 Amel Ayad1,2,3
Amel Ayad1,2,3 Mourad Drissi4
Mourad Drissi4 Claire de Curraize1
Claire de Curraize1 Chloé Dupont5
Chloé Dupont5 Alain Hartmann6Sébastien Solanas6
Alain Hartmann6Sébastien Solanas6 Eliane Siebor1
Eliane Siebor1 Lucie Amoureux1
Lucie Amoureux1 Catherine Neuwirth1*
Catherine Neuwirth1*- 1Laboratoire de Bactériologie Médicale, Plateau Technique de Biologie, Centre Hospitalier Universitaire de Dijon, Dijon, France
- 2Laboratoire Antibiotiques, Antifongiques: Physico-chimie, Synthèse et Activité Biologique, Faculté des Sciences de la Nature et de la Vie et Sciences de la Terre et de l’Univers, Université Abou Bekr Belkaid, Tlemcen, Algérie
- 3Département de Biologie, Faculté des Sciences de la Nature et de la Vie, Université Hassiba Benbouali, Chlef, Algérie
- 4Département de Biologie, Faculté des Sciences de la Nature et de la Vie et Sciences de la Terre et de l’Univers, Université Abou Bekr Belkaid, Tlemcen, Algérie
- 5UMR 5569, Equipe “Pathogénes hydriques, Santé, Environnements” PHySE, Université de Montpellier, France
- 6INRA, UMR 1347 Agroécologie, Dijon, France
The aim of this study, was to characterize the extended-spectrum-β-lactamases (ESBLs) producing clinical strains of Escherichia coli isolated between January 2009 and June 2012 from Algerian hospitals and to determine the prevalence of 16S rRNA methylase among them. Sixty-seven ESBL-producers were detected among the 239 isolates included: 52 CTX-M-15-producers, 5 CTX-M-3-producers, 5 CTX-M-1-producers, 2 CTX-M-14-producers, 2 SHV-12-producers and one TEM-167-producer. Among the ESBL–producing strains twelve harbored 16S rRNA methylase genes: 8 rmtB and 4 armA. rmtB was located on a IncFIA plasmid and armA was located either on a IncL/M or a IncFIA plasmid. RmtB-producing isolates were genotypically related and belonged to the sequence type ST 405 whereas ArmA-producing isolates belonged to ST10, ST 167, and ST 117. This first description of 16S rRNA methylases among E. coli in Algerian hospitals pointed out the necessity to establish control measures to avoid their dissemination.
Introduction
The emergence of β-lactam resistance in Escherichia coli which is the most common cause of Gram-negative infections has become a significant health concern, mainly due to the production of extended-spectrum beta-lactamases (ESBLs) (Pitout and Laupland, 2008). Indeed, CTX-M ESBLs (more than 150 enzymes) have increased significantly in Enterobacteriaceae, particularly in E. coli, in most regions of the world (Cantón and Coque, 2006). Another worrisome feature is the emergence of isolates resistant to all clinically important aminoglycosides related to the production of 16S rRNA methylases. In 2003, the first 16S rRNA methylase gene, armA, was identified (Galimand et al., 2003). To date eleven 16S rRNA methylases genes (rmtA, rmtB, rmtC, rmtD, rmtD2, rmtE, rmtF, rmtG, rmtH, armA, and npmA) have been identified, armA and rmtB being the most frequently described in Enterobacteriaceae (Galimand et al., 2003; Bogaerts et al., 2007; Yamane et al., 2007; Fritsche et al., 2008). As these genes are usually located on plasmids they can easily transfer to other bacteria (Nagasawa et al., 2014).
In Algeria, the first CTX-M enzymes have been reported in 2005 in clinical strains of Salmonella enterica from Constantine, East of Algeria (Naas et al., 2005). Since then, several surveys in the north of Algeria have reported the presence of CTX-M-type enzyme and associated resistance to non-β-lactam markers in various species of Enterobacteriaceae (Baba Ahmed-Kazi Tani and Arlet, 2014). Concerning 16S rRNA methylases only ArmA has been detected to date. It was identified for the first time in Enterobacteriaceae strains isolated from an Algerian patient transferred to Belgium (Bogaerts et al., 2007). It was subsequently identified in clinical isolates of S. enterica in Constantine associated with CTX-M-3 or CTX-M-15 (Naas et al., 2009, 2011) and in clinical strains of Salmonella non-Typhi in Annaba associated with CTX-M-15 and CMY-2 (Bouzidi et al., 2011). Recently, it was also described in clinical isolates of Klebsiella pneumoniae, Acinetobacter baumannii, Enterobacter cloacae, and Serratia marcescens (Bakour et al., 2014; Belbel et al., 2014; Batah et al., 2015; Khennouchi et al., 2015).
In this study, we have characterized the ESBL-producing clinical strains of E. coli isolated between October 2008 and June 2012 in three hospitals in Western Algeria and determined the prevalence of 16S rRNA methylase genes among them.
Materials and Methods
Bacterial Strains
Two hundred and thirty-nine isolates of E. coli collected between October 2008 and June 2012 from three different hospitals located in North-Western Algeria (Tlemcen, Sidi Bel Abbes and Oran) were included. Two hundred and thirty-seven strains were isolated from patients admitted to the intensive care unit (ICU), surgery and neurosurgery wards. Two isolates have been recovered from hospital environment. Identification was performed using the API 20E system (BioMerieux) and/or matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS).
Antibiotic Susceptibility Testing
Twenty three antibiotics (Bio-Rad) were tested (Table 1). Antibiotic susceptibility testing was determined by disk diffusion method and the minimum inhibitory concentration (MICs) of colistin was determined by a dilution method in Mueller-Hinton (MH) broth (Bio-Rad) as described by the European Committee on Antimicrobial Susceptibility Testing1. Detection of ESBL production was performed by double disk synergy test (Jarlier et al., 1988).
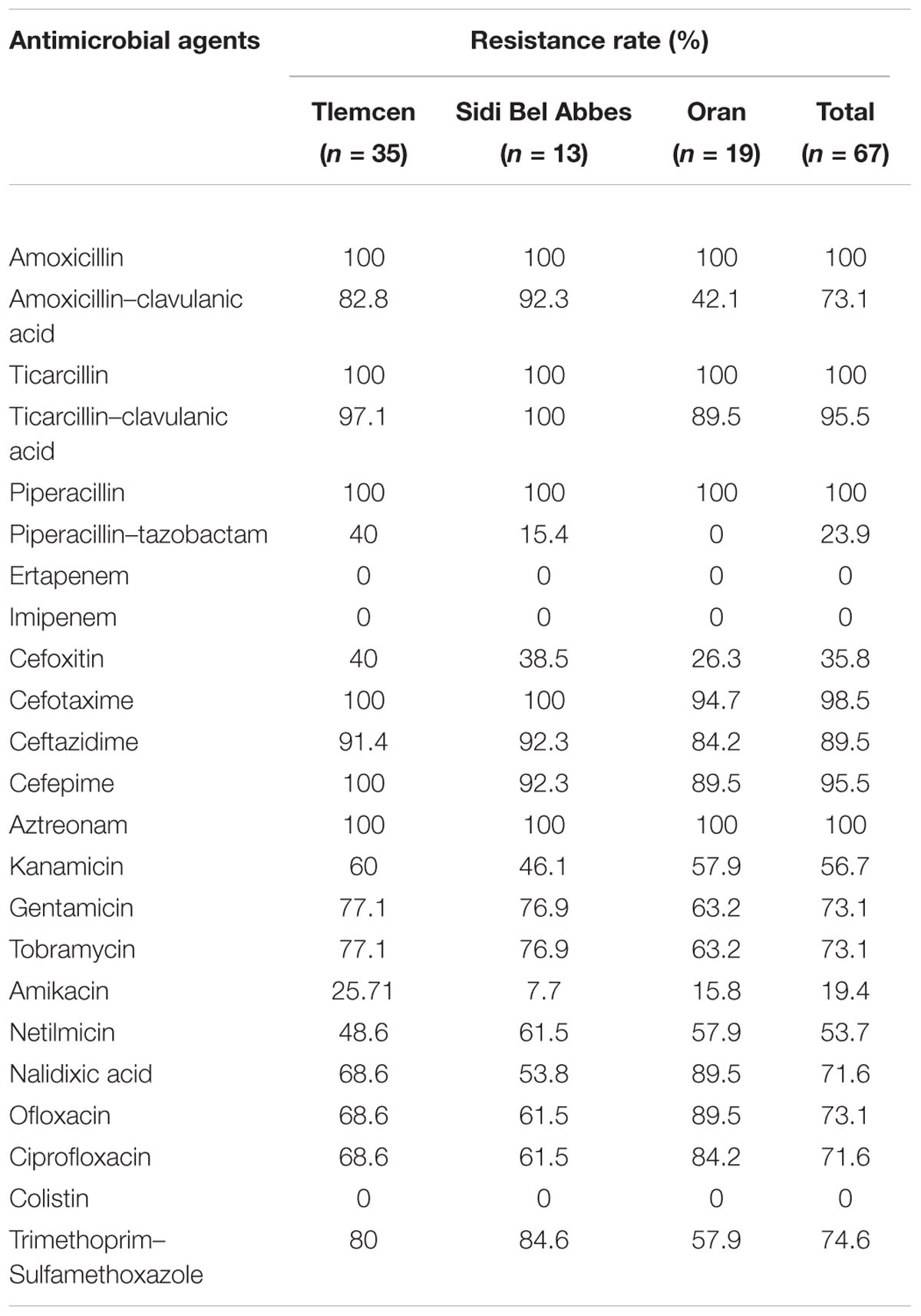
TABLE 1. Antimicrobial resistance of extended-spectrum-β-lactamases producing Escherichia coli isolated from three hospitals in Algeria.
Bacterial DNA Preparation, Gene Amplification, and Sequencing
Bacterial DNA preparation, gene amplification, sequencing, and nucleotide sequence analysis were performed as previously described (Siebor and Neuwirth, 2011). The primers used for the PCRs are listed in Table 2. All the isolates were analyzed for the presence of blaCTX-M, blaTEM, and blaSHV. The resistant strains to all aminoglycosides tested were further analyzed for the presence of armA and rmtB.

TABLE 2. Primers used for PCRs.
Genotyping of the Isolates Producing 16S rRNA Methylase
The strains were submitted to genotypic analysis by Pulsed-Field Gel Electrophoresis (PFGE) and Multilocus sequence typing (MLST). PFGE was performed following DNA digestion by XbaI as already described (Neuwirth et al., 1996). Restriction patterns were interpreted according to Tenover’s criteria (Tenover et al., 1995), and the isolates have been classified into different pulsotypes. Pulsotypes were compared by calculation of the Dice correlation coefficient with DendroUPGMA software2 and were clustered into a dendrogram by UPGMA. Strains were defined as having a clonal relationship at a criterium of 85% similarity (Ejrnaes et al., 2006). MLST was performed using seven conserved housekeeping genes of E. coli (adk, fumC, gyrB, icd, mdh, purA, and recA) as previously described (Wirth et al., 2006). The sequences types were determined by referring to the open-source software freely available at MLST Databases at the ERI, University College Cork website3.
Mating Experiments and Plasmids Analysis
Mating experiments to attempt to transfer resistance genes were carried out using rifampicin resistant E. coli K-12 C600 recipient strain as described previously (Chen et al., 2007). Transconjugants were selected on Drigalski agar plates containing amikacin (4 mg/L), gentamicin (4 mg/L), and rifampicin (200 mg/L), on Drigalski agar plates containing cefotaxime (4 mg/L) and rifampicin (200 mg/L), on Drigalski agar plates containing gentamicin (4 mg/L) and rifampicin (200 mg/L) and on Drigalski agar plates containing amikacin (30 mg/L) and rifampicin (200 mg/L).
Identification of plasmids by PCR-based replicon typing was performed on the donor strains and their transconjugants as described previously (Carattoli et al., 2005). Plasmid sizes were determined by digestion of whole-genome DNA with S1 endonuclease to linearize the plasmids and PFGE to separate them. Plasmids obtained were transferred to nylon membranes and hybridized with digoxigenin-labeled specific probes [blaCTX-M group 1, blaTEM, armA, rmtB, and replicon groups (IncL/M, IncFIA, IncFIB, and IncN)] (Barton et al., 1995).
Results
ESBL Producers
Sixty-seven ESBL-producing isolates have been detected: 35 from Tlemcen hospital, 13 from Sidi Bel Abbes hospital and 19 from Oran hospital. Isolates were mainly recovered from ICU (58.2%), followed by 26.9% from surgery department. Eighteen strains (26.9%) were recovered from wound pus, 17 (25.4%) from tracheal aspirate, 17 (25.4%) from urine and urinary catheters, 7 (10.4%) from rectal swabs, 4 (6%) from bedsores, 1 (1.5%) from catheter, 1 (1.5%) from gastric tube, and 2 (3%) from hospital environment. Among these strains 64 isolates (95.5%) carried blaCTX-M (52 blaCTX-M-15, 5 blaCTX-M-3, 5 blaCTX-M-1, 2 blaCTX-M-14), while 2 carried blaSHV-12 and 1 blaTEM-167.
All the ESBL-producing isolates remained susceptible to imipenem, ertapenem, and colistin and most of them to piperacillin-tazobactam (76%), cefoxitin (64.2%), and amikacin (80.6%). However most of the strains were resistant to all other antibiotics tested (>53%) (Table 1).
Characterization of 16S rRNA Methylase-Producers (Table 3; Supplementary Figure S1)
Twelve isolates were resistant to all aminoglycosides tested of which four strains harbored armA and 8 rmtB. ArmA was detected among CTX-M-15, CTX-M-1, and TEM-167-producers whereas rmtB was detected only among CTX-M-15-producers (Table 3). The eight RmtB-producing strains all originated from Tlemcen hospital belonged to pulsotype A and to ST 405. A wider diversity was observed among the four ArmA-producers that belonged to three different pulsotypes (B, C, and D) and three different ST: ST10 (one isolate from Sidi Bel Abbes hospital), ST167 (one from Tlemcen hospital), and ST117 (two isolates from Oran hospital).
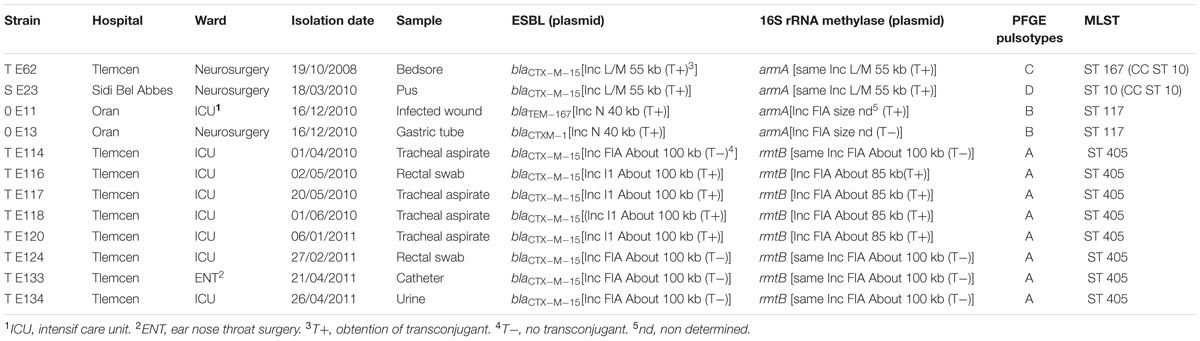
TABLE 3. Characteristics of the 16S rRNA methylase producing E. coli strains isolated in Tlemcen, Sidi Bel Abbes, and Oran hospitals, Algeria.
In the eight isolates, rmtB was located on a IncFIA plasmid. In four cases blaCTX-M-15 was located on the same plasmid while it was located on a IncI1 plasmid in the other four cases. IncFIA was transferred only in the presence of IncI1. armA was located either on a transferable IncL/M plasmid (with blaCTX-M-15) or on a IncFIA plasmid (in these two cases blaCTX-M-1 or blaTEM-167 were located on a IncN plasmid).
Discussion
Our results are in accord with previous studies and confirm that CTX-M-15 is the predominant ESBL encountered among E. coli in Algerian hospitals (Ramdani-Bouguessa et al., 2006; Baba Ahmed-Kazi Tani et al., 2013). The same situation is also observed in Tunisia (Mamlouk et al., 2006) and Morocco (Girlich et al., 2014) illustrating the large spread of CTX-M-15-producing E. coli in the north African countries. Nevertheless, the CTX-M-1, CTX-M-14, SHV-12, and TEM-167 ESBLs also detected in our study had never been reported to date among E. coli strains in Algeria. There is a single description of CTX-M-14-producing S. enterica collected from Draa-El-Mizan hospital, east of Algeria (Iabadene et al., 2009) whereas SHV-12 has already been detected in clinical isolates of K. pneumoniae and E. cloacae in Algeria (Baba Ahmed-Kazi Tani and Arlet, 2014). There is no description of TEM-167 to date, only blaTEM-167 is available in GenBank (FJ360884).
Our study further highlights the emergence of 16S rRNA methylase enzymes among ESBL-producing E. coli strains in the west of Algeria. Among our collection, the prevalence rate of 16S rRNA methylase determinants was 18%, which is very high compared to those reported in Japan (0.03%) (Yamane et al., 2007), in Turkey (0.7%) (Bercot et al., 2010), in France (1.3%) (Bercot et al., 2008), and in China (10%) (Wu et al., 2009). The detection of RmtB-producing E. coli is an interesting finding of our study. Indeed the RmtB enzyme has never been reported in Algeria. The eight isolates producing RmtB were genotypically related and belonged to ST405. Interestingly ST405 associated or not with production of CTX-M-15 ESBL is increasingly reported worldwide and belongs to the high-risk clones in the dissemination of antibiotic resistance (Woodford et al., 2011). The association of blaCTX-M-15 and rmtB on a conjugative plasmid (also carrying the plasmid-mediated fluoroquinolone efflux pump gene qepA) in one ST405 E. coli isolate has been reported in the United States (Tian et al., 2011) but the plasmid was not typed by PCR-based replicon.
All RmtB-producers have been recovered in Tlemcen Hospital and seven of them were collected in ICU suggesting patient cross contamination in this ward.
Therefore there is a need to reinforce the compliance of health-care workers with the infection control measures. The epidemiological results concerning ArmA-producers is quite different: the four isolates recovered in three hospitals in Western Algeria belonged to three different pulsotypes suggesting horizontal spread of the resistance determinants among E. coli. There are several reports of ArmA-producers among different species of Enterobacteriaceae and also A. baumannii from Algerian Hospitals (Bogaerts et al., 2007; Naas et al., 2009, 2011; Bouzidi et al., 2011; Bakour et al., 2014; Belbel et al., 2014; Batah et al., 2015; Khennouchi et al., 2015). The production of 16S rRNA methylase together with the production of ESBL observed in our study (RmtB plus CTX-M-15 or ArmA plus CTX-M-15) corresponds to an emerging and threatening combination reported worldwide (Bogaerts et al., 2007; Khennouchi et al., 2015). To our best knowledge two combinations detected in our study were not reported before: ArmA plus CTX-M-1 and ArmA plus TEM-167 in two isolates clonally related (OE11 and OE13) recovered simultaneously from Oran’s hospital.
Our data indicated that the diversity of ESBL among E. coli is growing in Algerian hospitals (CTX-M-1, CTX-M-14, CTX-M-15, TEM-167, and SHV-12) and that new mechanisms are emerging (16S rRNA methylases). In our study 11 out of 12 isolates that co-produced ESBL and methylase were resistant to all antibiotics tested except carbapenems and colistin, making of them the only available therapeutic option for these strains. Therefore the public health impact is potentially important.
Conclusion
There is a need for implementation of antimicrobial stewardship programs in Algerian hospitals to make the best use of the available antibiotics and to avoid as much as possible the unreasonable use of broad-spectrum molecules leading to the emergence of resistant isolates. Also strict control measures have to be elaborated to avoid the spread of multiresistant organisms such as those described here that co-produced ESBL and 16S rRNA methylase.
Author Contributions
AA: Selected the isolates, performed the characterization of the ESBL, of the methylase, the analysis by PFGE, the MLST analysis and manuscript preparation. MD: Designed the study. CdC and LA: Participated to genotyping and interpretation. ES: Sequence analysis and manuscript preparation. AH and SS: Performed plasmid analysis. CD: Performed PFGE analysis (dendrogram). CN: Choice of the isolates, choice of the tools, interpretation of the results and manuscript preparation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01409
Footnotes
References
Baba Ahmed-Kazi Tani, Z., and Arlet, G. (2014). News of antibiotic resistance among Gram-negative bacilli in Algeria. Pathol. Biol. 62, 169–178. doi: 10.1016/j.patbio.2014.01.005
Baba Ahmed-Kazi Tani, Z., Decré, D., Genel, N., Boucherit-Otmani, Z., Arlet, G., and Drissi, M. (2013). Molecular and epidemiological characterization of enterobacterial multidrug resistant strains in Tlemcen Hospital (Algeria) (2008–2010). Microb. Drug Resist. 19, 185–190. doi: 10.1089/mdr.2012.0161
Bakour, S., Touati, A., Bachiri, T., Sahli, F., Tiouit, D., Naim, M., et al. (2014). First report of 16S rRNA methylase ArmA-producing Acinetobacter baumannii and rapid spread of metallo-β-lactamase NDM-1 in Algerian hospitals. J. Infect. Chemother. 20, 696–701. doi: 10.1016/j.jiac.2014.07.010
Barton, B. M., Harding, G. P., and Zuccarelli, A. J. (1995). A general method for detecting and sizing large plasmids. Anal. Biochem. 226, 235–240. doi: 10.1006/abio.1995.1220
Batah, R., Loucif, L., Olaitan, A. O., Boutefnouchet, N., Allag, H., and Rolain, J. M. (2015). Outbreak of Serratia marcescens coproducing ArmA and CTX-M-15 mediated high levels of resistance to aminoglycoside and extended-spectrum beta-lactamases, Algeria. Microb. Drug Resist. 21, 470–476. doi: 10.1089/mdr.2014.0240
Belbel, Z., Chettibi, H., Dekhil, M., Ladjama, A., Nedjai, S., and Rolain, J. M. (2014). Outbreak of an armA methyltransferase-producing ST39 Klebsiella pneumoniae clone in a pediatric Algerian Hospital. Microb. Drug Resist. 20, 310–315. doi: 10.1089/mdr.2013.0193
Bercot, B., Poirel, L., and Nordmann, P. (2008). Plasmid-mediated 16S rRNA methylases among extended-spectrum β-lactamase-producing Enterobacteriaceae isolates. Antimicrob. Agents Chemother. 52, 4526–4527. doi: 10.1128/AAC.00882-08
Bercot, B., Poirel, L., Ozdamar, M., Hakko, E., Turkoglu, S., and Nordmann, P. (2010). Low prevalence of 16S methylases among extended-spectrum- β-lactamase-producing Enterobacteriaceae from a Turkish hospital. J. Antimicrob. Chemother. 65, 797–798. doi: 10.1093/jac/dkq003
Bogaerts, P., Galimand, M., Bauraing, C., Deplano, A., Vanhoof, R., De Mendonca, R., et al. (2007). Emergence of ArmA and RmtB aminoglycoside resistance 16S rRNA methylases in Belgium. J. Antimicrob. Chemother. 59, 459–464. doi: 10.1093/jac/dkl527
Bouzidi, N., Aoun, L., Dekhil, M., Granier, S. A., Poirel, L., Brisabois, A., et al. (2011). Co-occurrence of aminoglycoside resistance gene armA in non-Typhi Salmonella isolates producing CTX-M-15 in Algeria. J. Antimicrob. Chemother. 66, 2180–2181. doi: 10.1093/jac/dkr237
Cantón, R., and Coque, T. M. (2006). The CTX-M β-lactamase pandemic. Curr. Opin. Microbiol. 9, 466–475. doi: 10.1016/j.mib.2006.08.011
Carattoli, A., Bertini, A., Villa, L., Falbo, V., Hopkins, K. L., and Threlfall, E. J. (2005). Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 63, 219–228. doi: 10.1016/j.mimet.2005.03.018
Chen, L., Chen, Z. L., Liu, J. H., Zeng, Z. L., Ma, J. Y., and Jiang, H. X. (2007). Emergence of RmtB methylase-producing Escherichia coli and Enterobacter cloacae isolates from pigs in China. J. Antimicrob. Chemother. 59, 880–885. doi: 10.1093/jac/dkm065
Ejrnaes, K., Sandvang, D., Lundgren, B., Ferry, S., Holm, S., Monsen, T., et al. (2006). Pulsed-field gel electrophoresis typing of Escherichia coli strains from samples collected before and after pivmecillinam or placebo treatment of uncomplicated community-acquired urinary tract infection in women. J. Clin. Microbiol. 44, 1776–1781. doi: 10.1128/JCM.44.5.1776-1781.2006
Fritsche, T. R., Castanheira, M., Miller, G. H., Jones, R. N., and Armstrong, E. S. (2008). Detection of methyltransferases conferring high-level resistance to aminoglycosides in Enterobacteriaceae from Europe, North America, and Latin America. Antimicrob. Agents Chemother. 52, 1843–1845. doi: 10.1128/AAC.01477-07
Galimand, M., Courvalin, P., and Lambert, T. (2003). Plasmid-mediated highlevel resistance to aminoglycosides in Enterobacteriaceae due to 16S rRNA methylation. Antimicrob. Agents Chemother. 47, 2565–2571. doi: 10.1128/AAC.47.8.2565-2571.2003
Girlich, D., Bouihat, N., Poirel, L., Benouda, A., and Nordmann, P. (2014). High rate of faecal carriage of extended-spectrum b-lactamase and OXA-48 carbapenemase-producing Enterobacteriaceae at a University hospital in Morocco. Clin. Microbiol. Infect. 20, 350–354. doi: 10.1111/1469-0691.12325
Iabadene, H., Bakour, R., Messai, Y., Da Costa, A., and Arlet, G. (2009). Detection of blaCTX-M-14 and aac(3)-II genes in Salmonella enterica serotype Kedougou in Algeria. Med. Mal. Infect. 39, 806–807. doi: 10.1016/j.medmal.2009.04.003
Jarlier, V., Nicolas, M. H., Fournier, G., and Philippon, A. (1988). Extended broad-spectrum b-lactamases conferring transferable resistance to newer b-lactam agents in Enterobacteriaceae: hospital prevalence and susceptibility patterns. Rev. Infect. Dis. 10, 867–878. doi: 10.1093/clinids/10.4.867
Khennouchi, N. C., Loucif, L., Boutefnouchet, N., Alleg, H., and Rolain, J. M. (2015). MALDI-TOF MS as a tool to detect a nosocomial outbreak of extended-spectrum beta-lactamase- and ArmA methyltransferase-producing Enterobacter cloacae clinical isolates in Algeria. Antimicrob. Agents Chemother. 59, 6477–6483. doi: 10.1128/AAC.00615-15
Mamlouk, K., Boutiba-Ben Bakour, I., Gautie, V., Vimont, S., Picard, B., Redjeb, S. B., et al. (2006). Emergence and outbreaks of CTX-M extended spectrum β-lactamase producing Echerichia coli and Klebsiella pneumoniae strains in a Tunisian hospital. J. Clin. Microbiol. 44, 4049–4056. doi: 10.1128/JCM.01076-06
Naas, T., Bentchouala, C., Cuzon, G., Yaou, S., Lezzar, A., Smati, F., et al. (2011). Outbreak of Salmonella enterica serotype Infantis producing ArmA 16S RNA methylase and CTX-M-15 extended-spectrum β-lactamase in a neonatology ward in Constantine, Algeria. Int. J. Antimicrob. Agents 38, 135–139. doi: 10.1016/j.ijantimicag.2011.04.012
Naas, T., Bentchouala, C., Lima, S., Lezzar, A., Smati, F., Scheftel, J. M., et al. (2009). Plasmid-mediated 16S rRNA methylases among extended-spectrum-beta-lactamase-producing Salmonella enterica Senftenberg isolates from Algeria. J. Antimicrob. Chemother. 64, 866–868. doi: 10.1093/jac/dkp312
Naas, T., Lezzar, A., Bentchouala, C., Smati, F., Scheftel, J. M., Monteil, H., et al. (2005). Multidrug-resistant Salmonella enterica serotype Senftenberg isolates producing CTX-M beta-lactamases from Constantine, Algeria. J. Antimicrob. Chemother. 56, 439–440. doi: 10.1093/jac/dki216
Nagasawa, M., Kaku, M., Kamachi, K., Shibayama, K., Arakawa, Y., Yamaguchi, K., et al. (2014). Loop-mediated isothermal amplification assay for 16S rRNA méthylases genes in Gram-negative bacteria. J. Infect. Chemother. 20, 635–638. doi: 10.1016/j.jiac.2014.08.013
Neuwirth, C., Siebor, E., Lopez, J., Pechinot, A., and Kazmierczak, A. (1996). Outbreak of TEM-24-Producing Enterobacter aerogenes in an intensive care unit and dissemination of the extended-spectrum β-lactamase to other members of the family Enterobacteriaceae. J. Clin. Microbiol. 34, 76–79.
Pitout, J. D. D., and Laupland, K. B. (2008). Extended-spectrum-beta-lactamases producing Enterobacteriaceae: an emerging public-health concern. Lancet Infect. Dis. 8, 159–166. doi: 10.1016/S1473-3099(08)70041-0
Ramdani-Bouguessa, N., Mendonça, N., Leitao, J., Ferreira, E., Tazir, M., and Caniça, M. (2006). CTX-M-3 and CTXM- 15 extended-spectrum b-lactamases in isolates of Escherichia coli from a hospital in Algiers, Algeria. J. Clin. Microbiol. 44, 4584–4586. doi: 10.1128/JCM.01445-06
Siebor, E., and Neuwirth, C. (2011). The new variant of Salmonella genomic island 1 (SGI1-V) from a Proteus mirabilis French clinical isolate harbours blaVEB-6 and qnrA1 in the multiple antibiotic resistance region. J. Antimicrob. Chemother. 66, 2513–2520. doi: 10.1093/jac/dkr335
Tenover, F. C., Arbeit, R. D., Goering, R. V., Mickelsen, P. A., Murray, B. E., Persing, D. H., et al. (1995). Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol. 33, 2233–2239.
Tian, G. B., Rivera, J. I., Park, Y. S., Johnson, L. E., Hingwe, A., Adams-Haduch, J. M., et al. (2011). Sequence type ST405 Escherichia coli isolate producing QepA1, CTX-M-15, and RmtB from Detroit, Michigan. Antimicrob. Agents Chemother. 55, 3966–3967. doi: 10.1128/AAC.00652-11
Wirth, T., Falush, D., Lan, R., Colles, F., Mensa, P., Wieler, L. H., et al. (2006). Sex and virulence in Escherichia coli: an evolutionary perspective. Mol. Microbiol. 60, 1136–1151. doi: 10.1111/j.1365-2958.2006.05172.x
Woodford, N., Turton, J. F., and Livermore, D. M. (2011). Multiresistant Gram-negative bacteria: the role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbial. Rev. 35, 736–755. doi: 10.1111/j.1574-6976
Wu, Q., Zhang, Y., Han, L., Sun, J., and Ni, Y. (2009). Plasmid-mediated 16S rRNA méthylases in aminoglycoside-resistant Enterobacteriaceae isolates in Shanghai, China. Antimicrob. Agents Chemother. 53, 271–272. doi: 10.1128/AAC.00748-08
Keywords: Escherichia coli, extended-spectrum β-lactamase, 16S rRNA methylase, armA, rmtB, Algeria
Citation: Ayad A, Drissi M, de Curraize C, Dupont C, Hartmann A, Solanas S, Siebor E, Amoureux L and Neuwirth C (2016) Occurence of ArmA and RmtB Aminoglycoside Resistance 16S rRNA Methylases in Extended-Spectrum β-Lactamases Producing Escherichia coli in Algerian Hospitals. Front. Microbiol. 7:1409. doi: 10.3389/fmicb.2016.01409
Received: 25 February 2016; Accepted: 25 August 2016;
Published: 12 September 2016.
Edited by:
Manuela Caniça, National Institute of Health Dr. Ricardo Jorge, PortugalReviewed by:
Jian-Hua Liu, South China Agricultural University, ChinaLaurent Dortet, Bicêtre Hospital – South Paris University, France
Copyright © 2016 Ayad, Drissi, de Curraize, Dupont, Hartmann, Solanas, Siebor, Amoureux and Neuwirth. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Catherine Neuwirth, Y2F0aGVyaW5lLm5ldXdpcnRoQGNodS1kaWpvbi5mcg==