 Liliana Scorzoni
Liliana Scorzoni Ana C. A. de Paula e Silva
Ana C. A. de Paula e Silva Caroline M. Marcos
Caroline M. Marcos Patrícia A. Assato
Patrícia A. Assato Wanessa C. M. A. de Melo
Wanessa C. M. A. de Melo Haroldo C. de Oliveira
Haroldo C. de Oliveira Caroline B. Costa-Orlandi
Caroline B. Costa-Orlandi Maria J. S. Mendes-Giannini
Maria J. S. Mendes-Giannini Ana M. Fusco-Almeida*
Ana M. Fusco-Almeida*
- Laboratório de Micologia Clínica, Departamento de Análises Clínicas, Universidade Estadual Paulista (UNESP), Faculdade de Ciências Farmacêuticas, Araraquara, Brasil
The high rates of morbidity and mortality caused by fungal infections are associated with the current limited antifungal arsenal and the high toxicity of the compounds. Additionally, identifying novel drug targets is challenging because there are many similarities between fungal and human cells. The most common antifungal targets include fungal RNA synthesis and cell wall and membrane components, though new antifungal targets are being investigated. Nonetheless, fungi have developed resistance mechanisms, such as overexpression of efflux pump proteins and biofilm formation, emphasizing the importance of understanding these mechanisms. To address these problems, different approaches to preventing and treating fungal diseases are described in this review, with a focus on the resistance mechanisms of fungi, with the goal of developing efficient strategies to overcoming and preventing resistance as well as new advances in antifungal therapy. Due to the limited antifungal arsenal, researchers have sought to improve treatment via different approaches, and the synergistic effect obtained by the combination of antifungals contributes to reducing toxicity and could be an alternative for treatment. Another important issue is the development of new formulations for antifungal agents, and interest in nanoparticles as new types of carriers of antifungal drugs has increased. In addition, modifications to the chemical structures of traditional antifungals have improved their activity and pharmacokinetic parameters. Moreover, a different approach to preventing and treating fungal diseases is immunotherapy, which involves different mechanisms, such as vaccines, activation of the immune response and inducing the production of host antimicrobial molecules. Finally, the use of a mini-host has been encouraging for in vivo testing because these animal models demonstrate a good correlation with the mammalian model; they also increase the speediness of as well as facilitate the preliminary testing of new antifungal agents. In general, many years are required from discovery of a new antifungal to clinical use. However, the development of new antifungal strategies will reduce the therapeutic time and/or increase the quality of life of patients.
Introduction
When compared with antibacterial research, little progress has been made in the development of new antifungal agents, which has been justified by the low occurrence of fungal infections. However, the current increase in incidence of fungal infections has led to aggressive research on new antifungal agents as evidenced by the rise in the number of publications since the 1960s (Maertens, 2004; Ngo et al., 2016). Another reason for the slow development of antifungal agents is the fact that fungi are eukaryotic, with a close evolutionary relationship with human hosts, which complicates the search for antifungal targets. Nonetheless, detailed knowledge regarding the structure, composition and biochemistry of fungal cells, in addition to various facets of fungal infections, has contributed to our understanding about the mechanism of action of many antifungal agents (Borgers, 1980; Kanafani and Perfect, 2008). Typically a long period of 8 to 10 years is required for an antifungal to be approved for clinical use. Reducing toxicity, enhancing bioavailability, improving the antifungal spectrum and combating resistance are efforts that are expected to increase the efficacy of the available antifungals. Indeed, elucidation of the mode of action of a potential antifungal compound can shorten the time from lead to candidate drug. Small antifungal molecules from natural products could represent structural templates for structure-activity relationship studies, thus providing more information to optimize potential new antifungal agents (Sheng and Zhang, 2011). Overall, new strategies regarding antifungal therapy, target identification and rational drug design technologies can significantly accelerate the process of new antifungal development, reducing the time to cure or providing better quality of life to patients.
Antifungal Mechanism of Action: Old and New Targets of Antifungal Candidates
Although the commercially available antifungal agents to date have targets that are restricted to the plasma membrane and the cell wall (Odds et al., 2003; Sundriyal et al., 2006; Ngo et al., 2016), a certain diversity of targets has been discovered. To develop new therapies, recent studies have focused on the inhibition of fungal virulence factors. Some mechanisms of action are described below, and an overview is presented in Figure 1.
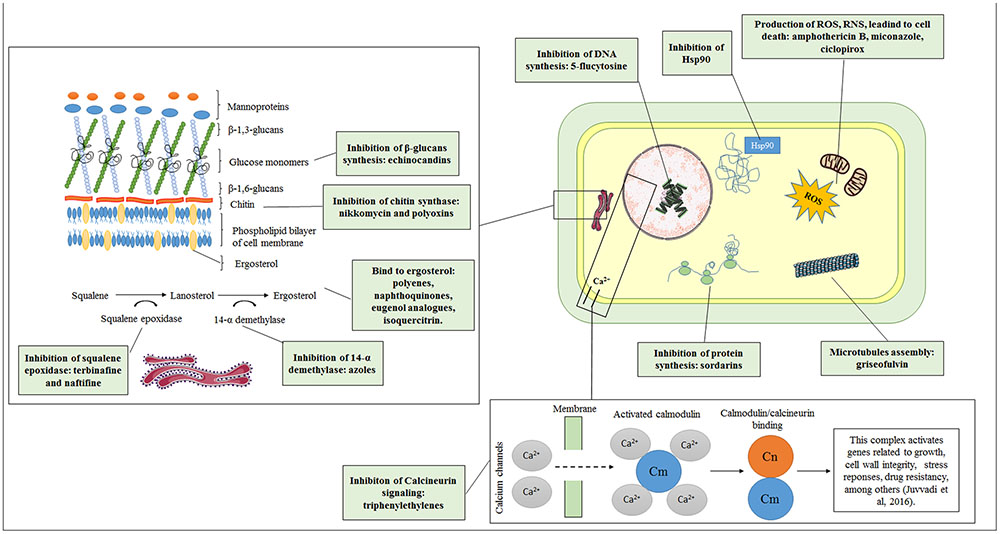
FIGURE 1. Old and new targets as antifungal candidates.
Ergosterol
Ergosterol is a lipid responsible for membrane fluidity and permeability and for the function of fungal integral membrane proteins; accordingly, this sterol is essential for cell viability (Leber et al., 2003; Tatsumi et al., 2013; Song et al., 2016). Several antifungals primarily target ergosterol, either by inhibiting its biosynthesis or by binding to it, causing formation of pores in the membrane.
Azole antifungals act by inhibiting ergosterol biosynthesis via the cytochrome P450 enzyme 14-α demethylase, which catalyzes the conversion of lanosterol to ergosterol (Kathiravan et al., 2012). Azoles affect the integrity of fungal membranes, altering their morphology and inhibiting growth (Kathiravan et al., 2012; Tatsumi et al., 2013).
The allylamine class, represented by terbinafine and naftifine, function by inhibiting the early steps of fungal ergosterol biosynthesis (Kathiravan et al., 2012) by targeting the enzyme squalene epoxidase, encoded by ERG1. This inhibition leads to accumulation of squalene and the absence of other sterol derivatives. Allylamines are highly effective against dermatophytes because they have been shown to accumulate more in the skin and nail beds relative to the blood, possibly due to their lipophilicity (Ngo et al., 2016).
Polyenes, such as nystatin and amphotericin B, exhibit fungicidal activity primarily by binding to ergosterol to form a complex capable of disrupting the membrane and leading to leakage of monovalent ions as well as other cytoplasmic contents (Odds et al., 2003; Walsh et al., 2008; Baginski and Czub, 2009). A second mechanism of polyene action involves a cascade of oxidation reactions and interactions with lipoproteins that impair membrane permeability through the release of free radicals (Sangalli-Leite et al., 2011; Mesa-Arango et al., 2012, 2014).
Cell Wall
The fungal cell wall, which is primarily composed of chitin, glucans, mannans, and glycoproteins, is essential for adhesion and fungal pathogenesis and also serves as a protective barrier, limiting the access of molecules to the plasma membrane (Bowman and Free, 2006; van der Weerden et al., 2013). The two main mechanisms of action of antifungals targeting the cell wall are related to the inhibition of chitin and β-glucan synthesis.
In the period between 2001 and 2006, the echinocandin class of drugs, represented by caspofungin, micafungin, and anidulafungin, was developed. This class has different mechanisms of action that are specific for the fungal cell wall. Echinocandins target the protein complex responsible for the synthesis of β-1,3 glucans by blocking the enzyme glucan synthase (Odds et al., 2003). This blockage causes a decrease in the incorporation of glucose monomers linking β-1,3 and β-1,6 glucans, thereby weakening the cell wall and leading to fungal cell lysis (Kathiravan et al., 2012; Song and Stevens, 2016).
Chitin, a β-1-4-linked N-acetylglucosamine polymer, is an essential component of the fungal cell wall, though it is only present in very small amounts in yeasts (1–2%) but in considerable quantities in filamentous fungi (10–20%) (Bowman and Free, 2006; Morozov and Likhoshway, 2016). Nikkomycin and polyoxins are antifungal agents that target chitin synthase, which is responsible for elongation of the chitin chain and, therefore, is considered an attractive target (Kathiravan et al., 2012).
Inhibition of Nucleic Acid, Protein, and Microtubule Syntheses
Inhibition of nucleic acid synthesis is related to the action of 5-flucytosine, which is converted primarily to 5-fluorouracil by the enzyme cytosine deaminase and then to 5-fluorouridylic acid by UMP pyrophosphorylase (Odds et al., 2003). Although 5-flucytosine was synthesized in 1957, its antifungal property was not discovered until 1964, 7 years later (Shukla et al., 2016). This acid can be incorporated into RNA, resulting in premature chain termination, thereby inhibiting DNA synthesis through effects on the enzyme thymidylate synthase (Polak and Scholer, 1975; Odds et al., 2003; Kathiravan et al., 2012).
With respect to the synthesis of microtubules, it is known that griseofulvin interferes with intracellular production, thus inhibiting fungal mitosis (Kathiravan et al., 2012).
Finally, sordarins suppress protein synthesis, which retards cell growth. Essentially, two fungal proteins have been described as target of sordarins: translation elongation factor 2 (eEF2) and the large ribosomal subunit protein rpP0 (Botet et al., 2008). Attempting to elucidate the mechanism of action, Justice et al. (1998) performed genetic assays using Saccharomyces cerevisiae mutants to demonstrate the fungal specificity of sordarins and proved that eEF2 is a target.
Reactive Oxygen Species (ROS)
It is known that treatment with some antifungals such as AmB and itraconazole can cause more than one effect on fungal cells (Ferreira et al., 2013; Mesa-Arango et al., 2014). According to Mesa-Arango et al. (2014), mitochondria naturally produce free radicals. However, under adverse conditions, such as in the presence of oxidants and UV light, these free radicals are produced in abundance, causing damage to proteins, lipids and DNA and leading to cell death. Accordingly, ROS production is also associated with apoptosis. Treatment with AmB is able to induce oxidative and nitrosative bursts in Candida, Cryptococcus, and Trichosporon, enhancing its fungicidal effect (Ferreira et al., 2013; Mesa-Arango et al., 2014).
Inhibition of Heat Shock Protein 90 (Hsp90)
Heat shock protein 90 (Hsp90) is a molecular chaperone of the heat shock protein (Hsp) family. Synthesized as an adaptive response to noxious conditions, these proteins contribute to the survival of pathogenic microorganisms in the host (Jacob et al., 2015). Hsp90 has been related to fungal pathogenicity, phase transition in dimorphic fungi and antifungal drug resistance, making it a potential target for antifungal therapy (Burnie et al., 2006; Brown et al., 2010; Jacob et al., 2015). Jacob et al. (2015) examined the transcription profiles of Trichophyton rubrum under different stress conditions, such as interaction with nail and skin cells and molecules, nutrients and treatment with antifungal drugs. In addition to suggesting the role of Hsp90 in the pathogenesis and susceptibility to dermatophytosis antifungal agents, the authors also related this protein to the regulation of other heat shock proteins.
Inhibition of Calcineurin Signaling
Calcineurin is defined as a conserved Ca2+-calmodulin(CaM)-activated protein phosphatase 2B that belongs to the phosphor-protein phosphatase family (Juvvadi et al., 2016). This protein is involved in calcium-dependent signaling and regulation of several important cellular processes in yeasts (Candida spp., Cryptococcus spp.) and filamentous fungi (Aspergillus fumigatus), including growth, cell wall integrity, transition between morphological states, cation homeostasis, stress responses, and drug resistance (Blankenship et al., 2003; Steinbach et al., 2006; Chen et al., 2013; Juvvadi et al., 2016). More important is the role of calcineurin in maintaining the integrity of the fungal cell wall by regulating downstream effectors and influencing the biosynthesis of ergosterol, chitin and β-glucans. Odom (2014) suggested that triphenylethylenes, described by Butts et al. (2014), are a novel class of antifungal drugs that act on calcium homeostasis in Cryptococcus neoformans via direct inhibition of the calcineurin activator calmodulin.
Molecular Mechanisms of Antifungal Resistance
The widespread use of antifungal agents and the limited arsenal associated with the increased number of opportunistic infections have resulted in the progression of resistance to available drugs. The antifungal resistance mechanism may occur through different conditions such as a decrease in the effective drug concentration, changes or overexpression of the drug targets, and metabolic bypasses (Sanglard, 2016). Figure 2 depicts an overview of several antifungal resistance mechanisms described for Candida spp.
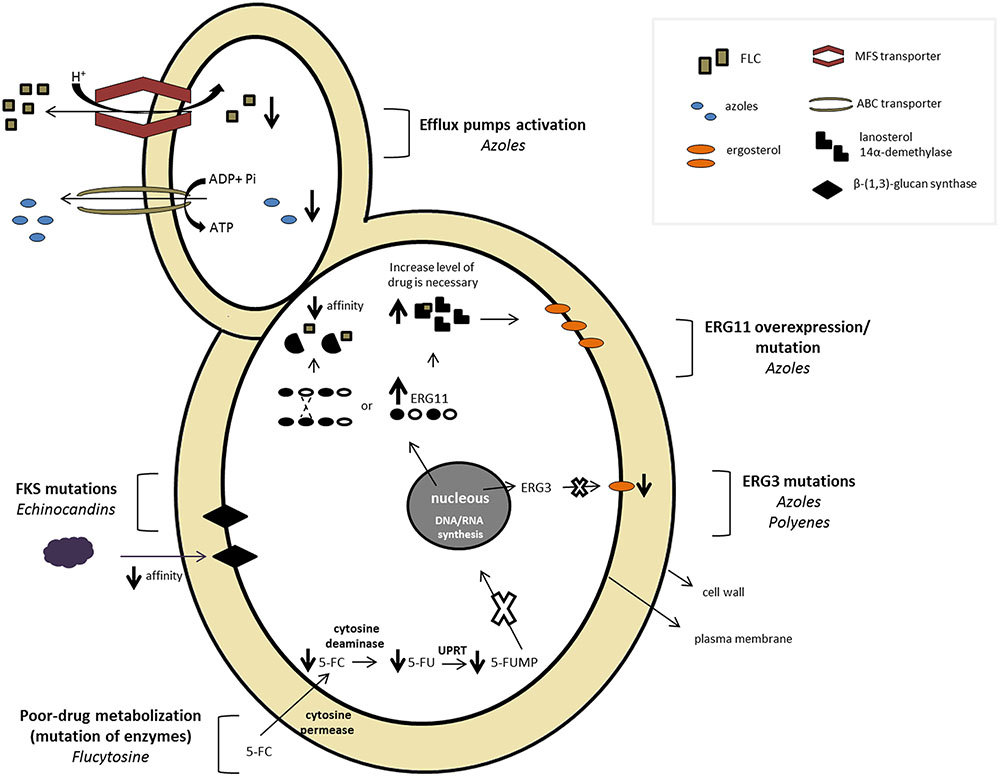
FIGURE 2. Candida sp. mechanisms of resistance to different antifungal classes.
Molecular Mechanisms of Resistance to Azoles
Azole resistance includes the following mechanisms: (1) activation of efflux pumps, (2) qualitative changes in the target enzyme, (3) quantitative changes caused by overexpression of ERG11, and (4) alterations in cell wall composition.
Activation of Efflux Pumps
Reduction in intracellular antifungal accumulation in Candida spp. is a consequence of the overexpression of membrane-associated transporters acting as multidrug efflux pumps (Prasad and Rawal, 2014). Two main classes of transporters are described as being involved in this resistance mechanism. The superfamily of ATP-binding cassette (ABC) proteins comprises the primary activity, hydrolyzing ATP to provide energy to drive the efflux of drugs. Transporters belonging to the major facilitator superfamily (MFS) constitute the secondary activity; these pumps utilize a proton electrochemical gradient across the plasma membrane to extrude substrates (Cannon et al., 2009). Although most Candida species are naturally susceptible to azoles, an increasing number of cases of acquired resistance have been reported in clinical isolates of patients exposed to prolonged treatment, especially to FLZ (White et al., 1998; Pfaller and Diekema, 2007; Arendrup, 2014; Espinel-Ingroff et al., 2014).
Candida albicans azole-resistant isolates can overexpress one or more efflux pumps (White, 1997; Franz et al., 1998; Lopez-Ribot et al., 1998). C. albicans possesses 28 putative types of ABC transporters, two of which, CDR1 and CDR2, are well characterized and overexpressed in resistant-isolates. Cdr1 has a greater contribution to FLZ resistance than Cdr2 (Sanglard et al., 1997; Sanglard et al., 2009; Prasad and Goffeau, 2012). Additionally, gain-of-function mutation of TAC1 (transcriptional activator of CDR genes) is related to increased levels of CDR1 and CRD2 (Coste et al., 2007). Regarding MFS transporters, C. albicans has 96 potential MFS transporters, though only one has been described with respect to azole resistance, the MDR1 member of the DHA1 family (Gaur et al., 2008).
It has been postulated that CDR genes in C. albicans are involved in the removal of different azoles. CDR1 efflux is associated with a wide range of substrates, whereas MDR1 appears to be specific for FLZ, as its overexpression results in moderate resistance to FLZ (Sanglard et al., 1995; White et al., 1998; Hiller et al., 2006; Cheng et al., 2007). The ABC and MFS pumps differ in relation to their structures and mechanisms, and differences in substrate specificity are expected (Keniya et al., 2015).
The exact mechanism underlying FLZ resistance in C. glabrata is not well defined; however, different studies indicate an essential role for ABC transporters (Abbes et al., 2013). FLZ-resistant C. glabrata clinical isolates present CDR1 and CDR2 upregulation (Sanglard et al., 1999; Bennett et al., 2004). Sanguinetti et al. (2005) observed increased expression of SQN2 (another ABC transporter) in two isolates (among 29) expressing normal levels of CRD1 and CRD2; SQN2 is possibly involved in the resistance of this species.
Studies have demonstrated that the primary mechanisms of azole resistance in clinical isolates of C. dubliniensis is upregulation of Mdr1 (Moran et al., 1998). Mdr1 is invariably overexpressed in C. dubliniensis strains with reduced susceptibility to FLZ (Perea et al., 2001). Moreover, among clinical isolates obtained from HIV-infected individuals with oropharyngeal candidiasis, Perea et al. (2002) reported upregulation of Mdr1 in all isolates with a high FLZ-resistance level, whereas less than half of the isolates presented CDR upregulation. Despite the 92% identity of the C. dubliniensis CDR1 gene with C. albicans, this may occur because C. dubliniensis genotype 1, previously described by Gee et al. (2002), possesses a non-sense mutation in CDR1 that converts a normal codon to a stop codon (TAG), resulting in expression of a truncated protein of 85 kDa instead of the wild-type 170 kDa (Perea et al., 2002).
Candida krusei carries two homologous genes for ABC transporters previously described for C. albicans: ABC1 and ABC2. The ABC1 gene is upregulated in cultures exposed to imidazole and cycloheximide. However, cycloheximide exhibits antagonist activity against FLZ, and this correlation between antagonism and upregulation suggests that FLZ and other azoles may be substrates for the ABC1 transporter (Katiyar and Edlind, 2001). Thereafter, Lamping et al. (2009) showed the involvement of ABC1 in the innate azole resistance of C. krusei, and Guinea et al. (2006) found that MDR proteins have a role in C. krusei resistance. However, expression of MFS transporters was not reported determined in FLZ-resistant C. krusei.
The FLZ resistance of C. parapsilosis has been linked to overexpression of MRR1 in high association with a mutation (Zhang et al., 2015) that concomitantly results in overexpression of MDR1 and overexpression of CRD1 (Souza et al., 2015; Zhang et al., 2015). The involvement of efflux pumps in C. tropicalis clinical isolate antifungal resistance has not yet been observed, but there is an in vitro study demonstrating the development of FLZ-resistance associated with the up-regulation of MDR1 and CDR1 (Barchiesi et al., 2000).
The involvement of efflux pumps has also been described in other pathogenic fungi. In C. neoformans, overexpression of the AFR1 gene, of the ABC transporter family, is associated with FLZ resistance (Posteraro et al., 2003; Sanguinetti et al., 2006). In addition, afr2p and Mdr1p, members of the ABC and MFS families, respectively, from C. neoformans and C. gattii can also promote resistance to FLZ and other azoles (Basso et al., 2015).
The ABC transporter Afr1 also acts in azole resistance in Aspergillus because overexpression of the AFR1 gene from A. fumigatus can confer itraconazole resistance (Slaven et al., 2002). Although Aspergillus fumigatus has four Mdr-like efflux pumps, itraconazole-resistant isolates or strains under itraconazole treatment only present overexpression of mdr3 and mdr4 (da Silva Ferreira et al., 2004).
Efflux appears to be the most prevalent mechanism of resistance in dermatophytes. Increased expression of the ABC transporters genes TruMDR1 and TruMDR2 from T. rubrum is observed in the presence of azoles (Cervelatti et al., 2006; Fachin et al., 2006). The use of efflux pumps by dermatophytes is also implicated in resistance to other antifungals, such as terbinafine, amphotericin B and griseofulvin (Cervelatti et al., 2006; Paião et al., 2007; Yu et al., 2007; Ghannoum, 2016; Martins et al., 2016).
ERG11: Overexpression and Changes in Drug Targets
In the presence of FLZ, C. albicans increases ERG11 expression, most likely via compensatory mechanisms, to deplete ergosterol (Albertson et al., 1996). However, Franz et al. (1998) showed that even in the absence of FLZ, resistant isolates express ERG11 at higher levels compared to susceptible isolates exposed to the drug. Overexpression of ERG11 results in increased concentrations of lanosterol 14α-demethylase; consequently, larger amounts of the antifungal are required to inhibit the enzyme (Morschhäuser, 2002). This mechanism has been described for many C. albicans FLZ-resistant isolates (White et al., 2002; Chau et al., 2004; Goldman et al., 2004). ERG11 overexpression in azole-resistant isolates can occur via two mechanisms of ERG11 amplification. One is formation of an isochromosome containing two copies of the left arm of chromosome 5, where ERG11 resides, and duplication of the entire chromosome (Selmecki et al., 2006). The second mechanism is mutation in the zinc cluster finger transcription factor Upc2, which results in overexpression of ERG11 in C. albicans (Dunkel et al., 2008). Overexpression of ERG11 has also been reported for other azole-resistant Candida species; however, the mechanisms remain unknown (Barchiesi et al., 2000; Redding et al., 2002; Vandeputte et al., 2005; Rogers, 2006; Jiang et al., 2013; Cowen et al., 2014).
Another mechanism of azole resistance among Candida strains involves non-synonymous point mutations in the ERG11 gene, which encodes the target enzyme lanosterol 14α-demethylase. Mutations in ERG11 can result in post-translational modifications in the amino acid sequence and consequently in the three dimensional structure of Erg11p, causing decreased binding affinity for azole components and also reducing ergosterol biosynthesis without impeding enzyme function but generating yeast with an altered phenotype resistant to azole (Xiang et al., 2013). Among 160 different amino acid substitutions, only 10 have been confirmed in FLZ resistance; four were obtained in the laboratory but not yet detected in clinical isolates (Chen et al., 2007). The substitutions R467K, I471T, G464S, S405F, and K143R were only related to azole-resistant C. albicans isolates (Sanglard et al., 1998; Lamb et al., 2000; Morio et al., 2010), with the most common being R467K and G464S (Morio et al., 2010). ERG11 mutations have been described for other azole-resistant clinical Candida isolates, including C. dubliniensis (Perea et al., 2002), C. krusei (Ricardo et al., 2014), C. tropicalis (Vandeputte et al., 2005; Jiang et al., 2013) and more recently C. parapsilosis (Grossman et al., 2015). However, the same finding has not yet described for C. glabrata (Gonçalves et al., 2016).
In non-Candida species, the most common mechanism of azole resistance is alteration of the target protein. Several studies describe point mutations in the ERG11 (CYP51) gene, encoding 14-α-demethylase, leading to amino acid substitution that decreases the affinity for azole (Xie et al., 2014).
In Cryptococcus spp., three point mutations in ERG11 have been described in association with azole resistance: point mutation G1855T leading to amino acid substitution of glycine 484 with a serine (G484S) (Rodero et al., 2003); substitution of tyrosine 132 by phenylalanine occurring in the catalytic domain (Sionov et al., 2012); and point mutation G1855A, also resulting in the amino acid substitution G484S (Bosco-Borgeat et al., 2016). Cryptococcus neoformans under FLZ stress is also able to adapt via duplication of chromosome 1, on which the genes ERG11 and AFR1 and that encoding the FLZ transporter protein are found (Sionov et al., 2010).
Aspergillus spp. have two genes encoding 14-α-demethylase: CYP51A and CYP51B. However, azole resistance is more associated with mutation in CYP51A (Gonçalves et al., 2016). Different point mutations and non-synonymous mutations have been described in CYP51A, and the different patterns of azole resistance depend on these mutations (Diaz-Guerra et al., 2003; Mann et al., 2003; Chowdhary et al., 2014a; Leonardelli et al., 2016). For example, non-synonymous mutations in codons 98,138, 220, 431, 434, and 448 may confer resistance to all azoles in Aspergillus spp. (Howard et al., 2009).
In drug-resistant Aspergillus, tandem repeat mutation associated with amino acid substitution in the CYP51A promoter have been described, such as TR34/L98H and TR46/Y121F/T289A (Verweij et al., 2007; Vermeulen et al., 2013). The most prevalent is TR34/L98H, which is found in different resistant Aspergillus isolates worldwide and seems to be the major mutation associated with azole resistance (Lockhart et al., 2011; Chowdhary et al., 2012, 2014b; Badali et al., 2013; Seyedmousavi et al., 2013; Chen et al., 2016).
In Histoplasma capsulatum, a Y136F substitution in CYP51Ap has also been implicated in decreased susceptibility to FLZ and voriconazole (Wheat et al., 2006).
Erg3 Mutations: Alteration of Ergosterol Biosynthesis
Fungi exposed to azoles suffer from ergosterol depletion and accumulation of toxic sterols, resulting in growth arrest (Kanafani and Perfect, 2008). Another less frequent mechanism of azole resistance is inactivation of the enzyme sterol Δ5,6-desaturase encoded by the gene ERG3, which is essential for ergosterol biosynthesis (Sanglard and Odds, 2002). Therefore, ERG3 protects yeast against toxic sterols; in contrast, deletions or mutations in ERG3 result in high levels of azole resistance once the production of toxic sterols is bypassed (Watson et al., 1989; Kelly et al., 1997). ERG3 mutants have been well studied in C. albicans and C. dubliniensis (Morio et al., 2012). Nonetheless, few of these mutations result in amino acid changes in Erg3 (Pinjon et al., 2003; Chau et al., 2005; Martel et al., 2010; Vale-Silva et al., 2012). Indeed, the exact mechanism by which a single substitution results in azole resistance needs to be investigated. Studies to date have demonstrated that cross-resistance between azoles and polyenes may occur because of ERG3 loss of function, which results in low ergosterol contents, protecting yeast against the toxic effects of AmB (Anderson et al., 2014; Sanglard, 2016). Some studies point to other mutations in genes of ergosterol biosynthesis suspected to be related to azole resistance, such as ERG6, ERG24, and ERG2 (Jensen-Pergakes et al., 1998; Jia et al., 2002; Vincent et al., 2013).
Molecular Mechanisms of Resistance to Flucytosine
In relation to 5-FC, approximately 10% of C. albicans isolates present primary resistance, even in the absence of drug exposure (Arikan and Rex, 2005). Different studies have shown that resistance to 5-FC is related to its metabolism. In C. glabrata and C. albicans, mutation in cytosine deaminase confers primary resistance to 5-FC, and deficiencies in cytosine permease activity have also been associated with resistance in Candida species (Hope et al., 2004; Vandeputte et al., 2011). Cytosine permease is involved in the uptake of 5-FC, after which cytosine deaminase produces 5-fluorouracil using cytosine; thus, 5-FC resistance is associated with deficiency in enzymes related to uptake, transport and transformation of 5-FC that result in failure to metabolize to the active drug (Bondaryk et al., 2013). A C. lusitaniae mutant lacking the enzyme uracil phosphoribosyl transferase, encoded by the gene FUR1 (Papon et al., 2007), exhibits secondary resistance to 5-FC. The enzyme UPRT converts 5-fluorouracil to 5-fluorouridine monophosphate and inhibits thymidylate synthetase by disrupting DNA synthesis.
The molecular mechanism of the resistance of Cryptococcus neoformans to 5-FC is not well established but is often related to mutation in pyrimidine salvage enzymes, as occurs in C. albicans (Whelan, 1987).
Song et al. (2012) demonstrated that C. neoformans might possess different mechanism of resistance against 5-FC that are based on sensor histidine kinases. C. neoformans possesses a two-component system, Tco2 and Tco1, to regulate 5-FC response; deletion of TCO2 leads to strong 5-FC resistance, and mutation in TCO1 increases susceptibility. In addition, through transcriptomic analysis, it was found that 5-FC-regulated genes in C. neoformans differ from those of S. cerevisiae. As most of these genes are of unknown function in other fungi, C. neoformans appears to have a unique mechanism of resistance against 5-FC.
Molecular Mechanisms of Resistance to Echinocandins
Mutations in the FKS1 gene lead to alterations in the conformation of the encoded enzyme, resulting in lower affinity between Fks1 and echinocandins and consequently resistance to these drugs (Gonçalves et al., 2016). Mutations in two hot spot regions of FKS1 are conserved in clinical isolates of C. albicans: the region between 641 and 648 (comprising a cytoplasmic domain/binding site of echinocandins) and 1345–1365 are hot spot 1 and hot spot 2 (HS2), respectively. These sites are responsible for the majority of mutations conferring resistance to echinocandins, including the most described: substitution of serine at position 645 (Balashov et al., 2006). Different studies have demonstrated alterations in FKS1 in other Candida species (Park et al., 2005; Desnos-Ollivier et al., 2008; Garcia-Effron et al., 2008, 2010), whereas mutations in Fks1 and its paralog Fks2 have been associated with resistance in C. glabrata (Garcia-Effron et al., 2009; Pham et al., 2014; Perlin, 2015).
Candida parapsilosis and C. guilliermondii show reduced susceptibility to echinocandins, most likely due to a natural polymorphism in the Fks1p hot spot region corresponding to mutations acquired in resistant isolates of other species: substitution of proline to alanine at positions 660 and 642 (Perlin, 2007; Garcia-Effron et al., 2008). Studies have shown that activation of cell wall recovery or compensatory pathways increases chitin production and that mutations that increase the chitin level result in reduced caspofungin susceptibility in C. albicans (Plaine et al., 2008).
Alterations in the Fks subunit have also been associated with echinocandin resistance in other fungi. A mutant with an amino acid substitution S678P in Fks1p resulted in resistance of A. fumigatus to echinocandins (Rocha et al., 2007). Alterations in FKS are also involved in Fusarium intrinsic resistance to echinocandin (Katiyar and Edlind, 2009).
Cryptococcus neoformans is intrinsically resistant to echinocandins without Fks alteration (Maligie and Selitrennikoff, 2005). Recently, Huang et al. (2016) investigated the molecular basis of C. neoformans resistance to echinocandins through a high-throughput genetic screen and found that the CDC50 gene may be involved in echinocandin resistance. CDC50 encodes the β-unit of membrane lipid flippase, which mediates the lipid trafficking pathway.
Molecular Mechanisms of Resistance to Polyenes
Although the development of acquired resistance to AmB rarely occurs in Candida, there are some reports to date (Favel et al., 2003; da Matta et al., 2007). Resistance to AmB generated a common phenotype with alterations in the membrane lipid composition and consequently a change in fluidly and permeability. A greater number of cases of therapy failure of AmB have been associated with C. lusitaniae (Minari et al., 2001; Favel et al., 2003; Atkinson et al., 2008). The main alterations involved in polyene resistance are in enzymes participating in ergosterol biosynthesis. Defects in ERG2 and ERG3, encoding C-8 sterol isomerase (converting fecosterol to episterol with low affinity for AmB) and Δ5,6-desaturase, respectively, result in quantitative and qualitative modifications in the membrane sterol content, influencing the amount of ergosterol or its availability for the action of polyenes (Arikan and Rex, 2005; Sheikh et al., 2013). A defective ERG3 gene resulted in low ergosterol levels in the fungal membrane of fungal, conferring azole/polyene cross-resistance to Candida isolates. Another likely AmB-resistance mechanism is via enhanced activity of catalases, which reduce oxidative damage (Sokol-Anderson et al., 1986; Kanafani and Perfect, 2008).
Although the mechanisms of resistance to AmB in Candida spp. are well described, these mechanisms in non-Candida species remain unclear. Resistance to AmB in C. neoformans isolated from AIDS patients was linked to alterations in sterol delta 8-7 isomerase (Kelly et al., 1994). Aspergillus strains are commonly resistant to AmB, though this varies among species, without alteration in ergosterol content. One of the mechanisms proposed for A. terreus AmB resistance is blockage of the Ras signaling pathway by Hsp90 and Hsp70, inhibiting the formation of aqueous pores (Blum et al., 2013; Blatzer et al., 2015).
Biofilm and Antifungal Resistance
The ability of many fungi to form biofilms is one of the reasons for antifungal drug resistance. Many medically important fungi are described as biofilm-forming organisms, such as Candida spp. (Chandra et al., 2005; Hinrichsen et al., 2008; Finkel and Mitchell, 2011; Pires et al., 2011), Pneumocystis spp. (Cushion et al., 2009), Coccidioides spp. (Davis et al., 2002), Aspergillus spp. (Mowat et al., 2009; Kaur and Singh, 2014), Zygomycetes (Singh et al., 2011), Malassezia spp. (Cannizzo et al., 2007; Figueredo et al., 2012), Trichosporon spp. (Di Bonaventura et al., 2006; Colombo et al., 2011), Cryptococcus (Walsh et al., 1986; Martinez and Casadevall, 2007), Histoplasma capsulatum (Pitangui et al., 2012), Trichophyton spp. (Costa-Orlandi et al., 2014) and Paracoccidioides spp. (Sardi et al., 2015).
Biofilms are highly structured and complex microbial communities embedded in a self-produced extracellular matrix (ECM) that attach to a wide range of surfaces and (Fanning and Mitchell, 2012). Several factors contribute to initial surface attachment, such as pH, temperature, osmolarity, flow of the surrounding biologic medium, host immune factors and even the presence of antimicrobial agents (Baillie and Douglas, 2000; Chandra et al., 2001; Ramage et al., 2008).
Fungal biofilm formation occurs through a sequential process including planktonic cell adhesion to an appropriate substratum, colonization, ECM production, biofilm maturation, and dispersion (Fanning and Mitchell, 2012). Despite these specific characteristics, all types of fungal biofilms have distinct properties from planktonic yeast cells and increase antifungal drug resistance up to 1000-fold (Ramage et al., 2001; Uppuluri et al., 2010). Indeed, several studies have shown the inefficacy of antifungal therapy against different fungal biofilms.
Multiple other biofilm-specific factors contribute simultaneously to the resistance of yeasts to antifungal drugs, including cell density, quorum sensing, efflux pump activity, persister cells, ECM presence, stress responses and overexpression of drug targets.
The cell density is an important factor that contributes to the antifungal resistance of biofilms. However, some studies show that this is not a biofilm-specific resistance mechanism because a similar trend was observed for planktonic cells. Perumal et al. (2007) studied the efficacy of different azoles, AmB and caspofungin on planktonic cells at densities similar to those found in biofilms. The susceptibility of dissociated biofilm cells was similar to that of planktonic cells at the same cell density, and this susceptibility decreased as the density of the cells increased (Perumal et al., 2007). This phenomenon was also demonstrated by Seneviratne et al. (2008), who noted the density-dependent susceptibility of planktonic or biofilm for ketoconazole and 5-FC. Lass-Florl et al. (2003) showed similar drug resistance results by increasing the inoculum sizes of Aspergillus species, supporting the idea that the physical density of the cells influences antifungal agent activity.
By providing microorganisms with the ability to communicate and coordinate population growth/morphology via the secretion of signaling molecules, cell density can be considered a key aspect of the quorum-sensing process (Ramage et al., 2009, 2012). QS for fungi was first described in C. albicans by Hornby et al. (2001), who identified farnesol as a molecule that inhibits the hyphal transitional stage of C. albicans and increases adhesion (Saville et al., 2003; Ramage et al., 2009). Exposing C. albicans to farnesol also resulted in alterations in gene expression involving hyphal developmental genes (TUP1 and CRK1), cell surface hydrophobicity genes and those involved in drug resistance (FCR1 and PDR16) (Cao et al., 2005; Enjalbert and Whiteway, 2005). In addition, farnesol induces apoptosis in both A. nidulans and Fusarium graminearum (Semighini et al., 2006, 2008). Tyrosol, the second QS molecule identified in C. albicans, promotes germ tube formation. According to Alem et al. (2006), tyrosol enhances the early phase of biofilm formation and may also inhibit farnesol activity, thereby controlling cell population morphology.
Khot et al. (2006) for the first time studied the mRNA levels of genes involved in ergosterol biosynthesis (ERG genes) and in β-1,6-glucan biosynthesis (SKN1 and KRE genes) in comparison between planktonic and biofilm-associated cells. The authors described the appearance of a unique transcript profile in a subpopulation of AmB-resistant blastospores with significant upregulation of ERG25, SKN1, and KRE1 and downregulation of ERG1. Mukherjee et al. (2003) reported that ergosterol levels were significantly decreased in intermediate and mature phases when compared to early-phase biofilms, suggesting that the biofilm resistance to azole might be explained by ergosterol alterations in biofilm membranes. Subsequent studies (Borecká-Melkusová et al., 2009; Nett et al., 2009; Nailis et al., 2010) compared the exposure of young and mature biofilms to fluconazole, concluding that both induce downregulation of genes encoding enzymes involved in ergosterol biosynthesis (CaERG1, CaERG3, CaERG11, and CaERG25). In addition, treatment of both young and mature biofilms with AmB predominantly resulted in overexpression of CaSKN1, with only modest upregulation of CaKRE1 (Nailis et al., 2010).
Induction of ergosterol pathway genes has been described in different biofilms of Candida species. For example, incubation with fluconazole caused upregulation of CdERG3 and CdERG25 in C. dubliniensis (Borecká-Melkusová et al., 2009) and of genes involved in ergosterol biosynthesis in C. parapsilosis, resulting in antifungal resistance (Rossignol et al., 2009).
Moreover, drug efflux pumps also participate in drug resistance of biofilms. Ramage et al. (2002) investigated the role of efflux pumps, ABC and MFS transporters, of C. albicans biofilm. Expression of CDR genes predominated at the beginning of biofilm formation (24 h), whereas MDR1 was solely overexpressed after 24 h. Expression of CDR1, CDR2, and MDR1 showed that C. albicans was susceptible to azoles when grown planktonically but exhibiting resistance when grown in a biofilm. Several subsequent studies confirmed these results, suggesting that the expression of these genes is necessary for biofilm resistance (Mukherjee et al., 2003; Perumal et al., 2007). Gao et al. (2014) showed that fluconazole increased the expression of CDR1, CDR2 and MDR1 and that the combination with doxycycline downregulated the gene overexpression induced by FLZ.
Later, Nett et al. (2009) performed an in vivo study of C. albicans biofilm formation on implanted catheters and showed upregulation of genes during biofilm development: CDR2 at 12 h and MDR1 at both 12 and 24 h. Similar results were reported for other Candida species, including C. glabrata, in which the expression of CDR1 and CDR2 was found at early (6 h) and intermediate (15 h) biofilm stages, even though neither gene was upregulated at the mature phase (48 h) (Song et al., 2009). C. tropicalis showed increased expression of MDR1 after 24 h of biofilm formation (Bizerra et al., 2008). In A. fumigatus, the initial phase of biofilm formation was associated with increased activity of the efflux pump MDR and gene upregulation at 8 h when treated with voriconazole (Rajendran et al., 2011). Thus, all reports support the idea that efflux pump expression is a mechanism of biofilm resistance, especially in the early phase of biofilm growth until ECM production.
The ECM is considered one of the essential mechanisms of biofilms resistance, conferring enhanced antimicrobial resistance and protection from host immune responses (Sutherland, 2001; Davies, 2003). The ECM may act as an adsorbent, reducing the amount of antimicrobial available to interact with the biofilm, and the structure physically reduces the penetration of antimicrobial agents by walling off access to regions of the biofilm (Taraszkiewicz et al., 2013). The matrix is composed of a variety of proteins, nucleic acids, phospholipids, lipids, amyloid fibers, humid substances, and in some cases, surprising amounts of extracellular DNA (e-DNA) (Estrela et al., 2009; Dogsa et al., 2013). The ECM confers important characteristics to biofilm, such as providing for mechanical stability, an external digestive system, and intense cell interactions, including cell–cell communication and synergistic microconsortia, and serving as a nutrient, energy, and recycling source (Flemming and Wingender, 2010).
During biofilm formation, β-1,3 glucan is one of the principle carbohydrate components of the ECM (Ramage et al., 2012). It is responsible for sequestering azoles, echinocandins, pyrimidines, and polyenes, acting as a “drug sponge,” and confers resistance to C. albicans biofilms (Nett et al., 2010a,b). In addition, the ECM of non-albicans Candida strains also contains β-1,3 glucan, which contributes to azole resistance via specific binding (Mitchell et al., 2013). FKS1 was the first gene described as encoding β-1,3 glucan synthase of C. albicans (Nett et al., 2010a). Other genes essential for the C. albicans ECM are SMI1 and RLM1, which are involved in the protein kinase C cell-wall integrity pathway, controlling the cell wall glucan content in response to stress. Recently, Taff et al. (2012) showed that glucan transferases and exoglucanase are crucial for the accumulation and delivery of β-1,3 glucan to the matrix.
Biofilm presents persister cells, which are directly correlated with accumulating high concentrations of antimicrobial agents. In essence, a persister is a dormant cell that with little or no cell wall synthesis; drugs bind to their target molecules but are unable to promote cell death (Lewis, 2007). The simplest route to form a dormant persister cell might be through the overproduction of proteins that are toxic to the cell and inhibit growth (Lewis, 2010). Different from bacteria, these cells have only been detected in yeast biofilms and not in planktonic populations (LaFleur et al., 2006). Due to the ECM, the persisters present in the biofilm can withstand both antifungal treatment and the immune system (Lewis, 2007). Persisters may be mainly responsible for re-infection once when the concentration of antimicrobial decreases, and they can repopulate the biofilm (Lewis, 2001). Recently, a dose-dependent study involving AmB and chlorhexidine against C. albicans in planktonic and biofilm forms reported complete elimination of planktonic cells in both exponential and stationary stages. However, biphasic killing occurred in the mature biofilm, suggesting the presence of persisters. After AmB treatment, surviving C. albicans in the biofilm capable of producing a new biofilm with a new subpopulation of persisters, suggesting that yeast persisters are not mutants but phenotypic variants of the wild-type population (LaFleur et al., 2006). In addition, Sun et al. (2016) showed that C. albicans AMB-tolerant persisters were produced mainly during the adhesion phase and that the maintenance of these cells was dependent of surface adhesion. Further studies showed the presence of persister cells after the treatment of C. krusei and C. parapsilosis biofilms with AmB. However, the biofilm of C. albicans SC5314 under the same treatment condition did not reveal surviving cells, indicating a lack of persister cells (Al-Dhaheri and Douglas, 2008).
Based on the above, it is possible to affirm that biofilm confers antifungal resistance and that this occurs through various mechanisms. Accordingly, the discovery of new strategies to overcome these microbial communities has been deeply researched.
The development of new drug formulations is one of the main options of studies, including echinocandins and AmB lipid forms that are inhibit fungal biofilm both in vitro (Kuhn et al., 2002; Ramage et al., 2013) and in vivo (Mukherjee et al., 2009; Kucharikova et al., 2013).
The association of drugs is another example of a strategy against fungal biofilms. Some studies have reported the efficacy of a combination of AmB and aspirin (Zhou et al., 2012), FLZ and doxycycline (Gao et al., 2013), and caspofungin and diclofenac (Bink et al., 2012). The sensitization of C. albicans biofilms to different antifungals by the immunosuppressant drug cyclosporine A also resulted in enhanced biofilm inhibition (Shinde et al., 2012).
The presence of e-DNA, which is an important component of the ECM, has prompted some strategies using DNase to decrease biofilm biomass and to enhance the activity of antifungal agents (Martins et al., 2010). Lactonases and α-amylases are also used to control fungal biofilms (Taraszkiewicz et al., 2013). Moreover, an interesting therapeutic option is to block persister survival. Bink et al. (2011) discovered that superoxide dismutase (SOD) may be an inhibitor of N,N’-diethyldithiocarbamate (DDC) in C. albicans biofilms, reducing the miconazole-resistant persister fraction by 18-fold (Bink et al., 2011).
The application of antimicrobial photodynamic therapy has been investigated for anti-fungal biofilm properties with regard to the efficiency of inhibiting several microorganisms, with minimal damage to the host cell (Biel, 2010). This therapy involves the combination of photosensitizer (PS), light and molecular oxygen (de Melo et al., 2013). In addition, strategies involving nanoparticles (Allaker, 2010; Ramasamy et al., 2016), plant extracts (Wojnicz et al., 2012), and chitosan (Carlson et al., 2008) have been applied against microbial biofilm, with significant results. Further studies are necessary for identifying means to overcome fungal biofilm resistance.
Drug Combinations
Based on the problems discussed above with regard to antifungal treatment, one of the options is the combination of drugs. Using more than one drug can increase efficacy due to the possibility of action on more than one target; in addition, toxicity is reduced because less of the drug is used (Chen et al., 2014). Drug combinations can lead to improved activity, such as synergist activity, or decrease antagonist action. Evaluation of the effect of a drug combination in vitro can be realized by the checkerboard method (Johnson et al., 2004).
Most studies on the in vitro and in vivo combination of azoles with AmB do not show synergistic activity, though previous treatment with azole can influence the action of polyenes. This can be explained by the fact that both drugs have the same target: ergosterol. However, because azoles inhibit ergosterol biosynthesis, less ergosterol is available for polyene to bind (Louie et al., 2001; Sanglard, 2002). Nonetheless, invasive mucormycosis has been successfully treat with an antifungal combination of AmB and posaconazole (Pagano et al., 2013).
There are numerous in vitro studies of antifungal combinations using triazole with echinocandin or triazole with AmB (Elefanti et al., 2013; Katragkou et al., 2014). Regardless, the clinical results of combinatory treatment remain unclear and are generally described for infections caused by fungi, which present difficulty in treatment, such as in aspergillosis and mucormycosis (Belanger et al., 2015). The combination of caspofungin and voriconazole showed in vitro synergistic activity against Aspergillus spp. (Walsh et al., 2008). Moreover, this combination showed efficacy in a mammalian model (Kirkpatrick et al., 2002). Treatment of patients with invasive aspergillosis with a combination of voriconazole and anidulafungin improved survival in comparison with treatment with voriconazole monotherapy (Marr et al., 2015). However, in another study, triazole and echinocandin combination showed the same effect as triazole alone (Raad et al., 2015).
The antifungal 5-FC in combination with AmB or with azole did not show synergistic activity for Candida sp. (Scheid et al., 2012). However, this was efficient for the treatment of cryptococcal meningitis. It should be noted that the lack of availability of 5-FC results in the use of less effective combinations in many countries (Perfect et al., 2010). Moreover, this compound is associated with rapid development of resistance (Vandeputte et al., 2011).
Interaction with non-antifungal agents has also been described as potentiating antifungal activity. Triclosan, a compound exhibiting antimicrobial activity, is widely used in soap, toothpaste, and other personal care products. This molecule showed in vitro synergistic activity with FLZ against C. albicans (Yu et al., 2011). Triclosan is also active against C. neoformans by activating the apoptosis pathway and also shows synergic activity with AmB and FLZ (Movahed et al., 2016). As ion homeostasis is a fundamental factor in the development of fungal disease, ion chelators have been used in the treatment of fungal infections. However, synergistic activity with antifungal drugs did not result in favorable responses in vitro (Lai et al., 2016). Treatment of cardio-vascular disorders has been realized with calcium channel blockers, which also demonstrate antifungal activity (Yu et al., 2014). Recent work shows that the calcium channel blockers amlodipine, nifedipine, benidipine, and flunarizine present synergistic activity with FLZ in C. albicans isolates resistant to FLZ via a mechanism not related to efflux pump inactivation (Liu S. et al., 2016).
Natural source molecules with antifungal activity are also described as having in vitro synergistic activity with traditional antifungal drugs, reducing the concentration of both substances (Soares et al., 2014; Sardi et al., 2016; Wang et al., 2016). Use of the natural substance beauvericin with traditional antifungals was able to inhibit efflux pumps and morphogenesis in FLZ-resistant C. albicans and potentiate the action of this azole (Shekhar-Guturja et al., 2016).
New Antifungal Formulations and New Antifungal Drug Structure Modification
Two different strategies have been developed to increase the therapeutic index of antifungal agents: chemical modifications and/or elaboration of new formulations of antifungal agents to obtain less toxic derivatives (Sheng and Zhang, 2011).
Theoretical and experimental studies on the mechanism of action of AmB and its derivatives were performed by Borowski (2000). Two generations of derivatives were developed. First-generation compounds are modified at the carboxyl group, which improves selective toxicity based on disturbance of the hydrogen bond network in complex with sterols. Second-generation compounds have introduction of a bulky substituent, resulting in an appropriate steric hindrance effect that disturbs interaction with cholesterol but not with ergosterol, leading to improved selective toxicity (Borowski, 2000). Among second-generation derivatives, N-methyl-N-D-fructosyl AMB methyl ester (MFAME) is considered the most interesting compound because it is able to form water-soluble salts, has a broad antifungal spectrum, and lower toxicity than AMB toward animal cells in in vitro and in vivo experiments (Szlinder-Richert et al., 2001).
Despite the huge effort made to decrease fungal resistance and the toxicity of AmB, the development of rational chemical modification of known antifungal agents was insufficient to solve these problems. Thus, new delivery systems have been evaluated to reach this goal (Borowski, 2000; Sheng and Zhang, 2011).
Since 1990, nanostructured systems have been studied as carriers of antifungal agents. Clinically, the intravenous dosage form of AmB-deoxycholate has adverse effects, mainly nephrotoxicity. The synthesis of AmB analogs such as AmB esters or a preparation including an AmB lipid complex, AmB colloidal dispersion, liposomal AmB and intralipid AmB have been generated to improve the therapeutic index and lower toxicity (Gupta and Tomas, 2003; Vyas and Gupta, 2006; Voltan et al., 2016).
In addition, other delivery systems, such as carriers based on solid and nanostructure lipids, synthetic and natural polymers, inorganic and metal nanostructure lipids, dendrimers, silica, and carbon materials (magnetic nanoparticles), have been pursued. These delivery systems are able to improve bioavailability and reduce toxicity and present specificity for target tissues; however, there is an associated high cost of production (Vyas and Gupta, 2006; Sato et al., 2015; Voltan et al., 2016).
Recent work shows that the composition of nanoparticles used as delivery vehicles is fundamental for increasing antifungal activity. Ahmad et al. (2016) produced a conjugate system with AmB and metal nanoparticles that displayed synergistic antifungal activity due to the antimicrobial property of silver against C. albicans and C. tropicalis. Niemirowicz et al. (2016) reported synergistic activity for the combination of polyenes and magnetic nanoparticles. This bio-active nano-sized formulation exhibited enhanced efficiency against two clinical isolates of Candida species in planktonic and biofilm states.
For many years, the only available antifungal for invasive fungal infections was AmB, which has been incorporated into three lipid formulations. However, the imidazole class offers new treatment options because it is proven to be less toxic and in some cases more effective than AmB (Shalini et al., 2011). Although the imidazole class has been available for a decade, improvements in safety were necessary. Accordingly, this class was also subjected to rational chemical modifications. The triazole class was generated by the addition of a nitrogen atom to a cyclic ring. This modification provided a broad spectrum of activity as well as improved safety and pharmacokinetic profile (Allen et al., 2015). In this sense, the introduction of triazoles accelerated the pace of drug development.
New azoles have been developed to combat resistant pathogens to improve the tolerability and administration. Voriconazole is structurally similar to FLZ, with the exception of a fluoropyrimidine group in place of a triazole moiety, which leads to better bioavailability. Alternatively, posaconazole has a spectrum of antifungal activity comparable to that of voriconazole, but the molecular structure with a hydroxylated analog is similar to that of itraconazole. Currently, isavuconazole, ravuconazole, albaconazole, and efinaconazole are the four best studied agents in research aiming to identify an ideal antifungal with a wide spectrum and reliable pharmacokinetics, parenteral and oral dosage forms, and a favorable adverse effect profile (Shalini et al., 2011; Allen et al., 2015).
In addition, Moazeni et al. (2016) reported that the combination of solid lipid nanoparticles with FLZ was able to avoid recognition by efflux pump proteins, preventing extrusion when tested against FLZ-resistant Candida isolates. In another study, an aqueous nano-suspension of itraconazole for the treatment of bronchopulmonary aspergillosis solved pharmacokinetic problem of adequate concentration (Rundfeldt et al., 2013).
In addition to all azole agents, echinocandins are among the newest class of antifungal agents and act by inhibiting glucan synthesis in the fungal cell wall. Echinocandins are composed of a complex hexapeptide core with an N-terminus acylated by a long hydrophobic chain. Although echinocandins are fungicidal with good selectivity, they cannot be orally administered because of their complex lipopeptide structure. To address the limitation, several small molecule glucan synthesis inhibitors have been discovered. However, none is under clinical evaluation thus far (Sheng and Zhang, 2011; Liu N. et al., 2016).
Antifungal Immunotherapy: Vaccines
Given their increasing frequency and unacceptably high morbidity and mortality rates, prevention of invasive fungal infections has become of vital importance (Spellberg, 2011; Medici and Del Poeta, 2015). Vaccination of high-risk groups is a particularly promising strategy to prevent invasive fungal infections because easily identifiable risk factors are clearly defined for many such infections (Perlroth et al., 2007; Spellberg, 2011).
Advances in our understanding of the host defense and pathogenic mechanisms underlying fungal infections has supported the development of effective vaccines to combat these diseases. Accordingly, researchers have dedicated studies to developing robust, durable and safe fungal vaccines, especially those that may be useful for endemic infections or in chronic or superimposed infections in intensive care patients (Santos and Levitz, 2014; Shahid, 2016).
Although very few clinical trials have been performed in humans, a growing number of antifungal vaccine candidates are being evaluated in pre-clinical studies. This may be due to the renewed interest in the potential use of vaccines, replacing or associated with chemotherapy, to reduce antifungal drug use and consequently limit drug resistance and toxicity.
For most active vaccines studied against invasive fungal infections, the key to protection has been the induction of cell-mediated, pro-inflammatory, Th1 or Th17 responses, which improve phagocytic killing of the fungus. It is also clear that antigens targeted for vaccination need not be restricted to virulence factors, markedly increasing the antigen repertoire available for testing. Additionally, the concept of niche vaccination of acutely at-risk patients or patients in restricted geographical areas is a new idea that opens doors in vaccinology (Spellberg, 2011).
The greatest advances in fungal vaccines are with regard to studies of invasive candidiasis, and two promising vaccines are presently in the clinical trial phase. The first, containing the rAls3p-N antigen, is in Phase IIa and prevents fungal adhesion and invasion in immunized hosts. Protection with this vaccine has been found to largely be mediated by T cells via neutrophil recruitment and a specific antibody in vaccinated hosts (Edwards, 2012; Shahid, 2016).
The second candidal vaccine in clinical trial phase is a virosome-based vaccine containing a Sap2 antigen/truncated recombinant Sap2 antigen. The truncated form is stable, immunogenic and harmless. The Sap2 vaccine administered intra-muscularly or intra-vaginally induces systemic and 100% mucosal protective immunity (Vecchiarelli et al., 2012; Shahid, 2016).
Another important molecule that has been studied in candidiasis vaccination is heat shock protein 90 (Hsp90-CA). Raska et al. (2005) tested an Hsp90-CA DNA vaccine and found that vaccinated mice exhibited survival times prolonged by 64% compared with the untreated control. In addition, vaccination with the recombinant protein r-hsp90-CA significantly increased survival compared to the control groups.
A monoclonal antibody that binds to the immunodominant epitope of C. albicans Hsp90p was tested pre-clinically. A synergistic action with AMB against FLZ-sensitive and -resistant C. albicans strains was demonstrated in in vivo and in vitro experiments. The same antibody was used in a multicenter double-blind placebo-controlled trial of patients with invasive candidiasis, and complete mycological resolution reached 84% in the combination therapy group compared to 48% in the AMB monotherapy group. Moreover, clearance of infections in the combination therapy group was twice that of AMB therapy alone, resulting in 4 and 18% mortality, respectively (Matthews et al., 2003; Bugli et al., 2013; Wang et al., 2015).
In a different approach, S. cerevisiae cells were genetically engineered to display Enolase 1 (Eno1p) antigens of C. albicans on their surface. Oral administration of these cells could elicit an immune response and aid the survival of mice challenged with C. albicans (Vecchiarelli et al., 2012; Shibasaki et al., 2013; Shahid, 2016). Interestingly, by sharing some elements with Aspergillus species, Candida enolase also exerts a protective function against aspergillosis.
Studies on vaccines protective against other fungi are still in the early pre-clinical phase (Fernandes et al., 2011; Hsieh et al., 2011; Edwards, 2012; Hole and Wormley, 2012; Shahid, 2016).
For protection against aspergillosis, studies have shown that the type 1, cell-mediated immune response, which is efficient in protecting against this disease, can be induced by recombinant protein antigens from Aspergillus, as observed by Bozza et al. (2002). Administration of recombinant allergen Asp 16 f in conjunction with CpG oligonucleotides improved the survival of mice infected by inhaled A. fumigatus. Vaccination with crude antigen preparations from A. fumigatus was also tested, and it was observed that such vaccination improved the survival of mice infected by inhaled and intravenously administered fungi, even in those that were subsequently immunocompromised (Cenci et al., 2000; Ito and Lyons, 2002).
In the case of Cryptococcus, Devi et al. (1991) proposed the use of an anti-phagocytic antigen from the capsule of C. neoformans, glucuronoxylomannan (GMX), as a vaccine to elicit antibody-mediated protection. In a different approach, Wozniak et al. (2011) administered T-cell depleted mice, mimicking a T-cell-deficient host, with an engineered strain of C. neoformans that could express IFN-g. After the immunization, a secondary pulmonary infection using a pathogenic strain was produced, and the mice were protected against the infection. The results demonstrate that it is possible to generate a protective immune response to Cryptococcus even after becoming immunocompromised, such as in cases of HIV.
Recently, Rella et al. (2015) proposed the use of a live attenuated mutant strain lacking sterol glucosidase enzyme (Δsgl1) as a vaccine. In this study, immunization of mice with Δsgl1 cells led to strong protection against challenge with C. neoformans and C. gattii and could also elicit protective immunity in mice deficient for T CD4+ cells, a frequent condition in cryptococcosis patients.
Another possible therapeutic agent in the prophylaxis of Cryptococcus is glycosphingolipid glucosylceramide (GlcCer), a C. neoformans virulence factor. In a recently study, Mor et al. (2016) demonstrated that administration of GlcCer prior to infection with C. neoformans in a murine model prevented dissemination of the fungi from the lungs to the brain and led to 60% mouse survival while preventing side effects such as hepatic injury, a common and great problem in the use of the antifungal therapy currently available.
A peptide derived from the gp43 adhesin of P. brasiliensis, named p10, protects mice from paracoccidioidomycosis (PCM), and combination of immunization with this peptide and different antifungal drugs showed an additive protective effect. The finding suggests that this is an important molecule to be tested in clinical trials against PCM (Taborda et al., 1998; Marques et al., 2006; Marques et al., 2008; Rittner et al., 2012).
To date, few vaccines have reached the clinical stage. However, the importance of mycosis to public health around the world indicates that efforts should be given to the discovery of new molecules and compounds that can be used in the prophylaxis and treatment of these diseases. The challenges are numerous, but the efforts offer hope for controlling these diseases that tend to increase in incidence over time.
Alternative Animal Models to Study Fungal Virulence and Antifungal Drugs
Classically, mammalian models are considered the gold standard for drug discovery, virulence and immune response studies. However, in 1959, the concept of the “alternative animal model” was described by Burch and Russell with “the three R’s: Refinement, Reduction, and Replacement” (Russell and Burch, 1959). In Table 1 are described the three R concept and the actions required for its application. In recent decades, the use of alternative animals, also called “mini hosts” or “non-conventional animal models” have been encouraging for in vivo testing. Amoeba, insects, nematodes, fish and chicken embryos are used for the following reasons: (1) because the neural systems of these animals are poorly developed, they are almost painless; (2) a large number of animals can be used for each experiment; (3) maintenance is cheaper than for traditional animals; (4) there is a good correlation between alternative animals and mammalian animals (Trevijano-Contador and Zaragoza, 2014).

TABLE 1. Burch and Russell: The three R’s concept and the action required for its application.
Chicken embryos have been used since 1971 for fungal virulence studies (Partridge et al., 1971), and many studies have been published using this model to evaluate virulence of different fungi such as Candida spp., Rhizopus, and A. fumigatus (Jacobsen et al., 2010, 2011; Kaerger et al., 2015). Moreover, with this model, it is possible to evaluate the role of genes using mutants and to study the immune response of the host during infection (Jacobsen et al., 2010). Despite the importance of this model to date, experiments of antifungal efficacy have not been validated. Although chicken embryos are not consider worldwide to be living organisms and the ethical issues for this type of experimentation are less complicated, it is important to highlight that the euthanasia performed after experiments should be administered properly using anesthesia (Aleksandrowicz and Herr, 2015).
Another non-conventional model widely used for in vivo testing is zebrafish embryos, larvae, and adults. The main advantage is that it is possible to analyze different biological processes due to the presence of organs and systems, providing complete raw data analysis for different fields of science (MacRae and Peterson, 2015). This model was useful for studying of the C. albicans virulence (Chao et al., 2010; Brothers et al., 2011; Chen et al., 2015) and C. neoformans pathogenesis (Tenor et al., 2015). Similar to chicken embryos, there is no description thus far of the evaluation of antifungal efficacy using zebrafish. However, toxicity assays and teratogenic testing have provided important information. It is essential to highlight that after 120 h post-fertilization (hpf), ethical regulations are required for zebrafish experiments (Strähle et al., 2012).
Insects can also be used as an alternative animal model. The fruit fly Drosophila melanogaster is described as a model for investigating the virulence of human pathogens, due to similarities between its immune system and that of mammals, and for verifying the efficacy of novel antifungal compounds. Infection can be achieved by injection, rolling contact or ingestion (Lionakis and Kontoyiannis, 2012). Different Candida species have been evaluated using this model; the virulence of C. parapsilosis was found to be lower than that of C. albicans, similar to observations in humans; FLZ also increased the survival of susceptible strains (Chamilos et al., 2006). This model is also suitable for virulence evaluation of C. neoformans mutants (Apidianakis et al., 2004). The virulence of clinical and environmental isolates of A. flavus have been evaluated, as well as different mating types (Ramírez-Camejo et al., 2014). In Drosophila melanogaster, antifungal treatment is realized by ingestion, mixed with food. This model showed a correlation with mammalian models with regard to synergism of tacrolimus with posaconazole for the treatment of Rhizopus oryzae (Lewis et al., 2013). Fusarium moniliforme and Scedosporium apiospermum are able to infect D. melanogaster, and ingestion of food containing voriconazole increased the survival of infected flies (Lamaris et al., 2007).
Galleria mellonella has been described in different fields of science for numerous purposes, and the number of studies is increasing every year. Concerning virulence fungal assays, studies have been reported using G. mellonella as an infection model for different Candida species (Scorzoni et al., 2013; Frenkel et al., 2016; Moralez et al., 2016), Cryptococcus spp. (Benaducci et al., 2016), Sporothrix schenckii (Clavijo-Giraldo et al., 2016), Paracoccidioides spp. (Thomaz et al., 2013; Scorzoni et al., 2015), Histoplasma capsulatum (Thomaz et al., 2013), and Fusarium sp. (Coleman et al., 2011) and also for biofilm formation and behavior studies (Fuchs et al., 2010a; Benaducci et al., 2016).
The advances of this model are due to the ease of manipulation, low cost, and large range of larval incubation temperatures (25–37°C) with the possibility of mimicking the human body temperature. However, one disadvantage is that the genome is not fully sequenced, resulting in a lack of mutant strains to study host responses. Most of the work related to infection and treatment of G. mellonella has been via injection through one of the prolegs (Fuchs et al., 2010b). The benefit of this is that the inoculum and the treatment are controlled. G. mellonella is useful for in vivo treatment with conventional antifungal drugs (Coleman et al., 2011; de Lacorte Singulani et al., 2016), for assessing antifungal synergistic activity (Gu et al., 2016; Sangalli-Leite et al., 2016) and for evaluating the in vivo efficacy of new drug candidates (Browne et al., 2014).
The nematode Caenorhabditis elegans is been used for decades in different fields of biology. This organism has a short life cycle, produces a large number of progeny, and is transparent; moreover, the genome is fully sequenced with the possibility of studying host pathways through RNA interference (RNAi) techniques or constructing transgenic strains. C. elegans is maintained on agar medium and fed non-pathogenic E. coli (Muhammed et al., 2016). C. elegans is susceptible to infection by human pathogenic yeasts and filamentous fungi (Johnson et al., 2009; Pukkila-Worley et al., 2009; Desalermos et al., 2015). For this reason, this organism can be used for high-throughput screening of antifungal drugs candidates. The C. elegans model of infection is well described in antifungal drug discovery. A study with 1,266 compounds identified 15 anti-candidal substances that prolonged the survival of infected larvae and inhibited in vivo filamentation of the yeast (Breger et al., 2007). In another screen with 3,228 candidate substances, nine compounds with potential antifungal activity were identified (Okoli et al., 2009). Synergistic antifungal activity of tyrocidines and caspofungin was successfully evaluated in vitro and in vivo using C. elegans (Troskie et al., 2014). Moreover, C. elegans is a suitable model for studying the immune response because it is able to produce different antimicrobial peptides that are regulated upon infection; these peptides can also serve as new antimicrobial candidates (Ewbank and Zugasti, 2011).
Although different types of animal models are discussed here, it is important to evaluate which is the best system for each study. Regardless, although alternative models have benefits, mammalian models are still considered the gold standard, and many additional studies are necessary before the complete substitution of mammalian models for alternative animals. At present, alternative animals are suitable for screening prior to further studies in mammals.
Conclusion
Despite the increasing number of reports regarding advances in antifungal therapy, the number of cases of infection and antifungal resistance are still alarmingly high, and control of antifungal disease is far from being achieved. Important new advances have been made in the discovery of antimicrobial fungal targets; however, many years are necessary from discovery to clinical use. Because of this, improving existing molecules and developing new formulations and alternative therapy for prevention and treatment are important for treating fungal infections and increasing treatment options and quality of life.
Author Contributions
All the authors contributed selecting the topics, reviewing the literature, selecting important and atual informations. Moreover the authors read and approved the final manuscript.
Funding
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo-FAPESP [2013/10917-9 (LS), 2015/03700-9 (MM-G), 2015/14023-8 (HdO), 2016/17048-4 (CM)], Coordenação de Aperfeiçoamento de Pessoal de Nível Superior and Programa de Apoio ao Desenvolvimento Científico da Faculdade de Ciências Farmacêuticas da UNESP and Conselho Nacional de Desenvolvimento Científico e Tecnológico 150261/2016-0 (CC-O).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abbes, S., Mary, C., Sellami, H., Michel-Nguyen, A., Ayadi, A., and Ranque, S. (2013). Interactions between copy number and expression level of genes involved in fluconazole resistance in Candida glabrata. Front. Cell Infect. Microbiol. 3:74. doi: 10.3389/fcimb.2013.00074
Ahmad, A., Wei, Y., Syed, F., Tahir, K., Taj, R., Khan, A. U., et al. (2016). Amphotericin B-conjugated biogenic silver nanoparticles as an innovative strategy for fungal infections. Microb. Pathog. 99, 271–281. doi: 10.1016/j.micpath.2016.08.031
Albertson, G. D., Niimi, M., Cannon, R. D., and Jenkinson, H. F. (1996). Multiple efflux mechanisms are involved in Candida albicans fluconazole resistance. Antimicrob. Agents Chemother. 40, 2835–2841.
Al-Dhaheri, R. S., and Douglas, L. J. (2008). Absence of amphotericin B-tolerant persister cells in biofilms of some Candida species. Antimicrob. Agents Chemother. 52, 1884–1887. doi: 10.1128/AAC.01473-07AAC.01473-07
Aleksandrowicz, E., and Herr, I. (2015). Ethical euthanasia and short-term anesthesia of the chick embryo. ALTEX 32, 143–147.
Alem, M. A., Oteef, M. D., Flowers, T. H., and Douglas, L. J. (2006). Production of tyrosol by Candida albicans biofilms and its role in quorum sensing and biofilm development. Eukaryot. Cell 5, 1770–1779. doi: 10.1128/EC.00219-06
Allaker, R. P. (2010). The use of nanoparticles to control oral biofilm formation. J. Dent. Res. 89, 1175–1186. doi: 10.1177/00220345103777940022034510377794
Allen, D., Wilson, D., Drew, R., and Perfect, J. (2015). Azole antifungals: 35 years of invasive fungal infection management. Expert Rev. Anti. Infect. Ther. 13, 787–798. doi: 10.1586/14787210.2015.1032939
Anderson, T. M., Clay, M. C., Cioffi, A. G., Diaz, K. A., Hisao, G. S., Tuttle, M. D., et al. (2014). Amphotericin forms an extramembranous and fungicidal sterol sponge. Nat. Chem. Biol. 10, 400–406. doi: 10.1038/nchembio.1496
Apidianakis, Y., Rahme, L. G., Heitman, J., Ausubel, F. M., Calderwood, S. B., and Mylonakis, E. (2004). Challenge of Drosophila melanogaster with Cryptococcus neoformans and role of the innate immune response. Eukaryot. Cell 3, 413–419. doi: 10.1128/EC.3.2.413-419.2004
Arendrup, M. C. (2014). Update on antifungal resistance in Aspergillus and Candida. Clin. Microbiol. Infect 20(Suppl. 6), 42–48. doi: 10.1111/1469-0691.12513
Arikan, S., and Rex, J. H. (2005). “Resistance to antifungal agents,” in Topley and Wilson’s Microbiology and Microbial Infec- tions: Medical Mycology, 10th Edn, eds W.G. Merz and R. J.Hay (London: Hodder Arnold), 168–181.
Atkinson, B. J., Lewis, R. E., and Kontoyiannis, D. P. (2008). Candida lusitaniae fungemia in cancer patients: risk factors for amphotericin B failure and outcome. Med. Mycol. 46, 541–546. doi: 10.1080/13693780801968571
Badali, H., Vaezi, A., Haghani, I., Yazdanparast, S. A., Hedayati, M. T., Mousavi, B., et al. (2013). Environmental study of azole-resistant Aspergillus fumigatus with TR34/L98H mutations in the cyp51A gene in Iran. Mycoses 56, 659–663. doi: 10.1111/myc.12089
Baginski, M., and Czub, J. (2009). Amphotericin B and its new derivatives – mode of action. Curr. Drug Metab. 10, 459–469. doi: 10.2174/138920009788898019
Baillie, G. S., and Douglas, L. J. (2000). Matrix polymers of Candida biofilms and their possible role in biofilm resistance to antifungal agents. J. Antimicrob. Chemother. 46, 397–403. doi: 10.1093/jac/46.3.397
Balashov, S. V., Park, S., and Perlin, D. S. (2006). Assessing resistance to the echinocandin antifungal drug caspofungin in Candida albicans by profiling mutations in FKS1. Antimicrob. Agents Chemother. 50, 2058–2063. doi: 10.1128/AAC.01653-05
Barchiesi, F., Schimizzi, A. M., Caselli, F., Novelli, A., Fallani, S., Giannini, D., et al. (2000). Interactions between triazoles and amphotericin B against Cryptococcus neoformans. Antimicrob. Agents Chemother. 44, 2435–2441. doi: 10.1128/AAC.44.9.2435-2441.2000
Basso, L. R., Gast, C. E., Bruzual, I., and Wong, B. (2015). Identification and properties of plasma membrane azole efflux pumps from the pathogenic fungi Cryptococcus gattii and Cryptococcus neoformans. J. Antimicrob. Chemother. 70, 1396–1407. doi: 10.1093/jac/dku554
Belanger, E. S., Yang, E., and Forrest, G. N. (2015). Combination antifungal therapy: when, where, and why. Curr. Clin. Microbiol. Rep. 2, 67–75. doi: 10.1007/s40588-015-0017-z
Benaducci, T., Sardi, J. E. C., Lourencetti, N. M., Scorzoni, L., Gullo, F. P., Rossi, S. A., et al. (2016). Virulence of Cryptococcus sp. Biofilms In Vitro and In Vivo using galleria mellonella as an alternative model. Front. Microbiol. 7:290. doi: 10.3389/fmicb.2016.00290
Bennett, J. E., Izumikawa, K., and Marr, K. A. (2004). Mechanism of increased fluconazole resistance in Candida glabrata during prophylaxis. Antimicrob. Agents Chemother. 48, 1773–1777. doi: 10.1128/AAC.48.5.1773-1777.2004
Biel, M. A. (2010). Photodynamic therapy of bacterial and fungal biofilm infections. Methods Mol. Biol. 635, 175–194. doi: 10.1007/978-1-60761-697-9_13
Bink, A., Kucharikova, S., Neirinck, B., Vleugels, J., Van Dijck, P., Cammue, B. P., et al. (2012). The nonsteroidal antiinflammatory drug diclofenac potentiates the in vivo activity of caspofungin against Candida albicans biofilms. J. Infect. Dis. 206, 1790–1797. doi: 10.1093/infdis/jis594jis594
Bink, A., Vandenbosch, D., Coenye, T., Nelis, H., Cammue, B. P., and Thevissen, K. (2011). Superoxide dismutases are involved in Candida albicans biofilm persistence against miconazole. Antimicrob. Agents Chemother. 55, 4033–4037. doi: 10.1128/AAC.00280-11AAC.00280-11
Bizerra, F. C., Nakamura, C. V., de Poersch, C., Estivalet Svidzinski, T. I., Borsato Quesada, R. M., Goldenberg, S., et al. (2008). Characteristics of biofilm formation by Candida tropicalis and antifungal resistance. FEMS Yeast Res. 8, 442–450. doi: 10.1111/j.1567-1364.2007.00347.xFYR347
Blankenship, J. R., Wormley, F. L., Boyce, M. K., Schell, W. A., Filler, S. G., Perfect, J. R., et al. (2003). Calcineurin is essential for Candida albicans survival in serum and virulence. Eukaryot. Cell 2, 422–430. doi: 10.1128/EC.2.3.422-430.2003
Blatzer, M., Blum, G., Jukic, E., Posch, W., Gruber, P., Nagl, M., et al. (2015). Blocking Hsp70 enhances the efficiency of amphotericin B treatment against resistant Aspergillus terreus strains. Antimicrob. Agents Chemother. 59, 3778–3788. doi: 10.1128/AAC.05164-14
Blum, G., Kainzner, B., Grif, K., Dietrich, H., Zelger, B., Sonnweber, T., et al. (2013). In vitro and in vivo role of heat shock protein 90 in Amphotericin B resistance of Aspergillus terreus. Clin. Microbiol. Infect. 19, 50–55. doi: 10.1111/j.1469-0691.2012.03848.x
Bondaryk, M., Kurzątkowski, W., and Staniszewska, M. (2013). Antifungal agents commonly used in the superficial and mucosal candidiasis treatment: mode of action and resistance development. Postepy Dermatol. Alergol. 30, 293–301. doi: 10.5114/pdia.2013.38358
Borecká-Melkusová, S., Moran, G. P., Sullivan, D. J., Kucharíková, S., Chorvát, D., and Bujdáková, H. (2009). The expression of genes involved in the ergosterol biosynthesis pathway in Candida albicans and Candida dubliniensis biofilms exposed to fluconazole. Mycoses 52, 118–128. doi: 10.1111/j.1439-0507.2008.01550.x
Borgers, M. (1980). Mechanism of action of antifungal drugs, with special reference to the imidazole derivatives. Rev. Infect. Dis. 2, 520–534. doi: 10.1093/clinids/2.4.520
Borowski, E. (2000). Novel approaches in the rational design of antifungal agents of low toxicity. Farmaco 55, 206–208. doi: 10.1016/S0014-827X(00)00024-0
Bosco-Borgeat, M. E., Mazza, M., Taverna, C. G., Córdoba, S., Murisengo, O. A., Vivot, W., et al. (2016). Amino acid substitution in Cryptococcus neoformans lanosterol 14-α-demethylase involved in fluconazole resistance in clinical isolates. Rev. Argent Microbiol. 48, 137–142. doi: 10.1016/j.ram.2016.03.003
Botet, J., Rodríguez-Mateos, M., Ballesta, J. P., Revuelta, J. L., and Remacha, M. (2008). A chemical genomic screen in Saccharomyces cerevisiae reveals a role for diphthamidation of translation elongation factor 2 in inhibition of protein synthesis by sordarin. Antimicrob. Agents Chemother. 52, 1623–1629. doi: 10.1128/AAC.01603-07
Bowman, S. M., and Free, S. J. (2006). The structure and synthesis of the fungal cell wall. Bioessays 28, 799–808. doi: 10.1002/bies.20441
Bozza, S., Gaziano, R., Lipford, G. B., Montagnoli, C., Bacci, A., Di Francesco, P., et al. (2002). Vaccination of mice against invasive aspergillosis with recombinant Aspergillus proteins and CpG oligodeoxynucleotides as adjuvants. Microbes Infect. 4, 1281–1290. doi: 10.1016/S1286-4579(02)00007-2
Breger, J., Fuchs, B. B., Aperis, G., Moy, T. I., Ausubel, F. M., and Mylonakis, E. (2007). Antifungal chemical compounds identified using a C. elegans pathogenicity assay. PLoS Pathog. 3:e18. doi: 10.1371/journal.ppat.0030018
Brothers, K. M., Newman, Z. R., and Wheeler, R. T. (2011). Live imaging of disseminated candidiasis in zebrafish reveals role of phagocyte oxidase in limiting filamentous growth. Eukaryot. Cell 10, 932–944. doi: 10.1128/EC.05005-11
Brown, A. J., Leach, M. D., and Nicholls, S. (2010). The relevance of heat shock regulation in fungal pathogens of humans. Virulence 1, 330–332. doi: 10.4161/viru.1.4.12364
Browne, N., Hackenberg, F., Streciwilk, W., Tacke, M., and Kavanagh, K. (2014). Assessment of in vivo antimicrobial activity of the carbene silver(I) acetate derivative SBC3 using Galleria mellonella larvae. Biometals 27, 745–752. doi: 10.1007/s10534-014-9766-z
Bugli, F., Cacaci, M., Martini, C., Torelli, R., Posteraro, B., Sanguinetti, M., et al. (2013). Human monoclonal antibody-based therapy in the treatment of invasive candidiasis. Clin. Dev. Immunol. 2013:403121. doi: 10.1155/2013/403121
Burnie, J. P., Carter, T. L., Hodgetts, S. J., and Matthews, R. C. (2006). Fungal heat-shock proteins in human disease. FEMS Microbiol. Rev. 30, 53–88. doi: 10.1111/j.1574-6976.2005.00001.x
Butts, A., Koselny, K., Chabrier-Roselló, Y., Semighini, C. P., Brown, J. C., Wang, X., et al. (2014). Estrogen receptor antagonists are anti-cryptococcal agents that directly bind EF hand proteins and synergize with fluconazole in vivo. MBio 5:e00765-13. doi: 10.1128/mBio.00765-13
Cannizzo, F. T., Eraso, E., Ezkurra, P. A., Villar-Vidal, M., Bollo, E., Castellá, G., et al. (2007). Biofilm development by clinical isolates of Malassezia pachydermatis. Med. Mycol. 45, 357–361. doi: 10.1080/13693780701225767
Cannon, R. D., Lamping, E., Holmes, A. R., Niimi, K., Baret, P. V., Keniya, M. V., et al. (2009). Efflux-mediated antifungal drug resistance. Clin. Microbiol. Rev. 22, 291–321. doi: 10.1128/CMR.00051-08
Cao, Y. Y., Cao, Y. B., Xu, Z., Ying, K., Li, Y., Xie, Y., et al. (2005). cDNA microarray analysis of differential gene expression in Candida albicans biofilm exposed to farnesol. Antimicrob. Agents Chemother. 49, 584–589. doi: 10.1128/AAC.49.2.584-589.2005
Carlson, R. P., Taffs, R., Davison, W. M., and Stewart, P. S. (2008). Anti-biofilm properties of chitosan-coated surfaces. J. Biomater. Sci. Polym. Ed. 19, 1035–1046. doi: 10.1163/156856208784909372
Cenci, E., Mencacci, A., Bacci, A., Bistoni, F., Kurup, V. P., and Romani, L. (2000). T cell vaccination in mice with invasive pulmonary aspergillosis. J. Immunol. 165, 381–388. doi: 10.4049/jimmunol.165.1.381
Cervelatti, E. P., Fachin, A. L., Ferreira-Nozawa, M. S., and Martinez-Rossi, N. M. (2006). Molecular cloning and characterization of a novel ABC transporter gene in the human pathogen Trichophyton rubrum. Med. Mycol. 44, 141–147. doi: 10.1080/13693780500220449
Chamilos, G., Lionakis, M. S., Lewis, R. E., Lopez-Ribot, J. L., Saville, S. P., Albert, N. D., et al. (2006). Drosophila melanogaster as a facile model for large-scale studies of virulence mechanisms and antifungal drug efficacy in Candida species. J. Infect. Dis. 193, 1014–1022. doi: 10.1086/500950
Chandra, J., Mukherjee, P. K., Leidich, S. D., Faddoul, F. F., Hoyer, L. L., Douglas, L. J., et al. (2001). Antifungal resistance of candidal biofilms formed on denture acrylic in vitro. J. Dent. Res. 80, 903–908. doi: 10.1177/00220345010800031101
Chandra, J., Patel, J. D., Li, J., Zhou, G., Mukherjee, P. K., McCormick, T. S., et al. (2005). Modification of surface properties of biomaterials influences the ability of Candida albicans to form biofilms. Appl. Environ. Microbiol. 71, 8795–8801. doi: 10.1128/AEM.71.12.8795-8801.2005
Chao, C. C., Hsu, P. C., Jen, C. F., Chen, I. H., Wang, C. H., Chan, H. C., et al. (2010). Zebrafish as a model host for Candida albicans infection. Infect. Immun. 78, 2512–2521. doi: 10.1128/IAI.01293-09
Chau, A. S., Gurnani, M., Hawkinson, R., Laverdiere, M., Cacciapuoti, A., and McNicholas, P. M. (2005). Inactivation of sterol Delta5,6-desaturase attenuates virulence in Candida albicans. Antimicrob. Agents Chemother. 49, 3646–3651. doi: 10.1128/AAC.49.9.3646-3651.2005
Chau, A. S., Mendrick, C. A., Sabatelli, F. J., Loebenberg, D., and McNicholas, P. M. (2004). Application of real-time quantitative PCR to molecular analysis of Candida albicans strains exhibiting reduced susceptibility to azoles. Antimicrob. Agents Chemother. 48, 2124–2131. doi: 10.1128/AAC.48.6.2124-2131.2004
Chen, S. H., Sheng, C. Q., Xu, X. H., Jiang, Y. Y., Zhang, W. N., and He, C. (2007). Identification of Y118 amino acid residue in Candida albicans sterol 14alpha-demethylase associated with the enzyme activity and selective antifungal activity of azole analogues. Biol. Pharm. Bull. 30, 1246–1253. doi: 10.1248/bpb.30.1246
Chen, X., Ren, B., Chen, M., Liu, M. X., Ren, W., Wang, Q. X., et al. (2014). ASDCD: antifungal synergistic drug combination database. PLoS ONE 9:e86499. doi: 10.1371/journal.pone.0086499
Chen, Y., Lu, Z., Zhao, J., Zou, Z., Gong, Y., Qu, F., et al. (2016). Epidemiology and molecular characterizations of azole resistance in clinical and environmental Aspergillus fumigatus isolates from China. Antimicrob. Agents Chemother. 60, 5878–5884. doi: 10.1128/AAC.01005-16
Chen, Y. L., Lehman, V. N., Lewit, Y., Averette, A. F., and Heitman, J. (2013). Calcineurin governs thermotolerance and virulence of Cryptococcus gattii. G3 (Bethesda) 3, 527–539. doi: 10.1534/g3.112.004242
Chen, Y. Z., Yang, Y. L., Chu, W. L., You, M. S., and Lo, H. J. (2015). Zebrafish egg infection model for studying Candida albicans adhesion factors. PLoS ONE 10:e0143048. doi: 10.1371/journal.pone.0143048
Cheng, S., Clancy, C. J., Nguyen, K. T., Clapp, W., and Nguyen, M. H. (2007). A Candida albicans petite mutant strain with uncoupled oxidative phosphorylation overexpresses MDR1 and has diminished susceptibility to fluconazole and voriconazole. Antimicrob. Agents Chemother. 51, 1855–1858. doi: 10.1128/AAC.00182-07
Chowdhary, A., Kathuria, S., Xu, J., Sharma, C., Sundar, G., Singh, P. K., et al. (2012). Clonal expansion and emergence of environmental multiple-triazole-resistant Aspergillus fumigatus strains carrying the TR34/L98H mutations in the cyp51A gene in India. PLoS ONE 7:e52871. doi: 10.1371/journal.pone.0052871
Chowdhary, A., Sharma, C., Hagen, F., and Meis, J. F. (2014a). Exploring azole antifungal drug resistance in Aspergillus fumigatus with special reference to resistance mechanisms. Future Microbiol. 9, 697–711. doi: 10.2217/fmb.14.27
Chowdhary, A., Sharma, C., van den Boom, M., Yntema, J. B., Hagen, F., Verweij, P. E., et al. (2014b). Multi-azole-resistant Aspergillus fumigatus in the environment in Tanzania. J. Antimicrob. Chemother. 69, 2979–2983. doi: 10.1093/jac/dku259
Clavijo-Giraldo, D. M., Matínez-Alvarez, J. A., Lopes-Bezerra, L. M., Ponce-Noyola, P., Franco, B., Almeida, R. S., et al. (2016). Analysis of Sporothrix schenckii sensu stricto and Sporothrix brasiliensis virulence in Galleria mellonella. J. Microbiol. Methods 122, 73–77. doi: 10.1016/j.mimet.2016.01.014
Coleman, J. J., Muhammed, M., Kasperkovitz, P. V., Vyas, J. M., and Mylonakis, E. (2011). Fusarium pathogenesis investigated using Galleria mellonella as a heterologous host. Fungal Biol. 115, 1279–1289. doi: 10.1016/j.funbio.2011.09.005
Colombo, A. L., Padovan, A. C., and Chaves, G. M. (2011). Current knowledge of Trichosporon spp. and Trichosporonosis. Clin. Microbiol. Rev. 24, 682–700. doi: 10.1128/CMR.00003-1124/4/682
Costa-Orlandi, C. B., Sardi, J. C., Santos, C. T., Fusco-Almeida, A. M., and Mendes-Giannini, M. J. (2014). in vitro characterization of Trichophyton rubrum and T. mentagrophytes biofilms. Biofouling 30, 719–727. doi: 10.1080/08927014.2014.919282
Coste, A., Selmecki, A., Forche, A., Diogo, D., Bougnoux, M. E., d’Enfert, C., et al. (2007). Genotypic evolution of azole resistance mechanisms in sequential Candida albicans isolates. Eukaryot. Cell 6, 1889–1904. doi: 10.1128/EC.00151-07
Cowen, L. E., Sanglard, D., Howard, S. J., Rogers, P. D., and Perlin, D. S. (2014). Mechanisms of antifungal drug resistance. Cold Spring Harb. Perspect. Med. 5:a019752. doi: 10.1101/cshperspect.a019752
Cushion, M. T., Collins, M. S., and Linke, M. J. (2009). Biofilm formation by Pneumocystis spp. Eukaryot. Cell 8, 197–206. doi: 10.1128/EC.00202-08EC.00202-08
da Matta, D. A., de Almeida, L. P., Machado, A. M., Azevedo, A. C., Kusano, E. J., Travassos, N. F., et al. (2007). Antifungal susceptibility of 1000 Candida bloodstream isolates to 5 antifungal drugs: results of a multicenter study conducted in São Paulo, Brazil, 1995-2003. Diagn. Microbiol. Infect. Dis. 57, 399–404. doi: 10.1016/j.diagmicrobio.2006.10.011
da Silva Ferreira, M. E., Capellaro, J. L., dos Reis Marques, E., Malavazi, I., Perlin, D., Park, S., et al. (2004). in vitro evolution of itraconazole resistance in Aspergillus fumigatus involves multiple mechanisms of resistance. Antimicrob. Agents Chemother. 48, 4405–4413. doi: 10.1128/AAC.48.11.4405-4413.2004
Davies, D. (2003). Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2, 114–122. doi: 10.1038/nrd1008nrd1008
Davis, L. E., Cook, G., and Costerton, J. W. (2002). Biofilm on ventriculo-peritoneal shunt tubing as a cause of treatment failure in coccidioidal meningitis. Emerg. Infect. Dis. 8, 376–379. doi: 10.3201/eid0804.010103
de Lacorte Singulani, J., Scorzoni, L., de Paula, E., Silva, A. C., Fusco-Almeida, A. M., and Mendes-Giannini, M. J. (2016). Evaluation of the efficacy of antifungal drugs against Paracoccidioides brasiliensis and Paracoccidioides lutzii in a Galleria mellonella model. Int. J. Antimicrob. Agents 48, 292–297. doi: 10.1016/j.ijantimicag.2016.05.012
de Melo, W. C., Avci, P., de Oliveira, M. N., Gupta, A., Vecchio, D., Sadasivam, M., et al. (2013). Photodynamic inactivation of biofilm: taking a lightly colored approach to stubborn infection. Expert Rev. Anti Infect. Ther. 11, 669–693. doi: 10.1586/14787210.2013.811861
Desalermos, A., Tan, X., Rajamuthiah, R., Arvanitis, M., Wang, Y., Li, D., et al. (2015). A multi-host approach for the systematic analysis of virulence factors in Cryptococcus neoformans. J. Infect. Dis. 211, 298–305. doi: 10.1093/infdis/jiu441
Desnos-Ollivier, M., Bretagne, S., Raoux, D., Hoinard, D., Dromer, F., Dannaoui, E., et al. (2008). Mutations in the fks1 gene in Candida albicans, C. tropicalis, and C. krusei correlate with elevated caspofungin MICs uncovered in AM3 medium using the method of the European Committee on Antibiotic Susceptibility Testing. Antimicrob. Agents Chemother. 52, 3092–3098. doi: 10.1128/AAC.00088-08
Devi, S. J., Schneerson, R., Egan, W., Ulrich, T. J., Bryla, D., Robbins, J. B., et al. (1991). Cryptococcus neoformans serotype A glucuronoxylomannan-protein conjugate vaccines: synthesis, characterization, and immunogenicity. Infect. Immun. 59, 3700–3707.
Di Bonaventura, G., Pompilio, A., Picciani, C., Iezzi, M., D’Antonio, D., and Piccolomini, R. (2006). Biofilm formation by the emerging fungal pathogen Trichosporon asahii: development, architecture, and antifungal resistance. Antimicrob. Agents Chemother. 50, 3269–3276. doi: 10.1128/AAC.00556-06
Diaz-Guerra, T. M., Mellado, E., Cuenca-Estrella, M., and Rodriguez-Tudela, J. L. (2003). A point mutation in the 14alpha-sterol demethylase gene cyp51A contributes to itraconazole resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 47, 1120–1124. doi: 10.1128/AAC.47.3.1120-1124.2003
Dogsa, I., Brloznik, M., Stopar, D., and Mandic-Mulec, I. (2013). Exopolymer diversity and the role of levan in Bacillus subtilis biofilms. PLoS ONE 8:e62044. doi: 10.1371/journal.pone.0062044
Dunkel, N., Liu, T. T., Barker, K. S., Homayouni, R., Morschhäuser, J., and Rogers, P. D. (2008). A gain-of-function mutation in the transcription factor Upc2p causes upregulation of ergosterol biosynthesis genes and increased fluconazole resistance in a clinical Candida albicans isolate. Eukaryot. Cell 7, 1180–1190. doi: 10.1128/EC.00103-08
Edwards, J. E. (2012). Fungal cell wall vaccines: an update. J. Med. Microbiol. 61(Pt. 7), 895–903. doi: 10.1099/jmm.0.041665-0
Elefanti, A., Mouton, J. W., Verweij, P. E., Tsakris, A., Zerva, L., and Meletiadis, J. (2013). Amphotericin B- and voriconazole-echinocandin combinations against Aspergillus spp.: effect of serum on inhibitory and fungicidal interactions. Antimicrob. Agents Chemother. 57, 4656–4663. doi: 10.1128/AAC.00597-13
Enjalbert, B., and Whiteway, M. (2005). Release from quorum-sensing molecules triggers hyphal formation during Candida albicans resumption of growth. Eukaryot. Cell 4, 1203–1210. doi: 10.1128/EC.4.7.1203-1210.2005
Espinel-Ingroff, A., Pfaller, M. A., Bustamante, B., Canton, E., Fothergill, A., Fuller, J., et al. (2014). Multilaboratory study of epidemiological cutoff values for detection of resistance in eight Candida species to fluconazole, posaconazole, and voriconazole. Antimicrob. Agents Chemother. 58, 2006–2012. doi: 10.1128/AAC.02615-13
Estrela, A. B., Heck, M. G., and Abraham, W. R. (2009). Novel approaches to control biofilm infections. Curr. Med. Chem. 16, 1512–1530. doi: 10.2174/092986709787909640
Ewbank, J. J., and Zugasti, O. (2011). C. elegans: model host and tool for antimicrobial drug discovery. Dis. Model. Mech. 4, 300–304. doi: 10.1242/dmm.006684
Fachin, A. L., Ferreira-Nozawa, M. S., Maccheroni, W., and Martinez-Rossi, N. M. (2006). Role of the ABC transporter TruMDR2 in terbinafine, 4-nitroquinoline N-oxide and ethidium bromide susceptibility in Trichophyton rubrum. J. Med. Microbiol. 55(Pt 8), 1093–1099. doi: 10.1099/jmm.0.46522-0
Fanning, S., and Mitchell, A. P. (2012). Fungal biofilms. PLoS Pathog. 8:e1002585. doi: 10.1371/journal.ppat.1002585PPATHOGENS-D-11-02842
Favel, A., Michel-Nguyen, A., Peyron, F., Martin, C., Thomachot, L., Datry, A., et al. (2003). Colony morphology switching of Candida lusitaniae and acquisition of multidrug resistance during treatment of a renal infection in a newborn: case report and review of the literature. Diagn. Microbiol. Infect. Dis. 47, 331–339. doi: 10.1016/S0732-8893(03)00094-4
Fernandes, V. C., Coitinho, J. B., Veloso, J. M., Araújo, S. A., Pedroso, E. P., and Goes, A. M. (2011). Combined use of Paracoccidioides brasiliensis recombinant rPb27 and rPb40 antigens in an enzyme-linked immunosorbent assay for immunodiagnosis of paracoccidioidomycosis. J. Immunol. Methods 367, 78–84. doi: 10.1016/j.jim.2011.02.006
Ferreira, G. F., Baltazar, L. E. M., Santos, J. R., Monteiro, A. S., Fraga, L. A., Resende-Stoianoff, M. A., et al. (2013). The role of oxidative and nitrosative bursts caused by azoles and amphotericin B against the fungal pathogen Cryptococcus gattii. J. Antimicrob. Chemother. 68, 1801–1811. doi: 10.1093/jac/dkt114
Figueredo, L. A., Cafarchia, C., Desantis, S., and Otranto, D. (2012). Biofilm formation of Malassezia pachydermatis from dogs. Vet. Microbiol. 160, 126–131. doi: 10.1016/j.vetmic.2012.05.012
Finkel, J. S., and Mitchell, A. P. (2011). Genetic control of Candida albicans biofilm development. Nat. Rev. Microbiol. 9, 109–118. doi: 10.1038/nrmicro2475nrmicro2475
Flemming, H. C., and Wingender, J. (2010). The biofilm matrix. Nat. Rev. Microbiol. 8, 623–633. doi: 10.1038/nrmicro2415nrmicro2415
Franz, R., Kelly, S. L., Lamb, D. C., Kelly, D. E., Ruhnke, M., and Morschhäuser, J. (1998). Multiple molecular mechanisms contribute to a stepwise development of fluconazole resistance in clinical Candida albicans strains. Antimicrob. Agents Chemother. 42, 3065–3072.
Frenkel, M., Mandelblat, M., Alastruey-Izquierdo, A., Mendlovic, S., Semis, R., and Segal, E. (2016). Pathogenicity of Candida albicans isolates from bloodstream and mucosal candidiasis assessed in mice and Galleria mellonella. J. Mycol. Med. 26, 1–8. doi: 10.1016/j.mycmed.2015.12.006
Fuchs, B. B., Eby, J., Nobile, C. J., El Khoury, J. B., Mitchell, A. P., and Mylonakis, E. (2010a). Role of filamentation in Galleria mellonella killing by Candida albicans. Microbes Infect. 12, 488–496. doi: 10.1016/j.micinf.2010.03.001
Fuchs, B. B., O’Brien, E., Khoury, J. B., and Mylonakis, E. (2010b). Methods for using Galleria mellonella as a model host to study fungal pathogenesis. Virulence 1, 475–482. doi: 10.4161/viru.1.6.12985
Gao, Y., Li, H., Liu, S., Zhang, X., and Sun, S. (2014). Synergistic effect of fluconazole and doxycycline against Candida albicans biofilms resulting from calcium fluctuation and downregulation of fluconazole-inducible efflux pump gene overexpression. J. Med. Microbiol. 63(Pt. 7), 956–961. doi: 10.1099/jmm.0.072421-0jmm.0.072421-0
Gao, Y., Zhang, C., Lu, C., Liu, P., Li, Y., Li, H., et al. (2013). Synergistic effect of doxycycline and fluconazole against Candida albicans biofilms and the impact of calcium channel blockers. FEMS Yeast Res. 13, 453–462. doi: 10.1111/1567-1364.12048
Garcia-Effron, G., Chua, D. J., Tomada, J. R., DiPersio, J., Perlin, D. S., Ghannoum, M., et al. (2010). Novel FKS mutations associated with echinocandin resistance in Candida species. Antimicrob. Agents Chemother. 54, 2225–2227. doi: 10.1128/AAC.00998-09
Garcia-Effron, G., Katiyar, S. K., Park, S., Edlind, T. D., and Perlin, D. S. (2008). A naturally occurring proline-to-alanine amino acid change in Fks1p in Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis accounts for reduced echinocandin susceptibility. Antimicrob. Agents Chemother. 52, 2305–2312. doi: 10.1128/AAC.00262-08
Garcia-Effron, G., Lee, S., Park, S., Cleary, J. D., and Perlin, D. S. (2009). Effect of Candida glabrata FKS1 and FKS2 mutations on echinocandin sensitivity and kinetics of 1,3-beta-D-glucan synthase: implication for the existing susceptibility breakpoint. Antimicrob. Agents Chemother. 53, 3690–3699. doi: 10.1128/AAC.00443-09
Gaur, M., Puri, N., Manoharlal, R., Rai, V., Mukhopadhayay, G., Choudhury, D., et al. (2008). MFS transportome of the human pathogenic yeast Candida albicans. BMC Genomics 9:579. doi: 10.1186/1471-2164-9-579
Gee, S. F., Joly, S., Soll, D. R., Meis, J. F., Verweij, P. E., Polacheck, I., et al. (2002). Identification of four distinct genotypes of Candida dubliniensis and detection of microevolution in vitro and in vivo. J. Clin. Microbiol. 40, 556–574. doi: 10.1128/JCM.40.2.556-574.2002
Ghannoum, M. (2016). Azole resistance in dermatophytes: prevalence and mechanism of action. J. Am. Podiatr. Med. Assoc. 106, 79–86. doi: 10.7547/14-109
Goldman, G. H., da Silva, Ferreira, M. E., dos Reis, Marques, E., Savoldi, M., et al. (2004). Evaluation of fluconazole resistance mechanisms in Candida albicans clinical isolates from HIV-infected patients in Brazil. Diagn. Microbiol. Infect. Dis. 50, 25–32. doi: 10.1016/j.diagmicrobio.2004.04.009
Gonçalves, S. S., Souza, A. C., Chowdhary, A., Meis, J. F., and Colombo, A. L. (2016). Epidemiology and molecular mechanisms of antifungal resistance in Candida and Aspergillus. Mycoses doi: 10.1111/myc.12469 [Epub ahead of print].
Grossman, N. T., Pham, C. D., Cleveland, A. A., and Lockhart, S. R. (2015). Molecular mechanisms of fluconazole resistance in Candida parapsilosis isolates from a U.S. surveillance system. Antimicrob. Agents Chemother. 59, 1030–1037. doi: 10.1128/AAC.04613-14
Gu, W., Guo, D., Zhang, L., Xu, D., and Sun, S. (2016). Synergistic effect of azoles and fluoxetine against resistant Candida albicans attributed to attenuating fungal virulence. Antimicrob. Agents Chemother. 60, 6179–6188. doi: 10.1128/AAC.03046-15
Guinea, J., Sánchez-Somolinos, M., Cuevas, O., Peláez, T., and Bouza, E. (2006). Fluconazole resistance mechanisms in Candida krusei: the contribution of efflux-pumps. Med. Mycol. 44, 575–578. doi: 10.1080/13693780600561544
Gupta, A. K., and Tomas, E. (2003). New antifungal agents. Dermatol. Clin. 21, 565–576. doi: 10.1016/S0733-8635(03)00024-X
Hiller, D., Sanglard, D., and Morschhäuser, J. (2006). Overexpression of the MDR1 gene is sufficient to confer increased resistance to toxic compounds in Candida albicans. Antimicrob. Agents Chemother. 50, 1365–1371. doi: 10.1128/AAC.50.4.1365-1371.2006
Hinrichsen, S. L., Falcão, E., Vilella, T. A., Colombo, A. L., Nucci, M., Moura, L., et al. (2008). Candidemia in a tertiary hospital in northeastern Brazil. Rev. Soc. Bras. Med. Trop. 41, 394–398. doi: 10.1590/S0037-86822008000400014
Hole, C. R., and Wormley, F. L. (2012). Vaccine and immunotherapeutic approaches for the prevention of cryptococcosis: lessons learned from animal models. Front. Microbiol. 3:291. doi: 10.3389/fmicb.2012.00291
Hope, W. W., Tabernero, L., Denning, D. W., and Anderson, M. J. (2004). Molecular mechanisms of primary resistance to flucytosine in Candida albicans. Antimicrob. Agents Chemother. 48, 4377–4386. doi: 10.1128/AAC.48.11.4377-4386.2004
Hornby, J. M., Jensen, E. C., Lisec, A. D., Tasto, J. J., Jahnke, B., Shoemaker, R., et al. (2001). Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl. Environ. Microbiol. 67, 2982–2992. doi: 10.1128/AEM.67.7.2982-2992.2001
Howard, S. J., Cerar, D., Anderson, M. J., Albarrag, A., Fisher, M. C., Pasqualotto, A. C., et al. (2009). Frequency and evolution of Azole resistance in Aspergillus fumigatus associated with treatment failure. Emerg. Infect. Dis. 15, 1068–1076. doi: 10.3201/eid1507.090043
Hsieh, S. H., Lin, J. S., Huang, J. H., Wu, S. Y., Chu, C. L., Kung, J. T., et al. (2011). Immunization with apoptotic phagocytes containing Histoplasma capsulatum activates functional CD8(+) T cells to protect against histoplasmosis. Infect. Immun. 79, 4493–4502. doi: 10.1128/IAI.05350-11
Huang, W., Liao, G., Baker, G. M., Wang, Y., Lau, R., Paderu, P., et al. (2016). Lipid flippase subunit Cdc50 mediates drug resistance and virulence in Cryptococcus neoformans. MBio 7:e00478-16. doi: 10.1128/mBio.00478-16
Ito, J. I., and Lyons, J. M. (2002). Vaccination of corticosteroid immunosuppressed mice against invasive pulmonary aspergillosis. J. Infect. Dis. 186, 869–871. doi: 10.1086/342509
Jacob, T. R., Peres, N. T., Martins, M. P., Lang, E. A., Sanches, P. R., Rossi, A., et al. (2015). Heat shock protein 90 (Hsp90) as a molecular target for the development of novel drugs against the dermatophyte Trichophyton rubrum. Front. Microbiol. 6:1241. doi: 10.3389/fmicb.2015.01241
Jacobsen, I. D., Grosse, K., Berndt, A., and Hube, B. (2011). Pathogenesis of Candida albicans infections in the alternative chorio-allantoic membrane chicken embryo model resembles systemic murine infections. PLoS ONE 6:e19741. doi: 10.1371/journal.pone.0019741
Jacobsen, I. D., Grosse, K., Slesiona, S., Hube, B., Berndt, A., and Brock, M. (2010). Embryonated eggs as an alternative infection model to investigate Aspergillus fumigatus virulence. Infect. Immun. 78, 2995–3006. doi: 10.1128/IAI.00268-10
Jensen-Pergakes, K. L., Kennedy, M. A., Lees, N. D., Barbuch, R., Koegel, C., and Bard, M. (1998). Sequencing, disruption, and characterization of the Candida albicans sterol methyltransferase (ERG6) gene: drug susceptibility studies in erg6 mutants. Antimicrob. Agents Chemother. 42, 1160–1167.
Jia, N., Arthington-Skaggs, B., Lee, W., Pierson, C. A., Lees, N. D., Eckstein, J., et al. (2002). Candida albicans sterol C-14 reductase, encoded by the ERG24 gene, as a potential antifungal target site. Antimicrob. Agents Chemother. 46, 947–957. doi: 10.1128/AAC.46.4.947-957.2002
Jiang, C., Dong, D., Yu, B., Cai, G., Wang, X., Ji, Y., et al. (2013). Mechanisms of azole resistance in 52 clinical isolates of Candida tropicalis in China. J. Antimicrob. Chemother. 68, 778–785. doi: 10.1093/jac/dks481
Johnson, C. H., Ayyadevara, S., McEwen, J. E., and Shmookler Reis, R. J. (2009). Histoplasma capsulatum and Caenorhabditis elegans: a simple nematode model for an innate immune response to fungal infection. Med. Mycol. 47, 808–813. doi: 10.3109/13693780802660532
Johnson, M. D., MacDougall, C., Ostrosky-Zeichner, L., Perfect, J. R., and Rex, J. H. (2004). Combination antifungal therapy. Antimicrob. Agents Chemother. 48, 693–715. doi: 10.1128/AAC.48.3.693-715.2004
Justice, M. C., Hsu, M. J., Tse, B., Ku, T., Balkovec, J., Schmatz, D., et al. (1998). Elongation factor 2 as a novel target for selective inhibition of fungal protein synthesis. J. Biol. Chem. 273, 3148–3151. doi: 10.1074/jbc.273.6.3148
Juvvadi, P. R., Lee, S. C., Heitman, J., and Steinbach, W. J. (2016). Calcineurin in fungal virulence and drug resistance: prospects for harnessing targeted inhibition of calcineurin for an antifungal therapeutic approach. Virulence doi: 10.1080/21505594.2016.1201250 [Epub ahead of print].
Kaerger, K., Schwartze, V. U., Dolatabadi, S., Nyilasi, I., Kovács, S. A., Binder, U., et al. (2015). Adaptation to thermotolerance in Rhizopus coincides with virulence as revealed by avian and invertebrate infection models, phylogeny, physiological and metabolic flexibility. Virulence 6, 395–403. doi: 10.1080/21505594.2015.1029219
Kanafani, Z. A., and Perfect, J. R. (2008). Antimicrobial resistance: resistance to antifungal agents: mechanisms and clinical impact. Clin. Infect. Dis. 46, 120–128. doi: 10.1086/524071
Kathiravan, M. K., Salake, A. B., Chothe, A. S., Dudhe, P. B., Watode, R. P., Mukta, M. S., et al. (2012). The biology and chemistry of antifungal agents: a review. Bioorg. Med. Chem. 20, 5678–5698. doi: 10.1016/j.bmc.2012.04.045S0968-0896(12)00329-X
Katiyar, S. K., and Edlind, T. D. (2001). Identification and expression of multidrug resistance-related ABC transporter genes in Candida krusei. Med. Mycol. 39, 109–116. doi: 10.1080/mmy.39.1.109.116
Katiyar, S. K., and Edlind, T. D. (2009). Role for Fks1 in the intrinsic echinocandin resistance of Fusarium solani as evidenced by hybrid expression in Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 53, 1772–1778. doi: 10.1128/AAC.00020-09
Katragkou, A., McCarthy, M., Meletiadis, J., Petraitis, V., Moradi, P. W., Strauss, G. E., et al. (2014). In vitro combination of isavuconazole with micafungin or amphotericin B deoxycholate against medically important molds. Antimicrob. Agents Chemother. 58, 6934–6937. doi: 10.1128/AAC.03261-14
Kaur, S., and Singh, S. (2014). Biofilm formation by Aspergillus fumigatus. Med. Mycol. 52, 2–9. doi: 10.3109/13693786.2013.81959213693786.2013.819592
Kelly, S. L., Lamb, D. C., Kelly, D. E., Manning, N. J., Loeffler, J., Hebart, H., et al. (1997). Resistance to fluconazole and cross-resistance to amphotericin B in Candida albicans from AIDS patients caused by defective sterol delta5,6-desaturation. FEBS Lett. 400, 80–82. doi: 10.1016/S0014-5793(96)01360-9
Kelly, S. L., Lamb, D. C., Taylor, M., Corran, A. J., Baldwin, B. C., and Powderly, W. G. (1994). Resistance to amphotericin B associated with defective sterol delta 8–>7 isomerase in a Cryptococcus neoformans strain from an AIDS patient. FEMS Microbiol. Lett. 122, 39–42. doi: 10.1111/j.1574-6968.1994.tb07140.x
Keniya, M. V., Fleischer, E., Klinger, A., Cannon, R. D., and Monk, B. C. (2015). Inhibitors of the Candida albicans major facilitator superfamily transporter Mdr1p responsible for fluconazole resistance. PLoS ONE 10:e0126350. doi: 10.1371/journal.pone.0126350
Khot, P. D., Suci, P. A., Miller, R. L., Nelson, R. D., and Tyler, B. J. (2006). A small subpopulation of blastospores in Candida albicans biofilms exhibit resistance to amphotericin B associated with differential regulation of ergosterol and beta-1,6-glucan pathway genes. Antimicrob. Agents Chemother. 50, 3708–3716. doi: 10.1128/AAC.00997-06
Kirkpatrick, W. R., Perea, S., Coco, B. J., and Patterson, T. F. (2002). Efficacy of caspofungin alone and in combination with voriconazole in a Guinea pig model of invasive aspergillosis. Antimicrob. Agents Chemother. 46, 2564–2568. doi: 10.1128/AAC.46.8.2564-2568.2002
Kucharikova, S., Sharma, N., Spriet, I., Maertens, J., Van Dijck, P., and Lagrou, K. (2013). Activities of systemically administered echinocandins against in vivo mature Candida albicans biofilms developed in a rat subcutaneous model. Antimicrob. Agents Chemother. 57, 2365–2368. doi: 10.1128/AAC.02288-12AAC.02288-12
Kuhn, D. M., George, T., Chandra, J., Mukherjee, P. K., and Ghannoum, M. A. (2002). Antifungal susceptibility of Candida biofilms: unique efficacy of amphotericin B lipid formulations and echinocandins. Antimicrob. Agents Chemother. 46, 1773–1780. doi: 10.1128/AAC.46.6.1773-1780.2002
LaFleur, M. D., Kumamoto, C. A., and Lewis, K. (2006). Candida albicans biofilms produce antifungal-tolerant persister cells. Antimicrob. Agents Chemother. 50, 3839–3846. doi: 10.1128/AAC.00684-06
Lai, Y. W., Campbell, L. T., Wilkins, M. R., Pang, C. N., Chen, S., and Carter, D. A. (2016). Synergy and antagonism between iron chelators and antifungal drugs in Cryptococcus. Int. J. Antimicrob. Agents 48, 388–394. doi: 10.1016/j.ijantimicag.2016.06.012
Lamaris, G. A., Chamilos, G., Lewis, R. E., and Kontoyiannis, D. P. (2007). Virulence studies of Scedosporium and Fusarium species in Drosophila melanogaster. J. Infect. Dis. 196, 1860–1864. doi: 10.1086/523765
Lamb, D. C., Kelly, D. E., White, T. C., and Kelly, S. L. (2000). The R467K amino acid substitution in Candida albicans sterol 14alpha-demethylase causes drug resistance through reduced affinity. Antimicrob. Agents Chemother. 44, 63–67. doi: 10.1128/AAC.44.1.63-67.2000
Lamping, E., Ranchod, A., Nakamura, K., Tyndall, J. D., Niimi, K., Holmes, A. R., et al. (2009). Abc1p is a multidrug efflux transporter that tips the balance in favor of innate azole resistance in Candida krusei. Antimicrob. Agents Chemother. 53, 354–369. doi: 10.1128/AAC.01095-08
Lass-Florl, C., Speth, C., Kofler, G., Dierch, M. P., Gunsilius, E., and Wurzner, R. (2003). Effect of increasing inoculum sizes of Aspergillus hyphae on MICs and MFCs of antifungal agents by broth microdilution method. Int. J. Antimicrob. Agents 21, 229–233. doi: 10.1016/S0924-8579(02)00189-9
Leber, R., Fuchsbichler, S., Klobucníková, V., Schweighofer, N., Pitters, E., Wohlfarter, K., et al. (2003). Molecular mechanism of terbinafine resistance in Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 47, 3890–3900. doi: 10.1128/AAC.47.12.3890-3900.2003
Leonardelli, F., Macedo, D., Dudiuk, C., Cabeza, M. S., Gamarra, S., and Garcia-Effron, G. (2016). Aspergillus fumigatus intrinsic fluconazole resistance is due to the naturally occurring T301I substitution in Cyp51Ap. Antimicrob. Agents Chemother. 60, 5420–5426. doi: 10.1128/AAC.00905-16
Lewis, K. (2001). Riddle of biofilm resistance. Antimicrob. Agents Chemother. 45, 999–1007. doi: 10.1128/AAC.45.4.999-1007.2001
Lewis, K. (2007). Persister cells, dormancy and infectious disease. Nat. Rev. Microbiol. 5, 48–56. doi: 10.1038/nrmicro1557
Lewis, K. (2010). Persister cells. Annu. Rev. Microbiol. 64, 357–372. doi: 10.1146/annurev.micro.112408.134306
Lewis, R. E., Ben-Ami, R., Best, L., Albert, N., Walsh, T. J., and Kontoyiannis, D. P. (2013). Tacrolimus enhances the potency of posaconazole against Rhizopus oryzae in vitro and in an experimental model of mucormycosis. J. Infect. Dis. 207, 834–841. doi: 10.1093/infdis/jis767
Lionakis, M. S., and Kontoyiannis, D. P. (2012). Drosophila melanogaster as a model organism for invasive aspergillosis. Methods Mol. Biol. 845, 455–468. doi: 10.1007/978-1-61779-539-8_32
Liu, N., Wang, C., Su, H., Zhang, W., and Sheng, C. (2016). Strategies in the discovery of novel antifungal scaffolds. Future Med Chem 8, 1435–1454. doi: 10.4155/fmc-2016-0020
Liu, S., Yue, L., Gu, W., Li, X., Zhang, L., and Sun, S. (2016). Synergistic Effect of Fluconazole and Calcium Channel Blockers against Resistant Candida albicans. PLoS ONE 11:e0150859. doi: 10.1371/journal.pone.0150859
Lockhart, S. R., Frade, J. P., Etienne, K. A., Pfaller, M. A., Diekema, D. J., and Balajee, S. A. (2011). Azole resistance in Aspergillus fumigatus isolates from the ARTEMIS global surveillance study is primarily due to the TR/L98H mutation in the cyp51A gene. Antimicrob. Agents Chemother. 55, 4465–4468. doi: 10.1128/AAC.00185-11
Lopez-Ribot, J. L., McAtee, R. K., Lee, L. N., Kirkpatrick, W. R., White, T. C., Sanglard, D., et al. (1998). Distinct patterns of gene expression associated with development of fluconazole resistance in serial candida albicans isolates from human immunodeficiency virus-infected patients with oropharyngeal candidiasis. Antimicrob. Agents Chemother. 42, 2932–2937.
Louie, A., Kaw, P., Banerjee, P., Liu, W., Chen, G., and Miller, M. H. (2001). Impact of the order of initiation of fluconazole and amphotericin B in sequential or combination therapy on killing of Candida albicans In Vitro and in a rabbit model of endocarditis and pyelonephritis. Antimicrob. Agents Chemother. 45, 485–494. doi: 10.1128/AAC.45.2.485-494.2001
MacRae, C. A., and Peterson, R. T. (2015). Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 14, 721–731. doi: 10.1038/nrd4627
Maertens, J. A. (2004). History of the development of azole derivatives. Clin. Microbiol. Infect. 10(Suppl. 1), 1–10. doi: 10.1111/j.1470-9465.2004.00841.x
Maligie, M. A., and Selitrennikoff, C. P. (2005). Cryptococcus neoformans resistance to echinocandins: (1,3)beta-glucan synthase activity is sensitive to echinocandins. Antimicrob. Agents Chemother. 49, 2851–2856. doi: 10.1128/AAC.49.7.2851-2856.2005
Mann, P. A., Parmegiani, R. M., Wei, S. Q., Mendrick, C. A., Li, X., Loebenberg, D., et al. (2003). Mutations in Aspergillus fumigatus resulting in reduced susceptibility to posaconazole appear to be restricted to a single amino acid in the cytochrome P450 14alpha-demethylase. Antimicrob. Agents Chemother. 47, 577–581. doi: 10.1128/AAC.47.2.577-581.2003
Marques, A. F., da Silva, M. B., Juliano, M. A., Munhõz, J. E., Travassos, L. R., and Taborda, C. P. (2008). Additive effect of P10 immunization and chemotherapy in anergic mice challenged intratracheally with virulent yeasts of Paracoccidioides brasiliensis. Microbes Infect. 10, 1251–1258. doi: 10.1016/j.micinf.2008.07.027
Marques, A. F., da Silva, M. B., Juliano, M. A., Travassos, L. R., and Taborda, C. P. (2006). Peptide immunization as an adjuvant to chemotherapy in mice challenged intratracheally with virulent yeast cells of Paracoccidioides brasiliensis. Antimicrob. Agents Chemother. 50, 2814–2819. doi: 10.1128/AAC.00220-06
Marr, K. A., Schlamm, H. T., Herbrecht, R., Rottinghaus, S. T., Bow, E. J., Cornely, O. A., et al. (2015). Combination antifungal therapy for invasive aspergillosis: a randomized trial. Ann. Intern. Med. 162, 81–89. doi: 10.7326/M13-2508
Martel, C. M., Parker, J. E., Bader, O., Weig, M., Gross, U., Warrilow, A. G., et al. (2010). Identification and characterization of four azole-resistant erg3 mutants of Candida albicans. Antimicrob. Agents Chemother. 54, 4527–4533. doi: 10.1128/AAC.00348-10
Martinez, L. R., and Casadevall, A. (2007). Cryptococcus neoformans biofilm formation depends on surface support and carbon source and reduces fungal cell susceptibility to heat, cold, and UV light. Appl. Environ. Microbiol. 73, 4592–4601. doi: 10.1128/AEM.02506-06
Martins, M., Uppuluri, P., Thomas, D. P., Cleary, I. A., Henriques, M., Lopez-Ribot, J. L., et al. (2010). Presence of extracellular DNA in the Candida albicans biofilm matrix and its contribution to biofilms. Mycopathologia 169, 323–331. doi: 10.1007/s11046-009-9264-y
Martins, M. P., Franceschini, A. C., Jacob, T. R., Rossi, A., and Martinez-Rossi, N. M. (2016). Compensatory expression of multidrug-resistance genes encoding ABC transporters in dermatophytes. J. Med. Microbiol. 65, 605–610. doi: 10.1099/jmm.0.000268
Matthews, R. C., Rigg, G., Hodgetts, S., Carter, T., Chapman, C., Gregory, C., et al. (2003). Preclinical assessment of the efficacy of mycograb, a human recombinant antibody against fungal HSP90. Antimicrob. Agents Chemother. 47, 2208–2216. doi: 10.1128/AAC.47.7.2208-2216.2003
Medici, N. P., and Del Poeta, M. (2015). New insights on the development of fungal vaccines: from immunity to recent challenges. Mem. Inst. Oswaldo Cruz 110, 966–973. doi: 10.1590/0074-02760150335
Mesa-Arango, A. C., Scorzoni, L., and Zaragoza, O. (2012). It only takes one to do many jobs: amphotericin B as antifungal and immunomodulatory drug. Front. Microbiol. 3:286. doi: 10.3389/fmicb.2012.00286
Mesa-Arango, A. C., Trevijano-Contador, N., Román, E., Sánchez-Fresneda, R., Casas, C., Herrero, E., et al. (2014). The production of reactive oxygen species is a universal action mechanism of Amphotericin B against pathogenic yeasts and contributes to the fungicidal effect of this drug. Antimicrob. Agents Chemother. 58, 6627–6638. doi: 10.1128/AAC.03570-14
Minari, A., Hachem, R., and Raad, I. (2001). Candida lusitaniae: a cause of breakthrough fungemia in cancer patients. Clin. Infect. Dis. 32, 186–190. doi: 10.1086/318473
Mitchell, K. F., Taff, H. T., Cuevas, M. A., Reinicke, E. L., Sanchez, H., and Andes, D. R. (2013). Role of matrix β-1,3 glucan in antifungal resistance of non-albicans Candida biofilms. Antimicrob. Agents Chemother. 57, 1918–1920. doi: 10.1128/AAC.02378-12Sz
Moazeni, M., Kelidari, H. R., Saeedi, M., Morteza-Semnani, K., Nabili, M., Gohar, A. A., et al. (2016). Time to overcome fluconazole resistant Candida isolates: solid lipid nanoparticles as a novel antifungal drug delivery system. Colloids Surf. B Biointerfaces 142, 400–407. doi: 10.1016/j.colsurfb.2016.03.013
Mor, V., Farnoud, A. M., Singh, A., Rella, A., Tanno, H., Ishii, K., et al. (2016). Glucosylceramide administration as a vaccination strategy in mouse models of Cryptococcosis. PLoS ONE 11:e0153853. doi: 10.1371/journal.pone.0153853
Moralez, A. T., Perini, H. F., Furlaneto-Maia, L., Almeida, R. S., Panagio, L. A., and Furlaneto, M. C. (2016). Phenotypic switching of Candida tropicalis is associated with cell damage in epithelial cells and virulence in Galleria mellonella model. Virulence 7, 379–386. doi: 10.1080/21505594.2016.1140297
Moran, G. P., Sanglard, D., Donnelly, S. M., Shanley, D. B., Sullivan, D. J., and Coleman, D. C. (1998). Identification and expression of multidrug transporters responsible for fluconazole resistance in Candida dubliniensis. Antimicrob. Agents Chemother. 42, 1819–1830.
Morio, F., Loge, C., Besse, B., Hennequin, C., and Le Pape, P. (2010). Screening for amino acid substitutions in the Candida albicans Erg11 protein of azole-susceptible and azole-resistant clinical isolates: new substitutions and a review of the literature. Diagn. Microbiol. Infect. Dis. 66, 373–384. doi: 10.1016/j.diagmicrobio.2009.11.006
Morio, F., Pagniez, F., Lacroix, C., Miegeville, M., and Le Pape, P. (2012). Amino acid substitutions in the Candida albicans sterol Δ5,6-desaturase (Erg3p) confer azole resistance: characterization of two novel mutants with impaired virulence. J. Antimicrob. Chemother. 67, 2131–2138. doi: 10.1093/jac/dks186
Morozov, A. A., and Likhoshway, Y. V. (2016). Evolutionary history of the chitin synthases of eukaryotes. Glycobiology 26, 635–639. doi: 10.1093/glycob/cww018
Morschhäuser, J. (2002). The genetic basis of fluconazole resistance development in Candida albicans. Biochim. Biophys. Acta 1587, 240–248. doi: 10.1016/S0925-4439(02)00087-X
Movahed, E., Tan, G. M., Munusamy, K., Yeow, T. C., Tay, S. T., Wong, W. F., et al. (2016). Triclosan demonstrates synergic effect with amphotericin B and fluconazole and Induces apoptosis-like cell death in Cryptococcus neoformans. Front. Microbiol. 7:360. doi: 10.3389/fmicb.2016.00360
Mowat, E., Williams, C., Jones, B., McChlery, S., and Ramage, G. (2009). The characteristics of Aspergillus fumigatus mycetoma development: is this a biofilm? Med. Mycol. 47(Suppl. 1), S120–S126. doi: 10.1080/13693780802238834
Muhammed, M., Arvanitis, M., and Mylonakis, E. (2016). Whole animal HTS of small molecules for antifungal compounds. Expert Opin. Drug Discov. 11, 177–184. doi: 10.1517/17460441.2016.1122591
Mukherjee, P. K., Chandra, J., Kuhn, D. M., and Ghannoum, M. A. (2003). Mechanism of fluconazole resistance in Candida albicans biofilms: phase-specific role of efflux pumps and membrane sterols. Infect. Immun. 71, 4333–4340. doi: 10.1128/IAI.71.8.4333-4340.2003
Mukherjee, P. K., Long, L., Kim, H. G., and Ghannoum, M. A. (2009). Amphotericin B lipid complex is efficacious in the treatment of Candida albicans biofilms using a model of catheter-associated Candida biofilms. Int. J. Antimicrob. Agents 33, 149–153. doi: 10.1016/j.ijantimicag.2008.07.030S0924-8579(08)00367-1
Nailis, H., Vandenbosch, D., Deforce, D., Nelis, H. J., and Coenye, T. (2010). Transcriptional response to fluconazole and amphotericin B in Candida albicans biofilms. Res. Microbiol. 161, 284–292. doi: 10.1016/j.resmic.2010.02.004
Nett, J. E., Crawford, K., Marchillo, K., and Andes, D. R. (2010a). Role of Fks1p and matrix glucan in Candida albicans biofilm resistance to an echinocandin, pyrimidine, and polyene. Antimicrob. Agents Chemother. 54, 3505–3508. doi: 10.1128/AAC.00227-10AAC.00227-10
Nett, J. E., Lepak, A. J., Marchillo, K., and Andes, D. R. (2009). Time course global gene expression analysis of an in vivo Candida biofilm. J. Infect. Dis. 200, 307–313. doi: 10.1086/599838
Nett, J. E., Sanchez, H., Cain, M. T., and Andes, D. R. (2010b). Genetic basis of Candida biofilm resistance due to drug-sequestering matrix glucan. J. Infect. Dis. 202, 171–175. doi: 10.1086/651200
Ngo, H. X., Garneau-Tsodikova, S., and Green, K. D. (2016). A complex game of hide and seek: the search for new antifungals. Medchemcomm 7, 1285–1306. doi: 10.1039/C6MD00222F
Niemirowicz, K., Durnaś, B., Tokajuk, G., Głuszek, K., Wilczewska, A. Z., Misztalewska, I., et al. (2016). Magnetic nanoparticles as a drug delivery system that enhance fungicidal activity of polyene antibiotics. Nanomedicine 12, 2395–2404. doi: 10.1016/j.nano.2016.07.006
Odds, F. C., Brown, A. J., and Gow, N. A. (2003). Antifungal agents: mechanisms of action. Trends Microbiol. 11, 272–279. doi: 10.1016/S0966-842X(03)00117-3
Odom, A. R. (2014). The triphenylethylenes, a novel class of antifungals. MBio 5, e1126–e1114. doi: 10.1128/mBio.01126-14
Okoli, I., Coleman, J. J., Tampakakis, E., Tempakakis, E., An, W. F., Holson, E., et al. (2009). Identification of antifungal compounds active against Candida albicans using an improved high-throughput Caenorhabditis elegans assay. PLoS ONE 4:e7025. doi: 10.1371/journal.pone.0007025
Pagano, L., Cornely, O. A., Busca, A., Caira, M., Cesaro, S., Gasbarrino, C., et al. (2013). Combined antifungal approach for the treatment of invasive mucormycosis in patients with hematologic diseases: a report from the SEIFEM and FUNGISCOPE registries. Haematologica 98, e127–e130. doi: 10.3324/haematol.2012.083063
Paião, F. G., Segato, F., Cursino-Santos, J. R., Peres, N. T., and Martinez-Rossi, N. M. (2007). Analysis of Trichophyton rubrum gene expression in response to cytotoxic drugs. FEMS Microbiol. Lett. 271, 180–186. doi: 10.1111/j.1574-6968.2007.00710.x
Papon, N., Noël, T., Florent, M., Gibot-Leclerc, S., Jean, D., Chastin, C., et al. (2007). Molecular mechanism of flucytosine resistance in Candida lusitaniae: contribution of the FCY2, FCY1, and FUR1 genes to 5-fluorouracil and fluconazole cross-resistance. Antimicrob. Agents Chemother. 51, 369–371. doi: 10.1128/AAC.00824-06
Park, S., Kelly, R., Kahn, J. N., Robles, J., Hsu, M. J., Register, E., et al. (2005). Specific substitutions in the echinocandin target Fks1p account for reduced susceptibility of rare laboratory and clinical Candida sp. isolates. Antimicrob. Agents Chemother. 49, 3264–3273. doi: 10.1128/AAC.49.8.3264-3273.2005
Partridge, B. M., Athar, M. A., and Winner, H. I. (1971). Chick embryo inoculation as a pathogenicity test for Candida species. J. Clin. Pathol. 24, 645–648. doi: 10.1136/jcp.24.7.645
Perea, S., López-Ribot, J. L., Kirkpatrick, W. R., McAtee, R. K., Santillán, R. A., Martínez, M., et al. (2001). Prevalence of molecular mechanisms of resistance to azole antifungal agents in Candida albicans strains displaying high-level fluconazole resistance isolated from human immunodeficiency virus-infected patients. Antimicrob. Agents Chemother. 45, 2676–2684. doi: 10.1128/AAC.45.10.2676-2684.2001
Perea, S., López-Ribot, J. L., Wickes, B. L., Kirkpatrick, W. R., Dib, O. P., Bachmann, S. P., et al. (2002). Molecular mechanisms of fluconazole resistance in Candida dubliniensis isolates from human immunodeficiency virus-infected patients with oropharyngeal candidiasis. Antimicrob. Agents Chemother. 46, 1695–1703. doi: 10.1128/AAC.46.6.1695-1703.2002
Perfect, J. R., Dismukes, W. E., Dromer, F., Goldman, D. L., Graybill, J. R., Hamill, R. J., et al. (2010). Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of america. Clin. Infect. Dis. 50, 291–322. doi: 10.1086/649858
Perlin, D. S. (2007). Resistance to echinocandin-class antifungal drugs. Drug Resist. Updat. 10, 121–130. doi: 10.1016/j.drup.2007.04.002
Perlin, D. S. (2015). Echinocandin resistance in Candida. Clin. Infect. Dis. 61(Suppl. 6), S612–S617. doi: 10.1093/cid/civ791
Perlroth, J., Choi, B., and Spellberg, B. (2007). Nosocomial fungal infections: epidemiology, diagnosis, and treatment. Med. Mycol. 45, 321–346. doi: 10.1080/13693780701218689
Perumal, P., Mekala, S., and Chaffin, W. L. (2007). Role for cell density in antifungal drug resistance in Candida albicans biofilms. Antimicrob. Agents Chemother. 51, 2454–2463. doi: 10.1128/AAC.01237-06
Pfaller, M. A., and Diekema, D. J. (2007). Epidemiology of invasive candidiasis: a persistent public health problem. Clin. Microbiol. Rev. 20, 133–163. doi: 10.1128/CMR.00029-06
Pham, C. D., Bolden, C. B., Kuykendall, R. J., and Lockhart, S. R. (2014). Development of a Luminex-based multiplex assay for detection of mutations conferring resistance to Echinocandins in Candida glabrata. J. Clin. Microbiol. 52, 790–795. doi: 10.1128/JCM.03378-13
Pinjon, E., Moran, G. P., Jackson, C. J., Kelly, S. L., Sanglard, D., Coleman, D. C., et al. (2003). Molecular mechanisms of itraconazole resistance in Candida dubliniensis. Antimicrob. Agents Chemother. 47, 2424–2437. doi: 10.1128/AAC.47.8.2424-2437.2003
Pires, R. H., Santos, J. M., Zaia, J. E., Martins, C. H., and Mendes-Giannini, M. J. (2011). Candida parapsilosis complex water isolates from a haemodialysis unit: biofilm production and in vitro evaluation of the use of clinical antifungals. Mem. Inst. Oswaldo Cruz 106, 646–654. doi: 10.1590/S0074-02762011000600002
Pitangui, N. S., Sardi, J. C., Silva, J. F., Benaducci, T., Moraes da Silva, R. A., Rodríguez-Arellanes, G., et al. (2012). Adhesion of Histoplasma capsulatum to pneumocytes and biofilm formation on an abiotic surface. Biofouling 28, 711–718. doi: 10.1080/08927014.2012.703659
Plaine, A., Walker, L., Da Costa, G., Mora-Montes, H. M., McKinnon, A., Gow, N. A., et al. (2008). Functional analysis of Candida albicans GPI-anchored proteins: roles in cell wall integrity and caspofungin sensitivity. Fungal Genet. Biol. 45, 1404–1414. doi: 10.1016/j.fgb.2008.08.003
Polak, A., and Scholer, H. J. (1975). Mode of action of 5-fluorocytosine and mechanisms of resistance. Chemotherapy 21, 113–130. doi: 10.1159/000221854
Posteraro, B., Sanguinetti, M., Sanglard, D., La Sorda, M., Boccia, S., Romano, L., et al. (2003). Identification and characterization of a Cryptococcus neoformans ATP binding cassette (ABC) transporter-encoding gene, CnAFR1, involved in the resistance to fluconazole. Mol. Microbiol. 47, 357–371.
Prasad, R., and Goffeau, A. (2012). Yeast ATP-binding cassette transporters conferring multidrug resistance. Annu. Rev. Microbiol. 66, 39–63. doi: 10.1146/annurev-micro-092611-150111
Prasad, R., and Rawal, M. K. (2014). Efflux pump proteins in antifungal resistance. Front. Pharmacol. 5:202. doi: 10.3389/fphar.2014.00202
Pukkila-Worley, R., Peleg, A. Y., Tampakakis, E., and Mylonakis, E. (2009). Candida albicans hyphal formation and virulence assessed using a Caenorhabditis elegans infection model. Eukaryot. Cell 8, 1750–1758. doi: 10.1128/EC.00163-09
Raad, I. I., Zakhem, A. E., Helou, G. E., Jiang, Y., Kontoyiannis, D. P., and Hachem, R. (2015). Clinical experience of the use of voriconazole, caspofungin or the combination in primary and salvage therapy of invasive aspergillosis in haematological malignancies. Int. J. Antimicrob. Agents 45, 283–288. doi: 10.1016/j.ijantimicag.2014.08.012
Rajendran, R., Mowat, E., McCulloch, E., Lappin, D. F., Jones, B., Lang, S., et al. (2011). Azole resistance of Aspergillus fumigatus biofilms is partly associated with efflux pump activity. Antimicrob. Agents Chemother. 55, 2092–2097. doi: 10.1128/AAC.01189-10AAC.01189-10
Ramage, G., Bachmann, S., Patterson, T. F., Wickes, B. L., and Lopez-Ribot, J. L. (2002). Investigation of multidrug efflux pumps in relation to fluconazole resistance in Candida albicans biofilms. J. Antimicrob. Chemother. 49, 973–980. doi: 10.1093/jac/dkf049
Ramage, G., Jose, A., Sherry, L., Lappin, D. F., Jones, B., and Williams, C. (2013). Liposomal amphotericin B displays rapid dose-dependent activity against Candida albicans biofilms. Antimicrob. Agents Chemother. 57, 2369–2371. doi: 10.1128/AAC.02344-12AAC.02344-12
Ramage, G., Mowat, E., Jones, B., Williams, C., and Lopez-Ribot, J. (2009). Our current understanding of fungal biofilms. Crit. Rev. Microbiol. 35, 340–355. doi: 10.3109/10408410903241436
Ramage, G., Rajendran, R., Sherry, L., and Williams, C. (2012). Fungal biofilm resistance. Int. J. Microbiol. 2012:528521. doi: 10.1155/2012/528521
Ramage, G., Vande Walle, K., Wickes, B. L., and Lopez-Ribot, J. L. (2001). Standardized method for in vitro antifungal susceptibility testing of Candida albicans biofilms. Antimicrob. Agents Chemother. 45, 2475–2479. doi: 10.1128/AAC.45.9.2475-2479.2001
Ramage, G., Wickes, B. L., and Lopez-Ribot, J. L. (2008). A seed and feed model for the formation of Candida albicans biofilms under flow conditions using an improved modified Robbins device. Rev. Iberoam. Micol. 25, 37–40. doi: 10.1016/S1130-1406(08)70009-3
Ramasamy, M., Lee, J. H., and Lee, J. (2016). Potent antimicrobial and antibiofilm activities of bacteriogenically synthesized gold-silver nanoparticles against pathogenic bacteria and their physiochemical characterizations. J. Biomater. Appl. 31, 366–378. doi: 10.1177/08853282166469100885328216646910
Ramírez-Camejo, L. A., Torres-Ocampo, A. P., Agosto-Rivera, J. L., and Bayman, P. (2014). An opportunistic human pathogen on the fly: strains of Aspergillus flavus vary in virulence in Drosophila melanogaster. Med. Mycol. 52, 211–219. doi: 10.1093/mmy/myt008
Raska, M., Beláková, J., Wudattu, N. K., Kafková, L., Rùzicková, K., Sebestová, M., et al. (2005). Comparison of protective effect of protein and DNA vaccines hsp90 in murine model of systemic candidiasis. Folia Microbiol. (Praha) 50, 77–82. doi: 10.1007/BF02931297
Redding, S. W., Kirkpatrick, W. R., Coco, B. J., Sadkowski, L., Fothergill, A. W., Rinaldi, M. G., et al. (2002). Candida glabrata oropharyngeal candidiasis in patients receiving radiation treatment for head and neck cancer. J. Clin. Microbiol. 40, 1879–1881. doi: 10.1128/JCM.40.5.1879-1881.2002
Rella, A., Mor, V., Farnoud, A. M., Singh, A., Shamseddine, A. A., Ivanova, E., et al. (2015). Role of Sterylglucosidase 1 (Sgl1) on the pathogenicity of Cryptococcus neoformans: potential applications for vaccine development. Front. Microbiol. 6:836. doi: 10.3389/fmicb.2015.00836
Ricardo, E., Miranda, I. M., Faria-Ramos, I., Silva, R. M., Rodrigues, A. G., and Pina-Vaz, C. (2014). In vivo and in vitro acquisition of resistance to voriconazole by Candida krusei. Antimicrob. Agents Chemother. 58, 4604–4611. doi: 10.1128/AAC.02603-14
Rittner, G. M., Muñoz, J. E., Marques, A. F., Nosanchuk, J. D., Taborda, C. P., and Travassos, L. R. (2012). Therapeutic DNA vaccine encoding peptide P10 against experimental paracoccidioidomycosis. PLoS Negl. Trop. Dis. 6:e1519. doi: 10.1371/journal.pntd.0001519
Rocha, E. M., Garcia-Effron, G., Park, S., and Perlin, D. S. (2007). A Ser678Pro substitution in Fks1p confers resistance to echinocandin drugs in Aspergillus fumigatus. Antimicrob. Agents Chemother. 51, 4174–4176. doi: 10.1128/AAC.00917-07
Rodero, L., Mellado, E., Rodriguez, A. C., Salve, A., Guelfand, L., Cahn, P., et al. (2003). G484S amino acid substitution in lanosterol 14-alpha demethylase (ERG11) is related to fluconazole resistance in a recurrent Cryptococcus neoformans clinical isolate. Antimicrob. Agents Chemother. 47, 3653–3656. doi: 10.1128/AAC.47.11.3653-3656.2003
Rogers, T. R. (2006). Antifungal drug resistance: limited data, dramatic impact? Int. J. Antimicrob. Agents 27(Suppl. 1), 7–11. doi: 10.1016/j.ijantimicag.2006.03.012
Rossignol, T., Ding, C., Guida, A., d’Enfert, C., Higgins, D. G., and Butler, G. (2009). Correlation between biofilm formation and the hypoxic response in Candida parapsilosis. Eukaryot. Cell 8, 550–559. doi: 10.1128/EC.00350-08EC.00350-08
Rundfeldt, C., Steckel, H., Scherliess, H., Wyska, E., and Wlaź, P. (2013). Inhalable highly concentrated itraconazole nanosuspension for the treatment of bronchopulmonary aspergillosis. Eur. J. Pharm. Biopharm. 83, 44–53. doi: 10.1016/j.ejpb.2012.09.018
Russell, W., and Burch, R. (1959). The Principles of Humane Experimental Technique. London: Methuen Publishing.
Sangalli-Leite, F., Scorzoni, L., Alves de Paula, E., Silva, A. C., da Silva, J. F., de Oliveira, H. C., et al. (2016). Synergistic effect of pedalitin and amphotericin B against Cryptococcus neoformans by in vitro and in vivo evaluation. Int. J. Antimicrob. Agents 48, 504–511. doi: 10.1016/j.ijantimicag.2016.07.025
Sangalli-Leite, F., Scorzoni, L., Cecilia Mesa-Arango, A., Casas, C., Herrero, E., Mendes Gianinni, M. J. S., et al. (2011). Amphotericin B mediates killing in Cryptococcus neoformans through the induction of a strong oxidative burst. Microbes Infect. 13, 457–467. doi: 10.1016/j.micinf.2011.01.015
Sanglard, D. (2002). Resistance of human fungal pathogens to antifungal drugs. Curr. Opin. Microbiol. 5, 379–385. doi: 10.1016/S1369-5274(02)00344-2
Sanglard, D. (2016). Emerging threats in antifungal-resistant fungal pathogens. Front. Med. (Lausanne) 3:11. doi: 10.3389/fmed.2016.00011
Sanglard, D., Coste, A., and Ferrari, S. (2009). Antifungal drug resistance mechanisms in fungal pathogens from the perspective of transcriptional gene regulation. FEMS Yeast Res. 9, 1029–1050. doi: 10.1111/j.1567-1364.2009.00578.x
Sanglard, D., Ischer, F., Calabrese, D., Majcherczyk, P. A., and Bille, J. (1999). The ATP binding cassette transporter gene CgCDR1 from Candida glabrata is involved in the resistance of clinical isolates to azole antifungal agents. Antimicrob. Agents Chemother. 43, 2753–2765.
Sanglard, D., Ischer, F., Koymans, L., and Bille, J. (1998). Amino acid substitutions in the cytochrome P-450 lanosterol 14alpha-demethylase (CYP51A1) from azole-resistant Candida albicans clinical isolates contribute to resistance to azole antifungal agents. Antimicrob. Agents Chemother. 42, 241–253.
Sanglard, D., Ischer, F., Monod, M., and Bille, J. (1997). Cloning of Candida albicans genes conferring resistance to azole antifungal agents: characterization of CDR2, a new multidrug ABC transporter gene. Microbiology 143(Pt 2), 405–416. doi: 10.1099/00221287-143-2-405
Sanglard, D., Kuchler, K., Ischer, F., Pagani, J. L., Monod, M., and Bille, J. (1995). Mechanisms of resistance to azole antifungal agents in Candida albicans isolates from AIDS patients involve specific multidrug transporters. Antimicrob. Agents Chemother. 39, 2378–2386. doi: 10.1128/AAC.39.11.2378
Sanglard, D., and Odds, F. C. (2002). Resistance of Candida species to antifungal agents: molecular mechanisms and clinical consequences. Lancet Infect. Dis. 2, 73–85. doi: 10.1016/S1473-3099(02)00181-0
Sanguinetti, M., Posteraro, B., Fiori, B., Ranno, S., Torelli, R., and Fadda, G. (2005). Mechanisms of azole resistance in clinical isolates of Candida glabrata collected during a hospital survey of antifungal resistance. Antimicrob. Agents Chemother. 49, 668–679. doi: 10.1128/AAC.49.2.668-679.2005
Sanguinetti, M., Posteraro, B., La Sorda, M., Torelli, R., Fiori, B., Santangelo, R., et al. (2006). Role of AFR1, an ABC transporter-encoding gene, in the in vivo response to fluconazole and virulence of Cryptococcus neoformans. Infect. Immun. 74, 1352–1359. doi: 10.1128/IAI.74.2.1352-1359.2006
Santos, E., and Levitz, S. M. (2014). Fungal vaccines and immunotherapeutics. Cold Spring Harb. Perspect. Med. 4:a019711. doi: 10.1101/cshperspect.a019711
Sardi, J. C., Gullo, F. P., Freires, I. A., Pitangui, N. S., Segalla, M. P., Fusco-Almeida, A. M., et al. (2016). Synthesis, antifungal activity of caffeic acid derivative esters, and their synergism with fluconazole and nystatin against Candida spp. Diagn. Microbiol. Infect. Dis. 86, 387–391. doi: 10.1016/j.diagmicrobio.2016.08.002
Sardi, J. E. C., Pitangui, N. E. S., Voltan, A. R., Braz, J. D., Machado, M. P., Fusco Almeida, A. M., et al. (2015). In vitro Paracoccidioides brasiliensis biofilm and gene expression of adhesins and hydrolytic enzymes. Virulence 6, 642–651. doi: 10.1080/21505594.2015.1031437
Sato, M. R., da Silva, P. B., de Souza, R. A., dos Santos, K. C., and Chorilli, M. (2015). Recent advances in nanoparticle carriers for coordination complexes. Curr. Top. Med. Chem 15, 287–297. doi: 10.2174/1568026615666150108145614
Saville, S. P., Lazzell, A. L., Monteagudo, C., and Lopez-Ribot, J. L. (2003). Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection. Eukaryot. Cell 2, 1053–1060. doi: 10.1128/EC.2.5.1053-1060.2003
Scheid, L. A., Mario, D. A., Kubiça, T. F., Santurio, J. M., and Alves, S. H. (2012). In vitro activities of antifungal agents alone and in combination against fluconazole-susceptible and -resistant strains of Candida dubliniensis. Braz. J. Infect. Dis. 16, 78–81. doi: 10.1016/S1413-8670(12)70279-9
Scorzoni, L., de Lucas, M. P., Mesa-Arango, A. C., Fusco-Almeida, A. M., Lozano, E., Cuenca-Estrella, M., et al. (2013). Antifungal efficacy during Candida krusei infection in non-conventional models correlates with the yeast in vitro susceptibility profile. PLoS ONE 8:e60047. doi: 10.1371/journal.pone.0060047
Scorzoni, L., de Paula, E., Silva, A. C., Singulani, J. L., Leite, F. S., de Oliveira, H. C., et al. (2015). Comparison of virulence between Paracoccidioides brasiliensis and Paracoccidioides lutzii using Galleria mellonella as a host model. Virulence 6, 766–776. doi: 10.1080/21505594.2015.1085277
Selmecki, A., Forche, A., and Berman, J. (2006). Aneuploidy and isochromosome formation in drug-resistant Candida albicans. Science 313, 367–370. doi: 10.1126/science.1128242
Semighini, C. P., Hornby, J. M., Dumitru, R., Nickerson, K. W., and Harris, S. D. (2006). Farnesol-induced apoptosis in Aspergillus nidulans reveals a possible mechanism for antagonistic interactions between fungi. Mol. Microbiol. 59, 753–764. doi: 10.1111/j.1365-2958.2005.04976.x
Semighini, C. P., Murray, N., and Harris, S. D. (2008). Inhibition of Fusarium graminearum growth and development by farnesol. FEMS Microbiol. Lett. 279, 259–264. doi: 10.1111/j.1574-6968.2007.01042.xFML1042
Seneviratne, C. J., Jin, L., and Samaranayake, L. P. (2008). Biofilm lifestyle of Candida: a mini review. Oral Dis. 14, 582–590. doi: 10.1111/j.1601-0825.2007.01424.xODI1424
Seyedmousavi, S., Hashemi, S. J., Zibafar, E., Zoll, J., Hedayati, M. T., Mouton, J. W., et al. (2013). Azole-resistant Aspergillus fumigatus. Iran. Emerg. Infect. Dis. 19, 832–834. doi: 10.3201/eid1905.130075
Shahid, S. K. (2016). Newer patents in antimycotic therapy. Pharm. Pat. Anal. 5, 115–134. doi: 10.4155/ppa-2015-0001
Shalini, K., Kumar, N., Drabu, S., and Sharma, P. K. (2011). Advances in synthetic approach to and antifungal activity of triazoles. Beilstein J. Org. Chem. 7, 668–677. doi: 10.3762/bjoc.7.79
Sheikh, N., Jahagirdar, V., Kothadia, S., and Nagoba, B. (2013). Antifungal drug resistance in Candida species. Eur. J. Gen. Med. 10, 254–258.
Shekhar-Guturja, T., Tebung, W. A., Mount, H., Liu, N., Köhler, J. R., Whiteway, M., et al. (2016). Beauvericin potentiates azole activity via inhibition of multidrug efflux, blocks C. albicans morphogenesis, and is effluxed via Yor1 and circuitry controlled by Zcf29. Antimicrob. Agents Chemother. 60, 7468–7480. doi: 10.1128/AAC.01959-16
Sheng, C., and Zhang, W. (2011). New lead structures in antifungal drug discovery. Curr. Med. Chem. 18, 733–766. doi: 10.2174/092986711794480113
Shibasaki, S., Aoki, W., Nomura, T., Miyoshi, A., Tafuku, S., Sewaki, T., et al. (2013). An oral vaccine against candidiasis generated by a yeast molecular display system. Pathog. Dis. 69, 262–268. doi: 10.1111/2049-632X.12068
Shinde, R. B., Chauhan, N. M., Raut, J. S., and Karuppayil, S. M. (2012). Sensitization of Candida albicans biofilms to various antifungal drugs by cyclosporine A. Ann. Clin. Microbiol. Antimicrob. 11:27. doi: 10.1186/1476-0711-11-271476-0711-11-27
Shukla, P. K., Singh, P., Yadav, R. K., Pandey, S., and Bhunia, S. S. (2016). Past, Present, and Future of Antifungal Drug Development. Berlin: Springer.
Singh, R., Shivaprakash, M. R., and Chakrabarti, A. (2011). Biofilm formation by zygomycetes: quantification, structure and matrix composition. Microbiology 157(Pt 9), 2611–2618. doi: 10.1099/mic.0.048504-0
Sionov, E., Chang, Y. C., Garraffo, H. M., Dolan, M. A., Ghannoum, M. A., and Kwon-Chung, K. J. (2012). Identification of a Cryptococcus neoformans cytochrome P450 lanosterol 14α-demethylase (Erg11) residue critical for differential susceptibility between fluconazole/voriconazole and itraconazole/posaconazole. Antimicrob. Agents Chemother. 56, 1162–1169. doi: 10.1128/AAC.05502-11
Sionov, E., Lee, H., Chang, Y. C., and Kwon-Chung, K. J. (2010). Cryptococcus neoformans overcomes stress of azole drugs by formation of disomy in specific multiple chromosomes. PLoS Pathog. 6:e1000848. doi: 10.1371/journal.ppat.1000848
Slaven, J. W., Anderson, M. J., Sanglard, D., Dixon, G. K., Bille, J., Roberts, I. S., et al. (2002). Increased expression of a novel Aspergillus fumigatus ABC transporter gene, atrF, in the presence of itraconazole in an itraconazole resistant clinical isolate. Fungal Genet. Biol. 36, 199–206. doi: 10.1016/S1087-1845(02)00016-6
Soares, L. A., Gullo, F. P., Sardi, J. E. C., Pitangui, N. E. S., Costa-Orlandi, C. B., Sangalli-Leite, F., et al. (2014). Anti-trichophyton activity of protocatechuates and their synergism with fluconazole. Evid. Based Complement. Alternat. Med. 2014:957860. doi: 10.1155/2014/957860
Sokol-Anderson, M. L., Brajtburg, J., and Medoff, G. (1986). Amphotericin B-induced oxidative damage and killing of Candida albicans. J. Infect. Dis. 154, 76–83. doi: 10.1093/infdis/154.1.76
Song, J., Zhai, P., Zhang, Y., Zhang, C., Sang, H., Han, G., et al. (2016). The Aspergillus fumigatus damage resistance protein family coordinately regulates ergosterol biosynthesis and azole susceptibility. MBio 7:e01919-15. doi: 10.1128/mBio.01919-15
Song, J. C., and Stevens, D. A. (2016). Caspofungin: pharmacodynamics, pharmacokinetics, clinical uses and treatment outcomes. Crit. Rev. Microbiol. 42, 813–846. doi: 10.3109/1040841X.2015.1068271
Song, J. W., Shin, J. H., Kee, S. J., Kim, S. H., Shin, M. G., Suh, S. P., et al. (2009). Expression of CgCDR1, CgCDR2, and CgERG11 in Candida glabrata biofilms formed by bloodstream isolates. Med. Mycol. 47, 545–548. doi: 10.1080/13693780802210726795279371
Song, M. H., Lee, J. W., Kim, M. S., Yoon, J. K., White, T. C., Floyd, A., et al. (2012). A flucytosine-responsive Mbp1/Swi4-like protein, Mbs1, plays pleiotropic roles in antifungal drug resistance, stress response, and virulence of Cryptococcus neoformans. Eukaryot. Cell 11, 53–67. doi: 10.1128/EC.05236-11
Souza, A. C., Fuchs, B. B., Pinhati, H. M., Siqueira, R. A., Hagen, F., Meis, J. F., et al. (2015). Candida parapsilosis resistance to fluconazole: molecular mechanisms and in vivo impact in infected Galleria mellonella larvae. Antimicrob. Agents Chemother. 59, 6581–6587. doi: 10.1128/AAC.01177-15
Spellberg, B. (2011). Vaccines for invasive fungal infections. F1000 Med. Rep. 3:13. doi: 10.3410/M3-13
Steinbach, W. J., Cramer, R. A., Perfect, B. Z., Asfaw, Y. G., Sauer, T. C., Najvar, L. K., et al. (2006). Calcineurin controls growth, morphology, and pathogenicity in Aspergillus fumigatus. Eukaryot. Cell 5, 1091–1103. doi: 10.1128/EC.00139-06
Strähle, U., Scholz, S., Geisler, R., Greiner, P., Hollert, H., Rastegar, S., et al. (2012). Zebrafish embryos as an alternative to animal experiments–a commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 33, 128–132. doi: 10.1016/j.reprotox.2011.06.121
Sun, J., Li, Z., Chu, H., Guo, J., Jiang, G., and Qi, Q. (2016). Candida albicans amphotericin B-tolerant persister formation is closely related to surface adhesion. Mycopathologia 181, 41–49. doi: 10.1007/s11046-015-9894-110.1007/s11046-015-9894-1
Sundriyal, S., Sharma, R. K., and Jain, R. (2006). Current advances in antifungal targets and drug development. Curr. Med. Chem. 13, 1321–1335. doi: 10.2174/092986706776873023
Sutherland, I. (2001). Biofilm exopolysaccharides: a strong and sticky framework. Microbiology 147(Pt 1), 3–9. doi: 10.1099/00221287-147-1-3
Szlinder-Richert, J., Mazerski, J., Cybulska, B., Grzybowska, J., and Borowski, E. (2001). MFAME, N-methyl-N-D-fructosyl amphotericin B methyl ester, a new amphotericin B derivative of low toxicity: relationship between self-association and effects on red blood cells. Biochim. Biophys. Acta 1528, 15–24. doi: 10.1016/S0304-4165(01)00166-0
Taborda, C. P., Juliano, M. A., Puccia, R., Franco, M., and Travassos, L. R. (1998). Mapping of the T-cell epitope in the major 43-kilodalton glycoprotein of Paracoccidioides brasiliensis which induces a Th-1 response protective against fungal infection in BALB/c mice. Infect. Immun. 66, 786–793.
Taff, H. T., Nett, J. E., Zarnowski, R., Ross, K. M., Sanchez, H., Cain, M. T., et al. (2012). A Candida biofilm-induced pathway for matrix glucan delivery: implications for drug resistance. PLoS Pathog. 8:e1002848. doi: 10.1371/journal.ppat.1002848
Taraszkiewicz, A., Fila, G., Grinholc, M., and Nakonieczna, J. (2013). Innovative strategies to overcome biofilm resistance. Biomed. Res. Int. 2013:150653. doi: 10.1155/2013/150653
Tatsumi, Y., Nagashima, M., Shibanushi, T., Iwata, A., Kangawa, Y., Inui, F., et al. (2013). Mechanism of action of efinaconazole, a novel triazole antifungal agent. Antimicrob. Agents Chemother. 57, 2405–2409. doi: 10.1128/AAC.02063-12
Tenor, J. L., Oehlers, S. H., Yang, J. L., Tobin, D. M., and Perfect, J. R. (2015). Live imaging of host-parasite interactions in a zebrafish infection model reveals cryptococcal determinants of virulence and central nervous system invasion. MBio 6:e01425-15. doi: 10.1128/mBio.01425-15
Thomaz, L., García-Rodas, R., Guimarães, A. J., Taborda, C. P., Zaragoza, O., and Nosanchuk, J. D. (2013). Galleria mellonella as a model host to study Paracoccidioides lutzii and Histoplasma capsulatum. Virulence 4, 139–146. doi: 10.4161/viru.23047
Trevijano-Contador, N., and Zaragoza, O. (2014). Expanding the use of alternative models to investigate novel aspects of immunity to microbial pathogens. Virulence 5, 454–456. doi: 10.4161/viru.28775
Troskie, A. M., Rautenbach, M., Delattin, N., Vosloo, J. A., Dathe, M., Cammue, B. P., et al. (2014). Synergistic activity of the tyrocidines, antimicrobial cyclodecapeptides from Bacillus aneurinolyticus, with amphotericin B and caspofungin against Candida albicans biofilms. Antimicrob. Agents Chemother. 58, 3697–3707. doi: 10.1128/AAC.02381-14
Uppuluri, P., Chaturvedi, A. K., Srinivasan, A., Banerjee, M., Ramasubramaniam, A. K., Kohler, J. R., et al. (2010). Dispersion as an important step in the Candida albicans biofilm developmental cycle. PLoS Pathog. 6:e1000828. doi: 10.1371/journal.ppat.1000828
Vale-Silva, L. A., Coste, A. T., Ischer, F., Parker, J. E., Kelly, S. L., Pinto, E., et al. (2012). Azole resistance by loss of function of the sterol Δ5,6-desaturase gene (ERG3) in Candida albicans does not necessarily decrease virulence. Antimicrob. Agents Chemother. 56, 1960–1968. doi: 10.1128/AAC.05720-11
van der Weerden, N. L., Bleackley, M. R., and Anderson, M. A. (2013). Properties and mechanisms of action of naturally occurring antifungal peptides. Cell. Mol. Life Sci. 70, 3545–3570. doi: 10.1007/s00018-013-1260-1
Vandeputte, P., Larcher, G., Bergès, T., Renier, G., Chabasse, D., and Bouchara, J. P. (2005). Mechanisms of azole resistance in a clinical isolate of Candida tropicalis. Antimicrob. Agents Chemother. 49, 4608–4615. doi: 10.1128/AAC.49.11.4608-4615.2005
Vandeputte, P., Pineau, L., Larcher, G., Noel, T., Brèthes, D., Chabasse, D., et al. (2011). Molecular mechanisms of resistance to 5-fluorocytosine in laboratory mutants of Candida glabrata. Mycopathologia 171, 11–21. doi: 10.1007/s11046-010-9342-1
Vecchiarelli, A., Pericolini, E., Gabrielli, E., and Pietrella, D. (2012). New approaches in the development of a vaccine for mucosal candidiasis: progress and challenges. Front. Microbiol. 3:294. doi: 10.3389/fmicb.2012.00294
Vermeulen, E., Lagrou, K., and Verweij, P. E. (2013). Azole resistance in Aspergillus fumigatus: a growing public health concern. Curr. Opin. Infect. Dis. 26, 493–500. doi: 10.1097/QCO.0000000000000005
Verweij, P. E., Mellado, E., and Melchers, W. J. (2007). Multiple-triazole-resistant aspergillosis. N. Engl. J. Med. 356, 1481–1483. doi: 10.1056/NEJMc061720
Vincent, B. M., Lancaster, A. K., Scherz-Shouval, R., Whitesell, L., and Lindquist, S. (2013). Fitness trade-offs restrict the evolution of resistance to amphotericin B. PLoS Biol. 11:e1001692. doi: 10.1371/journal.pbio.1001692
Voltan, A. R., Quindós, G., Alarcón, K. P., Fusco-Almeida, A. M., Mendes-Giannini, M. J., and Chorilli, M. (2016). Fungal diseases: could nanostructured drug delivery systems be a novel paradigm for therapy? Int. J. Nanomedicine 11, 3715–3730. doi: 10.2147/IJN.S93105
Vyas, S. P., and Gupta, S. (2006). Optimizing efficacy of amphotericin B through nanomodification. Int. J. Nanomedicine 1, 417–432. doi: 10.2147/nano.2006.1.4.417
Walsh, T. J., Anaissie, E. J., Denning, D. W., Herbrecht, R., Kontoyiannis, D. P., Marr, K. A., et al. (2008). Treatment of aspergillosis: clinical practice guidelines of the infectious diseases society of America. Clin. Infect. Dis. 46, 327–360. doi: 10.1086/525258
Walsh, T. J., Schlegel, R., Moody, M. M., Costerton, J. W., and Salcman, M. (1986). Ventriculoatrial shunt infection due to Cryptococcus neoformans: an ultrastructural and quantitative microbiological study. Neurosurgery 18, 373–375. doi: 10.1097/00006123-198603000-00025
Wang, X. J., Sui, X., Yan, L., Wang, Y., Cao, Y. B., and Jiang, Y. Y. (2015). Vaccines in the treatment of invasive candidiasis. Virulence 6, 309–315. doi: 10.4161/21505594.2014.983015
Wang, Y. H., Dong, H. H., Zhao, F., Wang, J., Yan, F., Jiang, Y. Y., et al. (2016). The synthesis and synergistic antifungal effects of chalcones against drug resistant Candida albicans. Bioorg. Med. Chem. Lett. 26, 3098–3102. doi: 10.1016/j.bmcl.2016.05.013
Watson, P. F., Rose, M. E., Ellis, S. W., England, H., and Kelly, S. L. (1989). Defective sterol C5-6 desaturation and azole resistance: a new hypothesis for the mode of action of azole antifungals. Biochem. Biophys. Res. Commun. 164, 1170–1175. doi: 10.1016/0006-291X(89)91792-0
Wheat, L. J., Connolly, P., Smedema, M., Durkin, M., Brizendine, E., Mann, P., et al. (2006). Activity of newer triazoles against Histoplasma capsulatum from patients with AIDS who failed fluconazole. J. Antimicrob. Chemother. 57, 1235–1239. doi: 10.1093/jac/dkl133
Whelan, W. L. (1987). The genetic basis of resistance to 5-fluorocytosine in Candida species and Cryptococcus neoformans. Crit. Rev. Microbiol. 15, 45–56. doi: 10.3109/10408418709104447
White, T. C. (1997). Increased mRNA levels of ERG16, CDR, and MDR1 correlate with increases in azole resistance in Candida albicans isolates from a patient infected with human immunodeficiency virus. Antimicrob. Agents Chemother. 41, 1482–1487.
White, T. C., Holleman, S., Dy, F., Mirels, L. F., and Stevens, D. A. (2002). Resistance mechanisms in clinical isolates of Candida albicans. Antimicrob. Agents Chemother. 46, 1704–1713. doi: 10.1128/AAC.46.6.1704-1713.2002
White, T. C., Marr, K. A., and Bowden, R. A. (1998). Clinical, cellular, and molecular factors that contribute to antifungal drug resistance. Clin. Microbiol. Rev. 11, 382–402.
Wojnicz, D., Kucharska, A. Z., Sokół-Łêtowska, A., Kicia, M., and Tichaczek-Goska, D. (2012). Medicinal plants extracts affect virulence factors expression and biofilm formation by the uropathogenic Escherichia coli. Urol. Res. 40, 683–697. doi: 10.1007/s00240-012-0499-6
Wozniak, K. L., Young, M. L., and Wormley, F. L. (2011). Protective immunity against experimental pulmonary cryptococcosis in T cell-depleted mice. Clin Vaccine Immunol. 18, 717–723. doi: 10.1128/CVI.00036-11
Xiang, M. J., Liu, J. Y., Ni, P. H., Wang, S., Shi, C., Wei, B., et al. (2013). Erg11 mutations associated with azole resistance in clinical isolates of Candida albicans. FEMS Yeast Res. 13, 386–393. doi: 10.1111/1567-1364.12042
Xie, J. L., Polvi, E. J., Shekhar-Guturja, T., and Cowen, L. E. (2014). Elucidating drug resistance in human fungal pathogens. Future Microbiol. 9, 523–542. doi: 10.2217/fmb.14.18
Yu, L., Ling, G., Deng, X., Jin, J., Jin, Q., and Guo, N. (2011). In vitro interaction between fluconazole and triclosan against clinical isolates of fluconazole-resistant Candida albicans determined by different methods. Antimicrob. Agents Chemother. 55, 3609–3612. doi: 10.1128/AAC.01313-10
Yu, L., Zhang, W., Wang, L., Yang, J., Liu, T., Peng, J., et al. (2007). Transcriptional profiles of the response to ketoconazole and amphotericin B in Trichophyton rubrum. Antimicrob. Agents Chemother. 51, 144–153. doi: 10.1128/AAC.00755-06
Yu, Q., Ding, X., Zhang, B., Xu, N., Jia, C., Mao, J., et al. (2014). Inhibitory effect of verapamil on Candida albicans hyphal development, adhesion and gastrointestinal colonization. FEMS Yeast Res. 14, 633–641. doi: 10.1111/1567-1364.12150
Zhang, L., Xiao, M., Watts, M. R., Wang, H., Fan, X., Kong, F., et al. (2015). Development of fluconazole resistance in a series of Candida parapsilosis isolates from a persistent candidemia patient with prolonged antifungal therapy. BMC Infect. Dis. 15:340. doi: 10.1186/s12879-015-1086-6
Keywords: antifungal drugs, antifungal resistance, biofilms, fungal vaccine, new antifungal therapy, alternative animal models, nanoparticles
Citation: Scorzoni L, de Paula e Silva ACA, Marcos CM, Assato PA, de Melo WCMA, de Oliveira HC, Costa-Orlandi CB, Mendes-Giannini MJS and Fusco-Almeida AM (2017) Antifungal Therapy: New Advances in the Understanding and Treatment of Mycosis. Front. Microbiol. 8:36. doi: 10.3389/fmicb.2017.00036
Received: 11 November 2016; Accepted: 06 January 2017;
Published: 23 January 2017.
Edited by:
Carlos Pelleschi Taborda, University of São Paulo, BrazilReviewed by:
Susana Frases, Federal University of Rio de Janeiro, BrazilPraveen Rao Juvvadi, Duke University, USA
Copyright © 2017 Scorzoni, de Paula e Silva, Marcos, Assato, de Melo, de Oliveira, Costa-Orlandi, Mendes-Giannini and Fusco-Almeida. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana M. Fusco-Almeida, YW5hLm1hcmlzYUB1b2wuY29tLmJy