 Gang-Ming Xu
Gang-Ming Xu- Key Laboratory of Experimental Marine Biology, Institute of Oceanology, Chinese Academy of Sciences, Qingdao, China
Cell–cell communications, known as quorum sensing (QS) in bacteria, involve the signal molecules as chemical languages and the corresponding receptors as transcriptional regulators. In Gram-negative bacteria, orphan LuxR receptors recognize signals more than just acylhomoserine lactones, and modulate interspecies and interkingdom communications. Whereas, in the Gram-positive Streptomyces, pseudo gamma-butyrolactones (GBLs) receptors bind antibiotics other than GBL signals, and coordinate antibiotics biosynthesis. By interacting with structurally diverse molecules like antibiotics, the TetR family receptors regulate multidrug resistance (MDR) by controlling efflux pumps. Antibiotics at subinhibitory concentration may act as signal molecules; while QS signals also have antimicrobial activity at high concentration. Moreover, the QS and MDR systems may share the same exporters to transport molecules. Among these orphan LuxR, pseudo GBL receptors, and MDR regulators, although only with low sequence homology, they have some structure similarity and function correlation. Therefore, perhaps there might be evolutionary relationship and biological relevance between the regulatory systems of QS and MDR. Since the QS systems become new targets for antimicrobial strategy, it would expand our understanding about the evolutionary history of these regulatory systems.
Introduction
Cell–cell communications among bacteria play vital roles for their adaption and survival in the environment (Waters and Bassler, 2005). Bacteria use chemical languages (signal molecules) and corresponding receptors (transcriptional regulators) during communications (Bassler and Losick, 2006). The signal molecules are structurally diverse, including acylhomoserine lactones (AHLs), gamma-butyrolactones (GBLs), and antibiotics (Ryan and Dow, 2008). Their corresponding receptors are usually transcriptional regulators, like LuxR, GBL receptors, and TetR family (Figure 1). They constitute widespread signal transduction systems, such as quorum sensing (QS) system, multidrug resistance (MDR) system, and two-component regulatory system (Ramos et al., 2005). These regulatory systems all have three essential components: signal molecule, synthase, and corresponding receptor (Cuthbertson and Nodwell, 2013).
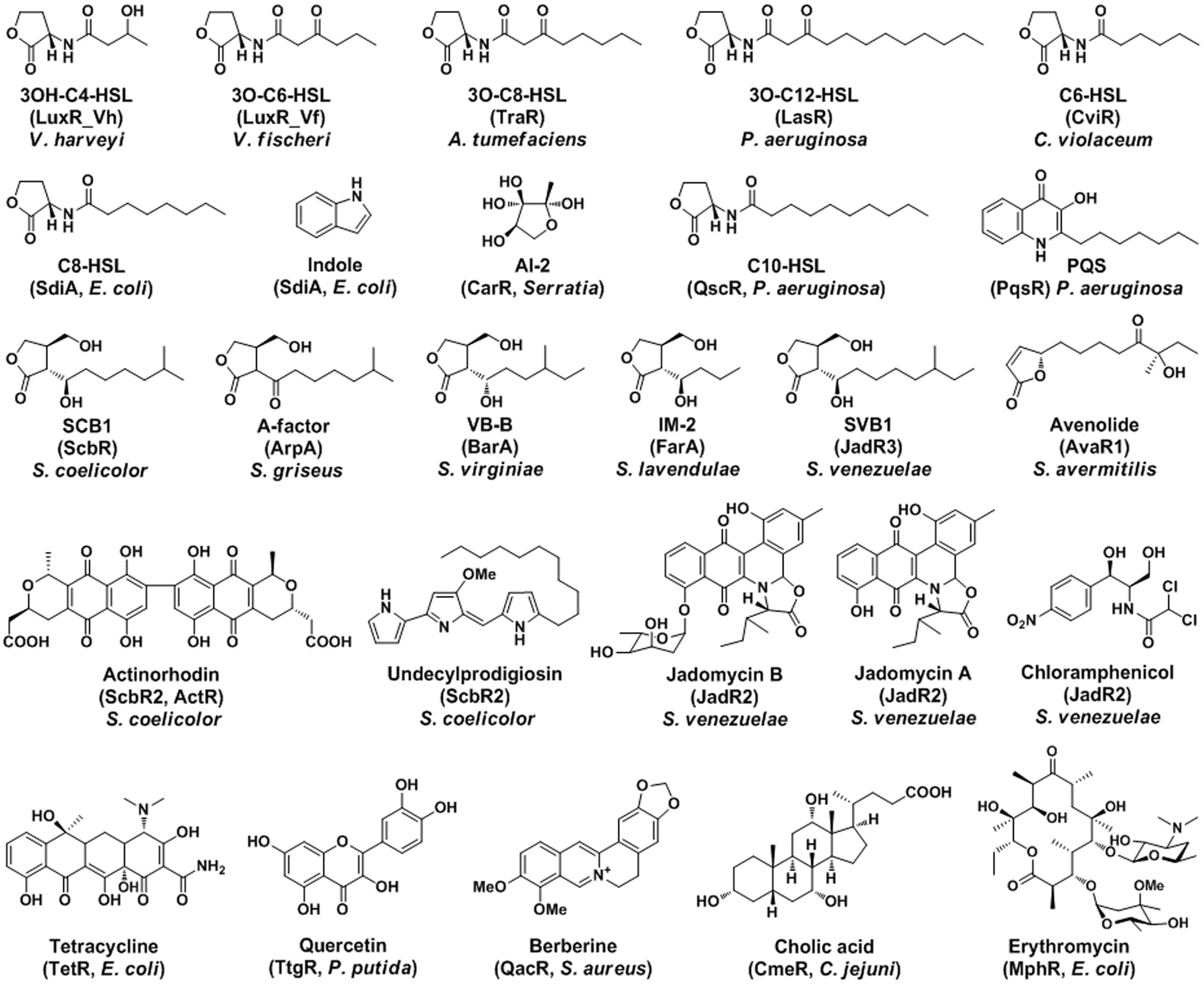
FIGURE 1. The signal molecules and the corresponding receptors of QS and MDR in bacteria. The representative ligands of transcriptional regulators are shown with chemical structures.
In Gram-negative bacteria, the autoinducer AHLs interact with cognate LuxR, and coordinate the bacterial quorum behaviors (Camilli and Bassler, 2006). The QS regulatory system has specific AHL synthase-receptor pairs, which possibly coevolved as functional cassettes (Gray and Garey, 2001; Lerat and Moran, 2004). However, some orphan LuxR receptors, which are not paired with synthase LuxI, have been identified later (Patankar and Gonzalez, 2009). Those LuxR homologues appeared to have different origins, which may due to horizontal gene transfer. They form various regulatory cascades and extend the bacterial regulatory networks (Patankar and Gonzalez, 2009; Subramoni and Venturi, 2009).
In Gram-positive Streptomyces, GBLs are used as signal molecules. By interacting with cognate receptors, GBLs activate the biosynthesis of secondary metabolites (Takano, 2006). In the GBL signaling system, cognate GBL receptors bind specific GBL molecule as ligand. The combination pairs of GBL synthase-receptor may have changed during evolution (Nishida et al., 2007). There are also some pseudo GBL receptors, whose coding genes are not adjacent to GBL synthase, and can not bind GBL molecules (Kitani et al., 2008). Streptomyces may reconstruct the regulatory system of secondary metabolisms for the adaption in complex habitats (Biarnes-Carrera et al., 2015).
The MDR regulatory system involves TetR family transcriptional regulators and multidrug efflux pumps (Paulsen, 2003). The MDR regulators can respond to structurally diverse compounds including antibiotics (Cuthbertson and Nodwell, 2013). Antibiotics abuse may lead to more efflux pumps, which expel various molecules (Nikaido, 2009). These molecules could act as antibiotics at high concentration, while as signals at low concentration (Romero et al., 2011). Moreover, the QS and MDR regulatory systems may share the same efflux pumps to transport molecules (Piddock, 2006; Martinez et al., 2009).
Evolution of the Transcriptional Regulators of QS and MDR
The orphan LuxR homologues and their functional characterizations in Gram-negative bacteria have been summarized previously (Patankar and Gonzalez, 2009). Based on the crystal structures and regulatory roles, the LuxR homologues include two families: family-A (LuxR_Vf, TraR, SdiA, CviR, LasR, and QscR) and family-B (LuxR_Vh, HapR, and SmcR; Kim et al., 2010; Lintz et al., 2011). The two LuxR families, without virtually homology, may have different evolutionary history (Lerat and Moran, 2004).
The GBL signals and cognate receptors are commonly used in the Gram-positive Streptomyces. Some pseudo GBL receptors, such as BarB, FarR2, JadR2, and ScbR2, were also reported (Nishida et al., 2007). These pseudo GBL receptors not only have high sequence homology with the genuine GBL receptors (ArpA, BarA, FarA, and ScbR), but also show similarity with the MDR regulators (Cuthbertson and Nodwell, 2013). Only the crystal structure of CprB (Streptomyces coelicolor) was determined and found to be highly similar with these MDR regulators (TtgR, QacR, and TetR; Bhukya et al., 2014).
TetR family regulators typically act as transcriptional repressors and involve in many biological processes, such as antibiotics resistance, cell–cell communication, and regulation of metabolisms (Ramos et al., 2005). Phylogenetic analyses of the TetR family regulators suggest that these GBL receptors, pseudo GBL receptors, LuxR homologues, and MDR regulators may cluster separately (Nishida et al., 2007; Cuthbertson and Nodwell, 2013). The chemical structures of representative ligands for these transcriptional regulators are also similar within each group accordingly (Figure 1). The ligand-binding specificity is in good agreement with these clustered receptors. The LuxR homologues receptors only recognize AHLs in Gram-negative bacteria; while the GBL receptors only bind GBLs in Gram-positive Streptomyces. However, some orphan LuxR and pseudo GBL receptors recognize more than just QS signals; and the MDR regulators accept even more diverse molecules. These regulators clustered together may have high sequence homology, structure similarity, and function relevance, which might suggest some evolutionary relationships (Cuthbertson and Nodwell, 2013).
Many crystal structures of the TetR family proteins have been determined, such as TtgR, QacR, CprB, HapR, and SmcR, whose overall structures are highly similar (Schumacher et al., 2001; Alguel et al., 2007; De Silva et al., 2007; Kim et al., 2010; Bhukya et al., 2014). Structure-based multiple sequence alignment analyses suggest that these proteins have similar secondary structures, with the helix-turn-helix motif highly conserved (Ramos et al., 2005). The N-terminal DNA-binding domain has relative high conservation for specific promoters; whereas the C-terminal ligand-binding domain has more variations for diverse ligands (Nishida et al., 2007; Yu et al., 2010). The ligand-binding pocket of QS regulators is very small to ensure its binding specificity; while that of the MDR regulators is relatively large to accommodate various molecules. Structural conservation of the TetR family regulators might determine their biological relevance (Cuthbertson and Nodwell, 2013).
Relationships Between the Regulatory Systems of QS and MDR
In Gram-negative bacteria, the LuxR receptors bind specific AHLs; whereas orphan LuxR receptors recognize signal molecules more than just AHLs, and involve in the interspecies and interkingdom communications among different bacteria and their hosts (Patankar and Gonzalez, 2009). Some orphan LuxR homologs have structural similarity with the MDR regulators of the TetR family. For example, the QS regulators HapR and SmcR, with no ligand reported, are very similar to the TetR family QacR, TtgR, and EthR (De Silva et al., 2007; Kim et al., 2010).
In Gram-positive Streptomyces, only several GBL molecules and cognate receptors are identified so far (Biarnes-Carrera et al., 2015). These pseudo GBL receptors can not bind endogenous GBL molecules (Nishida et al., 2007). The crystal structure of CprB was reported, with no ligand identified (Bhukya et al., 2014). Interestingly, two pseudo GBL receptors could recognize endogenous antibiotics: ScbR2 (S. coelicolor) respond to actinorhodin and undecylprodigiosin; JadR2 (S. venezuelae) binds jadomycin and chloramphenicol (Xu et al., 2010). And also, by interacting with ScbR2, the angucycline antibiotics jadomycin may function as signals to modulate the antibiotic production and morphological differentiation of S. coelicolor (Wang et al., 2014). The pseudo GBL receptors also negatively control the GBL biosynthesis, which expand the regulatory network (Wang et al., 2011; Liu et al., 2013). Moreover, the GBL molecule SVB1 (S. venezuelae) is identical to the SCB3 (S. coelicolor), which may suggest a novel signaling role for GBL molecules in the interspecies communication (Nodwell, 2014; Zou et al., 2014).
Microbes could produce structurally diverse natural products including antibiotics. Some investigators doubt whether the antibiotics in natural habitat can reach the killing concentration (Linares et al., 2006). Many antibiotics of subinhibitory concentration alter bacterial gene expression profiles but not inhibit bacterial growth, which is the traditional characteristics of QS signals (Davies et al., 2006; Fajardo and Martinez, 2008). Some gene expression alterations lead to significant interference in the transduction of QS signaling, which is similar to the inhibition of QS system (Linares et al., 2006). These phenomena may suggest us to rethink their physiological functions for microbes. The ecological roles of antibiotics might be as signal molecules among cell–cell communications in the natural environment (Romero et al., 2011).
The QS signals at high concentration have also been reported to have similar bioactivity like antibiotics (Schertzer et al., 2009). For instance, the 3-oxo-N-acylhomoserine lactones, Pseudomonas quinolone signal (PQS), and phenazines have been proved to have antimicrobial activities (Kaufmann et al., 2005; Dietrich et al., 2008; Schertzer et al., 2009). More and more studies have found that low concentration of antibiotics may act as signal molecules during cell–cell communication; while the QS signals also have antimicrobial activity at high concentration (Romero et al., 2011).
Moreover, the QS and MDR regulatory systems may share the same path to transport molecules. The MDR regulatory systems use the multidrug efflux pumps that also export QS signals (Yang et al., 2006). The drug molecules exported by these efflux pumps are similar to the QS signals. High concentration of antibiotics may induce overexpression of exporter genes for more efflux pumps. These molecules are recognized by the TetR family regulators, and these transcriptional regulators are in charge of the efflux transporters (Piddock, 2006; Cuthbertson and Nodwell, 2013). For example, the TtgR (Pseudomonas putida) regulates the efflux pump TtgABC, and the ActR (S. coelicolor) controls the exporter ActA (Alguel et al., 2007; Willems et al., 2008). The physiological role of these efflux pumps might be to export signal molecules for cell–cell communication (Piddock, 2006; Yang et al., 2006).
As summarized in Figure 2, the TetR family regulators play vital roles in the transcriptional regulation of cell–cell communication using chemical languages. The AHL receptors regulate the AHLs biosynthesis in Gram-negative bacteria; while the GBL receptors control the GBLs biosynthesis in Gram-positive Streptomyces. Some orphan LuxR and pseudo GBL receptors respond to molecules more than just QS signals. The QS signals may have antimicrobial activity; while antibiotics also act as signal molecules. The efflux pumps for antibiotics are also the exporters for QS signals. Therefore, the QS and MDR regulatory systems might have some evolutionary relationship and biological relevance. QS interference using inhibitors is proving to be a new strategy for antimicrobial therapy (Hirakawa and Tomita, 2013; LaSarre and Federle, 2013). Further understanding the evolutionary history and biological roles of these regulatory systems would have theoretical significance and potential application in future.
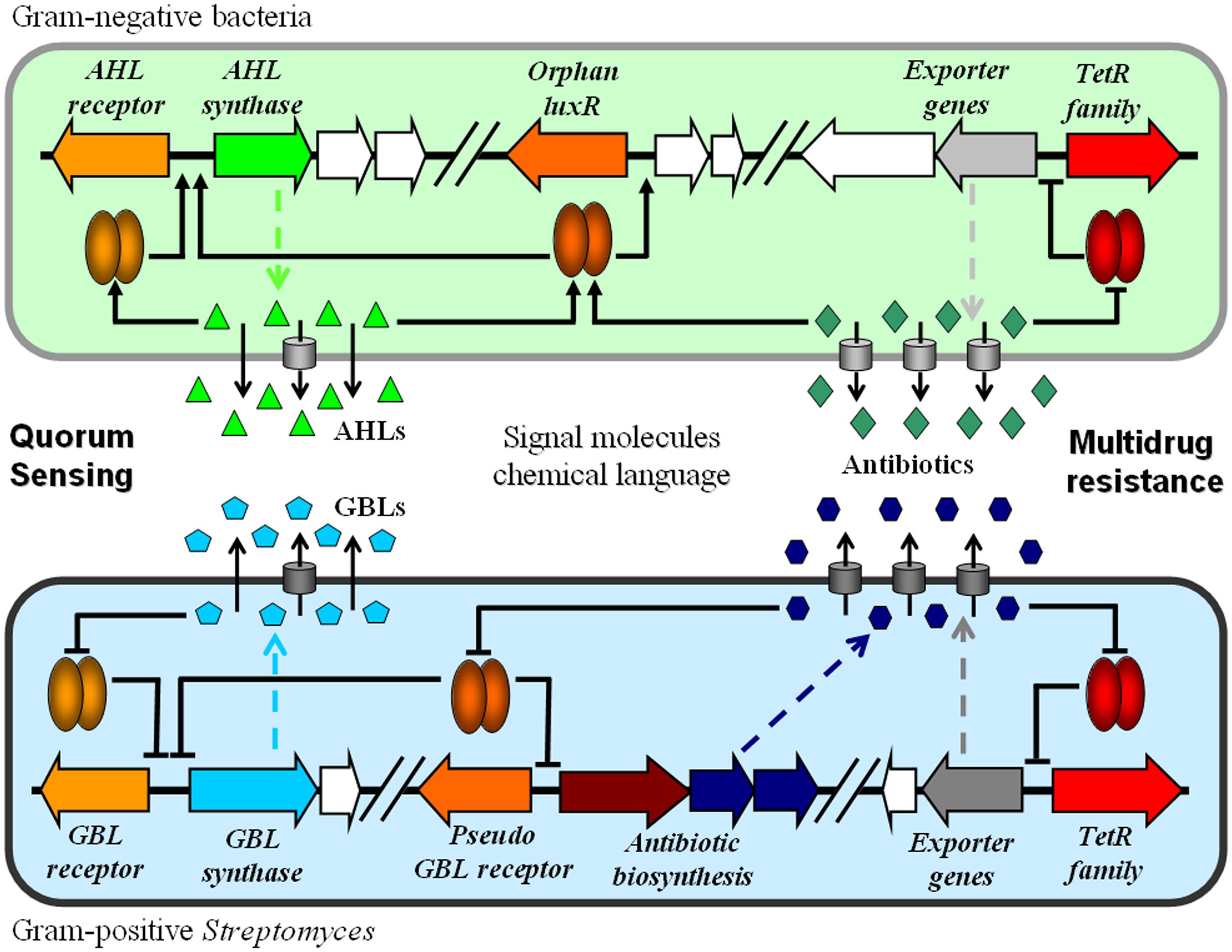
FIGURE 2. Relationships between the regulatory systems of QS and MDR in bacteria. Cell–cell communications involve the signal molecules (AHLs, GBLs, and antibiotics) as chemical languages and the corresponding receptors (LuxR, GBL receptor, and TetR family) as transcriptional regulators.
Author Contributions
GX developed the ideas, wrote and approved the final version of the manuscript.
Funding
This work is supported by grants from the National Natural Science Foundation of China (31570038 and 31200042).
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Alguel, Y., Meng, C., Teran, W., Krell, T., Ramos, J. L., Gallegos, M. T., et al. (2007). Crystal structures of multidrug binding protein TtgR in complex with antibiotics and plant antimicrobials. J. Mol. Biol. 369, 829–840. doi: 10.1016/j.jmb.2007.03.062
Bassler, B. L., and Losick, R. (2006). Bacterially speaking. Cell 125, 237–246. doi: 10.1016/j.cell.2006.04.001
Bhukya, H., Bhujbalrao, R., Bitra, A., and Anand, R. (2014). Structural and functional basis of transcriptional regulation by TetR family protein CprB from S. coelicolor A3(2). Nucleic Acids Res. 42, 10122–10133. doi: 10.1093/nar/gku587
Biarnes-Carrera, M., Breitling, R., and Takano, E. (2015). Butyrolactone signalling circuits for synthetic biology. Curr. Opin. Chem. Biol. 28, 91–98. doi: 10.1016/j.cbpa.2015.06.024
Camilli, A., and Bassler, B. L. (2006). Bacterial small-molecule signaling pathways. Science 311, 1113–1116. doi: 10.1126/science.1121357
Cuthbertson, L., and Nodwell, J. R. (2013). The TetR family of regulators. Microbiol. Mol. Biol. Rev. 77, 440–475. doi: 10.1128/MMBR.00018-13
Davies, J., Spiegelman, G. B., and Yim, G. (2006). The world of subinhibitory antibiotic concentrations. Curr. Opin. Microbiol. 9, 445–453. doi: 10.1016/j.mib.2006.08.006
De Silva, R. S., Kovacikova, G., Lin, W., Taylor, R. K., Skorupski, K., and Kull, F. J. (2007). Crystal structure of the Vibrio cholerae quorum-sensing regulatory protein HapR. J. Bacteriol. 189, 5683–5691. doi: 10.1128/JB.01807-06
Dietrich, L. E., Teal, T. K., Price-Whelan, A., and Newman, D. K. (2008). Redox-active antibiotics control gene expression and community behavior in divergent bacteria. Science 321, 1203–1206. doi: 10.1126/science.1160619
Fajardo, A., and Martinez, J. L. (2008). Antibiotics as signals that trigger specific bacterial responses. Curr. Opin. Microbiol. 11, 161–167. doi: 10.1016/j.mib.2008.02.006
Gray, K. M., and Garey, J. R. (2001). The evolution of bacterial LuxI and LuxR quorum sensing regulators. Microbiology 147(Pt 8), 2379–2387. doi: 10.1099/00221287-147-8-2379
Hirakawa, H., and Tomita, H. (2013). Interference of bacterial cell-to-cell communication: a new concept of antimicrobial chemotherapy breaks antibiotic resistance. Front. Microbiol. 4:114. doi: 10.3389/fmicb.2013.00114
Kaufmann, G. F., Sartorio, R., Lee, S. H., Rogers, C. J., Meijler, M. M., Moss, J. A., et al. (2005). Revisiting quorum sensing: discovery of additional chemical and biological functions for 3-oxo-N-acylhomoserine lactones. Proc. Natl. Acad. Sci. U.S.A. 102, 309–314. doi: 10.1073/pnas.0408639102
Kim, Y., Kim, B. S., Park, Y. J., Choi, W. C., Hwang, J., Kang, B. S., et al. (2010). Crystal structure of SmcR, a quorum-sensing master regulator of Vibrio vulnificus, provides insight into its regulation of transcription. J. Biol. Chem. 285, 14020–14030. doi: 10.1074/jbc.M109.100248
Kitani, S., Iida, A., Izumi, T. A., Maeda, A., Yamada, Y., and Nihira, T. (2008). Identification of genes involved in the butyrolactone autoregulator cascade that modulates secondary metabolism in Streptomyces lavendulae FRI-5. Gene 425, 9–16. doi: 10.1016/j.gene.2008.07.043
LaSarre, B., and Federle, M. J. (2013). Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 77, 73–111. doi: 10.1128/MMBR.00046-12
Lerat, E., and Moran, N. A. (2004). The evolutionary history of quorum-sensing systems in bacteria. Mol. Biol. Evol. 21, 903–913. doi: 10.1093/molbev/msh097
Linares, J. F., Gustafsson, I., Baquero, F., and Martinez, J. L. (2006). Antibiotics as intermicrobial signaling agents instead of weapons. Proc. Natl. Acad. Sci. U.S.A. 103, 19484–19489. doi: 10.1073/pnas.0608949103
Lintz, M. J., Oinuma, K., Wysoczynski, C. L., Greenberg, E. P., and Churchill, M. E. (2011). Crystal structure of QscR, a Pseudomonas aeruginosa quorum sensing signal receptor. Proc. Natl. Acad. Sci. U.S.A. 108, 15763–15768. doi: 10.1073/pnas.1112398108
Liu, G., Chater, K. F., Chandra, G., Niu, G., and Tan, H. (2013). Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 77, 112–143. doi: 10.1128/MMBR.00054-12
Martinez, J. L., Sanchez, M. B., Martinez-Solano, L., Hernandez, A., Garmendia, L., Fajardo, A., et al. (2009). Functional role of bacterial multidrug efflux pumps in microbial natural ecosystems. FEMS Microbiol. Rev. 33, 430–449. doi: 10.1111/j.1574-6976.2008.00157.x
Nikaido, H. (2009). Multidrug resistance in bacteria. Annu. Rev. Biochem. 78, 119–146. doi: 10.1146/annurev.biochem.78.082907.145923
Nishida, H., Ohnishi, Y., Beppu, T., and Horinouchi, S. (2007). Evolution of gamma-butyrolactone synthases and receptors in Streptomyces. Environ. Microbiol. 9, 1986–1994. doi: 10.1111/j.1462-2920.2007.01314.x
Nodwell, J. R. (2014). Are you talking to me? A possible role for gamma-butyrolactones in interspecies signalling. Mol. Microbiol. 94, 483–485. doi: 10.1111/mmi.12787
Patankar, A. V., and Gonzalez, J. E. (2009). Orphan LuxR regulators of quorum sensing. FEMS Microbiol. Rev. 33, 739–756. doi: 10.1111/j.1574-6976.2009.00163.x
Paulsen, I. T. (2003). Multidrug efflux pumps and resistance: regulation and evolution. Curr. Opin. Microbiol. 6, 446–451. doi: 10.1016/j.mib.2003.08.005
Piddock, L. J. (2006). Multidrug-resistance efflux pumps - not just for resistance. Nat. Rev. Microbiol. 4, 629–636. doi: 10.1038/nrmicro1464
Ramos, J. L., Martinez-Bueno, M., Molina-Henares, A. J., Teran, W., Watanabe, K., Zhang, X., et al. (2005). The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 69, 326–356. doi: 10.1128/MMBR.69.2.326-356.2005
Romero, D., Traxler, M. F., Lopez, D., and Kolter, R. (2011). Antibiotics as signal molecules. Chem. Rev. 111, 5492–5505. doi: 10.1021/cr2000509
Ryan, R. P., and Dow, J. M. (2008). Diffusible signals and interspecies communication in bacteria. Microbiology 154(Pt 7), 1845–1858. doi: 10.1099/mic.0.2008/017871-0
Schertzer, J. W., Boulette, M. L., and Whiteley, M. (2009). More than a signal: non-signaling properties of quorum sensing molecules. Trends Microbiol. 17, 189–195. doi: 10.1016/j.tim.2009.02.001
Schumacher, M. A., Miller, M. C., Grkovic, S., Brown, M. H., Skurray, R. A., and Brennan, R. G. (2001). Structural mechanisms of QacR induction and multidrug recognition. Science 294, 2158–2163. doi: 10.1126/science.1066020
Subramoni, S., and Venturi, V. (2009). LuxR-family ‘solos’: bachelor sensors/regulators of signalling molecules. Microbiology 155(Pt 5), 1377–1385. doi: 10.1099/mic.0.026849-0
Takano, E. (2006). Gamma-butyrolactones: Streptomyces signalling molecules regulating antibiotic production and differentiation. Curr. Opin. Microbiol. 9, 287–294. doi: 10.1016/j.mib.2006.04.003
Wang, J., Wang, W., Wang, L., Zhang, G., Fan, K., Tan, H., et al. (2011). A novel role of ‘pseudo’ gamma-butyrolactone receptors in controlling gamma-butyrolactone biosynthesis in Streptomyces. Mol. Microbiol. 82, 236–250. doi: 10.1111/j.1365-2958.2011.07811.x
Wang, W. S., Ji, J. J., Li, X., Wang, J., Li, S. S., Pan, G. H., et al. (2014). Angucyclines as signals modulate the behaviors of Streptomyces coelicolor. Proc. Natl. Acad. Sci. U.S.A. 111, 5688–5693. doi: 10.1073/pnas.1324253111
Waters, C. M., and Bassler, B. L. (2005). Quorum sensing: cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 21, 319–346. doi: 10.1146/annurev.cellbio.21.012704.131001
Willems, A. R., Tahlan, K., Taguchi, T., Zhang, K., Lee, Z. Z., Ichinose, K., et al. (2008). Crystal structures of the Streptomyces coelicolor TetR-like protein ActR alone and in complex with actinorhodin or the actinorhodin biosynthetic precursor (S)-DNPA. J. Mol. Biol. 376, 1377–1387. doi: 10.1016/j.jmb.2007.12.061
Xu, G., Wang, J., Wang, L., Tian, X., Yang, H., Fan, K., et al. (2010). “Pseudo” gamma-butyrolactone receptors respond to antibiotic signals to coordinate antibiotic biosynthesis. J. Biol. Chem. 285, 27440–27448. doi: 10.1074/jbc.M110.143081
Yang, S., Lopez, C. R., and Zechiedrich, E. L. (2006). Quorum sensing and multidrug transporters in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 103, 2386–2391. doi: 10.1073/pnas.0502890102
Yu, Z., Reichheld, S. E., Savchenko, A., Parkinson, J., and Davidson, A. R. (2010). A comprehensive analysis of structural and sequence conservation in the TetR family transcriptional regulators. J. Mol. Biol. 400, 847–864. doi: 10.1016/j.jmb.2010.05.062
Keywords: regulatory system, quorum sensing, multidrug resistance, transcriptional regulator, signal molecule
Citation: Xu G-M (2016) Relationships between the Regulatory Systems of Quorum Sensing and Multidrug Resistance. Front. Microbiol. 7:958. doi: 10.3389/fmicb.2016.00958
Received: 15 April 2016; Accepted: 02 June 2016;
Published: 16 June 2016.
Edited by:
José Eduardo González-Pastor, Centro de Astrobiologia-Instituto Nacional de Tecnica Aeroespacial, SpainReviewed by:
Luyan Zulie Ma, Chinese Academy of Sciences, ChinaMoshe Shemesh, Agricultural Research Organization, Volcani Center, Israel
Copyright © 2016 Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gang-Ming Xu, YWVyaWN4dUBnbWFpbC5jb20=