 Aida Kalantari1,2
Aida Kalantari1,2 Tao Chen1,3
Tao Chen1,3 Boyang Ji1
Boyang Ji1 Ivan A. Stancik1,4
Ivan A. Stancik1,4 Vaishnavi Ravikumar1
Vaishnavi Ravikumar1 Damjan Franjevic4Claire Saulou-Bérion5
Damjan Franjevic4Claire Saulou-Bérion5 Anne Goelzer6
Anne Goelzer6 Ivan Mijakovic1,7*
Ivan Mijakovic1,7*- 1Systems and Synthetic Biology Division, Department of Biology and Biological Engineering, Chalmers University of Technology, Gothenburg, Sweden
- 2Chaire Agro-Biotechnologies Industrielles, AgroParisTech, Reims, France
- 3Key Laboratory of Systems Bioengineering (Ministry of Education), Tianjin University, Tianjin, China
- 4Department of Biology, Faculty of Science, University of Zagreb, Zagreb, Croatia
- 5UMR Génie et Microbiologie des Procédés Alimentaires (GMPA), AgroParisTech, Institut National de la Recherche Agronomique, Université Paris-Saclay, Thiverval Grignon, France
- 6Mathématiques et Informatique Appliquuées du Génome à l’Environnement (MaIAGE), Institut National de la Recherche Agronomique, Université Paris-Saclay, Jouy-en-Josas, France
- 7Novo Nordisk Foundation Center for Biosustainability, Technical University of Denmark, Lyngby, Denmark
3-Hydroxypropanoic acid (3-HP) is an important biomass-derivable platform chemical that can be converted into a number of industrially relevant compounds. There have been several attempts to produce 3-HP from renewable sources in cell factories, focusing mainly on Escherichia coli, Klebsiella pneumoniae, and Saccharomyces cerevisiae. Despite the significant progress made in this field, commercially exploitable large-scale production of 3-HP in microbial strains has still not been achieved. In this study, we investigated the potential of Bacillus subtilis as a microbial platform for bioconversion of glycerol into 3-HP. Our recombinant B. subtilis strains overexpress the two-step heterologous pathway containing glycerol dehydratase and aldehyde dehydrogenase from K. pneumoniae. Genetic engineering, driven by in silico optimization, and optimization of cultivation conditions resulted in a 3-HP titer of 10 g/L, in a standard batch cultivation. Our findings provide the first report of successful introduction of the biosynthetic pathway for conversion of glycerol into 3-HP in B. subtilis. With this relatively high titer in batch, and the robustness of B. subtilis in high density fermentation conditions, we expect that our production strains may constitute a solid basis for commercial production of 3-HP.
Introduction
Concerns over usage of fossil fuels have led to the development of sustainable technologies to convert renewable materials into various compounds such as alcohols, acids, and chemicals (Hanai et al., 2007; Atsumi and Liao, 2008; Atsumi et al., 2008; McKenna and Nielsen, 2011; Borodina et al., 2015). One of the very important biomass-derivable platform chemicals is 3-hydroxypropanoic acid (3-HP; Choi et al., 2015). 3-HP can be used in the synthesis of bulk chemicals, such as the acrylic acid (patent: WO 2013192451 A1), and plastics (Valdehuesa et al., 2013). Production of 3-HP by chemical conversion is not desirable due to the high cost and toxic intermediates leading to environmental issues (Jiang et al., 2009; Kumar et al., 2013). Glycerol, a by-product of the biodiesel industry, is an attractive renewable starting material for production of 3-HP (Chen and Nielsen, 2016). The main bacterial pathway for the synthesis of 3-HP from glycerol consists of two enzymatic reactions. The first is catalyzed by a coenzyme B12-dependent glycerol dehydratase, which converts glycerol into 3-hydroxypropionaldehyde (3-HPA). The second reaction is the conversion of 3-HPA to 3-HP, catalyzed by an aldehyde dehydrogenase (Raj et al., 2008). This biosynthetic pathway is present in some natural producers of 3-HP, such as Klebsiella pneumoniae (Kumar et al., 2012; Ashok et al., 2013) and Lactobacillus reuteri (patent: US 20070148749 A1). The pathway has been heterologously expressed and studied in Escherichia coli and Saccharomyces cerevisiae (patent: WO 2011038364 A1; Chen et al., 2014; Jung et al., 2015; Kildegaard et al., 2015). Overexpression of the E. coli aldH gene in K. pneumoniae resulted in co-production of 1,3-propanediol (1,3-PDO) and 3-HP with a 3-HP titer of 24.4 g/L (Huang et al., 2012). In another study, 48.9 g/L of 3-HP was obtained in microaerobic fed-batch fermentation of K. pneumoniae WM3 expressing aldH from E. coli k12 (Huang et al., 2013). The recombinant E. coli BL21 expressing dhaB and gdrAB from K. pneumoniae and KGSADH from Azospirillum brasilense produced 38.7 g/L of 3-HP in fed-batch condition (Rathnasingh et al., 2009). In another report, deleting genes responsible for by products redirected the flux toward the 3-HP and resulted in higher titer of 57.3 g/L of 3-HP in fed-batch (Kim et al., 2014). Until now, the highest reported titer of 3-HP is 71.9 g/L using mutated aldehyde dehydrogenase (ALDH) (GabD4) from Cupriavidus necator in E. coli (Chu et al., 2015). Despite the noticeable progress in this area, microbial production of 3-HP has not yet reached industrial-scale productivity levels. Here we explored the possibility to use and alternative host for heterologous expression of the 3-HP synthetic pathway, the Gram-positive model organism Bacillus subtilis. It is non-pathogenic, generally recognized as safe (GRAS), suitable for large scale cultivation and has high growth rates. B. subtilis efficiently imports glycerol via a selective glycerol facilitator (GlpF; da Silva et al., 2009), and reaches a high growth rate on glycerol (μmax = 0.65 h-1; Kruyssen et al., 1980). When expressing the 3-HP synthetic pathway in a new host, the tolerance level toward 3-HP (Kildegaard et al., 2014) and the intermediate 3-HPA (Hao et al., 2008) can be a major consideration, resolved by a balanced expression of the two steps in the pathway (Sankaranarayanan et al., 2014). We expressed the 3-HP synthetic pathway from K. pneumoniae in B. subtilis. Since this particular glycerol dehydratase is sensitive to oxygen, micro-aerobic/anaerobic condition were required for optimal 3-HP production (Zhao et al., 2015). We carried out several rounds of strain and medium optimization, which resulted in a production strain capable of producing 10 g/L of 3-HP in shaking flasks, with the yield on glycerol ranging from 70 to 80%.
Materials and Methods
Strains and Plasmids
All the strains and plasmids used in this study are listed in Table 1. B. subtilis 168 trp+ was used as the host for genetic manipulations. Integrative plasmid pBS1C (BGSC, Columbus, USA) was used to clone dhaB, gdrAB, and puuC from K. pneumoniae. The codon-optimized construct containing the genes dhaB123, gdrAB, and puuC from K. pneumoniae was ordered as synthetic fragment (GenScript, Piscataway, NJ, USA) and cloned in the plasmid pBS1C. E. coli DH5α was used to amplify plasmids listed in Table 1 and the B. subtilis 168 trp+ cells were transformed with the same plasmids using a previously described method (Fabret et al., 2002).

TABLE 1. Overview of the plasmids and strains used in this study.
Genetic Manipulations
All synthetic genes (codon optimized) were under control of pHyperspank, a strong isopropyl β-D-1-thiogalactopyranoside (IPTG)-inducible promoter of B. subtilis. A strong ribosome binding site (RBS) from B. subtilis (ACATAAGGAGGAACTACT; kindly provided by Dominique Le Coq, INRA, France) was added before each synthetic fragment. The entire construct was ligated into the pBS1C integrative plasmid (BGSC, Columbus, USA) as an EcoRI–PstI fragment. The integrative construct was used to transform B. subtilis 168 trp+. The Δglpk strains were constructed either by using the single crossing over method described by Vagner et al. (1998) or a seamless and irreversible gene inactivation method (Arnaud et al., 2004). In the former, the pMUTIN2 plasmid carrying a portion of glpK is integrated into the B. subtilis genome through single cross-over event, resulting in disrupting the glpK gene. In the latter, the pMAD plasmid carrying a portion of the glpK gene is integrated at glpK locus in the B. subtilis genome in the first cross-over event, and the seamless and irreversible glpK knockout is obtained after the second single cross-over event. Stability of the recombinant constructs was tested by PCR using appropriate primers. All recombinant plasmids were cloned in E. coli DH5α and used to transform B. subtilis 168 trp+. All constructs were verified by restriction digestion and sequencing.
Media and Growth
Escherichia coli strains and wild type (WT) B. subtilis were cultured in Luria–Bertani (LB) broth at 37°C in shaking flask (200 rpm) for genetic manipulations. For E. coli strains, ampicillin was added to the medium when needed at 100 μg/mL. Recombinant B. subtilis strains were grown in the minimal medium (M9; Harwood and Cutting, 1990) complemented with 12 g/L glycerol, 12 g/L glucose, and/or 1 g/L yeast extract, when required, at 37°C in shaking flask (200 rpm). Chloramphenicol 5 mg/mL and erythromycin 1 mg/mL were used when required to select B. subtilis transformants. Coenzyme B12 (15 μM) and IPTG (500 μM) were added in the medium. Recombinant B. subtilis strains were cultured in rich media used in shaking flask which was composed of 2 × M9: 40 g/L glycerol, 10 g/L yeast extract, and 3 g/L peptone. Seeds medium contained M9Y (M9 medium supplemented with 1 g/L yeast extract) with addition of 1% glucose. Cultivation medium contained M9Y and/or 2 × M9Y with addition of different concentration of glucose and glycerol (the initial concentration for glucose was 0.8% and for glycerol was about 2.7%). For batch cultivation in flask, a single colony was transferred into 50 mL falcon tube containing 10 mL seeds medium and cultured in shaker at 37°C and 200 rpm. Once the cell OD600 reached about 2, 1 mL of seeds culture was inoculated into 250 mL flask containing 35 mL cultivation medium. In fed-batch cultivation in flask, five glucose tablets (FeedBead Glucose discs) were added in each flask to release the glucose slowly in the medium. Flasks were shaken at 37°C and 200 rpm, and were kept in darkness. B. subtilis transformants were selected using chloramphenicol 5 mg/mL and erythromycin 1 mg/mL when required. Coenzyme B12 (15 μM) and IPTG (500 μM) were added in the medium. 3-HP production was followed in this setup for 2–4 days.
Characterizing the Effect of 3-HP and 3-HPA on B. subtilis Growth
WT B. subtilis cells were grown in the M9 medium with 12 g/L glycerol as the sole carbon source. 3-HP was purchased from TCI Europe N.V. (Zwijndrecht, Belgium), and 3-HPA was chemically synthesized (Burgé et al., 2015a). 3-HP was tested in a range between 1 and 100 mM, while 3-HPA was tested between 0.02 and 20 mM. OD600 of the bacterial culture was continuously monitored for 24 h using a 96-well plate reader (Biotek, USA), at 37°C, with three biological replicates.
Proteomics Analysis
Cells were collected at four different time points during fermentation: 0, 24, 48, and 72 h. Cell pellets were re-suspended in an sodium dodecyl sulfate (SDS) lysis buffer containing 4% SDS in 100 mM triethylammonium bicarbonate pH 8.0, 10 mM ethylenediaminetetraacetic acid and a protease inhibitor cocktail (Roche). The cell extract was boiled at 90°C for 10 min followed by sonication for 30 s. The cell debris was removed by centrifugation. The protein pellet obtained after chloroform/methanol precipitation was dissolved in denaturation buffer containing 8 M urea in 10 mM Tris–HCl pH 8.0. Protein concentration was measured by Bradford protein assay. A total of 100 μg of each sample were separated on a Mini-Protean® TGXTM 4–20% gradient gel (Bio-Rad) and stained with Bio-SafeTM Coomassie (Bio-Rad). Regions corresponding to the size of “DhaB1” were cut and in-gel trypsin (PierceTM) digestion was performed as described (Shevchenko et al., 2007). Peptides were desalted using C-18 stage-tips and analyzed on the Orbitrap FusionTM TribridTM (Thermo Fischer Scientific). An inclusion list consisting of m/z values in the range of 400–1600 Da and charge states +2 and +3 only was incorporated into the instrument method. Acquired mass spectra were processed with the MaxQuant software suite (v.1.5.1.0; Cox et al., 2009).
In silico Simulation of the Metabolism
The genome-scale metabolic model iYO844 was obtained in a COBRA Toolbox (Schellenberger et al., 2011) compatible SBML format from http://systemsbiology.ucsd.edu. Two reactions (ALCD19_D and ALCD19_L) involving in alcohol dehydrogenases were removed from the model due to the determined roles of YhdN in methylglyoxal resistance (Chandrangsu et al., 2014). Although YhcW and YvoE had been identified as glycerol-3-phosphatase (Lindner et al., 2012), we did not include corresponding reaction into the model due to the low in vivo activity of these enzymes (data not shown). The relevant reactions for 3-HP production (Supplementary Figure S5) were added to the model. The 3-HP production envelopes were generated using COBRA toolbox in Matlab (MathWorks, Inc., Natick, MA, USA). For the simulation of aerobic growth in minimal medium M9, the carbon uptake (glycerol and/or glucose) had a limit of -10 mmol gDW-1 h-1 as the maximum uptake rate. Aerobic conditions were simulated by setting the lower and upper limits for the O2 exchange flux to free (-1000 to 1000 mmol gDW-1 h-1). In addition, the uptake rates of sulfate and ammonium were set to free, as were the K+, Na+, Mg2+, Ca2+, Fe3+, CO2, H2O, and H+ uptakes. For reaction knockout simulations, the minimum and maximum fluxes of the corresponding reactions were set to 0. The reactions available to knockout was reduced to a pool of approximately 400 reactions that excluded exchange reactions, transporter reactions, and essential reactions.
Metabolite Analyses
Glycerol, 3-HP, and lactate concentrations were measured using high performance liquid chromatography (HPLC) (Ultimate 3000, Dionex). 1 mL of culture samples were centrifuged at 12,000 × g for 10 min, then 200 μL of 8 mM H2SO4 was added to 800 μL of the supernatant and filtered through a 0.20 μm pore size nitrocellulose filter (Sartorius Stedim). Separation was performed on a Bio-Rad Aminex HPX-87H column (300 mm × 7.8 mm; Bio-Rad, Richmond, USA) connected to a VWD-3100 detector (Thermo Scientific Dionex) with 0.5 or 8 mM H2SO4 as the eluent, as described by Burgé et al. (2015b). All samples were analyzed in three replicates. Gas chromatography mass spectrometry (GC-MS) was also used to quantify the 3-HP and lactate. All samples were derivatized with methyl chloroformate. For quality control, a mixed pooled sample (QC sample) was created by taking an aliquot from each sample. Testing of matrix effects was performed by spiking/dilution of QC samples. The GC-MS data were processed by PARAFAC2 model from MS-Omics1.
Results and Discussion
Sensitivity of B. subtilis to 3-HPA and 3-HP
The product of the first step of conversion of glycerol to 3-HP is 3-HPA, known to be toxic to some bacteria (Hao et al., 2008). We examined the effect of 3-HPA and 3-HP on B. subtilis growing on the M9 minimum medium supplemented with glycerol. The 3-HPA exhibited a strong inhibitory effect from very low concentrations. The specific growth rate in the 0–0.1 mM range (red-colored group, Figure 1A) was 0.55 h-1. It decreased to 0.27 h-1 for the 0.2–1 mM range (green-colored group, Figure 1A), and the growth was completely abolished in the 2–20 mM range (blue-colored group, Figure 1A). The lag phase was also prolonged as the concentration of 3-HPA increased (9:30 h, 14:30 h, and 20 h for the three groups, respectively). B. subtilis cells were much more tolerant toward 3-HP. The specific growth rate in the 0–35 mM range (red-colored group, Figure 1B) was 0.59 h-1. It decreased slightly to 0.56 h-1 for the 50–80 mM range (green-colored group, Figure 1B) and further to μ = 0.54 h-1 for the 90–100 mM range (blue-colored group, Figure 1B). The lag phase was also affected as the concentration of 3-HP increased (7:30 h, 9:30 h, and 13:30 h for the three groups, respectively). It is important to note that the 3-HP sample was buffered, so the growth effects were not due to the acidification of the medium. Regarding the tolerance level of other microorganisms toward 3-HP, is has been reported that E. coli has normal growth in M9 with addition of 100 mM of 3-HP, while the growth significantly decreases with 3-HP concentrations in the range of 330–440 mM (Chun et al., 2014). There has also been a report on S. cerevisiae strains tolerant to 50 g/L 3-HP obtained through adaptive laboratory evolution (Kildegaard et al., 2014). Our results indicate that B. subtilis is fairly tolerant to 3-HP, but highlight the importance of preventing the accumulation of the 3-HPA in the cell.
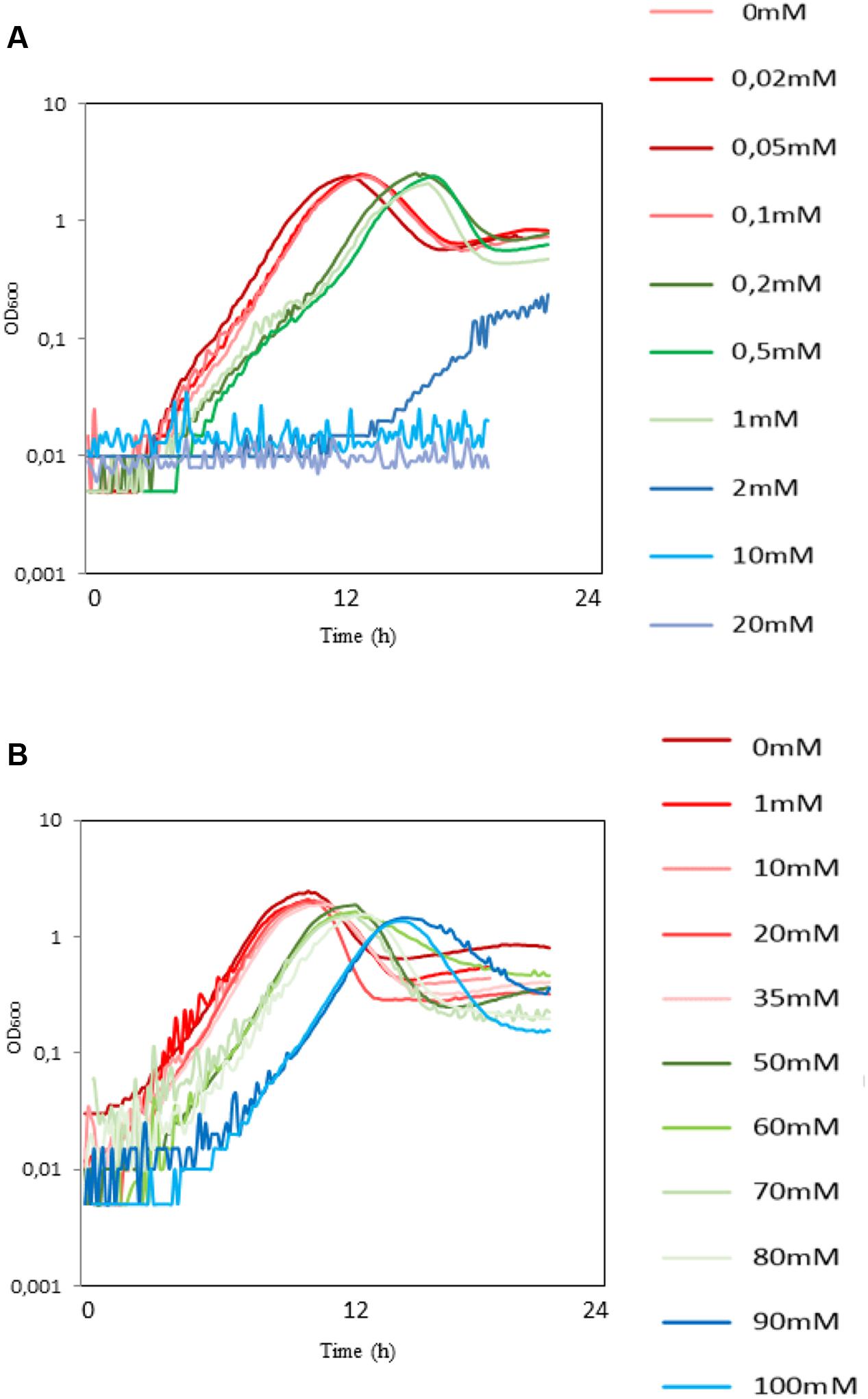
FIGURE 1. The toxic effect of 3-HPA and 3-HP on the growth of wild type (WT) B. subtilis. Growth curves of the WT B. subtilis growing for 24 h on the M9 minimal medium with glycerol as carbon source supplemented with (A) 3-HPA (0.02–20 mM) and (B) 3-HP (1–100 mM). Both growth rate and the duration of lag phase was affected as the concentration of either 3-HPA or 3-HP increased which are classified in three groups of red, green, and blue colors representing the effect of either of the compounds on growth of WT B. subtilis from low to high, respectively. Respective concentrations of 3-HP and 3-HPA are indicated next to each curve.
Expression of the Codon-optimized 3-HP Pathway from K. pneumoniae Leads to the Production of 3-HP
The dhaB and puuC, coding for the B12-dependent glycerol dehydratase and aldehyde dehydrogenase from K. pneumoniae, were codon optimized for B. subtilis (GenScript). The optimization concerned 23% of the codons. All modified codons can be seen in an alignment of non-optimized and optimized sequences provided in Supplementary Figure S1. The synthetic genes were expressed in B. subtilis from a single construct in pBS1C-3, resulting in the strain h-syn-KpDhaB-PuuC. The entire construct was under the control of a strong promoter pHyperspank (Vagner et al., 1998), and each gene was preceded by the strong RBS (ACATAAGGAGGAACTACT). The B. subtilis strain containing the empty pBS1C plasmid (pBS1C-E strain) was used as the control. We confirmed the expression of the glycerol dehydratase and aldehyde dehydrogenase in the h-syn-KpDhaB-PuuC crude extract by mass spectrometry (MS/MS spectra shown in Supplementary Figure S2), which were detected with excellent coverage and no ambiguity. The h-syn-KpDhaB-PuuC and pBS1C-E strains were cultured and samples were collected at regular intervals after induction to measure the growth rate, glycerol consumption and 3-HP production (Figure 2). The growth rates (Supplementary Figure S3) and glycerol consumption (Figure 2) were comparable for both strains. After 24 h, 3-HP was detected in the h-syn-KpDhaB-PuuC culture at 180 mg/L (Figure 2A). As expected, no 3-HP was detected in the control strain pBS1C-E (Figure 2B). Growth profiles of h-syn-KpDhaB-PuuC and pBS1C-E are shown in Figure 2C. Quantification of 3-HP was confirmed by an independent method using GC-MS (Supplementary Figure S4). Since the growth rate was not affected by the expression of the heterologous pathway, we concluded that 3-HPA is not accumulating in the production strain, and no further balancing of the expression of the two reaction steps was needed. The B12-dependent DhaB from K. pneumoniae is sensitive to excessive oxygen (Zhao et al., 2015), so we compared the production of 3-HP in micro-aerobic and fully aerobic conditions (Figure 3A) and their respective growth profile (Figure 3B). In the semi-aerobic condition the h-syn-KpDhaB-PuuC clearly performed better, so this condition was used throughout the following optimization steps. These results altogether indicate that the codon-optimized synthetic version of the 3-HP biosynthetic pathway was correctly expressed and functional in B. subtilis. There has been a report on positive effect of glucose addition to the 3-HP producing strain of E. coli grown on glycerol, increasing the 3-HP production by reducing the imbalance between the first and second step of the pathway (Niu et al., 2016). To test whether this effect applies to our strain, we grew the h-syn-KpDhaB-PuuC in two conditions: (1) 1.2% glycerol as the sole carbon source and (2) 1.2% glycerol supplemented with 1.2% glucose. Our results showed no significant increase in 3-HP production when adding glucose to the culture. The 3-HP production was identical in both conditions (approximately 200 mg/L with less than 3% difference). We concluded that this effect does not apply to our production strain.
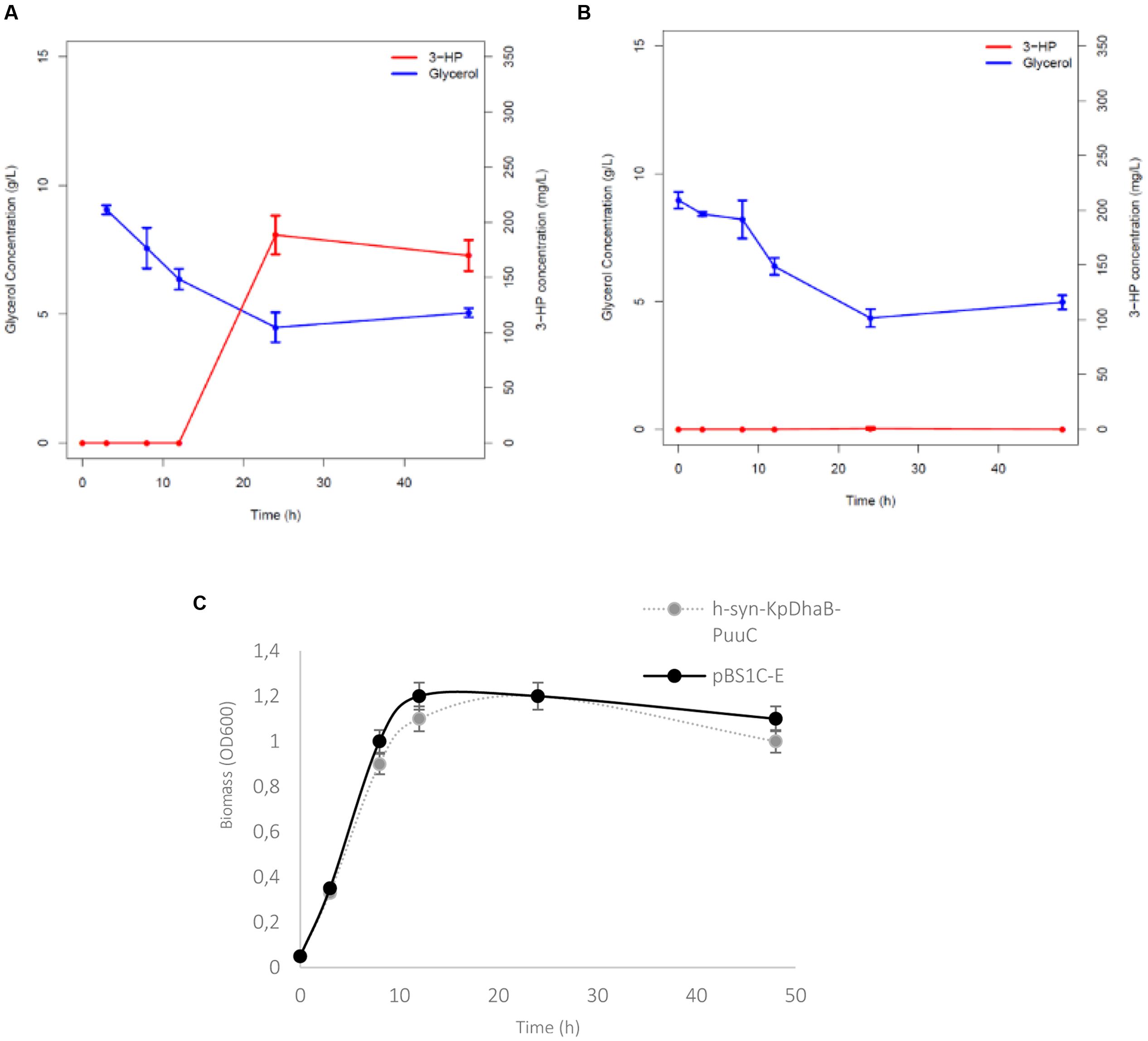
FIGURE 2. Time course of glycerol consumption and 3-HP production. (A) Recombinant strain overexpressing codon-optimized glycerol dehydratase and its activators (dhaB123, gdrAB) and the aldehyde dehydrogenase (PuuC) from K. pneumoniae under control of pHyperspank promoter (h-syn-KpDhaB-PuuC) and (B) the control strain with the empty plasmid (pBS1C-E), cultivated in M9 medium with 12 g/L glycerol, induced by IPTG and B12. (C) Growth profiles of h-syn-KpDhaB-PuuC and pBS1C-E strains. Glycerol consumption is shown in blue, and 3-HP production in red. Average values from three independent biological replicates are shown, with the standard deviation.
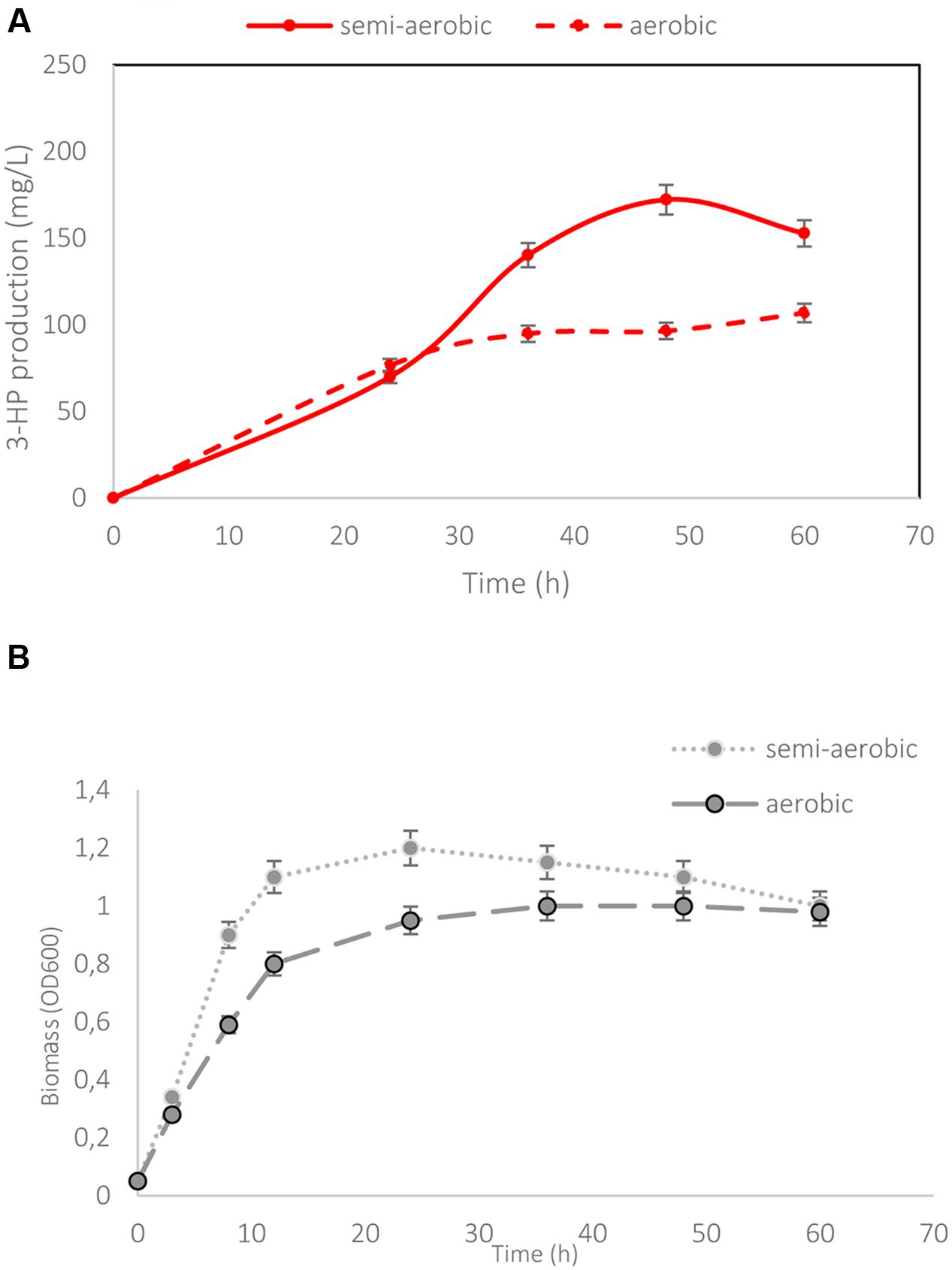
FIGURE 3. Effect of the aerobic and semi-aerobic condition on 3-HP production. (A) Cultivation of the strain h-syn-KpDhaB-PuuC in aerobic condition leads to decreased 3-HP production compared to semi-aerobic condition. (B) Growth profile of h-syn-KpDhaB-PuuC in aerobic and semi-aerobic condition. 3-HP production is represented in solid and dotted lines for semi-aerobic and aerobic conditions, respectively. Average values from two independent biological replicates are shown, with the standard deviation.
In silico Identification of Targets for Increased Production of 3-HP
The genome-scale metabolic model of B. subtilis (iYO844; Oh et al., 2007), with the addition of the heterologous 3-HP synthetic pathway (Supplementary Figure S5 and Figure 4A) was used to predict targets for optimizing the 3-HP production. As shown in Figure 4B, the predicted maximum growth rate was achieved when using glucose as the sole carbon source, while the predicted maximum 3-HP production rate was reached when using glycerol as the sole carbon source. When growing on both glucose and glycerol, the flux of glycerol to 3-HP biosynthesis is negatively related to the flux of glycerol to biomass, indicating that the reduction of glycerol conversion to biomass should improve the 3-HP yield. Our in silico analysis (Supplementary Table S1) was performed to identify single gene knockouts which are expected to improve 3-HP production under these conditions. As expected from the literature (Jung et al., 2014; Tsuruno et al., 2015), the deletion of the gene glpK, which encodes glycerol kinase, was the top candidate for redirecting the metabolic flux toward 3-HP biosynthesis when grown on a combination of glucose and glycerol. The glpK strain was the only knockout predicted to achieve high productivity, while maintaining an acceptable growth rate.
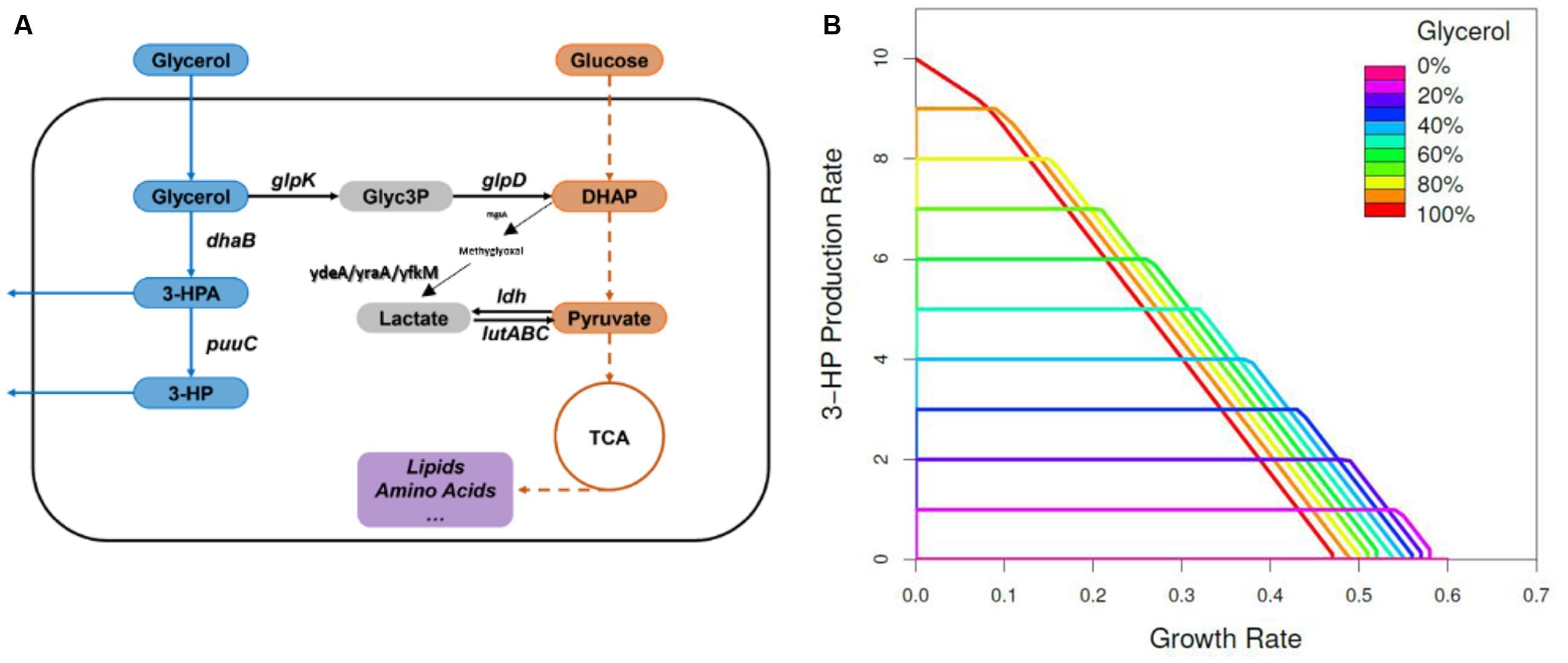
FIGURE 4. The metabolic pathways involved in 3-HP biosynthesis and prediction of maximum 3-HP production rate. (A) Biosynthesis of 3-HP was achieved by integration of a heterologous metabolic route into B. subtilis. The metabolite abbreviations are as follows: Glyc3P, glycerol 3-phosphate; DHAP, dihydroxyacetone phosphate. The gene names are as follows: glpK, glycerol kinase; glpD, glycerol-3-phosphate dehydrogenase; dhaB, glycerol dehydratase; dhaS, aldehyde dehydrogenase; ldh, L-lactate dehydrogenase; lutABC, lactate catabolic; mgsA, methylglyoxal synthase. (B) Predicted maximum 3-HP production rate limits as a function of glycerol proportion in the total carbon source, in the glucose- and glycerol-fed cultures, based on the iYO844 GEM. The color scale denotes the ratio of glycerol in the carbon source from 0 (velvet) to 100 (red).
Inactivation of glpK Leads to Increased 3-HP Production
GlpK catalyzes the formation of glycerol-3-phosphate from glycerol, which is further converted to dihydroxyacetone phosphate to enter glycolysis (Holmberg et al., 1990). Based on the in silico prediction presented above, the glpK gene was knocked out using the method reported by Vagner et al. (1998) resulting in strains h-syn-KpDhaB-PuuC-ΔglpK-i and pBS1C-E-ΔglpK, respectively. This deletion naturally suppressed the growth on glycerol as the sole carbon source (data not shown). We therefore cultured the strains h-syn-KpDhaB-PuuC-ΔglpK-i and pBS1C-E-ΔglpK in the M9 minimum medium supplemented with glucose up to the induction point at OD600 = 1.2. 3-HP was detected in the h-syn-KpDhaB-PuuC-ΔglpK-i culture 8 h after induction (50 mg/L) and its concentration increased to the maximum of 1 g/L after 60 h (Figure 5A). As expected, non-induced and pBS1C-E-Δglpk produced no 3-HP (Supplementary Figure S6). The glycerol uptake was not reduced in h-syn-KpDhaB-PuuC-ΔglpK-i (Figure 5A) compared to the control strain (Supplementary Figure S6A), indicating that the glpK knockout successfully redirected the flux toward 3-HP production. Next, we increased the glycerol concentration to 25 g/L, but this failed to increase the 3-HP titer (Figure 5B). This indicated that one (or more) of the pathway components does not operate at the maximum rate.
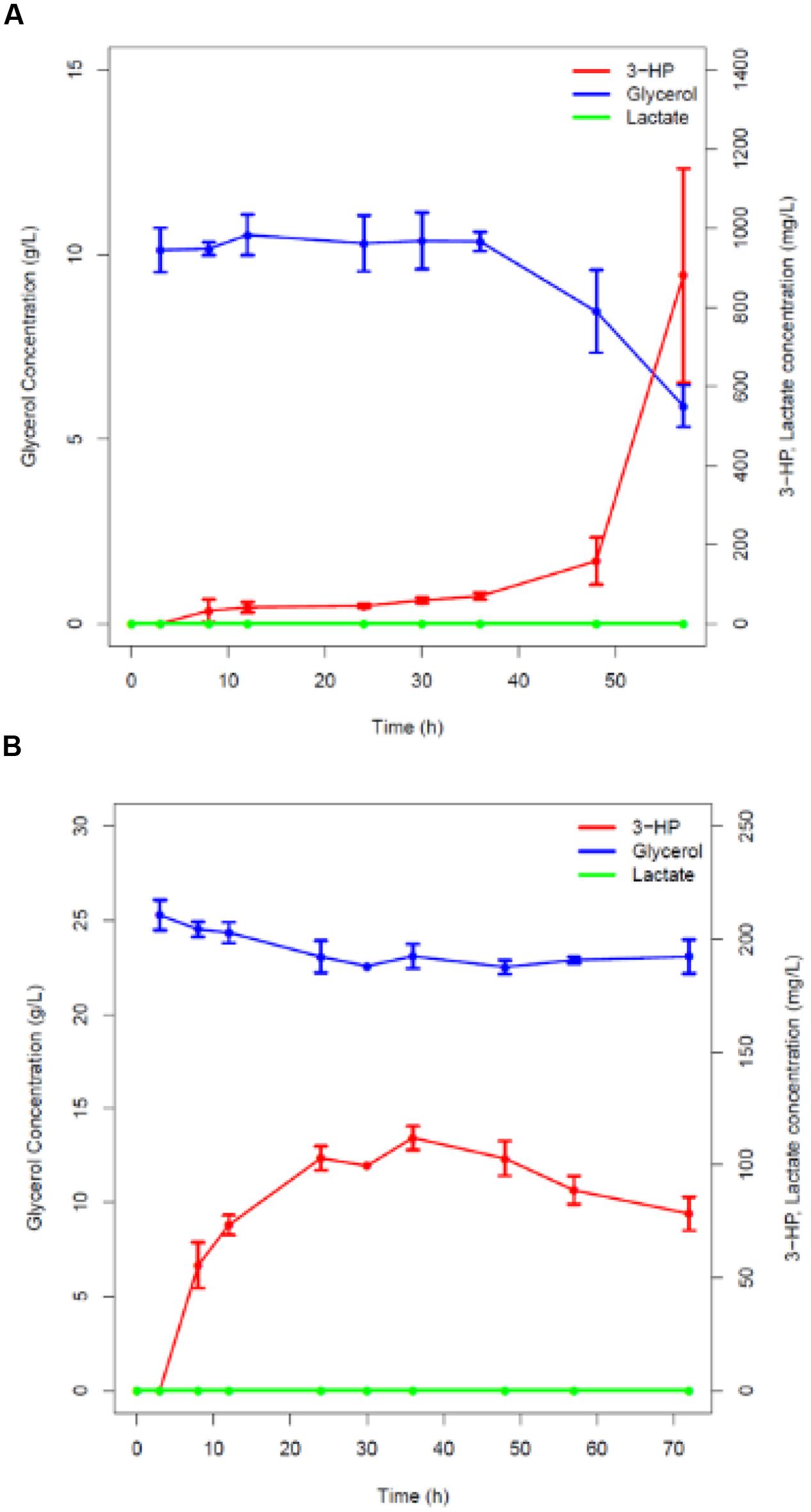
FIGURE 5. Time course of glycerol consumption and lactate and 3-HP production. (A) Recombinant strain with a knocked-out glycerol kinase (glpk) overexpressing codon-optimized glycerol dehydratase and its activators (dhaB123, gdrAB) and the aldehyde dehydrogenase (puuC) from K. pneumoniae under control of pHyperspank promoter (strain h-syn-KpDhaB-PuuC-ΔglpK-i) cultivated in the M9 medium with glucose, induced by IPTG, and supplemented with B12 and (A) 12 g/L glycerol, (B) 25 g/L glycerol. Glycerol consumption is shown in blue, lactate production in green and 3-HP production in red. Average values from three independent biological replicates are shown, with the standard deviation.
3-HP Production Increased to 10 g/L in the Semi-defined and Rich Media
The maximal OD600 reached by the strain h-syn-KpDhaB-PuuC-ΔglpK-i in the M9 medium was 3.8. To reach higher cell density, we further cultured the production strain in M9 supplemented with 1 g/L yeast extract. The maximal cell OD600 reached 4.7 (Figure 6A) after 20.5 h, which is about 24% higher than in the M9 medium. This strain produced 1.23 g/L of 3-HP and consumed 2.4 g/L glycerol at the end of the cultivation (after 44 h), with a yield of 0.51 g 3-HP/g glycerol. The less than 100% yield is common in other bacterial cell factories, and is usually due to competing pathways, such as 1,3-PDO production (Dishisha et al., 2015), or conversion of glycerol to lactate via the methylglyoxal pathway (Landmann et al., 2011). We also observed that glucose and glycerol were consumed simultaneously from 5 to 20.5 h, which suggested that the expression of glpF, the gene encoding glycerol transporter, was most probably not negatively regulated by the presence of glucose. Since there is only one copy of dhaB123, gdrAB, and puuC genes in h-syn-KpDhaB-PuuC-ΔglpK-i strain, we inferred that the expression of these genes brings no obvious metabolic burden. Therefore, we induced the expression of synthetic pathway at the beginning of the cultivation. As shown in the Figure 6B, the early induction had no obvious negative effect on cell growth. A titer of 1.56 and 1.76 g/L 3-HP was accumulated at 20.5 and 44 h, respectively, and the 3-HP yield on glycerol reached 0.83 g/g. The ability of h-syn-KpDhaB-PuuC-ΔglpK-i to produce 3-HP was also evaluated in a rich medium. As shown in Figure 6C, a final 3-HP titer of 7.6 g/L was achieved and 21.4 g/L glycerol was consumed after a 64 h cultivation. However, the average 3-HP yield was only 0.35 g/g glycerol, much lower than the one obtained in the semi-defined medium. The glycerol consumption rate significantly increased from 40 to 64 h, accompanied by an increase in growth rate and biomass accumulation. In the same period, the 3-HP yield decreased from 0.78 g/g (0–40 h) to 0.17 g/g (40–64 h) glycerol. We suspected that this growth transition and the decrease in yield was caused by the instability of the ΔglpK mutation. Under our cultivation conditions, it is possible for the pMUTIN2-2 to exit the chromosome, resulting in the restitution of the functional glpK. Before the glucose in the medium is exhausted, up to 40 h, glucose is converted into biomass and glycerol is efficiently converted to 3-HP (80% yield; Figure 6D). Once the glucose is exhausted, selection pressure restitutes the WT glpK, and glycerol is then mainly converted to biomass, with the 3-HP yield dropping to 40% (Figure 6D). To test this assumption, we plated the cultures grown in the production setup after 64 h, and counted the colony forming units (CFU) on M9 supplemented with either glucose or glycerol (10 g/L). The CFU counts of the strain h-syn-KpDhaB-PuuC-ΔglpK-i on M9-glycerol plate was 71 ± 11% of that on M9-glucose plates, indicating that a significant sub-population of cells have restituted WT glpK. This explains the reduced yield of 3-HP on glycerol after 40 h of cultivation, since the loss of the ΔglpK mutation diverts the flux from glycerol to biomass, and reduces the flux to 3-HP (Figure 6D). The loss of the glpK knockout during cultivation could also be detected by PCR, checked at four different time points (Supplementary Figure S7A). To counter this effect, we constructed an irreversible ΔglpK strain using the method described by Arnaud et al. (2004). Using this method, the pMAD plasmid is first integrated at the glpK locus in the B. subtilis genome, and then the deletion is achieved when the plasmid leaves the chromosome in the second crossing over event. The resulting strain was named h-syn-KpDhaB-PuuC-ΔglpK-ii. Stability of the glpK knockout in this strain was confirmed by PCR at four different time points throughout the cultivation (Supplementary Figure S7B). Moreover, full stability of the integrated heterologous pathway was also confirmed by PCR at the same time points (Supplementary Figure S7C). We also checked the expression of the heterologously expressed proteins in crude extracts by using mass spectrometry proteomics, and have consistently detected both the DhaB and PuuC at all time points throughout the cultivation (Supplementary Figure S2). Compared to h-syn-KpDhaB-PuuC-ΔglpK-i, the h-syn-KpDhaB-PuuC-ΔglpK-ii was superior in terms of 3-HP titer (10 g/L vs 7.6 g/L) (Figures 7A,B) and average yield on glycerol (0.79 g/g vs 0.35 g/g), both of which remained stable throughout the experiment. This confirmed the hypothesis of a positive selective pressure on restoring the glpK, and highlighted the importance of using a stable knockout.
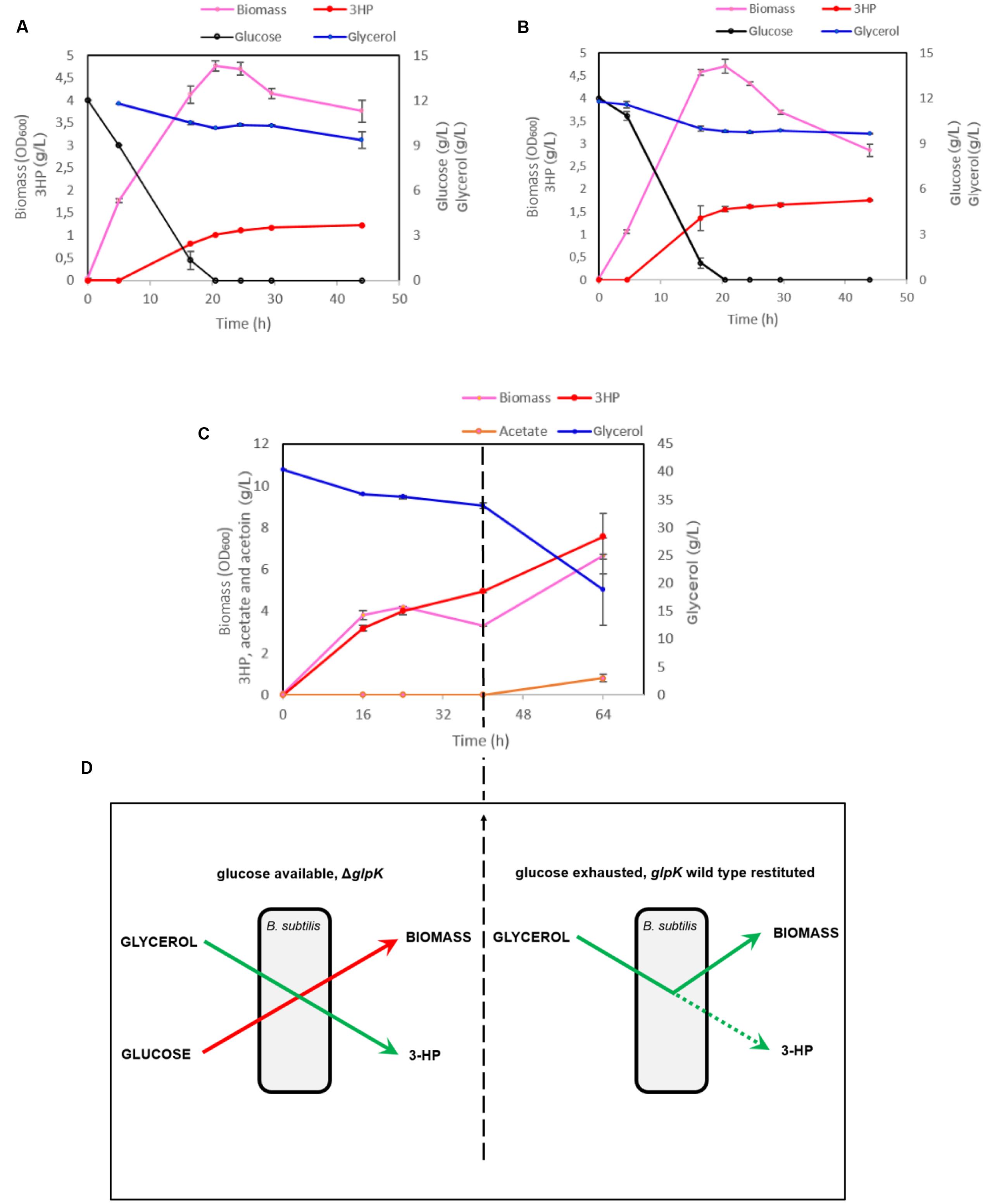
FIGURE 6. Cultivation of the strain h-syn-KpDhaB-PuuC-ΔglpK-i in the semi-defined and rich medium leads to increased 3-HP production. IPTG, glycerol and coenzyme B12 were added to the medium at 5 h (A) and at the beginning of cultivation (B,C). Glycerol and glucose consumption are shown in blue and black, respectively, 3-HP and acetate production are represented in red and orange, respectively, and biomass is shown in pink. Average values from three independent biological replicates are shown, with the standard deviation. (D) Schematic representation for explaining the loss of ΔglpK mutation after prolonged incubation of the production strain in the medium with glucose and glycerol. The restituted wild type (WT) glpK diverts the flux from glycerol to biomass, and reduces the flux to 3-HP, thus decreasing the 3-HP yield on glycerol. In the early stage of the experiment (left), glucose is used for biomass production, and a high yield of 3-HP on glycerol is achieved. At the late stage of the experiment (right), glucose is exhausted, glpK WT is restituted, and 3-HP yield on glycerol is diminished.
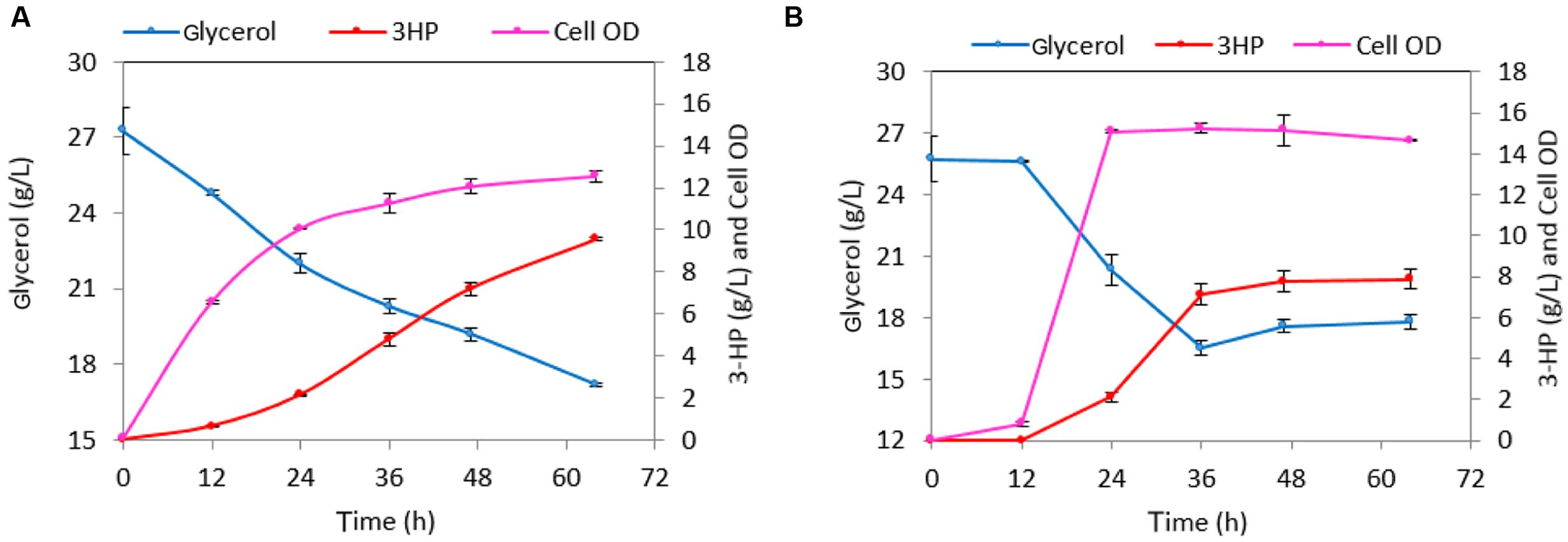
FIGURE 7. Cultivation of the strain h-syn-KpDhaB-PuuC-ΔglpK-ii in 2 × M9Y medium and rich medium for 3-HP production. (A) h-syn-KpDhaB-PuuC-ΔglpK-ii in 2 × M9Y medium, (B) h-syn-KpDhaB-PuuC-ΔglpK-ii in rich medium h-syn-KpDhaB-PuuC-ΔglpK-i in 2 × M9Y medium. Glycerol consumption is shown in blue, 3-HP production is represented in red, and biomass is shown in pink. Average values from two independent biological replicates are shown, with the standard deviation.
Conclusion and Perspectives
Bacillus subtilis has the potential to be considered as a microbial host for production of 3-HP due to several reasons including its efficient glycerol import system (da Silva et al., 2009) and high growth rate on glycerol (μmax = 0.65 h-1; Kruyssen et al., 1980) which compared to E. coli (μmax = 0.26 h-1; Chaudhary et al., 2012) is an advantage. It can reach high optical density (OD) in fermentation (Park et al., 1992) and has the ability to grow on simple media (Demain, 1958). B. subtilis can efficiently grow at high temperatures which reduces cooling costs during fermentation (Fu et al., 2016). In addition, there are number of developing novel and useful tools for genetic modification of B. subtilis (Dong and Zhang, 2014; Westbrook et al., 2016). Therefore, here we demonstrated that B. subtilis is a viable host for conversion of glycerol to 3-HP, with the production strain h-syn-KpDhaB-PuuC-ΔglpK-ii reaching a titer of 10 g/L in shake flasks. This is already comparable to the best benchmark strains of E. coli in shake flasks (Rathnasingh et al., 2009; Chu et al., 2015). Further optimization steps should include increasing the uptake rate for glycerol, by upregulating the expression of glpF, and export of 3-HP, which presumably takes place via the lactate transporter (Maris et al., 2004). More active dehydrogenases such as mutated ALDH (GabD4; Chu et al., 2015) could be used for further production improvement. Transferring the B12 production gene cluster to B. subtilis from its close relative Bacillus megaterium would significantly reduce the cost of production and should be considered. Adaptive laboratory evolution aimed at achieving higher glycerol uptake and utilization of chemostat conditions to neutralize the acidification from produced 3-HP, or on-line extractors to continuously remove 3-HP, should be reasonably expected to result in a robust and exploitable production strain.
Author Contributions
AK, TC, VR, and CS-B performed the experiments. BJ, IS, and AG performed the computational analyses. AK, TC, BJ, AG, DF, and IM analyzed the data. AK and IM wrote the manuscript.
Funding
We are thankful to the Region Champagne-Ardenne, the Conseil Général de la Marne, and Reims Métropole for their financial support.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
We are grateful to Carina Sihlbom at the Proteomics Core Facility, Sahlgrenska Academy, University of Gothenburg, Gothenburg, Sweden for assisting us with the proteome analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00638/full#supplementary-material
Footnotes
References
Arnaud, M., Chastanet, A., and Débarbouillé, M. (2004). New vector for efficient allelic replacement in naturally nontransformable, low-GC-content, gram-positive bacteria. Appl. Environ. Microbiol. 70, 6887–6891. doi: 10.1128/AEM.70.11.6887-6891.2004
Ashok, S., Mohan, S., Ko, Y., and Sankaranarayanan, M. (2013). Effect of puuC overexpression and nitrate addition on glycerol metabolism and anaerobic 3-hydroxypropionic acid production in recombinant Klebsiella pneumoniae ΔglpKΔdhaT. Metab. Eng. 15, 10–24. doi: 10.1016/j.ymben.2012.09.004
Atsumi, S., Hanai, T., and Liao, J. C. (2008). Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 451, 86–89. doi: 10.1038/nature06450
Atsumi, S., and Liao, J. C. (2008). Directed evolution of Methanococcus jannaschii citramalate synthase for biosynthesis of 1-propanol and 1-butanol by Escherichia coli. Appl. Environ. Microbiol. 74, 7802–7808. doi: 10.1128/AEM.02046-08
Borodina, I., Kildegaard, K. R., Jensen, N. B., Blicher, T. H., Maury, J., Sherstyk, S., et al. (2015). Establishing a synthetic pathway for high-level production of 3-hydroxypropionic acid in Saccharomyces cerevisiae via β-alanine. Metab. Eng. 27, 57–64. doi: 10.1016/j.ymben.2014.10.003
Burgé, G., Flourat, A. L., Pollet, B., Spinnler, H. E., and Allais, F. (2015a). 3-Hydroxypropionaldehyde (3-HPA) quantification by HPLC using a synthetic acrolein-free 3-hydroxypropionaldehyde system as analytical standard. RSC Adv. 5, 92619–92627. doi: 10.1039/C5RA18274C
Burgé, G., Saulou-Bérion, C., Moussa, M., Allais, F., Athes, V., and Spinnler, H. (2015b). Relationships between the use of Embden Meyerhof pathway (EMP) or Phosphoketolase pathway (PKP) and lactate production capabilities of diverse Lactobacillus reuteri strains. J. Microbiol. 53, 702–710. doi: 10.1007/s12275-015-5056-x
Chandrangsu, P., Dusi, R., Hamilton, C. J., and Helmann, J. D. (2014). Methylglyoxal resistance in Bacillus subtilis: contributions of bacillithiol-dependent and independent pathways. Mol. Microbiol. 91, 706–715. doi: 10.1111/mmi.12489
Chaudhary, N., Ngadi, M. O., and Simpson, B. (2012). Comparison of glucose, glycerol and crude glycerol fermentation by Escherichia Coli K12. J. Bioprocess. Biotech. S1, 001. doi: 10.4172/2155-9821.S1-001
Chen, Y., Bao, J., Kim, I., Siewers, V., and Nielsen, J. (2014). Coupled incremental precursor and co-factor supply improves 3-hydroxypropionic acid production in Saccharomyces cerevisiae. Metab. Eng. 22, 104–109. doi: 10.1016/j.ymben.2014.01.005
Chen, Y., and Nielsen, J. (2016). Biobased organic acids production by metabolically engineered microorganisms. Curr. Opin. Biotechnol. 37, 165–172. doi: 10.1016/j.copbio.2015.11.004
Choi, S., Song, C. W., Shin, J. H., and Lee, S. Y. (2015). Biorefineries for the production of top building block chemicals and their derivatives. Metab. Eng. 28, 223–239. doi: 10.1016/j.ymben.2014.12.007
Chu, H. S., Kim, Y. S., Lee, C. M., Lee, J. H., Jung, W. S., Ahn, J. H., et al. (2015). Metabolic engineering of 3-hydroxypropionic acid biosynthesis in Escherichia coli. Biotechnol. Bioeng. 112, 356–364. doi: 10.1002/bit.25444
Chun, A. Y., Yunxiao, L., Ashok, S., Seol, E., and Park, S. (2014). Elucidation of toxicity of organic acids inhibiting growth of Escherichia coli W. Biotechnol. Bioprocess Eng. 865, 858–865. doi: 10.1007/s12257-014-0420-y
Cox, J., Matic, I., Hilger, M., Nagaraj, N., Selbach, M., Olsen, J. V., et al. (2009). A practical guide to the MaxQuant computational platform for SILAC-based quantitative proteomics. Nat. Protoc. 4, 698–705. doi: 10.1038/nprot.2009.36
da Silva, G. P., Mack, M., and Contiero, J. (2009). Glycerol: a promising and abundant carbon source for industrial microbiology. Biotechnol. Adv. 27, 30–39. doi: 10.1016/j.biotechadv.2008.07.006
Demain, A. L. (1958). Minimal media for quantitative studies with Bacillus subtilis. J. Bacteriol. 75, 517–522.
Dishisha, T., Pyo, S. H., and Kaul, R. H. (2015). Bio-based 3-hydroxypropionic-and acrylic acid production from biodiesel glycerol via integrated microbial and chemical catalysis. Microb. Cell Fact. 14, 200. doi: 10.1186/s12934-015-0388-0
Dong, H., and Zhang, D. (2014). Current development in genetic engineering strategies of Bacillus species. Microb. Cell Fact. 13:63. doi: 10.1186/1475-2859-13-63
Fabret, C., Ehrlich, S. D., and Noirot, P. (2002). A new mutation delivery system for genome-scale approaches in Bacillus subtilis. Mol. Microbiol. 46, 25–36. doi: 10.1046/j.1365-2958.2002.03140.x
Fu, J., Huo, G., Feng, L., Mao, Y., Wang, Z., Ma, H., et al. (2016). Biotechnology for biofuels metabolic engineering of Bacillus subtilis for chiral pure meso-2,3-butanediol production. Biotechnol. Biofuels 9, 90. doi: 10.1186/s13068-016-0502-5
Hanai, T., Atsumi, S., and Liao, J. C. (2007). Engineered synthetic pathway for isopropanol production in Escherichia coli. Appl. Environ. Microbiol. 73, 7814–7818. doi: 10.1128/AEM.01140-07
Hao, J., Lin, R., Zheng, Z., Sun, Y., and Liu, D. (2008). 3-Hydroxypropionaldehyde guided glycerol feeding strategy in aerobic 1,3-propanediol production by Klebsiella pneumoniae. J. Ind. Microbiol. Biotechnol. 35, 1615–1624. doi: 10.1007/s10295-008-0405-y
Harwood, C. R., and Cutting, S. M. (eds). (1990). “Chemically defined growth media and supplements,” in Molecular Biological Methods for Bacillus (Chichester: Wiley), 548.
Holmberg, C., Beijer, L., Rutberg, B., and Rutberg, L. (1990). Glycerol catabolism in Bacillus subtilis: nucleotide sequence of the genes encoding glycerol kinase (glpK) and glycerol-3-phosphate dehydrogenase (glpD). J. Gen. Microbiol. 136, 2367–2375. doi: 10.1099/00221287-136-12-2367
Huang, Y., Li, Z., Shimizu, K., and Ye, Q. (2012). Simultaneous production of 3-hydroxypropionic acid and 1,3-propanediol from glycerol by a recombinant strain of Klebsiella pneumoniae. Bioresour. Technol. 103, 351–359. doi: 10.1016/j.biortech.2011.10.022
Huang, Y., Li, Z., Shimizu, K., and Ye, Q. (2013). Co-production of 3-hydroxypropionic acid and 1,3-propanediol by Klebseilla pneumoniae expressing aldH under microaerobic conditions. Bioresour. Technol. 128, 505–512. doi: 10.1016/j.biortech.2012.10.143
Jiang, X., Meng, X., and Xian, M. (2009). Biosynthetic pathways for 3-hydroxypropionic acid production. Appl. Microbiol. Biotechnol. 82, 995–1003. doi: 10.1007/s00253-009-1898-7
Jung, I., Lee, J., Min, W., Park, Y., and Seo, J. (2015). Bioresource technology simultaneous conversion of glucose and xylose to 3-hydroxypropionic acid in engineered Escherichia coli by modulation of sugar transport and glycerol synthesis. Bioresour. Technol. 198, 709–716. doi: 10.1016/j.biortech.2015.09.079
Jung, W. S., Kang, J. H., Chu, H. S., Choi, I. S., and Cho, K. M. (2014). Elevated production of 3-hydroxypropionic acid by metabolic engineering of the glycerol metabolism in Escherichia coli. Metab. Eng. 23, 116–122. doi: 10.1016/j.ymben.2014.03.001
Kildegaard, K. R., Hallstrom, B. M., Blicher, T. H., Sonnenschein, N., Jensen, N. B., Sherstyk, S., et al. (2014). Evolution reveals a glutathione-dependent mechanism of 3-hydroxypropionic acid tolerance. Metab. Eng. 26, 57–66. doi: 10.1016/j.ymben.2014.09.004
Kildegaard, K. R., Wang, Z., Chen, Y., Nielsen, J., and Borodina, I. (2015). Production of 3-hydroxypropionic acid from glucose and xylose by metabolically engineered Saccharomyces cerevisiae. Metab. Eng. Commun. 2, 132–136. doi: 10.1016/j.meteno.2015.10.001
Kim, K., Kim, S. K., Park, Y. C., and Seo, J. H. (2014). Enhanced production of 3-hydroxypropionic acid from glycerol by modulation of glycerol metabolism in recombinant Escherichia coli. Bioresour. Technol. 156, 170–175. doi: 10.1016/j.biortech.2014.01.009
Kruyssen, F. J., de Boer, W. R., and Wouters, J. T. (1980). Effects of carbon source and growth rate on cell wall composition of Bacillus subtilis subsp. niger. J. Bacteriol. 144, 238–246.
Kumar, V., Ashok, S., and Park, S. (2013). Recent advances in biological production of 3-hydroxypropionic acid. Biotechnol. Adv. 31, 945–961. doi: 10.1016/j.biotechadv.2013.02.008
Kumar, V., Sankaranarayanan, M., Ashok, S., Ko, Y., Sarkar, R., and Park, S. (2012). Co-production of 3-hydroxypropionic acid and 1, 3-propanediol from glycerol using resting cells of recombinant Klebsiella pneumoniae J2B strain overexpressing aldehyde dehydrogenase. Appl. Microbiol. Biotechnol. 96, 373–383. doi: 10.1007/s00253-012-4187-9
Landmann, J. J., Busse, R. A., Latz, J., Singh, K. D., Stülke, J., and Görke, B. (2011). Crh, the paralogue of the phosphocarrier protein HPr, controls the methylglyoxal bypass of glycolysis in Bacillus subtilis. Mol. Microbiol. 82, 770–787. doi: 10.1111/j.1365-2958.2011.07857.x
Lindner, S. N., Meiswinkel, T. M., Panhorst, M., Youn, J. W., Wiefel, L., and Wendisch, V. F. (2012). Glycerol-3-phosphatase of Corynebacterium glutamicum. J. Biotechnol. 159, 216–224. doi: 10.1016/j.jbiotec.2012.02.003
Maris, A. J. A., Konings, W. N., van Dijken, J. P., and Pronk, J. T. (2004). Microbial export of lactic and 3-hydroxypropanoic acid: implications for industrial fermentation processes. Metab. Eng. 6, 245–255. doi: 10.1016/j.ymben.2004.05.001
McKenna, R., and Nielsen, D. R. (2011). Styrene biosynthesis from glucose by engineered E. coli. Metab. Eng. 13, 544–554. doi: 10.1016/j.ymben.2011.06.005
Niu, K., Xiong, T., Qin, H. B., Wu, H., Liu, Z. Q., and Zheng, Y. G. (2016). 3-Hydroxypropionic acid production by recombinant Echerichia coli ZJU-3HP01using glycerol-glucose dual-substrate fermentative strategy. Biotechnol. Appl. Biochem.
Oh, Y. K., Palsson, B. O., Park, S. M., Schilling, C. H., and Mahadevan, R. (2007). Genome-scale reconstruction of metabolic network in Bacillus subtilis based on high-throughput phenotyping and gene essentiality data. J. Biol. Chem. 282, 28791–28799. doi: 10.1074/jbc.M703759200
Park, Y. S., Kai, K., and Kobayashi, T. (1992). Enhanced beta-galactosidase production by high cell-density culture of recombinant Bacillus subtilis with glucose concentration control. Biotechnol. Bioeng. 40, 686–696. doi: 10.1002/bit.260400607
Raj, S. M., Rathnasingh, C., Jo, J., and Park, S. (2008). Production of 3-hydroxypropionic acid from glycerol by a novel recombinant Escherichia coli BL21 strain. Process Biochem. 43, 1440–1446. doi: 10.1016/j.procbio.2008.04.027
Rathnasingh, C., Raj, S. M., Jo, J. E., and Park, S. (2009). Development and evaluation of efficient recombinant Escherichia coli strains for the production of 3-hydroxypropionic acid from glycerol. Biotechnol. Bioeng. 104, 729–739. doi: 10.1002/bit.22429
Sankaranarayanan, M., Ashok, S., and Park, S. (2014). Production of 3-hydroxypropionic acid from glycerol by acid tolerant Escherichia coli. J. Ind. Microbiol. Biotechnol. 41, 1039–1050. doi: 10.1007/s10295-014-1451-2
Schellenberger, J., Que, R., Fleming, R. M. T., Thiele, I., Orth, J. D., Feist, A. M., et al. (2011). Quantitative prediction of cellular metabolism with constraint-based models: the COBRA Toolbox v2.0. Nat. Protoc. 6, 1290–1307. doi: 10.1038/nprot.2011.308
Shevchenko, A., Thomas, H., Havlis, J., Olsen, J. V., and Mann, M. (2007). In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 1, 2856–2860. doi: 10.1038/nprot.2006.468
Tsuruno, K., Honjo, H., and Hanai, T. (2015). Enhancement of 3-hydroxypropionic acid production from glycerol by using a metabolic toggle switch. Microb. Cell Fact. 14:155. doi: 10.1186/s12934-015-0342-1
Vagner, V., Dervyn, E., and Ehrlich, D. (1998). A vector for systematic gene inactivation in Bacillus subtilis. Microbiology 144, 3097–3104.
Valdehuesa, K. N. G., Liu, H., Nisola, G. M., Chung, W. J., Lee, S. H., and Park, S. J. (2013). Recent advances in the metabolic engineering of microorganisms for the production of 3-hydroxypropionic acid as C3 platform chemical. Appl. Microbiol. Biotechnol. 97, 3309–3321. doi: 10.1007/s00253-013-4802-4
Westbrook, A. W., Moo-young, M., and Chou, C. P. (2016). Development of a CRISPR-Cas9 tool kit for comprehensive engineering of Bacillus subtilis. Appl. Environ. Microbiol. 82, 4876–4895. doi: 10.1128/AEM.01159-16.Editor
Keywords: 3-hydroxypropanoic acid, glycerol, Bacillus subtilis, metabolic engineering, glycerol kinase knock-out
Citation: Kalantari A, Chen T, Ji B, Stancik IA, Ravikumar V, Franjevic D, Saulou-Bérion C, Goelzer A and Mijakovic I (2017) Conversion of Glycerol to 3-Hydroxypropanoic Acid by Genetically Engineered Bacillus subtilis. Front. Microbiol. 8:638. doi: 10.3389/fmicb.2017.00638
Received: 17 January 2017; Accepted: 28 March 2017;
Published: 18 April 2017.
Edited by:
Michael Sauer, University of Natural Resources and Life Sciences, AustriaReviewed by:
Sunghoon Park, Pusan National University, South KoreaDong-Woo Lee, Kyungpook National University, South Korea
Binod Parameswaran, National Institute for Interdisciplinary Science and Technology (CSIR), India
Copyright © 2017 Kalantari, Chen, Ji, Stancik, Ravikumar, Franjevic, Saulou-Bérion, Goelzer and Mijakovic. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ivan Mijakovic, aXZhbi5taWpha292aWNAY2hhbG1lcnMuc2U=