 Peili Cao
Peili Cao Dongchun Guo
Dongchun Guo Jiasen Liu1
Jiasen Liu1 Zhuofei Xu
Zhuofei Xu Liandong Qu
Liandong Qu- 1State Key Laboratory of Veterinary Biotechnology, Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences, Harbin, China
- 2State Key Laboratory of Agricultural Microbiology, College of Veterinary Medicine, Huazhong Agricultural University, Wuhan, China
Pasteurella multocida, a Gram-negative opportunistic pathogen, has led to a broad range of diseases in mammals and birds, including fowl cholera in poultry, pneumonia and atrophic rhinitis in swine and rabbit, hemorrhagic septicemia in cattle, and bite infections in humans. In order to better interpret the genetic diversity and adaptation evolution of this pathogen, seven genomes of P. multocida strains isolated from fowls, rabbit and pigs were determined by using high-throughput sequencing approach. Together with publicly available P. multocida genomes, evolutionary features were systematically analyzed in this study. Clustering of 70,565 protein-coding genes showed that the pangenome of 33 P. multocida strains was composed of 1,602 core genes, 1,364 dispensable genes, and 1,070 strain-specific genes. Of these, we identified a full spectrum of genes related to virulence factors and revealed genetic diversity of these potential virulence markers across P. multocida strains, e.g., bcbAB, fcbC, lipA, bexDCA, ctrCD, lgtA, lgtC, lic2A involved in biogenesis of surface polysaccharides, hsf encoding autotransporter adhesin, and fhaB encoding filamentous haemagglutinin. Furthermore, based on genome-wide positive selection scanning, a total of 35 genes were subject to strong selection pressure. Extensive analyses of protein subcellular location indicated that membrane-associated genes were highly abundant among all positively selected genes. The detected amino acid sites undergoing adaptive selection were preferably located in extracellular space, perhaps associated with bacterial evasion of host immune responses. Our findings shed more light on conservation and distribution of virulence-associated genes across P. multocida strains. Meanwhile, this study provides a genetic context for future researches on the mechanism of adaptive evolution in P. multocida.
Introduction
Pasteurella multocida, a Gram-negative rod-shaped bacterium, is the type species of the genus Pasteurella (Christensen and Bisgaard, 2006). Pasteurella multocida is an opportunistic pathogen that can cause multihost diseases, characterized by fowl cholera in poultry, pneumonia and atrophic rhinitis in swine, hemorrhagic septicemia in cattle and rabbit, and bite infections in humans (Boyce et al., 2010). These diseases have led to huge economic losses to the livestock and poultry industry worldwide (Ghaffar and Tariq, 2016). Based on differences in capsular antigens, P. multocida is classified into five capsular serogroups A, B, D, E, and F (Carter, 1955). To date, a number of P. multocida virulence factors have been extensively studied, including capsule, lipopolysaccharides (LPSs), P. multocida toxin, surface adhesins and iron acquisition proteins (Harper et al., 2006).
To explore genetic content of virulence genes in P. multocida, the first complete genome sequence of the avian strain Pm70 has been reported in 2001 (May et al., 2001). The research on comparative genomics of P. multocida is subsequently fueled by rapid development of next-generation sequencing technology during the last decade. More epidemic strains of P. multocida have been whole-genome sequenced to investigate the associations of pathogenesis with the underlying genetic diversity among P. multocida genomes (Michael et al., 2012; Abrahante et al., 2014). Several studies on comparative genome analyses have been reported for this pathogen. Analyses of nine P. multocida genomes have indicated that there is no clear correlation between phylogenetic relatedness and host predilection or disease (Boyce et al., 2012). In addition, genome-wide comparison of three avian P. multocida strains has identified 336 unique genes present in both virulent strains P1059 and X73 but absent in the avirulent strain Pm70, some genes of which may be associated with enhanced virulence or fitness (Johnson et al., 2013). Using genome sequences of 11 haemorrhagic septicaemia (HS) strains and four other P. multocida strains, a recent study by Moustafa et al. (2015) has revealed 96 HS-specific genes which could promote the development of disease-specific diagnostic tests.
Except for large genome rearrangement and gene gain or lost, genetic variations referring to single nucleotide polymorphisms (SNPs) and insertions/deletions on the conserved genetic elements also play crucial roles on bacterial virulence, population diversification and adaptation to host niches (Toft and Andersson, 2010). Analyses of SNPs in three P. multocida genomes has indicated a higher dN/dS ratio is prone to occur on the genes encoding outer membrane proteins interacted with the host immune system (Johnson et al., 2013). The massive accumulation of genomic sequence data enables researchers to apply more robust dN/dS-based methods to trace evolutionary trajectories on the protein-coding genes under positive Darwinian selection exerted by the host immune system. Genome-scale positive selection scanning has been widely used to detect bacterial genes important for host adaptation in many pathogenic taxa, such as Escherichia coli (Chen et al., 2006; Petersen et al., 2007), Campylobacter (Lefébure and Stanhope, 2009), Streptococcus (Lefébure and Stanhope, 2007), Mycobacterium tuberculosis (Hongo et al., 2015), and Actinobacillus pleuropneumoniae (Xu et al., 2011a). To date, systematical analysis of positive selection on P. multocida protein-coding genome is still lacking.
In the present study, we sequenced the genomes of seven P. multocida strains isolated in China. Together with 26 public P. multocida genome assemblies, a comparative genome analysis was performed to characterize pangenome structure of this pathogenic bacterium. Genetic content and conservation of virulence genes were comprehensively analyzed. Furthermore, a genome-wide evolutionary analysis was carried out to investigate the effects of positive Darwinian selection acting on protein-coding genome. A number of genes and amino acid sites undergoing intensive positive selection were uncovered, which may be associated with the fitness and immunogenic properties of P. multocida. These findings provide a genetic context for future researches on diversity of virulence and adaptation evolution in P. multocida.
Methods
Bacterial Strains
Pasteurella multocida strains were cultured on brain heart infusion agar (BD Biosciences, San Jose, CA, USA) containing 5% sterile sheep blood and incubated at 37°C for 24~48 h. The P. multocida strains were identified via kmt1 gene and capsular multiplex PCR analysis using primers previously described by Townsend et al. (2001). The ST-type of these strains has been investigated by multi-locus sequence typing based on seven housekeeping genes (adk, est, pmi, zwf, mdh, gdh, and pgi) in our recent study (Wang et al., 2016). More information about seven P. multocida strains sequenced in this study is shown in Table 1. Total genomic DNA was extracted by using the DNeasy tissue kit (Qiagen).
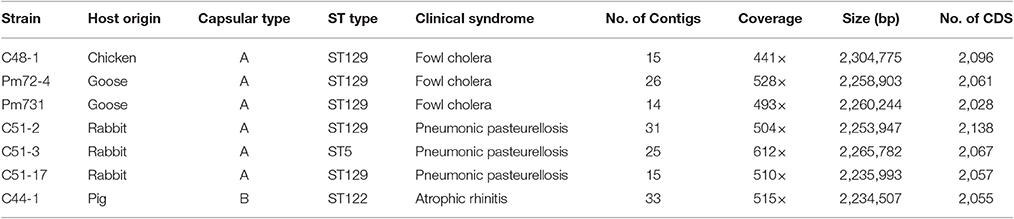
Table 1. Summary of genome assembly and gene calling of sequenced P. multocida strains.
Genome Sequencing, Annotation, and Alignment
Whole-genome shotgun sequencing was performed by using Illumina MiSeq platform. Illumina sequencing was done by Majorbio Bio-pharm Technology Co., Ltd, China. Genomic DNA library containing 400~500 bp fragments was constructed by TruSeq™ DNA Sample Prep Kit (Illumina). Sequencing experiments were conducted on an Illumina MiSeq sequencer, giving 250-bp paired-end reads and an average ~515-fold genome coverage per sample. Adaptor sequences and low quality ends per read were trimmed by using SeqPrep (https://github.com/jstjohn/SeqPrep) and Sickle (https://github.com/najoshi/sickle/). High quality reads were retained if satisfying the following criteria: minimum mean quality score of 20; minimum read length of 50 bp. Short reads were de novo assembled into contigs using the package SOAPdenovo v2.04 (Li et al., 2010), and contigs less than 500 bp were filtered. The genome assemblies were annotated by using an integrated computational pipeline Prokka v1.12 (Seemann, 2014). Briefly, protein-coding genes, tRNAs and rRNAs were predicted by Prodigal v2.6.2 (Hyatt et al., 2010), Aragorn v1.2.36 (Laslett and Canback, 2004), and RNAmmer v1.2 (Lagesen et al., 2007), respectively. Prophage sequences within the genome assemblies were identified by the PHAST web server (Zhou et al., 2011). Based on sequence similarity searching the genome assemblies against the complete genome sequence of P. multocida type strain ATCC 43137 using blastn (E-value cutoff set to 10−10) in the package BLAST 2.2.30+ (Camacho et al., 2009), a circular map of pairwise genome alignment was generated and visualized by the tools BRIG v0.95 (Alikhan et al., 2011) and CGview (Grant et al., 2012). The similarity between bacterial genomes was estimated by calculating the average nucleotide identity (ANI) based on BLAST analysis using JSpecies with default parameters (Goris et al., 2007).
Draft genome assemblies have been deposited in the GenBank database under the accession numbers MANI00000000, MAPP00000000, MAPQ00000000, MAPR00000000, MAPS00000000, MAPT00000000, and MBAG00000000.
Phylogenetic Analysis
To infer the phylogeny of P. multocida strains, Parsnp v1.2 was employed to perform whole-genome alignment and variant calling using the following parameters: -x -z 25 -c -C 1000 (Treangen et al., 2014). The genome of P. multocida type strain ATCC 43137 was used as reference. The core-genome multiple alignment was composed of locally collinear blocks (>25 bp) detected by Parsnp. The aligned columns with recombination signals were further detected by PhiPack (Bruen and Philippe, 2006) and filtered. A phylogenetic tree was reconstructed based on the final alignment of core-genome SNPs using FastTree2 (Price et al., 2010). The resulting tree of the whole-genome phylogeny was then visualized and annotated with the strain properties by using iTOL v3 (Letunic and Bork, 2011).
Pangenome Analysis
A pangenome analysis was carried out to compare genetic content between P. multocida genomes. To keep consistency of analysis procedures, we re-annotated the other collected genome assemblies using Prokka. Proteinortho v5.13 (Lechner et al., 2011) was then adopted to cluster orthologous genes according the criteria: more than 80% identity and 80% coverage of the best blast alignments. The clusters of orthologous genes (OGs) present in all strains were determined as the core genes of P. multocida. Distribution of OGs among strains was visualized by using FriPan (https://github.com/drpowell/FriPan). Using blastp functional annotation and classification of OGs were performed based on the UniRef50 database (Suzek et al., 2007), the cluster of orthologous groups (COG) database (Tatusov et al., 2000), and the virulence factor database VFDB (Chen et al., 2016). Gene annotation was replenished by the top hit with E-value cutoff of 10−20. Protein functional domain was predicted using the Pfam-A database v29.0 (Punta et al., 2012). OGs annotated to be virulence genes were further screened using tblastn searching against P. multocida genomes by the approach of BLAST Score Ratio (BSR) (Sahl et al., 2014). A data matrix consisting of the BSR values for each gene across genomes was obtained. Gene distribution and conservation patterns were visualized by using iTOL (Letunic and Bork, 2011). Biological pathway analysis was performed using BlastKOALA and KEGG database (Kanehisa et al., 2016).
Detection of Recombination
To eliminate the potential effect of recombination on positive selection scanning, intragenic homologous recombination was detected for the set of single-copy core OGs. Multiple sequence alignment for each OG cluster was initially performed with amino acid sequences by using T-Coffee v11 (Notredame et al., 2000). The resulting protein sequence alignments were then converted to the codon alignments for the subsequent analysis. Unreliable alignment regions were further removed using Gblocks v0.91 with the codon mode and the default relaxed settings as defined by Talavera (Talavera and Castresana, 2007). Recombination test was conducted using the stand-alone program Genetic Algorithm Recombination Detection (GARD) implemented by HYPHY v2.2 (Kosakovsky-Pond et al., 2006). Additionally, informative sites within each alignment were detected by the PhiPack package. The sequence alignments of the recombining OGs were then partitioned according to the breakpoints inferred by GARD.
Positive Selection Scanning
To detect adaptive evolution in the protein-coding genome of P. multocida, the rates of synonymous and non-synonymous substitutions were estimated using site-model of the codeml program in the PAML v4.8 package (Yang, 2007). Briefly, a phylogenetic tree for each gene was built using the maximum-likelihood approach implemented by PhyML v3.0 (Guindon and Gascuel, 2003). A general time-reversible (GTR) model of nucleotide substitutions with the estimated gamma distributed rate heterogeneity of four categories (Γ4) and a proportion of invariable sites was used in the tree reconstruction. Based on the resulting tree topology, two site-specific models that allow variable ratios (ω) of non-synonymous (dN) to synonymous substitutions (dS) among codons were used in our data set: M1a (NearlyNeutral) and M2a (PositiveSelection). The latter model adds an extra site class for a fraction of positively selected sites with ω > 1, indicating positive (adaptive) selection; whereas models M1a only allows site classes with ω varying between 0 and 1 (Wong et al., 2004). A likelihood ratio test was carried out to infer the occurrence of sites subject to positive selection through comparing M1a against M2a. The likelihood statistic (2Δℓ) was calculated and compared with the critical value from χ2 distribution with two degrees of freedom. The Bayes empirical bayes (BEB) method was employed to identify positively selected sites under the likelihood framework (Yang et al., 2005).
Functional Analysis of Positively Selected Genes
PSORTb v3.02 web server (Yu et al., 2010) was employed to predict protein subcellular localization. Protein Transmembrane helices were predicted by TMHMM 2.0 (Krogh et al., 2001). Integral β-barrel outer membrane proteins were screened by using BOMP (Berven et al., 2004) and the topology of transmembrane β-barrel was predicted by BOCTOPUS2 (Hayat et al., 2016). Signal peptide cleavage site was predicted by using SignalP v4.1 server (Petersen et al., 2011). Three dimensional structures of positively selected proteins were modeled using Phyre2 server with the intensive model (Kelley and Sternberg, 2015). The amino acid residues subject to positively selective pressure were mapped onto the structure and visualized by Chimera v1.11 (Pettersen et al., 2004).
Statistical Analyses
Multiple testing corrections were performed to control the Type I errors. For the analyses of homologous recombination, recombination breakpoints were inferred by the Shimodaira-Hasegawa test (Shimodaira and Hasegawa, 1999) with Bonferroni-corrected p-value and the threshold for significance was set at 0.05. For all genes tested for positive selection, the false discovery rate (FDR) was controlled by using the BY method (Benjamini and Yekutieli, 2001) and the significance level was set to 10%. Chi-square test and Fisher-exact test where appropriate were used to assess associations for the gene count data in the individual COG categories; Bonferroni corrections for multiple tests were applied. All computational analyses were carried out using in-house Perl scripts and R 3.2.1 (R Core Team, 2015).
Results
General Features of Newly Sequenced Genomes
In the present study, seven P. multocida strains were chosen for whole genome shotgun sequencing, including three strains C51-2, C51-3, C51-17 isolated from rabbit, two strains Pm72-4 and Pm731 isolated from goose, strain C48-1 isolated from chicken, and strain C44-1 isolated from pig (Table 1). After genome assembly, the average number of contigs per genome was 23 with a range from 14 to 33. The depth of genome coverage was 441- to 612-fold (Table 1). The average GC content of each genome was 40.3%, which was consistent with that of a complete P. multocida chromosome (Liu et al., 2012). The median number of coding sequences (CDSs) per strain was 2,061, the largest number was 2,138 for strain C51-2, and the smallest was 2,028 for strain Pm731. Additionally, the newly sequenced strains possessed pairwise ANI values of 98.06~99.96% compared with eight representative P. multocida strains (Table S1). The high ANI values of pairwise genome comparisons again confirmed these strains belonging to P. multocida. All genome assemblies of seven strains have been deposited in the GenBank database.
For a glimpse of genome similarity of P. multocida strains, pairwise sequence alignments between genomes of P. multocida reference strain ATCC 43137 and other strains is shown in Figure 1. We found the majority of genomic regions between the reference strain ATCC 43137 and the other P. multocida strains were highly conserved. Among seven newly sequenced P. multocida strains in our study, the proportion of all matched sequences (>95% identity) accounting for the ATCC 43137 genome (2,271,840 bp) was ranged from ~89.5% (C44-1 vs. ATCC 43137) to ~92.7% (C48-1 vs. ATCC 43137). Genomic fragments (>200 bp) that are present in the ATCC 43137 genome but absent in the newly sequenced genomes were summarized and shown in Table S2. In accordance with the type-A capsule of P. multocida strain ATCC 43137 (Davenport et al., 2014), the biosynthetic loci of capsular polysaccharide (CPS) were highly conserved in the newly sequenced genomes of six strains possessing type-A capsule. Unsurprisingly, the locus of type-A capsule was absent in strain C44-1 and the other strains bearing non-type-A capsule. Except the CPS locus, we also observed genomic islands relative to the other known VFs, including LPS locus, TonB receptor protein, autotransporter surface adhesins Hsf, and tad (tight adherence) locus (Figure 1). These particular genomic regions have been depicted by a recent pangenome analysis based on nine P. multocida genomes (Boyce et al., 2012). Gene content and conservation of these featured genomic regions were analyzed in details below. In addition, an intact prophage (41.6 kb), which was localized in the vicinity of the replication terminus of the ATCC 43137 genome, was also present in the genomes of three serotype-A strains (HB03, 3480, and X73) but not all. The prophage island contains 58 CDSs, 40 of which encode phage-related proteins.
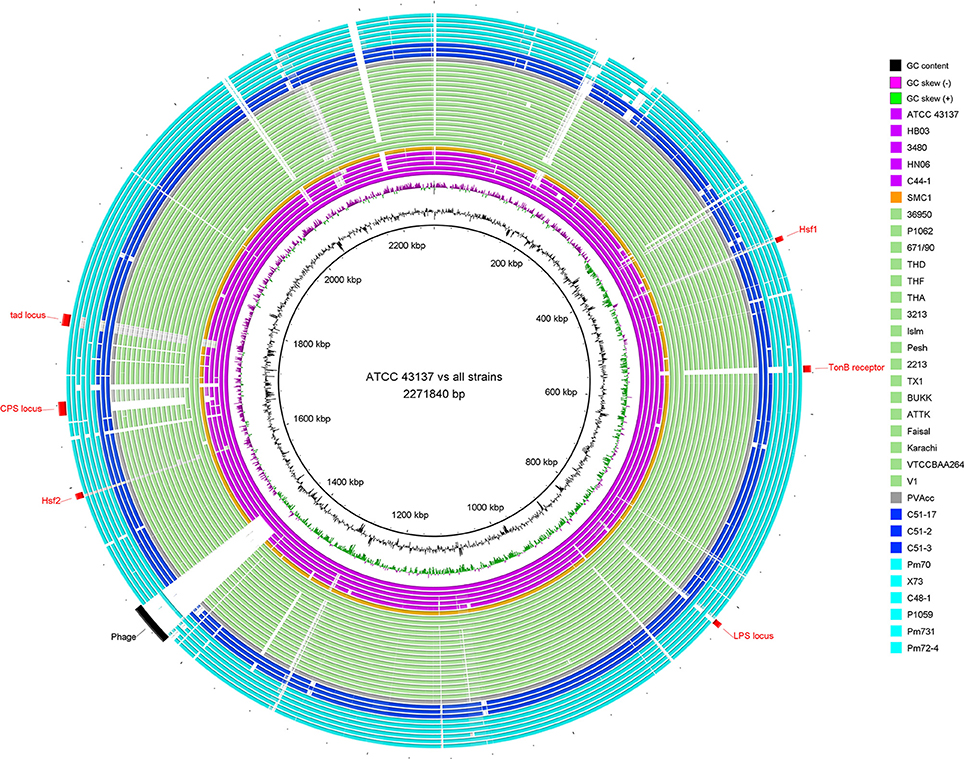
Figure 1. Circular diagram of genomic sequence conservation of 33 P. multocida strains. Rings are numbered from 1 (outermost ring) to 35 (innermost ring). The outermost 33 rings show pairwise nucleotide alignment between 32 genome assemblies of P. multocida and the reference genome of P. multocida type strain ATCC 43137. Each ring is color-coded according to host origins of isolates: magenta for swine source strains, light green for bovine source strains, blue for rabbit source strains, cyan for avian source strains, orange for human source strain, and gray for strain PVAcc without host metadata. BLAST matches (minimum sequence identity of 80% and E-value cutoff of 10−10) are colored from darkest to lightest shade. The innermost ring shows mean centered GC content (outward black bar: above mean; inward bar: below mean) of the ATCC 43137 genome. The second innermost ring shows GC skew plot [(G-C)/(G+C); green indicates ratio > 0 and purple indicates ratio < 0]. Red labels and arcs denote genomic regions encoding several known virulence factors of P. multocida; black label and arc denote a prophage region predicted by PHAST (Zhou et al., 2011).
Phylogeny of P. multocida
To understand the phylogenetic relationships among P. multocida isolates, 26 publicly available P. multocida genomes were collected and compared with seven sequenced genomes in this study (Table 2). As shown in Figure 2, a maximum-likelihood phylogeny of P. multocida was reconstructed based on SNPs detected in the core-genome of all strains. It was apparent that four major phylogenetic groups were found in the tree. All serotype-B strains, including fifteen bovine source strains and one swine source strain, were clustered into the same phylogenetic group IV. For strains in the phylogenetic group II, we observed that three poultry source stains (C48-1, Pm72-4, and Pm731) and two rabbit source strains (C51-17 and C51-2) were closely related, implicating these strains from both hosts probably derived from a recent common ancestor. Phylogenetic group I containing two swine source strains (ATCC 43137 and HB03) and two bovine source strains (36950 and P1062) was very distantly related to group II and III, in which serotype-A strains were also dominated. In addition, the pathogenic strain SMC1 isolated from an adult female in Malaysia and the virulent strain C51-3 isolated from rabbit in China were assigned to a common clade. Consistently, the ANI value between strains C51-3 and SMC1 was 99.31%, indicating both are closely related strains within P. multocida. It was noting that none of obvious associations between phylogenetic groups and phenotypic characteristics of P. multocida strains could be presented, perhaps due to the limited genome data collected in this study. This observation is in line with a previous study on comparative genomics analyses of nine P. multocida strains (Boyce et al., 2012).
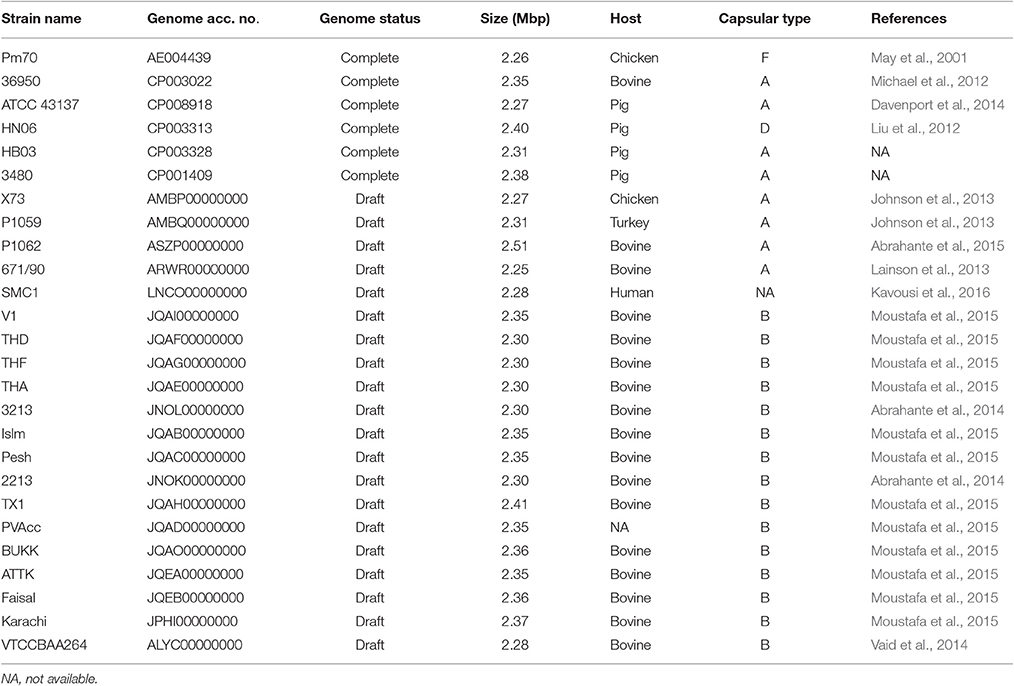
Table 2. Public P. multocida genome assemblies analyzed in this study.
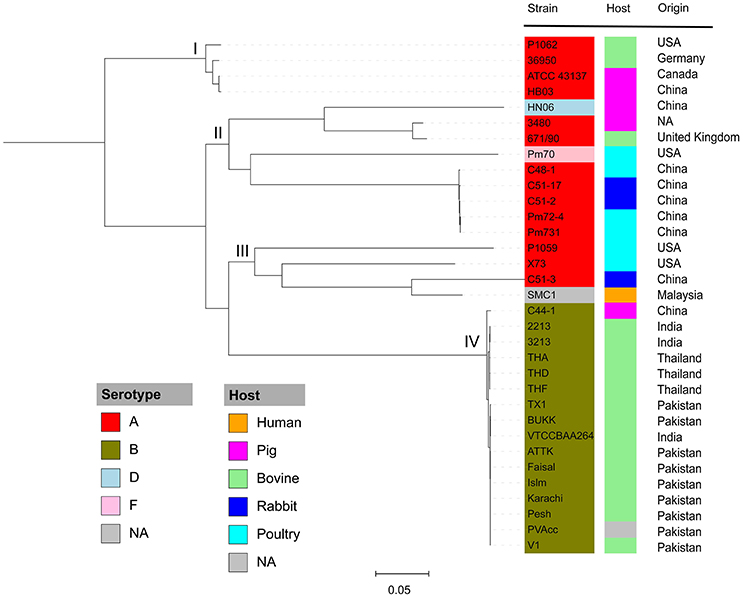
Figure 2. Maximum likelihood phylogeny of P. multocida. The phylogenetic tree was reconstructed based on SNPs in the core genomes of P. multocida strains using the Parsnp program (Treangen et al., 2014). Phylogenetic group was manually set and designated as groups I, II, III, and IV. All strains are color-coded according to their serotypes A, B, D, and F. The host origin of each strain is denoted by the right color strip with the same scheme as in Figure 1. The geographical location of isolation for each strain is shown. The gray box labeled as NA indicates the metadata is not available for the related strains.
Pangenome of P. multocida
To get a glimpse of distribution of protein clusters across P. multocida genomes, we performed pangenome analyses based on 70,565 CDSs from all 33 genomes tested in this study (Figure 3A). The total number of P. multocida orthologous gene (OG) clusters, also including unique genes specific to individual strain, was 4,036 (Table S3). Of these OGs, 39% were single-copy genes present in all tested strains and constituted the core genome of P. multocida, 33% were dispensable genes that were possessed by at least two strains but not all, 27% were unique genes per strain, and the remaining were multiple-copy genes (Figure S1). The dispensable genes together with the unique genes are likely to play crucial roles on strain-specific phenotypes or pathotypes. Genetic content and diversity on these accessory genes across strains were analyzed below.
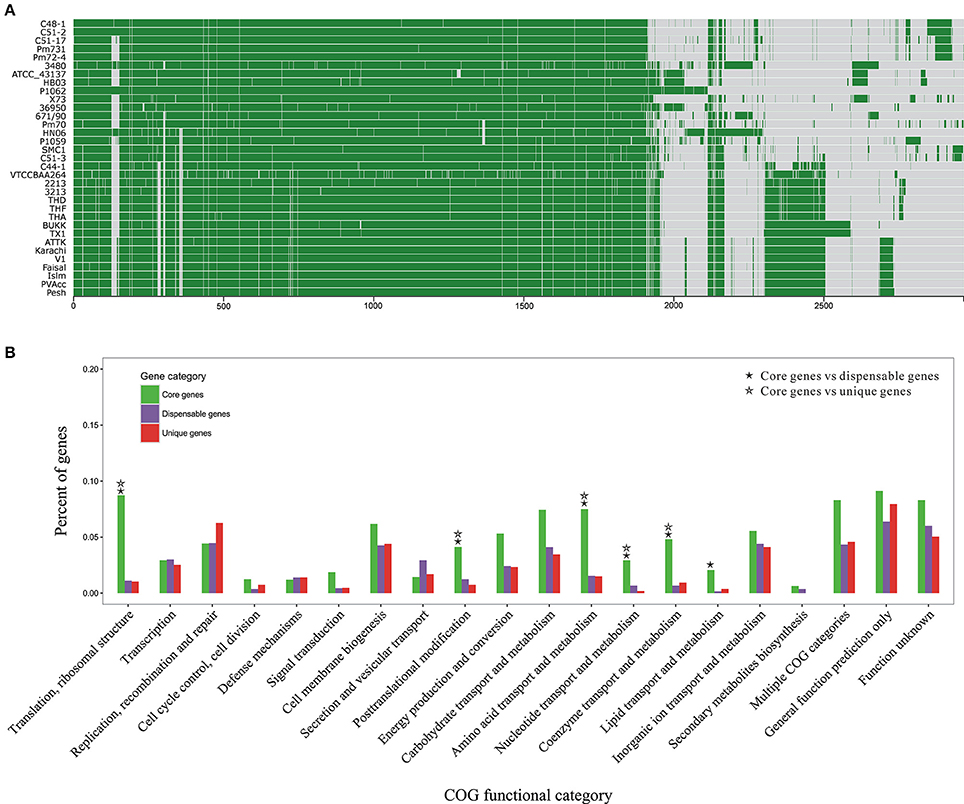
Figure 3. Pan-genome structure and function of P. multocida. (A) A pan-genome map containing 2,966 OG clusters in all P. multocida genomes detected by Proteinortho (Lechner et al., 2011). Each block strands for a gene: green for presence and gray for absence. (B) COG-based binning of core genes, dispensable genes, and strain-specific genes. The abscissa denotes different COG functional categories. The ordinate denotes the proportion of genes in each COG category and details are summarized in Table S3. Two COG functional categories (“RNA processing and modification” and “Cell motility”) including only one OG are not displayed, as well as the genes encoding proteins without homologs in the COG collection. Significant enrichment of gene occurrence in the individual category is marked by asterisks (FDR < 0.001; Chi-square test).
Functional classification of gene repertoire in the pangenome of P. multocida is shown in Figure 3B and details for COG-based binning are summarized in Table S4. The distribution of genes assigned to the major COG functional categories was remarkably different between core genes and dispensable/unique genes. In comparison to the dispensable and unique genes, the core genes encoding proteins involved in the fundamental metabolic activities were significantly abundant in the following COG categories: “Translation, ribosomal structure,” “Posttranslational modification,” “Amino acid metabolism,” “Nucleotide metabolism,” “Coenzyme metabolism” (FDR < 0.001; Chi-square test) (Figure 3B). Enrichment of genes involved in “Lipid metabolism” was also observed for the comparisons between core and dispensable genes (FDR < 0.001), and between core and unique genes (p-value < 0.001 and FDR = 0.13), respectively. Additionally, gene products of both categories “Carbohydrate metabolism” and “Energy metabolism” were more abundant in the set of core genes with less stringent FDR < 0.01 (Table S4). The core genes present in these functional categories, especially translation-associated genes, are essential for bacterial growth and survival. In contrast, genes coding for hypothetical proteins without homologs in the COG collection were accounting for about half of the dispensable genes and strain-specific genes, respectively (Table S4).
Gene Patterns of Virulence Factors
Based on BLAST searching against the VFDB database, we found 291 OGs encoding putative VFs in the pangenome of P. multocida. Details of these OGs together with Pfam functional domains were summarized in Table S5. Among these, 170 are core genes present in all P. multocida genomes tested and many of these encode products associated with common VFs. For instance, P. multocida LPS consist of lipid A, a core oligosaccharide, and a surface-exposed O-antigen (Christensen and Bisgaard, 2006). As is well known, O-antigen is the most variable component of the LPS, whereas, the structure of lipid A is highly conserved in general (Steimle et al., 2016). In accordance with the LPS structure, genes essential for biosynthesis of lipid A (lpxA, lpxB, lpxC, lpxD, lpxH, lpxK, htrB, msbB, kdsA, kdsB, kdtA, and kdkA) and core oligosaccharide (gmhA, rfaE, rfaF, and lgtF and waaQ) are uniformly present in all P. multocida strains (Table S5). In addition, pili have been observed with P. multocida (Christensen and Bisgaard, 2006) and consistently, five genes pilRQFBG involved in biogenesis of type IV pili were identified. In comparison to the core genes, accessory genes associated with bacterial virulence could be used as markers to distinguish pathovars (Rasko et al., 2008; Sahl et al., 2014). The distribution and conservation of virulence associated gene markers across P. multocida strains is shown in Figure 4 and discussed below.
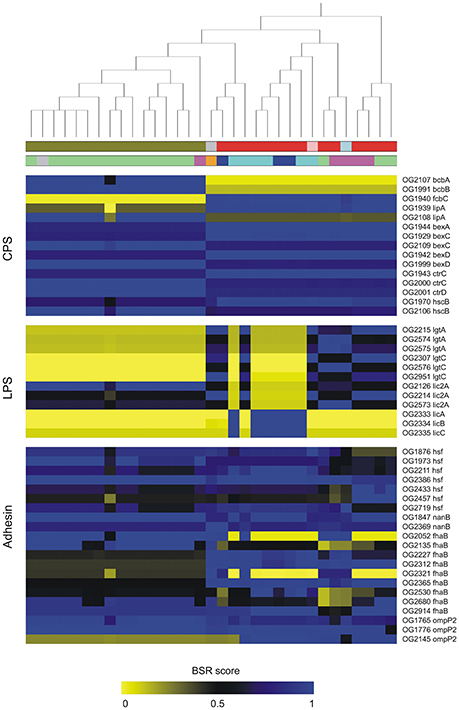
Figure 4. Distribution and conservation of selected genes coding for putative virulence factors in P. multocida. Putative virulence-associated genes were identified by blastp searching OGs against VFDB. The conservation of virulence markers in the pan-genome across all genomes is color-coded based on BLAST Score Ratio (BSR) values. The top dendrogram is the core SNP phylogeny. Color strips underneath the dendrogram represent serotypes and host origin with the same scheme shown in Figure 2. Gene annotations are detailed in Table S4.
Highly Conserved Genes in P. multocida
Among 1,559 single-copy core genes, 124 gene alignments containing few informative sites less than two were identified as highly conserved genes in P. multocida. Of the alignments of these conserved genes, 19 had no occurrence of nucleotide substitutions in the individual alignments. Notably, all these evolutionarily conversed genes were significantly abundant in the COG category “Translation” which 37 genes were affiliated to (p-value < 0.001; Chi-square test). It also confirms that genes involved in the translation machinery are evolving slowly with few nucleotide substitutions, perhaps due to functional constraints required by the fundamental cell cycle and bacterial survival (Jordan et al., 2002; Xu et al., 2011a).
Recombination of P. multocida Core Genes
Recombination test on the alignments of each single-copy core genes showed 7% (107 genes) were identified to exhibit significant evidence for homologous recombination in the individual alignments (FDR < 5%) (Table S6). Most of these genes had one recombinant breakpoint except for five genes (i.e., his7, his81, y112, dauA, and pstC) with multiple breakpoints. In comparison, Soyer reported that approximately 8% (270 of the 3,316 genes analyzed) of core genome genes present in five Salmonella genomes showed evidence for intragenic recombination (Soyer et al., 2009). Additionally, analysis of four E. coli and two Shigella genomes found 6% of core genes exhibited evidence for recombination (Petersen et al., 2007). Among the recombining genes in P. multocida, 13 are coding for putative virulence-associated proteins (Table S6). Besides, two genes (dnaB, replicative DNA helicase; glyS, Glycine–tRNA ligase beta subunit) have been experimentally confirmed to be preferentially expressed during acute P. multocida infection by selective capture of transcribed sequences (Guo et al., 2012).
Evidence for 35 P. multocida Genes Subject to Positive Selection
To detect potential genes of pathogenic P. multocida in response to host niches, positive selection analysis was performed on the alignments of non-recombinant genes/fragments using codeml-site-model with the nested models M1a/M2a, followed by multiple tests (FDR < 10%). A total of 35 genes were identified to be subject to strong selection pressure (Table 3). It was obvious that the genes were more abundant in the COG categories “Cell membrane biogenesis,” “General function prediction only,” “Function unknown,” and “Not in COG” (Table S7). Furthermore, based on the analyses of protein subcellular location, nearly half (n = 16) of all positively selected genes are coding for products localized on cell surface/membrane (Table 3). The enrichment of membrane-associated genes again supports a previous notion that positive selection driven by host immune and defense systems acts on surface/membrane structures in many pathogens (Fitzpatrick et al., 2005; Chen et al., 2006; Johnson et al., 2013). Some of the membrane-associated proteins have been implicated as crucial VFs associated with bacterial colonization and adaptation to host niches (Lin et al., 2002).
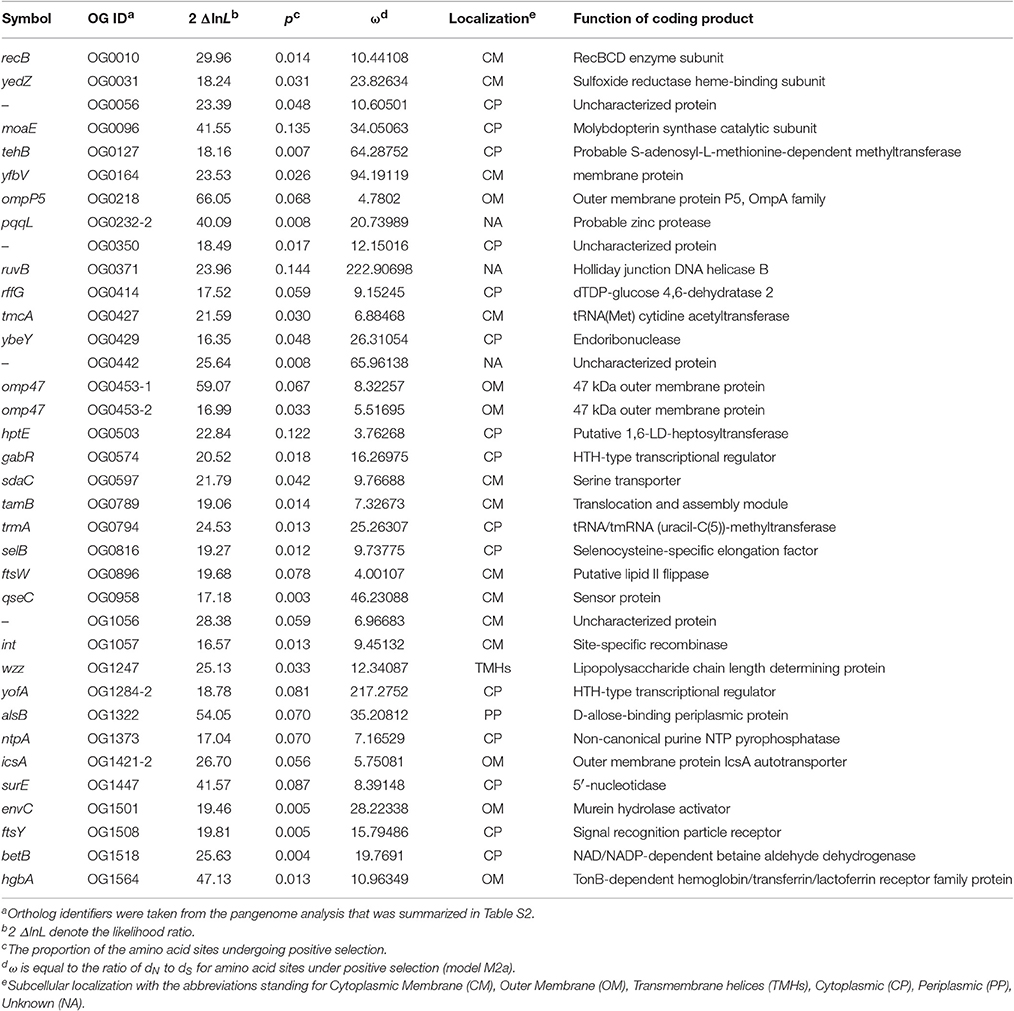
Table 3. P. multocida genes that show evidence for positive Darwinian selection.
Discussion
In this report, we performed deep genome sequencing for seven P. multocida strains. Together with 26 publicly available and high quality genome sequences of P. multocida, a comprehensive investigation was carried out to study pangenome characterization, genetic diversity of virulence genes, and evolutionary selection forces operating on the protein-coding genome of this zoonotic pathogen.
Diversity of Virulence Genes in P. multocida
Diversity of bacterial surface polysaccharides is often associated with genetic heterogeneity of the capsule and LPS loci (Howell et al., 2013). In the biosynthetic loci of P. multocida capsules, two genes bcbA and bcbB encoding enzymes that catalyze the conversion of UDP-N-acetylglucosamine to N-acetyl-D-mannosaminuronic acid (Townsend et al., 2001), were found to be serotype-B specific (Figure 4). In contrast, another capsule biosynthesis gene fcbC encoding UDP-glucose dehydrogenase is present in all strains except for the serotype-B strains (Figure 4). Two variants of lipA (OG1939, 696 aa; OG2108, 682 aa) encoding CPS modification protein are specific to serotype-B and non-serotype-B strains, respectively. Both products of lipA share 54% amino acid (aa) sequence identity (74% similarity) and 41% identity (61% similarity) with a LipA homolog from Neisseria meningitidis MC58, which is involved in the phospholipid modification of the CPS and its translocation to the cell surface (Frosch and Müller, 1993). In addition, it was apparent that five genes bexDCA and ctrCD involved in ATP-driven capsule export and gene hcs involved in post-polymerization (Satola et al., 2003) showed particular patterns of sequence conservation, which could be used to distinguish strains of serotype-B and non-serotype-B (Figure 4). On the other hand, glycosyltransferases participating in LPS biosynthesis have been considered to be serotype specific (Davies et al., 2013). Glycosyltransferases encoded by three accessary genes lgtA, lgtC, and lic2A were identified in the pangenome of P. multocida. Variants of both lgtA and lgtC are absent in all serotype-B strains and six serotype-A strains. Notably, three genes (OG2333–2335) that are present in the six type-A strains mentioned above share 41, 36, and 47% aa sequence identity, respectively, with the proteins LicA, LicB, and LicC encoded in the Haemophilus influenza lic operon responsible for the addition of phosphorylcholine to LPS (Humphries and High, 2002).
A dozen of genes coding for autotransporter (AT) proteins of the type V secretion pathway were identified in the pangenome of P. multocida (Figure 4), which are key players participating in virulence and pathogen-host interactions (Kline et al., 2009). Six variants of hsf encoding trimeric AT adhesins (TAAs) were found in the pangenome, all of which carry at least two of three typical TAA structure domains including an N-terminal head (PF05658), a stalk domain (PF05658), and a C-terminal membrane anchor (PF03895) (Table S5) (Mikula et al., 2012). Furthermore, two variants (OG1847, 1055 aa; OG2369, 1063 aa) of nanB encoding sialidase with 2-6′ and 2-3′ sialyl lactose specificity are present in the pangenome, which may contribute bacterial colonization on mucosal surfaces (Mizan et al., 2000). Both sialidases possess a signal peptide mediating protein transportation across the inner membrane and a C-terminal autotransporter domain (PF03797) responsible for transportation of the BNR repeat-like domain (PF13088) across the outer membrane (Leo et al., 2012). In addition, the protein products of nine fhaB variants are homologous to the Bordetella pertussis filamentous haemagglutinin (FHA), which is a surface-exposed protein functioning as both a primary adhesion and an immunomodulator (Inatsuka et al., 2005). It was worth nothing that the majority of these FHA-like proteins harbor an extended signal peptide of Type V secretion system (PF13018), a haemagglutination activity domain (PF05860) followed by the haemagluttinin repeats (PF13332) (Table S5). Mosaic patterns of sequence conservation revealed remarkable diversity of these virulence genes that could be used as genetic markers across the population of P. multocida. Detection of variants of virulence genes will facilitate the PCR-based epidemiological study to elucidate the associations between genotypes and phenotypes of P. multocida strains (Tang et al., 2009).
Positively Selected Genes in P. multocida
In Gram-negative bacteria, β-barrel outer membrane proteins are porins that mainly function in passive nutrient intake and active ion transport (Berven et al., 2004), as well as dynamic interactions with the host immune system (Massari et al., 2003). Bacterial transmembrane (TM) β-barrels consist of an even number of anti-parallel β-strands, generally 8–22 TM β-strands (Tamm et al., 2001). In our analysis, products of three genes (OG0453, omp47, 47 kDa outer membrane protein; OG1564, hgbA, TonB-dependent receptor family protein; OG0218, ompP5, outer membrane protein P5), which were predicted to be transmembrane β-barrel porins, showed strong evidence for positive selection (FDR < 10%; Table 3). The number of TM-strands in Omp47, HgbA, OmpP5 is 14, 20, and 8, respectively (Figure 5). The results of the BEB analyses showed that 14 amino acid residues of P. multocida Omp47 were subject to intensively positive selection (Table S7). Interestingly, all detected residues are located on the third and fourth extracellular loops, which are likely to be potential antigenic epitope. Omp47 is an adhesin binding to the host fibronectin and it has been experimentally validated to be immunogenic (Wheeler, 2009; Prasannavadhana et al., 2014). Genetic alterations on the extracellular region of Omp47 may be positively selected by recognition of fibronectin isoforms in different host niches. Notably, recombination signal was also observed in the alignment of omp47 (1,323 nt), with one breakpoint detected at position 651 (Table S6). Another β-barrrel porin, HgbA (971 aa) of P. multocida shares TonB dependent receptor family (PF00593) at the C-terminus and Plug domain (PF07715) at the N-terminus with H. influenza hemoglobin-haptoglobin binding protein HhuA (1025 aa) that is mostly involved in heme and iron acquisition (Maciver et al., 1996). Positive selection acting on hgbA may enable this pathogen to better scavenge host iron-binding complexes in various niches. OmpP5 is a major structural protein present in many gram-negative bacteria (Davies and Lee, 2004) and it has both immunodominant and host-adhesive domains (Novotny et al., 2000). Significant evidence for intensively positive selection on ompP5 was detected (FDR = 5.5e-11) and nine positively selected sites were all located in the extracellular space (Figure 5), perhaps associated with bacterial evasion of host immune responses. Pasteurella multocida OmpP5 (353 aa) shares high sequence similarity with the homologs outer membrane protein P5 (353 aa, 64% identity) of H. influenza and outer membrane protein A (OmpA, 346 aa, 47% identity) of E. coli. Similarly, a previous study has reported that E. coli OmpA that is a prime target of the host immune system during infection shows strong evidence of positive selection on the extracellular loops (Petersen et al., 2007).
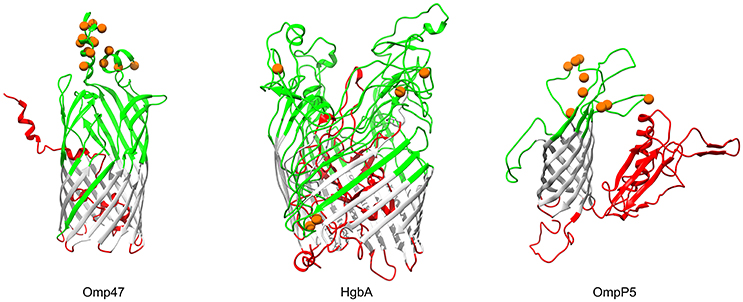
Figure 5. Three-dimensional structural models of β-barrel outer membrane proteins. Topology of three outer membrane proteins undergoing positive selection are color-coded according to the BOCTOPUS2 predictions (Hayat et al., 2016): red for inner-loops, gray for transmembrane β-strands, and green for outer-loops. Orange spheres stand for amino acid residues that are subject to strong positive selection (posterior probability > 95%).
Autotransporter proteins are typically VFs in P. multocida (Nicolay et al., 2015). The gene icsA (OG1421) encoding an autotransporter related to the Type V secretion system, was undergoing significantly positive selection (FDR = 0.002) (Table S7). It was worth noting that intragenic homologous recombination also occurred on icsA (2,556 nt), with one breakpoint at position 331 (Table S6). The outer membrane protein IcsA of P. multocida harbors a β-barrel domain (PF03797) at the C-terminal followed by an α-helical linker essential for outer membrane translocation of the passenger domain pertactin (PF03212) with characteristic β-helix (Nicolay et al., 2015). Structural mapping of seven positively selected sites on IcsA showed that the majority of sites (i.e., residues 159, 220, 315, 481, and 690) were located at the loops connecting the β-strands (Figure 6). Additionally, P. multocida IcsA shares 48% sequence similarity with the Shigella flexneri homolog that could interact with the host proteins vinculin and neural Wiskott-Aldrich syndrome protein implicated in the actin-based motility (Purdy et al., 2007).
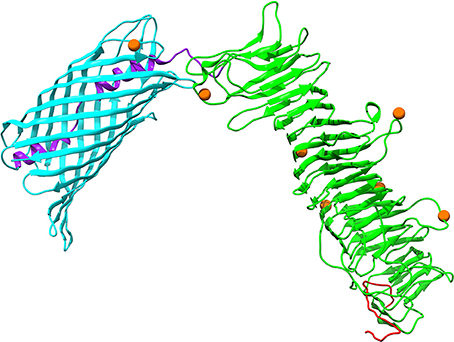
Figure 6. Three-dimensional structural models of the autotransporter protein IcsA. Protein secondary structure is color-coded based on the predicted PFAM domains and signal peptide cleavage site as well as modular organization previously described by Nicolay et al. (2015): an N-terminal signal peptide in red, a secreted mature passenger domain in green, and a C-terminal translocator domain in cyan with an upstream α-helical linker in purple. Orange spheres stand for amino acid residues that are subject to strong positive selection (posterior probability > 95%).
LPS in the outer membrane of P. multocida, which is a protective antigen stimulating humoral immunity, plays a critical role in the pathogenesis of disease (Harper et al., 2006). Three genes (rffG, OG0414, dTDP-glucose 4,6-dehydratase 2; ntpA, OG1373, non-canonical purine NTP pyrophosphatase; wzz, OG1247, LPS chain length determining protein) involved in biogenesis of LPS were subject to positive selection. The products of both genes ntpA and rffG share 83 and 64% identity with H. influenza OrfM and RffG, respectively, which are involved in the formation of bacterial endotoxin (Chen et al., 2016). Besides, the product of the positively selected gene wzz is an O-unit-processing enzyme, which was predicted to have two transmembrane helices (TMHs) (Figure S2) typical for the O-antigen chain length determining proteins of Gram-negative bacteria, such as H. parasuis and A. pleuropneumoniae (Xu et al., 2011b). Notably, three (77, 205, 241) out of four positively selected amino acid residues were located in the extracellular side (positions 36-247) of P. multocida Wzz, and the remaining site occurred in the TMH (positions 13-35) at the N-terminus.
The other gene ftsW coding for an integral membrane protein (Figure S2) functioning as a transporter of lipid-linked peptidoglycan precursors across the cytoplasmic membrane during cell division (Mohammadi et al., 2011), was also subject to positive selection (FDR = 0.028). Additionally, a functional unknown transmembrane protein encoded by the gene OG1056 (Figure S2) showed significant evidence for intensive positive selection (FDR < 0.001). The above analyses implicated that the adaptive changes is closely associated with the genes involved in the biosynthesis of cell surface/membrane components, perhaps due to interactions with the host immune and defense system.
Intriguingly, three P. multocida genes (recB, a subunit of RecBCD enzyme; int, site-specific recombinase; and ruvB, holliday junction DNA helicase B) with functional roles in DNA recombination and repair showed evidence for positive selection (Table 3). Similar phenomena have already been proposed for diversifying selection on the gene products constituting basal DNA repair machinery, such as recB in Neisseria (Yu et al., 2014), recC in E. coli (Chen et al., 2006) and basal DNA repair genes in ionizing-radiation-resistant bacteria (Sghaier et al., 2008). In addition, yedZ (OG0031) encoding a sulfoxide reductase heme-binding subunit with six TMHs (Figure S2), which was predicted to be an integral component of plasma membrane (GO: 0005887) and involved in protein repair (GO: 0030091), was also undergoing strong selection pressure. The precise function of these adaptive evolving genes associated with the metabolism of protein and DNA, to our knowledge, was not well studied in P. multocida and more experimental validation is desirable in the future.
Bacterial two-component regulatory systems and transcription factors are known to enable rapid adaptation to environmental conditions (Cases et al., 2003). We identified two genes gabR (OG0574) and yofA (OG1284) encoding HTH-type transcriptional regulators which showed significantly evidence for positive selection. Besides, the gene qseC (OG0958), which encodes a sensor kinase in the two-component QseBC quorum-sensing system, was also positively selected in P. multocida. Similar evidence for diversifying selection has been found in the PhoR/PhoB two component system in Brucella (Vishnu et al., 2015). These findings indicated positive selection of genes involved in transcriptional control may contribute to bacterial adaptation to various niches.
In summary, comparative genome analysis of 33 P. multocida strains revealed the pangenome compositions of P. multocida. Phylogeny of P. multocida was closely associated with serotyping rather than other factors like host origin or geographical location. Additionally, we performed a comprehensive analysis to investigate genetic content and diversification of virulence-associated genes in P. multocida. Our findings further indicated that evolutionary dynamics of pathogenic P. multocida were driven by positive Darwinian selection and homologous recombination. This study provides valuable targets for future researches on the mechanism of adaptive evolution and the host-pathogen interaction in P. multocida.
Author Contributions
Conceived and designed the experiment and wrote the paper: DG, ZX, and LQ. Performed the experiments: PC, DG, JL, and QJ. Contributed materials/analysis tools: DG and ZX. Analyzed the data: ZX, PC, and DG. All authors read and approved the final manuscript.
Funding
This work was supported by the National key research and development program (2016YFD0500800), and grants from the National Science Foundation of China (31302109).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00961/full#supplementary-material
References
Abrahante, J. E., Hunter, S. S., Maheswaran, S. K., Hauglund, M. J., Tatum, F. M., and Briggs, R. E. (2015). Draft genome sequence of Pasteurella multocida isolate P1062, isolated from bovine respiratory disease. Genome Announc. 3:e01254-15. doi: 10.1128/genomeA.01254-15
Abrahante, J. E., Veeregowda, B. M., Hogtapur, S. S., Briggs, R. E., Maheswaran, S. K., and Sreevatsan, S. (2014). Draft genome sequences of two Pasteurella multocida strains isolated from buffaloes in india with hemorrhagic septicemia disease. Genome Announc. 2:e00798-14. doi: 10.1128/genomeA.00798-14
Alikhan, N. F., Petty, N. K., Ben Zakour, N. L., and Beatson, S. A. (2011). BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics 12:402. doi: 10.1186/1471-2164-12-402
Benjamini, Y., and Yekutieli, D. (2001). The control of the false discovery rate in multiple testing under dependency. Ann. Statist. 29, 1165–1188. doi: 10.1186/1471-2105-9-114
Berven, F. S., Flikka, K., and Jensen, H. B. (2004). Eidhammer I. BOMP: a program to predict integral β-barrel outer membrane proteins encoded within genomes of Gram-negative bacteria. Nucleic Acids Res. 32, 394–399. doi: 10.1093/nar/gkh351
Boyce, J. D., Harper, M., Wilkie, I. W., and Adler, B. (2010). “Pasteurella,” in Pathogenesis of Bacterial Infections in Animals, ed J. F. Prescott (Ames, IA: Black-well Publishing), 327–328.
Boyce, J. D., Seemann, T., Adler, B., and Harper, M. (2012). Pathogenomics of Pasteurella multocida. Curr. Top. Microbiol. Immunol. 361, 23–38. doi: 10.1007/82_2012_203
Bruen, T. C., and Philippe, H. (2006). Bryant D: a simple and robust statistical test for detecting the presence of recombination. Genetics 172, 2665–2681. doi: 10.1534/genetics.105.048975
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., et al. (2009). BLAST+: architecture and applications. BMC Bioinformatics 10:421. doi: 10.1186/1471-2105-10-421
Carter, G. R. (1955). Studies on Pasteurella multocida. I. A haemagglutination test for the identification of serological types. Am. J. Vet. Res. 16, 481–484.
Cases, I., de Lorenzo, V., and Ouzounis, C. A. (2003). Transcription regulation and environmental adaptation in bacteria. Trends Microbiol. 11, 248–253. doi: 10.1016/S0966-842X(03)00103-3
Chen, L., Zheng, D., Liu, B., Yang, J., and Jin, Q. (2016). VFDB 2016: hierarchical and refined dataset for big data analysis-10 years on. Nucleic Acids Res. 44, 694–697. doi: 10.1093/nar/gkv1239
Chen, S. L., Hung, C. S., Xu, J., Reigstad, C. S., Magrini, V., Sabo, A., et al. (2006). Identification of genes subject to positive selection in uropathogenic strains of Escherichia coli: a comparative genomicsapproach. Proc. Natl. Acad. Sci. U.S.A. 103, 5977–5982. doi: 10.1073/pnas.0600938103
Christensen, H., and Bisgaard, M. (2006). “The Genus Pasteurella,” in The Prokaryotes, 3rd Edn. Vol. 6, eds M. Dworkin, S. Falkow, E. Rosenberg, K-H. Schleifer, and E. Stackebrandt (New York, NY: Springer), 1062–1090.
Davenport, K. W., Daligault, H. E., Minogue, T. D., Bishop-Lilly, K. A., and Bruce, D. C. (2014). Complete Genome Sequence of Type Strain Pasteurella multocida subsp. multocida ATCC 43137. Genome Announc. 2:e01070-14. doi: 10.1128/genomeA.01070-14
Davies, M. R., Broadbent, S. E., Harris, S. R., Thomson, N. R., and van der Woude, M. W. (2013). Horizontally acquired glycosyltransferase operons drive Salmonellae Lipopolysaccharide diversity. PLoS Genet. 9:e1003568. doi: 10.1371/journal.pgen.10035681
Davies, R. L., and Lee, I. (2004). Sequence diversity and molecular evolution of the heat-modifiable outer membrane protein gene (ompA) of Mannheimia (Pasteurella) haemolytica, Mannheimia glucosida, and Pasteurella trehalosi. J. Bacteriol. 186, 5741–5752. doi: 10.1128/JB.186.17.5741-5752.2004
Fitzpatrick, D. A., Creevey, C. J., and McInerney, J. O. (2005). Evidence of positive Darwinian selection in putative meningococcal vaccine antigens. J. Mol. Evol. 61, 90–98. doi: 10.1007/s00239-004-0290-6
Frosch, M., and Müller, A. (1993). Phospholipid substitution of capsular polysaccharides and mechanisms of capsule formation in Neisseria meningitidis. Mol. Microbiol. 8, 483–493. doi: 10.1111/j.1365-2958.1993.tb01592.x
Ghaffar, A., and Tariq, A. (2016). In-silico analysis of Pasteurella multocida to identify common epitopes between fowl, goat and buffalo. Gene 580, 58–66. doi: 10.1016/j.gene.2016.01.020
Goris, J., Konstantinidis, K. T., Klappenbach, J. A., Coenye, T., Vandamme, P., and Tiedje, J. M. (2007). DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 57, 81–91. doi: 10.1099/ijs.0.64483-0
Grant, J. R., Arantes, A. S., and Stothard, P. (2012). Comparing thousands of circular genomes using the CGView Comparison Tool. BMC Genomics 13:202. doi: 10.1186/1471-2164-13-202
Guindon, S., and Gascuel, O. (2003). A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 52, 696–704. doi: 10.1080/10635150390235520
Guo, D., Lu, Y., Zhang, A., Liu, J., Yuan, D., Jiang, Q., et al. (2012). Identification of genes transcribed by Pasteurella multocida in rabbit livers through the selective capture of transcribed sequences. FEMS Microbiol. Lett. 331, 105–112. doi: 10.1111/j.1574-6968.2012.02559.x
Harper, M., Boyce, J. D., and Adler, B. (2006). Pasteurella multocida pathogenesis:125 years after Pasteur. FEMS Microbiol. Lett. 265, 1–10. doi: 10.1111/j.1574-6968.2006.00442.x
Hayat, S., Peters, C., Shu, N., Tsirigos, K. D., and Elofsson, A. (2016). Inclusion of dyad-repeat pattern improves topology prediction of transmembrane β-barrel proteins. Bioinformatics. 32, 1571–1573. doi: 10.1093/bioinformatics/btw025
Hongo, J. A., de Castro, G. M., Cintra, L. C., Zerlotini, A., and Lobo, F. P. (2015). POTION: an end-to-end pipeline for positive Darwinian selection detection in genome-scale data through phylogenetic comparison of protein-coding genes. BMC Genomics 16:567. doi: 10.1186/s12864-015-1765-0
Howell, K. J., Weinert, L. A., Luan, S. L., Peters, S. E., and Chaudhuri, R. R. (2013). Gene content and diversity of the loci encoding biosynthesis of Capsular Polysaccharides of the 15 serovar reference strains of Haemophilus parasuis. J. Bacteriol. 195, 4264–4273. doi: 10.1128/JB.00471-13
Humphries, H. E., and High, N. J. (2002). The role of licA phase variation in the pathogenesis of invasive disease by Haemophilus influenzae type b. FEMS Immunol. Med. Microbiol. 34, 221–230. doi: 10.1111/j.1574-695X.2002.tb00628.x
Hyatt, D., Chen, G. L., Locascio, P. F., Land, M. L., Larimer, F. W., and Hauser, L. J. (2010). Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics. 11:119. doi: 10.1186/1471-2105-11-119
Inatsuka, C. S., Julio, S. M., and Cotter, P. A. (2005). Bordetella filamentous hemagglutinin plays a critical role in immunomodulation, suggesting a mechanism for host specificity. Proc. Natl. Acad. Sci. U.S.A. 102, 18578–18583. doi: 10.1073/pnas.0507910102
Johnson, T. J., Abrahante, J. E., Hunter, S. S., Hauglund, M., Tatum, F. M., Maheswaran, S. K., et al. (2013). Comparative genome analysis of an avirulent and two virulent strains of avian Pasteurella multocida reveals candidate genes involved in fitness and pathogenicity. BMC Microbiol. 13:106. doi: 10.1186/1471-2180-13-106
Jordan, I. K., Rogozin, I. B., Wolf, Y. I., and Koonin, E. V. (2002). Essential genes are more evolutionarily conserved than are nonessential genes in bacteria. Genome Res. 12, 962–968. doi: 10.1101/gr.87702
Kanehisa, M., Sato, Y., and Morishima, K. (2016). BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 428, 726–731. doi: 10.1016/j.jmb.2015.11.006
Kavousi, N., Eng, W. W. H., Lee, Y. P., Tan, L. H., Thuraisingham, R., Yule, C. M., et al. (2016). First high-quality draft genome sequence of Pasteurella multocida sequence type 128 isolated from infected bone. Genome Announc. 4:e00023-16. doi: 10.1128/genomeA.00023-16
Kelley, L. A., and Sternberg, M. J. (2015). Protein structure prediction on the Web: a case study using the Phyre server. Nat. Protoc. 4, 363–371. doi: 10.1038/nprot.2009.2
Kline, K. A., Fälker, S., Dahlberg, S., Normark, S., and Henriques-Normark, B. (2009). Bacterial adhesins in host-microbe interactions. Cell Host Microbe 5, 580–592. doi: 10.1016/j.chom.2009.05.011
Kosakovsky-Pond, S. L., Posada, D., Gravenor, M. B., Woelk, C. H., and Frost, S. D. (2006). GARD: a genetic algorithm for recombination detection. Bioinformatics 22, 3096–3098. doi: 10.1093/bioinformatics/btl474
Krogh, A., Larsson, B., von Heijne, G., and Sonnhammer, E. L. (2001). Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305, 567–580. doi: 10.1006/jmbi.2000.4315
Lagesen, K., Hallin, P., Rødland, E. A., Staerfeldt, H. H., Rognes, T., and Ussery, D. W. (2007). RNAmmer: consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 35, 3100–3108. doi: 10.1093/nar/gkm160
Lainson, F. A., Dagleish, M. P., Fontaine, M. C., Bayne, C., and Hodgson, J. C. (2013). Draft genome sequence of Pasteurella multocida A:3 strain 671/90. Genome Announc. 1:e00803-13. doi: 10.1128/genomeA.00803-13
Laslett, D., and Canback, B. (2004). ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 32, 11–16. doi: 10.1093/nar/gkh152
Lechner, M., Findeiss, S., Steiner, L., Marz, M., Stadler, P. F., and Prohaska, S. J. (2011). Proteinortho: detection of (co-)orthologs in large-scale analysis. BMC Bioinformatics 12:124. doi: 10.1186/1471-2105-12-124
Lefébure, T., and Stanhope, M. J. (2007). Evolution of the core and pan-genome of Streptococcus: positive selection, recombination, and genome composition. Genome Biol. 8:R71. doi: 10.1186/gb-2007-8-5-r71
Lefébure, T., and Stanhope, M. J. (2009). Pervasive, genome-wide positive selection leading to functional divergence in the bacterial genus Campylobacter. Genome Res. 19, 1224–1232. doi: 10.1101/gr.089250.108
Leo, J. C., Grin, I., and and Linke, D. (2012). Type V secretion: mechanism(s) of autotransport through the bacterial outer membrane. Philos. Trans. R. Soc. Lond B Biol. Sci. 367, 1088–1101. doi: 10.1098/rstb.2011.0208
Letunic, I., and Bork, P. (2011). Interactive Tree Of Life (iTOL): an online tool for phylogenetic treedisplay and annotation. Bioinformatics. 23, 127–128. doi: 10.1093/bioinformatics/btl529
Li, R., Zhu, H., Ruan, J., Qian, W., Fang, X., Shi, Z., et al. (2010). De novo assembly of human genomes with massively parallel short read sequencing. Genome. Res. 20, 265–272. doi: 10.1101/gr.097261.109
Lin, J., Huang, S., and Zhang, Q. (2002). Outer membrane proteins: key players for bacterial adaptation in host niches. Microb. Infect. 4, 325–331. doi: 10.1016/S1286-4579(02)01545-9
Liu, W., Yang, M., Xu, Z., Zheng, H., Liang, W., Zhou, R., et al. (2012). Complete genome sequence of Pasteurella multocida HN06, a toxigenic strain of serogroup D. J. Bacteriol. 194, 3292–3293. doi: 10.1128/JB.00215-12
Maciver, I., Latimer, J. L., Liem, H. H., Muller-Eberhard, U., Hrkal, Z., and Hansen, E. J. (1996). Identification of an outer membrane protein involved in utilization of hemoglobin-haptoglobin complexes by nontypeable Haemophilus influenzae. Infect. Immun. 64, 3703–3712.
Massari, P., Ram, S., Macleod, H., and Wetzler, L. M. (2003). The role of porins in neisserial pathogenesis and immunity. Trends Microbial. 11, 87–93. doi: 10.1016/S0966-842X(02)00037-9
May, B. J., Zhang, Q., Li, L. L., Paustian, M. L., Whittam, T. S., and Kapur, V. (2001). Complete genomic sequence of Pasteurella multocida, Pm70. Proc. Natl. Acad. Sci. U.S.A. 98, 3460–3465. doi: 10.1073/pnas.051634598
Michael, G. B., Kadlec, K., Sweeney, M. T., Brzuszkiewicz, E., Liesegang, H., Daniel, R., et al. (2012). ICEPmu1, an integrative conjugative element (ICE) of Pasteurella multocida: structure and transfer. J. Antimicrob. Chemother. 67, 91–100. doi: 10.1093/jac/dkr411
Mikula, K. M., Leo, J. C., Łyskowski, A., Kedracka-Krok, S., Pirog, A., and Goldman, A. (2012). The translocation domain in trimeric autotransporter adhesins is necessary and sufficient for trimerization and autotransportation. J. Bacteriol. 194, 827–838. doi: 10.1128/JB.05322-11
Mizan, S., Henk, A., Stallings, A., Maier, M., and Lee, M. D. (2000). Cloning and characterization of sialidases with 2-6′ and 2-3′ sialyl lactose specificity from Pasteurella multocida. J. Bacteriol. 182, 6874–6883. doi: 10.1128/JB.182.24.6874-6883.2000
Mohammadi, T., van Dam, V., Sijbrandi, R., Vernet, T., Zapun, A., Bouhss, A., et al. (2011). Identification of FtsW as a transporter of lipid-linked cell wall precursors across the membrane. EMBO J. 30, 1425–1432. doi: 10.1038/emboj.2011.61
Moustafa, A. M., Seemann, T., Gladman, S., Adler, B., Harper, M., Boyce, J. D., et al. (2015). Comparative genomic analysis of asian Haemorrhagic Septicaemia- associated strains of Pasteurella multocida identifies More than 90 Haemorrhagic Septicaemia- Specific Genes. PLoS ONE 10:e0130296. doi: 10.1371/journal.pone.0130296
Nicolay, T., Vanderleyden, J., and Spaepen, S. (2015). Autotransporter-based cell surface display in Gram-negative bacteria. Crit. Rev. Microbiol. 41, 109–123. doi: 10.3109/1040841X.2013.804032
Notredame, C., Higgins, D. G., and Heringa, J. (2000). T-Coffee: a novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 302, 205–217. doi: 10.1006/jmbi.2000.4042
Novotny, L. A., Jurcisek, J. A., Pichichero, M. E., and Bakaletz, L. O. (2000). Epitope mapping of the outer membrane protein P5-homologous fimbrin adhesin of nontypeable Haemophilus influenzae. Infect. Immun. 68, 2119–2128. doi: 10.1128/IAI.68.4.2119-2128.2000
Petersen, L., Bollback, J. P., Dimmic, M., Hubisz, M., and Nielsen, R. (2007). Genes under positive selection in Escherichia coli. Genome Res. 17, 1336–1343. doi: 10.1101/gr.6254707
Petersen, T. N., Brunak, S., von Heijne, G., and Nielsen, H. (2011). SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat. Methods 8, 785–786. doi: 10.1038/nmeth.1701
Pettersen, E. F., Goddard, T. D., Huang, C. C., Couch, G. S., Greenblatt, D. M., Meng, E. C., et al. (2004). UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612. doi: 10.1002/jcc.20084
Prasannavadhana, A., Kumar, S., Thomas, P., Sarangi, L. N., Gupta, S. K., Priyadarshini, A., et al. (2014). Outer membrane proteome analysis of Indian strain of Pasteurella multocida serotype B:2 by MALDI-TOF/MS analysis. Sci. World J. 2014:617034. doi: 10.1155/2014/617034
Price, M. N., Dehal, P. S., and Arkin, A. P. (2010). FastTree 2–approximately maximumlikelihood trees for large alignments. PLoS ONE 5:e9490. doi: 10.1371/journal.pone.0009490
Punta, M., Coggill, P. C., Eberhardt, R. Y., Mistry, J., Tate, J., Boursnell, C., et al. (2012). The Pfam protein families database. Nucleic Acids Res. 40, 290–301. doi: 10.1093/nar/gkr1065
Purdy, G. E., Fisher, C. R., and Payne, S. M. (2007). IcsA surface presentation in Shigella flexneri requires the periplasmic chaperones DegP, Skp, and SurA. J. Bacteriol. 189, 5566–5573. doi: 10.1128/J.B.00483-07
Rasko, D. A., Rosovitz, M. J., Myers, G. S., Mongodin, E. F., Fricke, W. F., Gajer, P., et al. (2008). The pangenome structure of Escherichia coli: comparative genomic analysis of E. coli commensal and pathogenic isolates. J. Bacteriol. 190, 6881–6893. doi: 10.1128/JB.00619-08
R Core Team (2015). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Sahl, J. W., Caporaso, J. G., Rasko, D. A., and Keim, P. (2014). The large-scale blast score ratio (LS-BSR) pipeline: a method to rapidly compare genetic content between bacterial genomes. Peer J. 2:e332. doi: 10.7717/peerj.332
Satola, S. W., Schirmer, P. L., and Farley, M. M. (2003). Complete sequence of the cap locus of Haemophilus influenzae serotype b and nonencapsulated b capsule-negative variants. Infect. Immun. 71, 3639–3644. doi: 10.1128/IAI.71.6.3639-3644.2003
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Sghaier, H., Ghedira, K., Benkahla, A., and Barkallah, I. (2008). Basal DNA repair machinery is subject to positive selection in ionizing-radiation-resistant bacteria. BMC Genomics 9:297. doi: 10.1186/1471-2164-9-297
Shimodaira, H., and Hasegawa, M. (1999). Multiple comparisons of log-likelihoods with applications to phylogenetic inference. Mol. Biol. Evol. 13, 964–969.
Soyer, Y., Orsi, R. H., Rodriguez-Rivera, L. D., Sun, Q., and Wiedmann, M. (2009). Genome wide evolutionary analyses reveal serotype specific patterns of positive selection in selected Salmonella serotypes. BMC Evol. Biol. 9:264. doi: 10.1186/1471-2148-9-264
Steimle, A., Autenrieth, I. B., and Frick, J. S. (2016). Structure and function: lipid A modifications in commensals and pathogens. Int. J. Med. Microbiol. 306, 290–301. doi: 10.1016/j.ijmm.2016.03.001
Suzek, B. E., Huang, H., McGarvey, P., Mazumder, R., and Wu, C. H. (2007). UniRef: comprehensive and non-redundant UniProt reference clusters. Bioinformatics 23, 1282–1288. doi: 10.1093/bioinformatics/btm098
Talavera, G., and Castresana, J. (2007). Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 56, 564–577. doi: 10.1080/10635150701472164
Tamm, L. K., Arora, A., and Kleinschmidt, J. H. (2001). Structure and assembly of beta-barrel membrane proteins. J. Biol. Chem. 276, 32399–32402. doi: 10.1074/jbc.R100021200
Tang, X., Zhao, Z., Hu, J., Wu, B., Cai, X., He, Q., et al. (2009). Isolation, antimicrobial resistance, and virulence genes of Pasteurella multocida strains from Swine in China. J. Clin. Microbiol. 47, 951–958. doi: 10.1128/JCM.02029-08
Tatusov, R. L., Galperin, M. Y., Natale, D. A., and Koonin, E. V. (2000). The COG database: a tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 28, 33–36. doi: 10.1093/nar/28.1.33
Toft, C., and Andersson, S. G. (2010). Evolutionary microbial genomics: insights into bacterial host adaptation. Nat. Rev. Genet. 11, 465–475. doi: 10.1038/nrg2798
Townsend, K. M., Boyce, J. D., Chung, J. Y., Frost, A. J., and Adler, B. (2001). Genetic organization of Pasteurella multocida cap Loci and development of a multiplex capsular PCR typing system. J. Clin. Microbiol. 39, 924–929. doi: 10.1128/JCM.39.3.924-929.2001
Treangen, T. J., Ondov, B. D., Koren, S., and Phillippy, A. M. (2014). The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 15:524. doi: 10.1186/s13059-014-0524-x
Vaid, R. K., Shanmugasundaram, K., Boora, A., Bera, B. C., Shukla, B. N., Anand, T., et al. (2014). Draft genome sequence of Pasteurella multocida subsp. multocida B:2 strain VTCCBAA264 isolated from Bubalus bubalis in North India. Genome Announc. 2:e00755-14. doi: 10.1128/genomeA.00755-14
Vishnu, U. S., Sankarasubramanian, J., Sridhar, J., Gunasekaran, P., and Rajendhran, J. (2015). Identification of recombination and positively selected genes in Brucella. Indian J. Microbiol. 55, 384–391. doi: 10.1007/s12088-015-0545-5
Wang, L., Sun, J., Guo, D., Cao, P., Liu, J., Liu, C., et al. (2016). Identification of capsule serotype and genotype of Pasteurella multocida in some areas of China. Chinese J. Prev. Vet. Med. 38, 116–119. doi: 10.3969/j.issn.1008-0589.2016.02.07 (In Chinese)
Wheeler, R. (2009). Outer membrane proteomics of Pasteurella multocida isolates to identify putative host-specificity determinants. Biosci. Horiz. 2, 1–12. doi: 10.1093/biohorizons/hzp002
Wong, W. S., Yang, Z., Goldman, N., and Nielsen, R. (2004). Accuracy and power of statistical methods for detecting adaptive evolution in protein coding sequences and for identifying positively selected sites. Genetics 168, 1041–1051. doi: 10.1534/genetics.104.031153
Xu, Z., Chen, H., and Zhou, R. (2011a). Genome-wide evidence for positive selection and recombination in Actinobacillus pleuropneumoniae. BMC Evol. Biol. 11:203. doi: 10.1186/1471-2148-11-203
Xu, Z., Yue, M., Zhou, R., Jin, Q., Fan, Y., Bei, W., et al. (2011b). Genomic characterization of Haemophilus parasuis SH0165, a highly virulent strain of serovar 5 prevalent in China. PLoS ONE 6:e19631. doi: 10.1371/journal.pone.0019631
Yang, Z. (2007). PAML 4: phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 24, 1586–1591. doi: 10.1093/molbev/msm088
Yang, Z., Wong, W. S., and Nielsen, R. (2005). Bayes empirical bayes inference of amino acid sites under positive selection. Mol. Biol. Evol. 22, 1107–1118. doi: 10.1093/molbev/msi097
Yu, D., Jin, Y., Yin, Z., Ren, H., Zhou, W., Liang, L., et al. (2014). A genome-wide identification of genes undergoing recombination and positive selection in Neisseria. BioMed. Res. Int. 2014:815672. doi: 10.1155/2014/815672
Yu, N. Y., Wagner, J. R., Laird, M. R., Melli, G., Rey, S., Lo, R., et al. (2010). PSORTb 3.0: improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 26, 1608–1615. doi: 10.1093/bioinformatics/btq249
Keywords: P. multocida, comparative genome analysis, virulence, genetic diversity, positive Darwinian selection
Citation: Cao P, Guo D, Liu J, Jiang Q, Xu Z and Qu L (2017) Genome-Wide Analyses Reveal Genes Subject to Positive Selection in Pasteurella multocida. Front. Microbiol. 8:961. doi: 10.3389/fmicb.2017.00961
Received: 06 February 2017; Accepted: 15 May 2017;
Published: 30 May 2017.
Edited by:
Rakesh Sharma, Institute of Genomics and Integrative Biology (CSIR), IndiaReviewed by:
Prabhu B. Patil, Institute of Microbial Technology (CSIR), IndiaBaojun Wu, Clark University, United States
Copyright © 2017 Cao, Guo, Liu, Jiang, Xu and Qu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhuofei Xu, emh1b2ZlaS54dUBnbWFpbC5jb20=
Liandong Qu, cWxkQGh2cmkuYWMuY24=
†These authors have contributed equally to this work.