 Abdoul-Salam Ouedraogo1,2,3,4*
Abdoul-Salam Ouedraogo1,2,3,4* Catherine Dunyach-Remy5,6Aimée Kissou1Soufiane Sanou1Armel Poda1Carole G. Kyelem1
Catherine Dunyach-Remy5,6Aimée Kissou1Soufiane Sanou1Armel Poda1Carole G. Kyelem1 Jérôme Solassol7,8Anne-Laure Bañuls9Philippe Van De Perre2,3,4
Jérôme Solassol7,8Anne-Laure Bañuls9Philippe Van De Perre2,3,4 Rasmata Ouédraogo1Hélène Jean-Pierre2
Rasmata Ouédraogo1Hélène Jean-Pierre2 Jean-Philippe Lavigne5,6†Sylvain Godreuil2,3,4†
Jean-Philippe Lavigne5,6†Sylvain Godreuil2,3,4†- 1Centre Hospitalier Universitaire Souro Sanou, Bobo Dioulasso, Burkina Faso
- 2Département de Bactériologie-Virologie, Centre Hospitalier Universitaire de Montpellier, Montpellier, France
- 3Université de Montpellier, Montpellier, France
- 4Institut National de la Santé et de la Recherche Médicale U1058, Infection by HIV and by Agents with Mucocutaneous Tropism: From Pathogenesis to Prevention, Montpellier, France
- 5Institut National de la Santé et de la Recherche Médicale U1047, Université de Montpellier, Nîmes, France
- 6Service de Microbiologie, Centre Hospitalier Universitaire Caremeau, Nîmes, France
- 7Department of Biopathology, Centre Hospitalier Universitaire Montpellier, Montpellier, France
- 8Department of Clinical Oncoproteomics, Montpellier Cancer Institute, Montpellier, France
- 9UMR MIVEGEC (IRD 224 - Centre National de la Recherche Scientifique 5290 - Université de Montpellier), Montpellier, France
The objectives of the present study were to investigate the rate of S.aureus nasal carriage and molecular characteristics in hospital and community settings in Bobo Dioulasso, Burkina Faso. Nasal samples (n = 219) were collected from 116 healthy volunteers and 103 hospitalized patients in July and August 2014. Samples were first screened using CHROMagar Staph aureus chromogenic agar plates, and S. aureus strains were identified by mass spectrometry. Antibiotic susceptibility was tested using the disk diffusion method on Müller-Hinton agar. All S. aureus isolates were genotyped using DNA microarray. Overall, the rate of S. aureus nasal carriage was 32.9% (72/219) with 29% in healthy volunteers and 37% in hospital patients. Among the S. aureus isolates, only four methicillin-resistant S. aureus (MRSA) strains were identified and all in hospital patients (3.9%). The 72 S. aureus isolates from nasal samples belonged to 16 different clonal complexes, particularly to CC 152-MSSA (22 clones) and CC1-MSSA (nine clones). Two clones were significantly associated with community settings: CC1-MSSA and CC45-MSSA. The MRSA strains belonged to the ST88-MRSA-IV or the CC8-MRSA-V complex. A very high prevalence of toxinogenic strains 52.2% (36/69), containing Panton-Valentine leucocidin- and EDIN-encoding genes, was identified among the S. aureus isolates in community and hospital settings. This study provides the first characterization of S. aureus clones and their genetic characteristics in Burkina Faso. Altogether, it highlights the low prevalence of antimicrobial resistance, high diversity of methicillin-sensitive S. aureus clones and high frequency of toxinogenic S. aureus strains.
Introduction
Staphylococcus aureus is both a human commensal and a frequent cause of clinically important infections in hospital and community settings. It colonizes about one third of healthy humans and is most often found in the nose (Kaspar et al., 2016). As pathogenic agent, S. aureus causes a wide range of infections (e.g., skin and soft tissue infections, sepsis, pneumonia, osteomyelitis, endocarditis; Shittu et al., 2011). Epidemiological studies of staphylococcal infections showed that nasal carriage is a risk factor and usually the origin of the infection (Wertheim et al., 2004). S. aureus treatment is complicated by the global spread of methicillin-resistant S. aureus (MRSA) strains (Holden et al., 2013).
Studies on the worldwide spread of S. aureus indicated that clone predominance varies according to the continent: ST300 in USA, ST59 in Asia, ST30/USA1100 in Southwest Pacific Oceania, ST93 in Queensland and ST80 in European countries (Fluit et al., 2015). In Africa, the limited epidemiological data suggest that S. aureus clone distribution is particularly heterogeneous, possibly due to the huge cultural and geographical diversity. Similarly, toxinogenic strains are unequally distributed in the different countries. For example, the prevalence of S. aureus strains that produce Panton-Valentine Leukocidin (PVL) is of 0.3% in South Africa and of 100% in Tunisia (Schaumburg et al., 2014; Abdulgader et al., 2015). Conversely, no data are available on the distribution of S. aureus strains that produce epidermal differentiation inhibitor (EDIN), one of the main virulence factors involved in S. aureus diffusion. As several pathogenic S. aureus clones, such as the European clone ST80-MRSA-IV (Messad et al., 2013), express EDIN or EDIN-like exotoxins, EDIN-positive strains could also be present in the African continent, especially in Maghreb where the ST80 clone is very common.
Few studies focused on S. aureus in hospital or community settings in Burkina Faso, a low-income country of West Africa, and nothing is known about clone circulation. The objectives of the present study were to characterize the S. aureus isolates in nasal specimens from hospitalized and healthy volunteers in Bobo Dioulasso, Burkina Faso.
Materials and Methods
Patients, Specimen Collection, and Ethical Clearance
The study was approved by the local ethics committee (Souro Sanou University Hospital board; authorization number: MS/SG/CHUSS/DG/DL 2014-171, 2 July 2014). All participants signed a written informed consent for participation in the study. From 1st July to 31th August 2014, 116 randomly selected healthy volunteers from the community and 103 patients hospitalized (not for S. aureus infection) at the Souro Sanou University Hospital (Bobo Dioulasso, Burkina Faso) for more than 48 h were included prospectively and consecutively in the study. Souro Sanou University Hospital is the major healthcare and referral center for the southern and western regions of Burkina Faso. During this period only one nasal swab sample by individuals was collected excluding the possibility to study the dynamics of SA nasal carriage over time (distinction between intermittent and persistent carriers).
Nasal specimens from healthy volunteers and hospitalized patients were collected using double-head flocked swabs (Copan®, Italy). The following epidemiological and clinical data were recorded for all participants: age, gender, antibiotic treatment during the past 3 months and hospital stays in the previous year.
Microbiological Study of S. aureus
Nasal swab samples were plated on CHROMagar Staph aureus chromogenic medium (BD Diagnostics, Sparks, Md.) and examined after 24 and 48 h of incubation at 37⋅C, following the manufacturer's recommendations. All mauve to orange/mauve colonies were considered as positive for the presence of S. aureus. The identification was confirmed by matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry (Bruker Daltonics, Germany). A multiplex real-time PCR assay that targets bacterial nuc gene to distinguish S. aureus from other Staphylococcus spp., and the mecA gene for the detection of methicillin resistance was used. In all confirmed S. aureus isolates, antibiotic susceptibilities were then determined using the disk diffusion method on Müller–Hinton agar with the following antibiotics: amoxicillin, amoxicillin-clavulanic acid, oxacillin, gentamicin, tobramycin, tetracycline, erythromycin, lincomycin, trimethoprim/sulfamethoxazole, ofloxacin, fusidic acid, fosfomycin, and vancomycin. Results were interpreted following the European Committee on Antimicrobial Susceptibility Testing (EUCAST) clinical breakpoints (Version 5.0; http://www.eucast.org/clinical_breakpoints/). Susceptibility to methicillin was screened with the cefoxitin disk diffusion method.
Oligonucleotide DNA Arrays and Genotyping
Each S. aureus strain was analyzed at the INSERM laboratory in Nîmes, France. The S. aureus genotyping kit (Alere Technologies GmbH, Germany) was used according to previously detailed protocols and procedures (Monecke et al., 2008). The DNA microarray covers 332 different target sequences, including species, capsule and agr group typing markers, resistance genes (SCCmec, mecA, mecC) and genes encoding exotoxins or Microbial Surface Components Recognizing Adhesive Matrix Molecules (MSCRAMM). The affiliation of isolates to clonal complexes (CCs) or sequence types (STs), defined based on MLST and spa-typing (Monecke et al., 2008), was determined by automated comparison of the hybridization profiles to a collection of previously characterized reference strains (Monecke et al., 2008, 2011).
Statistical Analysis
The nasal carriage rate and the frequency of MRSA strains were analyzed relative to the demographic and clinical characteristics of to the clonal complexes (CCs) and the expression of the different virulence genes, using the Fisher's exact test. Statistical analyses were performed using the S-Plus 2000 software package (Insightful Corporation, Seattle, WA, USA) and results were considered significant at p < 0.05.
Results
Prevalence and Risk Factors of S. aureus Nasal Carriage
During the study period, 219 subjects (116 healthy volunteers and 103 hospitalized patients) were enrolled. The demographic characteristics and possible risk factors for S. aureus colonization in the two groups are shown in Table 1. Overall, the participants' mean age was 24.1 years (±14.6) and 69.4% of them were males. Among healthy volunteers, 13 (11%) had been hospitalized in the previous year and 18 (16%) received antibiotics within the 3 months prior to inclusion in the study: fluoroquinolones (n = 9/18; 50%), ß-lactams (n = 7/18; 39%), or others (n = 2/18, 11%). Among the hospitalized patients, 14 (14%) had been hospitalized in the previous year. Significantly more patients than healthy volunteers had received antibiotics within the last 3 months (37 vs. 16%, p < 0.001). No significant difference could be observed concerning the type of antibiotic treatment in the two populations (Table 1).
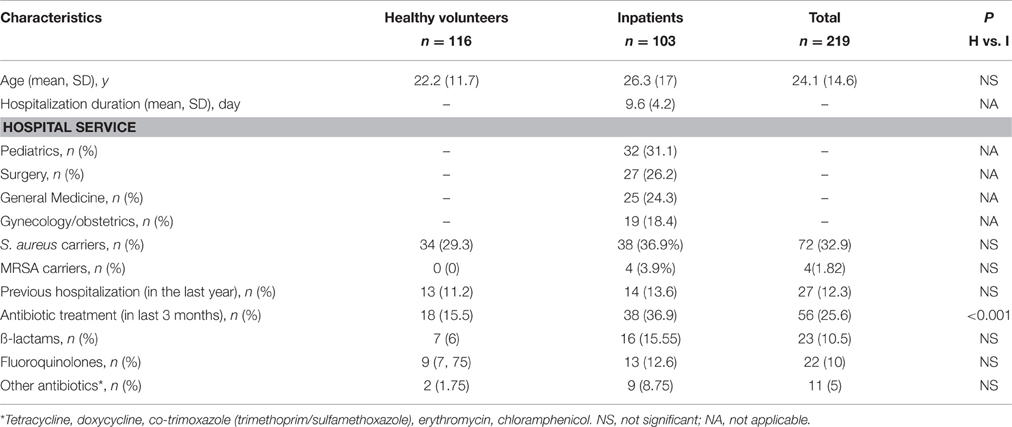
Table 1. Demographic characteristics, prevalence of Staphylococcus aureus nasal carriage and risk factors in the study population.
S. aureus was identified in the nasal samples of 32.9% (72/219) participants (29.3% of healthy volunteers and 37% of hospitalized patients; p = 0.23). Moreover, the rate of S. aureus carriage varied in the different hospital services: 59% (16 of the 27 patients from this department) in the Surgery Department, 37% (12/32) in the Pediatric, 32% (8/25) in the General Medicine, and 11% (2/19) in the Gynecology/Obstetric Department. Univariate analysis did not identify any specific risk factor associated with S. aureus nasal carriage in the whole population and the two groups separately. Among the S. aureus isolates, MRSA frequency was 0 (n = 0) in healthy volunteers and 3.9% (n = 4) in hospitalized patients (p = 0.25), representing a total occurrence of 1, 82%.
S. aureus Antimicrobial Susceptibility Testing
Analysis of the in vitro activities of antimicrobial agents against the 72 S. aureus isolates highlighted a low level of resistance to all classical anti-staphylococcal treatments (Figure 1). All isolates were susceptible to vancomycin, fosfomycin, and fusidic acid. In strains isolated from hospitalized patients, the trimethoprim/sulfamethoxazole association was always active; in strains from healthy volunteers, oxacillin, ofloxacin, and aminoglycosides (kanamycin, gentamicin, and tobramycin) were always active. The highest resistance rates were observed for amoxicillin (90%) followed by tetracycline (61%) and erythromycin (54%), Single one MRSA isolate was resistant to aminoglycosides and also to fluoroquinolones.
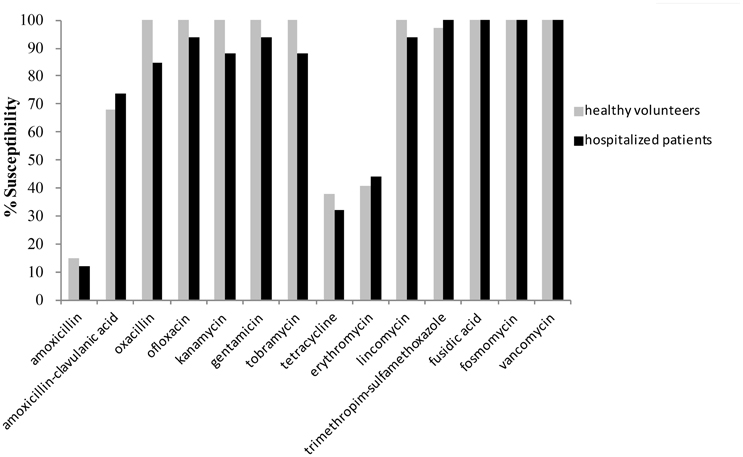
Figure 1. Percentage of antimicrobial susceptibility for the Staphylococcus aureus strains isolated from healthy volunteers (n = 34) and hospitalized patients (n = 38) (no significant differences between groups).
Distribution of S. aureus Clonal Complexes
Clonal complexes (CC) characterization in the 72 isolates allowed comparing the S. aureus populations (diversity and origin) of hospitalized patients and healthy volunteers and highlighted the great diversity of S. aureus strains circulating in the studied region (Table 2). ST 152-MSSA (n = 15) and CC1-MSSA (n = 8) were the main clones isolated in hospitalized patients and healthy volunteers, respectively. Overall, methicillin-susceptible S. aureus (MSSA) strains belonged to many different CCs: ST 152 (n = 22), CC5 (n = 14), CC1 (n = 9), ST 121 (n = 5), CC45 (n = 5), ST 707 (n = 2), ST 15 (n = 2), CC22 (n = 1), CC30 (n = 1), ST5 (n = 1), ST 30 (n = 1), CC9 (n = 1), ST 152 (n = 1). The Four MRSA strains belonged either to the CC88-MRSA-IV or to the CC8-MRSA-V lineage (no significant difference. Three isolates were classified in unknown CCs.
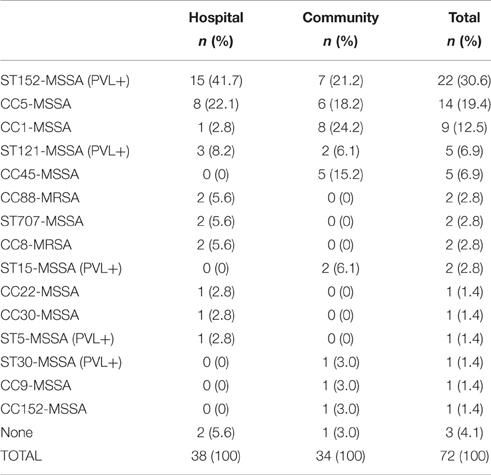
Table 2. Clonal complex distribution of the Staphylococcus aureus strains isolated from nasal samples of hospital patients and community volunteers in Burkina Faso.
Antibiotic Resistance and Virulence Gene Profiles
The antibiotic resistance gene profiles obtained by DNA array analysis were in full agreement with the data obtained by conventional susceptibility testing. All MRSA isolates detected with the cefoxitin disk diffusion test carried the mecA gene within the SCCmec mobile genetic element. MecC was not detected in any of our samples. The most prevalent macrolide resistance gene was ermC (25 isolates, 36.2%), whereas ermA was never found. Only one aminoglycoside resistance gene (aacA-aphD) was detected and only in one MRSA strain, confirming the still high antimicrobial activity of this antibiotic family. The tetK efflux gene (tetracycline resistance) was detected in 36 isolates (52.2%). No van gene was detected in agreement with the in vitro susceptibility data.
Some distinctive features regarding the virulence factor profiles were observed. Between 14 and 42% of strains expressed enterotoxin-coding genes [seg (n = 29, 42%), sek and seq (n = 12, 17.4% for each), and sea (n = 10, 14.5%)]. Conversely, genes encoding haemolysins (hlg, hlgv, hlgA, hld), intracellular adhesion proteins (icaA, icaC, icaD) or MSCRAMMs (ebpS, clfA, clfB, fnbA, fnbB, and bbp) were detected in more than half and often all strains. The agr1 allele was found in 42% of isolates (n = 29). Toxinogenic genes were frequently present in our strains. Although eta and etb (the genes encoding exfoliatin A and B) were never detected, 44.9% (n = 31) and 29% (n = 20) of isolates expressed the genes that encode PVL (lukS-PV/lukF-PV) and toxic shock syndrome toxin 1 (TSST-1; tst), respectively. Moreover, the prevalence of edin-positive isolates was 46.4% (n = 32) and specifically, 14.5% (n = 10) of all isolates were edinA-positive and 31.9% (n = 22) edin-B positive. The edin-B and etD genes were always co-expressed.
Main Characteristics of the edin+ and PVL+ S. aureus Isolates
The edin-B+ strains (n = 22) belonged to the ST152 clone (spa type t355). In this clone, edin-B, etD, and lukF/lukS-PV genes are associated. All 22 strains expressed the sarA regulon that controls edin-B expression, and belonged to the agrI cluster. No enterotoxin gene was detected. However, all edin-B+ strains expressed one or more genes encoding haemolysins (hlgv, hlgA hla, and hlb), intracellular adhesion proteins (icaA, icaD), type 5 capsular polysaccharides (cap5), immune evasion proteins (sak, scn), proteases (sspA, sspB), some MSCRAMMs (bbp, clfA, clfB, cna, ebpS, fnbA, fnbB, sdrD), defensin (mprF), and hyaluronidases (hysA1, hysA2).
The edin-A+ strains (n = 10) belonged to the clonal complexes ST73 (CC5-MSSA, n = 8), ST5 (n = 1), and ST707 (n = 1). The ST5 strain co-expressed edin-A and lukF/lukS-PV. All carried the sak, chp, scn hl, hlgA, hlgv, and hlA lukDE genes. The strains belonging to the ST73 and ST5 clones expressed many virulence genes (e.g., the egc cluster, agrII, cap5, bbp, clfA, clfB, ebpS, fib, fnbA, and fnbB). Finally, the ST707 strain harbored distinctive virulence traits (sek, seq, agrIII, fnbB alone, sdrC and sdrD).
Concerning the PVL+ strains (n = 31), they belonged to the ST152 (n = 23), ST121 (n = 4), ST15 (n = 2), ST30 (n = 1), and ST5 (n = 1) clones. They all expressed agrI and cap5, with the exception of the ST121 strains that carried agrIV and cap8. Moreover, all PVL+ strains expressed genes encoding haemolysins (hlgv, hlgA hla), immune evasion proteins (sak, scn) and MSCRAMMs (bbp, clfA, clfB, ebpS, fnbA, fnbB).
Discussion
This study provides the first characterization (clones and genetic traits) of S. aureus samples isolated from nasal specimens in Burkina Faso. Our findings highlight the low prevalence of antimicrobial resistance, the high diversity of MSSA clones and the high frequency of toxinogenic isolates among the S. aureus strains that circulate in community and hospital settings.
In Africa, S. aureus epidemiological data remain scarce. The distribution of MRSA clones is relatively heterogeneous on this continent with CC5 as the predominant clonal complex in healthcare settings. The ≪Brazilian/Hungarian≫ clone (hospital-associated MRSA ST239/ST241-III) has been identified in Maghreb, West Africa, and South African countries (Schaumburg et al., 2014; Fluit et al., 2015). The European ST80-IV clone is limited to Algeria, Egypt, and Tunisia (Schaumburg et al., 2014; Fluit et al., 2015). The community clones ST8-IV and ST88-IV have been reported both in hospital and community settings in West Africa. The ST88-MRSA-IV was also found in our population associated with the ST8-MRSA-V clone (Abdulgader et al., 2015). However, the percentage of MRSA strains (2.3%) was very low in our panel, as previously reported in Gabon (3%) (Kraef et al., 2015). On the other hand (and this is the first major finding of this study), our data highlight the high prevalence of strains harboring PVL-encoding genes (45%). These genes encode a bi-component cytotoxin that binds to the complement receptor C5a of neutrophils and that leads to severe tissue necrosis due to its cytotoxic effect on granulocytes (Dufour et al., 2002; Shallcross et al., 2013). Sub-Saharan Africa is characterized by a high dissemination of PVL+ S. aureus strains that mainly belong to three major clones: ST152, ST121, and ST15 (Rasigade et al., 2010; Schaumburg et al., 2014). Our study confirms this trend. The ST152 clone seems to be frequent and widespread in West Africa (40–60% of all strains) both in the community and in hospital settings (Ruimy et al., 2008; Shittu et al., 2012; Kraef et al., 2015). This clone has also been detected in Europe (Jappe et al., 2008; Perez-Roth et al., 2010; Krziwanek et al., 2011), Turkey (Sudagidan and Aydin, 2010), and Haiti (Rosenthal et al., 2014).
The high proportion of edin-positive MSSA (46%) strains in our population is the second main finding of this study. Indeed, such high prevalence has never been described before (14% in a previous study) (Munro et al., 2011). Three edin alleles have been characterized (Sugai et al., 1992; Yamaguchi et al., 2001, 2002). They all share similar biochemical activities toward Rho proteins, but are characterized by different genetic backgrounds. For instance, the edin-A gene is located on a plasmid, like edin-C, and is closely associated with the gene encoding exfoliative toxin B (Yamaguchi et al., 2001). EDIN proteins promote S. aureus invasiveness, by favoring its translocation to the bloodstream (Munro et al., 2010; Messad et al., 2013; Courjon et al., 2015). In our population, edin-B was the most prevalent edin gene associated with S. aureus nasal carriage. Many different S. aureus clones harbor these genes, demonstrating the wide geographical distribution of EDINs. In our population, edinB was more frequently detected in isolates belonging to the ST152 clone, like for the PVL+ strains. Therefore, our data indicate that strains belonging to this clone can co-express edinB/etD/lukSF-PV, which are the prevalent virulence markers detected in infectious strains. More worrying, in Nigeria, the ST152 clone has been associated with the SCCmec cassette (Shittu et al., 2012). These findings suggest that these strains could pose serious problems in the future, as potential reservoirs for resistance and virulence factors, and they could lead to the emergence and diffusion of PVL+ MRSA clones in Burkina Faso, with a risk of severe infections, as recently discussed (Vignaroli et al., 2016).
Nasal colonization often precedes clinical infection, frequently caused by the same isolate (von Eiff et al., 2001). Therefore, nasal carriage rates and the molecular characterization of the colonizing isolates provide important insights into the risk of S. aureus infection in Burkina Faso. The high proportion of EDIN+ and PVL+ S. aureus observed in this study indicates that conditions that increase the risk of inter-individual transmission (e.g., skin-to-skin and skin-to-fomite contacts) could represent important spreading routes in Burkina Faso. One the principal limitation of this study was the absence of distinction between intermittent and persistent carriers. It is now well-established that this distinction is important as persistent carriers are at a higher risk of developing active auto-infections than intermittent and non-carriers (Wertheim et al., 2005; Muthukrishnan et al., 2013). A larger sample of strains to study the dynamics of SA nasal carriage over time in Burkina would provide a more accurate measure of risk factors for infection according to patient status (intermittent and persistent carriers).
Classically, mortality associated with severe S. aureus infections is particularly high in emerging countries. Recent studies have identified S. aureus as the main etiological agent of many infections in Sub-Saharan Africa (Shittu et al., 2011). Microbiologists and clinicians should be aware of the threat represented by the ST152 clone and implement the necessary control measures to prevent its spread.
Author Contributions
Conceived and designed the experiments: AO, CD, HJ, PV, AB, JS, RO, JL, and SG. Performed the experiments: AO, CD, AK, SS, AP, and CK. Contributed reagents/materials/analysis tools: AO, AP, CD, AK, JL. Contributed to the writing of the manuscript: AO, HJ, AB, JS, RO, JL, and SG.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the National Institute of Health and Medical Research (INSERM). AO is a PhD student supported by the Infectiopole Sud Foundation.
References
Abdulgader, S. M., Shittu, A. O., Nicol, M. P., and Kaba, M. (2015). Molecular epidemiology of Methicillin-resistant Staphylococcus aureus in Africa: a systematic review. Front. Microbiol. 6:348. doi: 10.3389/fmicb.2015.00348
Courjon, J., Munro, P., Benito, Y., Visvikis, O., Bouchiat, C., Boyer, L., et al. (2015). EDIN-B promotes the translocation of Staphylococcus aureus to the bloodstream in the course of pneumonia. Toxins (Basel) 7, 4131–4142. doi: 10.3390/toxins7104131
Dufour, P., Gillet, Y., Bes, M., Lina, G., Vandenesch, F., Floret, D., et al. (2002). Community-acquired methicillin-resistant Staphylococcus aureus infections in France: emergence of a single clone that produces Panton-Valentine leukocidin. Clin. Infect. Dis. 35, 819–824. doi: 10.1086/342576
Fluit, A. C., Carpaij, N., Majoor, E. A., Weinstein, R. A., Aroutcheva, A., Rice, T. W., et al. (2015). Comparison of an ST80 MRSA strain from the USA with European ST80 strains. J. Antimicrob. Chemother. 70, 664–669. doi: 10.1093/jac/dku459
Holden, M. T., Hsu, L. Y., Kurt, K., Weinert, L. A., Mather, A. E., Harris, S. R., et al. (2013). A genomic portrait of the emergence, evolution, and global spread of a methicillin-resistant Staphylococcus aureus pandemic. Genome Res. 23, 653–664. doi: 10.1101/gr.147710.112
Jappe, U., Heuck, D., Strommenger, B., Wendt, C., Werner, G., Altmann, D., et al. (2008). Staphylococcus aureus in dermatology outpatients with special emphasis on community-associated methicillin-resistant strains. J. Invest. Dermatol. 128, 2655–2664. doi: 10.1038/jid.2008.133
Kaspar, U., Kriegeskorte, A., Schubert, T., Peters, G., Rudack, C., Pieper, D. H., et al. (2016). The culturome of the human nose habitats reveals individual bacterial fingerprint patterns. Environ. Microbiol. 18, 2130–2142. doi: 10.1111/1462-2920.12891
Kraef, C., Alabi, A. S., Peters, G., Becker, K., Kremsner, P. G., Rossatanga, E. G., et al. (2015). Co-detection of Panton-Valentine leukocidin encoding genes and cotrimoxazole resistance in Staphylococcus aureus in Gabon: implications for HIV-patients' care. Front. Microbiol. 6:60. doi: 10.3389/fmicb.2015.00060
Krziwanek, K., Metz-Gercek, S., and Mittermayer, H. (2011). Trends in the occurrence of MRSA strains in Upper Austria from 2006 to 2009. Clin. Microbiol. Infect. 17, 920–923. doi: 10.1111/j.1469-0691.2010.03376.x
Messad, N., Landraud, L., Canivet, B., Lina, G., Richard, J. L., Sotto, A., et al. (2013). Distribution of edin in Staphylococcus aureus isolated from diabetic foot ulcers. Clin. Microbiol. Infect. 19, 875–880. doi: 10.1111/1469-0691.12084
Monecke, S., Coombs, G., Shore, A. C., Coleman, D. C., Akpaka, P., Borg, M., et al. (2011). A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 6:e17936. doi: 10.1371/journal.pone.0017936
Monecke, S., Jatzwauk, L., Weber, S., Slickers, P., and Ehricht, R. (2008). DNA microarray-based genotyping of methicillin-resistant Staphylococcus aureus strains from Eastern Saxony. Clin. Microbiol. Infect. 14, 534–545. doi: 10.1111/j.1469-0691.2008.01986.x
Munro, P., Benchetrit, M., Nahori, M. A., Stefani, C., Clément, R., Michiels, J. F., et al. (2010). The Staphylococcus aureus epidermal cell differentiation inhibitor toxin promotes formation of infection foci in a mouse model of bacteremia. Infect. Immun. 78, 3404–3411. doi: 10.1128/IAI.00319-10
Munro, P., Clément, R., Lavigne, J. P., Pulcini, C., Lemichez, E., and Landraud, L. (2011). High prevalence of edin-C encoding RhoA-targeting toxin in clinical isolates of Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 30, 965–972. doi: 10.1007/s10096-011-1181-6
Muthukrishnan, G., Lamers, R. P., Ellis, A., Paramanandam, V., Persaud, A. B., Tafur, S., et al. (2013). Longitudinal genetic analyses of Staphylococcus aureus nasal carriage dynamics in a diverse population. BMC Infect Dis. 13:221. doi: 10.1186/1471-2334-13-221
Pérez-Roth, E., Alcoba-Flórez, J., López-Aguilar, C., Gutiérrez-González, I., Rivero-Pérez, B., and Méndez-Alvarez, S. (2010). Familial furunculosis associated with community-acquired leukocidin-positive methicillin-susceptible Staphylococcus aureus ST152. J. Clin. Microbiol. 48, 329–332. doi: 10.1128/JCM.00622-09
Rasigade, J. P., Laurent, F., Lina, G., Meugnier, H., Bes, M., Vandenesch, F., et al. (2010). Global distribution and evolution of Panton-Valentine leukocidin-positive methicillin-susceptible Staphylococcus aureus, 1981–2007. J. Infect. Dis. 201, 1589–1597. doi: 10.1086/652008
Rosenthal, M. E., Mediavilla, J., Chen, L., Sonnenfeld, J., Pierce, L., Shannon, A., et al. (2014). Molecular epidemiology of Staphylococcus aureus in post-earthquake northern Haiti. Int. J. Infect. Dis. 29, 146–151. doi: 10.1016/j.ijid.2014.08.007
Ruimy, R., Maiga, A., Armand-Lefevre, L., Maiga, I., Diallo, A., Koumaré, A. K., et al. (2008). The carriage population of Staphylococcus aureus from Mali is composed of a combination of pandemic clones and the divergent Panton-Valentine leukocidin-positive genotype ST152. J. Bacteriol. 190, 3962–3968. doi: 10.1128/JB.01947-07
Schaumburg, F., Alabi, A. S., Peters, G., and Becker, K. (2014). New epidemiology of Staphylococcus aureus infection in Africa. Clin. Microbiol. Infect. 20, 589–596. doi: 10.1111/1469-0691.12690
Shallcross, L. J., Fragaszy, E., Johnson, A. M., and Hayward, A. C. (2013). The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: a systematic review and meta-analysis. Lancet Infect. Dis. 13, 43–54. doi: 10.1016/S1473-3099(12)70238-4
Shittu, A., Oyedara, O., Abegunrin, F., Okon, K., Raji, A., Taiwo, S., et al. (2012). Characterization of methicillin-susceptible and -resistant staphylococci in the clinical setting: a multicentre study in Nigeria. BMC Infect. Dis. 12:286. doi: 10.1186/1471-2334-12-286
Shittu, A. O., Okon, K., Adesida, S., Oyedara, O., Witte, W., Strommenger, B., et al. (2011). Antibiotic resistance and molecular epidemiology of Staphylococcus aureus in Nigeria. BMC Microbiol. 11:92. doi: 10.1186/1471-2180-11-92
Sudagidan, M., and Aydin, A. (2010). Virulence properties of methicillin-susceptible Staphylococcus aureus food isolates encoding Panton-Valentine Leukocidin gene. Int. J. Food Microbiol. 138, 287–291. doi: 10.1016/j.ijfoodmicro.2010.01.023
Sugai, M., Hashimoto, K., Kikuchi, A., Inoue, S., Okumura, H., Matsumoto, K., et al. (1992). Epidermal cell differentiation inhibitor ADP-ribosylates small GTP-binding proteins and induces hyperplasia of epidermis. J. Biol. Chem. 267, 2600–2604.
Vignaroli, C., Sante, L. D., Stano, P., Varaldo, P. E., and Camporese, A. (2016). Recurrent skin infection associated with nasal carriage of Panton-Valentine leukocidin-positive methicillin-susceptible Staphylococcus aureus closely related to the EMRSA-15 clone. Future Microbiol. 11, 17–21. doi: 10.2217/fmb.15.125
von Eiff, C., Becker, K., Machka, K., Stammer, H., and Peters, G. (2001). Nasal carriage as a source of Staphylococcus aureus bacteremia. Study Group. N. Engl. J. Med. 344, 11–16. doi: 10.1056/NEJM200101043440102
Wertheim, H. F., Melles, D. C., Vos, M. C., van Leeuwen, W., van Belkum, A., Verbrugh, H. A., et al. (2005). The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 5, 751–762. doi: 10.1016/S1473-3099(05)70295-4
Wertheim, H. F., Vos, M. C., Ott, A., van Belkum, A., Voss, A., Kluytmans, J. A., et al. (2004). Risk and outcome of nosocomial Staphylococcus aureus bacteraemia in nasal carriers versus non-carriers. Lancet 364, 703–705. doi: 10.1016/S0140-6736(04)16897-9
Yamaguchi, T., Hayashi, T., Takami, H., Ohnishi, M., Murata, T., Nakayama, K., et al. (2001). Complete nucleotide sequence of a Staphylococcus aureus exfoliative toxin B plasmid and identification of a novel ADP-ribosyltransferase, EDIN-C. Infect. Immun. 69, 7760–7771. doi: 10.1128/IAI.69.12.7760-7771.2001
Keywords: carriage, EDIN, oligonucleotide array, PVL, Staphylococcus aureus
Citation: Ouedraogo A-S, Dunyach-Remy C, Kissou A, Sanou S, Poda A, Kyelem CG, Solassol J, Bañuls A-L, Van De Perre P, Ouédraogo R, Jean-Pierre H, Lavigne J-P and Godreuil S (2016) High Nasal Carriage Rate of Staphylococcus aureus Containing Panton-Valentine leukocidin- and EDIN-Encoding Genes in Community and Hospital Settings in Burkina Faso. Front. Microbiol. 7:1406. doi: 10.3389/fmicb.2016.01406
Received: 13 February 2016; Accepted: 24 August 2016;
Published: 13 September 2016.
Edited by:
Leonard Peruski, US Centers for Disease Control and Prevention, USAReviewed by:
Haider Abdul-Lateef Mousa, University of Basrah, IraqBruno Pozzetto, Jean Monnet University, France
Copyright © 2016 Ouedraogo, Dunyach-Remy, Kissou, Sanou, Poda, Kyelem, Solassol, Bañuls, Van De Perre, Ouédraogo, Jean-Pierre, Lavigne and Godreuil. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Abdoul-Salam Ouedraogo, YWJkb3VzYWwyMDAwQHlhaG9vLmZy
†Co-last authors.