 Shui-Xiu Li1,2†
Shui-Xiu Li1,2† Yan-Jun Song1,2†Yi-Shan Zhang1,2†Hao-Tian Wu1,2Hui Guo1,2
Yan-Jun Song1,2†Yi-Shan Zhang1,2†Hao-Tian Wu1,2Hui Guo1,2 Kun-Ju Zhu1,2*Dong-Mei Li3*Hong Zhang1,2*
Kun-Ju Zhu1,2*Dong-Mei Li3*Hong Zhang1,2*
- 1The First Affiliated Hospital of Jinan University, Guangzhou, China
- 2Institute of Mycology, Jinan University, Guangzhou, China
- 3Department of Microbiology and Immunology, Georgetown University Medical Center, Washington, DC, USA
The α subunit (ATP1) is a vital component of mitochondrial complex V which counts for the majority of cellular ATP production in a living organism. Nevertheless, how the α subunit influences other cellular processes such as pathogenicity in Candida albicans remains poorly understood. To address this question, ATP1 mutant (atp1Δ/Δ) and the gene-reconstituted strain (atp1Δ/ATP1) have been constructed in this study and their pathogenicity-related traits are compared to those of wild type (WT). In a murine model of disseminated candidiasis, atp1Δ/Δ infected mice have a significantly higher survival rate and experience a lower fungal burden in tissues. In in vitro studies atp1Δ/Δ lose a capability to damage or destroy macrophages and endothelial cells. Furthermore, atp1Δ/Δ is not able to grow under either glucose-denial conditions or high H2O2 conditions, both of which are associated with the potency of the macrophages to kill C. albicans. Defects in filamentation and biofilm formation may impair the ability of atp1Δ/Δ to penetrate host cells and establish robust colonies in the host tissues. In concert with these pathogenic features, intracellular ATP levels of atp1Δ/Δ can drop to 1/3 of WT level. These results indicate that the α subunit of Complex V play important roles in C. albicans pathogenicity.
Introduction
Candida albicans is the most common fungal pathogen capable of producing severe systemic infection in immune compromised individuals (Calderone et al., 2014). The overall morbidity and mortality for systemic candidiasis has been reported to be greater than 30% (Leroy et al., 2009; Wisplinghoff et al., 2014).
During hematogenous dissemination, blood-borne C. albicans must adhere to, colonize and invade the endothelial cells of blood vessels or the internal organs. Therefore, these cells need to develop a strategy that can counteract the stresses induced by immune-derived nutritional starvation, reactive oxygen intermediates and antimicrobial peptides (Grubb et al., 2008; Frohner et al., 2009; Walker et al., 2009). Mitochondria play an important role in each of these activities. For example, mitochondrial activity regulates the Ras1-cAMP-PKA signaling pathway that in turn induces a filamentation response for better invasion of the tissue (Dalle et al., 2010; Zhu and Filler, 2010; Grahl et al., 2015). In accordance with these morphological changes, the metabolic adaptability of fungal cells effectively facilitates the assimilation of nutrients available from a variety of sources (Brock, 2009; Fleck et al., 2011).
Mitochondrial complex V (CV) is the key enzyme in the final steps of oxidative phosphorylation. In living organisms, this ATP synthesis accounts for the majority of the cellular ATP yield required to drive the many energy-consuming reactions and processes of the organism (Senior, 1990; Harold and Maloney, 1996). This protein complex is an assemblage of complex F1 and complex F0. The former is composed of the subunits α3β3γδ𝜀 and the latter is composed of the subunits ab2c10–15. The fundamentals of oxidative phosphorylation rely on the α3β3 hexamer, of which the α subunit is encoded by ATP1 in C. albicans.
Genetic abnormality of the α subunit in humans can cause a fatal infantile mitochondrial encephalopathy (Jonckheere et al., 2013). The vital roles of the α subunit are also reflected in other organisms. In Saccharomyces cerevisiae, deletion of the α subunit renders the organism unable to grow (Lai-Zhang et al., 1999). Knockdown of the α subunit in Trypanosoma brucei during its infectious stage also results in a decreased growth rate due to suppressed ATP synthesis. In C. albicans, the α subunit has been found at the surface of hyphae but not in yeast cells (Hernández et al., 2004). Since, the yeast-hyphae transition in C. albicans is known to play a role in host tissue invasion, the emergence of ATP1 on the hyphal form gives rise to the question of what the α subunit contributes to C. albicans pathogenicity in vivo and how the essential energetic pathway gets involved with fungal growth in the host.
To answer these questions, we construct a null ATP1 mutant (atp1Δ/Δ) and a gene-reconstituted strain (atp1Δ/ATP1). The α subunit (ATP1) of C. albicans was characterized from these strains and compared to the wild strain (WT), and pathogenic roles of ATP1 were then firstly analyzed in vivo. We demonstrate that deletion of ATP1 diminishes the ability of C. albicans to cause systemic infections in animal model. The attenuated pathogenicity in these mutants may be due to some defects in the process of host invasion and compromised capability to form biofilm. We conclude that the α subunit is essential for C. albicans pathogenicity.
Materials and Methods
Strains and Growth Conditions
A null mutant of ATP5 (atp1Δ/Δ) was constructed by PCR-mediated homologous recombination as described previously (Sasse and Morschhäuser, 2012). C. albicans SC5314 (wild type; Gillum et al., 1984) was used to generate the atp1Δ/Δ mutant strain (atp1Δ::FRT/atp1Δ::FRT) and atp1Δ/ATP1 gene-reconstituted strain (atp1Δ::FRT /ATP1::FRT). The primers used for gene deletion in this study are listed in Supplementary Table S1. The confirmation of the atp1Δ/Δ mutant and reconstituted strain (atp1Δ/ATP1) are performed by PCR amplicons using primer pairs as shown in Supplementary Figures S1 and S2. Strains were routinely grown in YPD broth or on YPD agar (1% yeast extract, 2% peptone, and 2% glucose) with or without compounds as indicated.
Effect of ATP1 on the Virulence in Mice
A mouse model of disseminated candidiasis was used to evaluate the virulence of the strains (Spellberg et al., 2003, 2005). Female BALB/c mice (18–22 g; Guangdong Medical Laboratory Animal Center, Foshan, Guangdong, China) were used for all experiments. Mice were injected via the lateral tail vein with either a suspension of 1 × 105 cells or 1 × 106 cells from each strain. Survival rate was calculated from 10 infected mice per strain. For determination of fungal burden, another three mice from each group were euthanized after 1, 24, 48, and 72 h infection. Kidney, spleen and liver were harvested, weighed, homogenized, and quantitatively cultured. In addition, at day 1 of infection, mice were killed and organs removed to fix in 10% buffered formalin, then embed in paraffin, sectioned and stained with Periodic Acid-Schiff for histological study. Mortality was represented with Kaplan–Meier survival curves and quantitative tissue burdens were marked in the log scale and compared in the Mann–Whitney test.
Ethics Statement
The animal experiments were performed under the guidance of a protocol approved by the Animal Study Committee of the Institute of Dermatology, CAMS, according to the National Guidelines for Animal Care. All animal experiments were carried out with permission from the Ethical Committee of Institute of Zoonosis, Jinan University, Guangdong, China (Ref no. 20080101).
Interaction between C. albicans and Macrophages
The mouse macrophage cell line RAW264.7 (ATCC) was used for all assays. To investigate the susceptibility of C. albicans strains to the macrophages, macrophage cells were seeded in a 96-well plate at 5 × 104 cells/ml in 150 μl of DMEM with 10% FBS overnight at 37°C in 5% CO2. The C. albicans were diluted into DMEM at 2 × 106 cells/ml, and 50 μl was added into the first column wells, mixed and then serially diluted 1:4 a total of six times. Plates were incubated at 37°C and 5% CO2 for 24 h. Colonies were counted. The survival rate was calculated as the numbers of colonies with macrophages divided by the number of colonies in the absence of macrophages. P-values were determined using the unpaired Student’s t-test (She et al., 2015). For evaluating macrophage survival, macrophage cells were seeded in 24-well plate at 1 × 106 cells in 1 ml of DMEM with 10% FBS, and cultured at 37°C and 5% CO2 for 24 h. The C. albicans cells were suspended in DMEM with 10% FBS to 1 × 106 cells/ml, then added into the wells. After 1 h of co-incubation, supernatant was removed from each wells, washed and stained with Hoecheset33258 for 1 h, and observed by a fluorescent microscope (Olympus, Japan) with a TRIT-C/Texas red filter set. Hoecheset-positive (damaged) and total macrophages were counted, and the percentage of damaged macrophages was calculated as the number of damaged macrophages divided by that of total macrophages (Yu et al., 2014).
Effect of ATP1 on Endothelial Cells Damage
The extent of damage of endothelial cells by the C. albicans was quantified by measuring lactate dehydrogenase (LDH) activity (Mayer et al., 2012). The human umbilical vein endothelial cell line HUVEC (ATCC CRL-1730, LGC Standards, Promocell) cells were seeded in a 96-well plate at 1 × 105 cells/ml in 200 μl of DMEM with 10% FBS overnight at 37°C and 5% CO2 for 2 days. Cells were washed and 100 μl DMEM with 2% FBS were added. The C. albicans were diluted into DMEM at 5 × 105 cells/ml, and 100 μl was added to infect endothelial cells. Incubation was carried out at 37°C and 5% CO2 for 15 or 24 h. Measurement of LDH activity with the Cytotoxicity Detection Kit was performed according to the manufacturer’s manual. Absorbance of the samples was measured at 490 nm. Medium only and low control values were subtracted from all sample values. Damage was expressed as percentage of the control strain. Each experiment was performed at least in triplicate.
Effect of ATP1 on C. albicans Adhesion
The adhesion was observed visually (Nobile et al., 2006). The 24-well flat bottomed pre-sterilized microtiter plates were incubated in fetal bovine serum overnight at 37°C. A suspension of 1 × 107 cells of C. albicans in Spider media was added to the wells and incubated at 37°C for 2 h. Non-adherent cells were removed by washing with PBS. Fresh Spider media was added to the corresponding wells and the plates were incubated at 37°C for 24 h. The wells were washed and photographed.
Effect of ATP1 on C. albicans Filamentation
A 1 × 106 cells/ml suspension of each strain in YPD, YPD+10% FBS, Spider and Lee’s liquid media was added to the wells of a 12-well flat-bottomed pre-sterilized microtiter plate and incubated at 37°C for 2 h. The plates were visualized under inverted microscope and photographed. In addition, cells of each strain were serial diluted and spotted on YPD, YPD+10% FBS, Spider, Lee’s and SLAD agar, and incubated at 37°C for 7 days. The colonies were photographed.
Effect of ATP1 on C. albicans Biofilm Formation
The biofilms were observed by Confocal laser scanning microscopy (CSLM) (Nobile et al., 2006). Briefly, all strains were grown in Glass Bottom Cell Culture Dish. After 24 h of incubation at 37°C, the resulting biofilms were washed and stained with 25 μg/ml Concanavalin A-Alexa Fluor 594 conjugate (C-11253; Molecular Probes, Eugene, OR) at 37°C for 1 h. CSLM was performed with a Zeiss LSM 510 upright confocal microscope (Carl Zeiss, Thornwood, NY, USA), using a Zeiss Achroplan 40×, 0.8-W objective, and a HeNe1 laser with an excitation wavelength of 543 nm. Moreover, biofilms activity was assessed by XTT reduction assay and the crystal violet assay, accordingly to previously described protocols (Peeters et al., 2008; Pierce et al., 2008; Krom and Willems, 2016). Each experiment was performed at least in triplicate.
Quantitative Real Time Polymerase Chain Reaction (qRT-PCR) Assay
Overnight cultures of tested strains in 5 ml of YPD at 30°C were collected, then diluted into 100 ml of YPD or Spider medium to obtain an OD600 of 0.2 and incubated at 37°C for additional 6 h. Total RNA was extracted using the E.Z.N.A. Yeast RNA kit (Omega Bio-tek) following the manufacturer’s instruction. cDNA was synthesized and qRT-PCR was done as previously described (Guo et al., 2014).
Stress Susceptibility
To investigate the sensitivity of C. albicans strains to different stress agents, cells were overnight cultured, washed and suspended in PBS with an initial OD600 of 1.0. Five microliters of 10-fold dilutions was spotted onto YPD agar without or with different stress agents. For testing the sensitivity of C. albicans strains to heat stress, plates were cultured at indicated temperatures for 2 days and then photographed. The minimum inhibitory concentrations (MICs) were determined for each strain according to the broth microdilution method of the Clinical and Laboratory Standards Institute (CLSI) M27-A3 protocol (Method M27-A3, CLSI, 2008).
Growth Curve Assay
All strains were routinely grown in 5 ml of YPD at 30°C overnight, washed with PBS, then inoculated in 100 ml of YPD with an OD600 of 0.02. Shake cultures were grown at 30°C and OD600 of each strain was measured every two hours.
Mitochondrial Function Assay
All strains were grown in YPD overnight at 30°C. Mitochondrial complex V activity was measured by an assay kit according to the manufacturer’s protocol. For determinations of intracellular ATP concentrations, an aliquot of 1 × 106 cells from each strain was mixed with the same volume of BacTiter-GloTM reagent (Promega Corporation, Madison, WI, USA) and incubated for 5 min at room temperature as described previously (Zhang et al., 2013). ROS measurement was performed by using a oxidation-sensitive fluorescent dye DCFDA (Guo et al., 2014). Briefly, a suspension of 2.5 × 106 cells was stained with DCFDA (20 μg/ml) at 37°C for 20 min. The emission spectra at 488 nm and excitation spectra at 595 nm were determined by FACScan flow cytometer (Becton Dickinson). The cyanine dye JC-1 was used for determination of mitochondrial membrane potential of each strain (She et al., 2015). A suspension of 2.0 × 106 cells was incubated with 5 μM JC-1 at 37°C for 15 min and excitation spectra at 595 nm was determined by FACScan flow cytometer with emission spectra at 488 nm (Becton Dickinson).
Results
Deletion of ATP1 Results in Avirulence in a Mouse Model of Hematogenously Disseminated Candidiasis
The atp1Δ/Δ mutant displayed moderately reduced growth rate in vitro even when grown in rich medium (Figure 1A, Supplementary Table S3), which is similar to other C. albicans mutants with reduced virulence (Murad et al., 2001; Hwang et al., 2003; Chauhan et al., 2005; Lopes da Rosa et al., 2010; Chang et al., 2015; Lee et al., 2015). We speculate that such slow growth of atp1Δ/Δ in vitro may affect its virulence. A murine model of hematogenously disseminated candidiasis was first used to measure virulence changes in atp1Δ/Δ. All of the mice infected intravenously with either 1 × 105 cells or 1 × 106 cells of atp1Δ/Δ survived post 30 days infection (Figure 1B). By contrast, mice infected with WT began to die on days 7 and 1 post infection, and all mice were moribund on days 14 and 3, respectively. On the other hand, mice infected with 1 × 105 cells and 1 × 106 cells of atp1Δ/ATP1 began to die on days 8 and 4 and all mice were moribund on days 15 and 8, respectively. Clearly, ATP1 is required for C. albicans virulence, and the loss of virulence in atp1Δ/Δ is correlated with is not slow growth in vitro.
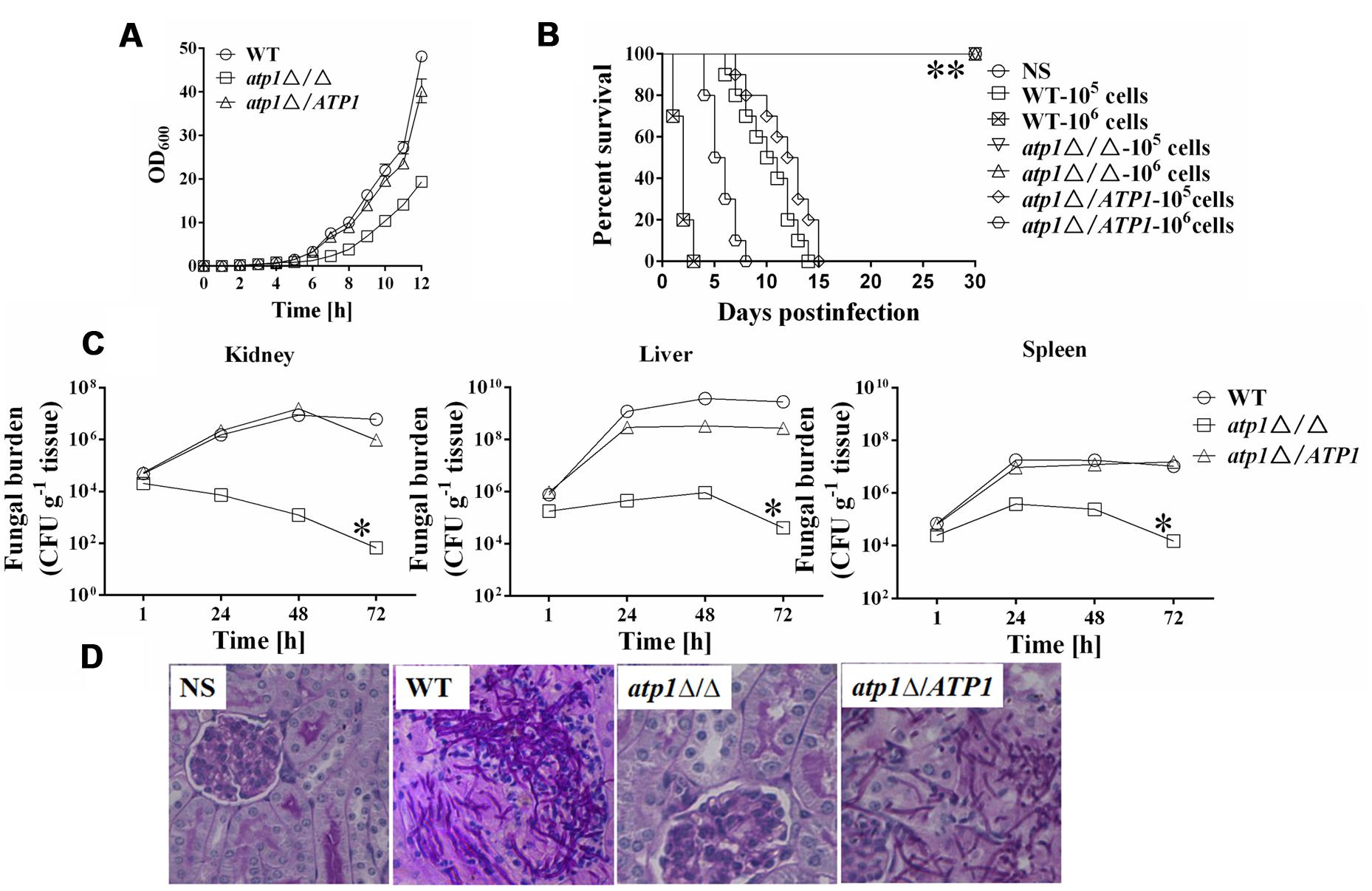
FIGURE 1. Deletion of ATP1 results in avirulence in a mouse model of hematogenously disseminated candidiasis. (A) Reduced growth rate of the atp1Δ/Δ was determined by measuring the OD600 of cells growing in YPD at 30°C. (B) Mice survival following infection with 1 × 105 and 1 × 106 cells of the Candida albicans. ∗∗ P < 0.0001 compared to mice infected with WT. (C) Fungal burden (CFU/g tissue) of mice infected with 1 × 106 cells of the C. albicans. Following infection, at 1, 24, 48, and 72 h, kidney, liver and spleens were removed, weighed, homogenized and samples plated on YPD agar medium. The total CFUs in tissue for each strain were shown over time. Data are from two experiments with three mice per time point for each strain (mean ± SD). ∗P < 0.05 compared to mice infected with WT. (D) Periodic acid Schiff staining of kidney sections from mice after 1 day of infection with C. albicans. Pictures were taken at 40× magnification.
Next, to correlate the avirulence of atp1Δ/Δ with its growth in host tissues, the fungal load was evaluated in kidney, liver and spleen (Figure 1C). The colony forming unit (CFU) was the same for all strains 1 h post infection. However, tissue loads in either WT or atp1Δ/ATP1 increased significantly (by about 2 orders of magnitude) in kidney while atp1Δ/Δ load gradually reduced post infection and dropped by about one order of magnitude at 48 h post infection. The fungal loads in liver and spleen reached their peaks at 48 h for all three strains but reduced fungal cell populations were observed in mutant during 72 h post infection when compared to the control strains. At 72 h post infection with atp1Δ/Δ, fungal cells continued to be cleared from the kidney, liver and spleen. The significant reduction of fungal burden in the kidneys infected with the atp1Δ/Δ within 3 days of infection suggests that ATP1 is required for kidney invasion and is necessary for the organism to persist in other tissues at later time points.
The low fungal burden in the kidneys of mice infected with the atp1Δ/Δ was verified by histopathology. After 24 h of infection, the kidneys infected with atp1Δ/Δ were scant to detect in tissue sections, as demonstrated by a absence of fungal hyphae and inflammatory cells, even though the fungal load was still positive according to CFU count. By contrast, kidneys infected with either WT or atp1Δ/ATP1 displayed the expected micro-abscesses consisting of numerous inflammatory cells surrounded by abundant hyphae or pseudohyphae (Figure 1D), suggesting that the avirulence of the atp1Δ/Δ is likely due to defects in tissue invasion and colonization ability.
Deletion of ATP1 Results in Hypersensitivity to Macrophages
Macrophages play a key role in host defense against C. albicans infections (Mayer et al., 2012). In order to invade the host organs and begin colonization, blood-borne C. albicans must evade the attacks from the immune system prior to crossing the endothelial cell barrier of blood vessels (Grubb et al., 2008). We therefore investigate whether ATP1 is required for C. albicans sensitivity to host killing events. The sensitivity of the atp1Δ/Δ to macrophages and its capability to damage macrophages were both measured. Macrophage killing tests revealed that atp1Δ/Δ is more vulnerable to macrophages, as demonstrated by the survival of fewer atp1Δ/Δ cells after an overnight incubation with macrophages than those from either WT or atp1Δ/ATP1 (Figure 2A). On the other hand, far fewer macrophages were damaged when incubated with atp1Δ/Δ cells than when incubated with the control strains (Figure 2B). Evanescence of atp1Δ/Δ cells and insufficient killing of macrophages suggest that the ATP1 is required for C. albicans to evade macrophage attacks.
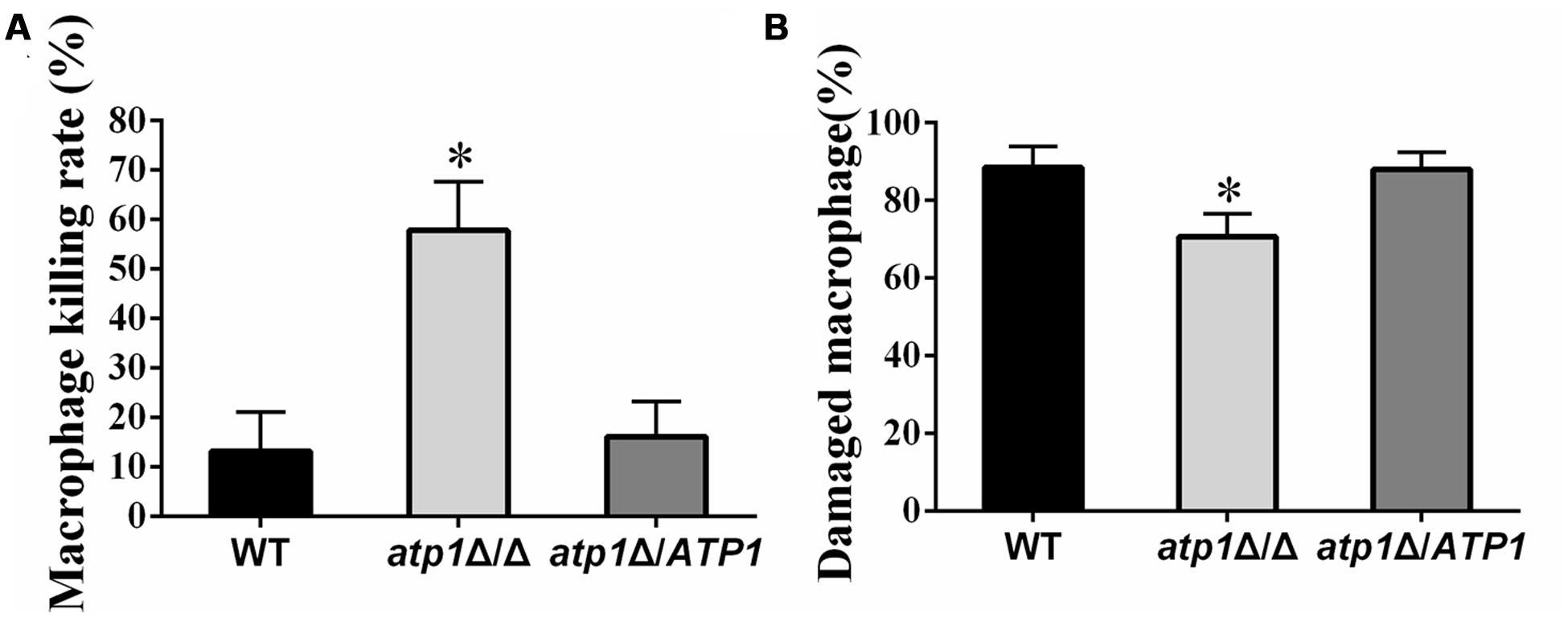
FIGURE 2. Interaction between C. albicans and macrophages. (A) Viability of all strains is shown after an overnight incubation with macrophages (RAW264.7). Viable colonies of each strain were counted under microscopy. Data are averages of three separate experiments. ∗P < 0.05 compared with WT. (B) Macrophages were incubated with the indicated strains of C. albicans for 1 h and the extent of macrophages damage was then determined. Data are averages of three separate experiments. ∗P < 0.05 compared with WT.
Deletion of ATP1 Disables Utilization of Non-glucose Carbon Sources and Increases Sensitivity to Oxidative Stress in C. albicans
The capability of C. albicans to utilize non-glucose carbon sources and thereby counteract elevated reactive oxygen species (ROS) within macrophages are believed to be two elemental factors for the establishment of candidiasis (Frohner et al., 2009). We find that growth of the atp1Δ/Δ on spot assay is slightly less pronounced than those of WT and atp1Δ/ATP1 on YPD agar, but show no growth on YPG (glycerol), YPE (ethanol), YPO (oleic acid), or YPA (acetate). The last four media are YP-based media integrated with a non-glucose carbon source (Figure 3A). For oxidative stress response, the atp1Δ/Δ showed a reduced growth when exposed to H2O2, but no response to menadione (Figure 3B). These data indicate a vulnerable response of the atp1Δ/Δ to macrophage attacks may be due largely to an inability to consume non-glucose carbons as well as poor management of oxidative stress.
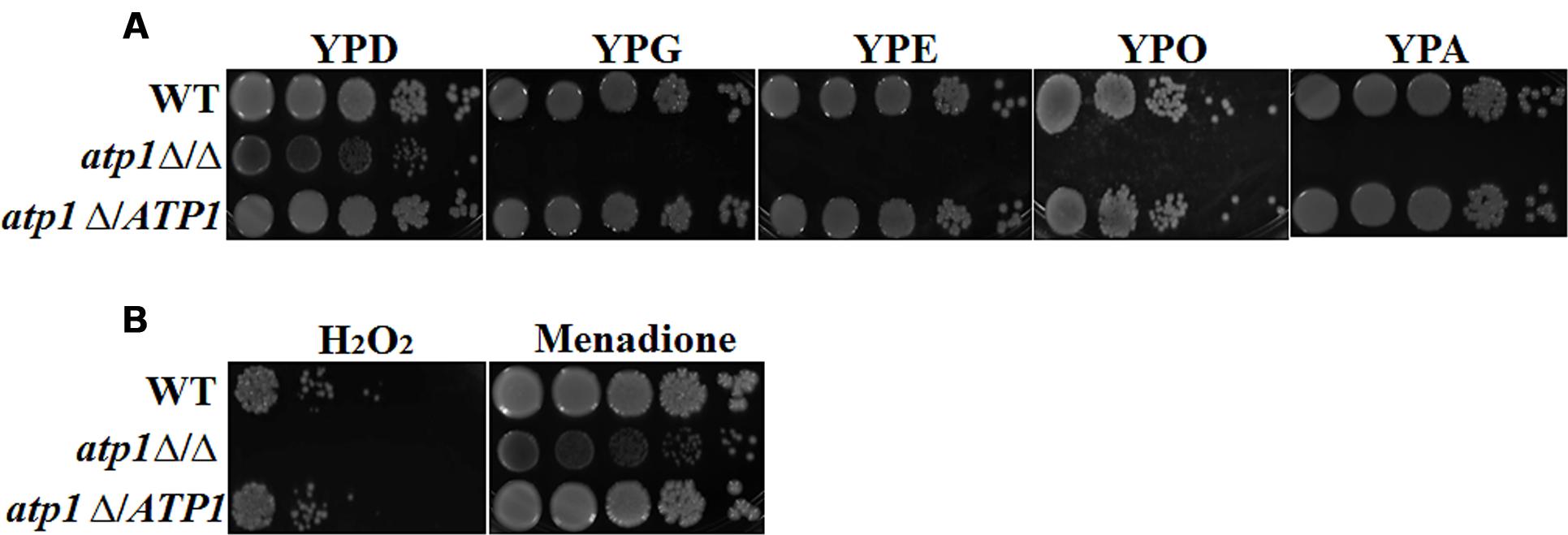
FIGURE 3. Deletion of ATP1 disables utilization of non-glucose carbon sources and elevates oxidative stress. (A) C. albicans strains were grown overnight in YPD medium, washed and serially diluted in PBS. Three microlitres of each serial dilution was plated on the YP medium supplemented with 2% glucose (YPD), 2% glycerol (YPG), 3% oleic acid (YPO), 2% ethanol (YPE), or 2% acetate (YPA) agar plates. (B) Oxidative stress response. Strains were grown in the presence of 8 mM hydrogen peroxide and 0.05 mM menadione. All cultures were incubated for 2 days at 30°C.
Deletion of ATP1 Results in Less Damage to Endothelial Cells in vitro
Endothelial cells of blood vessels provide a physical barrier for C. albicans to penetrate and invade the internal organs. We find that the atp1Δ/Δ has a significantly reduced capacity to destroy endothelial cells. The cell lysis rates were 77% less and 24% less at 15 and 24 h of infection (Figure 4), respectively, when compared to WT. These results indicate that ATP1 is required for endothelial cell penetration.
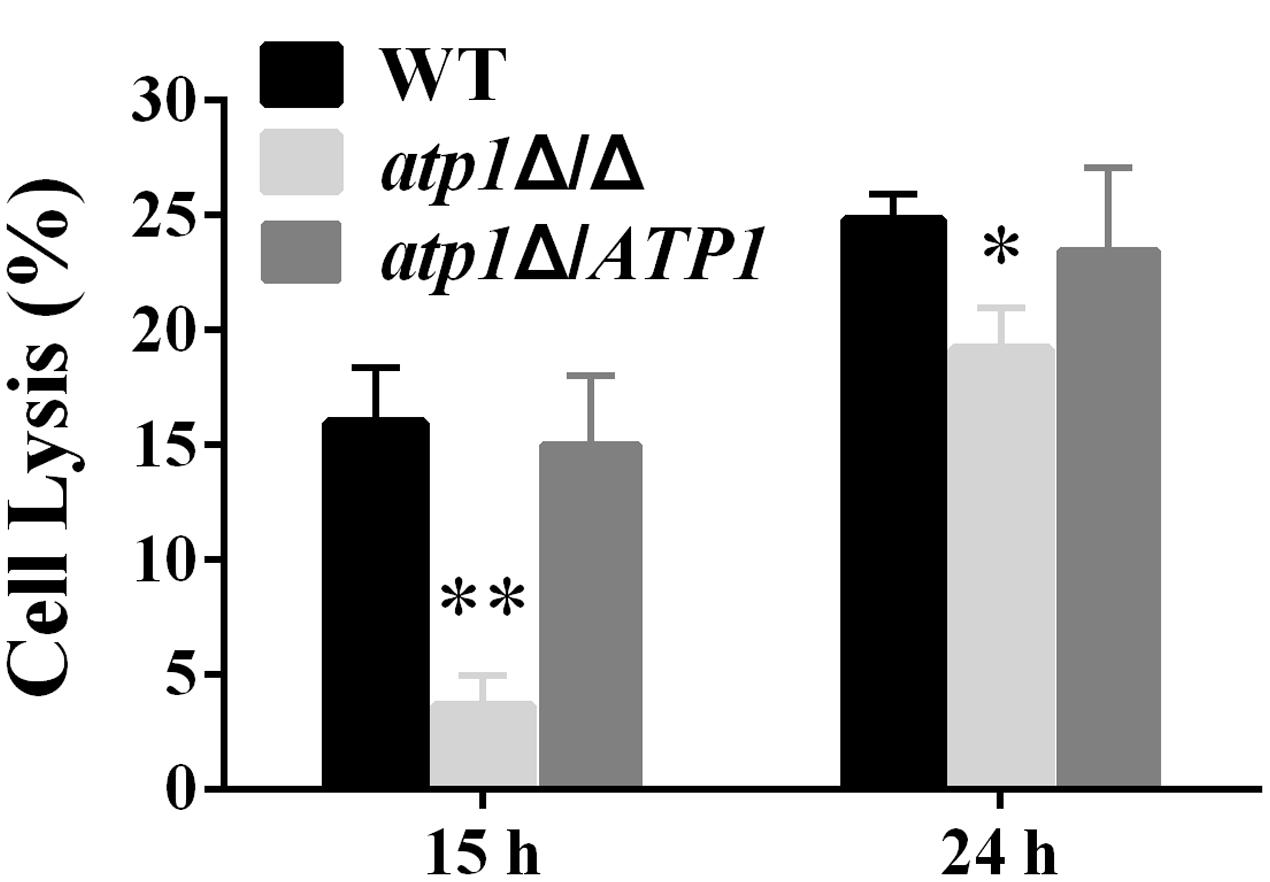
FIGURE 4. Deletion of ATP1 causes less damage of human endothelial cells. Monolayers of HUVEC endothelial cells were infected with C. albicans strains for 15 or 24 h. Damage of host cells was determined by measuring lactate dehydrogenase (LDH) levels. Results are the mean ± SD of at least three independent experiments, each performed in triplicate. ∗P < 0.05 and ∗∗P < 0.01 compared to WT.
Deletion of ATP1 Displays Defective Adhesion and Invasion
The contribution of ATP1 to adhesion events has also been tested in microtiter plates in vitro in order to understand whether less lysis of macrophages or endothelial cells may be due to the inefficiency of adhesion (Filler et al., 1991; Park et al., 2005; Wächtler et al., 2011). Adhesion assay reveals that atp1Δ/Δ fails to adhere to the bottom of the microtiter plate when compared with WT (Figure 5A). In qRT-PCR assay, the gene expression levels of SSA1 and ALS3 were markedly down-regulated in the atp1Δ/Δ when compared with WT (Figure 5B). These genes have long been known for their prominent roles in host tissue adhesion and invasion (Phan et al., 2007; Sun et al., 2010).
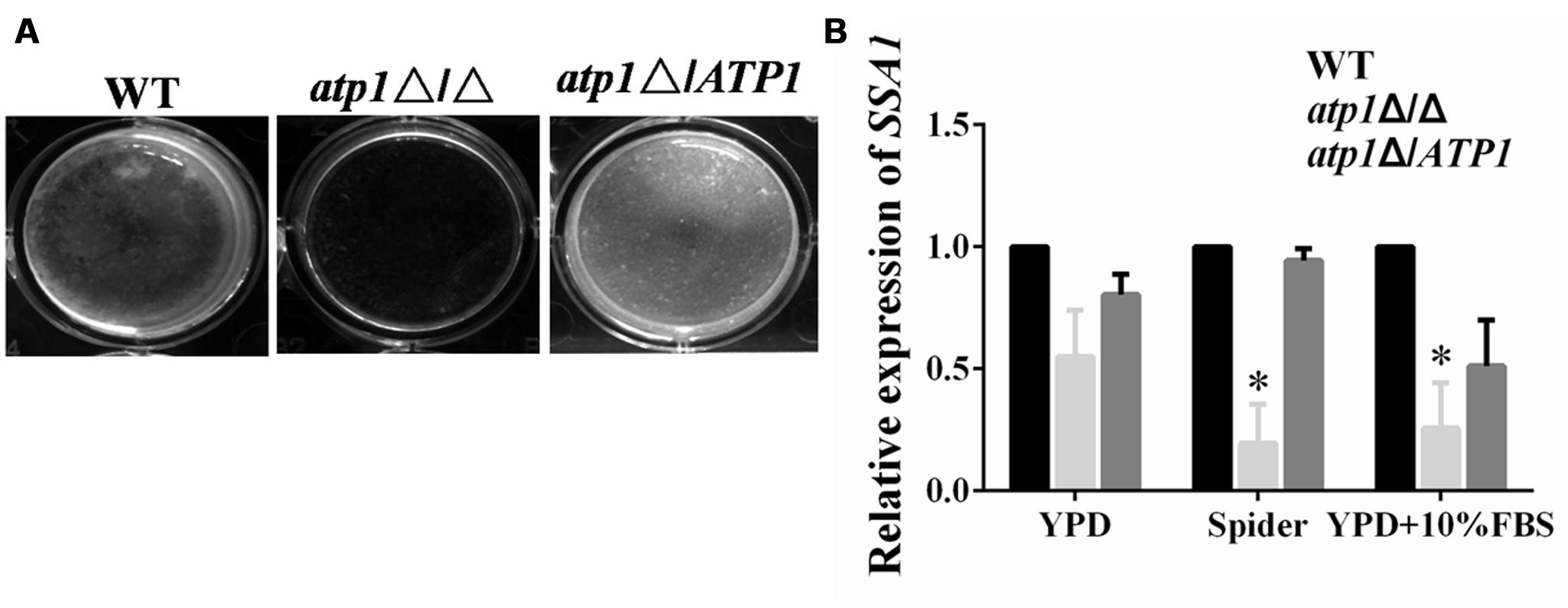
FIGURE 5. Deletion of ATP1 displays defective adhesion and invasion. (A) Effect of ATP1 on C. albicans adhesion. 1 × 107 cells suspension of C. albicans in spider media was added to 24-well flat bottomed plates and incubated for 24 hours at 37°C. The wells were washed with PBS and taken photos. (B) Effect of ATP1 on the expression of C. albicans adhesion gene. Effect of ATP1 on the expression of C. albicans invasion gene. C. albicans cells were incubated in YPD, Spider, YPD+10%FBS media at 37°C for 6 h. Following incubation expression of the indicated genes were determined by qRT-PCR. Expression level of each gene is displayed after normalization with internal control housekeeping gene 18S. The histogram shows the relative expression fold change of genes. Results represent the average of three independent experiments ±SD. ∗P < 0.05 when compared with WT.
Deletion of ATP1 Displays Defective Filamentation and Colony Formation
Filamentous growth enables C. albicans to penetrate host immune cells and tissue more efficiently after endocytosis (Phan et al., 2000; Dalle et al., 2010; Zhu and Filler, 2010). The requirement of ATP1 for filamentation has been tested under hyphal inducing conditions. In each hyphal inducing medium (10% FBS, Spider and Lee’s medium), the atp1Δ/Δ displays shorter or even no filamentous growth at 2 h growth point compared with WT, which has massive and long filamentous growth under microscopy (Figures 6A,B, Supplementary Figure S3). Similar results were also present on each agar medium. The atp1Δ/Δ displays a more severe defective phenotype in filamentous growth and forms small downy or even smooth colonies which lack the peripheral and invasive filaments, while WT produced large colonies with florid and invasive filaments on the edges (Figure 6C). The atp1Δ/ATP1 has intermediate effects in these matters between WT and atp1Δ/Δ. The filamentation defect in the atp1Δ/Δ is consistent with marked down-regulation of hyphal-specific genes such as HWP1, ALS3, ECE1, HGC1 in qRT-PCR test (Figure 6D). These results indicate that ATP1 is required for the filamentation process in C. albicans.
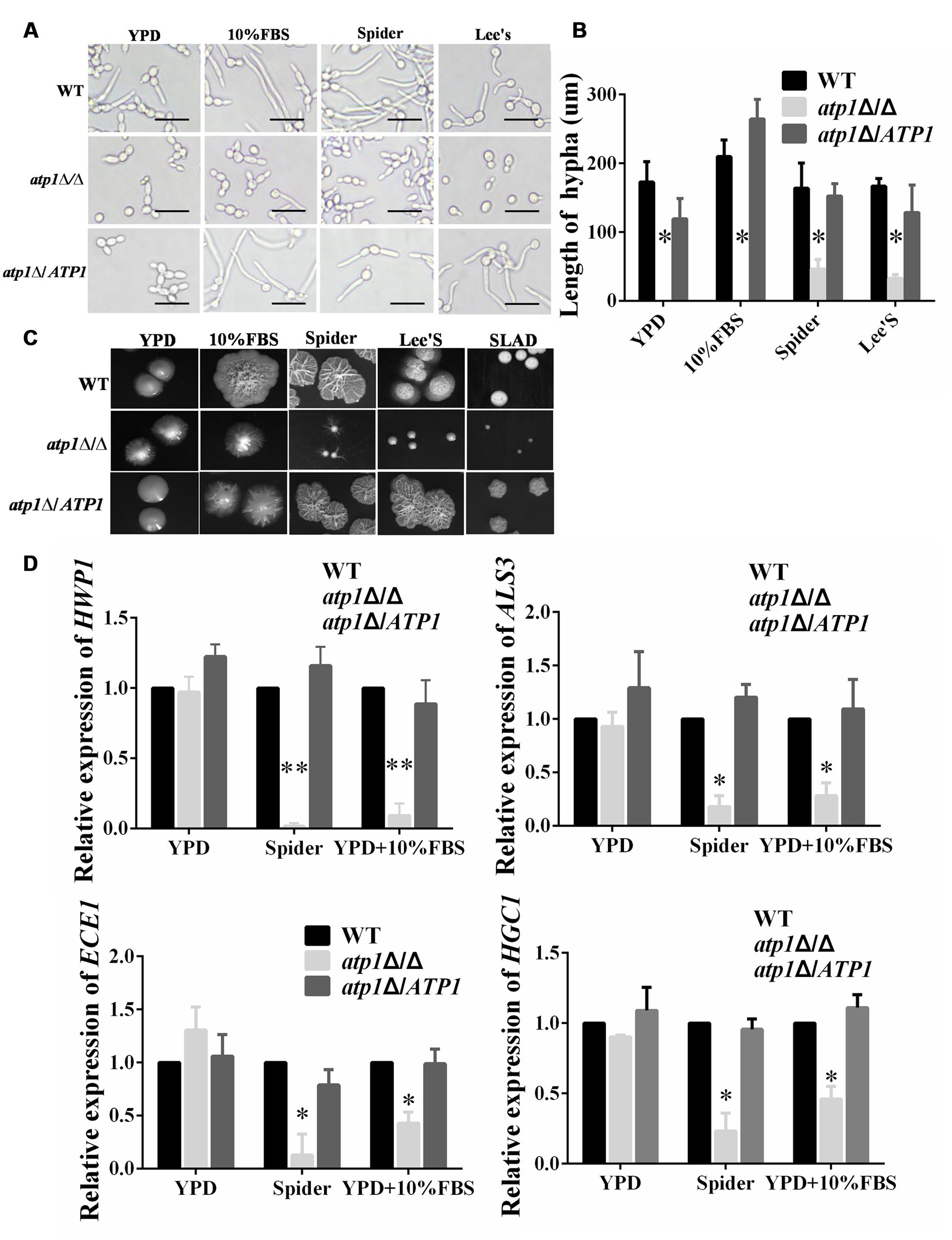
FIGURE 6. Deletion of ATP1 displays defective filamentation and colony formation. (A) Overnight cultures of the strains were resuspended to 1.0 × 106 cells/ml, and added into 12-well microtiter plates with filaments-inducing media. All the plates were cultured at 37°C for 2 h before photographed. (B) Length of filaments was measured. (C) Overnight cultures of the strains were resuspended, and 500 cells were spotted onto the indicated filaments-inducing agar plates. All the plates were cultured at 37°C for 7 days before photographed. (D) Effect of ATP1 on the expression of C. albicans hyphal-specific genes. C. albicans cells were incubated in YPD, Spider, YPD+10%FBS media at 37°C for 6 h. Following incubation expression of the indicated genes were determined by qRT-PCR. Results represent the average of three independent experiments ± SD. ∗P < 0.05 and ∗∗P < 0.01 when compared with WT.
Deletion of ATP1 Displays Defective Biofilm Formation
Aside from filament propensity, Candida biofilm also carries important clinical consequences because it will encase organism from antifungal therapy and withstand host immune defense (Douglas, 2003; Nett et al., 2010). Confocal scanning laser microscopy (CSLM) imaging was used to visualize biofilm formation in the atp1Δ/Δ and meanwhile, the biomass and cell metabolic activity within biofilm were measured. After 24 h culture, the atp1Δ/Δ forms a rudimentary biofilm with a thickness of less than 20 μm, while WT produces biofilm with thickness of over 50 μm, and atp1Δ/ATP1 produces biofilm of intermediate thicknesses (>20 μm and <50 μm) (Figure 7A). Under CSLM, the rudimentary atp1Δ/Δ biofilm consists mainly of yeast cells, with few filaments, whereas the biofilms of WT and atp1Δ/ATP1 have abundant filaments (Figure 7A). Consistently, the metabolic activity and total biomass of the atp1Δ/Δ are obviously reduced by XTT assay (Figure 7B) and by crystal violet assay, respectively when compared with WT and atp1Δ/ATP1 (Figure 7C). These results suggest that ATP1 deletion may be responsible for the failure of C. albicans to form biofilms.
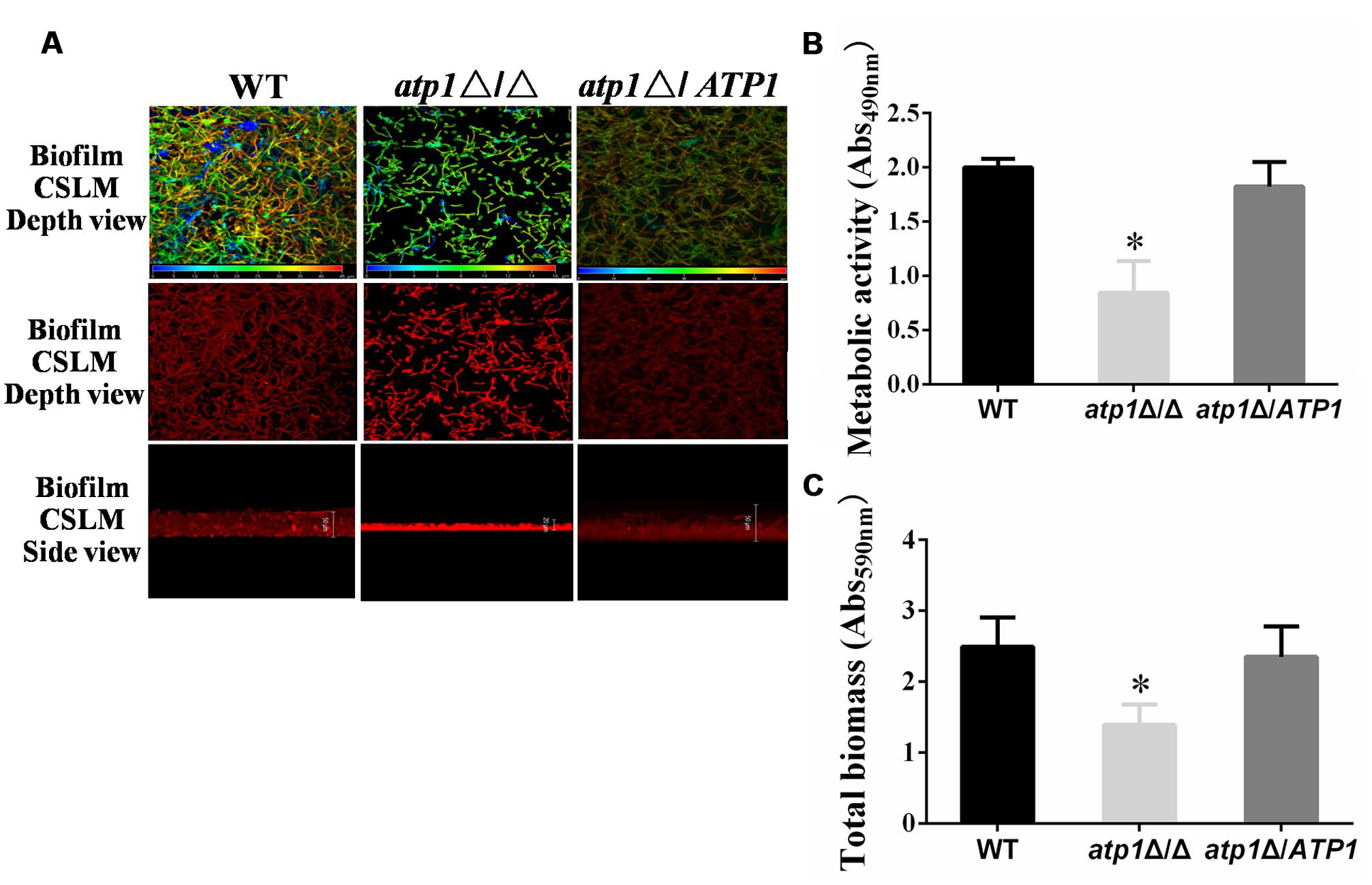
FIGURE 7. Deletion of ATP1 displays defective biofilm formation. (A) Biofilms of strains were stained with Concanavalin A-Alexa Fluorconjugate and analyzed by CSLM visualization. The top two panels show the visual appearance and depth over top view, in which blue color represents cells close to the glass bottom and red color represents cells far from the glass bottom. The bottom panel is side view of biofilm depth. At depth views, WT, blue = 0 μm and red = 45 μm; atp1Δ/Δ, blue = 0 μm and red = 16 μm;atp1Δ/ATP1, blue = 0 μm and red = 55 μm. At the side views, the biofilm depths are 50 μm for WT and atp1Δ/ATP1 but 20 μm for atp1Δ/Δ. (B) Biofilm was quantified colorimetrically by XTT assay, which measured biofilm metabolic activity. (C) Biofilm was quantified colorimetrically by crystal violet assay, which measured biofilm total biomass. Error bars represent the standard deviation among results for different isolates. ∗P < 0.05 when compared with WT.
Deletion of ATP1 Results in a Suppression of Mitochondrial Activity
The association of mitochondrial activity with C. albicans filamentation has been noted previously (Bambach et al., 2009; Grahl et al., 2015). Enzymatic assay of mitochondrial CV and intracellular ATP content were used to evaluate mitochondrial activity in the atp1Δ/Δ mutant. In the absence of the α subunit of Complex V, we find that the activity of Complex V is only 50% of the WT level (Figure 8A) and intracellular ATP drops to 1/3 of WT levels (Figure 8B). Meanwhile, the membrane potential of the atp1Δ/Δ is significantly decreased to nearly 50% of WT (Figure 8D). These data therefore confirm a critical role of ATP1 on mitochondrial function. However, in spite of these suppressed mitochondrial functions, we find that atp1Δ/Δ has a decreased ROS when compared with the WT (Figure 8C). Unlike other mitochondrial CI mutants with high ROS (Li et al., 2011; She et al., 2015), such reduced ROS levels in atp1Δ/Δ may be due to the fact that CV is not a direct electron acceptor. However, the contribution of ROS scavengers in atp1Δ/Δ needs to be clarified with further study.
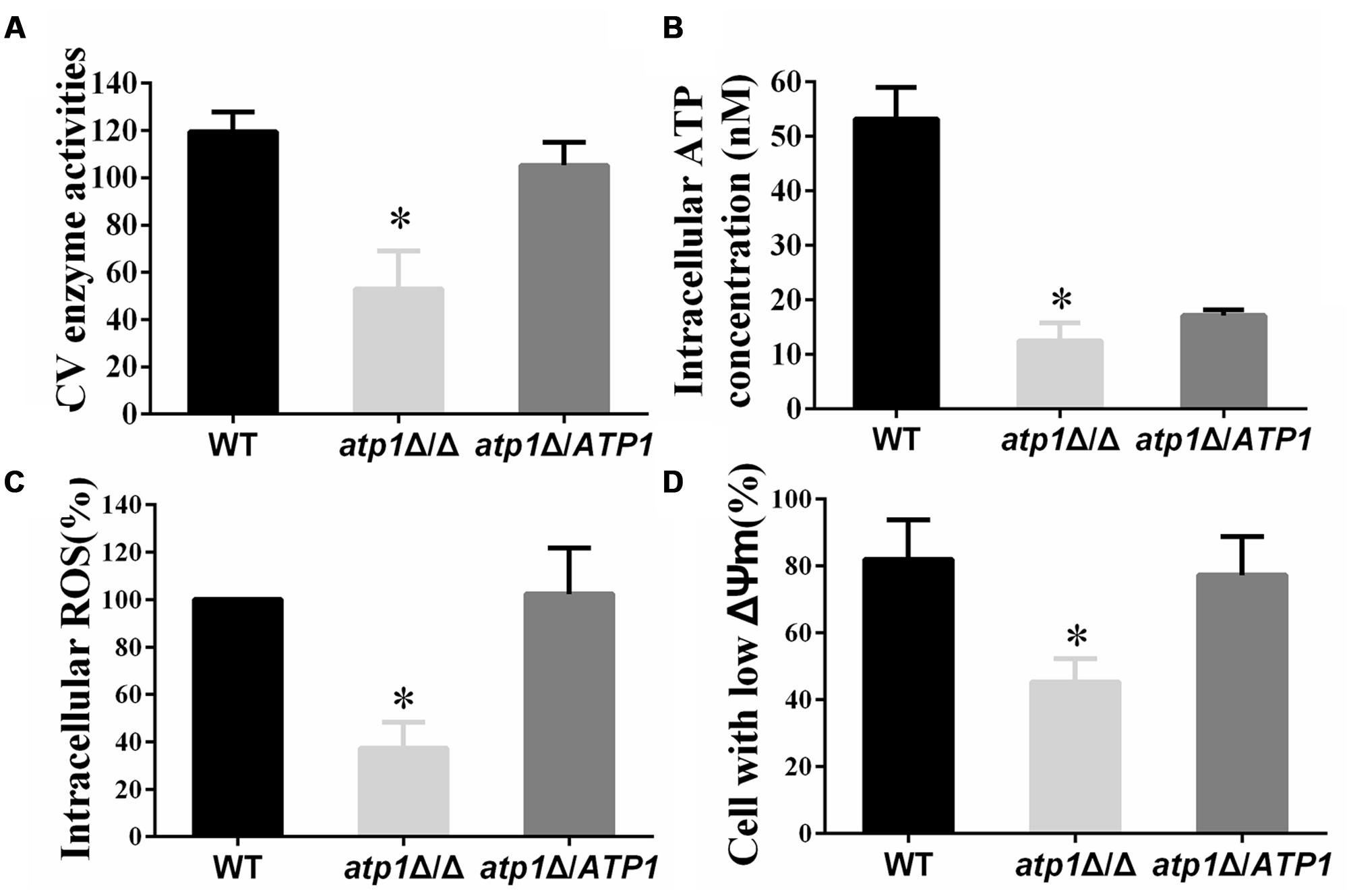
FIGURE 8. Deletion of ATP1 results in decreasing mitochondrial activity. (A) Mitochondrial Complex V enzyme activities. (B) The intracellular ATP content was measured by microplate reader. (C) The ROS levels were measured with DCFDA dye by flow cytometry. Cells were collected, resuspended in PBS and incubated with DCFDA at 37°C for 20 min. (D) Mitochondrial membrane potential was measured with JC-1 dye by flow cytometry. Cells were collected, resuspended in PBS and incubated with JC-1 for 15 min at 37°C. Gated region R1 includes cells with intact mitochondrial membranes and gated region R2 depicts cells with loss of mitochondrial membrane potential. ∗P < 0.01.
Deletion of ATP1 Changes the Stress Response in C. albicans
The fate of invasive Candida cells also depends on how well they handle stress in the host environment. To evaluate these adaptive skills, we measured the susceptibility of the atp1Δ/Δ to various stresses. As shown in Figure 9A, atp1Δ/Δ is susceptible to plasma membrane stress (SDS), but the susceptibilities to cell wall (Calcofluor White, Congo Red), heat, calcium (CaCl2) and osmotic stress (NaCl) are less pronounced in mutant when compared to its growth on YPD agar (Supplementary Figure S4). Interestingly, atp1Δ/Δ shows resistance to FLC, but not to ITR, KCZ and VRC (Figure 9B, Supplementary Table S2) even though the membrane of this mutant is more susceptible to SDS. The FLC resistance in the absence of ATP1 highlights the possible link between ATP1 function and FLC target or uptake process.
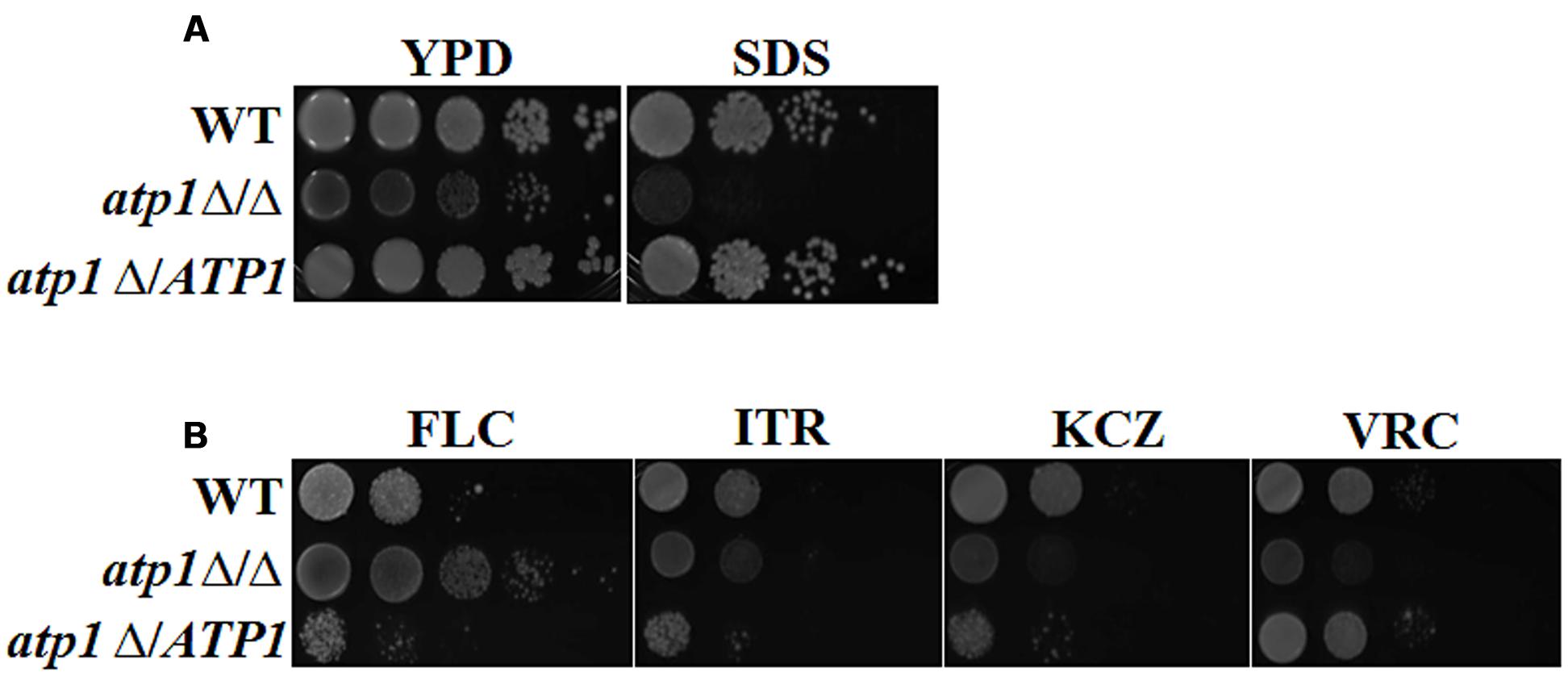
FIGURE 9. Deletion of ATP1 diverge stress response. (A) C. albicans strains were grown overnight in YPD medium, washed and serially diluted in PBS. Three microlitres of each serial dilution were grown in the presence of 0.06% SDS. (B) Susceptibility to different antifungal agents. Strains were grown in the presence of 2 μg/ml FLC, 2 μg/ml ITR, 2 μg/ml KCZ, and 2 μg/ml VRC.
The Expression of ATP1 in Yeast and Hyphal forms of C. albicans
In C. albicans, the α subunit has been found at the surface of hyphae but not in yeast cells (Hernández et al., 2004). To better understand the pathogenic roles of ATP1 in this organism, we also measured the gene expression levels of ATP1 at the yeast and hyphal stages of WT. We find ATP1 expression levels at the hyphal form are similar to those of yeast form (Supplementary Figure S5). Likely, one possible explanation is that Atplp is regulated at the post-translational level rather than transcriptional level. Whether ATP1 is hyphal-specific requires further investigation.
Discussion
The previous studies have demonstrated that mitochondrial activity is prerequisite for C. albicans virulence (Bambach et al., 2009; Grahl et al., 2015). In this study, the α subunit of mitochondrial Complex V was deleted from C. albicans and its pathogenic association was firstly assessed in animal model and in vitro immune cell lines. Our results with the atp1Δ/Δ confirm that ATP1 is required for C. albicans virulence by interfering with adhesion, filamentation, biofilm formation and stress adaptation.
Candida albicans mutants with slow growth in vitro often show defects in virulence as well, but this is not always true (Noble et al., 2010). Clearly, the atp1Δ/Δ grows more slowly in vitro. In order to minimize this slower growth effect, mice infected with ten times more atp1Δ/Δ cells were also included for virulence study. No matter how many fungal cells were introduced, all mice infected with the mutant survived without any clinical signs of the disease, whereas mice infected with WT were always killed. It is therefore likely that the loss of virulence in the atp1Δ/Δ is not simply due to slow growth, but rather that the lack of Candida cells in the kidneys of the atp1Δ/Δ infected mice is due to the defects of the mutants in colonizing and invading the host tissues.
After reaching the bloodstream, C. albicans must evade immune attacks and then penetrate the endothelial cells of the blood vessels (Mayer et al., 2012). The results of interactions between C. albicans and macrophages show that the atp1Δ/Δ is more vulnerable to be killed by macrophages. Among the macrophages, highly reactive intermediates like ROS that are generated during interaction with pathogens, and nutrient starvation are believed to be the strategies used by macrophages to kill C. albicans (Frohner et al., 2009). Obviously, the atp1Δ/Δ has no advantages against macrophages, which are demonstrated by no growth of the atp1Δ/Δ on non-glucose carbon source agar as well as increased sensitivity to H2O2. These findings may also explain an avirulent phenotype of the mutant. On the other hand, stress response is almost intact with the atp1Δ/Δ mutant, except that it is susceptible to SDS and resistant to FLC. Unlike Complex I subunit mutants that often display a hypersensitivity to FLC (Sun et al., 2013), the resistance to FLC over other azoles suggests that ATP1 may be specific for FLC target binding or uptake processes.
Once C. albicans cells survive the attacks of the immune cells, the next step is for them to penetrate the endothelial lining of the blood vessels and then colonize the internal organs. The defects of the atp1Δ/Δ in adhesion, invasion, and damage to endothelial cells may reflect an impaired capacity to disseminate through blood vessels during systemic candidiasis. Indeed, C. albicans mutants which have defects in invading and damaging endothelial cells frequently display attenuated virulence during candidiasis in mice (Phan et al., 2000; Sanchez et al., 2004; Chiang et al., 2007; Nobile et al., 2008). Another factor to affect tissue invasion is hyphal formation (Dalle et al., 2010; Zhu and Filler, 2010), which is also required for C. albicans escaping from host macrophages after phagocytosis (Lorenz et al., 2004; McKenzie et al., 2010). Filamentation is the major virulence factor of candidiasis (Lo et al., 1997; Mitchell, 1998). One of the more striking phenotypes of the atp1Δ/Δ is its short filaments under microscopy and minimal colonies on hyphae-inducing media. To date, the direct link between mitochondrial function and morphological switches in C. albicans is still unknown; however, some have suggested that mitochondrial activity regulates the Ras1-cAMP-PKA signaling pathway that is responsible for the yeast-hyphae transition (Grahl et al., 2015).
Biofilm formation provides a useful platform for C. albicans to form robust colonies in the host tissues and protect C. albicans from environmental stresses, such as antifungal agents, oxidative stress and immune attacks (Douglas, 2003; Seneviratne et al., 2008). Moreover, the filaments in biofilm have a strong propensity for tissue invasion (Nett et al., 2010). By consequence of defective filamentation, CSLM imaging reveals that the atp1Δ/Δ forms a rudimentary biofilm that is composed of yeast cells in contrast to a mature biofilm of WT that is made of yeast, pseudohyphae and hyphae. Therefore, the defects of the atp1Δ/Δ on hyphal formation and biofilm may further degrade the virulence of C. albicans in a mice disseminated model.
Atp1p has been found at the surface of hyphae but not in yeast cells (Hernández et al., 2004). Its antigenic property has also been convincingly shown by the detection of Atp1 antibodies both in human sera from systemic candidiasis patients and in serum from mice systemically infected with C. albicans (Pitarch et al., 2004; Martínez-López et al., 2008). Our observation on defective hyphae of the atp1Δ/Δ aligns well with the enrichment of Atp1p at the hyphal phase. However, ATP1 expression in the hyphal form of WT is the same as for the yeast form in this study. Also, the experiments regarding mitochondrial function in this study were all carried out in Candida cells grown in YPD (yeast phase growth), and we speculate that the dysfunctional mitochondria of the mutants would not exclude the possible roles of ATP1 in the yeast growth phase as well. In fact, such a possibility is currently being investigated.
The critical roles of ATP1 in cellular activity have been described previously. In a large scale gene annotation study with transposon mutants, haploinsufficiency of this gene resulted in detectable phenotypes (Oh et al., 2010). This agrees with our findings that intracellular ATP level in reconstituted strain is as low as gene deletion mutant. However, most phenotypes of atp1Δ/ATP1 observed in this study are either compatible to WT or in a manner of intermediate effect, the mechanism of such haploinsufficiency is not yet clear.
In summary, we first identified that the α subunit is essential for pathogenicity in C. albicans. The avirulence phenotype of the atp1Δ/Δ may be caused by a combination of decreased sensitivity to macrophages, together with a reduced capacity to damage endothelial cells and defects in filamentation and biofilm formation.
Author Contributions
SL, YS, and YZ contributed equally to the article; SL, YS, and HZ designed the research plan; SL, YS, YZ, and HW executed the experiments. SL, YS, YZ, HW, HG, KZ, DL, and HZ performed the data analyses and writing. All authors read and approved the final manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (81471995/81171542).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00285/full#supplementary-material
References
Bambach, A., Fernandes, M. P., Ghosh, A., Kruppa, M., Alex, D., Li, D., et al. (2009). Goa1p of Candida albicans localizes to the mitochondria during stress and is required for mitochondrial function and virulence. Eukaryot. Cell 8, 1706–1720. doi: 10.1128/EC.00066-09
Brock, M. (2009). Fungal metabolism in host niches. Curr. Opin. Microbiol. 12, 371–376. doi: 10.1016/j.mib.2009.05.004
Calderone, R., Sun, N., Gay-Andrieu, F., Groutas, W., Weerawarna, P., Prasad, S., et al. (2014). Antifungal drug discovery: the process and outcomes. Future Microbiol. 9, 791–805. doi: 10.2217/fmb.14.32
Chang, P., Fan, X., and Chen, J. (2015). Function and subcellular localization of Gcn5, a histone acetyltransferase in Candida albicans. Fungal Genet. Biol. 81, 132–141. doi: 10.1016/j.fgb.2015.01.011
Chauhan, N., Ciudad, T., Rodríguez-Alejandre, A., Larriba, G., Calderone, R., and Andaluz, E. (2005). Virulence and karyotype analyses of rad52 mutants of Candida albicans: regeneration of a truncated chromosome of a reintegrant strain (rad52/RAD52) in the host. Infect. Immun. 73, 8069–8078. doi: 10.1128/IAI.73.12.8069-8078.2005
Chiang, L. Y., Sheppard, D. C., Bruno, V. M., Mitchell, A. P., and Edwards, J. E. (2007). Candida albicans protein kinase CK2 governs virulence during oropharyngeal candidiasis. Cell Microbiol. 9, 233–245. doi: 10.1111/j.1462-5822.2006.00784.x
Dalle, F., Wächtler, B., L’Ollivier, C., Holland, G., Bannert, N., Wilson, D., et al. (2010). Cellular interactions of Candida albicans with human oral epithelial cells and enterocytes. Cell Microbiol. 12, 248–271. doi: 10.1111/j.1462-5822.2009.01394.x
Douglas, L. J. (2003). Candida biofilms and their role in infection. Trends Microbiol. 11, 30–36. doi: 10.1016/S0966-842X(02)00002-1
Filler, S. G., Ibe, B. O., Luckett, P. M., Raj, J. U., and Edwards, J. E. Jr. (1991). Candida albicans stimulates endothelial cell eicosanoid production. J. Infect. Dis. 164, 928–935.
Fleck, C. B., Schöbel, F., and Brock, M. (2011). Nutrient acquisition by pathogenic fungi: nutrient availability, pathway regulation, and differences in substrate utilization. Int. J. Med. Microbiol. 301, 400–407. doi: 10.1016/j.ijmm.2011.04.007
Frohner, I. E., Bourgeois, C., Yatsyk, K., Majer, O., and Kuchler, K. (2009). Candida albicans cell surface superoxide dismutases degrade host-derived reactive oxygen species to escape innate immune surveillance. Mol. Microbiol. 71, 240–252. doi: 10.1111/j.1365-2958.2008.06528.x
Gillum, A. M., Tsay, E. Y., and Kirsch, D. R. (1984). Isolation of the Candida albicans gene for orotidine-5’-phosphate decarboxylase by complementation of S. cerevisiae ura3 and E. coli pyrF mutations. Mol. Gen. Genet. 198, 179–182.
Grahl, N., Demers, E. G., Lindsay, A. K., Harty, C. E., Willger, S. D., Piispanen, A. E., et al. (2015). Mitochondrial activity and Cyr1 are key regulators of Ras1 activation of C. albicans virulence pathways. PLoS Pathog. 11:e1005133. doi: 10.1371/journal.ppat.1005133
Grubb, S. E., Murdoch, C., Sudbery, P. E., Saville, S. P., Lopez-Ribot, J. L., and Thornhill, M. H. (2008). Candida albicans-endothelial cell interactions: a key step in the pathogenesis of systemic candidiasis. Infect. Immun. 76, 4370–4377. doi: 10.1128/IAI.00332-08
Guo, H., Xie, S. M., Li, S. X., Song, Y. J., Lv, X. L., and Zhang, H. (2014). Synergistic mechanism for tetrandrine on fluconazole against Candida albicans through the mitochondrial aerobic respiratory metabolism pathway. J. Med. Microbiol. 63, 988–996. doi: 10.1099/jmm.0.073890-0
Harold, F. M., and Maloney, P. C. (1996). “Energy transduction by ion currents,” in Escherichia coli and Salmonella: Cellular and Molecular Biology, ed. F. C. Neidhart, R. Curtiss, III, J. L. Ingraham, E. C. C. Lin, K. B. Low, B. Magasanik, et al. (Washington, DC: American Society for Microbiology), k283–306.
Hernández, R., Nombela, C., Diez-Orejas, R., and Gil, C. (2004). Two-dimensional reference map of Candida albicans hyphal forms. Proteomics 4, 374–382. doi: 10.1002/pmic.200300608
Hwang, C. S., Oh, J. H., Huh, W. K., Yim, H. S., and Kang, S. O. (2003). Ssn6, an important factor of morphological conversion and virulence in Candida albicans. Mol. Microbiol. 47, 1029–1043. doi: 10.1046/j.1365-2958.2003.03353.x
Jonckheere, A. I., Renkema, G. H., Bras, M., van den Heuvel, L. P., Hoischen, A., Gilissen, C., et al. (2013). A complex V ATP5A1 defect causes fatal neonatal mitochondrial encephalopathy. Brain 136, 1544–1554. doi: 10.1093/brain/awt086
Krom, B. P., and Willems, H. M. (2016). In vitro models for Candida biofilm development. Methods Mol. Biol. 1356, 95–105. doi: 10.1007/978-1-4939-3052-4_8
Lai-Zhang, J., Xiao, Y., and Mueller, D. M. (1999). Epistatic interactions of deletion mutants in the genes encoding the F1-ATPase in yeast Saccharomyces cerevisiae. EMBO J. 18, 58–64. doi: 10.1093/emboj/18.1.58
Lee, J. E., Oh, J. H., Ku, M., Kim, J., and Lee, J. S. (2015). Ssn6 has dual roles in Candida albicans filament development through the interaction with Rpd31. FEBS Lett. 589, 513–520. doi: 10.1016/j.febslet.2015.01.011
Leroy, O., Gangneux, J. P., Montravers, P., Mira, J. P., Gouin, F., Sollet, J. P., et al. (2009). Epidemiology, management, and risk factors for death of invasive Candida infections in critical care: a multicenter, prospective, observational study in France (2005-2006). Crit. Care Med. 37, 1612–1618. doi: 10.1097/CCM.0b013e31819efac0
Li, D., Chen, H., Florentino, A., Alex, D., Sikorski, P., Fonzi, W. A., et al. (2011). Enzymatic dysfunction of mitochondrial complex I of the Candida albicans goa1 mutant is associated with increased reactive oxidants and cell death. Eukaryot. Cell 10, 672–682. doi: 10.1128/EC.00303-10
Lo, H. J., Köhler, J. R., DiDomenico, B., Loebenberg, D., Cacciapuoti, A., and Fink, G. R. (1997). Nonfilamentous C. albicans mutants are avirulent. Cell 90, 939–949. doi: 10.1016/S0092-8674(00)80358-X
Lopes da Rosa, J., Boyartchuk, V. L., Zhu, L. J., and Kaufman, P. D. (2010). Histone acetyltransferase Rtt109 is required for Candida albicans pathogenesis. Proc. Natl. Acad. Sci. U.S.A. 107, 1594–1599. doi: 10.1073/pnas.0912427107
Lorenz, M. C., Bender, J. A., and Fink, G. R. (2004). Transcriptional response of Candida albicans upon internalization by macrophages. Eukaryot. Cell 3, 1076–1087. doi: 10.1128/EC.3.5.1076-1087.2004
Martínez-López, R., Nombela, C., Diez-Orejas, R., Monteoliva, L., and Gil, C. (2008). Immunoproteomic analysis of the protective response obtained from vaccination with Candida albicans ecm33 cell wall mutant in mice. Proteomics 8, 2651–2664. doi: 10.1002/pmic.200701056
Mayer, F. L., Wilson, D., Jacobsen, I. D., Miramón, P., Große, K., and Hube, B. (2012). The novel Candida albicans transporter Dur31 is a multi-stage pathogenicity factor. PLoS Pathog. 8:e1002592. doi: 10.1371/journal.ppat.1002592
McKenzie, C. G., Koser, U., Lewis, L. E., Bain, J. M., Mora-Montes, H. M., Barker, R. N., et al. (2010). Contribution of Candida albicans cell wall components to recognition by and escape from murine macrophages. Infect. Immun. 78, 1650–1658. doi: 10.1128/IAI.00001-10
Method M27-A3, CLSI (2008). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard, 3rd Edn, CLSI Document M27-A3. Wayne, PA: Clinical and Laboratory Standards Institute.
Mitchell, A. P. (1998). Dimorphism and virulence in Candida albicans. Curr. Opin. Microbiol. 1, 687–692. doi: 10.1016/S1369-5274(98)80116-1
Murad, A. M., Leng, P., Straffon, M., Wishart, J., Macaskill, S., MacCallum, D., et al. (2001). NRG1 represses yeast-hypha morphogenesis and hypha-specific gene expression in Candida albicans. EMBO J. 20, 4742–4752. doi: 10.1093/emboj/20.17.4742
Nett, J. E., Marchillo, K., Spiegel, C. A., and Andes, D. R. (2010). Development and validation of an in vivo Candida albicans biofilm denture model. Infect. Immun. 78, 3650–3659. doi: 10.1128/IAI.00480-10
Nobile, C. J., Andes, D. R., Nett, J. E., Smith, F. J., Yue, F., Phan, Q. T., et al. (2006). Critical role of Bcr1-dependent adhesins in C. albicans biofilm formation in vitro and in vivo. PLoS Pathog. 2:e63. doi: 10.1371/journal.ppat.0020063
Nobile, C. J., Solis, N., Myers, C. L., Fay, A. J., Deneault, J. S., Nantel, A., et al. (2008). Candida albicans transcription factor Rim101 mediates pathogenic interactions through cell wall functions. Cell Microbiol. 10, 2180–2196. doi: 10.1111/j.1462-5822.2008.01198.x
Noble, S. M., French, S., Kohn, L. A., Chen, V., and Johnson, A. D. (2010). Systematic screens of Candida albicans homozygous deletion library decouple morphogenetic switching and pathogenicity. Nat. Genet. 42, 590–598. doi: 10.1038/ng.605
Oh, J., Fung, E., Schlecht, U., Davis, R. W., Giaever, G., and St Onge, R. P. (2010). Gene annotation and drug target discovery in Candida albicans with a tagged transposon mutant collection. PLoS Pathog. 7:e1001140. doi: 10.1371/journal.ppat.1001140
Park, H., Myers, C. L., Sheppard, D. C., Phan, Q. T., Sanchez, A. A., E Edwards, J., et al. (2005). Role of the fungal Ras-protein kinase A pathway in governing epithelial cell interactions during oropharyngeal candidiasis. Cell Microbiol. 7, 499–510. doi: 10.1111/j.1462-5822.2004.00476.x
Peeters, E., Nelis, H. J., and Coenye, T. (2008). Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 72, 157–165. doi: 10.1016/j.mimet.2007.11.010
Phan, Q. T., Belanger, P. H., and Filler, S. G. (2000). Role of hyphal formation in interactions of Candida albicans with endothelial cells. Infect. Immun. 68, 3485–3490.
Phan, Q. T., Myers, C. L., Fu, Y., Sheppard, D. C., Yeaman, M. R., Welch, W. H., et al. (2007). Als3 is a Candida albicans invasin that binds to cadherins and induces endocytosis by host cells. PLoS Biol. 5:e64. doi: 10.1371/journal.pbio.0050064
Pierce, C. G., Uppuluri, P., Tristan, A. R., Wormley, F. L. Jr., Mowat, E., Ramage, G., et al. (2008). A simple and reproducible 96-well plate-based method for the formation of fungal biofilms and its application to antifungal susceptibility testing. Nat. Protoc. 3, 1494–1500. doi: 10.1038/nport.2008.141
Pitarch, A., Abian, J., Carrascal, M., Sánchez, M., Nombela, C., and Gil, C. (2004). Proteomics-based identification of novel Candida albicans antigens for diagnosis of systemic candidiasis in patients with underlying hematological malignancies. Proteomics 4, 3084–3106. doi: 10.1002/pmic.200400903
Sanchez, A. A., Johnston, D. A., Myers, C., Edwards, J. E. Jr., Mitchell, A. P., and Filler, S. G. (2004). Relationship between Candida albicans virulence during experimental hematogenously disseminated Infection and endothelial cell damage in vitro. Infect. Immun. 72, 598–601. doi: 10.1128/IAI.72.1.598-601.2004
Sasse, C., and Morschhäuser, J. (2012). Gene deletion in Candida albicans wild-type strains using the SAT1-flipping strategy. Methods Mol. Biol. 845, 3–17. doi: 10.1007/978-1-61779-539-8_1
Seneviratne, C. J., Wang, Y., Jin, L., Abiko, Y., and Samaranayake, L. P. (2008). Candida albicans biofilm formation is associated with increased anti-oxidative capacities. Proteomics 8, 2936–2947. doi: 10.1002/pmic.200701097
Senior, A. E. (1990). The proton-translocating ATPase of Escherichia coli. Annu. Rev. Biophys. Biophys. Chem. 19, 7–41. doi: 10.1146/annurev.bb.19.060190.000255
She, X., Khamooshi, K., Gao, Y., Shen, Y., Lv, Y., Calderone, R., et al. (2015). Fungal-specific subunits of the Candida albicans mitochondrial complex I drive diverse cell functions including cell wall synthesis. Cell Microbiol. 17, 1350–1364. doi: 10.1111/cmi.12438
Spellberg, B., Ibrahim, A. S., Edwards, J. E. Jr., and Filler, S. G. (2005). Mice with disseminated candidiasis die of progressive sepsis. J. Infect. Dis. 192, 336–343. doi: 10.1086/430952
Spellberg, B., Johnston, D., Phan, Q. T., Edwards, J. E. Jr., French, S. W., Ibrahim, A. S., et al. (2003). Parenchymal organ, and not splenic, immunity correlates with host survival during disseminated candidiasis. Infect. Immun. 71, 5756–5764. doi: 10.1128/IAI.71.10.5756-5764.2003
Sun, J. N., Solis, N. V., Phan, Q. T., Bajwa, J. S., Kashleva, H., Thompson, A., et al. (2010). Host cell invasion and virulence mediated by Candida albicans Ssa1. PLoS Pathog. 6:e1001181. doi: 10.1371/journal.ppat.1001181
Sun, N., Fonzi, W., Chen, H., She, X., Zhang, L., Zhang, L., et al. (2013). Azole susceptibility and transcriptome profiling in Candida albicans mitochondrial electron transport chain complex I mutants. Antimicrob. Agents Chemother. 57, 532–542. doi: 10.1128/AAC.01520-12
Wächtler, B., Wilson, D., Haedicke, K., Dalle, F., and Hube, B. (2011). From attachment to damage: defined genes of Candida albicans mediate adhesion, invasion and damage during interaction with oral epithelial cells. PLoS ONE 6:e17046. doi: 10.1371/journal.pone.0017046
Walker, L. A., Maccallum, D. M., Bertram, G., Gow, N. A., Odds, F. C., and Brown, A. J. (2009). Genome-wide analysis of Candida albicans gene expression patterns during infection of the mammalian kidney. Fungal Genet. Biol. 46, 210–219. doi: 10.1016/j.fgb.2008.10.012
Wisplinghoff, H., Ebbers, J., Geurtz, L., Stefanik, D., Major, Y., Edmond, M. B., et al. (2014). Nosocomial bloodstream infections due to Candida spp. in the USA: species distribution, clinical features and antifungal susceptibilities. Int. J. Antimicrob. Agents 43, 78–81. doi: 10.1016/j.ijantimicag.2013.09.005
Yu, Q., Zhang, B., Yang, B., Chen, J., Wang, H., Jia, C., et al. (2014). Interaction among the vacuole, the mitochondria, and the oxidative stress response is governed by the transient receptor potential channel in Candida albicans. Free Radic. Bio. Med. 77, 152–167. doi: 10.1016/j.freeradbiomed.2014.09.011
Zhang, X., Guo, H., Gao, L., Song, Y., Li, S., and Zhang, H. (2013). Molecular mechanisms underlying the tetrandrine-mediated reversal of the fluconazole resistance of Candida albicans. Pharm. Biol. 51, 749–752. doi: 10.3109/13880209.2013.764537
Keywords: ATP1, α subunit, Candida albicans, pathogenicity, virulence properties
Citation: Li S-X, Song Y-J, Zhang Y-S, Wu H-T, Guo H, Zhu K-J, Li D-M and Zhang H (2017) Mitochondrial Complex V α Subunit Is Critical for Candida albicans Pathogenicity through Modulating Multiple Virulence Properties. Front. Microbiol. 8:285. doi: 10.3389/fmicb.2017.00285
Received: 01 January 2017; Accepted: 13 February 2017;
Published: 23 February 2017.
Edited by:
Dominique Sanglard, University of Lausanne, SwitzerlandReviewed by:
David Kadosh, University of Texas Health Science Center at San Antonio, USADerek Sullivan, Dublin Dental University Hospital, Ireland
Copyright © 2017 Li, Song, Zhang, Wu, Guo, Zhu, Li and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kun-Ju Zhu, emh1a3VuanUxMTExQDE2My5jb20= Dong-Mei Li, ZGwzM0BnZW9yZ2V0b3duLmVkdQ== Hong Zhang, dHpoYW5naEBqbnUuZWR1LmNu
†These authors have contributed equally to this work.