 Tanja Obermajer
Tanja Obermajer Tomislav Pogačić
Tomislav Pogačić- 1Department of Animal Science, Biotechnical Faculty, Institute of Dairy Science and Probiotics, University of Ljubljana, Domžale, Slovenia
- 2Dairy Farm–Hanne Storm Brems, Faaborg, Denmark
A commentary on
Relationship between Milk Microbiota, Bacterial Load, Macronutrients, and Human Cells during Lactation
by Boix-Amorós, A., Collado, M. C. and Mira, A. (2016). Front. Microbiol. 7:492. doi: 10.3389/fmicb.2016.00492
The recent contribution by Boix-Amorós et al. (2016) reveals certain associations among the microbiota, bioactive components and nutrients found in human colostrum and breast milk of 21 healthy Spanish lactating mothers. We would like to emphasize the importance of their findings, discuss some phenomena and highlight the relevant future research directions (Figure 1).
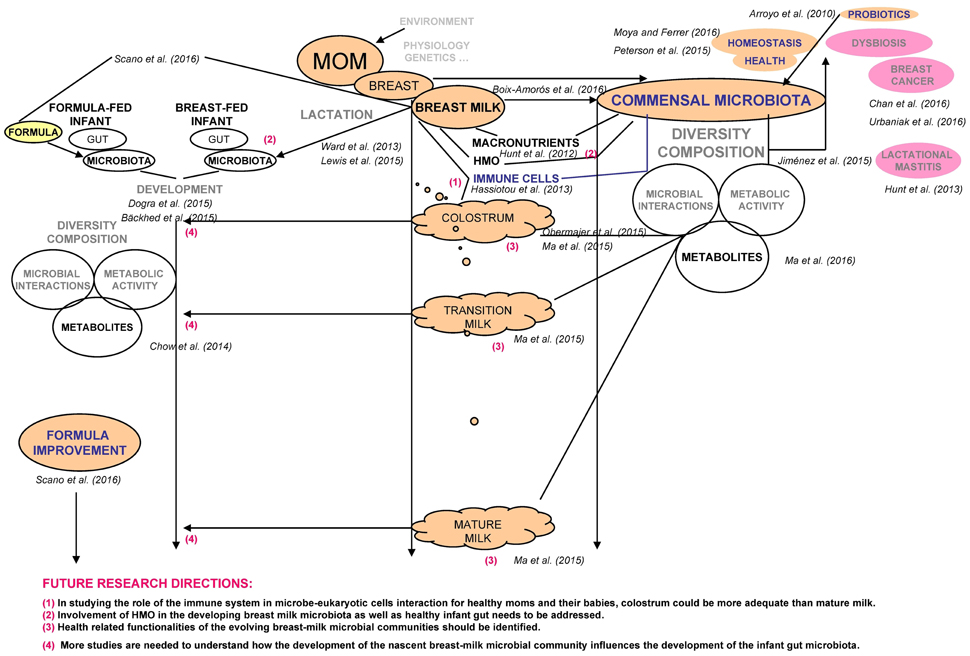
Figure 1. Schematic presentation summarizing the discussed current research work and proposed future research directions.
The commensal microbiota, as part of epigenetic landscape, reflects human health status by its diversity, composition and metabolic activity (Moya and Ferrer, 2016). An improved understanding of the factors controlling health associated bacterial interactions may, upon perturbation, define novel therapeutic or preventive measures to restore individual's dynamic microbial equilibria and immune homeostasis (Peterson et al., 2015). Probiotics were demonstrated as an effective tool in lactational mastitis interventions (Arroyo et al., 2010), and may even hold potential in breast cancer prevention (Chan et al., 2016; Urbaniak et al., 2016).
In the study by Boix-Amorós et al. (2016), microbial communities, again, were confirmed as complex and individual-specific. High inter-subject variability was disclosed for the composition and the number of bacteria per ml, while higher total bacterial load was not associated with lower diversity of milk sample. Namely, as reported previously, the milk microbiome of mastitis-affected women suffered a loss of bacterial diversity, while an increase in sequences related to the presumptive etiological agents was also observed (Jiménez et al., 2015). Staphylococci and streptococci were classified in dominant core of genera, existing at all three lactation sampling points (Boix-Amorós et al., 2016). The study further suggested that the specific microbiota composition, rather than total sample bacterial load, might trigger the mammary gland immune responses. Moreover, the positive correlation was only established between the staphylococci proportion and somatic cell count (SCC). Specifically, the SCC biomarker was earlier reported (Hunt et al., 2013) to be roughly 10-fold increased in milk from mastitic compared to milk from healthy breasts. Similarly to the work by Boix-Amorós et al. (2016), our recent publication studied the colostrum bioactive potential of 45 healthy Slovenian mothers (Obermajer et al., 2015) and qPCR approach was applied on the automatedly extracted colostrum microbial DNA. The interactions among target bacterial groups were investigated. As in previous reports concerning microbial biofilms, but in oral specimens (Benítez-Páez et al., 2014; Loozen et al., 2014), here, we would like to underline the positive abundance correlations found between the specific microbial components in colostrum samples of different mothers (e.g., Staphylococcus and total bacterial load; Staphylococcus and Staphylococcus epidermidis), testifying that some common issues in microbial consortia truly exist and may play a role in the maintenance of the ecosystem homeostasis (Obermajer et al., 2015). As suggested by Delgado et al. (2008), the trend toward the overgrowth of one component (e.g., staphylococci and streptococci strains as a common mastitis causing agents) could provoke a dysbiotic process in a predisposed host. Interestingly enough, staphylococci and streptococci were also quantitatively correlated (Spearman's rank correlation coefficient 0.478; p < 0.01) (Obermajer et al., 2015), even though their quantities varied considerably among samples of different mothers. In contrast, Boix-Amorós et al. (2016) reported a trend toward the negative association between the total bacterial load and the staphylococci proportion. While the difference in this correlation remains to be elucidated, we could hypothesize that a significantly higher level of immune cells in colostrum than in mature milk (up to 70% of the total milk cells for colostrum vs. 0–2% of total cells for mature milk; Hassiotou et al., 2013) could favor different associations between specific microbial populations in both human fluids. Thus, differential characteristics of colostrum, mainly related to immune cells, could make this biological samples more suitable to study bacteria–human immune cells interactions.
The study by Boix-Amorós et al. (2016) presented different compositional and diversity patterns in milk microbiome at three lactation stages. Considering few bacterial genera present at all-time points, future research should focus on the bacterial species interaction networks as a research tool to decipher possible functionalities changing during evolution of milk microbial communities. The robust microbiotas, able to regain their original function upon disturbances, should be identified. The proposed networking approach on the breast milk dataset was recently employed for the geographically distinct healthy cohort (Ma et al., 2015) and was also applied (Ma et al., 2016) in the preliminary comparative analysis of Hodgkin's lymphoma milk microbiome after drug administration. Moreover, the authors suggested the metabolites-microbial species networks to be very useful in identifying the active bacterial taxa really involved in the milk microbial communities conferred to infants.
Boix-Amorós et al. (2016) successfully identified some associations among macronutrients and specific microbial genera, however, the authors failed to address an important prebiotic component: human milk oligosaccharides (HMO). Hunt et al. (2012) demonstrated that HMO stimulated the growth of staphylococci isolates from breast milk. But, the observed effect was only associated with the increased amino acid metabolism and not with the depletion of HMO. As proposed, the specific HMO profiles in women may be involved in the development of lactational mastitis. Lewis et al. (2015) extended the importance of the maternal synthesis of human milk glycans on the bifidobacterial species content in the evolving infant gut.
Daily consumption of 107–108 bacterial cells by a breast-fed infant (Boix-Amorós et al., 2016) is a valuable information from the perspective of competitive early supplementation of infant formulae. Ward et al. (2013) suggested that a broad array of microbes within the human milk likely contributes toward an effective infant gut colonization, while diverse microbial DNA sequences as well as mother's DNA may create a balanced immune stimulatory and suppressive DNA exposures, leading infants to eventual tolerance of the immense number of gut bacteria. Following birth, the microbial gut consortia form in a non-random, successive fashion (Dogra et al., 2015). While the developing microbiome of formula-fed infants fails to resemble the compositional and functional capacity of the microbiome of breast-fed infants (Bäckhed et al., 2015), the proposed research directions (Figure 1) with network analyses (nutrients-microbes-metabolites; Chow et al., 2014; Scano et al., 2016) may open new frontiers to formulate the next generation of innovative infant foods.
Author Contributions
TO drafted the work and TP critically revised the work. Both authors approved the final version.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Arroyo, R., Martín, V., Maldonado, A., Jiménez, E., Fernández, L., and Rodríguez, J. M. (2010). Treatment of infectious mastitis during lactation: antibiotics versus oral administration of lactobacilli isolated from breast milk. Clin. Infect. Dis. 50, 1551–1558. doi: 10.1086/652763
Bäckhed, F., Roswall, J., Peng, Y., Feng, Q., Jia, H., Kovatcheva-Datchary, P., et al. (2015). Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 17, 690–703. doi: 10.1016/j.chom.2015.04.004
Benítez-Páez, A., Belda-Ferre, P., Simón-Soro, A., and Mira, A. (2014). Microbiota diversity and gene expression dynamics in human oral biofilms. BMC Genomics 15:311. doi: 10.1186/1471-2164-15-311
Boix-Amorós, A., Collado, M. C., and Mira, A. (2016). relationship between milk microbiota, bacterial load, macronutrients, and human cells during lactation. Front. Microbiol. 7:492. doi: 10.3389/fmicb.2016.00492
Chan, A. A., Bashir, M., Rivas, M. N., Duvall, K., Sieling, P. A., Pieber, T. R., et al. (2016). Characterization of the microbiome of nipple aspirate fluid of breast cancer survivors. Sci. Rep. 6:28061. doi: 10.1038/srep28061
Chow, J., Panasevich, M. R., Alexander, D., Vester Boler, B. M., Rossoni Serao, M. C., Faber, T. A., et al. (2014). Fecal metabolomics of healthy breast-fed versus formula-fed infants before and during in vitro batch culture fermentation. J. Proteome Res. 13, 2534–2542. doi: 10.1021/pr500011w
Delgado, S., Arroyo, R., Martín, R., and Rodríguez, J. M. (2008). PCR-DGGE assessment of the bacterial diversity of breast milk in women with lactational infectious mastitis. BMC Infect. Dis. 8:51. doi: 10.1186/1471-2334-8-51
Dogra, S., Sakwinska, O., Soh, S.-E., Ngom-Bru, C., Brück, W. M., Berger, B., et al. (2015). Rate of establishing the gut microbiota in infancy has consequences for future health. Gut Microbes 6, 321–325. doi: 10.1080/19490976.2015.1078051
Hassiotou, F., Hepworth, A. R., Metzger, P., Tat Lai, C., Trengove, N., Hartmann, P. E., et al. (2013). Maternal and infant infections stimulate a rapid leukocyte response in breastmilk. Clin. Transl. Immunol. 2:e3. doi: 10.1038/cti.2013.1
Hunt, K. M., Preuss, J., Nissan, C., Davlin, C. A., Williams, J. E., Shafii, B., et al. (2012). Human milk oligosaccharides promote the growth of staphylococci. Appl. Environ. Microbiol. 78, 4763–4770. doi: 10.1128/AEM.00477-12
Hunt, K. M., Williams, J. E., Shafii, B., Hunt, M. K., Behre, R., Ting, R., et al. (2013). Mastitis is associated with increased free fatty acids, somatic cell count, and interleukin-8 concentrations in human milk. Breastfeed. Med. Off. J. Acad. Breastfeed. Med. 8, 105–110. doi: 10.1089/bfm.2011.0141
Jiménez, E., de Andrés, J., Manrique, M., Pareja-Tobes, P., Tobes, R., Martínez-Blanch, J. F., et al. (2015). Metagenomic analysis of milk of healthy and mastitis-suffering women. J. Hum. Lact. 31, 406–415. doi: 10.1177/0890334415585078
Lewis, Z. T., Totten, S. M., Smilowitz, J. T., Popovic, M., Parker, E., Lemay, D. G., et al. (2015). Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome 3:13. doi: 10.1186/s40168-015-0071-z
Loozen, G., Ozcelik, O., Boon, N., De Mol, A., Schoen, C., Quirynen, M., et al. (2014). Inter-bacterial correlations in subgingival biofilms: a large-scale survey. J. Clin. Periodontol. 41, 1–10. doi: 10.1111/jcpe.12167
Ma, Z. S., Guan, Q., Ye, C., Zhang, C., Foster, J. A., and Forney, L. J. (2015). Network analysis suggests a potentially “evil” alliance of opportunistic pathogens inhibited by a cooperative network in human milk bacterial communities. Sci. Rep. 5:8275. doi: 10.1038/srep08275
Ma, Z. S., Li, L., Li, W., Li, J., and Chen, H. (2016). Integrated network-diversity analyses suggest suppressive effect of Hodgkin's lymphoma and slightly relieving effect of chemotherapy on human milk microbiome. Sci. Rep. 6:28048. doi: 10.1038/srep28048
Moya, A., and Ferrer, M. (2016). Functional redundancy-induced stability of gut microbiota subjected to disturbance. Trends Microbiol. 24, 402–413. doi: 10.1016/j.tim.2016.02.002
Obermajer, T., Lipoglavšek, L., Tompa, G., Treven, P., Lorbeg, P. M., Matijašić, B. B., et al. (2015). Colostrum of healthy slovenian mothers: microbiota composition and bacteriocin gene prevalence. PLoS ONE 10:e0123324. doi: 10.1371/journal.pone.0123324
Peterson, C. T., Sharma, V., Elmén, L., and Peterson, S. N. (2015). Immune homeostasis, dysbiosis and therapeutic modulation of the gut microbiota. Clin. Exp. Immunol. 179, 363–377. doi: 10.1111/cei.12474
Scano, P., Murgia, A., Demuru, M., Consonni, R., and Caboni, P. (2016). Metabolite profiles of formula milk compared to breast milk. Food Res. Int. 87, 76–82. doi: 10.1016/j.foodres.2016.06.024
Urbaniak, C., Gloor, G. B., Brackstone, M., Scott, L., Tangney, M., and Reid, G. (2016). The microbiota of breast tissue and its association with breast cancer. Appl. Environ. Microbiol. 82, 5039–5048. doi: 10.1128/AEM.01235-16
Keywords: human colostrum, human milk, microbiota, diversity, microbial interactions, ecosystem homeostasis, functionality
Citation: Obermajer T and Pogačić T (2016) Commentary: Relationship between Milk Microbiota, Bacterial Load, Macronutrients, and Human Cells during Lactation. Front. Microbiol. 7:1281. doi: 10.3389/fmicb.2016.01281
Received: 12 June 2016; Accepted: 03 August 2016;
Published: 17 August 2016.
Edited by:
Martin G. Klotz, Queens College of The City University of New York, USAReviewed by:
David Andrew Mills, University of California, Davis, USAClara G. De Los Reyes-Gavilan, Spanish National Research Council, Spain
Maria Carmen Collado, Spanish National Research Council, Spain
Copyright © 2016 Obermajer and Pogačić. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tanja Obermajer, dGFuamEub2Jlcm1hamVyQGJmLnVuaS1sai5zaQ==