., and Dittmann, E. (2014). Metabolomic analysis indicates a pivotal role of the hepatotoxin microcystin in high light adaptation of Microcystis. Environ. Microbiol. 17, 1497–1509. doi: 10.1111/1462-2920.12565
Moreira, C., Vasconcelos, V., and Antunes, A. (2013). Phylogeny of microcystins: evidence of a biogeographical trend? Curr. Microbiol. 66, 214–221. doi: 10.1007/s00284-012-0258-5
Moreira, C., Spillane, C., Fathalli, A., Vasconcelos, V., and Antunes, A. (2014). African origin and Europe-Mediated global dispersal of The cyanobacterium Microcystis aeruginosa. Curr. Microbiol. 69, 628–633. doi: 10.1007/s00284-014-0628-2
Nalewajko, C., and Murphy, T. P. (2001). Effects of temperature, and availability of nitrogen and phosphorus on the abundance of Anabaena and Microcystis in Lake Biwa, Japan: an experimental approach. Limnology 2, 45–48. doi: 10.1007/s102010170015
Naselli-Flores, L., Barone, R., Chorus, I., and Kurmayer, R. (2007). Toxic cyanobacterial blooms in reservoirs under a semiarid mediterranean climate: the magnification of a problem. Environ. Toxicol. 22, 399–404. doi: 10.1002/tox.20268
Nübel, U., Garcia-Pichel, F., and Muyzer, G. (1997). PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 63, 3327–3332.
O'Neil, J. M., Davis, T. W., Burford, M. A., and Gobler, C. J. (2012). The rise of harmful cyanobacteria blooms: the potential roles of eutrophication and climate change. Harmful Algae 14, 313–334. doi: 10.1016/j.hal.2011.10.027
Padilla, D. K., and Williams, S. L. (2004). Beyond ballast water: aquarium and ornamental trades as sources of invasive species in aquatic ecosystems. Front. Ecol. Environ. 2, 131–138. doi: 10.1890/1540-9295(2004)002[0131:BBWAAO]2.0.CO;2
Paerl, H., and Huisman, J. (2009). Climate change: a catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 1, 27–37. doi: 10.1111/j.1758-2229.2008.00004.x
Paerl, H. W., and Huisman, J. (2008). Blooms like it hot. Science 320, 57–58. doi: 10.1126/science.1155398
Paerl, H. W., Hall, N. S., and Calandrino, E. S. (2011a). Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total Environ. 409, 1739–1745. doi: 10.1016/j.scitotenv.2011.02.001
Paerl, H. W., Xu, H., McCarthy, M. J., Zhu, G., Qin, B., Li, Y., et al. (2011b). Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): the need for a dual nutrient (N & P) management strategy. Water Res. 45, 1973–1983. doi: 10.1016/j.watres.2010.09.018
Paerl, H. W., and Paul, V. J. (2012). Climate change: links to global expansion of harmful cyanobacteria. Water Res. 46, 1349–1363. doi: 10.1016/j.watres.2011.08.002
Paerl, H. W., and Otten, T. G. (2013). Harmful Cyanobacterial Blooms: causes, consequences, and controls. Environ. Microbiol. 65, 995–1010. doi: 10.1007/s00248-012-0159-y
Paerl, H. W., Gardner, W. S., McCarthy, M. J., Peierls, B. L., and Wilhelm, S. W. (2014). Algal blooms: noteworthy nitrogen. Science 346:175. doi: 10.1126/science.346.6206.175-a
Pearson, L. A., Hisbergues, M., Börner, T., Dittmann, E., and Neilan, B. A. (2004). Inactivation of an ABC transporter gene, mcyH, results in loss of microcystin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Appl. Environ. Microbiol. 70, 6370–6378. doi: 10.1128/AEM.70.11.6370-6378.2004
Pouria, S., Andrade, A. D., Barbosa, J., Cavalcanti, R. L., Barreto, V. T. S., Ward, C. J., et al. (1998). Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 352, 21–26. doi: 10.1016/S0140-6736(97)12285-1
Rinta-Kanto, J. M., Ouellette, A. J. A., Boyer, G. L., Twiss, M. R., Bridgeman, T. B., and Wilhelm, S. W. (2005). Quantification of Toxic Microcystis spp. during the 2003 and 2004 Blooms in Western Lake Erie using Quantitative Real-Time PCR. Environ. Sci. Technol. 39, 4198–4205. doi: 10.1021/es048249u
Ritchie, R. J. (2008). Universal chlorophyll equations for estimating chlorophylls a, b, c, and d and total chlorophylls in natural assemblages of photosynthetic organisms using acetone, methanol, or ethanol solvents. Photosynthetica 46, 115–126. doi: 10.1007/s11099-008-0019-7
Schinder, D. W., and Valentyne, J. R. (2008). The Algal Bowl: Overfertilization of the World's Freshwater and Estuaries. Edmonton, AB: The University of Alberta Press.
Shen, H., Niu, Y., Xie, P., Tao, M. I. N., and Yang, X. I. (2011). Morphological and physiological changes in Microcystis aeruginosa as a result of interactions with heterotrophic bacteria. Freshw. Biol. 56, 1065–1080. doi: 10.1111/j.1365-2427.2010.02551.x
Sinang, S. C., Reichwaldt, E. S., and Ghadouani, A. (2013). Spatial and temporal variability in the relationship between cyanobacterial biomass and microcystins. Environ. Monit. Assess. 185, 6379–6395. doi: 10.1007/s10661-012-3031-0
Smil, V. (2000). Phosphorus in the environment: natural flows and human interferences. Annu. Rev. Ener. Environ. 25, 53–88. doi: 10.1146/annurev.energy.25.1.53
Stone, R. (2011). China aims to turn tide against toxic lake pollution. Science 333, 1210–1211. doi: 10.1126/science.333.6047.1210
Ståhl-Delbanco, A., Hansson, L. A., and Gyllström, M. (2003). Recruitment of resting stages may induce blooms of Microcystis at low N:P ratios. J. Plankton Res. 25, 1099–1106. doi: 10.1093/plankt/25.9.1099
Sukenik, A., Hadas, O., Kaplan, A., and Quesada, A. (2012). Invasion of Nostocales (cyanobacteria) to subtropical and temperate freshwater lakes – physiological, regional, and global driving forces. Front. Microbiol. 3:86. doi: 10.3389/fmicb.2012.00086
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
Tanabe, Y., Sano, T., Kasai, F., and Watanabe, M. (2009). Recombination, cryptic clades and neutral molecular divergence of the microcystin synthetase (mcy) genes of toxic cyanobacterium Microcystis aeruginosa. BMC Evol. Biol. 9:115. doi: 10.1186/1471-2148-9-115
Tillett, D., Parker, D. L., and Neilan, B. A. (2001). Detection of toxigenicity by a probe for the microcystin synthetase a gene (mcyA) of the cyanobacterial genus Microcystis: comparison of toxicities with 16S rRNA and phycocyanin operon (phycocyanin intergenic spacer) phylogenies. Appl. Environ. Microbiol. 67, 2810–2818. doi: 10.1128/AEM.67.6.2810-2818.2001
Tzahor, S., Man-Aharonovich, D., Kirkup, B., Yogev, T., Berman-Frank, I., Polz, M., et al. (2009). A supervised learning approach for taxonomic classification of core-photosystem-II genes and transcripts in the marine environment. BMC Genomics 10:229. doi: 10.1186/1471-2164-10-229
Van de Waal, D. B., Verspagen, J. M. H., Finke, J. F., Vournazou, V., Immers, A. K., Kardinaal, W. E. A., et al. (2011). Reversal in competitive dominance of a toxic versus non-toxic cyanobacterium in response to rising CO2. ISMEJ. 5, 1438–1450. doi: 10.1038/ismej.2011.28
Van Gremberghe, I., Vanormelingen, P., Van der Gucht, K., Souffreau, C., Vyverman, W., and De Meester, L. (2009). Priority effects in experimental populations of the cyanobacterium Microcystis. Environ. Microbiol. 11, 2564–2573. doi: 10.1111/j.1462-2920.2009.01981.x
van Gremberghe, I., Leliaert, F., Mergeay, J., Vanormelingen, P., Van der Gucht, K., Debeer, A., et al. (2011). Lack of phylogeographic structure in the freshwater cyanobacterium Microcystis aeruginosa suggests global dispersal. PLoS ONE 6:e19561. doi: 10.1371/journal.pone.0019561
van Leeuwen, C. H. A., van der Velde, G., van Groenendael, J. M., and Klaassen, M. (2012). Gut travellers: internal dispersal of aquatic organisms by waterfowl. J. Biogeogr. 39, 2031–2040. doi: 10.1111/jbi.12004
Valério, E. (2010). Diversity and impact of prokaryotic toxins on aquatic environments: a review. Toxins 2, 2359–2410. doi: 10.3390/toxins2102359
Vardi, A., Schatz, D., Beeri, K., Motro, U., Sukenik, A., Levine, A., et al. (2002). Dinoflagellate-cyanobacterium communication may determine the composition of phytoplankton assemblage in a mesotrophic lake. Curr. Biol. 12, 1767–1772. doi: 10.1016/S0960-9822(02)01217-4
Vasconcelos, V., Martinis, A., Vale, M., Antunes, A., Azevedo, J., Welker, M., et al. (2010). First report on the occurrence of microcystins in planktonic cyanobacteria from Central Mexico. Toxicon 56, 425–431. doi: 10.1016/j.toxicon.2010.04.011
Vézie, C., Rapala, J., Vaitomaa, J., Seitsonen, J., and Sivonen, K. (2002). Effect of nitrogen and phosphorus on growth of toxic and nontoxic Microcystis strains and on intracellular microcystin concentrations. Microb. Ecol. 43, 443–454. doi: 10.1007/s00248-001-0041-9
Warwick, R. M., and Clarke, K. R. (1993). Comparing the severity of disturbance: a meta-analysis of marine macrobenthic community data. Mar. Ecol. Prog. Ser. 92, 221–231. doi: 10.3354/meps092221
Wilson, A. E., Sarnelle, O., Neilan, B. A., Salmon, T. P., Gehringer, M. M., and Hay, M. E. (2005). Genetic variation of the bloom-forming cyanobacterium Microcystis aeruginosa within and among Lakes: implications for harmful algal blooms. Appl. Environ. Microbiol. 71, 6126–6133. doi: 10.1128/AEM.71.10.6126-6133.2005
Wu, J. T., Kuo-Huang, L. L., and Lee, J. (1998). Algicidal effect of peridinium bipes on Microcystis aeruginosa. Curr. Microbiol. 37, 257–261. doi: 10.1007/s002849900375
Xie, L., Xie, P., Li, S., Tang, H., and Liu, H. (2003). The low TN:TP ratio, a cause or a result of Microcystis blooms? Water Res. 37, 2073–2080. doi: 10.1016/S0043-1354(02)00532-8
Xu, H., Paerl, H. W., Qin, B., Zhu, G., and Gaoa, G. (2010). Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China. Limnol. Oceanogr. 55, 420–432. doi: 10.4319/lo.2010.55.1.0420
Keywords: cyanobacteria, distribution, mcyD, mcyA, microcystins, Microcystis
Citation: Marmen S, Aharonovich D, Grossowicz M, Blank L, Yacobi YZ and Sher DJ (2016) Distribution and Habitat Specificity of Potentially-Toxic Microcystis across Climate, Land, and Water Use Gradients. Front. Microbiol. 7:271. doi: 10.3389/fmicb.2016.00271
Received: 15 November 2015; Accepted: 19 February 2016;
Published: 15 March 2016.
Edited by:
Anton F. Post, University of Rhode Island, USAReviewed by:
Bas Ibelings, University of Geneva, SwitzerlandGeorge S. Bullerjahn, Bowling Green State University, USA
Copyright © 2016 Marmen, Aharonovich, Grossowicz, Blank, Yacobi and Sher. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel J. Sher, ZHNoZXJAdW5pdi5oYWlmYS5hYy5pbA==