 Tao Sun
Tao Sun Le Xu1,2,3
Le Xu1,2,3 Lei Chen
Lei Chen Weiwen Zhang
Weiwen Zhang- 1Laboratory of Synthetic Microbiology, School of Chemical Engineering and Technology, Tianjin University, Tianjin, China
- 2Key Laboratory of Systems Bioengineering, Ministry of Education of China, Tianjin, China
- 3Collaborative Innovation Center of Chemical Science and Engineering, Tianjin, China
- 4Center for Biosafety Research and Strategy, Tianjin University, Tianjin, China
Survival of photosynthetic cyanobacteria is challenged by environmental contaminations like heavy metals. Among them, deciphering the regulatory mechanisms for cadmium (Cd) in cyanobacteria would facilitate the construction of Cd-resistant strains. In this study, the DNA-Affinity-Purified-chromatin immunoprecipitation assay was employed to identify the direct targets of Sll0649, which was a Cd2+-related response regulator identified in our previous work in model cyanobacteria Synechocystis sp. PCC 6803. As a result, the promoter region of slr0946 encoding the arsenate reductase was enriched fourfolds by quantitative real time PCR analysis. Further, deletion of slr0946 led to a sensitive phenotype to Cd2+ stress compared with the wild type (WT) and the sensitive phenotype of Δslr0946 could be rescued by complementation assay via introducing slr0946 back into Δslr0946. Finally, individually overexpression of slr0946 as well as two Cd2+-related genes identified priviously (i.e., sll1598 and slr0798) in WT could significantly improve the tolerance of Synechocystis sp. PCC 6803 to Cd2+. This study provided a better understanding of the tolerance mechanism to Cd2+ in cyanobacteria and also feasible strategies for tolerance modifications to heavy metals in the future.
Introduction
Photosynthetic cyanobacteria are a large group of Gram-negative prokaryotes able to utilize CO2 and sunlight directly for growth (Gao et al., 2016), playing a pivotal role in the global carbon and nitrogen cycling as well as in some bioremediation applications (Rahman et al., 2011). Notably, cyanobacteria have also been utilized as “photosynthetic microbial factories” and dozens of biofuels and chemicals have been successfully synthesized in recent years (Gao et al., 2016). Though various metals were required for growth, survival of cyanobacteria is challenged by heavy metals like arsenic (As), cadmium (Cd), mercury (Hg), and lead (Pb), which are increasingly spread out in the environment through human activities (Cassier-Chauvat and Chauvat, 2015). Among them, Cd2+ is toxic to cyanobacteria even at a low concentration thus it is important to investigate the response mechanisms of cyanobacteria to Cd2+ and then modified the tolerance to Cd2+. Sas et al. (2006) monitored the effect of Cd2+ on the photosynthetic activity of model cyanobacteria Synechocystis sp. PCC 6803 (hereafter Synechocystis), founding that Cd2+ could penetrate rapidly into the cells and blocked the photosynthetic activity by altering the whole-chain electron transport (Sas et al., 2006). In addition, Toth et al. (2012) claimed that the toxic effect of Cd2+ on Synechocystis could be a cascade mechanism, in which the primary effect involved the rapid inhibition of CO2-dependent electron transport while the secondary effect related with inhibitory influences on PS II electron transport as well as the degradation of the reaction center protein D1 (Toth et al., 2012). Nevertheless, the response mechanisms of cyanobacteria to Cd2+ were rarely elucidated (Chen et al., 2014b). A better understanding of the regulation mechanisms of cyanobacteria to Cd2+ would benefit the tolerance modifications of cyanobacteria in the future.
Two-component systems (TCSs) are important regulatory mechanisms allowing microorganisms to sense and respond to environmental changes and stress conditions (Los et al., 2010). Each of them contains a histidine kinase (HK) serving as a sensor to perceive a specific environmental stimulus and a corresponding response regulator (RR) to regulate the expression level of target genes (West and Stock, 2001). Besides the participation of TCSs in biological processes such as membrane porin regulation (Yuan et al., 2011) and cell communications (von Bodman et al., 2008), recent studies have found the crucial roles of TCSs in bacterial stress responses like ion stress (Los et al., 2010; Prabhakaran et al., 2016). For example, the ColRS operon composed of a HK ColS and a RR ColR was found related with Cd2+ and Mn2+ resistance as a lack of ColRS caused a five-fold reduction in resistance to Mn2+ in Pseudomonas putida CD2 (Hu and Zhao, 2007). In addition, the TCSs of CzcRS in P. aeruginosa and two regulatory systems (i.e., CusRS and CueR) in Escherichia coli were proved related with Zn2+ and Cu2+ stress response, respectively (Grass and Rensing, 2001; Caille et al., 2007). These studies suggested the important roles of TCSs in bacterial ion stress regulation and its potential application in tolerance modifications.
In Synechocystis, more than 90 genes were believed to encode a HK or RR protein (Gao et al., 2016). Among them, several proteins have been functionally characterized and proved to be related with various biological processes as well as abiotic stresses (Los et al., 2010; Liu et al., 2015). Our recent efforts using functional genomics strategies to study the metabolic responses of Synechocystis to various abiotic stresses also discovered several stress-responsive genes including a RR gene slr1909 involving acid stress and two RR genes (i.e., sll0039 and slr1037) directly related with 1-butanol stress (Chen et al., 2014a; Ren et al., 2014; Niu et al., 2015). Notably, our previous study also identified a RR gene sll0649 involved in Cd2+ tolerance in Synechocystis (Chen et al., 2014b). Besides, sll1598 and slr0798 were proved to be the target of sll0649 via electrophoretic mobility shift assays (EMSAs) (Chen et al., 2014b). In this study, to further explore the Cd2+ resistance mechanism in Synechocystis, DNA-affinity-purified chip (DAP-chip) assays was employed to identify new targets of Sll0649. The DAP-chip assay successfully identified another new target, i.e., slr0946, related with Cd2+ stress response. In addition, individually overexpression of all three targets of sll0649 (i.e., sll1598, slr0798, and slr0946) could improve the tolerance of Synechocystis to Cd2+. Our work here provided new insights about the Cd2+ regulatory mechanisms in cyanobacteria and also feasible strategies for tolerance modifications to heavy metals.
Materials and Methods
Bacterial Culture Conditions
Escherichia coli BL21 (DE3) and E. coli DH5α were grown in the standard liquid LB medium or on agar plate with appropriate antibiotic (i.e., 10 μg/mL kanamycin) at 37°C using a shaking incubator at 130 rpm or incubator (Honour, Tianjin, China). Wild type Synechocystis (WT), mutants and the constructed strains were grown on agar plate or in BG11 medium at pH 7.5 using an illuming incubator or shaking incubator at a light intensity of approximately 50 μmol photons m-2s-1 and 130 rpm at 30°C (Honour, Tianjin, China). Medium for mutants and constructed strains was supplemented with appropriate antibiotic(s) (i.e., 10 μg/mL chloramphenicol and/or 10 μg/mL kanamycin). All strains and plasmids used in this study were listed in Table 1.

TABLE 1. Strains and plasmids used in this study.
Strains Construction
All primers used in this study were listed in Supplementary Table S1.
For gene deletion, the homologous recombination method was employed for the construction of gene knockout fragments for slr0946 (Chen et al., 2014b). Briefly, the chloramphenicol resistance cassette (amplified from pACYC184), two flanking homologous arms (about 1 kb) were employed for overlapping PCR and replacing the target gene of Synechocystis by natural transformation. The successful knockout mutant was confirmed by PCR and purified via successive passages.
For gene complementation and overexpression, a replicative vector pJA2 kindly provided by Prof. Paul Hudson (KTH Royal Institute of Technology of Sweden) was employed to overexpress sll0649, sll1598, slr0798, and slr0946, respectively (Huang et al., 2010; Kaczmarzyk et al., 2014). The resulting plasmid pJA2-sll0649 and pJA2-slr0946 was, respectively, back introduced into Δsll0649 and Δslr0946, leading to complementation strains Δsll0649-pJA0649 and Δslr0946-pJA0946. In addition, the resulting plasmid pJA2-slr0946, pJA2-sll0649, pJA2-sll1598, and pJA2-slr0798 were, respectively, introduced into the WT, leading to the overexpression strains WT-pJA0946, WT-pJA0649, WT-pJA1598, and WT-pJA0798. The transformation was performed using GenePulser Xcell (Bio-Rad, Hercules, CA, United States) (Sun et al., 2017). The positive colonies were validated by PCR.
Growth Patterns under Cd2+ Stress
For growth patterns, 5 mL fresh cells at OD630 nm = 0.2 were collected by centrifugation (4°C, 3000 × g for 15 min) and then were inoculated into 25 mL BG11 liquid medium in a 100 mL flask with or without CdSO4, each with three replicates (the concentration of CdSO4 was 4.6 μM for WT and deletion mutants but 5.0 μM for WT and overexpression strains). Cell density was measured on an ELx808 Absorbance Microplate Reader (BioTek, Winooski, VT, United States) at OD630 (Sun et al., 2017). Growth experiments were repeated at least three times to confirm the phenotype.
Overexpression and Purification of His6-Sll0649 Protein
Overexpression and purification of His6-Sll0649 protein were carried out as described previously (Chen et al., 2014b). Briefly, the sll0649 gene was amplified and then cloned to pET46 Ek/LIC vector, resulting in the plasmid pET46-sll0649. The pET46-sll0649 plasmid was then transformed into E. coli BL21 (DE3). The expression of His6-Sll0649 was induced by 0.1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) and followed by incubation at 22°C overnight. His6-Sll0649 was purified by the Ni-NTA agarose chromatography (GE healthcare, Uppsala, Sweden).
DAP-Chip Assay
DNA-affinity-purified chip assays were employed to identify the genes that directly regulated by Sll0649. Promoter regions of 10 selected genes were amplified by PCR and incubated with the purified recombinant His6-Sll0649 to allow the possible enrichment after elution according to the protocols described in the literature (Rajeev et al., 2011). Briefly, the binding reactions (500 μL) were set up with 12 to 18 μg of sheared Synechocystis genomic DNA (with an average length of 500 bp) and purified protein in the incubation buffer [20 mM Tris-HCl, pH 7.5; 1 mM dithiothreitol (DTT); 5 mM MgCl2; 0.04 mg/mL BSA and 25% glycerol (v/v)]. The reactions were incubated at 25°C in a thermal cycler for 30 min; 50 μL of the reaction was then cleaned up by Qiaquick PCR purification columns (Qiagen, Hilden, Germany) and saved as input DNA. The rest was loaded to the Ni-NTA agarose chromatography that had been washed in the binding/wash buffer [20 mM Tris-HCl, pH 7.5; 10 mM MgCl2; 50 mM KCl; 25% glycerol (v/v)]. The enriched DNA was specifically eluted from the resin with 500 μL elution buffer [20 mM Tris-HCl, pH 7.5; 500 mM NaCl; 600 mM imidazole; 10% glycerol (v/v)]. The enriched DNA fractions were cleaned up and saved as output DNA. Input DNA and output DNA were quantified using the Nanodrop 2000 (Thermo, CA, United States).
Quantitative Real Time PCR Analysis (qRT-PCR)
The qRT-PCR analysis was used to examine the enrichment fold of promoter regions of different genes after incubation with His6-Sll0649. Primers for qRT-PCR analysis were designed using Primer Express 2.0. To differentiate PCR products from primer dimers, primers were selected to generate amplicons with sizes around 100–200 bp. Experimental steps are based on the description previously (Sun et al., 2017). Three technical replicates were performed for each sample. Data analysis was carried out using the StepOnePlus analytical software (Applied Biosystems, Foster City, CA, United States) and the 2-ΔΔCT method (Livak and Schmittgen, 2001). The rnpB gene encoding RNase P subunit B was used as an internal control (Chen et al., 2014a). Then the enrichment fold of output DNA was relatively quantified compared to that of input DNA. All primers were provided in Supplementary Table S1.
Electrophoretic Mobility Shift Assay (EMSAs)
The EMSAs were performed as described previously (Chen et al., 2014b). Briefly, the promoter regions of slr0946 and slr1204 were amplified using the genomic DNA of Synechocystis and labeled with Cy5-labeled primer (5′-AGCCAGTGGCGATAAG-3′). The labeled PCR products were purified by QIAquick PCR Purification Kit (Qiagen, Hilden, Germany). In each EMSA reaction, ∼10 ng of Cy5-labeled DNA probes was incubated with varying amount of His6-Sll0649 protein in incubation buffer [1 mg/mL poly(dI–dC) (Roche, Basel, Switzerland), 20 mM Tris-base (pH 7.9), 1 mM DTT, 10 mM MgCl2, 0.2 mg/mL BSA and 5% glycerol (v/v)] for 20 min at 25°C. After incubation, protein-bound DNA and free DNA were separated by 6% Native-PAGE and viewed under Typhoon (GE healthcare, Uppsala, Sweden).
Results
Complementation of sll0649 in Δsll0649
In our previous work, a RR encoding gene sll0649 was identified involved in Cd2+ stress response in Synechocystis and Δsll0649 exhibited more sensitive phenotype to Cd2+ than WT (Chen et al., 2014b). In this work, to further confirm the involvement of sll0649 in Cd2+ tolerance, the sll0649 gene was placed under the control of the PpsbA2 promoter using a shuttle vector pJA2 and introduced back into the Δsll0649 mutant. The growth patterns among WT, Δsll0649 and Δsll0649-pJA0649 strains were then tested under normal medium and medium with 4.6 μM Cd2+. As illustrated in Figure 1, no obvious growth differences were observed for all three strains under both normal BG11 medium. Under 4.6 μM Cd2+ stress condition, the complementation stain (i.e., Δsll0649-pJA0649) was able to rescue the sensitive phenotype of the Δsll0649 to Cd2+, further suggesting the participation of sll0649 in Cd2+ regulation (Figure 1).
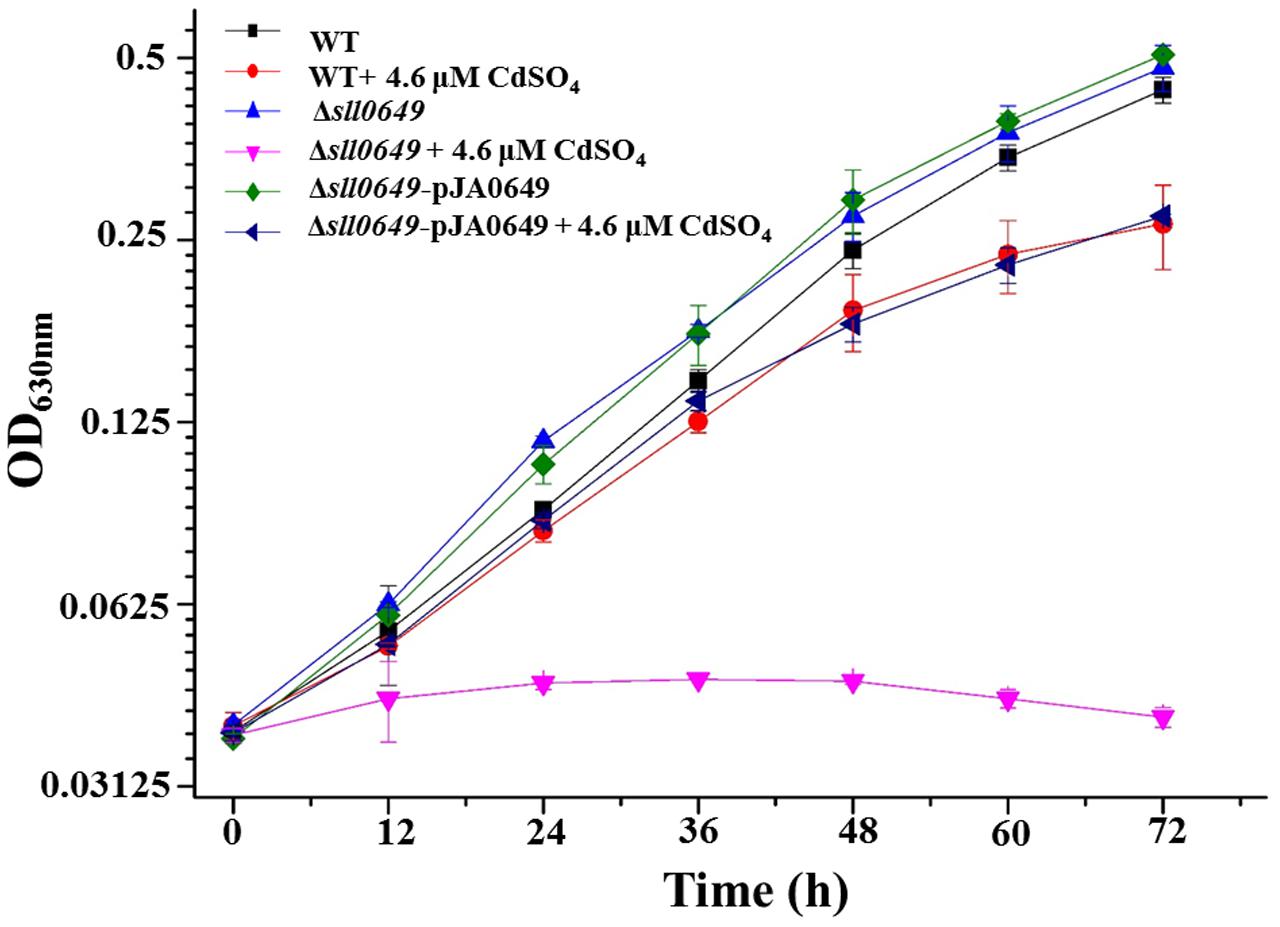
FIGURE 1. Growth patterns of WT, Δsll0649 and Δsll0649-pJA0649 strains in BG11 medium with or without 4.6 μM cadmium sulfate. The error bars represented the calculated standard deviation of the three biological replicates.
Identification of New Binding Targets of Sll0649 by DAP-Chip Assays
In our previous work, sll1598 and slr0798 have been identified as target genes of Sll0649 via EMSAs (Chen et al., 2014b). To further identify the new binding target(s) of Sll0649, DAP-chip strategy was employed. The full-length Sll0649 protein was first expressed in E. coli BL21(DE3) with a His6-tag at its N-termini. Extractive Synechocystis genomic DNA was sheared into 500–600 bp by sonification as input DNA. Purified His6-tagged Sll0649 proteins were incubated with sheared genomic DNA, and protein-bound DNA was purified using Ni-NTA resin to obtain the output DNA.
Then qRT-PCR was employed to determine the enrichment folds of different DNA regions. Ten candidate genes from upstream regions of sll0649 were selected according to the previous results of quantitative proteomics analysis (Chen et al., 2014b). Among these ten candidates, seven of them (i.e., sll0247, sll0248, slr0513, slr1204, slr0944, slr0945, and slr0946) were found down-regulated in Δsll0649 compared to WT under Cd2+ stress, and the other three (i.e., sll0041, sll0507, and sll0819) were randomly selected as negative controls. The rnpB gene was used as a control for normalization in this study. The results of qRT-PCR were listed in Table 2, in which slr0946 encoding the arsenate reductase was found enriched fourfolds among output Sll0649-bound DNAs compared to input DNA, suggesting it could be a new target of Sll0649.
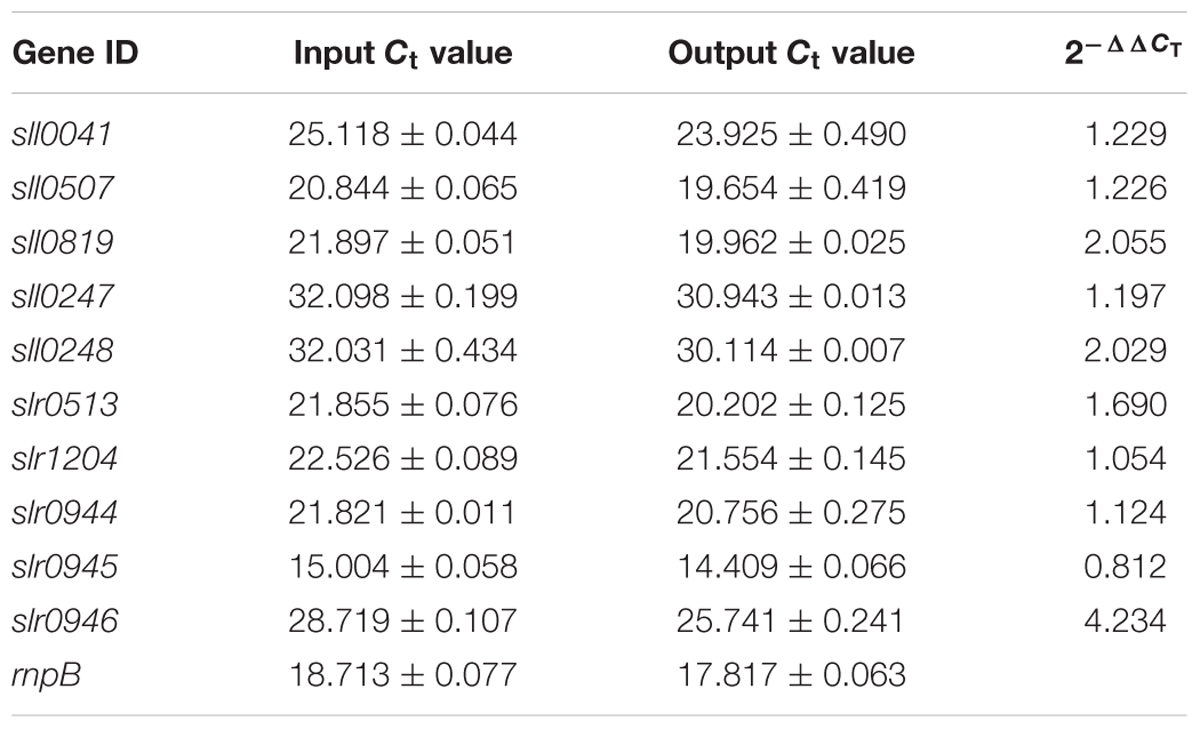
TABLE 2. The results of quantitative real time PCR analysis (qRT-PCR).
Validation of Binding Target of Sll0649 through EMSAs
In order to further verify the reliability of the new target slr0946, we performed EMSAs using purified His6-Sll0649 and the promoter region of slr0946. Meanwhile, the slr1204 gene encoding degP was selected as the negative control. As shown in Figure 2, clear gel-shift pattern for the purified His6-Sll0649 with Pslr0946 was investigated while no direct binding was observed for the His6-Sll0649 with Pslr1204 under the testing condition, suggesting that Sll0649 was able to bind directly to the promoter region of slr0946.
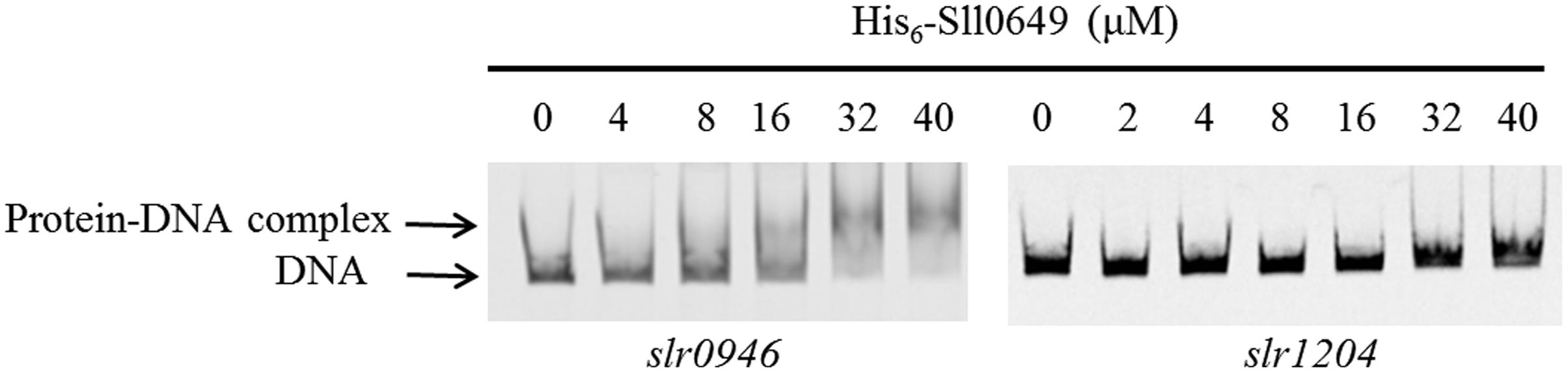
FIGURE 2. Electrophoretic mobility shift assays (EMSAs) to investigate the interaction of Sll0649 with promoter regions of slr0946 and slr1204. The amounts of His6-Sll0649 (μM) used were as indicated and 10 ng each of 5′-cy5-labeled probes was added in the reaction of EMSAs.
Functional Analysis of slr0946 by Knockout and Complementation Assays
To investigate the relationship of slr0946 with Cd2+ stress response, knockout mutant was generated by inserting the chloramphenicol resistance cassettes to the opening reading frame (ORF) of slr0946. The Δslr0946 mutant was viable and its growth rate in the normal BG11 medium was similar to that of the WT (Figure 3). However, under 4.6 μM Cd2+ stress condition, Δslr0946 was found more sensitive to Cd2+ than WT (Figure 3), indicating its involvement in Cd2+ stress response. We further constructed a complementary mutant named Δslr0946-pJA0946 by introducing the gene slr0946 back into Δslr0946 using a shuttle vector pJA2. As expected, the Δslr0946-pJA0946 strain was able to rescue the sensitive phenotype of Δslr0946 to Cd2+ in 4.6 μM Cd2+ stress (Figure 3), further confirming the participation of slr0946 in Cd2+ stress response.
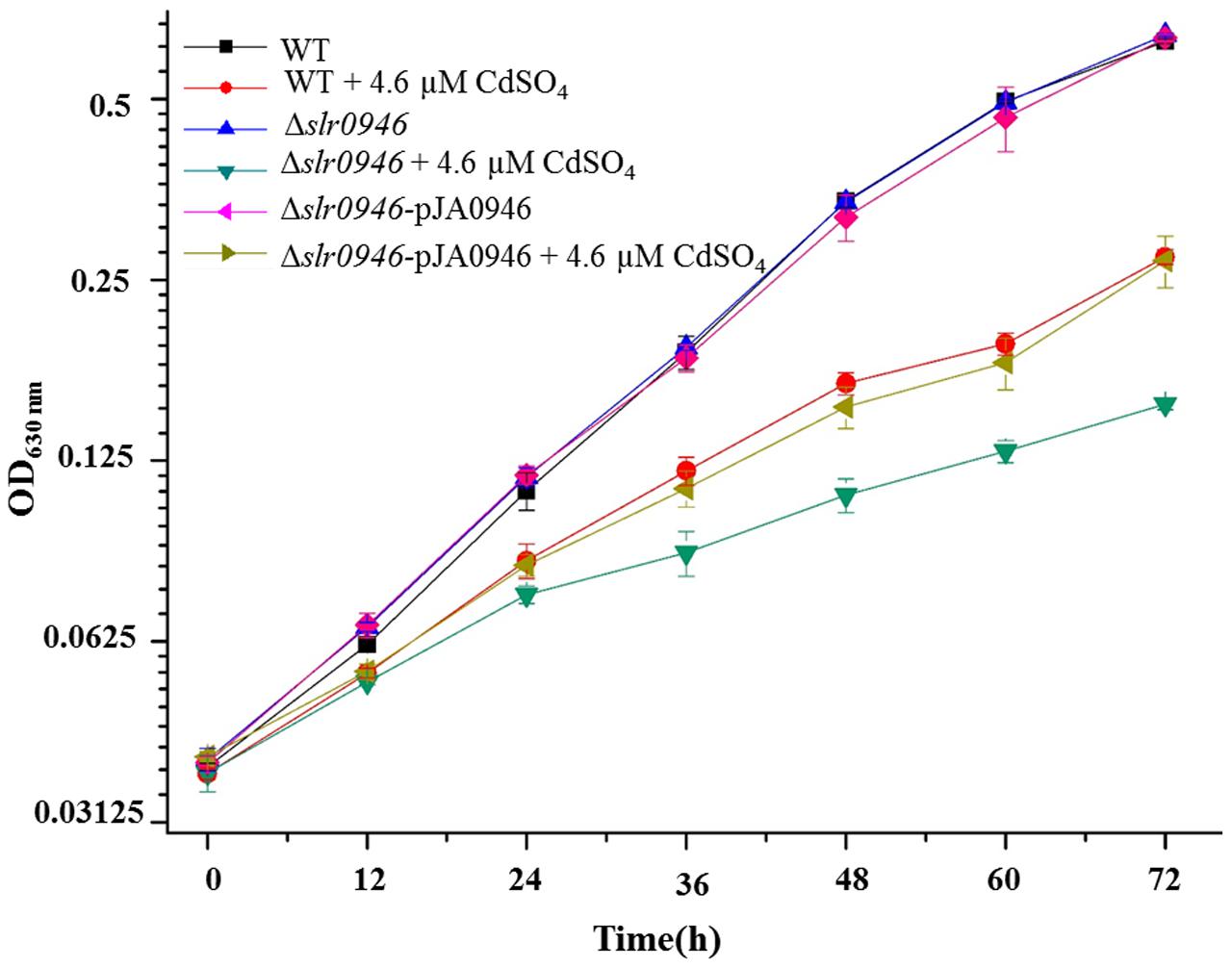
FIGURE 3. Growth patterns of WT, Δslr0946 and Δslr0946-pJA0946 in BG11 medium with or without 4.6 μM cadmium sulfate. The error bars represented the calculated standard deviation of the three biological replicates.
Tolerance Modifications to Cd2+ via Overexpressing sll0649, sll1598, slr0798, and slr0946
Engineered Cd2+-resistant strains in Synechocystis could be promising and useful for further Cd2+ tolerance modifications in other cyanobacterial chassis. In this study, aiming to improve the Cd2+ resistance of Synechocystis, we respectively, overexpressed four genes related to Cd2+ resistance, i.e., sll0649, sll1598, slr0798, and slr0946 in WT. The constructed strains were named as WT-pJA0649, WT-pJA1598, WT-pJA0798, and WT-pJA0946, respectively.
Growth patterns showed no visible differences among all the four overexpression strains in the normal BG11 medium compared to WT (Figure 4). Excitingly, three of the four overexpression strains, i.e., WT-pJA1598, WT-pJA0798, and WT-pJA0946 had significant tolerance improvement compared to WT under 5.0 μM Cd2+ stress condition (Figures 4B–D). This indicated that overexpression of any of the three target genes of Sll0649 (i.e., sll1598, slr0798, and slr0946) could improve the tolerance of WT to Cd2+. However, we found that overexpression of sll0649 can’t improve the tolerance of WT to Cd2+ due to some unknown reason (Figure 4A). To address this issue, the expression level of sll0649 was measured by qRT-PCR in WT and WT-pJA0649. The result showed that the transcriptional level of sll0649 gene in WT-pJA0649 was over 10-folds than that in WT (data not shown), suggesting that overexpressing sll0649 gene can’t improve Cd2+ tolerance in Synechocystis.
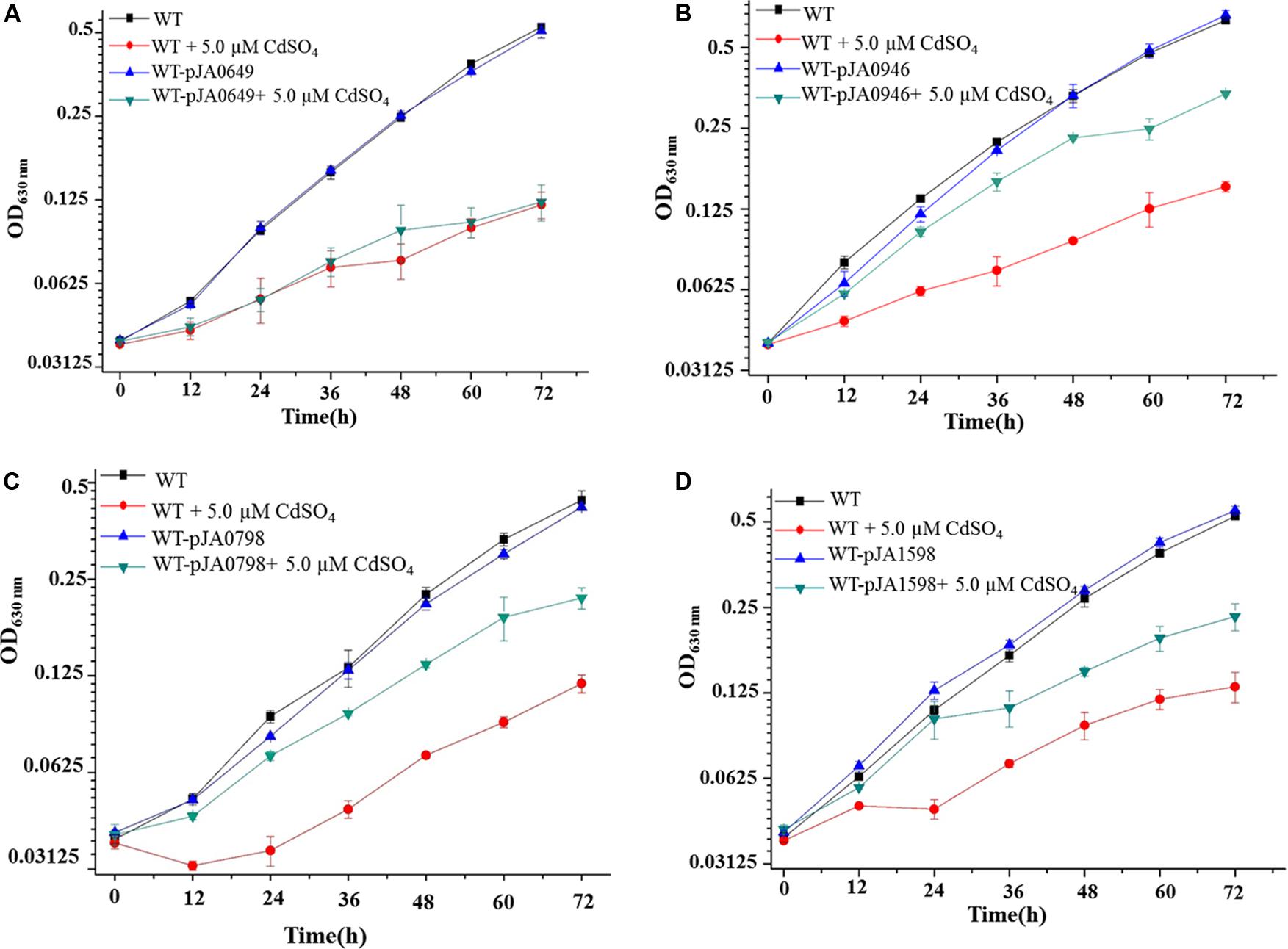
FIGURE 4. Growth patterns between WT and constructed strains. (A) Growth patterns between WT and WT-pJA0649 with or without 5.0 μM cadmium sulfate. (B) Growth patterns between WT and WT-pJA1598 with or without 5.0 μM cadmium sulfate. (C) Growth patterns between WT and WT-pJA0798 with or without 5.0 μM cadmium sulfate. (D) Growth patterns between WT and WT-pJA0946 with or without 5.0 μM cadmium sulfate. The error bars represented the calculated standard deviation of the three biological replicates.
Discussion
It is well known that microbes tend to employ multiple resistance mechanisms in dealing with a single stress (Prabhakaran et al., 2016). Thus, it could be challenging to achieve tolerance improvement by sequentially engineering multiple genes. As manipulation of a regulatory gene might achieve simultaneous modifications of series of genes related to tolerance (Los et al., 2010), it has been proposed as an alternative strategy to focus on regulatory genes for tolerance modifications. In our previous study, a RR Sll0649 was found related with Cd2+ stress response (Chen et al., 2014b). In this study, complementation of sll0649 in Δsll0649 rescued the sensitive phenotype though overexpression of sll0649 can’t improve the tolerance of WT to Cd2+. As qRT-PCR showed overexpressed transcripts of sll0649 in WT-pJA0649, we deduced that sll0649 could control the Cd2+ response mechanism in Synechocystis but its expression level might already be saturated in WT.
Biochemical and/or genetic approaches have historically been used to study DNA-protein interactions. Among them, ChiP was a powerful and useful tool to obtain information of binding sites for RR. For example, by employing DAP-chip, Rajeev et al. (2011) presented a systematic experimental determination of the genes regulated by two RRs in Desulfovibrio vulgaris Hildenborough (Rajeev et al., 2011). In our DAP-chip assays along with qRT-PCR, Sll0649 was found to directly bind to the promoter region of slr0946. The slr0946 gene appears to be located at the end of a gene cluster of slr0944-slr0945-slr0946 belonging to the arsBHC operon. However, our results showed that there might be different regulation mechanisms for these three genes, since only the promoter region of slr0946 was enriched (Table 2). Early studies have shown that the operon of slr0944-slr0945-slr0946 was involved in arsenic sensing and resistance system in Synechocystis (Li et al., 2003; Lopez-Maury et al., 2003). In addition, slr0946 encoding the arsenate reductase was found triggered by Cd in Synechocystis under the control of the regulator Slr1738 (Houot et al., 2007). In our previous study, Slr0946 was also among the down-regulated proteins in Δsll0649 after Cd2+ treatment (Chen et al., 2014b), which was consistent with the result that Slr0946 was triggered by Cd2+ stress (Houot et al., 2007). Notably, though overexpression of sll0649 had no tolerance improvement of WT to Cd2+, overexpression of any of its three target genes including slr0946, sll1598, and slr0798 could enhance the resistance to Cd2+, suggesting their relevance with Cd2+ stress response.
In E. coli, OmpR serving as a transcriptional factor promoted the transcription of ompF in conditions of low osmolality while repressing the transcription of ompF and activating the transcription of ompC at high osmolality (Martinez-Hackert and Stock, 1997). In addition, the DNA binding sequences for OmpR have been elucidated, which were in a tandem arrangement and conserved bases were separated from each other by ten base pairs, or roughly one helical turn (Martinez-Hackert and Stock, 1997). For Sll0649 of Synechocystis, it shares a high identity up to 41% to OmpR of E. coli using Blastp1, suggesting the potential similarity for their target DNA sequences. Thus, we tried to find the potential conserved target sequences among the promoter regions of slr0946, sll1598, and slr0798 according to the previous study (Martinez-Hackert and Stock, 1997). Interestingly, similar target sequences were found in all three genes (Figure 5 and Supplementary Table S2), further suggesting their reliability as the targets of Sll0649. The Cd2+ stress could be sensed by Sll0649, then leading to the transcriptional activation of slr0946, sll1598, and slr0798 related with stress response. In addition, Δsll0649 could hardly grew under 4.6 μM Cd2+ condition while growth of Δslr0946 was partially inhibited compared to Δsll0649. This could be due to two possible reasons: (i) functional redundancy existed between slr0946 and the other two targets thus partial function of slr0946 could be replaced by sll1598 and/or slr0798; (ii) the functional roles of sll0649 was more important than that of slr0946 as multiple genes could be activated by sll0649 thus deletion of sll0649 could cause a large deficiency of genes related with stress response.
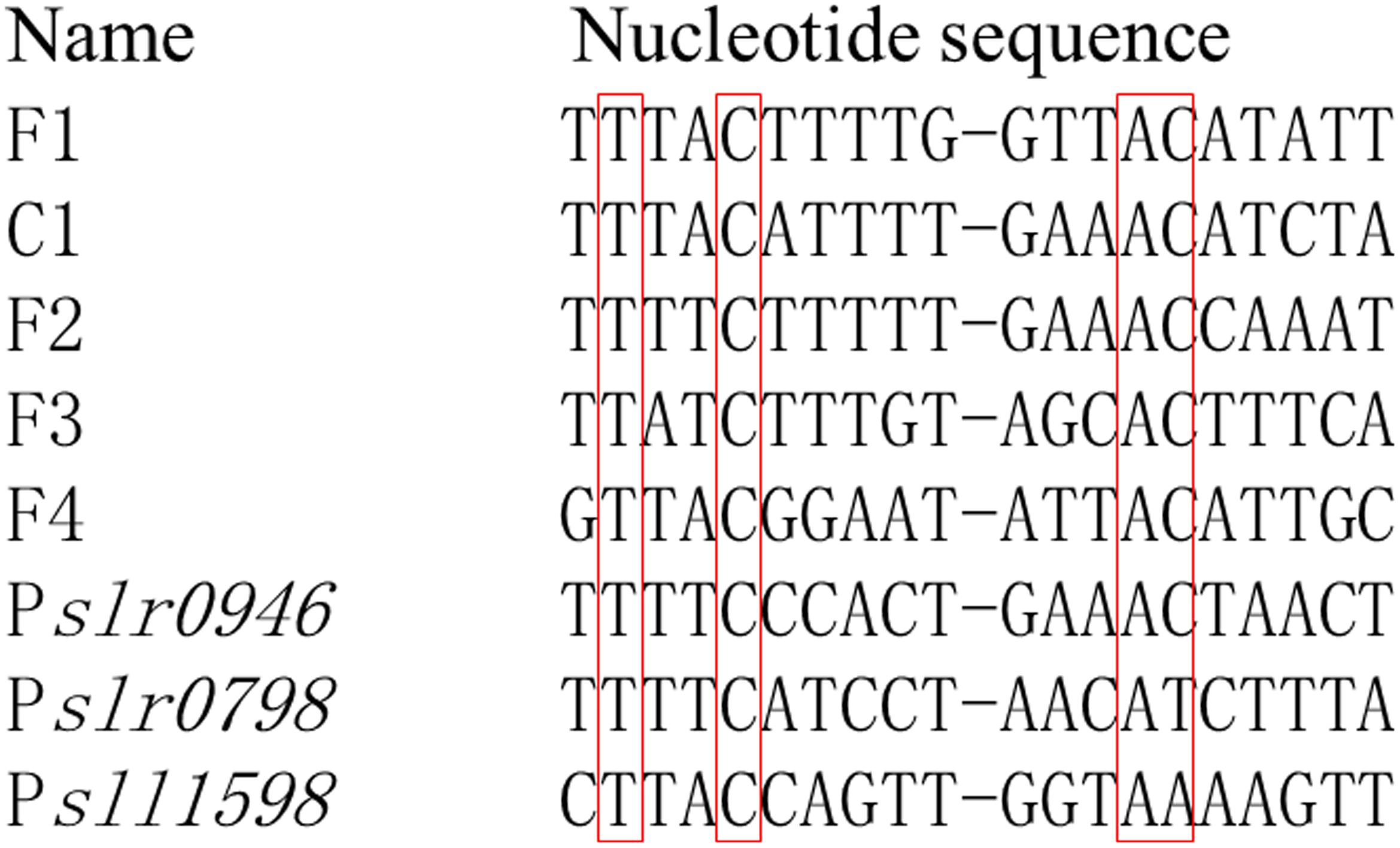
FIGURE 5. Potential conserved DNA-binding sequences for OmpR family RR Sll0649 existing in promoter regions of slr0946, slr0798, and sll1598. F1-F4 and C1 were reported previously as binding sequences for OmpR in Escherichia coli (Martinez-Hackert and Stock, 1997). Promoter regions for slr0946, slr0798, and sll1598 were selected from the intergenic sequence between each target gene and its flanking gene. The conserved nucleotides were shown in red box.
In this study, a new target gene of the RR Sll0649, slr0946, was identified using DAP-chip and EMSAs. In addition, tolerance of Synechocystis was enhanced through overexpression any of the three target genes of Sll0649. This study deepened the tolerance mechanism of cyanobacteria to heavy metals and provided feasible strategies for tolerance modifications.
Author Contributions
TS, LX, ZS, and LW performed the experiments. TS and LX wrote the manuscript. TS, LX, and LC analyzed the data. LC and WZ designed the study and revised the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The research was supported by grants from Natural Science Foundation of China (NSFC) (No. 31470217 and No. 21621004) and National Basic Research Program of China (National “973” program, project No. 2014CB745101) and the Tianjin Municipal Science and Technology Commission (No. 15JCZDJC32500).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.01582/full#supplementary-material
Footnotes
References
Caille, O., Rossier, C., and Perron, K. (2007). A copper-activated two-component system interacts with zinc and imipenem resistance in Pseudomonas aeruginosa. J. Bacteriol. 189, 4561–4568. doi: 10.1128/JB.00095-07
Cassier-Chauvat, C., and Chauvat, F. (2015). responses to oxidative and heavy metal stresses in cyanobacteria: recent advances. Int. J. Mol. Sci. 16, 871–886. doi: 10.3390/ijms16010871
Chen, L., Wu, L., Wang, J., and Zhang, W. (2014a). Butanol tolerance regulated by a two-component response regulator Slr1037 in photosynthetic Synechocystis sp. PCC 6803. Biotechnol. Biofuels 7:89. doi: 10.1186/1754-6834-7-89
Chen, L., Zhu, Y., Song, Z., Wang, J., and Zhang, W. (2014b). An orphan response regulator Sll0649 involved in cadmium tolerance and metal homeostasis in photosynthetic Synechocystis sp. PCC 6803. J. Proteomics 103, 87–102. doi: 10.1016/j.jprot.2014.03.029
Gao, X., Sun, T., Pei, G., Chen, L., and Zhang, W. (2016). Cyanobacterial chassis engineering for enhancing production of biofuels and chemicals. Appl. Microbiol. Biotechnol. 100, 3401–3413. doi: 10.1007/s00253-016-7374-2
Grass, G., and Rensing, C. (2001). Genes involved in copper homeostasis in Escherichia coli. J. Bacteriol. 183, 2145–2147. doi: 10.1128/JB.183.6.2145-2147.2001
Houot, L., Floutier, M., Marteyn, B., Michaut, M., Picciocchi, A., Legrain, P., et al. (2007). Cadmium triggers an integrated reprogramming of the metabolism of Synechocystis PCC6803, under the control of the Slr1738 regulator. BMC Genomics 8:350. doi: 10.1186/1471-2164-8-350
Hu, N., and Zhao, B. (2007). Key genes involved in heavy-metal resistance in Pseudomonas putida CD2. FEMS Microbiol. Lett. 267, 17–22. doi: 10.1111/j.1574-6968.2006.00505.x
Huang, H. H., Camsund, D., Lindblad, P., and Heidorn, T. (2010). Design and characterization of molecular tools for a synthetic biology approach towards developing cyanobacterial biotechnology. Nucleic Acids Res. 38, 2577–2593. doi: 10.1093/nar/gkq164
Kaczmarzyk, D., Anfelt, J., Sarnegrim, A., and Hudson, E. P. (2014). Overexpression of sigma factor SigB improves temperature and butanol tolerance of Synechocystis sp. PCC6803. J. Biotechnol. 18, 54–60. doi: 10.1016/j.jbiotec.2014.04.017
Li, R., Haile, J. D., and Kennelly, P. J. (2003). An arsenate reductase from Synechocystis sp. strain PCC 6803 exhibits a novel combination of catalytic characteristics. J. Bacteriol. 185, 6780–6789. doi: 10.1128/JB.185.23.6780-6789.2003
Liu, Z. X., Li, H. C., Wei, Y. P., Chu, W. Y., Chong, Y. L., Long, X. H., et al. (2015). Signal transduction pathways in Synechocystis sp. PCC 6803 and biotechnological implications under abiotic stress. Crit. Rev. Biotechnol. 35, 269–280. doi: 10.3109/07388551.2013.838662
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lopez-Maury, L., Florencio, F. J., and Reyes, J. C. (2003). Arsenic sensing and resistance system in the cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol. 185, 5363–5371.
Los, D. A., Zorina, A., Sinetova, M., Kryazhov, S., Mironov, K., and Zinchenko, V. V. (2010). stress sensors and signal transducers in cyanobacteria. Sensors 10, 2386–2415. doi: 10.3390/s100302386
Martinez-Hackert, E., and Stock, A. M. (1997). The DNA-binding domain of OmpR: crystal structures of a winged helix transcription factor. Structure 5, 109–124. doi: 10.1016/S0969-2126(97)00170-6
Niu, X., Zhu, Y., Pei, G., Wu, L., Chen, L., and Zhang, W. (2015). Elucidating butanol tolerance mediated by a response regulator Sll0039 in Synechocystis sp. PCC 6803 using a metabolomic approach. Appl. Microbiol. Biotechnol. 99, 1845–1857. doi: 10.1007/s00253-015-6374-y
Prabhakaran, P., Ashraf, M. A., and Aqma, W. S. (2016). Microbial stress response to heavy metals in the environment. RSC Adv. 6, 109862–109877. doi: 10.1039/C6RA10966G
Rahman, M. A., Soumya, K. K., Tripathi, A., Sundaram, S., Singh, S., and Gupta, A. (2011). Evaluation and sensitivity of cyanobacteria, Nostoc muscorum and Synechococcus PCC 7942 for heavy metals stress – a step toward biosensor. Toxicol. Environ. Chem. 93, 1982–1990. doi: 10.1080/02772248.2011.606110
Rajeev, L., Luning, E. G., Dehal, P. S., Price, M. N., Arkin, A. P., and Mukhopadhyay, A. (2011). Systematic mapping of two component response regulators to gene targets in a model sulfate reducing bacterium. Genome Biol. 12:R99. doi: 10.1186/gb-2011-12-10-r99
Ren, Q., Shi, M., Chen, L., Wang, J., and Zhang, W. (2014). Integrated proteomic and metabolomic characterization of a novel two-component response regulator Slr1909 involved in acid tolerance in Synechocystis sp. PCC 6803. J. Proteomics 109, 76–89. doi: 10.1016/j.jprot.2014.06.021
Sas, K. N., Kovacs, L., Zsiros, O., Gombos, Z., Garab, G., Hemmingsen, L., et al. (2006). Fast cadmium inhibition of photosynthesis in cyanobacteria in vivo and in vitro studies using perturbed angular correlation of gamma-rays. J. Biol. Inorg. Chem. 11, 725–734. doi: 10.1007/s00775-006-0113-x
Sun, T., Pei, G., Wang, J., Chen, L., and Zhang, W. (2017). A novel small RNA CoaR regulates coenzyme A biosynthesis and tolerance of Synechocystis sp. PCC6803 to 1-butanol possibly via promoter-directed transcriptional silencing. Biotechnol. Biofuels 10, 42. doi: 10.1186/s13068-017-0727-y
Toth, T., Zsiros, O., Kis, M., Garab, G., and Kovacs, L. (2012). Cadmium exerts its toxic effects on photosynthesis via a cascade mechanism in the cyanobacterium, Synechocystis PCC 6803. Plant Cell Environ. 35, 2075–2086. doi: 10.1111/j.1365-3040.2012.02537.x
von Bodman, S. B., Willey, J. M., and Diggle, S. P. (2008). Cell-cell communication in bacteria: united we stand. J. Bacteriol. 190, 4377–4391. doi: 10.1128/JB.00486-08
West, A. H., and Stock, A. M. (2001). Histidine kinases and response regulator proteins in two-component signaling systems. Trends Biochem. Sci. 26, 369–376. doi: 10.1016/S0968-0004(01)01852-7
Keywords: cyanobacteria, cadmium tolerance, response regulator, Sll0649, Slr0946
Citation: Sun T, Xu L, Wu L, Song Z, Chen L and Zhang W (2017) Identification of a New Target slr0946 of the Response Regulator Sll0649 Involving Cadmium Tolerance in Synechocystis sp. PCC 6803. Front. Microbiol. 8:1582. doi: 10.3389/fmicb.2017.01582
Received: 23 June 2017; Accepted: 04 August 2017;
Published: 15 August 2017.
Edited by:
Qiang Wang, Institute of Hydrobiology (CAS), ChinaReviewed by:
Weimin Ma, Shanghai Normal University, ChinaWenli Chen, Huazhong Agricultural University, China
Copyright © 2017 Sun, Xu, Wu, Song, Chen and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lei Chen, bGNoZW5AdGp1LmVkdS5jbg==