 Estefânia F. Garcia1Winnie A. Luciano1
Estefânia F. Garcia1Winnie A. Luciano1 Danilo E. Xavier1Whyara C. A. da Costa2
Danilo E. Xavier1Whyara C. A. da Costa2 Kleber de Sousa Oliveira3
Kleber de Sousa Oliveira3 Octávio L. Franco3Marcos A. de Morais Júnior4
Octávio L. Franco3Marcos A. de Morais Júnior4 Brígida T. L. Lucena4Renata C. Picão5
Brígida T. L. Lucena4Renata C. Picão5 Marciane Magnani2
Marciane Magnani2 Maria Saarela6
Maria Saarela6 Evandro L. de Souza1*
Evandro L. de Souza1*- 1Laboratório de Microbiologia de Alimentos, Departamento de Nutrição, Universidade Federal da Paraíba, João Pessoa, Brazil
- 2Laboratório de Processos Microbianos em Alimentos, Departamento de Engenharia de Alimentos, Universidade Federal da Paraíba, João Pessoa, Brazil
- 3Laboratório de Genômica e Proteômica, Universidade Católica de Brasília, Brasília, Brazil
- 4Grupo Interdepartamental de Pesquisa em Engenharia Metabólica, Departamento de Genética, Universidade Federal de Pernambuco, Recife, Brazil
- 5Instituto de Microbiologia Paulo Góes, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
- 6VTT Technical Research Centre of Finland, Espoo, Finland
This study aimed to identify lactic acid bacteria (LAB) in byproducts of fruit (Malpighia glabra L., Mangifera indica L., Annona muricata L., and Fragaria vesca L.) pulp processing. Fifty strains of LAB were identified using matrix-assisted laser desorption/ionization–time of flight mass spectrometry (MALDI-TOF MS) and 16S rRNA gene sequence (16S rRNA) analysis. Species belonging to Lactobacillus genus were the predominant LAB in all fruit pulp processing byproducts. The average congruency between the MALDI-TOF MS and 16S rRNA in LAB species identification reached 86%. Isolates of L. plantarum, L. brevis, L. pentosus, L. lactis and L. mesenteroides were identified with 100% congruency. MALDI-TOF MS and 16S rRNA analysis presented 86 and 100% efficiency of LAB species identification, respectively. Further, five selected Lactobacillus strains (L. brevis 59, L. pentosus 129, L. paracasei 108, L. plantarum 49, and L. fermentum 111) were evaluated for desirable probiotic-related properties and growth behavior on two different cultivation media. The exposure to pH 2.0 sharply decreased the counts of the different Lactobacillus strains after a 1 or 2 h incubation, while varied decreases were noted after 3 h of exposure to pH 3.0. Overall, the exposure to pH 5.0 and to bile salts (0.15, 0.30, and 1.00%) did not decrease the counts of the Lactobacillus strains. All tested Lactobacillus strains presented inhibitory activity against Staphylococcus aureus, Salmonella Typhimurium, Salmonella Enteritidis, Listeria monocytogenes and Escherichia coli, and presented variable susceptibility to different antibiotics. The selected Lactobacillus strains presented satisfactory and reproducible growth behavior. In conclusion, MALDI-TOF MS and 16S rRNA analysis revealed high efficiency and congruency for LAB species identification, and the selected Lactobacillus strains may be candidates for further investigation of novel probiotic strains.
Introduction
Consumption of fruit and fruit products (mostly low-processed juices and frozen pulps) has been increasing due to growing recognition of their nutritional value associated with their high content of minerals, vitamins, and secondary phytochemical compounds (Rufino et al., 2010; Silva et al., 2014). The processing of fruit generates a great amount of industrial byproducts, representing 10–60% of the total fruit weight (Ayala-Zavala et al., 2010). These byproducts comprise peels, rinds, seeds, and unused flesh, which are usually inappropriately discarded in the environment, leading to waste accumulation, and negative environmental impacts (Ajila et al., 2007; Araújo et al., 2014).
The economics of processing tropical fruit could be improved by developing higher value use for their byproducts (Silva et al., 2014). In addition to the known potential use of tropical fruit pulps and their byproducts for the isolation of phytochemicals for application in nutraceutical supplements (Ayala-Zavala et al., 2011), these byproducts also exhibit a wide variety of microorganisms of interest to the food industry (Yang et al., 2010), especially lactic acid bacteria (LAB). Each particular type of fruit provides a unique environment in terms of chemical composition, buffering capacity, competitive microbiota, and natural antagonist compounds (Naeem et al., 2012). The microbial populations of raw fruit commonly vary between 5 and 7 log CFU/g, where LAB constitute a small part (2–4 log CFU/g) of the autochthonous microbiota (Di Cagno et al., 2010a,b, 2011a,b).
Most probiotic bacteria are LAB, and among them, Lactobacillus is the most common genera (Argyri et al., 2013). According to FAO/WHO (2006), probiotics are non-pathogenic microorganisms, which exert a positive health benefit on the host when ingested in an adequate amount. The majority of the commercialized and most studied probiotics have been isolated from dairy products and human gastrointestinal tract (García-Ruiz et al., 2014). Although, dairy foods are recognized to be the best vehicle for the delivery of viable probiotics to the human gut, the increasing number of individuals with lactose intolerance, dyslipidemia, and vegetarianism reinforces the importance of the development of non-dairy probiotic products (Ranadheera et al., 2010; Peres et al., 2012), such as fruit juices. In fruit juices, the low pH (approx. 3.7) compared to the fairly neutral pH of milk (approx. 6.7) is possibly the chief determinant for the poor viability of probiotics in these matrices (Saarela et al., 2006). Raw fruit and their byproducts possess intrinsic physicochemical parameters that resemble those of the human gastrointestinal tract for some traits, such as the acidic environment and presence of anti-nutritional factors (tannins and phenols, Vitali et al., 2012). The natural adaptation to the intrinsic characteristics of fruit may help fruit-originating bacteria to survive during the processing and storage of fruit-based probiotic formulations as well as in the human stomach.
Various LAB have been isolated from fruit as follows: Lactobacillus rossiae from pineapple (Di Cagno et al., 2010a,b); L. plantarum from tomato, pineapple, plum, kiwi, papaya, grape, strawberry, and cherries (Di Cagno et al., 2008a,b, 2010a, 2011a,b; Naeem et al., 2012); L. brevis from tomato (Di Cagno et al., 2008b); and Leuconostoc mesenteroides subsp. mesenteroides and Pediococcus pentosaceus from cherries (Di Cagno et al., 2011b). The identification of LAB species in fruit is typically performed using molecular tools, particularly polymerase chain reaction (PCR)-based methods and 16S rRNA gene sequencing (Dusková et al., 2012). Matrix-assisted laser desorption/ionization–time of flight mass spectrometry (MALDI-TOF MS) has been recently introduced with marked success into routine clinical microbiological diagnosis of human pathogens (Bizzini et al., 2011; Welker, 2011; Nomura, 2015). However, studies reporting the application of MALDI-TOF MS for bacterial identification in food microbiology are still uncommon. The capability of this technique to identify bacteria isolated from food matrices not only at the genus and species level but also at the subspecies level reveals that MALDI-TOF MS could become a key tool in food microbiology and safety (Angelakis et al., 2011; Dusková et al., 2012).
This study aimed (i) to isolate and then identify LAB in fruit pulp processing byproducts using MALDI-TOF MS and 16S rRNA gene sequence analysis, as well as to verify the identification congruency between the two techniques; (ii) to assess the probiotic properties of selected Lactobacillus strains in vitro, including acid tolerance, bile tolerance, and capability to inhibit pathogenic food-related bacteria; and (iii) to verify the antibiotic resistance and growth behavior of the selected Lactobacillus strains in different cultivation media.
Materials and Methods
Isolation of LAB
Samples (250 g) of fruit pulp processing byproducts of Malpighia glabra L. (barbados cherry), Mangifera indica L. (mango), Annona muricata L. (soursop), and Fragaria vesca L. (strawberry) were obtained from a company producing frozen fruit pulps located at the city of João Pessoa (Paraíba, Brazil). These byproducts were composed mostly of mashed peels and seeds as well as small amounts of mashed flesh. Initially, 25 g of each sample was suspended in 225 mL of sterile peptone water (0.1 g/100 mL) and homogenized using a stomacher (Model A440, Marconi Equip. Lab. Ltda., Piracicaba, Brazil) for 3 min at room temperature. Subsequently, serial dilutions (10−2–10−5) were performed using the same diluent, and 100 μL aliquots from each dilution were spread plated onto de Man, Rogosa, and Sharpe (MRS) agar (HiMedia, Mumbai, India) containing cysteine HCl (0.05 g/100 mL) and incubated anaerobically (Anaerobic System Anaerogen, Oxoid Ltda., Wade Road, UK) at 37°C for 48–72 h. At least five colonies presenting different morphologies were randomly isolated from MRS agar plates spread with the two highest serial dilutions of each type of fruit pulp byproduct. These isolates were maintained on MRS agar slants under refrigeration and further submitted to analysis of Gram staining, morphology, catalase production, and motility using standard procedures previously described (American Public Health (APHA), 2015). All of these analyses comprised a presumptive LAB identification step. All isolates presumptively identified as LAB (non-motile, catalase negative, Gram-positive cocci or rods) were stored at −20°C in MRS broth (HiMedia, Mumbai, India) containing glycerol (15 mL/100 mL) for further studies.
Identification of LAB Isolates
Identification Using 16S rRNA Gene Sequence Analysis
The bacterial genomic DNA was extracted using a Genomic DNA extraction kit (Promega Cooporation, Wisconsin, USA) according to the manufacturer's instructions. For the detection of 16S rRNA gene sequences, the following primers were used: 27F, 50-AGAGTTTGATCCTGGCTCAG-30, and 1492R, 50-GGTTACCTTGTTACGACTT-30. PCR was performed using a DNA thermocycler (Applied Biosystems, USA), and the reactions contained 0.5 μM of each primer, 0.2 mM dNTP mix, 1.5 mM MgCl2, and 1 U Taq DNA polymerase (Invitrogen, Germany) in a 50 μL final volume. The PCR was run under the following conditions: initial activation at 94°C for 2 min; denaturation step cycles at 94°C for 30 s; annealing step at 55°C for 1 min; extension step at 72°C for 1 min; and final cycle at 72°C for 10 min (Guo et al., 2010). The PCR products were purified using a DNA purification kit (Invitrogen, Germany) and sequenced using the 27F and 1492R primers in a sequencing reaction using the ABI Prism™ Bigdye™ terminator cycle sequencing reaction kit (Applied Biosystems, USA).
The resulting 1465 bp sequences were analyzed using the Pregap4 and Gap4 tools in the STADEN 1.6 software package and submitted to a search for similarity in the National Center for Biotechnology Information (NCBI) database using the blastn (nucleotide database) tool (Altschul et al., 1997; Guo et al., 2010) and Ribosomal Database Project (RDP). Bacterial identification was assumed when the query sequence showed similarity >97% for the 16S rRNA gene sequence (Gevers et al., 2005; Guo et al., 2010). Parcial 16S rRNA sequence was compared to known sequences in the NCBI Genbank database using the Local Alignment Search Tool (BLAST) algorithm (Altschul et al., 1990).
Identification Using MALDI-TOF MS Analysis
Initially, a standard protein extraction protocol adapted from Freiwald and Sauer (2009) was followed. Approximately 20 colonies of each LAB isolate culture were resuspended in 1.2 mL of 75% EtOH. After centrifugation (14,000 × g, 2 min, 4°C) and removal of the supernatant, proteins were extracted with 50 μL of an acetonitrile/formic acid/water mixture by vortexing for 1 min. The supernatant was then deposited in three wells of the sample plate at a volume of 1 μL and dried at room temperature, and the samples were then overlaid with 1 μL of a saturated alpha-cyano-4-hydroxycinnamic acid solution in acetonitrile:water:TFA (10 mg/mL; Bruker Daltonics, Germany).
The MALDI-TOF mass spectra measurements of samples were performed using a Bruker Biotyper 3.1 (Bruker Daltonics, Germany). External calibration of mass spectra was performed using Escherichia coli DH5 alpha standard peaks. Mass spectra were processed using MALDI Biotyper™ 3.1 software (Bruker Biotyper 3.1, Bruker Daltonics, Germany). The identification results were expressed by BioTyper log (scores) indicating the similarity of the unknown MALDI-TOF MS profile to available database entries. BioTyper logs (score) ≥2.3 and ≤3.0 indicate a highly probable identification at the species level; logs (score) ≥2.0 and ≤2.3 indicate secure genus identification, and probable species identification; logs (score) ≥1.7 and ≤2.0 imply probable genus identification; and logs (score) <1.7 imply no significant similarity between the unknown profile and any of the database entries. MALDI-TOF MS profile spectra for bacterial identification were automatelly compared to the BioTyper reference library of MALDI-TOF mass spectra by MALDI Biotype™ 3.1 software (Bruker Daltonics, Germany).
After the identification of LAB isolates, five different isolates with congruency of identification by the MALDI-TOF MS and 16S rRNA gene sequencing techniques that belonged to different species from the Lactobacillus genus, namely, L. brevis, L. pentosus, L. paracasei, L. plantarum, and L. fermentum, and that are commonly studied for probiotic properties, were selected for use in further assays of potential probiotic properties and growth behavior.
Inoculum of Lactobacilli and Pathogenic Bacteria
Initially, each Lactobacillus strain was grown anaerobically (Anaerobic System Anaerogen, Oxoid) in MRS broth at 37°C for 20–24 h (stationary growth phase), harvested through centrifugation (4500 g, 15 min, and 4°C), washed twice in sterile saline solution (0.85 g/100 mL) and resuspended in sterile saline solution to obtain cell suspensions with an OD reading at 660 nm (OD660) of 0.5. This suspension provided viable counts of approximately 8 log CFU/mL for each strain when pour plated in MRS agar.
The strains of the pathogenic bacteria Staphylococcus aureus (INCQS 00015, originally ATCC 25923), Salmonella enterica subsp. enterica serovar Typhimurium (INCQS 00150, originally ATCC 14028), S. enterica subsp. enterica serovar Enteritidis (INCQS 00258, originally 13076), L. monocytogenes (INCQS 00266, originally ATCC 7644), and E. coli (INCQS 00219, originally ATCC 8739) were obtained from the National Institute for Quality Control in Health (Oswaldo Cruz Foundation, Rio de Janeiro, Brazil). The stock cultures were maintained in Brain Heart Infusion (BHI) broth (HiMedia, Mumbai, India) containing glycerol (15 g/100 mL) at −20°C. Prior to use in antagonistic assays, each strain was aerobically grown in BHI broth at 37°C for 20–24 h, harvested through centrifugation (4500 g, 15 min, and 4°C), washed twice in sterile saline solution and resuspended in sterile saline solution to obtain cell suspensions with an OD625 of 0.1. This suspension provided viable cell counts of approximately 8 log CFU/mL for each strain when pour plated in BHI agar (HiMedia, Mumbai, India).
Acid and Bile Salt Tolerance Assays
The tolerance to different pH values and bile salt concentrations was assessed by inoculating 1 mL aliquots of each tested Lactobacillus strain suspension in 10 mL of PBS (final viable cell counts of approximately 7 log CFU/mL) with pH adjusted to 2.0, 3.0 or 5.0 (using 1 M HCl) or supplemented with bile salts (Sigma-Aldrich Co., St. Louis, USA) at 1.0, 2.0 or 3.0% (w/v). The cells were incubated aerobically at 37°C under stirring (150 rpm). At different incubation periods (1, 2, and 3 h), 1 mL aliquots were removed from each system, serially diluted in sterile peptone water (10−1–10−5) and spread plated onto MRS agar for enumeration of viable cells. After an incubation period of 48 h at 37°C under anaerobiosis (Anaerobic System Anaerogen, Oxoid), the viable cells were counted, and the results were expressed as the log of the colony forming units per mL (log CFU/mL). For controls, Lactobacillus strains were cultivated in PBS at pH 7.2 (adjusted using 1 M HCl) and in MRS without bile salts (Jacobsen et al., 1999; Monteagudo-Mera et al., 2012).
Antagonistic Activity against Pathogens
The antagonistic activity of the Lactobacillus strains against the indicator foodborne pathogenic bacteria was evaluated using the spot agar and well diffusion methods. For the spot agar test, a 2 μL-aliquot from each Lactobacillus strain suspension (approximately 7 log CFU/mL) cultivated overnight in MRS broth under anaerobiosis (Anaerobic System Anaerogen, Oxoid) was spotted on the surface of MRS agar containing 0.2% (w/v) glucose and 1.2% (w/v) agar and incubated anaerobically for 24 h at 37°C. A 1 mL-aliquot of each indicator bacterium suspension was then mixed with 18 mL of soft BHI agar (0.7% agar) (final viable count of approx. 5 log CFU/mL) and poured over the spot-inoculated MRS agar. The plates were incubated aerobically at 37°C for 48 h. The antagonistic activity was recorded as the diameter (mm) of growth inhibition zones around each spot (Jacobsen et al., 1999). Uninoculated MRS agar was used as a negative control.
For the well diffusion method, Lactobacillus strains were first cultivated for 18 h in MRS broth under anaerobiosis (Anaerobic System Anaerogen, Oxoid), and the supernatants of these cultures were collected by centrifugation (15,000 g, 15 min, and 4°C). A 1 mL aliquot of each indicator bacterium suspension was then incorporated into 20 mL BHI soft agar plates (final viable counts of approx. 5 log CFU/mL), and 50 μL aliquots of lactobacilli supernatants were dispensed into wells (5 mm diameter and 5 mm depth; drilled using sterile glass cannulas) in BHI agar. The plates were aerobically incubated at 37°C for 48 h. After the incubation period, the antagonistic activity was recorded as the diameter (mm) of growth inhibition zones around each well. In this assay, MRS broth was used as the negative control (Vitali et al., 2012).
In both, the spot agar and well diffusion assays, a free growth inhibition zone with a diameter greater than 1 mm (around the spot or well) was considered as positive inhibitory activity (Jacobsen et al., 1999). All the five tested lactobacilli isolates were tested for the capability to inhibit each other.
Antibiotic Susceptibility Testing
The minimum inhibitory concentrations of ampicillin, chloramphenicol, clindamycin, erythromycin, gentamycin, kanamycin, streptomycin, and tetracycline (European Food Safety Authority (EFSA), 2012) against the selected lactobacilli strains were determined using a broth microdilution test previously described (CLSI, 2012) with a minor modification related to the growth media and incubation atmosphere. Approximately 50 μL of each antibiotic solution was dispensed into each well of a 96-well microplate containing 100 μL of MRS broth. Subsequently, a 50 μL-culture aliquot of each test lactobacilli isolate was added to each well (final viable cell count of approximately 7 log CFU/mL). The microplate was loosely wrapped with cling wrap to prevent bacterial dehydration. Each plate included a control (without antibiotic), an inoculated sample (positive control) or an uninoculated sample (negative control). The system was anaerobically (Anaerobic System Anaerogen, Oxoid) and statically incubated at 37°C for 48 h. Subsequently, the bacterial growth was visually observed, and the MIC of each antibiotic was confirmed as its lowest concentration capable of inhibiting visible bacterial growth. The MIC cut-off values of European Food Safety Authority (EFSA) (2012) were considered to categorize the lactobacilli strains as susceptible or resistant to each tested antibiotic. Each isolate was defined as susceptible when it was inhibited at a concentration (μg/mL) of a specific antibiotic equal to or lower than the established cut-off value, and each isolate was defined as resistant when it was inhibited at a concentration (μg/mL) of a specific antibiotic higher than the established cut-off value (European Food Safety Authority (EFSA), 2012).
Growth Kinetics in MRS Broth and General Edible Medium Broth
The growth kinetics of Lactobacillus strains were assessed in MRS broth and in general edible medium (GEM) broth (40 g/L glucose, 30 g/L soya peptone, 7 g/L yeast extract, and 1 g/L MgSO4.7H2O in 0.01 mol/L K-phosphate buffer; pH 6.3 ± 0.2; Saarela et al., 2004) using two different volume scales, i.e., 200 and 2000 mL. Aliquots of each lactobacilli strain suspension were inoculated (1% v/v; final viable cell count of approximately 7 log CFU/mL) in MRS or GEM broth and incubated aerobically at 37°C under stirring (150 rpm) for 48 h. At different incubation time intervals (16, 24, and 48 h), samples were taken (1 mL) and serially diluted (10−1–10−5) in sterile peptone (0.1 g/100 mL), spread plated onto MRS agar, and incubated anaerobically (Anaerobic System Anaerogen, Oxoid) at 37°C for 48 h. After the incubation period, the viable cells were counted, and the results were expressed as the log CFU/mL.
Reproducibility and Statistical Analysis
All assays were performed in triplicate in two independent experiments (repetitions), and the results are expressed as the average of the tests. Statistical analyses were performed to determine significant differences (P ≤ 0.05) among obtained results using the Student's t-test or ANOVA followed by Tukey's post hoc test. These analyses were performed using Graphpad Prism 6.0 software.
Results
Identification of LAB
A total of 50 isolates of LAB, comprising 10 isolates from each type of pulp fruit processing byproduct source, were randomly selected for identification (Table 1 and Table S1). Species belonging to Lactobacillus genus were the most predominant (41/50 isolates; 82%) identified LAB. In pineapple and barbados cherry pulp byproducts only a few isolates were identified as Lactococcus lactis (one and two isolates, respectively) and in the mango pulp byproduct, one isolate was identified as P. pentosaceus, and two isolates were identified as L. mesenteroides. The following Lactobacillus species were found for each type of fruit pulp byproduct: L. fermentum and L. nagelii in pineapple pulp byproduct; L. plantarum, L. brevis, L. fermentum, and L. nagelii in barbados cherry pulp byproduct; L. fermentum, L. casei, L. paracasei, and L. nagelii in soursop pulp byproduct; L. plantarum, L. pentosus, and L. nagelii in mango pulp byproduct; and L. fermentum in strawberry pulp byproduct. Considering the total number of isolates identified as belonging to Lactobacillus genus, the following rank for frequency of species identification was observed: L. fermentum > L. plantarum/L. nagelii > L. brevis > L. pentosus/L. paracasei/L. casei.
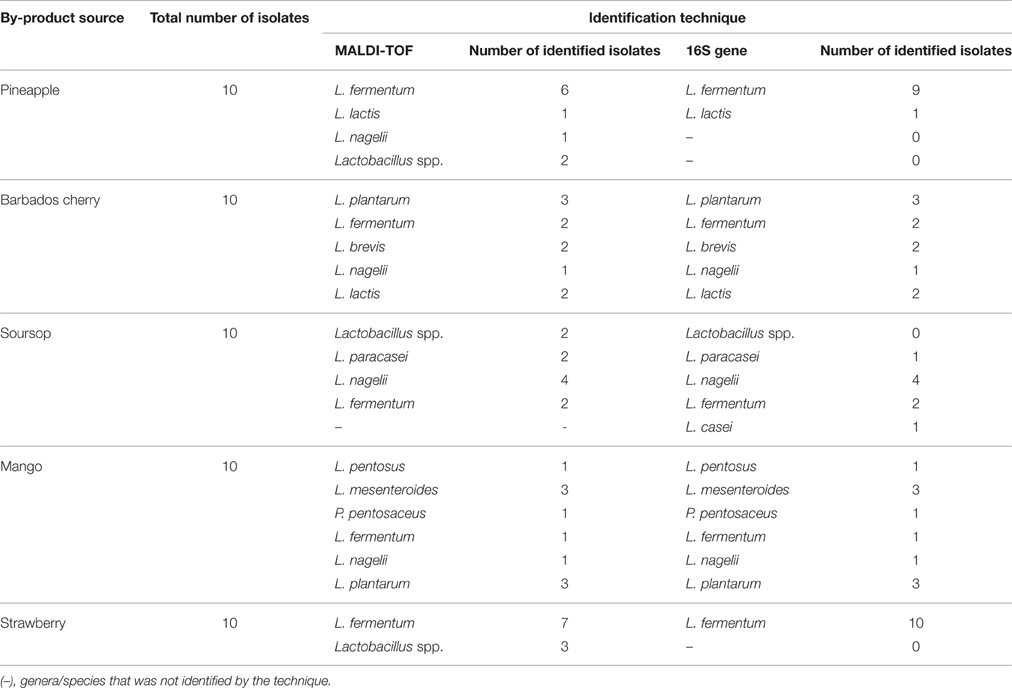
Table 1. Identification of lactic acid bacteria isolates from different fruit pulp processing byproducts (total number of isolates from each source, technique applied for identification and number of isolates identified by each applied identification technique).
The average congruency between MALDI-TOF MS and 16S rRNA gene sequence analysis in the identification of LAB species reached 86.1% (Table 2). Isolates of L. plantarum, L. brevis, L. pentosus, L. lactis, and L. mesenteroides were identified with 100% congruency. The lowest congruency (50%) was obtained for L. casei and L. paracasei. The low identification congruency (75%) for L. fermentum was due to the inability of MALDI-TOF MS to identify this species beyond the genus level (Lactobacillus spp.). All isolates that were not identified to the species level by MALDI-TOF MS were further identified as L. fermentum by 16S rRNA gene sequence analysis. In this study, there was an 86 and 100% efficiency of LAB species identification by the MALDI-TOF MS and 16S rRNA gene analysis techniques, respectively. Without considering the L. casei and L. paracasei isolates, which were distinctly identified in MALDI-TOF MS and 16S-rRNA gene analysis, both techniques showed similar 100% efficiency in LAB species identification.
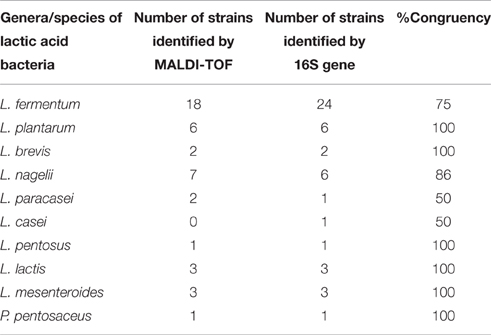
Table 2. Number of identified lactic acid species from fruit pulp processing byproducts according to each applied identification technique and congruency of identification between the applied identification techniques.
To perform the second stage of this study, five strains identified as different Lactobacillus species, namely, L. brevis 59, L. pentosus 129, L. paracasei 108, L. plantarum 49, and L. fermentum 111, were selected for inclusion in further assays.
Tolerance to Acidic Conditions and Bile Salt Concentrations
The testing of the tolerance of the Lactobacillus strains to different pH values revealed sharp decreases in viable counts (low survival rate) during the 3 h assessed incubation period. At pH 3.0, the tested Lactobacillus strains presented variable declines in viable counts over time. In this condition, L. brevis 59 presented the highest (P ≤ 0.05) viable counts during the 3 h assessed incubation period (good tolerance), while L. pentosus 129, L. paracasei 108, and L. plantarum 111 displayed sharp decreases in viable counts after the 2 or 3 h incubation. At pH 5.0, the five tested strains survived during the 3 h incubation (good tolerance) (Table 3). In most cases, the strains displayed similar (P > 0.05) viable counts when cultivated in both pH 5.0 and 7.2.
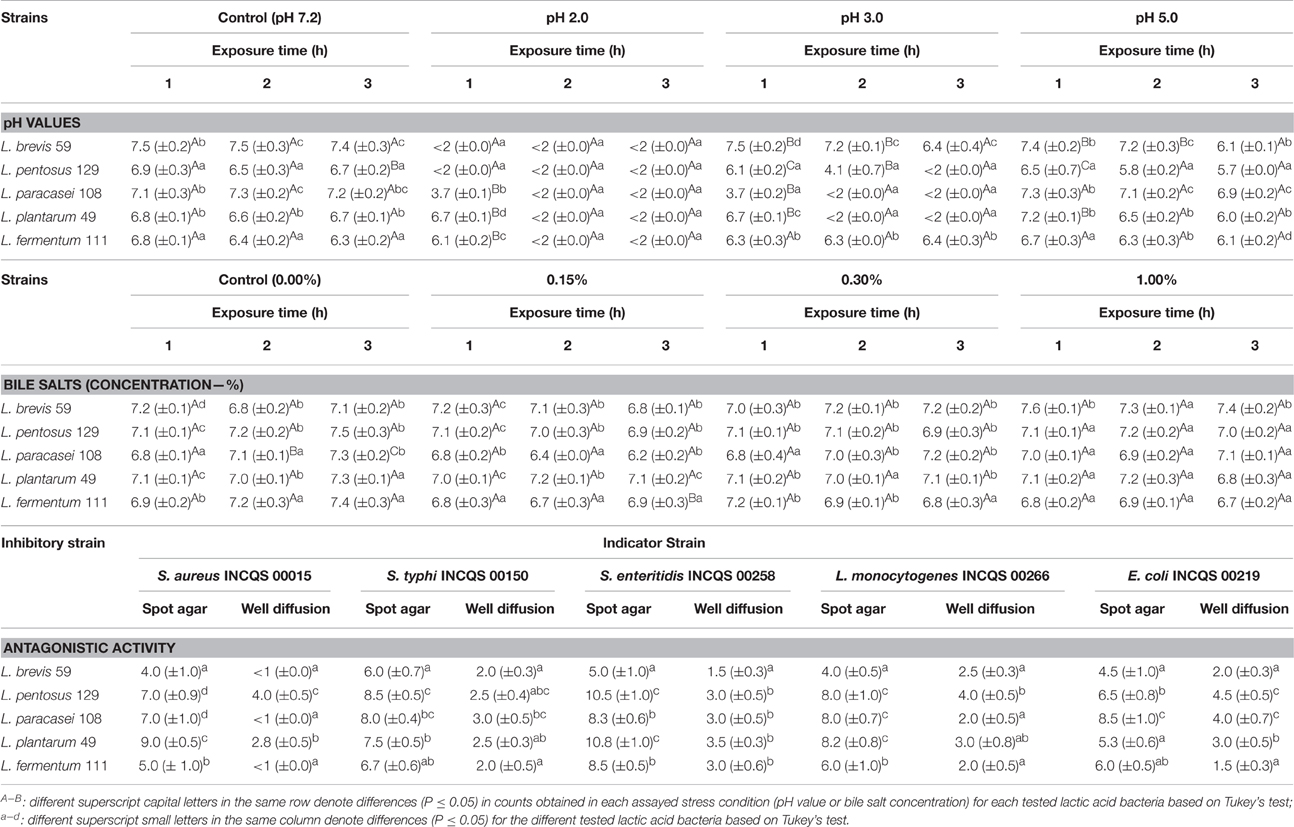
Table 3. Viable cell counts (n = 3, mean values ± standard deviation; log CFU/g) of different lactobacilli strains from pulp fruit processing byproducts when challenged with different pH values and bile salt concentrations for different exposure time periods and antagonistic activities, as expressed in diameter (mm) of growth inhibition zones (± standard deviation), toward pathogenic bacteria as measured by spot agar and well diffusion assays.
Overall, the exposure to the different bile salt concentrations did not result in decreases (P > 0.05) in the initial viable counts of the tested Lactobacillus strains during the 3 h incubation (good tolerance), and no differences (P > 0.05) were found compared to the systems not exposed to bile salts (Table 3).
Antagonistic Activity against Pathogenic Bacteria
The five tested Lactobacillus strains presented inhibitory activities against all the selected target pathogenic bacteria strains in spot agar assay, and the inhibitory effects (diameter of growth inhibition zones) varied among inhibitors and indicator strains. In the spot agar assay, the growth inhibition zone diameters displayed by the five tested Lactobacillus strains were ≥4 mm against all target pathogenic bacteria and reached ≥8.0 mm in most cases. Similarly, the cell-free supernatants of all Lactobacillus strains presented inhibitory activity against the target bacteria in the well diffusion assay with growth inhibition zone diameters varying from 1.5 to 4.5 mm. The only exceptions were the cell-free supernatants of L. brevis 59 and L. fermentum 111, which did not inhibit the growth of S. aureus INCQS 00015 (Table 3). The diameters of the growth inhibition zones were always greater in spot agar assays than in well diffusion assays.
The strongest antagonistic activities in spot agar and well diffusion assays were displayed by L. pentosus 129 and L. plantarum 49. Considering the average diameter of growth inhibition zones detected by the spot agar and well diffusion assays, the following rank of sensitivity among the target foodborne pathogenic bacteria was observed: Salmonella Enteritidis > Salmonella Typhimurium > L. monocytogenes > E. coli > S. aureus.
Antibiotic Resistance
The studied lactobacilli strains did not show resistance to ampicillin, chloramphenicol, or streptomycin, except for L. paracasei 108 that presented resistance to ampicillin. Three out of the five strains were resistant to gentamycin (L. plantarum 49, L. paracasei 108, and L. pentosus 129) and tetracycline (L. brevis 59, L. paracasei 108, and L. fermentum 111), and two strains were resistant to clindamycin (L. brevis 59 and L. paracasei 108). All of the strains were resistant to kanamycin and erythromycin (Table 4). Overall, the resistance profiles to antibiotics varied among the lactobacilli strains.
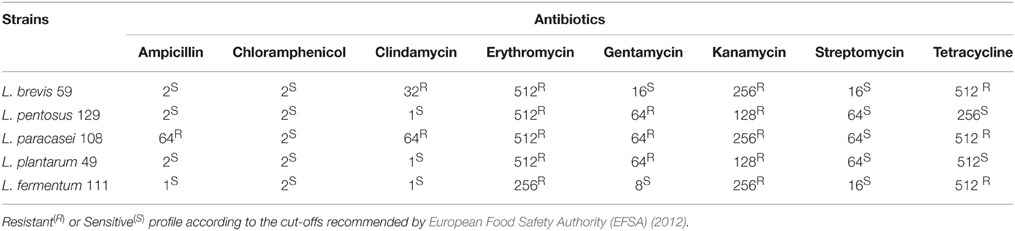
Table 4. MIC of different antibiotics against different lactobacilli strains from pulp fruit processing byproducts.
Growth in MRS and GEM Broth
The viable counts of Lactobacillus strains in MRS and GEM broth at two different medium volume scales (200 and 2000 mL) during 48 h are shown in Table 5. In most cases, the viable counts were greater (P ≤ 0.05) in MRS broth than in GEM broth during the assessed incubation period. However, the viable counts of the five strains in MRS and GEM broths were maintained at ≥6.7 log CFU/mL (6.7–7.2 CFU/mL) during the monitored incubation period, and the viable counts were close (P > 0.05) to those observed at the beginning of the experiments (approx. 7 log CFU/mL). Overall, at the two different assessed MRS and GEM broth volumes, the Lactobacillus strains presented similar viable counts (P > 0.05) during the 48 h cultivation period with increasing counts up to 16 h of cultivation followed by a decline at the later assessed cultivation periods.
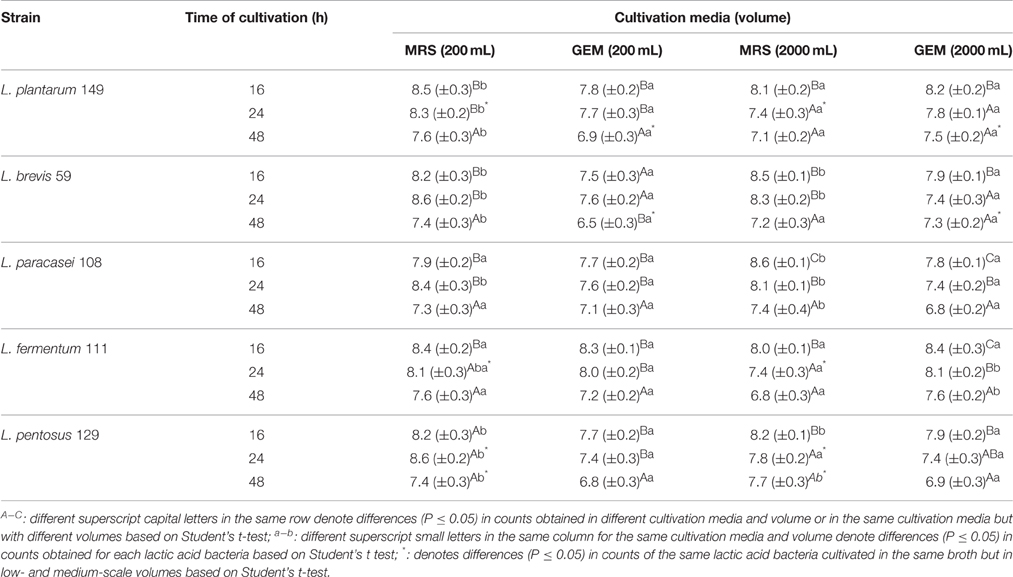
Table 5. Viable cell counts (n = 3, mean values ± standard deviation; log CFU/g) of lactobacilli isolates from pulp fruit byproducts when cultivated in de Man, Rogosa, and Sharpe (MRS) and in general edible medium (GEM) broths at low-scale (200 mL) and medium-scale (2000 mL) volumes at 37°C during 48 h (initial viable cell counts of approx. 7 log CFU/mL).
Discussion
Recently, MALDI-TOF MS has been introduced for the identification of microorganisms, and it has been suggested as a key tool in food safety and control (Jadhav et al., 2015; Nomura, 2015). However, studies showing the performance and reproducibility of this method to identify LAB species from food sources are still scarce. It has been reported that a polyphasic strategy based on molecular techniques is necessary for accurate species designation within the LAB group (Singh et al., 2009). Thus, 16S RNA gene sequence analysis has become a routine tool in LAB species identification (Dusková et al., 2012). In the present study, the results showed 100% congruency between MALDI-TOF MS and 16S rRNA gene analysis in identification of L. plantarum, L. brevis, L. pentosus, L. lactis, and L. mesenteroides, but divergence was found between the two methods for the identification of L. casei, L. paracasei, and L. fermentum. The difficulty in the identification of L. casei/paracasei by MALDI-TOF MS or 16S rRNA gene sequence analysis and cross reactions for these species in PCR-based methods have already been reported (Sisto et al., 2009; Dusková et al., 2012). Angelakis et al. (2011) found discrepancy in identification of L. paracasei instead of L. casei by MALDI-TOF MS, and this discordance was explained to be probably related with the number of species (L. casei, L. paracasei, L. rhamnosus, L. zeae) forming the L. casei group, which cannot be distinguished by conventional phenotypic properties (Klein et al., 1998; Holzapfel et al., 2001). Taxonomic controversies on to reject (Dicks et al., 1996) or to retain (Dellaglio et al., 2002) the species name L. paracasei may also difficult the denomination of a same strain. These difficulties in correct identification of lactobacilli have led to difficulties in classification of Lactobacillus strains (Schillinger et al., 2003).
Considering the view of the controversy on the nomenclature and taxonomy of L. casei-related taxa, it is possible that isolates assigned to L. casei should actually be classified as L. paracasei (Sisto et al., 2009; Dusková et al., 2012). Another interesting finding in the present study was the genus-level identification (Lactobacillus spp.) by MALDI-TOF MS of seven isolates, which were further assigned by 16S rRNA gene sequence analysis as L. fermentum. An earlier study observed similar results and reported a MALDI-TOF MS log (score) indicating probable genus identification for one Lactobacillus strain, which was further assigned by PCR as L. fermentum (Dusková et al., 2012). Thus, the discriminatory inability of MALDI-TOF MS to identify species of some Lactobacillus strains, which were further identified as L. fermentum by 16S rRNA gene analysis, suggests that clarification of MALDI-TOF MS outputs using well-known molecular techniques is still required.
The observed prevalence of Lactobacillus species in the autochthonous LAB microbiota of fruit pulp processing byproducts has already been reported for raw fruit and vegetables (Vitali et al., 2012; Argyri et al., 2013). Although, the epiphytic microbial population of plants is largely subjected to fluctuations of physicochemical and nutritional conditions, each fruit and vegetable harbors a dominant and constant microbiota (Yang et al., 2000; Dusková et al., 2012). Supporting our data, L. plantarum, L. brevis, L. fermentum, and L. paracasei are cited among the most frequent lactobacilli isolated from fruit and vegetables (Vitali et al., 2012; Argyri et al., 2013).
In vitro studies of tolerance to harsh acidic conditions and bile contents normally found in the stomach and upper parts of the intestine, respectively, together with the capability to inhibit pathogenic bacteria, have been successfully used to select potentially probiotic LAB (Mättö et al., 2006; Tuo et al., 2013). In the present study, none of the five selected Lactobacillus strains (L. brevis 59, L. pentosus 129, L. paracasei 108, L. plantarum 49, and L. fermentum 111) survived at pH 2.0 after 2 h of exposure. The tolerance assays at pH 3.0 revealed a clear separation of a group composed of three strains (L. brevis 59, L. pentosus 129, and L. fermentum 111) that presented good tolerance and another group of two strains (L. paracasei 108 and L. plantarum 49) that showed a sharp loss of cell viability after 2 h of exposure. In agreement with these results, some studies have verified that the capability of different Lactobacillus strains to maintain cell viability varies greatly between pH 2.0 and 3.0 (Jacobsen et al., 1999; Monteagudo-Mera et al., 2012). In the stomach, probiotic strains do not necessarily encounter as low pH as 2.0 because the environment can be buffered by various food components, increasing the gastric pH (Zarate et al., 2000). Components of gastric juice may also confer some protective effect on bacterial cell viability (Conway et al., 1987). In this sense, the survival of probiotic strains during gastric transit depends on both their intrinsic tolerance to the hostile conditions found therein and the food matrix harboring them.
The studied Lactobacillus strains presented good tolerance to 0.15, 0.30, and 1.00% bile salt concentrations with no delay in growth. Bile tolerance is an important characteristic in Lactobacillus species enabling them to survive, metabolize and grow during the gastrointestinal transit as well as to exert their beneficial effects on the host (Charteris et al., 1998; Argyri et al., 2013). Interestingly, some studies have found a relationship between high bile salt tolerance and the capability to hydrolyze bile salts in Lactobacillus strains (Sridev et al., 2009; Argyri et al., 2013), and this later property has been correlated to cholesterol lowering effects (Begley et al., 2006).
The five tested Lactobacillus strains displayed the capability to inhibit pathogenic bacteria, including E. coli, L. monocytogenes, Salmonella Enteritidis, Salmonella Typhimurium, and S. aureus, in the spot agar and/or well diffusion assays, but the nature of the inhibitory substance(s) remains unknown. The capability to produce antimicrobial compounds, such as organic acids, short chain fatty acids and bacteriocins, is one of the functional properties used to characterize probiotics (Argyri et al., 2013). The production of some metabolites with antimicrobial properties by probiotic bacteria can be beneficial for food preservation and the prevention of the growth of foodborne pathogens (Monteagudo-Mera et al., 2012).
Among the required properties by which specific strains can be considered as potential probiotics is that they do not harbor acquired and transferable (added genes) antibiotic resistances (European Food Safety Authority (EFSA), 2012; Gueimonde et al., 2013). The resistance of all five strains to erythromycin and kanamycin, as well as the resistance of two strains to clindamycin, are of concern. Resistance to tetracycline or gentamycin was also observed for three strains. The nature of the resistance warrants further studies before any of the resistant strains can be considered safe for human use.
Studies of the growth behavior of the selected Lactobacillus strains revealed a repeatedly similar growth pattern in both MRS and GEM broths. The similar growth behavior of the strains in MRS broth, a well-known expensive laboratory medium for Lactobacillus cultivation, and in GEM broth, an inexpensive media containing only food-grade ingredients (Saarela et al., 2006; Pimentel et al., 2012), is noteworthy. Particularly, the verified growth behavior in GEM broth encourages further studies for scale-up and optimization of the production of highly concentrated forms of the tested Lactobacillus strains in this medium for possible direct vat applications (as highly concentrated frozen cultures or as freeze-dried cultures) in formulations.
In conclusion, the results of this study showed that the LAB forming the microbiota of the studied fruit pulp processing byproducts mostly belonged to the Lactobacillus genus and included the L. fermentum, L. plantarum, L. nagelii, L. brevis, L. pentosus, L. paracasei, and L. casei species. MALDI-TOF MS and 16S rRNA gene sequence analysis revealed high congruency of LAB species identification. Overall, the selected Lactobacillus strains revealed the following desirable probiotic-related properties: tolerance to different acidic conditions and bile salt concentrations; good growth in both laboratory and edible growth media; and capacity to inhibit distinct pathogenic bacteria despite their variable susceptibility to different antibiotics. Thus, these data suggested that most of the assessed Lactobacillus strains are good candidates for further studies including fermentation and technological characteristics, survival in different food matrices and impacts on quality characteristics, as well as for in vivo studies to verify their potential health benefits. Finally, fruit pulp processing byproducts may be considered potential sources of Lactobacillus strains possessing interesting probiotic-related properties.
Author Contributions
Conceived and designed the experiments: ED, MS. Performed the experiments: EG, WL, DX, WD, KD, MD, BL, RP, OF. Analyzed the data: ED, BL, EG, MM, MS. Drafted the paper: ED, EG, MM, MS.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to CNPq (Brazil) and CAPES for the financial support (Science without Borders Program - Call Special Visiting Research - Grant 400384/2013-2).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01371
References
Ajila, C. M., Bhat, S. G., and Prasada Rao, U. J. S. (2007). Valuable components of raw and ripe peels from two Indian mango varieties. Food Chem. 102, 1006-01011. doi: 10.1016/j.foodchem.2006.06.036
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucl. Acid Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
American Public Health (APHA) (2015). Compendium for the Microbiological Examination of Foods. Washington, DC: American Public Health.
Angelakis, E., Million, M., Henry, M., and Raoult, D. (2011). Rapid and accurate bacterial identification in probiotics and yoghurts by MALDI-TOF mass spectrometry. J. Food Sci. 76, 568–572. doi: 10.1111/j.1750-3841.2011.02369.x
Araújo, K. L. G. V., Magnani, M., Nascimento, J. A., Souza, A. L., Epaminondas, P. S., Souza, A. L., et al. (2014). Antioxidant activity of co-products from guava, mango and barbados cherry produced in the Brazilian Northeast. Molecules 19, 3110–3119. doi: 10.3390/molecules19033110
Argyri, A. A., Zoumpopoulou, G., Karatizas, K. A., Tsakalidou, E., Nychas, G. J. E., Paganou, E. Z., et al. (2013). Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 33, 282–291. doi: 10.1016/j.fm.2012.10.005
Ayala-Zavala, J. F., Rosas-Domínguez, C., Vega-Vega, V., and González-Aguilar, G. A. (2010). Antioxidant enrichment and antimicrobial protection of fresh-cut fruits using their own byproducts: looking for integral exploitation. J. Food Sci. 25, 175-181. doi: 10.1111/j.1750-3841.2010.01792.x
Ayala-Zavala, J. F., Vega-Vega, V., Rosas-Domínguez, C., Palafox-Carlos, H., Villa-Rodriguez, J. A., Siddiqui, W., et al. (2011). Agro-industrial potential of exotic fruit byproducts as a source of food additives. Food Res. Int. 44, 1866–1874. doi: 10.1016/j.foodres.2011.02.021
Begley, M., Hill, C., and Cahan, C. G. (2006). Bile salts hydrolase activity in probiotics. Appl. Environ. Microbiol. 72, 1729–1738. doi: 10.1128/AEM.72.3.1729-1738.2006
Bizzini, A., Jaton, K., Romo, D., Bille, J., Prod'hom, G., and Greub, G. (2011). Matrix-assisted laser desorption ionization–time of flight mass spectrometry as an alternative to 16S rRNA gene sequencing for identification of difficult-to-identify bacterial strains. J. Clin. Microbiol. 49, 693–696. doi: 10.1128/JCM.01463-10
Charteris, W. P., Kelly, P. M., Morelli, L., and Collins, J. K. (1998). Antibiotic susceptibility of potentially probiotic Lactobacillus species. J. Food Prot. 61, 1636–1643.
CLSI (2012). Performance Standards of Antimicrobial Susceptibility Testing; Twenty-Second Informational Supplement. CLSI document M100-22. Wayne, PA: Clinical and Laboratory Standards Institute.
Conway, P. L., Gorbach, M. S. I., and Goldin, B. R. (1987). Survival of lactic acid bacteria in the human stomach and adhesion to intestinal cells. J. Dairy Sci. 70, 1–12. doi: 10.3168/jds.S0022-0302(87)79974-3
Dellaglio, F., Felis, G. E., and Torriani, S. (2002). The status of the species Lactobacillus casei (Orla-Jensen 1916) Hansen and Lessel 1971 and Lactobacillus paracasei Collins et al. 1989. Request for an opinion. Int. J. Syst. Evol. Microbiol. 52, 285–287. doi: 10.1099/00207713-52-1-285
Di Cagno, R., Cardinali, G., Minervini, G., Antonielli, L., Rizzello, C. G., Ricciuti, P., et al. (2010a). Taxonomic structure of the yeasts and lactic acid bacteria microbiota of pineapple (Ananas comosus L. Merr.) and use of autochthonous starters for minimally processing. Food Microbiol. 27, 381–389. doi: 10.1016/j.fm.2009.11.012
Di Cagno, R., Minervini, G., Rizzello, C. G., De Angelis, M., and Gobbetti, M. (2011a). Effect of lactic acid fermentation on antioxidant, texture, color and sensory properties of red and green smoothies. Food Microbiol. 28, 1062–1071. doi: 10.1016/j.fm.2011.02.011
Di Cagno, R., Minervini, G., Rizzello, C. G., Lovino, R., Servili, M., Taticchi, A., et al. (2011b). Exploitation of sweet cherry (Prunus avium L.) puree added of stem infusion through fermentation by selected autochthonous lactic acid bacteria. Food Microbiol. 28, 900–909. doi: 10.1016/j.fm.2010.12.008
Di Cagno, R., Minervini, G., Sgarbi, E., Lazzi, C., Bernini, V., Neviani, E., et al. (2010b). Comparison of phenotypic (Biolog System) and genotypic (random amplified polymorphic DNA-polymerase chain reaction, RAPD-PCR, and amplified fragment length polymorphism, AFLP) methods for typing Lactobacillus plantarum isolates from raw vegetables and fruits. Int. J. Food Microbiol. 143, 246–253. doi: 10.1016/j.ijfoodmicro.2010.08.018
Di Cagno, R., Surico, R. F., Paradiso, A., De Angelis, M., Salmon, J. C., Buchin, S., et al. (2008b). Effect of autochthonous lactic acid bacteria starters on health-promoting and sensory properties of tomato juices. Int. J. Food Microbiol. 128, 473–483. doi: 10.1016/j.ijfoodmicro.2008.10.017
Di Cagno, R., Surico, R. F., Siragusa, S., De Angelis, M., Paradiso, A., Minervini, F., et al. (2008a). Selection and use of autochthonous mixed starter for lactic acid fermentation of carrots, French beans or marrows. Int. J. Food Microbiol. 127, 220–228. doi: 10.1016/j.ijfoodmicro.2008.07.010
Dicks, L. M., Du Plessis, E. M., Dellaglio, F., and Lauer, E. (1996). Reclassification of Lactobacillus casei subsp. casei ATCC 393 and Lactobacillus rhamnosus ATCC 15820 as Lactobacillus zeae nom. rev., designation of ATCC 334 as the neotype of L. casei subsp. casei, and rejection of the name Lactobacillus paracasei. Int. J. Syst. Bacteriol. 46, 337–340. doi: 10.1099/00207713-46-1-337
Dusková, M., Sedo, O., KsiCová, K., Zdrahál, Z., and Karpísková, R. (2012). Identification of lactobacilli isolated from food by genotypic methods and MALDI-TOF MS. Int. J. Food Microbiol. 159, 107–114. doi: 10.1016/j.ijfoodmicro.2012.07.029
European Food Safety Authority (EFSA) (2012). Scientific opinion. Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 10:2740. doi: 10.2903/j.efsa.2012.2740
FAO/WHO (2006). Probiotics in Food, Health and Nutritional Properties and Guidelines for Evaluation. Rome: FAO Food and Nutritional Paper.
Freiwald, A., and Sauer, S. (2009). Phylogenetic classification and identification of bacteria by mass spectrometry. Nat. Protoc. 4, 732–742. doi: 10.1038/nprot.2009.37
García-Ruiz, A., de Llano, D. G., Esteban-Fernandez, A., Requena, T., Bartolome, B., and Moreno-Arribas, M. V. (2014). Assessment of probiotic properties in lactic acid bacteria isolated from wine. Food Microbiol. 44, 220–225. doi: 10.1016/j.fm.2014.06.015
Gevers, D., Cohan, F. M., Lawrence, J. G., Spratt, B. G., Coenye, T., Feil, E. J., et al. (2005). Re-evaluating prokaryotic species. Nat. Rev. Microbiol. 3, 733–739. doi: 10.1038/nrmicro1236
Gueimonde, M., Sánchez, B. G., de Los Reyes-Gavilán, C., and Margolles, A. (2013). Antibiotic resistance in probiotic bacteria. Front. Microbiol. 18:202. doi: 10.3389/fmicb.2013.00202
Guo, X. H., Kim, J. M., Nam, H. M., Park, S. Y., and Kim, J. M. (2010). Screening lactic acid bacteria from swine origins for multistrain probiotics based on in vitro functional properties. Anaerobe 16, 321–326. doi: 10.1016/j.anaerobe.2010.03.006
Holzapfel, W. H., Haberer, P., Geisen, R., Bjorkroth, J., and Schillinger, U. (2001). Taxonomy and important features of probiotic microorganisms in food and nutrition. Am. J. Clin. Nutr. 73, 365S–373S.
Jacobsen, C. N., Nielsen, V. R., Hayford, A. E., Møller, P. L., Michaelsen, K. F., Paerregaard, A., et al. (1999). Screening of probiotic activities of forty-seven strains of Lactobacillus spp. by in vitro techniques and evaluation of the colonization ability of five selected strains in humans. Appl. Environ. Microbiol. 65, 4949–4956.
Jadhav, S., Gulati, V., Fox, E. M., Karpe, A., Beale, D. J., Sevior, D., et al. (2015). Rapid identification and source-tracking of Listeria monocytogenes using MALDI-TOF mass spectrometry. Int. J. Food Microbiol. 202, 1–9. doi: 10.1016/j.ijfoodmicro.2015.01.023
Klein, G., Pack, A., Bonaparte, C., and Reuter, G. (1998). Taxonomy and physiology of probiotic lactic acid bacteria. Int. J. Food Microbiol. 41, 103–125. doi: 10.1016/S0168-1605(98)00049-X
Mättö, J., Alakomi, H.-L., Virkajärvi, I., and Saarela, M. (2006). Influence of processing conditions on Bifidobacterium animalis sbsp. lactis functionality with a special focus on acid tolerance and factors affecting it. Int. Dairy J. 16, 1029–1037. doi: 10.1016/j.idairyj.2005.10.014
Monteagudo-Mera, A., Rodrígues-Aparício, L., Rúa, J., Martínez-Blanco, H., Navasa, N., García-Armesto, M. R., et al. (2012). In vitro evaluation of physiological probiotic properties of different lactic acid bacteria strains of dairy and human origin. J. Funct. Foods 4, 531–541. doi: 10.1016/j.jff.2012.02.014
Naeem, M., Ilyas, M., Haider, S., Baig, S., and Saleem, M. (2012). Isolation characterization and identification of lactic acid bacteria from fruit juices and their efficacy against antibiotics. Pak. J. Bot. 44, 323–328.
Nomura, F. (2015). Proteome-based bacterial identification using matrix-assisted laser desorption ionization–time of flight mass spectrometry (MALDI-TOFMS): a revolutionary shift in clinical diagnostic microbiology. Biochem. Biophys. Acta 1854, 528–537. doi: 10.1016/j.bbapap.2014.10.022
Peres, C. M., Peres, C., Hernández-Mendoza, A., and Malcata, F. X. (2012). Review on fermented plant materials as carriers and sources of potentially probiotic lactic acid bacteria - With an emphasis on table olives. Trends Food Sci. Technol. 26, 31–42. doi: 10.1016/j.tifs.2012.01.006
Pimentel, L. L., Mättö, J., Malcata, F. X., Pintado, M. E., and Saarela, M. (2012). Survival of potentially probiotic enterococci in dairy matrices and in the human gastrointestinal tract. Int. Dairy J. 27, 53–57. doi: 10.1016/j.idairyj.2012.07.008
Ranadheera, R. D. C. S., Baines, S. K., and Adams, M. C. (2010). Importance of food in probiotic efficacy. Food Res. Int. 43, 1–7. doi: 10.1016/j.foodres.2009.09.009
Rufino, M. S. M., Alves, R. E., de Brito, E. S., Perez-Jimenez, J., Saura-Calixto, F., and Mancini-Filho, J. (2010). Bioactive compounds and antioxidant capacities of 18 non-traditional tropical fruits from Brazil. Food Chem. 121, 996–1002. doi: 10.1016/j.foodchem.2010.01.037
Saarela, M., Rantala, M., Hallamaa, K., Nohynek, L., Virkajärvi, I., and Mättö, J. (2004). Stationary-phase acid and heat treatments for improvement of the viability of probiotic lactobacilli and bifidobacteria. J. Appl. Microbiol. 96, 1205–1214. doi: 10.1111/j.1365-2672.2004.02286.x
Saarela, M., Virkajärvi, I., Alakomi, H.-L., Sigvart-Mattila, P., and Mätto, J. (2006). Stability and functionality of freeze-dried probiotic Bifidobacterium cells during storage in juice and milk. Int. Dairy J. 16, 1477–1482. doi: 10.1016/j.idairyj.2005.12.007
Schillinger, U., Yousif, N. M., Sesar, L., and Franz, C. M. (2003). Use of group-specific and RAPD-PCR analyses for rapid differentiation of Lactobacillus strains from probiotic yogurts. Curr. Microbiol. 47, 453–456. doi: 10.1007/s00284-003-4067-8
Silva, L. M. R., Figueiredo, E. A. T., Ricardo, N. M. P. S., Vieira, I. G. P., Figueiredo, R. W., Brasil, I. M., et al. (2014). Quantification of bioactive compounds in pulps and by-products of tropical fruits from Brazil. Food Chem. 143, 398–404. doi: 10.1016/j.foodchem.2013.08.001
Singh, S., Goswami, P., Singh, R., and Heller, K. J. (2009). Application of molecular identification tools for Lactobacillus, with a focus on discrimination between closely related species: a review. LWT Food Sci. Technol. 42, 448–457. doi: 10.1016/j.lwt.2008.05.019
Sisto, A., De Bellis, P., Visconti, A., Morelli, L., and Lavermicocca, P. (2009). Development of a PCR assay for the strain-specific identification of probiotic strain Lactobacillus paracasei IMPC2.1. Int. J. Food Microbiol. 136, 59–65. doi: 10.1016/j.ijfoodmicro.2009.09.017
Sridev, N., Vishwe, P., and Prabhune, A. (2009). Hypocholesterolemic effect of bile salt hydrolase from Lactobacillus buchneri ATCC 4005. Food Res. Int. 42, 516–520. doi: 10.1016/j.foodres.2009.02.016
Tuo, Y., Zhang, W., Zhang, L., Ai, L., Zhang, Y., and Han, X. (2013). Study of probiotic potential of four wild Lactobacillus strains. Anaerobe 21, 22–27. doi: 10.1016/j.anaerobe.2013.03.007
Vitali, B., Minervini, G., Rizzello, C. G., Spisni, E., Maccaferri, S., Brigidi, P., et al. (2012). Novel probiotic candidates for humans isolated from raw fruits and vegetables. Food Microbiol. 31, 116–125. doi: 10.1016/j.fm.2011.12.027
Welker, M. (2011). Proteomics for routine identification of microorganisms. Proteomics 11, 3143–3153. doi: 10.1002/pmic.201100049
Yang, C. H., Crowley, D. E., Borneman, J., and Keen, N. T. (2000). Microbial phyllosphere populations are more complex than previously realized. Microb. Ecol. 98, 3889–3894. doi: 10.1073/pnas.051633898.
Yang, J., Cao, Y., Cai, Y., and Terada, F. (2010). Natural populations of lactic acid bacteria isolated from vegetable residues and silage fermentation. J. Dairy Sci. 93, 3136–3145. doi: 10.3168/jds.2009-2898
Keywords: fruit, 16S rRNA gene sequencing, MALDI-TOF profiling, Lactobacillus, probiotic
Citation: Garcia EF, Luciano WA, Xavier DE, da Costa WCA, de Sousa Oliveira K, Franco OL, de Morais Júnior MA, Lucena BTL, Picão RC, Magnani M, Saarela M and de Souza EL (2016) Identification of Lactic Acid Bacteria in Fruit Pulp Processing Byproducts and Potential Probiotic Properties of Selected Lactobacillus Strains. Front. Microbiol. 7:1371. doi: 10.3389/fmicb.2016.01371
Received: 02 June 2016; Accepted: 18 August 2016;
Published: 30 August 2016.
Edited by:
Andrea Gomez-Zavaglia, Center for Research and Development in Food Cryotechnology (CIDCA, CONICET), ArgentinaReviewed by:
Fausto Gardini, University of Bologna, ItalyLucia Victoria Cassani, National University of Mar del Plata, Argentina
Copyright © 2016 Garcia, Luciano, Xavier, da Costa, de Sousa Oliveira, Franco, de Morais Júnior, Lucena, Picão, Magnani, Saarela and de Souza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Evandro L. de Souza, ZXZhbmRyb2xlaXRlc291emFAY2NzLnVmcGIuYnI=