 Caroline M. Mota1
Caroline M. Mota1 Ana C. M. Oliveira1
Ana C. M. Oliveira1 Marcela Davoli-Ferreira2
Marcela Davoli-Ferreira2 Murilo V. Silva1Fernanda M. Santiago1Santhosh M. Nadipuram3
Murilo V. Silva1Fernanda M. Santiago1Santhosh M. Nadipuram3 Ajay A. Vashisht4James A. Wohlschlegel4
Ajay A. Vashisht4James A. Wohlschlegel4 Peter J. Bradley3,5
Peter J. Bradley3,5 João S. Silva2
João S. Silva2 José R. Mineo1
José R. Mineo1 Tiago W. P. Mineo1*
Tiago W. P. Mineo1*- 1Laboratory of Immunoparasitology “Dr. Mário Endsfeldz Camargo,” Department of Immunology, Institute of Biomedical Sciences, Federal University of Uberlândia, Uberlândia, Brazil
- 2Department of Biochemistry and Immunology, School of Medicine of Ribeirão Preto, University of São Paulo, Ribeirão Preto, Brazil
- 3Department of Microbiology, Immunology and Molecular Genetics, University of California, Los Angeles, Los Angeles, CA, USA
- 4Department of Biological Chemistry and Institute of Genomics and Proteomics, University of California, Los Angeles, Los Angeles, CA, USA
- 5Molecular Biology Institute, University of California, Los Angeles, Los Angeles, CA, USA
Due to the high prevalence and economic impact of neosporosis, the development of safe and effective vaccines and therapies against this parasite has been a priority in the field and is crucial to limit horizontal and vertical transmission in natural hosts. Limited data is available regarding factors that regulate the immune response against this parasite and such knowledge is essential in order to understand Neospora caninum induced pathogenesis. Mitogen-activated protein kinases (MAPKs) govern diverse cellular processes, including growth, differentiation, apoptosis, and immune-mediated responses. In that sense, our goal was to understand the role of MAPKs during the infection by N. caninum. We found that p38 phosphorylation was quickly triggered in macrophages stimulated by live tachyzoites and antigen extracts, while its chemical inhibition resulted in upregulation of IL-12p40 production and augmented B7/MHC expression. In vivo blockade of p38 resulted in an amplified production of cytokines, which preceded a reduction in latent parasite burden and enhanced survival against the infection. Additionally, the experiments indicate that the p38 activation is induced by a mechanism that depends on GPCR, PI3K and AKT signaling pathways, and that the phenomena here observed is distinct that those induced by Toxoplasma gondii’s GRA24 protein. Altogether, these results showed that N. caninum manipulates p38 phosphorylation in its favor, in order to downregulate the host’s innate immune responses. Additionally, those results infer that active interference in this signaling pathway may be useful for the development of a new therapeutic strategy against neosporosis.
Introduction
Neospora caninum is an obligate intracellular protozoan from the phylum Apicomplexa, closely related to Toxoplasma gondii. This parasite has a worldwide distribution and causes relevant economic impact in dairy and beef industries (Cardoso et al., 2011) due to negative effects such as abortion or reproductive disorders in cattle, in addition to neuromuscular disease in dogs (Goodswen et al., 2012). Cattle are an important intermediate host species that may acquire the infection through horizontal transmission after ingestion of oocysts excreted by canine, which are the parasite’s definitive hosts. Infection may also occur during gestation, due to the immune response regulation, which leads to the recrudescence of chronic infections into subsequent parasitemia (Mineo et al., 2010a; Pinheiro et al., 2010). Afterward, the fetus may be infected by parasites that cross the placenta, causing abortions or congenital infections, depending on the period of gestation and parasite virulence (Eastick and Elsheikha, 2010).
The immune response against N. caninum is predominantly Th1-based, with the sequential production of interleukin-12 (IL-12), interferon gamma (IFN-γ) and nitric oxide (NO) by cells of the immune system, with a notable highlight to macrophages (MO), which underline the importance of cellular effector responses in the reduction of tissue parasitism and host survival, along with IgG2 immunoglobulin production (Abe et al., 2015; Donahoe et al., 2015; Hecker et al., 2015). IL-12 is a key cytokine that links the innate and adaptive compartments of the immune system, and is triggered by microbial products during early host–pathogen interactions (Aliberti, 2005; Yarovinsky et al., 2005; Debierre-Grockiego et al., 2007; Jenkins et al., 2010; Donahoe et al., 2015).
The usual production of this cytokine is initiated through recognition of highly conserved sets of molecular patterns (pathogen-associated molecular patterns, PAMPs) through a limited number of germline-encoded receptors in innate cells called pattern-recognition receptors (PRRs). The contact of these PAMPs with PRRs on macrophages triggers intracellular signaling pathways that result in the induction of an appropriated inflammatory response (Kawai and Akira, 2010). However, the host-specific intracellular signaling pathways triggered by innate recognition of N. caninum have not yet been fully elucidated. Previous studies demonstrated that initial N. caninum recognition includes TLR2, TLR3, and TLR11 (Jenkins et al., 2010; Mineo et al., 2010b; Beiting et al., 2014). Engagement of these receptors triggers activation of MyD88 or TRIF-dependent pathways, respectively, enhancing the immune response against this parasite (Mineo et al., 2009; Beiting et al., 2014). CCR5 is also a key player in the immune response against N. caninum through the production of cyclophilin, a parasite protein that modulates migration and activation of innate cells during the early phase of the infection (Mineo et al., 2010a; Abe et al., 2015).
The sustained synthesis of IL-12 in tissues and cells hosting Apicomplexan parasites is not controlled only by NF-κB, and also requires activation of MAPK pathways (Kim et al., 2005; Masek et al., 2006). MAPK pathways are composed by several kinases that activate an orchestrated cascade (MAP4K, the MAP3K, the MAP2K), which culminates in the phosphorylation of specific proteins of MAPK pathways that regulate the expression of sets of genes and effector proteins. The three most studied MAPK pathways are c-Jun-activated kinases (JNKs), extracellular signal-related kinases (ERKs) and p38 MAPK. MAPKs are activated by dual phosphorylation of the threonine and tyrosine residues, mediated by upstream MAPK kinases (MKK) (Kim et al., 2005; Yang et al., 2013). Usually, the ERK pathway induces growth factor signals, whereas the JNK and p38 pathways may be activated by a variety of extracellular stress signals (Cuadrado and Nebreda, 2010; Yang et al., 2013). Studies have been proposed that T. gondii induces phosphorylation of p38α MAPK in order to promote IL-12p70 production (Cuadrado and Nebreda, 2010). T. gondii secretes GRA24, a dense granule protein that has no ortholog expressed in N. caninum, that has the unique ability to trigger autophosphorylation and nuclear translocation of host p38, which correlates with the up-regulation of the transcription factors Egr-1 and c-Fos, and consequent synthesis of key proinflammatory cytokines, including IL-12 (Braun et al., 2013). Additionally, previous studies have reported that T. gondii also exploits heterotrimeric Gi-protein-mediated signaling to activate phosphoinositide 3-kinases (PI3Ks), leading to phosphorylation of PI3K and MAPK pathways, that results in inhibition of apoptosis and regulation of cytokine production (Kim and Denkers, 2006; Quan et al., 2013).
Within this context, the present study aimed to characterize the effect of MAPK signaling in the activation of immune responses against N. caninum and its antigens. Our results provide evidence that the parasite evades effective host responses through the activation of p38 MAPK-dependent G protein-coupled receptor (GPCR)/PI3K/AKT pathway.
Materials and Methods
Ethics Statement
All animal studies were approved by the animal research committee at UFU (Comitê de Ética na Utilização de Animais da Universidade Federal de Uberlândia – CEUA/UFU), under protocol number 029/12. All procedures including housing and welfare were carried out in accordance with the recommendations in the Guiding Principles for Biomedical Research Involving Animals of the International Council for Laboratory Animal Science (ICLAS), countersigned by the Conselho Nacional de Controle de Experimentação Animal (CONCEA; Brazilian National Consul for the Control of Animal Experimentation) in its E-book1. The UFU animal facility (Centro de Bioterismo e Experimentação Animal – CBEA/UFU) is accredited by the CONCEA (CIAEP: 01.0105.2014) and Comissão Técnica Nacional de Biossegurança (CTNBio, Brazilian National Commission on Biossecurity; CQB: 163/02).
Parasite and Antigens
Wild type (Nc-1) and genetically modified (NcLivΔHPT and NcLiv_GRA24-BirA∗) strains of N. caninum and T. gondii tachyzoites (PRU isolate) were maintained by serial passages in HeLa cell line (CCL-2, ATCC, Manassas, VA, USA), cultured in RPMI 1640 medium supplemented with 2 mM glutamine, 100 U/mL penicillin and 100 μg/mL streptomycin, at 37°C in 5% CO2 atmosphere. Parasite suspensions were obtained as previously described (Ribeiro et al., 2009). Briefly, tachyzoites were harvested by scraping off the cell monolayer after 2–3 days of infection, passed through a 26-gauge needle and centrifuged at low speed (45 × g) for 1 min at 4°C, for debris removal. The supernatant containing parasite suspension was collected, washed in phosphate buffered saline (PBS, pH 7.2), and the pellet was resuspended in PBS for antigen preparation.
N. caninum lysate antigen (NLA) was prepared as previously described (Ribeiro et al., 2009). Briefly, parasite suspension was treated with protease inhibitors (Roche Diagnostics GmbH, Mannheim, Germany) and then lysed by freeze-thaw cycles, followed by ultrasound cycles (60 H/30 s) on ice. After centrifugation (10,000 × g, 30 min, 4°C), the supernatant was collected, filtered on 0.22 μm membranes and its protein concentration determined by Bradford assay (Sigma–Aldrich, St. Louis, MO, USA). NLA aliquots were stored at -20°C until used in vitro assays.
Preparation of N. caninum excreted-secreted antigen (NcESA) was carried out as described elsewhere (Ribeiro et al., 2009). Freshly egressed tachyzoites (108 parasites/ml) were washed twice in PBS, resuspended in Hanks saline solution and incubated for 30 min at 37°C, at mild agitation. Parasites were then centrifuged (720 × g, 10 min, 4°C), the supernatant was collected and again centrifuged (10,000 × g, 4°C, 30 min). The final supernatant was filtered on 0.22 μm membranes and its protein concentration determined by Bradford (1976). Different batches of NcESA were prepared and pooled together to obtain the required protein concentration. NcESA aliquots were stored at -20°C until use.
To exclude that experiments conduced with different strains and p38 inhibitors would generate distinct results, we have tested both parental N. caninum strains used (Nc-1 and NcLivΔHPT), well as both p38 specific chemical inhibitors (SB203580 – for in vitro use only; and SB239063 – preconized for in vivo use) and observed that there are no notable phenotype differences (Supplementary Figure 1).
Differentiation of Bone Marrow-Derived Macrophages (BMDMs)
Bone-marrow stem cells were obtained from femurs and tibiae of 6- to 8-week-old wild-type and PI3K, MyD88 and CCR5 deficient C57BL/6 mice, according to previous description (Marim et al., 2010). Briefly, mice were euthanized and the bones were collected and flushed to extrude bone marrow. The cell suspension was used to generate bone marrow derived macrophages (BMDMs) using L929-cell conditioned medium (LCCM) as a source of granulocyte/macrophage colony stimulating factor. Cells were seeded in non-tissue culture treated petri dishes (Optilux, BD Biosciences) and incubated at 37°C in a 5% CO2 atmosphere. Four days after seeding the cells, an extra of fresh R20/30 (RPMI1640 supplemented with 20% FBS, 30% LCCM, 100 U/ml penicillin, 100 μg/mL streptomycin, and 2 mM L-glutamine) were added per plate. After 7 days, adherent cells were removed and analyzed by flow cytometry. Once the positive phenotype was confirmed (CD11b expression over 95%), BMDMs were plated at a concentration of 1.106 cells/ml of RPMI, supplemented with 10% FBS, before some further experimental procedure.
Measurement of MAPK Activation
Bone marrow derived macrophages were added to 12-well plate at a density of 1 × 106 cells per ml and infected with live tachyzoites (Nc-1 or NcLivΔHPT; 1:1 ratio of parasites to cells). The cells were lysed in denaturation buffer 5x containing protease and phosphatase inhibitors (Roche), 0, 15, 30, 60, and 120 min post-infection.
Preparation of samples was performed according to the manufacturer’s protocol for adherent cells (BD Biosciences; Schubert et al., 2009). Briefly, cells were lysed and collected from the plates. Samples were then placed immediately in a boiling water bath for 5 min. Cell lysates were centrifuged at 15.000 × g for 5 min and supernatants were stored at -80°C until measurement. pJNK1/2 (T183/Y185), pp38 (T180/Y182), and pERK1/2 (T202/Y204) were quantitatively determined using antibodies from the multiplex Flex Set Cytometric Bead Array (pCBA, BD Biosciences). Serial dilutions (1/2 v/v) of the standards were prepared; cell lysates were (1/2 v/v) diluted using assay diluent and transferred to a FACS tube. Then, 25 μl of mixed capture beads were transferred to each tube. After a 3 h incubation at room temperature, 25 μL PE detection reagent was added and samples were incubated for another hour at room temperature. FACS tube was washed with buffer and after final centrifugation, 200 × g for 10 min, 150 μL wash buffer was added. Flow cytometric analysis was performed using a flow cytometer (FACSCantoII, BD Biosciences) and dedicated software (FCAP array v3.0, BD Biosciences). A total of 900 events were acquired following the protocol supplied. The minimum detection levels for each phospho-protein were: pJNK = 0.38 U/ml; pp38 = 0.64 U/ml; pERK = 0.64 U/ml.
Western Blotting
For Western blotting, BMDMs were added to 12-well plate at a density of 1 × 106 cells/ml and infected with N. caninum tachyzoites or stimulated with LPS or PMA. After 30 min, the cells were collected in lysis buffer containing a mixture of protease and phosphatase inhibitors (Roche). The lysates were resolved by SDS-PAGE and transferred onto nitrocellulose membranes. Blots were probed with the primary antibodies against p-p38, total p38, p-pERK1/2, total pERK1/2,p-JNK and total JNK (R&D Systems), followed by secondary antibodies conjugated to horseradish peroxidase (HRP). Target proteins were visualized by chemiluminescence for the detection of HRP activity (ChemiDoc XRS, Bio-Rad, Hercules, CA, USA). Additionally, whole-parasite lysates from parental and modified strains were separated by 12% SDS-PAGE. Samples were transferred to nitrocellulose overnight and probed with primary antibodies. For all secondary antibody incubations, HRP-conjugated goat anti-mouse or goat anti-rabbit antibodies were used at a 1:2000 dilution. Following secondary incubation, a chemiluminescent substrate was used for the detection of HRP activity.
Phenotyping of Costimulatory Molecules (B7) and Major Histocompability Complex (MHC)
Bone marrow derived macrophages were added to 12-well plates at a density of 1 × 106 cells/mL, pretreated for 3 h with p38 inhibitor (SB203580, 10 μM) or medium before antigenic stimulus (NLA, 10 μg/mL). After, 24 h of stimulated cells were labeled with antibodies conjugated to fluorochromes, following the protocol previously described (Mineo et al., 2010b). For these tests, we used the following commercial antibodies: CD11b-APC.Cy7 (clone M1/70), MHC II-APC (clone AF-120.1), MHC I-PE.Cy7 (clone AF-88.5), CD80-PE (clone 16-10A1), and CD86-FITC (clone GL1) (BD Biosciences). Phenotyping of BMDMs was performed in a flow cytometer (FACSCanto II, BD Biosciences) and the data was processed and analyzed using dedicated software (FlowJo, TreeStar, USA).
Antigenic Recall Using Primed Spleen Cells
For measurement of ex vivo cytokine production, spleens retrieved from mice infected by N. caninum (5 × 106 Nc-1 tachyzoites/mouse) or infected and treated with p38 inhibitor SB239063 (0.5 mg/Kg), along with its appropriated controls, were dissociated 30 days post infection (dpi) in RPMI medium and cell suspensions were washed in medium and added a lysis buffer (0.16 M NH4Cl and 0.17M Tris–HCl, pH 7.5), cells were washed again and resuspended in complete RPMI medium containing 10% FBS. Viable cells (2 × 105 cells/200 μl/well) were cultured in triplicate in 96-well plates in the presence of antigen (NLA, 10 μg/ml), mitogen (Concanavalin A – ConA, 2.5 μg/ml), Nc-1 tachyzoites (1:10 parasite per cells) or medium alone and incubated at 37°C in 5% CO2. After 48 h, cell-free supernatants were collected and stored at -80°C for the measurement of cytokines.
Challenge
In vivo experiments were conducted with two different parasite doses, depending on the aim of each experiment: (i) WT mice (at least six mice/group) were challenged intraperitoneally with lethal doses (1 × 107 Nc-1 tachyzoites/mouse), treated or not for 5 days with the p38 inhibitor SB239063 (0.5 mg/Kg), in order to observe the effect of the p38 inhibitor treatment in the survival of the challenged mice. Animals were observed daily for morbidity and mortality during 30 dpi. (ii) In other sets of experiments, WT mice (six mice/group) were infected intraperitoneally with sub-lethal doses (5 × 106 Nc-1 tachyzoites/mouse), treated or not for 5 days with SB239063 (0.5 mg/Kg), for the evaluation of immunological parameters and chronic phase parasite burden. After 7 days of infection, blood samples were collected and analyzed for the concentration of cytokines. After 30 days of infection, brain tissues and spleen cells were used for the quantification of chronic phase parasite burden and evaluation of cytokine production upon antigenic recall, respectively.
Cytokine Quantification
Bone marrow derived macrophages were added to 96-well plate at a density of 1 × 106 cells/mL. Stimulation was performed with live N. caninum tachyzoites (1:1 ratio of parasites to cells), NLA (10 μg/mL) or ESA (10 μg/mL). In some experiments, BMDMs were pretreated for 3 h with p38 inhibitor (SB203580, 10 μM), JNK inhibitor (SP600125, 10 μM), ERK1/2 inhibitor (PD98059, 10 μM), GPCRs inhibitor (Pertussis toxin, 200 ng/ml), PI3K inhibitor (Wortmannin, 500 nM), AKT inhibitor (AKT quinase VIII, 10 μM) or medium before infection. The concentration of each drug was chosen based on previously published manuscripts (Kim et al., 2005; Ruhland and Kima, 2009; Zong et al., 2012). p38 inhibition by SB203580 was verified in BMDMs by pCBA (Supplementary Figure 2). After, 24 h of infection or antigenic stimulus, the culture supernatant were collected for the measurement of IL-12p40 cytokine concentration. Quantification of IL-12p40 and IFN-γ in BMDMs and splenocytes was performed by ELISA, according to the manufacturer instructions (BD Biosciences). The assays were read at 450 nm and the OD values obtained were converted to pg/mL by the extrapolation of the standard curve (M2e plate reader, Molecular Devices, Sunnyvale, CA, USA). Serum of WT mice infected with Nc-1 strain (5 × 106 tachyzoites/mouse), submitted or not to SB239063 treatment (0.5 mg/Kg), was collected after 7 dpi. The effect on the cytokine profile during SB239063 treatment was evaluated using a mouse Th1/Th2/Th17 cytometric bead array kit (CBA), according to the manufacturers’ instructions (BD Biosciences). The assays were read in a flow cytometer (FACSCanto II, BD Biosciences), and the concentration of each analyte was extrapolated from standard curves of recombinant cytokines. The minimum detection levels for each cytokine were: IL-2 = 0.1 pg/mL; IL-4 = 0.03 pg/mL; IL-6 = 1.4 pg/mL; IFN-γ = 0.5 pg/mL; TNF = 0.9 pg/mL; IL-17A = 0.8 pg/mL; IL-10 = 16.8 pg/mL.
Determination of Parasite Burden
Brain parasite burden was determined by quantitative real-time PCR as previously described (Ribeiro et al., 2009), by the use of primer pairs (sense 3′-GCTGAACACCGTATGTCGTAAA-5′; antisense 3′-AGAGGAATGCCACATAGAAGC-5′) to detect the N. caninum Nc-5 sequence. DNA extraction was performed from 20 mg of murine brain tissues (Genomic DNA kit, Promega Co., USA) and parasite loads were calculated by interpolation from a standard curve of Nc-1 tachyzoite DNA included in each run (StepOne Plus, Applied Biosystems, USA). As a negative control, DNA obtained from brain tissues of non-immunized and unchallenged mice was analyzed in parallel.
Generation of GRA24 Knock-in Parasites
Targeting constructs were generated using the pNeoGra7-3xHA-BirA∗-HPT vector. Briefly, Type II GRA24 gene locus is defined by TGME49_230180 in the T. gondii genome database2 (version 6.0). The gene was cloned by PCR with engineered Nsi I 5′ sites and Not1 3′ sites. After amplification in a pJET vector, the GRA24 insert was fused in frame with a plasmid using the same restriction sites (Figure 5A) (manuscript in preparation). The construct was linearized by Hind III restriction digest and 75 μg of the DNA was transfected by electroporation into NcLivΔHPT parasites. Stable transformants were selected using medium containing 25 μg/mL of mycophenolic acid plus 25 μg/mL of xanthine (for HXGPRT plasmids) and cloned by limited dilution in 96 well plates. To confirm proper Gra24 gene targeting and expression, transgenic parasites were analyzed by IFA and the HA-tagged construct was shown to be co-localized in the parasitophorous vacuole with GRA2 (Supplementary Figure 3).
Immunofluorescence Assay
For IFA, HFFs were grown to confluence on coverslips with medium containing or not 150 μM of biotin and infected with N. caninum parasites expressing HA-tag and BirA∗ fusions. After 36 h, the coverslips were fixed and processed for indirect immunofluorescence as previously described (Chen et al., 2015). The coverslips were mounted in Vectashield (Vector Labs) and viewed with an Axio Imager.Z1 fluorescence microscope (Zeiss).
Affinity Capture of Biotinylated Proteins
HFF monolayers infected with parasites expressing BirA∗ fusions, or its respective parental line, were grown in medium containing 150 μM biotin for 24 h prior to parasite egress (Chen et al., 2015). Cells infected were collected, washed in PBS, and lysed in RIPA buffer (50 mM Tris [pH 7.5], 150 mM NaCl, 0.1% SDS, 0.5% sodium deoxycholate, 1% NP-40) supplemented with protease inhibitor cocktail (Roche) for 30 min on ice. Lysates were centrifuged for 15 min at 14,000 × g to pellet insoluble debris and then incubated with streptavidin magnetic beads (Pierce, USA), at room temperature for 4 h, under gentle agitation. Beads were collected using magnets and washed five times in RIPA buffer, followed by three washes in 8 M urea buffer (50 mM Tris-HCl [pH 7.4], 150 mM NaCl). Ten percent of each sample was boiled in Laemmli sample buffer, and eluted proteins were analyzed by Western blotting with streptavidin-HRP prior to mass spectrometry analysis.
Mass Spectrometry
By sequential addition of Lys-C and trypsin proteases the purified biotinylated proteins bound to streptavidin beads were reduced, alkylated, and digested (Chen et al., 2015). The peptide mixture was desalted by the use of C18 tips and fractionated by a 75 μm inner diameter filter fused to silica capillary column with a 5 μm pulled electrospray tip, and packed in house with 15 cm of Luna C18 column with 3 μm reversed-phase particles. Delivering of the gradient was performed by an easy-nLC 1000 ultrahigh-pressure liquid chromatography (UHPLC) system (Thermo Scientific). Tandem mass spectrometry (MS/MS) spectra were collected on a Q-Exactive mass spectrometer (Thermo Scientific). Data analysis was done using the ProLuCID and DTASelect2 implemented in the Integrated Proteomics pipeline IP2 (Integrated Proteomics Applications, Inc., San Diego, CA, USA). Protein and peptide identifications were filtered using DTASelect and required a minimum of two unique peptides per protein and a peptide-level false-positive rate of less than 5%, as estimated by a decoy database strategy. Normalized spectral abundance factor (NSAF) values were calculated as previously described (Chen et al., 2015).
Statistical Analysis
Differences amongst the groups were analyzed using ANOVA followed by Bonferroni multiple comparison post-tests to examine all possible pairwise comparisons, with few exceptions: (i) Comparisons involving parasite burden quantification by qPCR were analyzed by Kruskal–Wallis test followed by Dunn multiple comparison post-test; (ii) Student t-test was used for comparison between IgG1 and IgG2abc isotypes between different individual groups; (iii) The Kaplan–Meier method was applied to estimate the percentage of mice surviving at each time point after challenge and survival curves were compared using the Log-rank test. Analysis was carried out using GraphPad Prism 6.0 (GraphPad Software Inc., La Jolla, CA, USA). Values of P < 0.05 were considered statistically significant. Each experiment was independently conducted at least two times, and each condition was analyzed in triplicates, at least.
Results
N. caninum Induced Rapid Phosphorylation of p38 MAPK
To determine the effect of N. caninum infection on MAPK activation in macrophages, the phosphorylation of major MAPK signaling components was examined by CBA and Western Blotting (Figure 1). CBA experiments revealed that N. caninum induced strong phosphorylation of p38 MAPK (p-p38) between 15 and 30 min after contact with live parasites, with a tendency to reduce the reactivity after 60 min of infection. ERK1/2 and JNK did not show similar activation upon exposure to live parasites (Figure 1A). The same profile was observed by western blotting of BMDMs after 30 min of infection (Figure 1B).

FIGURE 1. Neospora caninum induces phosphorylation of p38 MAPK in macrophages. BMDMs (1 × 106 cells/ml) were infected with live Nc-1 isolate tachyzoites of N. caninum (Nc, 1:1 parasite to cell ratio) or stimulated with NLA (10 μg/mL) or ESA (10 μg/mL) and incubated under controlled temperature and atmosphere. (A) Representative dot plots of the kinetics of MAPKs phosphorylation after exposure of BMDMs to Nc (0, 15, 30, 60, and 120 min post infection) by CBA; (B) Expression of the activated MAPKs after 30 min of infection by Western Blotting. Uninfected cells were used as negative controls and, as a positive control, BMDMs stimulated with 1 μg/mL of Lipopolysaccharides (LPS) or 100 nM of Phorbol-12-myristate-13-acetate (PMA); (C) Representative dot plots of the phosphorylation of the different MAPKs tested after stimuli with live N. caninum tachyzoites or its soluble lysate (NLA) and excreted/secreted (ESA) antigens by flow cytometry. Results are representative of three independent experiments, with at least three technical replicates in each, which were extrapolated in relation the standard curve and are expressed as units/mL.
We next evaluated MAPK activation in macrophages exposed to different antigenic stimuli. For that purpose, BMDMs were exposed to N. caninum tachyzoites, soluble lysate antigen (NLA) or excreted-secreted antigens (ESA) and analyzed after 30 min (Figure 1C). It was observed that all antigens induced p-p38, although the highest levels were observed after infection with live N. caninum tachyzoites.
In reference to those findings, we suggest that N. caninum activates predominantly p38 MAPK after infection of BMDMs, probably through a component of its excreted/secreted antigens.
Parasite-Triggered p38 MAPK Activation Dampens Antigen Presentation Features of Macrophages
We first verified the role of p38, JNK and ERK in IL-12p40 production in N. caninum-infected BMDMs. Inhibition of p38 MAPK by SB203580 induced IL-12p40 upregulation after 24 h of infection with live parasites or its components (Figure 2A), while chemical inhibition of ERK 1/2 and JNK did not yield notable differences in relation to controls.
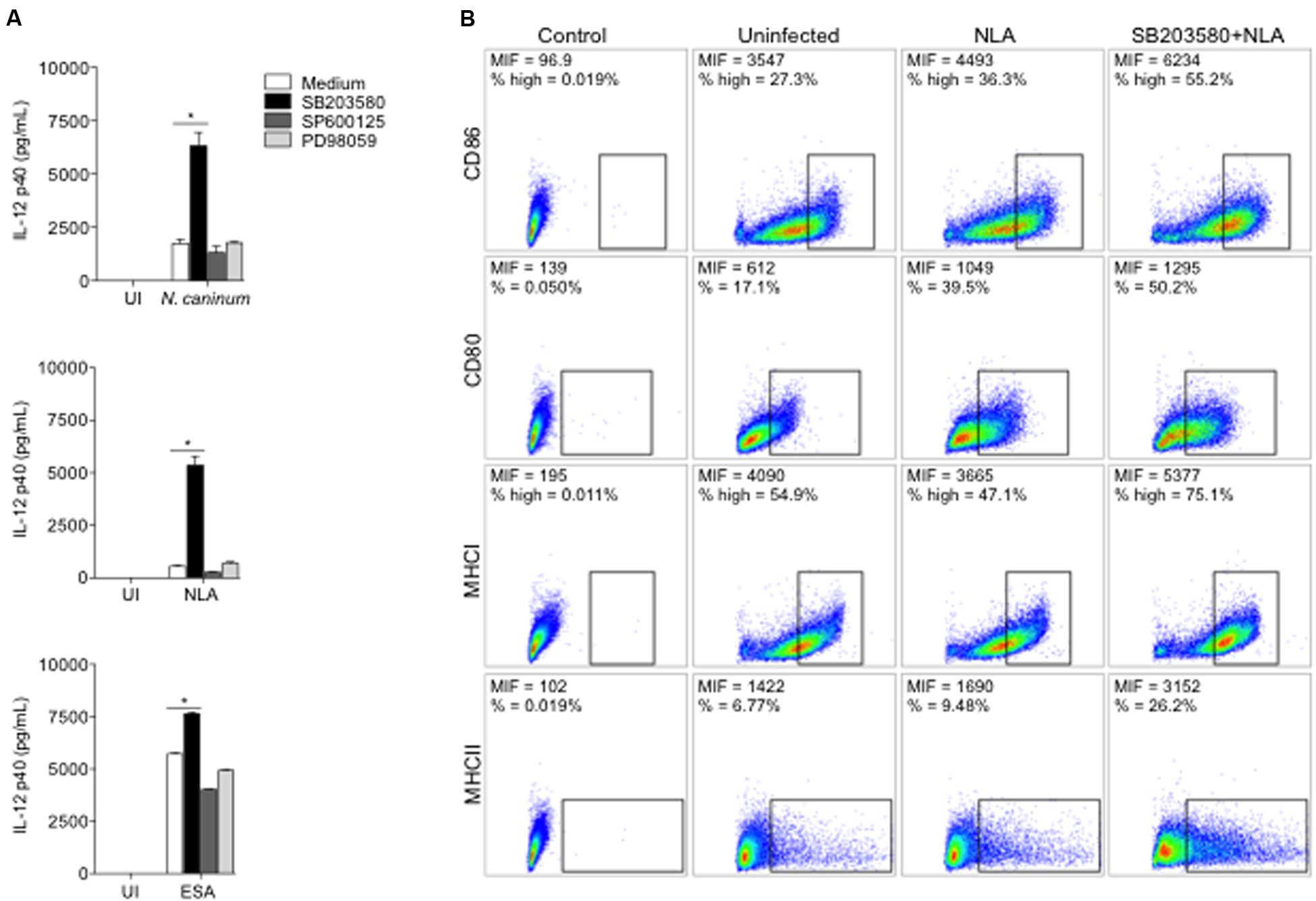
FIGURE 2. Neospora-induced p38 activation downregulates the production of IL-12 and the expression of antigen presentation molecules. BMDMs (1 × 106 cells/mL) were pretreated for 3 h with p38 inhibitor (SB203580, 10 μM), SAPK/JNK inhibitor (SP600125, 10 μM), ERK1/2 inhibitor (PD98059, 10 μM). (A) Cells were infected with N. caninum Nc-1 strain (Nc; 1:1 parasite to cell ratio) or stimulated with NLA (10 μg/mL) or ESA (10 μg/mL) or medium alone (UI). After 24 h, supernatants were collected and analyzed for IL-12p40 concentration. Results were expressed as mean ± SEM. ∗Indicates statistically significant differences between untreated cells and SB203580-treated cells, upon antigenic stimulation; P < 0.05, assessed by ANOVA followed by Bonferroni multiple comparison post-test to examine all possible pairwise comparisons. Results are representative of at least five independent experiments, with five technical replicates each. (B) BMDMs were stimulated with NLA (10 μg/mL) for 24 h and then checked for population positivity (%) and mean intensity of fluorescence (MIF) of surface markers CD80, CD86, MHCI, and MHCII. Results are representative of two independent experiments, with three technical replicates each, and dot plots were represented as percentage of positives cells, where warmer colors indicate higher cell densities.
To characterize the phenotype of N. caninum-infected BMDMs, the expression of B7 co-stimulatory (CD80 and CD86) and Major Histocompability Complex (MHCI and MHCII) molecules were observed, upon MAPK chemical inhibition. Macrophages pretreated with SB203580 and exposed to NLA showed higher expression of B7 and MHC molecules, percentage wise or by mean intensity of fluorescence (MIF), if compared to uninfected or infected untreated cells (Figure 2B). Similar results were observed in experiments with live tachyzoites (Supplementary Figure 4).
Based on these experiments, we have observed that p38 MAPK activation in infected macrophages is associated with pronounced decrease in IL-12p40 production, alongside with the expression of MHC and B7 co-stimulatory molecules.
p38 Inhibition Is Effective in Therapeutic and Immune-Prophylactic Strategies against N. caninum through the Upregulation of Cellular Responses
To assess whether p38 inhibition had a therapeutic effect in vivo, we first infected groups of mice with a lethal dose (1 × 107 Nc-1 tachyzoites/mouse) for 100% of the animals (LD100), in order to evaluate clinical parameters as survival (Figure 3A). Infected mice treated with p38 inhibitor SB239063 did not show significant differences in the body weight or temperature loss, if compared to the other groups (P > 0.05, data not shown). Strikingly, the treatment rescued 84% of the mice from LD100, yielding a significant statistical increase in the survival rate (P = 0.0042), if compared to the untreated group of animals.
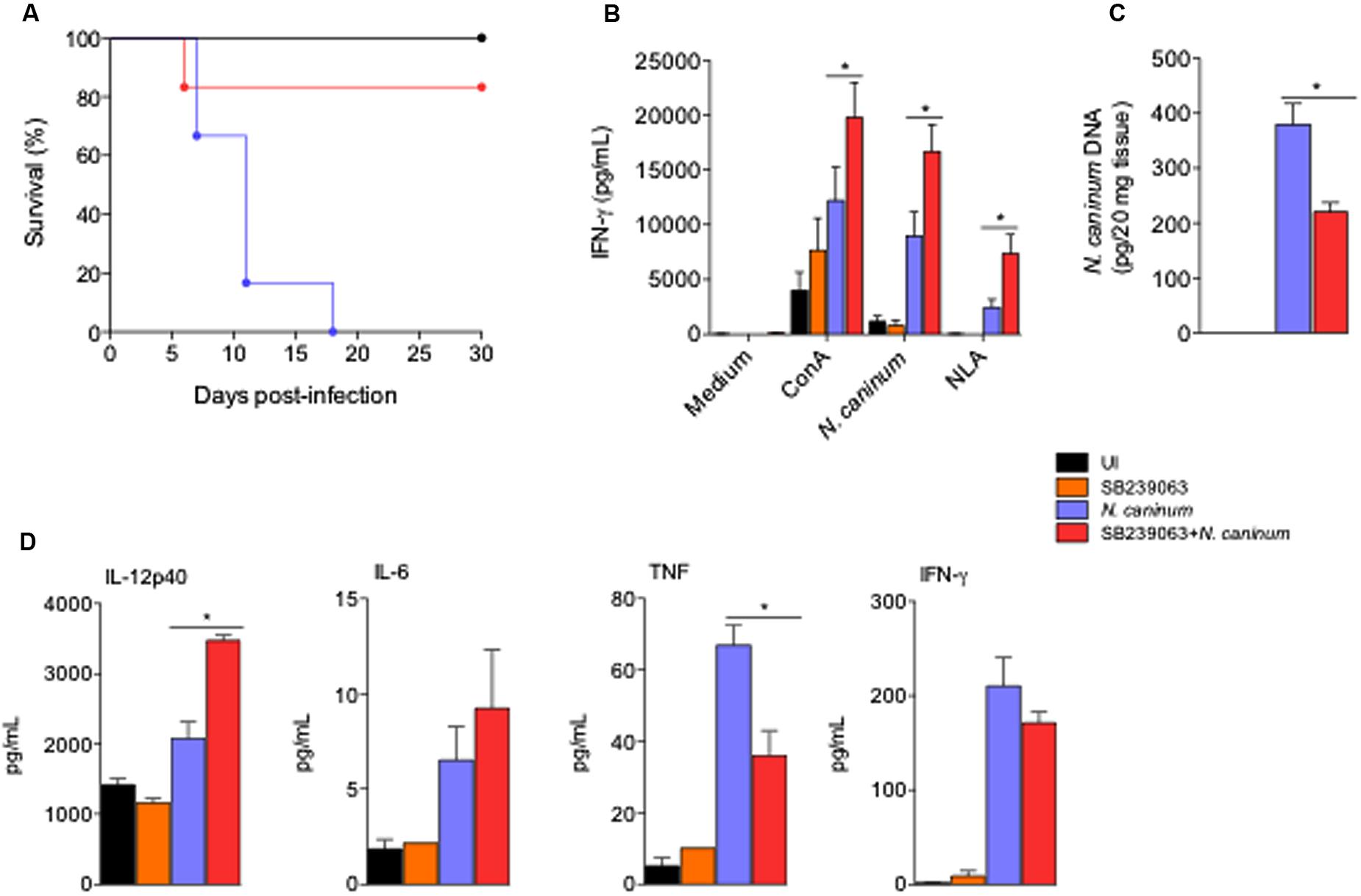
FIGURE 3. Treatment with p38 inhibitor improved immune responses and increased survival of mice infected with N. caninum. WT mice were treated or not with p38 inhibitor (SB239063, 0.5 mg/Kg) for 5 days post infection in order to observe survival. As controls, mice were inoculated with SB239063 alone (inhibitor control), or PBS (uninfected - UI). (A) Survival curves (six mice/group). (B) IFN-γ production during antigenic recall of spleen cells recovered from mice after 30 days of sub-lethal infection. Spleen cells (1 × 106/ml; five technical replicates) were cultured in the presence of mitogen (ConA, 2.5 μg/ml); live Nc-1 tachyzoites (1:10 parasite to cell ratio), soluble lysate antigen (NLA, 10 μg/ml) or medium alone. Supernatants were collected after 48 h and analyzed by ELISA. ∗Indicates statistically significant differences (P < 0.05) between untreated mice and SB239063-treated mice, upon antigenic stimulation, assessed by ANOVA followed by Bonferroni multiple comparison post-test to examine all possible pairwise comparisons. Results are representative of two independent experiments, with five technical replicates each. (C) Brain parasite load after 30 days of sub-lethal infection with N. caninum in the different test groups (five mice/group), analyzed by real-time PCR. ∗Indicates statistically significant differences (P < 0.05) between untreated mice and SB239063-treated mice, assessed by Kruskal–Wallis followed by Dunn’s multiple comparison post-test. Results are representative of two independent experiments, with three technical replicates each. (D) Cytokine production in the serum of mice after 7 days of sub-lethal infection treated or not with SB239063, analyzed by ELISA and CBA. Results are expressed as mean ± SEM of cytokine levels (five mice/group), extrapolated in relation to each assays’ standard curve, and representative of two independent experiments, with two technical replicates/each. ∗Indicates statistically significant differences (P < 0.05) between untreated mice and SB239063-treated mice; assessed by ANOVA followed by Bonferroni multiple comparison post-test to examine all possible pairwise comparisons.
We also examined whether the treatment with p38 inhibitor would affect the antigenic recall in chronically infected mice. With that intent, we observed the production of IFN-γ in ex vivo stimulation of spleen cells obtained from the different groups of mice during chronic phase of the infection (30 days after inoculation) with sub-lethal tachyzoite doses (5 × 106 Nc-1 tachyzoites/mouse). Spleen cells from SB239063-treated and infected mice produced statistically higher concentration of IFN-γ upon stimulation with the mitogen Concanavalin A (ConA, P = 0.0029), live tachyzoites (1/10 parasites/cell ratio, P < 0.0001) and NLA (20 μg/mL, P = 0.0014), if compared to cells extracted from untreated and infected littermates (Figure 3B; Means and SEM are represented in Supplementary Table 1). In agreement, the group of mice treated with the p38 MAPK inhibitor presented a significantly reduced concentration of parasite genomic DNA in the central nervous system (P < 0.0001) than the infected mice group (Figure 3C). In another set of experiments, mice were infected with sub-lethal doses of tachyzoites and treated for 5 days with p38 inhibitor SB239063, in order to test how inhibition of p38 MAPK in vivo would affect the outcome of disease, through immune and parasitological parameters. We observed that mice infected and treated with the chemical inhibitor had higher serum concentration of pro-inflammatory cytokines as IL-12p40 than the untreated group (Figure 3D) during the acute phase of infection (7 days after inoculation).
To assess whether p38 inhibition would also be a target for immune-prophylactic strategy, we immunized mice with NLA (50 μg/mouse) and used SB239063 as adjuvant in the preparation of the test group. We observed that mice immunized with NLA plus SB239063 presented mean reduction of 75% in the concentration of N. caninum genomic DNA in the central nervous system compared to the PBS (P < 0.05, Supplementary Figure 5). We also sought to observe if p38 MAPK activation interfered with the production of specific antibodies against N. caninum. With that intent, we measured the production kinetics of specific IgG antibodies and its subclasses IgG1 and IgG2abc, in the serum of mice 0, 15, 30, and 45 days after immunization. The ELISA assays determined that mice immunized with SB239063 presented similar production of specific antibody against N. caninum (Supplementary Figure 6), indicating that p38 signaling blockade does not affect antibody production after immunization against N. caninum.
GPCR, PI3K, and AKT Are Required for Neospora-Induced p38 MAPK Downregulation of IL-12 Production
To define the molecular mechanisms that underlie the regulatory effects of parasite mediated p38 activation, we tested distinct signaling pathways with known (direct or indirect) ability of inducing p38 phosphorylation. Neospora-infected BMDMs were treated with chemical/biological inhibitors of G-coupled protein receptors (GPCRs, pertussis toxin, Figure 4A), PI3K (Wortmannin, Figure 4B) and AKT (AKT inhibitor VIII, Figure 4C), combined or not with p38 inhibition (SB203580), in order to observe a possible additive effect in the IL-12p40 production. That experimental approach led to the observation that those inhibitors yielded exactly the same increase in cytokine production as SB203580, as well as the double inhibition of infected BMDMs, strongly suggesting that Neospora-induced p38 activation was dependent on upstream signaling of GPCR, PI3K, and AKT. To confirm that PI3K played a crucial role in the activation of p38 MAPK, we observed the cytokine production of BMDMs genetically depleted of PI3K (PI3K-/-) and infected by N. caninum (Figure 4D). In this set of experiments, we observed that the absence of PI3K induced similar increase of IL-12p40 production as that induced by inhibition of p38 MAPK. Also, the pretreatment of the chemical inhibitors SB203580 and Wortmannin in PI3K-/- BMDMs did not alter the increased IL-12p40 production after exposure to tachyzoites. In order to provide direct evidence to corroborate that Neospora-induced p38 activation was dependent of PI3K signaling, we also observed that PI3K inhibition by Wortmannin abrogated p38 MAPK phosphorylation in BMDMs after 30 min of infection with N. caninum tachyzoites (Figure 4E).
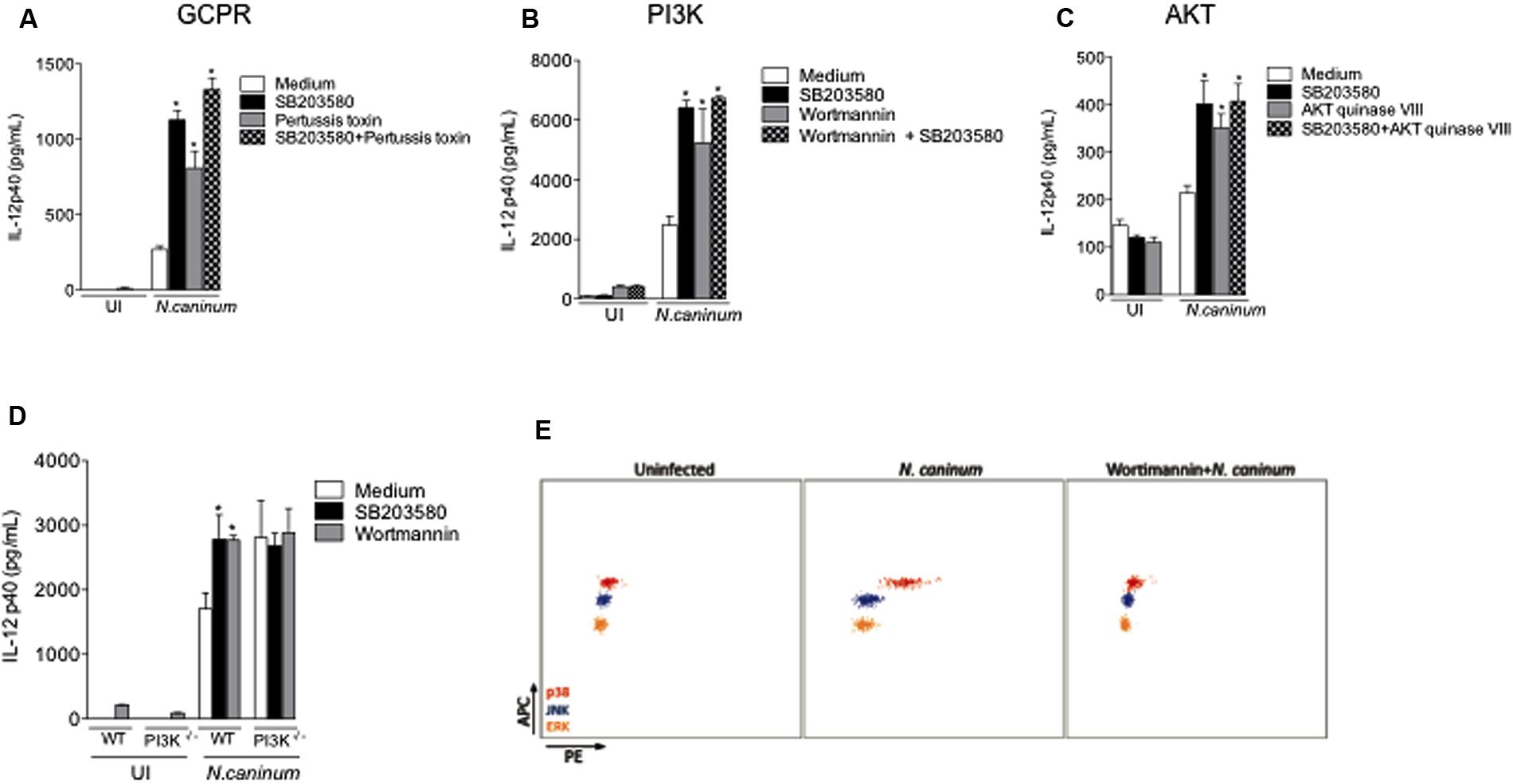
FIGURE 4. N. caninum infection activates p38 MAPK signaling through GPCR and PI3K/AKT pathways. Bone marrow-derived macrophages (BMDMs) of WT mice were pretreated with inhibitors to (A) GPCR (Pertussis toxin, 200 ng/ml), (B) PI3K (Wortmannin, 500 nM), (C) AKT (AKT inhibitor VIII, 10 μM), in addition or not to p38 inhibitor (SB203580, 10 μM), for 3 h, and sequentially infected by N. caninum live tachyzoites (Nc-1; 1:1 ratio of parasites to cells), or left uninfected (UI). After 24 h of infection, the supernatants were collected for IL-12p40 measurement. (D) BMDMs generated from WT mice and genetically deficient littermates in PI3K (PI3K-/-) were inhibited by SB203580 and/or Wortmannin, prior to infection with N. caninum live tachyzoites (Nc-1; 1:1 parasite to cell ratio) for the measurement of IL-12p40. ∗Indicates statistically significant differences between untreated cells and SB203580-treated cells, upon antigenic stimulation; P < 0.05, assessed by ANOVA followed by Bonferroni multiple comparison post-test to examine all possible pairwise comparisons. Results are representative of at least five independent experiments, with five technical replicates each. (E) Representative dot plots of p38 phosphorylation in WT BMDMs inhibited or not by Wortmannin for 3 h, and subsequently infected with N. caninum (Nc-1; 1:1 ratio) for 30 min, measured by CBA. Results are representative of three independent experiments, with three technical replicates/each.
We also assessed whether CCR5, MyD88, NF-κB, AP-1, mTOR, and JAK2 were involved in p38 activation in BMDMs during N. caninum infection, through the measurement of the concentration of IL-12p40 after 24 h. As previously shown by our group (Mineo et al., 2009), genetic deficiency in MyD88 (MyD88-/-) completely abolished IL-12 production after exposure to live parasites (Supplementary Figure 7A). Genetically deficient BMDMs in CCR5 (CCR5-/-) or treatment with specific inhibitors to JAK2 (Tyrphostin AG490), NF-κB (CAPE), AP-1 (Tanshinone) or mTOR (Rapamycin) did not yield significant differences in cytokine production after infection, if compared to WT or untreated controls. Therefore, we conclude that the phenomena herein described, induced by p38 activation, are independent of those molecules (Supplementary Figures 7B–F).
N. caninum-Triggered p38 Activation Is Induced by a Distinct Mechanism than T. gondii’s GRA24 Protein
To assess whether the mechanisms herein described are similar to those driven by T. gondii’s type II exclusive protein GRA24 (TgGRA24), we have genetically modified N. caninum tachyzoites to express this secreted antigen. For that purpose, we have generated a construct containing TgGRA24 with a C-terminal hemagglutinin (HA) tag plus BirA∗, driven by N. caninum GRA7 promoter (Figure 5A). TgGRA24 was integrated into the genome of NcLivΔHPT (parental) parasites, and localization of the gene fusion was assessed by immunofluorescence assay (IFA) using BirA/streptavidin interaction, as previously described (Chen et al., 2015). As expected, IFA showed that tagged GRA24 was correctly targeted to the dense granules, as it colocalized with GRA2 protein dispersion in the parasitophorous vacuole (data not shown), and the BirA∗-streptavidin complex further corroborated this pattern (Figure 5B), demonstrating that the construct for heterologous protein expression was successful.
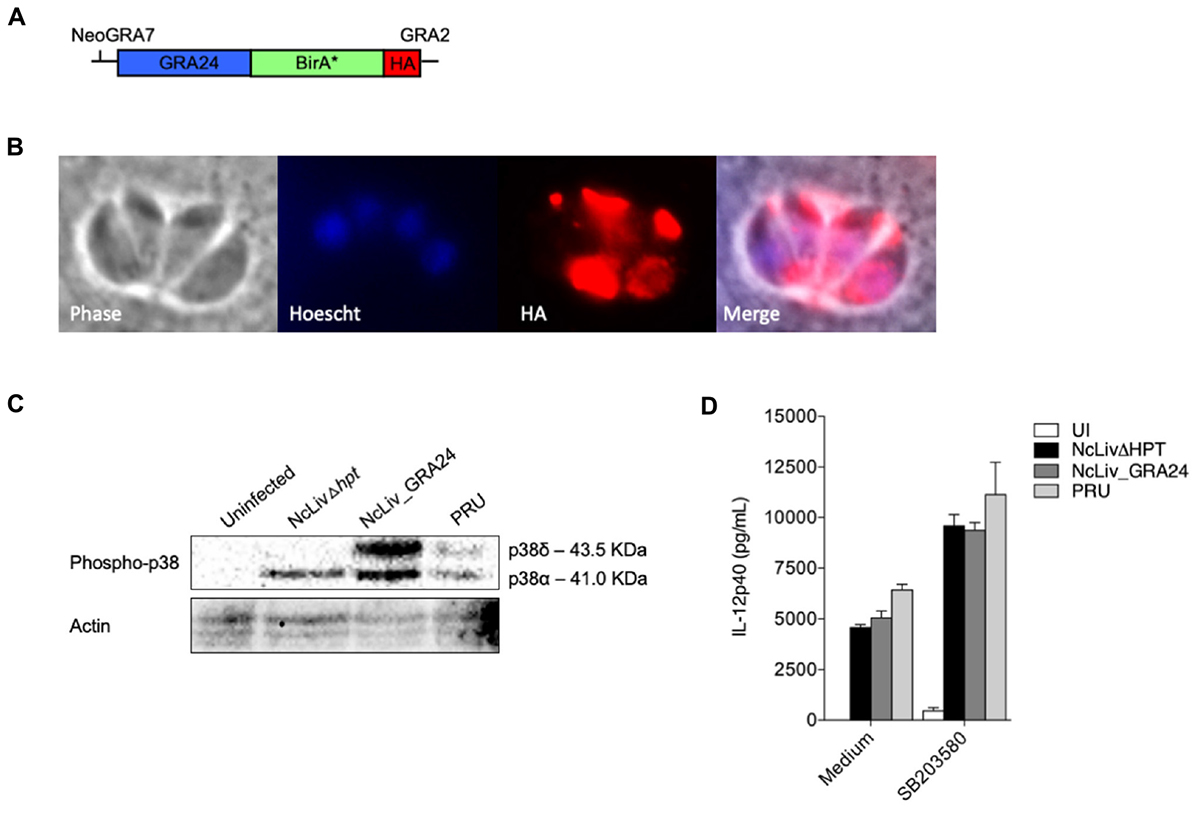
FIGURE 5. N. caninum-induced p38 phosphorylation in macrophages is triggered by a distinct mechanism than T. gondii’s GRA24 protein. (A) Diagram of the expression cassette encoding GRA24 fused to BirA∗, plus a 1C-terminal 3 HA epitope tag, driven by the N. caninum GRA7 promoter; (B) IFA of GRA24-BirA∗-expressing parasites, grown for 48 h. GRA24-BirA∗ localizes to the parasitophorous vacuole. Red, mouse anti-HA antibody; Blue, Hoescht; (C) BMDMs (1 × 106 cells/ml) were infected with live tachyzoites of N. caninum and T. gondii (NcLivΔHPT, NcLiv_GRA24, PRU; 1:1 parasite to cell ratio). After 30 min or 18 h, the cells were lysed and submitted to Western blot of p38 phosphorylation. Results are representative of at least two independent experiments. (D) BMDMs (1 × 106 cells/ml) were pretreated for 3 h with p38 inhibitor (SB203580, 10 μM) and infected with live tachyzoites of N. caninum and T. gondii (NcLivΔHPT, NcLiv_GRA24, PRU; 1:1 ratio) for 24 h. The supernatants were collected and the concentration of IL-12p40 was measured by ELISA. Results were expressed as mean ± SEM, and are representative of at least two independent experiments, with five technical replicates each.
Although MS experiments with N. caninum GRA24-BirA∗ expressing tachyzoites did not retrieve biotinylated p38 MAPK within its results, we continued to investigate whether the mechanism behind p38 pathway triggered by N. caninum shares common features with those described for TgGRA24, BMDMs were infected for 30 min and 18 h by parental (NcLivΔHPT), TgGRA24+ N. caninum (NcLiv_GRA24) or type II T. gondii (PRU) tachyzoites. As seen in Figure 5C, NcLivΔHPT induced a significantly less robust p38 activation compared to parasites that expressed type II TgGRA24 (NcLiv_GRA24 and PRU), independently if observed after 30 min or 18 h of exposure to the tachyzoites. Finally, we assessed if the addition of TgGRA24 in N. caninum tachyzoites would further enhance IL-12 production. For that purpose, cells were treated with p38 inhibitor SB203580 and infected with NcLivΔHPT, NcLiv_GRA24 or PRU tachyzoites. This assay demonstrated that all tested parasites induced similar cytokine production, as inhibition of p38 MAPK induced higher IL-12p40 production in all infected BMDMs, if compared to infected and untreated cells (Figure 5D). These results show that TgGRA24 does not further negatively interfere on IL-12p40 production in macrophages infected with N. caninum, demonstrating that the mechanisms herein reported—downregulation of IL-12 by activation of the p38 MAPK pathway by Neospora’s antigens—are distinct from those previously described for T. gondii (Braun et al., 2013), although it also makes us speculate whether the ability to evade innate immune responses through the GCPR/PI3K/AKT/p38 pathway is preserved between the parasites.
Discussion
Control of neosporosis involves the introduction of standard biosecurity measures; actually control programs are usually based on decreasing the vertical transmission in cattle by reducing the number of seropositive herd and/or decreasing the risk of horizontal transmission of N. caninum, mainly by the control of domestic definitive host population as a source of oocyst contamination. Afterward, different control measures have been suggested, however, the strategies have been ineffective (Dubey et al., 2007; Reichel et al., 2015). Thus, the increase in knowledge about the host immune response after infection by this parasite is essential to develop effective vaccine and treatment protocol.
Macrophages are the most important phagocytic cells in mammals and play a key role in the pathogens detection and elimination. Together with DCs, they provide the first line of cell-mediated defense and are pivotal in controlling the initial dissemination and/or growth of intracellular parasites (Donahoe et al., 2015; Hecker et al., 2015). Macrophage functions include synthesis of cytokines, such as IL-12, co-stimulatory molecules and presentation of antigens to T cells through the MHC that trigger adaptive immune responses, as well as antimicrobial mechanisms such as phagocytosis and NO synthase (Mineo et al., 2009; Monney and Hemphill, 2014). Earlier studies have convincingly shown that intracellular parasites like T. gondii and Leishmania sp. induces the MAPK pathway as modulatory mechanism in the IL-12 production in macrophages, and that this response is dependent of the phosphorylation of p38 MAPK (Rincón et al., 1998; Gigley et al., 2009; Yang et al., 2010; Dion et al., 2011; Quan et al., 2015). Therefore, MAPK pathway plays a key role in the control of intracellular infection, resulting in regulation of inflammatory cytokines that compose the Th1 immune responses. Therefore, we assessed for the first time the role of MAPK pathway in N. caninum-infected BMDMs, and we observed that N. caninum induced early p38 MAPK phosphorylation, as previously described in infection of macrophages by other Apicomplexa parasites (Kim et al., 2005; Horton et al., 2011; Klotz et al., 2011). We also observed that the robust p38 phosphorylation was opposed to little to absent induction of ERK1/2 and JNK. This conclusion maybe drawn regardless of the technique applied, since western blotting and CBA yielded similar results.
We analyzed if distinct antigenic fractions of N. caninum would be capable of inducing of the same p38 activation profile than live tachyzoites, and found that all antigens tested (NLA and ESA) were able to induce the activation of p38. Apicomplexan parasites are able to actively invade host cells using the action of apical secretory organelles named the micronemes, rhoptries, and dense granules. The proteins impacts host signaling pathways to serve as “master regulators” of the immune system (Melo et al., 2011). Thus, the induction of p38 here observed could be induced by ESA, since recent work by others has emphasized that early phase parasite secretion of kinases and phosphatases co-opt host cells by interfering with their signaling pathways (Dubremetz, 2007; Hajagos et al., 2012; Jensen et al., 2013; Bougdour et al., 2014). Braun et al. (2013) identified GRA24 as a strong agonist of host p38-alpha. However, there are no apparent GRA24 orthologs within the N. caninum genome. The use of N. caninum as a heterologous system for the expression of foreign genes such as T. gondii has been explored, and it may prove to be a tool for the parasite factors identification that are involved in the phenotypic differences stated between these apicomplexa parasites (Lei et al., 2014). The results showed that N. caninum expressing TgGRA24 induced higher activation of the subunit p38-alpha than the parental line, although it is noteworthy to point out that the BirA∗ bared within the construct was not tested to whether it would interfere with the observed phenomena. We actually speculate that N. caninum may activate a distinct subunit of p38 MAPK, since the other biological phenotypes assessed were distinct than that induced by NcLiv_GRA24 or PRU. Previous studies reported about some surface genes and secreted proteins, which are known to control virulence and host interactions in T. gondii infection, are altered in Neospora genome with different expression and functionality, which imply that there have been significant changes in the evolution of host-interacting genes between these species (Reid et al., 2012; Ramaprasad et al., 2015; Silmon de Monerri and Weiss, 2015). Hence, the pursuit for such antigenic targets with potential p38 activation is desired and should be better investigated.
The p38 pathway is associated with cytokine production, inflammation, cell growth, differentiation, and death (Terrazas et al., 2011). Studies with other infectious parasites, as Leishmania, have considered the role of MAP kinases in the regulation of IL-12 production (Yang et al., 2007, 2010; Ruhland and Kima, 2009), however, there is no clear consensus on the role of p38 activation in the regulation of this key cytokine. In the present study, we have identified that p38 plays a crucial role in dampening the cellular immune responses of mice infected by N. caninum, since mice treated with specific p38 inhibitors had increased pro-inflammatory cytokine production, lower chronic phase parasite burden and higher survival than untreated mice. Also, the use of p38 inhibitors as an adjuvant in vaccine protocols could block the immunosuppressant effects of N. caninum antigens, by significantly reducing the parasite load of challenged mice. It is noteworthy that inhibition of p38 alone during the immunization schemes was able to completely abrogate chronic phase parasitism, fact which should be further explored in future experiments. An additional remark should be made toward the possible direct effects of the p38 inhibitor in the tachyzoites, during in vivo treatment. Wei et al. (2002) reported that SB203580 or SB202190 treatment in T. gondii-infected fibroblasts significantly inhibited intracellular tachyzoite replication. Also, in a Leishmania major model, mice vaccinated with heat-killed L. major plus CpG and SB203580 elicited complete protection against the infection (Horton et al., 2011), and a similar protection was observed when p38 MAPK blockade inhibited the replication of Plasmodium falciparum in human erythrocytes cultured ex vivo (Brumlik et al., 2011). Thus, this work corroborates and adds new information to the literature that p38 MAPK inhibitors may be a potential active principle to improve treatment and vaccines against intracellular protozoa.
The precise mechanism underlying the phenomena described in this work is yet to be unveiled. Our data show that one of the interactions between N. caninum and macrophages is induced through activation of the GPCR/PI3K/AKT pathway, which by its turn induces p38 phosphorylation and consequent blockade of crucial mediators required for an accurate immune response against the parasite. In agreement with this finding, studies demonstrated that inhibition of PI3K/Akt signaling resulted in increased IL-12 production against infections by intracellular parasites, although these studies were inconclusive on how this signaling pathway could be mediated by p38 MAPK (Ruhland and Kima, 2009; Quan et al., 2015). In order to describe this pathway as that responsible for this downregulation of the innate immune responses against the parasite, we have also checked if other known signaling pathways were involved. None yielded results that agree with the phenotype observed. To rationalize our choices for the pathways, we initially assessed MyD88, a major adaptor protein for Toll-like receptor (TLR) signaling, which our group previously described as critical for acute resistance to N. caninum infection (Mineo et al., 2009), especially regarding the induction of IL-12p40. We have also assessed the participation of CCR5, a GCPR-dependent chemokine receptor that is known to induce MyD88-independent inflammatory cell migration during N. caninum infection (Mineo et al., 2010a; Bonfá et al., 2014; Abe et al., 2015). During T. gondii and N. caninum infections, parasite-derived cyclophilin 18 induces IL-12 and IFN-γ production by binding to CCR5. Moreover, N. caninum cyclophilin caused CCR5-dependent migration of murine and bovine cells (Aliberti et al., 2000; Ibrahim et al., 2009; Kameyama et al., 2012). We also showed that none of the classical transcription factors is directly involved with upstream phosphorylation of p38 MAPK, since IL-12p40 production was not affected by the inhibition of JAK2, NF-κB, AP-1 and mTOR. In this sense, p38 MAPK may regulate alternative transcription factors, as the ATF family - nuclear factor-erythroid 2-related factor 2 (Nrf2), Egr-1 and c-Fos which induce a wide variety of physical, chemical and biological effect that should be further investigated (Chakrabarti et al., 2014; Chi et al., 2015).
Conclusion
N. caninum downregulates, possibly through parasite secreted-excreted antigens, the signaling pathway composed of PI3K/AKT and p38 MAPK pathways downstream of GPCR signaling. Our results indicate that N. caninum manipulates those pathways in its favor, in order to evade the host’s innate immune responses, increasing acute phase replication and consequently chronic phase parasite burden. In that sense, that pathway may be a potential target for therapeutic intervention against neosporosis.
Author Contributions
Conceived and designed the experiments: CM, TM, and PB. Performed the experiments: CM, AO, MD-F, MS, SN, and AV. Analyzed the data: CM, TM, and PB. Contributed reagents/materials/analysis tools: CM, TM, FS, JW, PB, JdS, and JM. Wrote the paper: CM, TM, and PB.
Funding
This work was supported by Brazilian Funding Agencies Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES; AUX-PE-PARASITOLOGIA 1348/2011), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; 309011/2013-2), and Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG; CDS-RED-00013-14, CVZ-PPM-00784-15). We hereby state that the funding agencies had no role in study design, data collection and interpretation, or in the decision to submit the work for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
The authors thank to Marley Dantas Barbosa, Zilda Mendonça da Silva, and Ana Claúdia Pajuaba for their technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01456
Footnotes
References
Abe, C., Tanaka, S., Nishimura, M., Ihara, F., Xuan, X., and Nishikawa, Y. (2015). Role of the chemokine receptor CCR5-dependent host defense system in Neospora caninum infections. Parasit Vectors. 8:5. doi: 10.1186/s13071-014-0620-5
Aliberti, J. (2005). Host persistence: exploitation of anti-inflammatory pathways by Toxoplasma gondii. Nat. Rev. Immunol. 5, 162–170. doi: 10.1038/nri1547
Aliberti, J., Reis e Sousa, C., Schito, M., Hieny, S., Wells, T., Huffnagle, G. B., et al. (2000). CCR5 provides a signal for microbial induced production of IL-12 by CD8 alpha+ dendritic cells. Nat. Immunol. 1, 83–87. doi: 10.1038/76957
Beiting, D. P., Peixoto, L., Akopyants, N. S., Beverley, S. M., Wherry, E. J., Christian, D. A., et al. (2014). Differential induction of TLR3-dependent innate immune signaling by closely related parasite species. PLoS ONE 9:e88398. doi: 10.1371/journal.pone.0088398
Bonfá, G., Benevides, L., Souza, M. C., Fonseca, D. M., Mineo, T. W., Rossi, M. A., et al. (2014). CCR5 controls immune and metabolic functions during Toxoplasma gondii infection. PLoS ONE 9:e104736. doi: 10.1371/journal.pone.0104736
Bougdour, A., Tardieux, I., and Hakimi, M. A. (2014). Toxoplasma exports dense granule proteins beyond the vacuole to the host cell nucleus and rewires the host genome expression. Cell Microbiol. 16, 334–343. doi: 10.1111/cmi.12255
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1006/abio.1976.9999
Braun, L., Brenier-Pinchart, M. P., Yogavel, M., Curt-Varesano, A., Curt-Bertini, R. L., Hussain, T., et al. (2013). A Toxoplasma dense granule protein, GRA24, modulates the early immune response to infection by promoting a direct and sustained host p38 MAPK activation. J. Exp. Med. 210, 2071–2086. doi: 10.1084/jem.20130103
Brumlik, M. J., Pandeswara, S., Ludwig, S. M., Murthy, K., and Curiel, T. J. (2011). Parasite mitogen-activated protein kinases as drug discovery targets to treat human protozoan pathogens. J. Signal Transduct. 2011, 971968. doi: 10.1155/2011/971968
Cardoso, M. R. D., Mota, C. M., Ribeiro, D. P., Santiago, F. M., Carvalho, J. V., Araujo, E. C. B., et al. (2011). ArtinM, a d-mannose-binding lectin from Artocarpus integrifolia, plays a potent adjuvant and immunostimulatory role in immunization against Neospora caninum. Vaccine 29, 9183–9193. doi: 10.1016/j.vaccine.2011.09.136
Chakrabarti, S., Poidevin, M., and Lemaitre, B. (2014). The Drosophila MAPK p38c regulates oxidative stress and lipid homeostasis in the intestine. PLoS Genet. 10:e1004659. doi: 10.1371/journal.pgen.1004659
Chen, A. L., Kim, E. W., Toh, J. Y., Vashisht, A. A., Rashoff, A. Q., Van, C., et al. (2015). Novel components of the Toxoplasma inner membrane complex revealed by BioID. MBio 6, e2357–e2314. doi: 10.1128/mBio.02357-14
Chi, P. L., Lin, C. C., Chen, Y. W., Hsiao, L. D., and Yang, C. M. (2015). CO induces Nrf2-dependent heme oxygenase-1 transcription by cooperating with Sp1 and c-Jun in rat brain astrocytes. Mol. Neurobiol. 52, 277–292. doi: 10.1007/s12035-014-8869-4
Cuadrado, A., and Nebreda, A. R. (2010). Mechanisms and functions of p38 MAPK signaling. Biochem. J. 429, 403–417. doi: 10.1042/BJ20100323
Debierre-Grockiego, F., Campos, M. A., Azzouz, N., Schmidt, J., Bieker, U., Resende, M. G., et al. (2007). Activation of TLR2 and TLR4 by glycosylphosphatidylinositols derived from Toxoplasma gondii. J. Immunol. 179, 1129–1137. doi: 10.4049/jimmunol.179.2.1129
Dion, S., Germon, S., Guiton, R., Ducournau, C., and Dimier-Poisson, I. (2011). Functional activation of T cells by dendritic cells and macrophages exposed to the intracellular parasite Neospora caninum. Int. J. Parasitol. 41, 685–695. doi: 10.1016/j.ijpara.2011.01.008
Donahoe, S. L., Lindsay, S. A., Krockenberger, M., Phalen, D., and Šlapeta, J. (2015). A review of neosporosis and pathologic findings of Neospora caninum infection in wildlife. Int. J. Parasitol. Parasites Wildl. 4, 216–238. doi: 10.1016/j.ijppaw.2015.04.002
Dubey, J. P., Schares, G., and Ortega-Mora, L. M. (2007). Epidemiology and control of neosporosis and Neospora caninum. Clin. Microbiol. Rev. 20, 323–367. doi: 10.1128/CMR.00031-06
Dubremetz, J. F. (2007). Rhoptries are major players in Toxoplasma gondii invasion and host cell interaction. Cell Microbiol. 9, 841–848. doi: 10.1111/j.1462-5822.2007.00909.x
Eastick, F. A., and Elsheikha, H. M. (2010). Stress-driven stage transformation of Neospora caninum. Parasitol. Res. 106, 1009–1014. doi: 10.1007/s00436-010-1788-z
Gigley, J. P., Fox, B. A., and Bzik, D. J. (2009). Cell-mediated immunity to Toxoplasma gondii develops primarily by local Th1 host immune responses in the absence of parasite replication. J. Immunol. 182, 1069–1078. doi: 10.4049/jimmunol.182.2.1069
Goodswen, S., Kennedy, P., and Ellis, J. (2012). A review of the infection, genetics, and evolution of Neospora caninum: from the past to the present. Infect. Genet. Evol. 13, 133–150. doi: 10.1016/j.meegid.2012.08.012
Hajagos, B. E., Turetzky, J. M., Peng, E. D., Cheng, S. J., Ryan, C. M., Souda, P., et al. (2012). Molecular dissection of novel trafficking and processing of the Toxoplasma gondii rhoptry metalloprotease toxolysin-1. Traffic 13, 292–304. doi: 10.1111/j.1600-0854.2011.01308.x
Hecker, Y. P., Cantón, G., Regidor-Cerrillo, J., Chianini, F., Morrell, E., Lischinsky, L., et al. (2015). Cell mediated immune responses in the placenta following challenge of vaccinated pregnant heifers with Neospora caninum. Vet. Parasitol. 214, 247–254. doi: 10.1016/j.vetpar.2015.10.015
Horton, A. A., Wang, B., Camp, L., Price, M. S., Arshi, A., Nagy, M., et al. (2011). The mitogen-activated protein kinome from Anopheles gambiae: identification, phylogeny and functional characterization of the ERK, JNK and p38 MAP kinases. BMC Genomics 12:574. doi: 10.1186/1471-2164-12-574
Ibrahim, H. M., Bannai, H., Xuan, X., and Nishikawa, Y. (2009). Toxoplasma gondii cyclophilin 18-mediated production of nitric oxide induces Bradyzoite conversion in a CCR5-dependent manner. Infect. Immun. 77, 3686–3695. doi: 10.1128/IAI.00361-09
Jenkins, M. C., Tuo, W., Feng, X., Cao, L., Murphy, C., and Fetterer, R. (2010). Neospora caninum: cloning and expression of a gene coding for cytokine-inducing profiling. Exp. Parasitol. 125, 357–362. doi: 10.1016/j.exppara.2010.03.001
Jensen, K. D., Hu, K., Whitmarsh, R. J., Hassan, M. A., Julien, L., Lu, D., et al. (2013). Toxoplasma gondii rhoptry 16 kinase promotes host resistance to oral infection and intestinal inflammation only in the context of the dense granule protein GRA15. Infect. Immun. 81, 2156–2167. doi: 10.1128/IAI.01185-12
Kameyama, K., Nishimura, M., Punsantsogvoo, M., Ibrahim, H. M., Xuan, X., Furuoka, H., et al. (2012). Immunological characterization of Neospora caninum cyclophilin. Parasitology 39, 294–301. doi: 10.1017/S0031182011002022
Kawai, T., and Akira, S. (2010). The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat. Immunol. 11, 373–384. doi: 10.1038/ni.1863
Kim, L., Del Rio, L., Butcher, B. A., Mogensen, T. H., Paludan, S. R., Flavell, R. A., et al. (2005). p38 MAPK autophosphorylation drives macrophage IL-12 production during intracellular infection. J. Immunol. 174, 4178–4184. doi: 10.4049/jimmunol.174.7.4178
Kim, L., and Denkers, E. Y. (2006). Toxoplasma gondii triggers Gi-dependent PI 3-kinase signaling required for inhibition of host cell apoptosis. J. Cell Sci. 119, 2119–2126. doi: 10.1242/jcs.02934
Klotz, C., Ziegler, T., Figueiredo, A. S., Rausch, S., Hepworth, M. R., Obsivac, N., et al. (2011). A helminth immunomodulator exploits host signaling events to regulate cytokine production in macrophages. PLoS Pathog. 7:e1001248. doi: 10.1371/journal.ppat.1001248
Lei, T., Wang, H., Liu, J., Nan, H., and Liu, Q. (2014). ROP18 is a key factor responsible for virulence difference between Toxoplasma gondii and Neospora caninum. PLoS ONE 9:e99744. doi: 10.1371/journal.pone.0099744
Marim, F. M., Silveira, T. N., Lima, D. S. Jr., and Zamboni, D. S. (2010). A method for generation of bone marrow-derived macrophages from cryopreserved mouse bone marrow cells. PLoS ONE 5:e15263. doi: 10.1371/journal.pone.0015263
Masek, K. S., Fiore, J., Leitges, M., Yan, S. F., Freedman, B. D., and Hunter, C. A. (2006). Host cell Ca2+ and protein kinase C regulate innate recognition of Toxoplasma gondii. J. Cell Sci. 119, 4565–4573. doi: 10.1242/jcs.03319
Melo, M. B., Jensen, K. D., and Saeij, J. P. (2011). Toxoplasma gondii effectors are master regulators of the inflammatory response. Trends Parasitol. 27, 487–495. doi: 10.1016/j.pt.2011.08.001
Mineo, T. W., Benevides, L., Silva, N. M., and Silva, J. S. (2009). Myeloid differentiation factor 88 is required for resistance to Neospora caninum infection. Vet. Res. 40:32. doi: 10.1051/vetres/2009015
Mineo, T. W., Oliveira, C. J., Silva, D. A., Oliveira, L. L., Abatepaulo, A. R., Ribeiro, D. P., et al. (2010a). Neospora caninum excreted/secreted antigens trigger CC-chemokine receptor 5-dependent cell migration. Int. J. Parasitol. 40, 797–805. doi: 10.1016/j.ijpara.2009.12.003
Mineo, T. W., Oliveira, C. J. F., Gutierrez, F. R. S., and Silva, J. S. (2010b). Recognition by toll-like receptor 2 induces antigen presenting cell activation and Th1 programming during infection by Neospora caninum. Immunol. Cell Biol. 88, 1–9. doi: 10.1038/icb.2010.52
Monney, T., and Hemphill, A. (2014). Vaccines against neosporosis: what can we learn from the past studies? Exp. Parasitol. 140, 52–70. doi: 10.1016/j.exppara.2014.02.015
Pinheiro, A. M., Costa, S. L., Freire, S. M., Ribeiro, C. S. O., Tardy, M., El-Bachá, R. S., et al. (2010). Neospora caninum: early immune response of rat mixed glial cultures after tachyzoites infection. Exp. Parasitol. 124, 442–447. doi: 10.1016/j.exppara.2009.12.018
Quan, J. H., Cha, G. H., Zhou, W., Chu, J. Q., Nishikawa, Y., and Lee, Y. H. (2013). Involvement of PI 3 kinase/Akt-dependent bad phosphorylation in Toxoplasma gondii-mediated inhibition of host cell apoptosis. Exp. Parasitol. 133, 462–471. doi: 10.1016/j.exppara.2013.01.005
Quan, J. H., Chu, J. Q., Kwon, J., Choi, I. W., Ismail, H. A., Zhou, W., et al. (2015). Intracellular networks of the PI3K/AKT and MAPK pathways for regulating Toxoplasma gondii-induced IL-23 and IL-12 production in human THP-1 cells. PLoS ONE 10:e0141550. doi: 10.1371/journal.pone.0141550
Ramaprasad, A., Mourier, T., Naeem, R., Malas, T. B., Moussa, E., Panigrahi, A., et al. (2015). Comprehensive evaluation of Toxoplasma gondii VEG and Neospora caninum LIV genomes with tachyzoite stage transcriptome and proteome defines novel transcript features. PLoS ONE 10:e0124473. doi: 10.1371/journal.pone.0124473
Reichel, M. P., Moore, D. P., Hemphill, A., Ortega-Mora, L. M., Dubey, J. P., and Ellis, J. T. (2015). A live vaccine against Neospora caninum abortions in cattle. Vaccine 33, 1299–1301. doi: 10.1016/j.vaccine.2015.01.064
Reid, A. J., Vermont, S. J., Cotton, J. A., Harris, D., Hill-Cawthorne, G. A., Könen-Waisman, S., et al. (2012). Comparative genomics of the apicomplexan parasites Toxoplasma gondii and Neospora caninum: coccidia differing in host range and transmission strategy. PLoS Pathog. 8:e1002567. doi: 10.1371/journal.ppat.1002567
Ribeiro, D. P., Freitas, M. M., Cardoso, M. R., Pajuaba, A. C., Silva, N. M., Mineo, T. W., et al. (2009). CpG-ODN combined with Neospora caninum lysate, but not with excreted-secreted antigen, enhances protection against infection in mice. Vaccine 27, 2570–2579. doi: 10.1016/j.vaccine.2009.02.028
Rincón, M., Enslen, H., Raingeaud, J., Recht, M., Zapton, T., Su, M. S., et al. (1998). Interferon-gamma expression by Th1 effector T cells mediated by the p38 MAP kinase signaling pathway. EMBO J. 17, 2817–2829. doi: 10.1093/emboj/17.10.2817
Ruhland, A., and Kima, P. E. (2009). Activation of PI3K/Akt signaling has a dominant negative effect on IL-12 production by macrophages infected with Leishmania amazonensis promastigotes. Exp. Parasitol. 122, 28–36. doi: 10.1016/j.exppara.2008.12.010
Schubert, R., Geiger, H., Zielen, S., and Baer, P. C. (2009). Simultaneous detection of ERK-, p38-, and JNK-MAPK phosphorylation in human adipose-derived stem cells using the Cytometric Bead Array technology. J. Immunol. Methods 350, 200–204. doi: 10.1016/j.jim.2009.08.015
Silmon de Monerri, N. C., and Weiss, L. M. (2015). Integration of RNA-seq and proteomics data with genomics for improved genome annotation in Apicomplexan parasites. Proteomics 15, 2557–2559. doi: 10.1002/pmic.201500253
Terrazas, C. A., Huitron, E., Vazquez, A., Juarez, I., Camacho, G. M., Calleja, E. A., et al. (2011). MIF synergizes with Trypanosoma cruzi antigens to promote efficient dendritic cell maturation and IL-12 production via p38 MAPK. Int. J. Biol. Sci. 7, 1298–1310. doi: 10.7150/ijbs.7.1298
Wei, S., Marches, F., Daniel, B., Sonda, S., Heidenreich, K., and Curiel, T. (2002). Pyridinylimidazole p38 mitogen-activated protein kinase inhibitors block intracellular Toxoplasma gondii replication. Int. J. Parasitol. 32, 969–977. doi: 10.1016/S0020-7519(02)00061-9
Yang, S. H., Sharrocks, A. D., and Whitmarsh, A. J. (2013). MAP kinase signalling cascades and transcriptional regulation. Gene 513, 1–13. doi: 10.1016/j.gene.2012.10.033
Yang, Z., Mosser, D. M., and Zhang, X. (2007). Activation of the MAPK, ERK, following Leishmania amazonensis infection of macrophages. J. Immunol. 178, 1077–1085. doi: 10.4049/jimmunol.178.2.1077
Yang, Z., Zhang, X., Darrah, P. A., and Mosser, D. M. (2010). The regulation of Th1 responses by the p38 MAPK. J. Immunol. 185, 6205–6213. doi: 10.4049/jimmunol.1000243
Yarovinsky, F., Zhang, D., Andersen, J. F., Bannenberg, G. L., Serhan, C. N., Hayden, M. S., et al. (2005). TLR11 activation of dendritic cells by a protozoan profilin-like protein. Science 308, 1626–1629. doi: 10.1126/science.1109893
Keywords: N. caninum, immune response, p38/MAPk, evasion, IL-12
Citation: Mota CM, Oliveira ACM, Davoli-Ferreira M, Silva MV, Santiago FM, Nadipuram SM, Vashisht AA, Wohlschlegel JA, Bradley PJ, Silva JS, Mineo JR and Mineo TWP (2016) Neospora caninum Activates p38 MAPK as an Evasion Mechanism against Innate Immunity. Front. Microbiol. 7:1456. doi: 10.3389/fmicb.2016.01456
Received: 16 June 2016; Accepted: 31 August 2016;
Published: 13 September 2016.
Edited by:
Alexandre Morrot, Federal University of Rio de Janeiro, BrazilReviewed by:
Celio Geraldo Freire De Lima, Federal University of Rio de Janeiro, BrazilLuis Gustavo Corbellini, Universidade Federal do Rio Grande do Sul, Brazil
Copyright © 2016 Mota, Oliveira, Davoli-Ferreira, Silva, Santiago, Nadipuram, Vashisht, Wohlschlegel, Bradley, Silva, Mineo and Mineo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tiago W. P. Mineo, dGlhZ28ubWluZW9AdWZ1LmJy