 María G. Cárdenas-Mondragón1†
María G. Cárdenas-Mondragón1† Miguel A. Ares1†
Miguel A. Ares1† Leonardo G. Panunzi2
Leonardo G. Panunzi2 Sabino Pacheco3Margarita Camorlinga-Ponce1Jorge A. Girón4Javier Torres1Miguel A. De la Cruz1*
Sabino Pacheco3Margarita Camorlinga-Ponce1Jorge A. Girón4Javier Torres1Miguel A. De la Cruz1*- 1Unidad de Investigación Médica en Enfermedades Infecciosas y Parasitarias, Hospital de Pediatria, Centro Médico Nacional Siglo XXI, IMSS, Mexico City, Mexico
- 2Centre d’Immunologie de Marseille-Luminy, Aix Marseille Université UM2, Inserm, U1104, CNRS UMR7280, Marseille, France
- 3Departamento de Microbiología Molecular, Instituto de Biotecnología UNAM, Cuernavaca, Mexico
- 4Centro de Detección Biomolecular, Benemérita Universidad Autónoma de Puebla, Puebla, Mexico
Helicobacter pylori is a Gram-negative bacterium that colonizes the human gastric mucosa and is responsible for causing peptic ulcers and gastric carcinoma. The expression of virulence factors allows the persistence of H. pylori in the stomach, which results in a chronic, sometimes uncontrolled inflammatory response. Type II toxin–antitoxin (TA) systems have emerged as important virulence factors in many pathogenic bacteria. Three type II TA systems have previously been identified in the genome of H. pylori 26695: HP0315–HP0316, HP0892–HP0893, and HP0894–HP0895. Here we characterized a heretofore undescribed type II TA system in H. pylori, HP0967–HP0968, which is encoded by the bicistronic operon hp0968–hp0967 and belongs to the Vap family. The predicted HP0967 protein is a toxin with ribonuclease activity whereas HP0968 is an antitoxin that binds to its own regulatory region. We found that all type II TA systems were expressed in H. pylori during early stationary growth phase, and differentially expressed in the presence of urea, nickel, and iron, although, the hp0968–hp0967 pair was the most affected under these environmental conditions. Transcription of hp0968–hp0967 was strongly induced in a mature H. pylori biofilm and when the bacteria interacted with AGS epithelial cells. Kanamycin and chloramphenicol considerably boosted transcription levels of all the four type II TA systems. The hp0968–hp0967 TA system was the most frequent among 317 H. pylori strains isolated from all over the world. This study is the first report on the transcription of type II TA genes in H. pylori under different environmental conditions. Our data show that the HP0967 and HP0968 proteins constitute a bona fide type II TA system in H. pylori, whose expression is regulated by environmental cues, which are relevant in the context of infection of the human gastric mucosa.
Introduction
Helicobacter pylori is a Gram-negative bacterium, member of the Epsilon proteobacteria that colonizes the human gastric mucosa (Marshall and Warren, 1984). A hallmark of H. pylori is its ability to survive in the hostile environment found in the stomach, which is attributed to the expression of various virulence factors, such as secretion systems, cytotoxins, flagella, and adhesins. In this regard, the H. pylori genome encodes a type IV secretion system, which is responsible for the translocation of the CagA effector protein into host epithelial gastric cells (Amieva and El-Omar, 2008; Tegtmeyer et al., 2011). In addition to CagA, H. pylori produces the vacuolating cytotoxin VacA, which is present in all H. pylori strains (Palframan et al., 2012). The toxin–antitoxin (TA) systems have emerged as important virulence factors in many pathogenic bacteria (Kedzierska and Hayes, 2016). At present, six types of TA systems have been described (Page and Peti, 2016) among which, the type II is highly prevalent in prokaryote genomes (Makarova et al., 2009). The functionality of these systems has been described to be beneficial in bacterial fitness, persistence, and virulence (Pandey and Gerdes, 2005; Lewis, 2007; Leplae et al., 2011; Kedzierska and Hayes, 2016). These systems are formed by a couple of TA proteins, where the toxin exerts a bacteriostatic effect on bacterial growth and the antitoxin neutralizes the toxin through a protein-protein interaction. A remarkable characteristic of the type II TA system is that the antitoxin acts as a repressor of its own transcription (Yamaguchi and Inouye, 2011; Yamaguchi et al., 2011). In terms of virulence, type II TA systems have been described to be important in the host–pathogen interactions, particularly in Gram-negative bacteria. In uropathogenic Escherichia coli, a type II TA module promotes the colonization of the mouse bladder or kidneys (Norton and Mulvey, 2012). The SehAB system was shown to be important for the survival of Salmonella enterica serotype Typhimurium in the mesenteric lymphoid nodes of mice (De la Cruz et al., 2013). Also, the RelBE TA system of Vibrio cholerae affects biofilm formation and intestinal colonization (Wang et al., 2015). Moreover, in both Leptospira interrogans and Rickettsia spp., type II TA mutants were affected in their induction of apoptosis in eukaryotic cells (Audoly et al., 2011; Komi et al., 2015). Thus, a growing body of evidence suggests that these emerging virulence factors display functional and regulatory roles in the expression of multiple virulence traits.
Three type II TA systems have been structurally and functionally characterized in H. pylori 26695: HP0315–HP0316, HP0892–HP0893, and HP0894–HP0895 (Han et al., 2011, 2013; Kwon et al., 2012; Pathak et al., 2013; Im et al., 2014). These TA systems belong to the Vap (virulence-associated protein) family, found in Bacteria and Archaea (Makarova et al., 2006). However, no transcriptional analyses have been reported for these TA systems in H. pylori under environmental conditions such as acidic pH or in the presence of urea, nickel, and iron, which affect the expression of virulence factors in this bacterium (Sachs et al., 2003; Haley and Gaddy, 2015; Keilberg and Ottemann, 2016).
In this work we identified and characterized a new type II TA system (HP0967–HP0968) in the genome of H. pylori strain 26695. Whilst HP0967 turned out to be a toxin with ribonuclease activity, HP0968 was found to work as an antitoxin that bound to its own regulatory region. Compared to the other three known type II TA systems, the hp0968–hp0967 system was highly expressed when H. pylori 26695 was grown in Brucella broth, particularly during the early stationary phase. Interestingly, metals such as nickel and iron repressed the expression of hp0968–hp0967, while urea had a moderate positive effect. Transcription of hp0968–hp0967 was highly induced in a mature biofilm and when H. pylori was interacting with AGS epithelial cells, whereas the other type II TA systems showed a weaker induction, suggesting that particularly this TA system might be activated in the environment of the host gastric mucosa prior to host colonization. The presence of antibiotics such as kanamycin and chloramphenicol dramatically boosted the transcription of hp0968–hp0967. The hp0968–hp0967 genes were present in both clinical isolates and in H. pylori strains whose genomes are publicly available. Our data show that HP0967 and HP0968 proteins form a type II TA system whose expression activates known virulence factors of H. pylori. This is the first report about regulation of transcription of H. pylori type II TA systems under different environmental conditions.
Materials and Methods
Bacterial Strains and Culture Conditions
Helicobacter pylori 26695 (Tomb et al., 1997) was grown for 2 days on blood agar culture plates containing 10% defibrinated sheep blood at 37°C under microaerophilic conditions, and a bacterial suspension was prepared in Brucella broth and adjusted to an optical density of 0.1 at 600 nm. H. pylori was grown at 37°C for 24 h (exponential phase) and 48 h (stationary phase) in Brucella broth supplemented with 10% fetal bovine serum (FBS; Invitrogen). In addition, Brucella broth adjusted to pH 5.5 or containing either urea [5 μM CO(NH2)2], nickel [250 μM NiCl2] or iron [150 μM (NH4)2Fe(SO4)2⋅6H2O], were tested as previously described (Contreras et al., 2003; Wen et al., 2003; Vannini et al., 2014). In all cases, the bacteria were incubated at 37°C with shaking and samples were collected 48 h post-inoculation for RNA extraction. Fold-change transcription was determined by calculating the relative expression of TA genes under different environmental conditions as compared to bacteria growing in Brucella broth.
Cloning and Purification of HP0967 and HP0968 Proteins
For cloning of hp0967 and hp0968 coding regions, specific primers (Table 1) containing the NcoI (5′)/HindIII (3′) restriction sites were used to obtain PCR products, which were digested with NcoI and HindIII and then ligated into pBAD-Myc-HisA previously digested with the same restriction enzymes. HP0967-Myc-His6 and HP0968-Myc-His6 proteins were purified with Ni-nitrilotriacetic acid resin. Briefly, E. coli BL21(DE3) carrying the pBAD-HP0967-Myc-His6 and pBAD-HP0968-Myc-His6 plasmids were grown to mid-logarithmic phase. L(+)-arabinose (Sigma-Aldrich) was added at final concentration of 0.1%, and the bacteria were grown at 6 h at 30°C. The cells were then pelleted by centrifugation, resuspended in PBS buffer and ruptured by sonication. The suspension was centrifuged and the supernatant was filtered through a Ni-nitrilotriacetic acid agarose column (QIAExpress, Qiagen) pre-equilibrated with PBS buffer. After extensive washing with binding buffer containing 50 μM imidazole (100 ml), the bound protein was eluted with 500 μM imidazole. Fractions were analyzed by SDS-PAGE. Protein concentration was determined by the Bradford procedure. Aliquots of the purified protein were stored at -70°C.
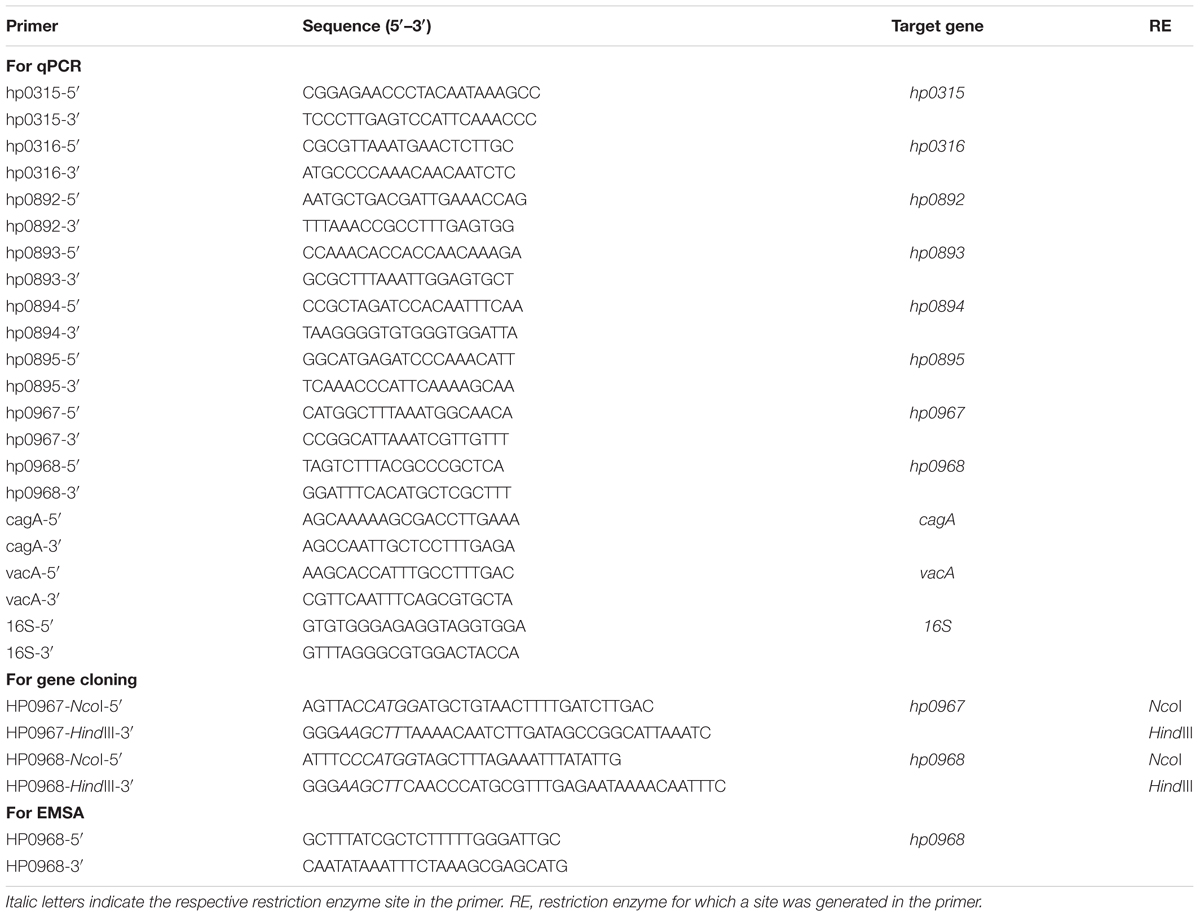
TABLE 1. Primers used in this study.
Electrophoretic Mobility Shift Assay (EMSA)
Electrophoretic mobility shift assay (EMSA) experiments were performed as previously described (De la Cruz et al., 2007, 2013). Specific primers (Table 1) were used to amplify the hp0968–hp0967 promoter region by PCR. The PCR product (100 ng) was mixed with increasing concentrations of HP0968-Myc-His6 at room temperature in the presence of the binding buffer 10X (400 μM HEPES, 80 μM MgCl2, 500 μM KCl, 10 μM DTT, 0.5% NP40, and 1 μg/ml BSA). The reactions were incubated during 30 min at room temperature and then separated in 6% SDS-PAGE gels in Tris-Borate-EDTA buffer. The DNA bands were visualized with ethidium bromide staining.
Ribonuclease Activity Assay
HP0967-Myc-His6 protein (0, 0.6, and 1.2 μM, respectively) was incubated with 1 μg of total H. pylori RNA in the presence of 50 μM Tris (pH 8.0) and 10 μM MgCl2 in a final reaction volume of 20 μl. When necessary, HP0968-Myc-His6 protein was added to analyze the antagonic effect of the antitoxin on the toxin activity. The reaction was allowed to proceed for 30 min at 25°C, and the samples were then loaded in a bleach agarose gel 2% as previously described (Aranda et al., 2012). RNA was visualized with ethidium bromide staining.
RNA Isolation and Quantitative RT-PCR (qRT-PCR)
Total RNA was extracted from the different culture conditions using the hot phenol method (Jahn et al., 2008). Purification of RNA, and qRT-PCR were performed as previously reported (Ares et al., 2016). Briefly, DNA was removed with TURBO DNA-free (Ambion, Inc.) and the quality of RNA was assessed using a NanoDrop (ND-1000; Thermo Scientific) and a bleach agarose gel 2% as previously described (Aranda et al., 2012). cDNA was prepared using 1 μg of RNA, random hexamer primers (0.2 μg/μl), and M-MulV-RT (20 U/μl, reverse transcriptase of Moloney Murine leukemia Virus; Thermo Scientific). Specific primers were designed with the Primer3Plus software1 and are listed in Table 1. The absence of contaminating DNA was controlled by lack of amplification products after 35 qPCR cycles. Control reactions with no template (water) and minus-Reverse Transcriptase were run with all reactions. 16S rRNA (HPrrnA16S) was used as a reference gene for normalization and the relative gene expression was calculated using the 2-ΔCt method (Livak and Schmittgen, 2001).
TA Gene Expression during H. pylori Infection of AGS Cells
AGS gastric epithelial cells were grown to about 75% confluence in RPMI-1640 medium containing 10% FBS, and washed thrice with PBS before adding fresh RPMI media with 10% FBS. H. pylori 26695 was grown in Brucella broth for 24 h, suspended in RPMI, and added to the AGS cell culture at a multiplicity of infection (MOI) of 100. Infected cells were incubated at 37°C under microaerophilic conditions for 0 and 6 h, and bacteria were recovered. RNA was extracted from bacteria to determine gene expression. Fold-change in gene transcription was determined by calculating the relative expression of TA genes with respect to bacteria at time 0 of infection.
Biofilm Formation Assay on Abiotic Surface and TA Gene Expression
Adhesion to abiotic surface (polystyrene) was analyzed using 96-well plates as previously described (Ares et al., 2016). H. pylori 26695 was grown for 2 days on blood agar culture plates containing 10% defibrinated sheep blood at 37°C under microaerophilic conditions, and a bacterial suspension was prepared in Brucella broth supplemented with 10% decomplemented FBS and adjusted to an optical density of 0.1 at 600 nm. Two-hundred μl of the bacterial suspensions were added per well into 96-well plates in octuplicate and incubated at different times at 37°C under microaerophilic conditions. Unbound bacteria were removed by washing the wells three times with PBS, and bound bacteria were stained with 1% Crystal Violet for 20 min. Wells were thoroughly rinsed thrice with PBS and the dye was solubilized in 100 μl of ethanol 70%. Finally, the amount of extracted Crystal Violet was determined by measuring color absorbance at OD600 using a spectrophotometer plate reader (Thermo Scientific).
For determination of TA gene expression, the biofilm assay was performed in 6 well plates (3 ml) and supernatant (planktonic) and adhered (sessile) bacteria were recovered for RNA extraction. Fold-change in gene transcription was determined by calculating the relative expression of TA genes within biofilms (sessile bacteria) as compared to the planktonic bacteria.
Transcription of TA Genes in the Presence of Antibiotics
Helicobacter pylori was grown in Brucella broth supplemented with 10% decomplemented FBS at 37°C for 48 h (stationary phase), with gentle shaking under microaerophilic conditions. The antibiotics ampicillin (Ap, 100 μg/mL), kanamycin (Km, 50 μg/mL), chloramphenicol (Cm, 30 μg/mL), or tetracycline (Tc, 10 μg/mL) were added to the liquid culture and incubated for 1 h as previously described (Christensen-Dalsgaard et al., 2010). Antibiotics concentrations were used as reported for E. coli and Salmonella enterica (Christensen-Dalsgaard et al., 2010; Maisonneuve et al., 2011; Silva-Herzog et al., 2015; Li et al., 2016). Fold-change in gene transcription was determined by calculating the relative expression of TA genes in the presence of each antibiotic as compared to bacteria growing in Brucella broth without antibiotics.
In silico Identification of TA Modules
Analyzed sequence data and loci annotations of 260 H. pylori genomes were retrieved from the NCBI database2 by a series of custom Perl scripts. In addition, the genomes of 57 Helicobacter non-pylori strains were included in the comparative analysis.
We also selected the sequences from known toxin and antitoxin modules that were classified into super-families as stated previously (Leplae et al., 2011) and compared their homology with previously referred Helicobacter genomes. Briefly, each TA protein pair was queried using PSI-BLAST (Altschul et al., 1997) under the following parameters: matrix = BLOSUM62, word size = 3, PSI-BLAST threshold = 0.005, expect threshold = 10, and without filtering low complexity regions. Hits were carefully examined and selected according to their functional annotation.
Furthermore, we employed the Toxin–Antitoxin Data Base (TADB3) (Shao et al., 2011) to extend our findings.
Heatmap Construction
In order to illustrate the presence of every toxin and antitoxin module in all Helicobacter genomes, a gene content matrix (“heatmap” function) was built using the R program4 v3.2.4. Next, these paired loci were hierarchical clustered (“hclust” function, “ward.D” method) according to their loci-content similarity to build a sidelong dendogram.
Results
The hp0968–hp0967 H. pylori Genes Code for a New Type II TA System
Three pairs of type II TA proteins have been described in H. pylori: HP0315–HP0316, HP0892–HP0893, and HP0894–HP0895 (Han et al., 2011, 2013; Kwon et al., 2012; Im et al., 2014). Searching for TA-encoding homolog genes of different families, we identified hp0967, a gene that codes for a type II toxin belonging to the VapD protein family. Of note, an intergenic region was identified between hp0967 and hp0969 genes and was designated herein as antitoxin gene hp0968. In contrast, by analyzing the genome sequence of H. pylori J99 strain, a homolog of hp0968 (jhp0902) antitoxin gene was found. The analysis of the H. pylori 26695s genome revealed a stop codon in the coding region of hp0968 gene resulting in a peptide of 19 aminoacids. However, by comparing the nucleotidic sequences of all H. pylori strains deposited in the GenBank, no stop codon was found in this gene, suggesting a problem of annotation in the strain 26695. To demonstrate that hp0968 antitoxin gene is present and transcribed in 26695 strain, we performed RT-PCR experiments using primers that hybridize with TA genes as shown in Figure 1A. We found that both hp0968 and hp0967 genes were transcriptionally expressed and as expected, they presented a single mRNA similar to other type II TA systems (Figure 1B). The analyses of the internal controls for hp0316–hp0315, hp0893–hp0892, and hp0895–hp0894 TA genes also showed that they are genetically organized as bicistronic operons (Figure 1B).
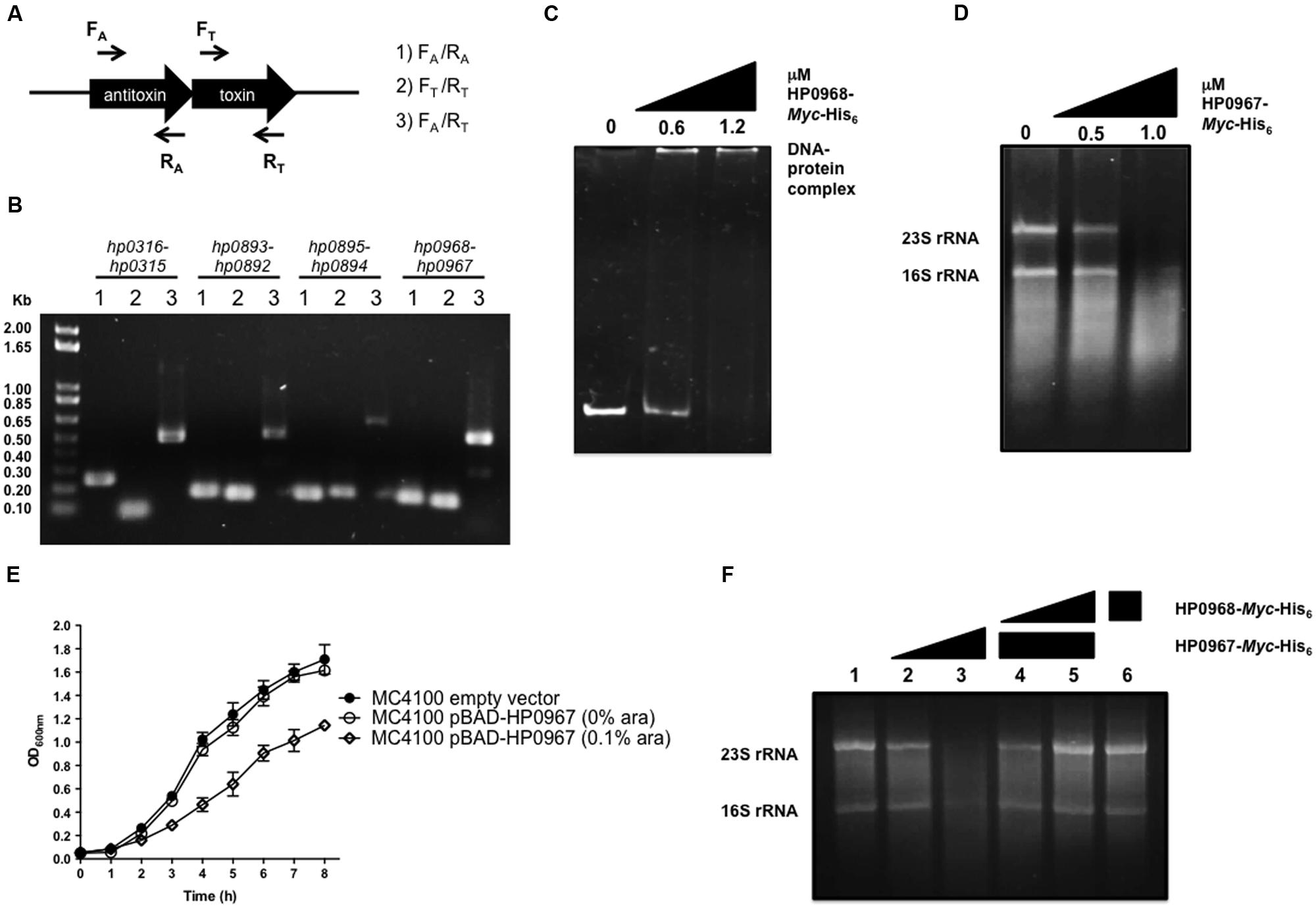
FIGURE 1. HP0967–HP0968 is a bona fide type II TA system. (A) Schematic representation of the type II TA operons amplified for RT-PCR analysis using specific primers for toxin and antitoxin genes (arrows). (B) Qualitative RT-PCR assays carried out with RNA extracted from Helicobacter pylori 26695 grown in Brucella broth during 48 h. This is a representative result of two independent experiments. (C) EMSA exemplifying the binding of HP0968 Myc-His6 to hp0968–hp0967 promoter region. DNA-protein complex is indicated in the figure. (D) Ribonuclease assay using amounts of HP0967-Myc-His6 toxin with 1 μg total RNA. 23S rRNA and 16S rRNA are shown in the figure. (E) Growth kinetic of wild-type Escherichia coli (MC4100) harboring the empty vector pBAD-Myc-HisA and the pBAD-HP0967-Myc-His6 plasmid without and with induction of L-arabinose (ara). Bacterial cultures were grown for 8 h in LB medium at 37°C. (F) Ribonuclease inhibition assay using HP0967-Myc-His6 toxin in presence of HP0968-Myc-His6 antitoxin with 1 μg of total RNA. Alone RNA (lane 1); RNA with 0.5 (lane 2) and 1.0 μM (lane 3) of HP0967-Myc-His6 toxin; RNA with 1.0 μM of HP0967-Myc-His6 toxin and in presence of 0.6 (lane 4) and 1.2 μM (lane 5) of HP0968-Myc-His6 antitoxin; RNA with 1.2 μM of HP0968-Myc-His6 antitoxin (lane 6).
We also amplified the coding region of both hp0967 and hp0968 genes of H. pylori 26695 and cloned them in a pBAD-Myc/HisA vector in order to overexpress and purify the proteins. The sequence analysis of the amplified hp0968 gene revealed that it did not present the stop codon, and that it encodes a functional protein of 93 aminoacids (12.8 kDa). In order to demonstrate the functionality of this antitoxin, EMSA experiments were performed incubating the promoter region of hp0968–hp0967 with the HP0968-Myc-His6 protein. The DNA-protein complex was observed at 1.2 μM of HP0968-Myc-His6 antitoxin (Figure 1C). In contrast, the aminoacid sequence of HP0967 presented a PIN domain associated with ribonuclease activity. Total RNA of H. pylori 26695 was incubated with 0.5 and 1.0 μM of HP0967-Myc-His6 and RNA stability was determined. The assay showed that HP0967-Myc-His6 (1.0 μM) degraded H. pylori RNA, suggesting that it is indeed a toxin (Figure 1D). In addition, the overexpression of HP0967 toxin affected the growth of E. coli (Figure 1E). To demonstrate the inhibitory effect of HP0968 antitoxin on HP0967 toxin, we added HP0968-Myc-His6 protein to the ribonuclease activity assay. Ribonuclease activity of HP0967-Myc-His6 toxin was inhibited in the presence of HP0968-Myc-His6 antitoxin (Figure 1F). HP0968-Myc-His6 alone protein did not affect RNA stability. Thus, our data strongly suggests that both HP0967 and HP0968 proteins constitute a new type II TA system of the Vap family in H. pylori.
Environmental Cues Differentially Regulate Expression of H. pylori Type II TA Systems
In order to elucidate the environmental cues that may affect expression of type II TA systems we performed qRT-PCR experiments. Relative expression was determined during both exponential (24 h) and stationary phase (48 h). Interestingly, expression of all the type II TA genes was higher during stationary phase than in exponential phase (Figure 2A); although hp0316–hp0315 and hp0968–hp0967 showed the highest transcriptional levels (Figure 2A), and hp0893–hp0892 and hp0895–hp0894 were poorly expressed in this growth condition. Similarly, expression of cagA and vacA was found increased during stationary growth phase (Figure 2B), in agreement to previously reported data (Thompson et al., 2003; Boonjakuakul et al., 2005).
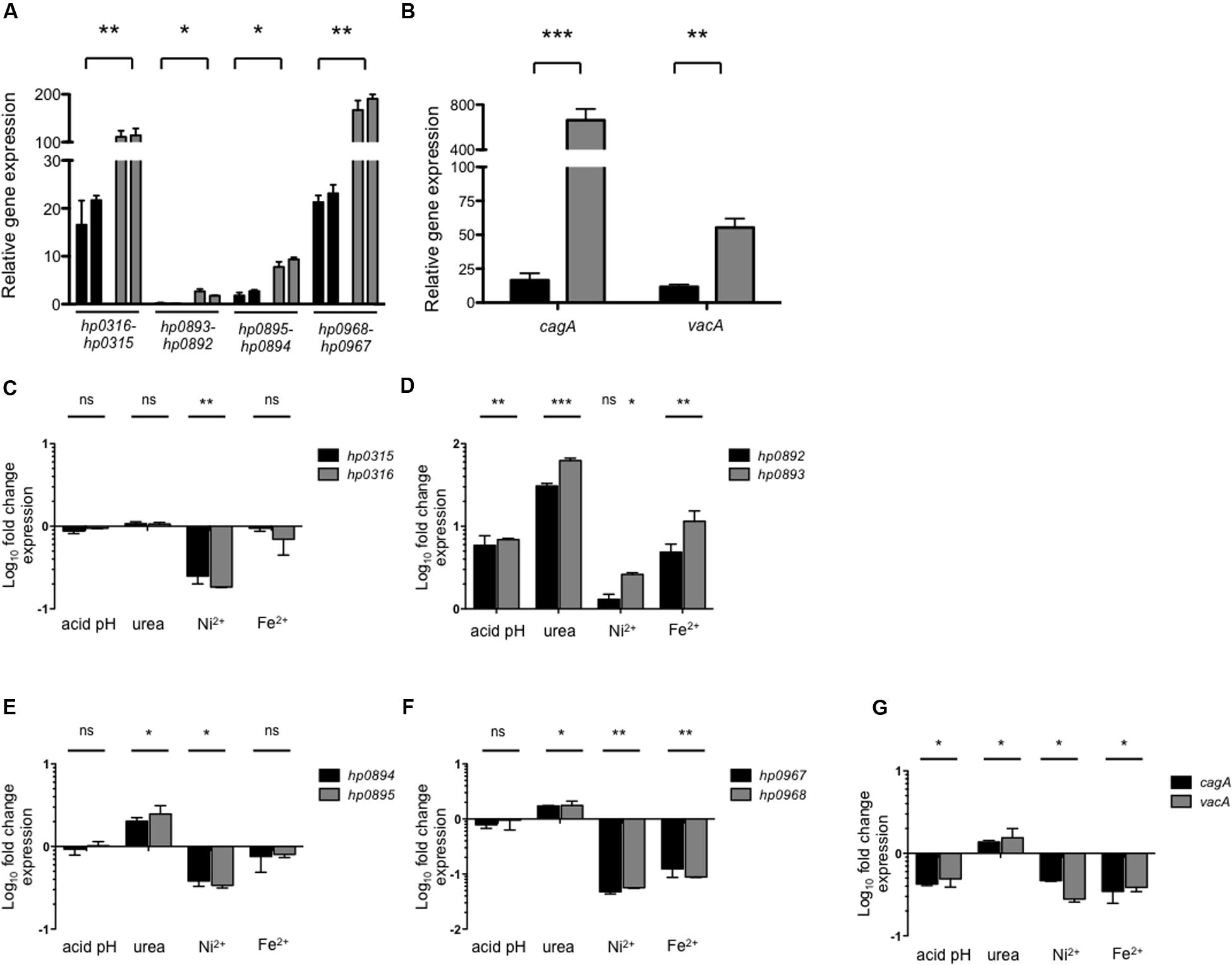
FIGURE 2. Environmental cues and expression of type II TA genes. Expression (qRT-PCR) of the type II TA genes (A) and cagA/vacA (B) of H. pylori 26695 in exponential (black bars) and stationary phase (gray bars). Fold-change expression (qRT-PCR) of the type II TA genes (C–F) and cagA/vacA (G) under different environmental conditions with respect to Brucella broth at 48 h. Data represent the mean of at least three independent experiments (mean ± SD). ns, not significant; statistically significant ∗∗∗p < 0.001; ∗∗p < 0.01; ∗p < 0.05.
Of note, the effect of the environmental cues such as pH, urea, nickel, and iron in transcription of type II TA systems has not been reported for H. pylori, although, they are known to affect the expression of other virulence factors (Sachs et al., 2003; Haley and Gaddy, 2015; Keilberg and Ottemann, 2016). We found that during stationary growth phase type II TA genes showed different levels of expression in the presence of the tested cues. While nickel repressed the expression of hp0316–hp0315 (fivefold) and hp0895–hp0894 (threefold) (Figures 2C,E), all conditions tested triggered expression of hp0893–hp0892 TA genes (Figure 2D). Expression of hp0968–hp0967 was repressed in the presence of nickel (20-fold) and iron (10-fold), while urea caused a twofold increase in its expression (Figure 2F). As reaction controls, the transcription of cagA and vacA genes was determined under the same growth conditions. We found that transcription of both virulence genes was differentially regulated: urea slightly boosted transcription (1.5-fold), while acidic pH (2.2-fold), nickel (2.7-fold), and iron (2.7-fold) repressed them (Figure 2G).
Biofilm Formation and Contact with Gastric Epithelial Cells Stimulate the Expression of hp0968–hp0967
Type II TA systems are reported to influence biofilm formation (Wang and Wood, 2011; Wen et al., 2014; Kedzierska and Hayes, 2016). We quantitatively analyzed the biofilm formation of H. pylori on a polystyrene surface at different time points measuring uptake of Crystal Violet. After 72 h of incubation, a mature biofilm was observed (Figure 3A). Thus, in order to evaluate the expression of type II TA genes in a mature biofilm, we determined their expression in both H. pylori planktonic and sessile cells at 72 h. Expression of all type II TA genes was found to be induced in biofilms-associated H. pylori, as compared to planktonic bacteria (Figure 3B). It should be noted that hp0968–hp0967 showed the highest induction in this environment (500-fold). In a mature biofilm, vacA transcription was significantly increased (23-fold), whereas cagA expression was not affected (Figure 3B). On the other hand, the contact with AGS cells significantly increased transcription of type II TA genes. Of note, hp0968–hp0967 genes presented the highest levels of expression over the other TA systems, even higher than expression of cagA and vacA (Figure 3C). These data strongly suggest that hp0968–hp0967 genes might be expressed in the host during stomach colonization.
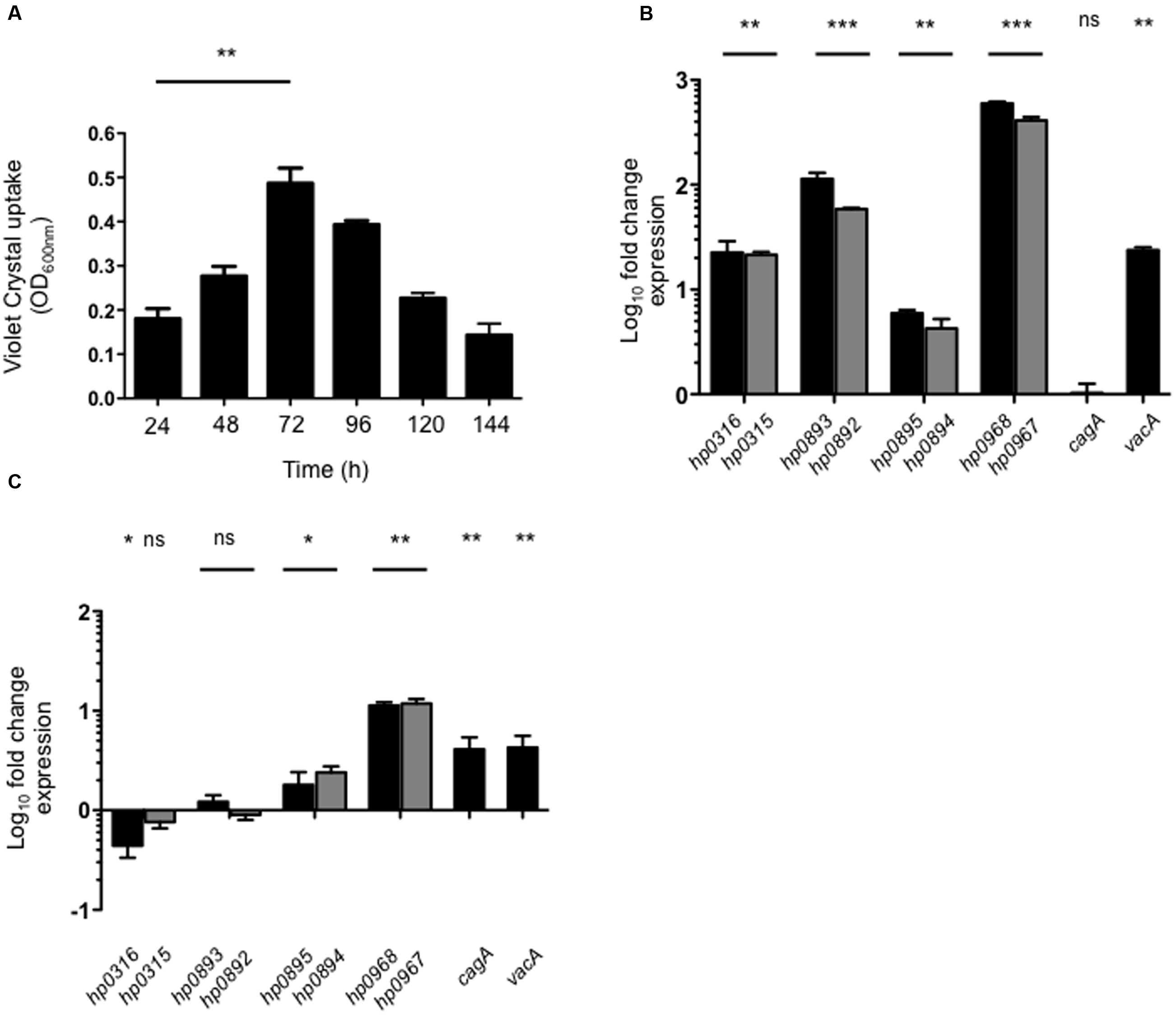
FIGURE 3. Expression of type II TA genes in biofilm formation and in AGS cells. (A) Quantification of biofilm formation of H. pylori 26695 by measuring Crystal Violet uptake at different times. Fold-change expression (qRT-PCR) of the type II TA genes and cagA/vacA in a mature biofilm after 72 h (B) and in contact with AGS cells (C). Data represent the mean of at least three independent experiments (mean ± SD). ns, not significant; statistically significant ∗∗∗p < 0.001; ∗∗p < 0.01; ∗p < 0.05.
The Presence of Antibiotics Affects the Transcription of H. pylori Type II TA Genes
We analyzed the expression of type II TA genes in the presence of antibiotics, and found that kanamycin and chloramphenicol (Km/Cm) increased the expression of hp0316–hp0315 (6.2/9.9-fold), hp0893–hp0892 (3.1/9.2-fold), and hp0895–hp0894 (4.34/9-fold) systems (Figure 4). In particular, chloramphenicol, kanamycin, and tetracycline stimulated the expression of hp0968–hp0967 genes (50-, 12-, and 2.5-fold, respectively) while ampicillin repressed them (2.8-fold) (Figure 4D). Interestingly, similar transcriptional profiling was found between the hp0893–hp0892 and hp0968–hp0967 genes in presence of chloramphenicol, kanamycin, and tetracycline (Figures 4B,D). Concerning the effect of antibiotics on the expression of cytotoxin genes, only vacA expression was boosted in the presence of kanamycin (10-fold) and chloramphenicol (23-fold) (Figure 4E). These results suggest that type II TA genes may have an important role in the response of H. pylori to the presence of antibiotics by H. pylori, particularly to chloramphenicol and kanamycin.
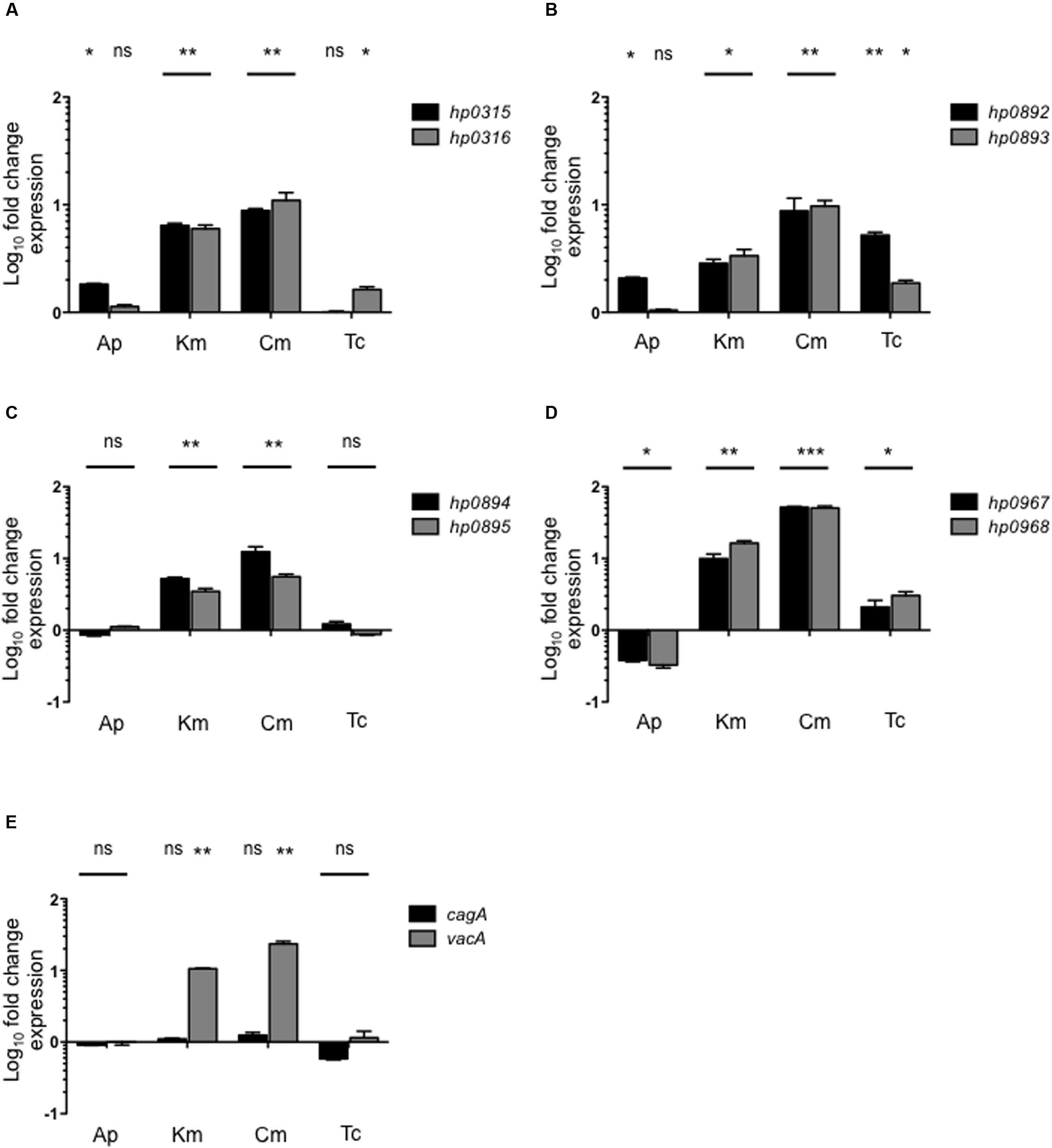
FIGURE 4. Effect of antibiotics on type II TA expression. Fold-change expression (qRT-PCR) of the type II TA genes (A–D) and cagA/vacA (E) in presence of different antibiotics concentrations such as: Ap (Ampicillin, 100 μg/ml), Km (Kanamycin, 30 μg/ml), Cm (Chloramphenicol, 30 μg/ml), and Tc (Tetracycline, 10 μg/ml) by 1 h. Data represent the mean of at least three independent experiments (mean ± SD). ns, not significant; statistically significant ∗∗∗p < 0.001; ∗∗p < 0.01; ∗p < 0.05.
The hp0968–hp0967 Genes Are Highly Prevalent among Helicobacter Strains
We performed PCR experiments to identify type II TA genes in a collection of 114 clinical H. pylori isolates. These bacteria were isolated from patients with diverse pathologies, such as non-atrophic gastritis, premalignant lesions, gastric ulcer, and gastric cancer. Whereas hp0968–hp0967 was the most prevalent type II TA system in these isolates (in over 70% of all isolates), hp0895–hp0894 genes were the least prevalent (in less than 50%) (Figure 5A). In addition, a Blast search in sequences deposited in GenBank5 using sequences of different TA families (included Vap), revealed the presence of type II TA genes in other H. pylori strains. TA genes of the Vap family were highly prevalent, presenting high identity and constituting a genetic signature inside H. pylori species (Figure 5B). In agreement with our findings in the clinical isolates, the hp0968–hp0967 genes presented the highest prevalence as compared to the other type II TA genes (Figures 5A–C). Interestingly, in both groups of H. pylori clinical and sequenced strains, the hp0968 antitoxin gene was more conserved than hp0967 toxin gene (Figures 5A–C).
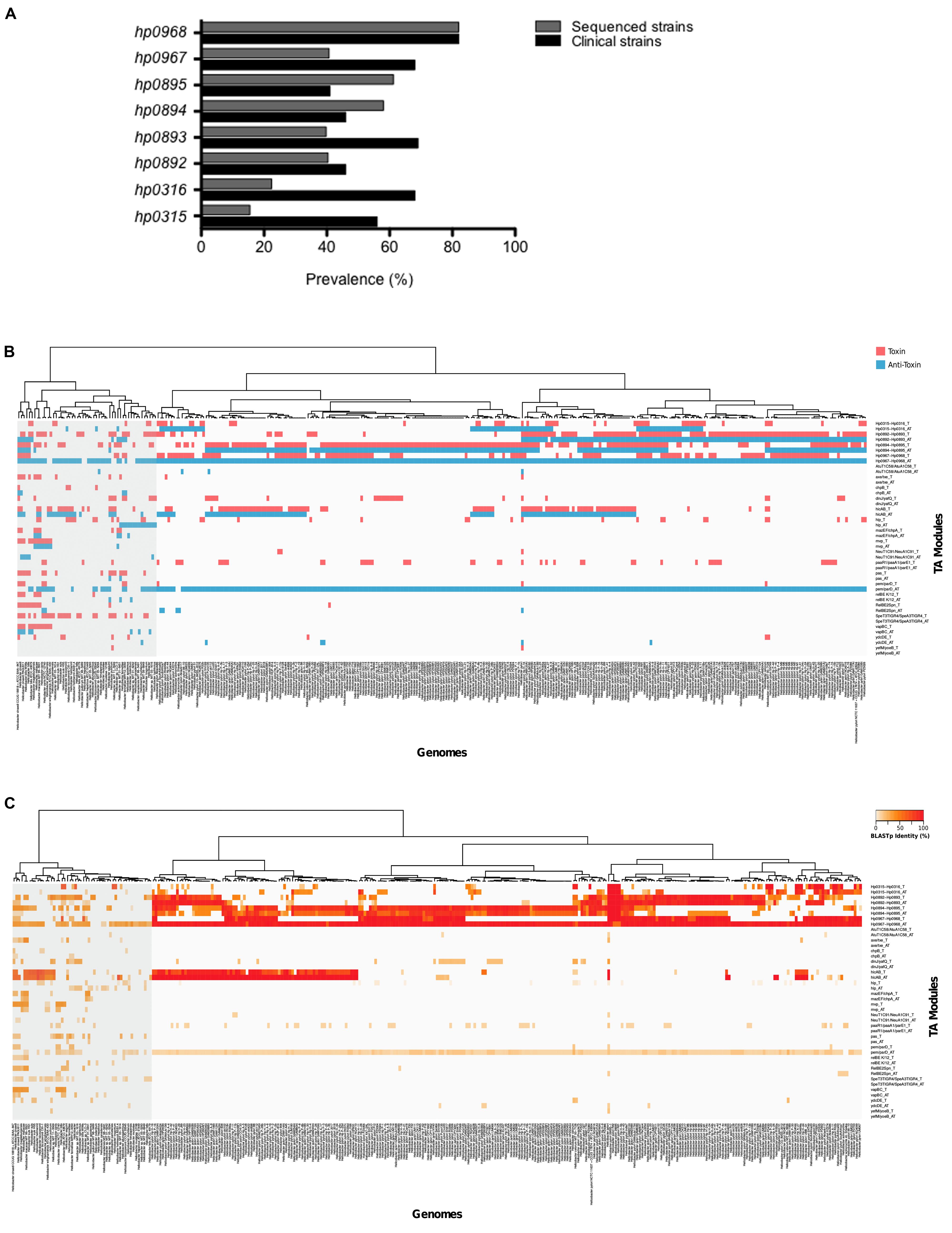
FIGURE 5. Prevalence of type II TA genes. (A) The clinical isolates tested (n = 114) comprised strains of H. pylori with different pathologies. Bars show the prevalence of type II TA genes in clinic (black) and sequenced (gray) strains. The heatmaps and hierarchical clustering of selected genomes for H. pylori and Helicobacter species showing both the presence (B) and identity (C) of toxin–antitoxin genes were determined in R (version 3.2.4) using the hclust function with the “ward.D” method. The red and blue colors indicate the presence of the toxin and the anti-toxin gene.
Since the number of type II TA systems has been associated to bacterial pathogenicity (Georgiades and Raoult, 2011; De la Cruz et al., 2013), we studied the presence of the type II TA systems of different families in other Helicobacter species, and found that the presence of type II TA genes varied from one species to another. Species phylogenetically related to H. pylori such as H. acinonychis (big cats) and H. cetorum (marin mammals), contain the four type II TA systems (hp0316–hp0315, hp0893–hp0892, hp0895–hp0894, and hp0968–hp0967) and show high identity to the H. pylori sequences (Figures 5B,C). The presence of TA genes marked a separation of H. acinonychis and H. cetorum from the rest of Helicobacter species and they were grouped together with the H. pylori strains.
Discussion
Helicobacter pylori produces a number of virulence-associated proteins such as secretion systems, flagella, adhesins, and cytotoxins that enable successful colonization of its human host. Expression of virulence factors also allows for the survival and persistence of H. pylori in the hostile environment found in the stomach (Salama et al., 2013). This persistence is mediated by a myriad of bacterial strategies to evade the immune response, such as detoxifying enzymes for reactive species of oxygen and nitrogen, DNA repair pathways, and natural competence (Wang et al., 2006; Dorer et al., 2011, 2013). In this sense, type II TA systems have emerged as potential virulence factors, not only affecting pathogenicity, but they are also involved in biofilm formation and persistence (Gerdes and Maisonneuve, 2012; Wen et al., 2014; Kedzierska and Hayes, 2016). In this work, we describe the transcriptional profiling of type II TA genes present in the genome of H. pylori 26695: hp0316–hp0315, hp0893–hp0892, hp0895–hp0894, and hp0968–hp0967, since the hp0968–hp0967 system had not been identified and it was characterized in this work. These genes are transcriptionally organized as bicistronic operons, which is a hallmark of the type II TA systems. Type II toxins present different biochemical activities such as ribonucleases, kinases, adenylyl- and acetyl-transferases, and act also as inhibitors of protein synthesis and DNA supercoiling (Jiang et al., 2002; Mutschler et al., 2011; Yamaguchi et al., 2011; Harms et al., 2015; Cheverton et al., 2016; Rocker and Meinhart, 2016). In contrast, type II antitoxins are DNA-binding proteins that regulate their own expression and directly interact with their cognate toxins (Yamaguchi and Inouye, 2011; Chan et al., 2016). Despite having more specific functions, antitoxins show more complexity by possessing two functional domains: a DNA-binding domain and a toxin-interaction domain. Our characterization of both H. pylori HP0967 and HP0968 proteins revealed that they display ribonuclease and DNA-binding activities, respectively. HP0967 and HP0968 proteins belong to the Vap family, which is the type II TA pair most widespread in Bacteria and Archaea (Pandey and Gerdes, 2005; Sevin and Barloy-Hubler, 2007; Arcus et al., 2011; Jin et al., 2015).
In the stomach, H. pylori is exposed to frequent changes in pH, for example to acidic pH when present in the lumen and to neutral pH as it crosses the mucus to interact with epithelial cells. Presumably, these changes alter the expression of genes related to colonization and persistence in its human host. We found that an acidic pH boosted the expression of hp0893–hp0892, while no effect was observed for the other type II TA systems, highlighting the importance of this system in the response to the acidic pH, which H. pylori frequently finds prior to colonization of the stomach mucosa. Previous reports regarding the effect of acidic pH on the expression of the most thoroughly studied virulence factors cagA and vacA have been controversial (Karita et al., 1996; Allan et al., 2001; Merrell et al., 2003a; Bury-Mone et al., 2004; Vannini et al., 2014). Our results support the observation that an acidic pH represses both cagA and vacA, in agreement to the notion that CagA and VacA are produced when H. pylori is in close contact to epithelial cells, where the pH is neutral (Salama et al., 2013). Urea, nickel, and iron are components present in the human stomach and it has been suggested that they act as regulatory networks whose purpose is the survival and persistence of H. pylori (Haley and Gaddy, 2015). Urea and nickel are interconnected cues because transcription/activity of urease is nickel-dependent (Contreras et al., 2003; Khan et al., 2009). While high concentrations of nickel repress virulence genes (Haley and Gaddy, 2015), the presence of urea is essential for the survival and colonization of H. pylori (Nakazawa, 2002; Sachs et al., 2003). We found that urea strongly stimulated the expression of hp0893–hp0892, while it had a mild effect in the expression of hp0895–hp0894 and hp0968–hp0967. With the exception of hp0893–hp0892, nickel and iron repressed the type II TA systems in H. pylori. Our results suggest that the high concentration of urea and low concentrations of both nickel and iron present in the stomach, would induce transcription of type II TA systems in preparation for successful colonization and persistent infection. Our study represents the first report on the transcriptional analysis of type II TA genes in H. pylori under several environmental conditions that mimic those prevailing in the gastric mucosa.
Contrasting data have been reported concerning metal regulation of vacA and cagA (Merrell et al., 2003b; Ernst et al., 2005; Pich et al., 2012; Noto et al., 2013; Vannini et al., 2014). Iron exposure is associated with repression of numerous factors involved in the repertoire of virulence in H. pylori (Haley and Gaddy, 2015). The role of nickel on the expression of vacA and cagA genes has not been previously studied. Transcriptional data showed that both, nickel and iron repressed both cagA and vacA genes, supporting the notion that the presence of both metals affect the virulence of H. pylori.
Biofilm formation is a bacterial strategy to counteract the stress caused by the limitation of nutrients and the effect of antibiotics. It is well-established that type II TA systems are required for the biofilm formation (Wang and Wood, 2011; Wen et al., 2014; Kedzierska and Hayes, 2016). Since environmental and clinical strains of H. pylori produce biofilm to persist and colonize both abiotic and biotic surfaces (Cammarota et al., 2012; Percival and Suleman, 2014), we studied the expression of the four type II TA genes in H. pylori growing in a mature biofilm. In this condition, the four type II TA systems were induced, supporting the notion that these systems are expressed and may be required for the biofilm formation.
In contact with AGS gastric epithelial cells, transcription of hp0968–hp0967 was considerably stimulated, presenting values higher than cagA and vacA cytotoxin genes. These results would support a probable role of hp0968–hp0967 genes in the context of infection of H. pylori.
Kenn Gerdes’s lab has clearly shown that antibiotics such as chloramphenicol, positively affect the expression of TA genes (Christensen et al., 2001; Christensen-Dalsgaard and Gerdes, 2006; Jorgensen et al., 2009; Winther and Gerdes, 2009; Christensen-Dalsgaard et al., 2010). We found that the presence of both chloramphenicol and kanamycin boosted transcription of all type II TA systems in H. pylori. In the presence of both kanamycin and chloramphenicol, bacterial translation is inhibited and proteases such as Lon specifically degrades the type II antitoxins, generating an increase in the transcription of TA genes due to the lack of repression of the antitoxins on their own expressions (Christensen et al., 2001; Winther and Gerdes, 2009; Christensen-Dalsgaard et al., 2010). We found that the presence of both kanamycin and chloramphenicol, increased expression of type II TA genes, suggesting a probable role of type II TA systems in the bacterial fitness during biofilm formation and in the presence of antibiotics. In agreement, recent reports show that type II TA modules are necessary for the development of persister cells in the presence of antibiotics (Maisonneuve et al., 2011; Wen et al., 2014; Kedzierska and Hayes, 2016). Current experiments in our group are aimed to further analyze the expression of type II TA systems in persister H. pylori.
In most reports, expression of cagA and vacA has been found to be co-regulated, corroborating the concept that both cytotoxins are required and co-expressed in the context of infection (Yokoyama et al., 2005; Singh et al., 2012; Raghwan and Chowdhury, 2014). However, we found that in a mature biofilm and in the presence of kanamycin or chloramphenicol, transcription of vacA, but not cagA was significantly increased. In contrast to CagA which is translocated into host cells by the type 4 secretion system, VacA is secreted by a type 5 secretion system (autotransporter) and it can also be located on the bacterial surface (Ilver et al., 2004). Based on the above notions, we suggest that VacA may have a role in bacterium-bacterium interactions, as it occurs during biofilm formation.
In terms of prevalence, hp0968–hp0967 system was the most prevalent in both clinical and sequenced H. pylori strains. Moreover, the hp0968–hp0967 system was also widely distributed in all Helicobacter species. It was recently reported the retention of the antitoxin gene but not the toxin gene in TA modules of Xanthomonas genomes (Martins et al., 2016). Both clinical and sequenced H. pylori strains revealed the same profile, loss of toxin and retention of antitoxin genes. In H. pylori and non-pylori genomes deposited in GenBank, hp0968 antitoxin gene was much more prevalent than hp0967 toxin. The retention of antitoxin genes in bacterial genomes has been postulated due to the toxin-independent functions of type II antitoxins such as MqsA and DinJ (Wang and Wood, 2011; Hu et al., 2012).
The presence and homology of type II TA genes in Helicobacter strains generated two main clusters: H. pylori and other Helicobacter species, showing that these TA systems represent a DNA signature for H. pylori. Moreover, species phylogenetically related to H. pylori such as H. acinonychis and H. cetorum were separated from the rest of Helicobacter species and grouped with H. pylori strains, supporting the concept that TA genes are part of the prokaryotic mobilome and that they are involved in the bacterial evolution (Koonin and Wolf, 2008). Searching type II TA genes by using Toxin–Antitoxin Data Base (TADB3) we found two TA systems for the H. pylori strains 26695 and J99 (HP0892–HP0893 and HP0894–HP0895). In the case of other Helicobacter species, H. hepaticus was the only species showing one TA system (HH0272–HH0273), which clearly indicates the need for an update and construction of a new database that integrates more bacterial genomes.
It is worth noticing that hp0968–hp0967 genes presented the same transcriptional profile than cagA and vacA under growth in urea, nickel, iron and in contact with AGS cells. These findings together with the high prevalence of this TA system in H. pylori strains, suggest that the hp0968–hp0967 may represent a very important virulence factor. We cannot discard a probable virulence role for the other type II TA system, since these genes presumably display redundant and specific functions. The generation of null mutants will help to elucidate the role of type II TA genes in different phenotypes such as biofilm formation, persistence and mainly in the virulence of H. pylori.
Author Contributions
Conceived and designed the experiments: MDC. Performed the experiments: MC-M, MA, LP, SP. Analyzed the data: MDC, MC-M, MA, MC-P, JG, JT. Wrote the paper: MDC, JG, JT.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was supported by grant FIS/IMSS/PROT/G14/1329 (to MC-M) from the Fondo de Investigación en Salud (FIS)-IMSS, México. We thank Areli Altamira-Velasco, Mariana Chávez-Montaño, Diana Márquez-Delfín, and Mariana Ramírez-Etienne for technical assistance.
Footnotes
- ^http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi/
- ^ftp://ftp.ncbi.nlm.nih.gov/genomes/genbank/bacteria/
- ^http://202.120.12.135/TADB2/index.php
- ^http://www.R-project.org/
- ^http://www.ncbi.nlm.nih.gov/genbank/
References
Allan, E., Clayton, C. L., Mclaren, A., Wallace, D. M., and Wren, B. W. (2001). Characterization of the low-pH responses of Helicobacter pylori using genomic DNA arrays. Microbiology 147, 2285–2292. doi: 10.1099/00221287-147-8-2285
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Amieva, M. R., and El-Omar, E. M. (2008). Host-bacterial interactions in Helicobacter pylori infection. Gastroenterology 134, 306–323. doi: 10.1053/j.gastro.2007.11.009
Aranda, P. S., Lajoie, D. M., and Jorcyk, C. L. (2012). Bleach gel: a simple agarose gel for analyzing RNA quality. Electrophoresis 33, 366–369. doi: 10.1002/elps.201100335
Arcus, V. L., Mckenzie, J. L., Robson, J., and Cook, G. M. (2011). The PIN-domain ribonucleases and the prokaryotic VapBC toxin-antitoxin array. Protein Eng. Des. Sel. 24, 33–40. doi: 10.1093/protein/gzq081
Ares, M. A., Fernandez-Vazquez, J. L., Rosales-Reyes, R., Jarillo-Quijada, M. D., Von Bargen, K., Torres, J., et al. (2016). H-NS nucleoid protein controls virulence features of Klebsiella pneumoniae by regulating the expression of type 3 pili and the capsule polysaccharide. Front. Cell. Infect. Microbiol. 6:13. doi: 10.3389/fcimb.2016.00013
Audoly, G., Vincentelli, R., Edouard, S., Georgiades, K., Mediannikov, O., Gimenez, G., et al. (2011). Effect of rickettsial toxin VapC on its eukaryotic host. PLoS ONE 6:e26528. doi: 10.1371/journal.pone.0026528
Boonjakuakul, J. K., Canfield, D. R., and Solnick, J. V. (2005). Comparison of Helicobacter pylori virulence gene expression in vitro and in the Rhesus macaque. Infect. Immun. 73, 4895–4904. doi: 10.1128/IAI.73.8.4895-4904.2005
Bury-Mone, S., Thiberge, J. M., Contreras, M., Maitournam, A., Labigne, A., and De Reuse, H. (2004). Responsiveness to acidity via metal ion regulators mediates virulence in the gastric pathogen Helicobacter pylori. Mol. Microbiol. 53, 623–638. doi: 10.1111/j.1365-2958.2004.04137.x
Cammarota, G., Sanguinetti, M., Gallo, A., and Posteraro, B. (2012). Review article: biofilm formation by Helicobacter pylori as a target for eradication of resistant infection. Aliment. Pharmacol. Ther. 36, 222–230. doi: 10.1111/j.1365-2036.2012.05165.x
Chan, W. T., Espinosa, M., and Yeo, C. C. (2016). Keeping the wolves at Bay: antitoxins of prokaryotic type II toxin-antitoxin systems. Front. Mol. Biosci. 3:9. doi: 10.3389/fmolb.2016.00009
Cheverton, A. M., Gollan, B., Przydacz, M., Wong, C. T., Mylona, A., Hare, S. A., et al. (2016). A Salmonella toxin promotes persister formation through acetylation of tRNA. Mol. Cell 63, 86–96. doi: 10.1016/j.molcel.2016.05.002
Christensen, S. K., Mikkelsen, M., Pedersen, K., and Gerdes, K. (2001). RelE, a global inhibitor of translation, is activated during nutritional stress. Proc. Natl. Acad. Sci. U.S.A. 98, 14328–14333. doi: 10.1073/pnas.251327898
Christensen-Dalsgaard, M., and Gerdes, K. (2006). Two higBA loci in the Vibrio cholerae superintegron encode mRNA cleaving enzymes and can stabilize plasmids. Mol. Microbiol. 62, 397–411. doi: 10.1111/j.1365-2958.2006.05385.x
Christensen-Dalsgaard, M., Jorgensen, M. G., and Gerdes, K. (2010). Three new RelE-homologous mRNA interferases of Escherichia coli differentially induced by environmental stresses. Mol. Microbiol. 75, 333–348. doi: 10.1111/j.1365-2958.2009.06969.x
Contreras, M., Thiberge, J. M., Mandrand-Berthelot, M. A., and Labigne, A. (2003). Characterization of the roles of NikR, a nickel-responsive pleiotropic autoregulator of Helicobacter pylori. Mol. Microbiol. 49, 947–963. doi: 10.1046/j.1365-2958.2003.03621.x
De la Cruz, M. A., Fernandez-Mora, M., Guadarrama, C., Flores-Valdez, M. A., Bustamante, V. H., Vazquez, A., et al. (2007). LeuO antagonizes H-NS and StpA-dependent repression in Salmonella enterica ompS1. Mol. Microbiol. 66, 727–743. doi: 10.1111/j.1365-2958.2007.05958.x
De la Cruz, M. A., Zhao, W., Farenc, C., Gimenez, G., Raoult, D., Cambillau, C., et al. (2013). A toxin-antitoxin module of Salmonella promotes virulence in mice. PLoS Pathog. 9:e1003827. doi: 10.1371/journal.ppat.1003827
Dorer, M. S., Cohen, I. E., Sessler, T. H., Fero, J., and Salama, N. R. (2013). Natural competence promotes Helicobacter pylori chronic infection. Infect. Immun. 81, 209–215. doi: 10.1128/IAI.01042-12
Dorer, M. S., Sessler, T. H., and Salama, N. R. (2011). Recombination and DNA repair in Helicobacter pylori. Annu. Rev. Microbiol. 65, 329–348. doi: 10.1146/annurev-micro-090110-102931
Ernst, F. D., Bereswill, S., Waidner, B., Stoof, J., Mader, U., Kusters, J. G., et al. (2005). Transcriptional profiling of Helicobacter pylori Fur- and iron-regulated gene expression. Microbiology 151, 533–546. doi: 10.1099/mic.0.27404-0
Georgiades, K., and Raoult, D. (2011). Genomes of the most dangerous epidemic bacteria have a virulence repertoire characterized by fewer genes but more toxin-antitoxin modules. PLoS ONE 6:e17962. doi: 10.1371/journal.pone.0017962
Gerdes, K., and Maisonneuve, E. (2012). Bacterial persistence and toxin-antitoxin loci. Annu. Rev. Microbiol. 66, 103–123. doi: 10.1146/annurev-micro-092611-150159
Haley, K. P., and Gaddy, J. A. (2015). Metalloregulation of Helicobacter pylori physiology and pathogenesis. Front. Microbiol. 6:911. doi: 10.3389/fmicb.2015.00911
Han, K. D., Ahn, D. H., Lee, S. A., Min, Y. H., Kwon, A. R., Ahn, H. C., et al. (2013). Identification of chromosomal HP0892-HP0893 toxin-antitoxin proteins in Helicobacter pylori and structural elucidation of their protein-protein interaction. J. Biol. Chem. 288, 6004–6013. doi: 10.1074/jbc.M111.322784
Han, K. D., Matsuura, A., Ahn, H. C., Kwon, A. R., Min, Y. H., Park, H. J., et al. (2011). Functional identification of toxin-antitoxin molecules from Helicobacter pylori 26695 and structural elucidation of the molecular interactions. J. Biol. Chem. 286, 4842–4853. doi: 10.1074/jbc.M109.097840
Harms, A., Stanger, F. V., Scheu, P. D., De Jong, I. G., Goepfert, A., Glatter, T., et al. (2015). Adenylylation of gyrase and topo IV by FicT toxins disrupts bacterial DNA topology. Cell Rep. 12, 1497–1507. doi: 10.1016/j.celrep.2015.07.056
Hu, Y., Benedik, M. J., and Wood, T. K. (2012). Antitoxin DinJ influences the general stress response through transcript stabilizer CspE. Environ. Microbiol. 14, 669–679. doi: 10.1111/j.1462-2920.2011.02618.x
Ilver, D., Barone, S., Mercati, D., Lupetti, P., and Telford, J. L. (2004). Helicobacter pylori toxin VacA is transferred to host cells via a novel contact-dependent mechanism. Cell. Microbiol. 6, 167–174. doi: 10.1046/j.1462-5822.2003.00349.x
Im, H., Jang, S. B., Pathak, C., Yang, Y. J., Yoon, H. J., Yu, T. K., et al. (2014). Crystal structure of toxin HP0892 from Helicobacter pylori with two Zn(II) at 1.8 A resolution. Protein Sci. 23, 819–832. doi: 10.1002/pro.2465
Jahn, C. E., Charkowski, A. O., and Willis, D. K. (2008). Evaluation of isolation methods and RNA integrity for bacterial RNA quantitation. J. Microbiol. Methods 75, 318–324. doi: 10.1016/j.mimet.2008.07.004
Jiang, Y., Pogliano, J., Helinski, D. R., and Konieczny, I. (2002). ParE toxin encoded by the broad-host-range plasmid RK2 is an inhibitor of Escherichia coli gyrase. Mol. Microbiol. 44, 971–979. doi: 10.1046/j.1365-2958.2002.02921.x
Jin, G., Pavelka, M. S. Jr., and Butler, J. S. (2015). Structure-function analysis of VapB4 antitoxin identifies critical features of a minimal VapC4 toxin-binding module. J. Bacteriol. 197, 1197–1207. doi: 10.1128/JB.02508-14
Jorgensen, M. G., Pandey, D. P., Jaskolska, M., and Gerdes, K. (2009). HicA of Escherichia coli defines a novel family of translation-independent mRNA interferases in bacteria and archaea. J. Bacteriol. 191, 1191–1199. doi: 10.1128/JB.01013-08
Karita, M., Tummuru, M. K., Wirth, H. P., and Blaser, M. J. (1996). Effect of growth phase and acid shock on Helicobacter pylori cagA expression. Infect. Immun. 64, 4501–4507.
Kedzierska, B., and Hayes, F. (2016). Emerging roles of toxin-antitoxin modules in bacterial pathogenesis. Molecules 21:E790. doi: 10.3390/molecules21060790
Keilberg, D., and Ottemann, K. M. (2016). How Helicobacter pylori senses, targets and interacts with the gastric epithelium. Environ. Microbiol. 18, 791–806. doi: 10.1111/1462-2920.13222
Khan, S., Karim, A., and Iqbal, S. (2009). Helicobacter urease: niche construction at the single molecule level. J. Biosci. 34, 503–511. doi: 10.1007/s12038-009-0069-4
Komi, K. K., Ge, Y. M., Xin, X. Y., Ojcius, D. M., Sun, D., Hu, W. L., et al. (2015). ChpK and MazF of the toxin-antitoxin modules are involved in the virulence of Leptospira interrogans during infection. Microbes Infect. 17, 34–47. doi: 10.1016/j.micinf.2014.10.010
Koonin, E. V., and Wolf, Y. I. (2008). Genomics of bacteria and archaea: the emerging dynamic view of the prokaryotic world. Nucleic Acids Res. 36, 6688–6719. doi: 10.1093/nar/gkn668
Kwon, A. R., Kim, J. H., Park, S. J., Lee, K. Y., Min, Y. H., Im, H., et al. (2012). Structural and biochemical characterization of HP0315 from Helicobacter pylori as a VapD protein with an endoribonuclease activity. Nucleic Acids Res. 40, 4216–4228. doi: 10.1093/nar/gkr1305
Leplae, R., Geeraerts, D., Hallez, R., Guglielmini, J., Dreze, P., and Van Melderen, L. (2011). Diversity of bacterial type II toxin-antitoxin systems: a comprehensive search and functional analysis of novel families. Nucleic Acids Res. 39, 5513–5525. doi: 10.1093/nar/gkr131
Lewis, K. (2007). Persister cells, dormancy and infectious disease. Nat. Rev. Microbiol. 5, 48–56. doi: 10.1038/nrmicro1557
Li, T., Yin, N., Liu, H., Pei, J., and Lai, L. (2016). Novel inhibitors of toxin HipA reduce multidrug tolerant persisters. ACS Med. Chem. Lett. 7, 449–453. doi: 10.1021/acsmedchemlett.5b00420
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Maisonneuve, E., Shakespeare, L. J., Jorgensen, M. G., and Gerdes, K. (2011). Bacterial persistence by RNA endonucleases. Proc. Natl. Acad. Sci. U.S.A. 108, 13206–13211. doi: 10.1073/pnas.1100186108
Makarova, K. S., Grishin, N. V., and Koonin, E. V. (2006). The HicAB cassette, a putative novel, RNA-targeting toxin-antitoxin system in archaea and bacteria. Bioinformatics 22, 2581–2584. doi: 10.1093/bioinformatics/btl418
Makarova, K. S., Wolf, Y. I., and Koonin, E. V. (2009). Comprehensive comparative-genomic analysis of type 2 toxin-antitoxin systems and related mobile stress response systems in prokaryotes. Biol. Direct 4:19. doi: 10.1186/1745-6150-4-19
Marshall, B. J., and Warren, J. R. (1984). Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 1, 1311–1315. doi: 10.1016/S0140-6736(84)91816-6
Martins, P. M., Machado, M. A., Silva, N. V., Takita, M. A., and De Souza, A. A. (2016). Type ii toxin-antitoxin distribution and adaptive aspects on Xanthomonas genomes: focus on Xanthomonas citri. Front. Microbiol. 7:652. doi: 10.3389/fmicb.2016.00652
Merrell, D. S., Goodrich, M. L., Otto, G., Tompkins, L. S., and Falkow, S. (2003a). pH-regulated gene expression of the gastric pathogen Helicobacter pylori. Infect. Immun. 71, 3529–3539. doi: 10.1128/IAI.71.6.3529-3539.2003
Merrell, D. S., Thompson, L. J., Kim, C. C., Mitchell, H., Tompkins, L. S., Lee, A., et al. (2003b). Growth phase-dependent response of Helicobacter pylori to iron starvation. Infect. Immun. 71, 6510–6525. doi: 10.1128/IAI.71.11.6510-6525.2003
Mutschler, H., Gebhardt, M., Shoeman, R. L., and Meinhart, A. (2011). A novel mechanism of programmed cell death in bacteria by toxin-antitoxin systems corrupts peptidoglycan synthesis. PLoS Biol. 9:e1001033. doi: 10.1371/journal.pbio.1001033
Nakazawa, T. (2002). Growth cycle of Helicobacter pylori in gastric mucous layer. Keio J. Med. 51(Suppl. 2), 15–19. doi: 10.2302/kjm.51.supplement2_15
Norton, J. P., and Mulvey, M. A. (2012). Toxin-antitoxin systems are important for niche-specific colonization and stress resistance of uropathogenic Escherichia coli. PLoS Pathog. 8:e1002954. doi: 10.1371/journal.ppat.1002954
Noto, J. M., Gaddy, J. A., Lee, J. Y., Piazuelo, M. B., Friedman, D. B., Colvin, D. C., et al. (2013). Iron deficiency accelerates Helicobacter pylori-induced carcinogenesis in rodents and humans. J. Clin. Invest. 123, 479–492. doi: 10.1172/JCI64373
Page, R., and Peti, W. (2016). Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 12, 208–214. doi: 10.1038/nchembio.2044
Palframan, S. L., Kwok, T., and Gabriel, K. (2012). Vacuolating cytotoxin A (VacA), a key toxin for Helicobacter pylori pathogenesis. Front. Cell. Infect. Microbiol. 2:92. doi: 10.3389/fcimb.2012.00092
Pandey, D. P., and Gerdes, K. (2005). Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res. 33, 966–976. doi: 10.1093/nar/gki201
Pathak, C., Im, H., Yang, Y. J., Yoon, H. J., Kim, H. M., Kwon, A. R., et al. (2013). Crystal structure of apo and copper bound HP0894 toxin from Helicobacter pylori 26695 and insight into mRNase activity. Biochim. Biophys. Acta 1834, 2579–2590. doi: 10.1016/j.bbapap.2013.09.006
Percival, S. L., and Suleman, L. (2014). Biofilms and Helicobacter pylori: dissemination and persistence within the environment and host. World J. Gastrointest. Pathophysiol. 5, 122–132.
Pich, O. Q., Carpenter, B. M., Gilbreath, J. J., and Merrell, D. S. (2012). Detailed analysis of Helicobacter pylori Fur-regulated promoters reveals a Fur box core sequence and novel Fur-regulated genes. Mol. Microbiol. 84, 921–941. doi: 10.1111/j.1365-2958.2012.08066.x
Raghwan and Chowdhury, R. (2014). Host cell contact induces fur-dependent expression of virulence factors CagA and VacA in Helicobacter pylori. Helicobacter 19, 17–25. doi: 10.1111/hel.12087
Rocker, A., and Meinhart, A. (2016). Type II toxin: antitoxin systems. More than small selfish entities? Curr. Genet. 62, 287–290. doi: 10.1007/s00294-015-0541-7
Sachs, G., Weeks, D. L., Melchers, K., and Scott, D. R. (2003). The gastric biology of Helicobacter pylori. Annu. Rev. Physiol. 65, 349–369. doi: 10.1146/annurev.physiol.65.092101.142156
Salama, N. R., Hartung, M. L., and Muller, A. (2013). Life in the human stomach: persistence strategies of the bacterial pathogen Helicobacter pylori. Nat. Rev. Microbiol. 11, 385–399. doi: 10.1038/nrmicro3016
Sevin, E. W., and Barloy-Hubler, F. (2007). RASTA-Bacteria: a web-based tool for identifying toxin-antitoxin loci in prokaryotes. Genome Biol. 8:R155. doi: 10.1186/gb-2007-8-8-r155
Shao, Y., Harrison, E. M., Bi, D., Tai, C., He, X., Ou, H. Y., et al. (2011). TADB: a web-based resource for Type 2 toxin-antitoxin loci in bacteria and archaea. Nucleic Acids Res. 39, D606-611. doi: 10.1093/nar/gkq908
Silva-Herzog, E., Mcdonald, E. M., Crooks, A. L., and Detweiler, C. S. (2015). Physiologic stresses reveal a Salmonella persister state and TA family toxins modulate tolerance to these stresses. PLoS ONE 10:e0141343. doi: 10.1371/journal.pone.0141343
Singh, A., Hodgson, N., Yan, M., Joo, J., Gu, L., Sang, H., et al. (2012). Screening Helicobacter pylori genes induced during infection of mouse stomachs. World J. Gastroenterol. 18, 4323–4334. doi: 10.3748/wjg.v18.i32.4323
Tegtmeyer, N., Wessler, S., and Backert, S. (2011). Role of the cag-pathogenicity island encoded type IV secretion system in Helicobacter pylori pathogenesis. FEBS J. 278, 1190–1202. doi: 10.1111/j.1742-4658.2011.08035.x
Thompson, L. J., Merrell, D. S., Neilan, B. A., Mitchell, H., Lee, A., and Falkow, S. (2003). Gene expression profiling of Helicobacter pylori reveals a growth-phase-dependent switch in virulence gene expression. Infect. Immun. 71, 2643–2655. doi: 10.1128/IAI.71.5.2643-2655.2003
Tomb, J. F., White, O., Kerlavage, A. R., Clayton, R. A., Sutton, G. G., Fleischmann, R. D., et al. (1997). The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature 388, 539–547. doi: 10.1038/41483
Vannini, A., Roncarati, D., Spinsanti, M., Scarlato, V., and Danielli, A. (2014). In depth analysis of the Helicobacter pylori cag pathogenicity island transcriptional responses. PLoS ONE 9:e98416. doi: 10.1371/journal.pone.0098416
Wang, G., Alamuri, P., and Maier, R. J. (2006). The diverse antioxidant systems of Helicobacter pylori. Mol. Microbiol. 61, 847–860. doi: 10.1111/j.1365-2958.2006.05302.x
Wang, X., and Wood, T. K. (2011). Toxin-antitoxin systems influence biofilm and persister cell formation and the general stress response. Appl. Environ. Microbiol. 77, 5577–5583. doi: 10.1128/AEM.05068-11
Wang, Y., Wang, H., Hay, A. J., Zhong, Z., Zhu, J., and Kan, B. (2015). Functional RelBE-family toxin-antitoxin pairs affect biofilm maturation and intestine colonization in Vibrio cholerae. PLoS ONE 10:e0135696. doi: 10.1371/journal.pone.0135696
Wen, Y., Behiels, E., and Devreese, B. (2014). Toxin-Antitoxin systems: their role in persistence, biofilm formation, and pathogenicity. Pathog. Dis. 70, 240–249. doi: 10.1111/2049-632X.12145
Wen, Y., Marcus, E. A., Matrubutham, U., Gleeson, M. A., Scott, D. R., and Sachs, G. (2003). Acid-adaptive genes of Helicobacter pylori. Infect. Immun. 71, 5921–5939. doi: 10.1128/IAI.71.10.5921-5939.2003
Winther, K. S., and Gerdes, K. (2009). Ectopic production of VapCs from Enterobacteria inhibits translation and trans-activates YoeB mRNA interferase. Mol. Microbiol. 72, 918–930. doi: 10.1111/j.1365-2958.2009.06694.x
Yamaguchi, Y., and Inouye, M. (2011). Regulation of growth and death in Escherichia coli by toxin-antitoxin systems. Nat. Rev. Microbiol. 9, 779–790. doi: 10.1038/nrmicro2651
Yamaguchi, Y., Park, J. H., and Inouye, M. (2011). Toxin-antitoxin systems in bacteria and archaea. Annu. Rev. Genet. 45, 61–79. doi: 10.1146/annurev-genet-110410-132412
Yokoyama, K., Higashi, H., Ishikawa, S., Fujii, Y., Kondo, S., Kato, H., et al. (2005). Functional antagonism between Helicobacter pylori CagA and vacuolating toxin VacA in control of the NFAT signaling pathway in gastric epithelial cells. Proc. Natl. Acad. Sci. U.S.A. 102, 9661–9666. doi: 10.1073/pnas.0502529102
Keywords: H. pylori, HP0967, HP0968, toxin–antitoxin system, environmental cues
Citation: Cárdenas-Mondragón MG, Ares MA, Panunzi LG, Pacheco S, Camorlinga-Ponce M, Girón JA, Torres J and De la Cruz MA (2016) Transcriptional Profiling of Type II Toxin–Antitoxin Genes of Helicobacter pylori under Different Environmental Conditions: Identification of HP0967–HP0968 System. Front. Microbiol. 7:1872. doi: 10.3389/fmicb.2016.01872
Received: 19 August 2016; Accepted: 07 November 2016;
Published: 22 November 2016.
Edited by:
Dongsheng Zhou, Beijing Institute of Microbiology and Epidemiology, ChinaReviewed by:
Jennifer Angeline Gaddy, Vanderbilt University, USAAlessandra Alves De Souza, Secretaria de Agricultura e Abastecimento, Brazil
Copyright © 2016 Cárdenas-Mondragón, Ares, Panunzi, Pacheco, Camorlinga-Ponce, Girón, Torres and De la Cruz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miguel A. De la Cruz, bWlndWVsX2FuZ2VsXzgxQGxpdmUuY29t
†These authors have contributed equally to this work.